
Structural and histomorphometric evaluations of ferutinin effects on the uterus of ovariectomized...
8
Structural and histomorphometric evaluations of ferutinin effects on the uterus of ovariectomized rats during osteoporosis treatment Marzia Ferretti a , Laura Bertoni b , Francesco Cavani a , Marta Benincasa a , Paola Sena a , Gianluca Carnevale b , Manuela Zavatti a , Vittoria Di Viesti b , Paola Zanoli b , Carla Palumbo a, ⁎ a Section of Human Morphology of the Department of Biomedical Sciences, University of Modena and Reggio Emilia, Modena, Italy b Section of Pharmacology of the Department of Biomedical Sciences, University of Modena and Reggio Emilia, Modena, Italy abstract article info Article history: Received 5 July 2011 Accepted 27 October 2011 Keywords: Ferutinin Phytoestrogen Rat uterus Ovariectomy Apoptosis Aims: The effects of chronic administration of Ferutinin (phytoestrogen found in the plants of genus Ferula), compared with those elicited by estradiol benzoate, were evaluated, following ovariectomy, on the uterus of ovariectomized rats as regard weight, size, structure and histomorphometry. Main methods: The experimental study included 40 female Sprague–Dawley rats, assigned to two different pro- tocols, i.e. preventive and recovering. In the preventive protocol, ferutinin (2 mg/kg/day) was orally administered for 30 days, starting from the day after ovariectomy; in the recovering protocol, ferutinin was administered, at the same dosage, for 30 days starting from the 60th day after ovariectomy, when osteoporosis was clearly estab- lished. Its effects were compared with those of estradiol benzoate (1.5 μg per rat twice a week, subcutaneously injected) vs. vehicle-treated ovariectomized controls and vehicle-treated sham-operated controls. Uteri were re- moved, weighed and analysed under both the structural and histomorphometrical points of view. Key findings: Our data show that ferutinin acts, similarly to estradiol benzoate, on the uterus stimulating endo- metrial and myometrial hypertrophy; this notwithstanding, the phytoestrogen ferutinin, in contrast to estrogen treatment, appears to increase apoptosis in uterine luminal and glandular epithelia. Significance: Ferutinin, used in osteoporosis treatment primarily for bone mass recovering, seems in line with an eventual protective function against uterine carcinoma, unlike estrogens so far employed in hormone replace- ment therapy (HRT). © 2011 Elsevier Inc. All rights reserved. Introduction Ferutinin (jaeschkeanadiol p-hydroxybenzoate) is a phytoestro- genic compound found in plant roots of genus Ferula (Abourashed et al., 2001). Its estrogenic property was firstly demonstrated in vitro (Appendino et al., 2002). Specifically, ferutinin displayed the ability to bind estrogen receptors α (ERα) with a higher affinity (IC50 = 33.1 nM) in comparison with estrogen receptor β (ERβ) (IC50 = 180.5 nM) (Ikeda et al., 2002). Ferutinin was also found to act as agonist for ERα and as agonist/antagonist for ERβ: therefore it could be considered a selective estrogen receptor modulator (SERM) (Appendino et al., 2002; Ikeda et al., 2002). Recently, we eval- uated the ferutinin effects on sexual behavior of hormone-primed and non-hormone-primed female rats; in hormone-primed females, the acute administration of ferutinin significantly inhibited female recep- tivity (Zavatti et al., 2006), whereas in non hormone-primed rats the chronic administration of the compound was able to restore a normal sex- ual function, previously suppressed by ovariectomy (Zavatti et al., 2009). Further experiments performed administering ferutinin, alone or with es- tradiol benzoate (EB), in ovariectomized progesterone primed rats Life Sciences 90 (2012) 161–168 ⁎ Corresponding author at: University of Modena and Reggio Emilia, Department of Biomedical Sciences, Section of Human Morphology, c/o Istituti Anatomici, Area Policli- nico, Via del Pozzo 71, 41125, Modena, Italy. Tel.: +39 059 4224850; fax: +39 059 4224861. E-mail address: [email protected] (C. Palumbo). SHAM C-OVX EB-OVX F-OVX 0.0 0.1 0.2 0.3 0.4 0.5 0.6 ### *** *** Uterine weight (g) Fig. 1. Mean values of uterine weight in the preventive study. Values are expressed as mean ± SEM. ***P b 0.001 versus C-OVX; ### P b 0.001 versus SHAM (ANOVA followed by Newman–Keuls test). SHAM sham-operated controls receiving vehicle; C-OVX ovariectomized controls receiving vehicle; F-OVX ovariectomized treated with feruti- nin; EB-OVX ovariectomized treated with estradiol benzoate. 0024-3205/$ – see front matter © 2011 Elsevier Inc. All rights reserved. doi:10.1016/j.lfs.2011.11.001 Contents lists available at SciVerse ScienceDirect Life Sciences journal homepage: www.elsevier.com/locate/lifescie
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Structural and histomorphometric evaluations of ferutinin effects on the uterus of ovariectomized...

Life Sciences 90 (2012) 161–168
Contents lists available at SciVerse ScienceDirect
Life Sciences
j ourna l homepage: www.e lsev ie r .com/ locate / l i fesc ie
Structural and histomorphometric evaluations of ferutinin effects on the uterus ofovariectomized rats during osteoporosis treatment
Marzia Ferretti a, Laura Bertoni b, Francesco Cavani a, Marta Benincasa a, Paola Sena a, Gianluca Carnevale b,Manuela Zavatti a, Vittoria Di Viesti b, Paola Zanoli b, Carla Palumbo a,⁎a Section of Human Morphology of the Department of Biomedical Sciences, University of Modena and Reggio Emilia, Modena, Italyb Section of Pharmacology of the Department of Biomedical Sciences, University of Modena and Reggio Emilia, Modena, Italy
⁎ Corresponding author at: University of Modena andBiomedical Sciences, Section of Human Morphology, c/onico, Via del Pozzo 71, 41125, Modena, Italy. Tel.: +394224861.
E-mail address: [email protected] (C. Palum
0024-3205/$ – see front matter © 2011 Elsevier Inc. Alldoi:10.1016/j.lfs.2011.11.001
a b s t r a c t
a r t i c l e i n f oArticle history:
Received 5 July 2011Accepted 27 October 2011Keywords:FerutininPhytoestrogenRat uterusOvariectomyApoptosis
Aims: The effects of chronic administration of Ferutinin (phytoestrogen found in the plants of genus Ferula),compared with those elicited by estradiol benzoate, were evaluated, following ovariectomy, on the uterus ofovariectomized rats as regard weight, size, structure and histomorphometry.Main methods: The experimental study included 40 female Sprague–Dawley rats, assigned to two different pro-tocols, i.e. preventive and recovering. In the preventive protocol, ferutinin (2 mg/kg/day) was orally administeredfor 30 days, starting from the day after ovariectomy; in the recovering protocol, ferutinin was administered, atthe same dosage, for 30 days starting from the 60th day after ovariectomy, when osteoporosis was clearly estab-lished. Its effects were compared with those of estradiol benzoate (1.5 μg per rat twice a week, subcutaneouslyinjected) vs. vehicle-treated ovariectomized controls and vehicle-treated sham-operated controls. Uteri were re-
moved, weighed and analysed under both the structural and histomorphometrical points of view.Key findings: Our data show that ferutinin acts, similarly to estradiol benzoate, on the uterus stimulating endo-metrial and myometrial hypertrophy; this notwithstanding, the phytoestrogen ferutinin, in contrast to estrogentreatment, appears to increase apoptosis in uterine luminal and glandular epithelia.Significance: Ferutinin, used in osteoporosis treatment primarily for bonemass recovering, seems in line with aneventual protective function against uterine carcinoma, unlike estrogens so far employed in hormone replace-ment therapy (HRT).© 2011 Elsevier Inc. All rights reserved.
0.1
0.2
0.3
0.4
0.5
0.6
###
*** ***
Ute
rine
wei
ght (
g)
Introduction
Ferutinin (jaeschkeanadiol p-hydroxybenzoate) is a phytoestro-genic compound found in plant roots of genus Ferula (Abourashedet al., 2001). Its estrogenic property was firstly demonstratedin vitro (Appendino et al., 2002). Specifically, ferutinin displayed theability to bind estrogen receptors α (ERα) with a higher affinity(IC50=33.1 nM) in comparison with estrogen receptor β (ERβ)(IC50=180.5 nM) (Ikeda et al., 2002). Ferutinin was also found toact as agonist for ERα and as agonist/antagonist for ERβ: thereforeit could be considered a selective estrogen receptor modulator(SERM) (Appendino et al., 2002; Ikeda et al., 2002). Recently, we eval-uated the ferutinin effects on sexual behavior of hormone-primedand non-hormone-primed female rats; in hormone-primed females,the acute administration of ferutinin significantly inhibited female recep-tivity (Zavatti et al., 2006), whereas in non hormone-primed rats the
Reggio Emilia, Department ofIstituti Anatomici, Area Policli-059 4224850; fax: +39 059
bo).
rights reserved.
chronic administration of the compoundwas able to restore a normal sex-ual function, previously suppressed by ovariectomy (Zavatti et al., 2009).Further experiments performed administering ferutinin, alone orwith es-tradiol benzoate (EB), in ovariectomized progesterone primed rats
SHAM C-OVX EB-OVX F-OVX0.0
Fig. 1. Mean values of uterine weight in the preventive study. Values are expressed asmean±SEM. ***Pb0.001 versus C-OVX; ###Pb0.001 versus SHAM (ANOVA followedby Newman–Keuls test). SHAM sham-operated controls receiving vehicle; C-OVXovariectomized controls receiving vehicle; F-OVX ovariectomized treated with feruti-nin; EB-OVX ovariectomized treated with estradiol benzoate.

SHAM C-OVX EB-OVX F-OVX0.0
0.1
0.2
0.3
0.4
0.5
0.6
###
***
***
Ute
rine
wei
ght (
g)
Fig. 2. Mean values of uterine weight in the recovering study. Values are expressed asmean±SEM. ***Pb0.001 versus C-OVX; ###Pb0.001 versus SHAM (ANOVA followedby Newman–Keuls test). SHAM sham-operated controls receiving vehicle; C-OVXovariectomized controls receiving vehicle; F-OVX ovariectomized treated with feruti-nin; EB-OVX ovariectomized treated with estradiol benzoate.
162 M. Ferretti et al. / Life Sciences 90 (2012) 161–168
demonstrated ferutinin property in increasing ERα expression in the hy-pothalamuswhenadministered alone, like estradiol, but in decreasing theresponse to estradiol when administered in combination (Zanoli et al.,2009). Thus ferutinin seems to display estrogenic or antiestrogenic activ-ity, through hypothalamic ERα, depending on absence/presence of estro-gen priming. From a clinical viewpoint our findings suggest potentialtherapeutic effects of ferutinin in estrogen-deficiency like menopause.This hypothesis was confirmed by our recent results from osteoporoticovariectomized rats; ferutinin was found to counteract the reduction inbone density due to estrogen deficiency, following chronic treatmentstarting the day after ovariectomy (Palumbo et al., 2009) or 2 monthslater, when osteoporosis was clearly evident (Ferretti et al., 2010). Inboth studies, ferutinin displayed the same effects on bonemass observed
Fig. 3. LM micrographs of Gomori trichrome–stained cross sections showing the uterine struovariectomized controls receiving vehicle; F-OVX ovariectomized treated with ferutinin; EB
with estradiol benzoate, thus suggesting that it could prevent osteoporo-sis and enhance bone loss recovery in osteoporotic ovariectomized rats.
Overall, our in vivo findings confirm ferutinin estrogenic property,suggesting its potential benefit in reducing symptoms and degenerativeprocesses associated tomenopause. Now it is crucial to evaluate ferutininside effects, specifically on the organs which are reputed to be the targetof estrogen effects, like uterus, vagina, mammary glands. It is well knownthat estrogens stimulate endometrial proliferation and their administra-tion in HRTwas associated to an increased risk of cancer. Phytoestrogensare claimed to have beneficial effectswith aminor incidence of undesiredside effects in comparison with estrogen therapy. In general, phytoestro-gens showed a higher binding affinity for ERβ than ERα (Kuiper et al.,1998) and displayed both agonist and antagonist effects (Patisaul et al.,2005). Proliferative activity in estrogen-responsive cells can be either en-hanced or suppressed by phytoestrogens depending on their concentra-tion and relative potency (Whitten and Patisaul, 2001). Clinical reportsabout phytoestrogen effect on endometrial cancer are limited to case-controlled observational studies (Johnson et al., 2001).
The present study was designed to compare the effects of thechronic ferutinin treatment with those induced by estradiol benzoateon the uterus of ovariectomized rats.
Materials and methods
Animals and treatments
All animal handling and experimental conditions were carried outaccording to the Italian law (D.L. n. 116/1992) and European legisla-tion (EEC n. 86/609). The experimental design and procedures wereconducted under protocols approved by the Bioethical Committee ofthe Italian National Institute of Health.
cture in the preventive study. SHAM sham-operated controls receiving vehicle; C-OVX-OVX ovariectomized treated with estradiol benzoate.
163M. Ferretti et al. / Life Sciences 90 (2012) 161–168
Forty female Sprague–Dawley rats, aged 7 weeks according to thegeneral age-models used by Kalu (1991) and Fanti et al. (1998) andweighing 170–190 g at the beginning of the experiments, wereobtained from Harlan Italy (Udine, Italy). They were housed two percage and maintained in standard laboratory conditions at a tempera-ture of 22±1 °C, relative humidity 55%–60% and a 12 h light/darkcycle. The animals had free access to commercial rat pellets free of es-trogenic substances (Global Diet 2018, Mucedola s.r.l., Milan, Italy)and water. After a 7-day adaptation period to the controlled laborato-ry conditions, the animals were randomly assigned to two differentprotocols in order to study ferutinin effects on the uterus in two con-ditions, i.e. in preventive (Palumbo et al., 2009) and recovering(Ferretti et al., 2010) estrogen deficiency osteoporosis. Five rats ofeach protocol were sham-operated, while the others were bilaterallyovariectomized (OVX) under anesthesia induced by intraperitonealinjections of ketamin hydrochloride (Ketavet 100®, Farmaceutici Gel-lini S.p.a, Italy) plus xylazine hydrochloride (Rompun®, Bayer,Germany).
The animals of the preventive study were submitted to the follow-ing treatment for 30 days:
Group 1 (SHAM): Sham-operated controls receiving vehicle (5%Tween 80 in water);Group 2 (C-OVX): Ovariectomized controls receiving vehicle (5%Tween 80 in water);Group 3 (F-OVX): Ovariectomized treated with ferutinin 2 mg/kg/day;Group 4 (EB-OVX): Ovariectomized treated with estradiol benzo-ate 1.5 μg/rat twice a week.
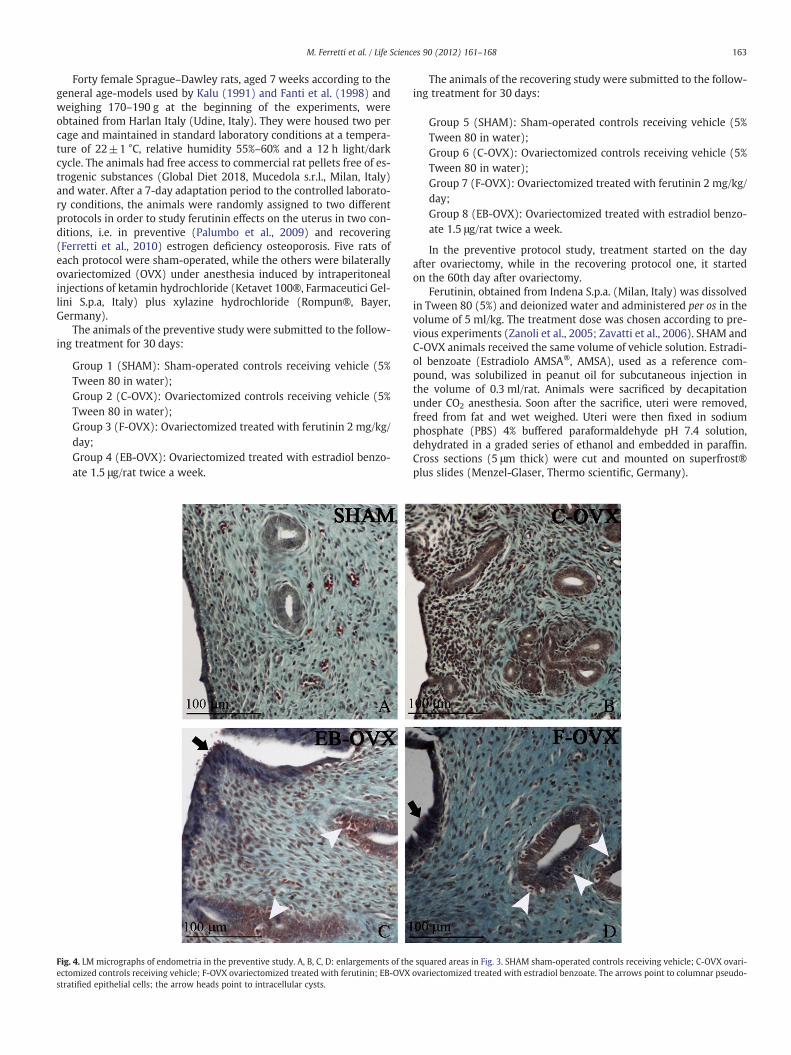
Fig. 4. LM micrographs of endometria in the preventive study. A, B, C, D: enlargements of theectomized controls receiving vehicle; F-OVX ovariectomized treated with ferutinin; EB-OVXstratified epithelial cells; the arrow heads point to intracellular cysts.
The animals of the recovering study were submitted to the follow-ing treatment for 30 days:
Group 5 (SHAM): Sham-operated controls receiving vehicle (5%Tween 80 in water);Group 6 (C-OVX): Ovariectomized controls receiving vehicle (5%Tween 80 in water);Group 7 (F-OVX): Ovariectomized treated with ferutinin 2 mg/kg/day;Group 8 (EB-OVX): Ovariectomized treated with estradiol benzo-ate 1.5 μg/rat twice a week.
In the preventive protocol study, treatment started on the dayafter ovariectomy, while in the recovering protocol one, it startedon the 60th day after ovariectomy.
Ferutinin, obtained from Indena S.p.a. (Milan, Italy) was dissolvedin Tween 80 (5%) and deionized water and administered per os in thevolume of 5 ml/kg. The treatment dose was chosen according to pre-vious experiments (Zanoli et al., 2005; Zavatti et al., 2006). SHAM andC-OVX animals received the same volume of vehicle solution. Estradi-ol benzoate (Estradiolo AMSA®, AMSA), used as a reference com-pound, was solubilized in peanut oil for subcutaneous injection inthe volume of 0.3 ml/rat. Animals were sacrificed by decapitationunder CO2 anesthesia. Soon after the sacrifice, uteri were removed,freed from fat and wet weighed. Uteri were then fixed in sodiumphosphate (PBS) 4% buffered paraformaldehyde pH 7.4 solution,dehydrated in a graded series of ethanol and embedded in paraffin.Cross sections (5 μm thick) were cut and mounted on superfrost®plus slides (Menzel-Glaser, Thermo scientific, Germany).
squared areas in Fig. 3. SHAM sham-operated controls receiving vehicle; C-OVX ovari-ovariectomized treated with estradiol benzoate. The arrows point to columnar pseudo-
164 M. Ferretti et al. / Life Sciences 90 (2012) 161–168
Uterine structure and morphometry
The general morphology of the tissues, as well as the cellularshape of the luminal and glandular epithelia, was analyzed in theGomori trichrome-stained sections following standard procedures.Additionally, cross-sectional areas of endometrium and myometriumwere measured using a light microscope (Zeiss Axiophot—Jena, WestGermany) equipped with an image analysis system (Nikon DS-5Mccamera connected to a personal computer—NIS Elements AR 2.20Nikon software). Measurements were performed on three differentsections for each animal.
In situ end-labelling analysis (TUNEL—ApopTag® Peroxidase Kits,Millipore)
After being deparaffinated and rehydrated, the sectionswere treatedin a humidified chamber with proteinase K (Boehringer, Mannheim,Germany) at 20 μg ml−1 for 15 min at room temperature, washed inbidistilled water, treated with 2% H2O2 in methanol for 10 min at roomtemperature and then washed in bidistilled water. The slides werepre-incubated with terminaldeoxynucleotidyl transferase (TdT) bufferand 1 mM CoCl2 for 5 min at room temperature and then incubated for60 min. in a humidified chamber at 37 °Cwith 50 μl TdT and biotinylateddeoxyuridine triphosphate (Bio dUTP) (Boehringer) (TdT 0.3 U μl−1,Bio dUTP 8 μM in TdT buffer and CoCl2 1 mM). The sections were thenwashed four times in bidistilled apyrogen water (2 min each), twice inphosphate-buffered saline (PBS) (5 min each), in human serumalbumin2% (5 min) and in PBS (5 min) then covered with streptavidin-biotinylated peroxidise complex (Boehringer) diluted 1:100 in a humid-ified chamber for 45 min at room temperature, washed in PBS and
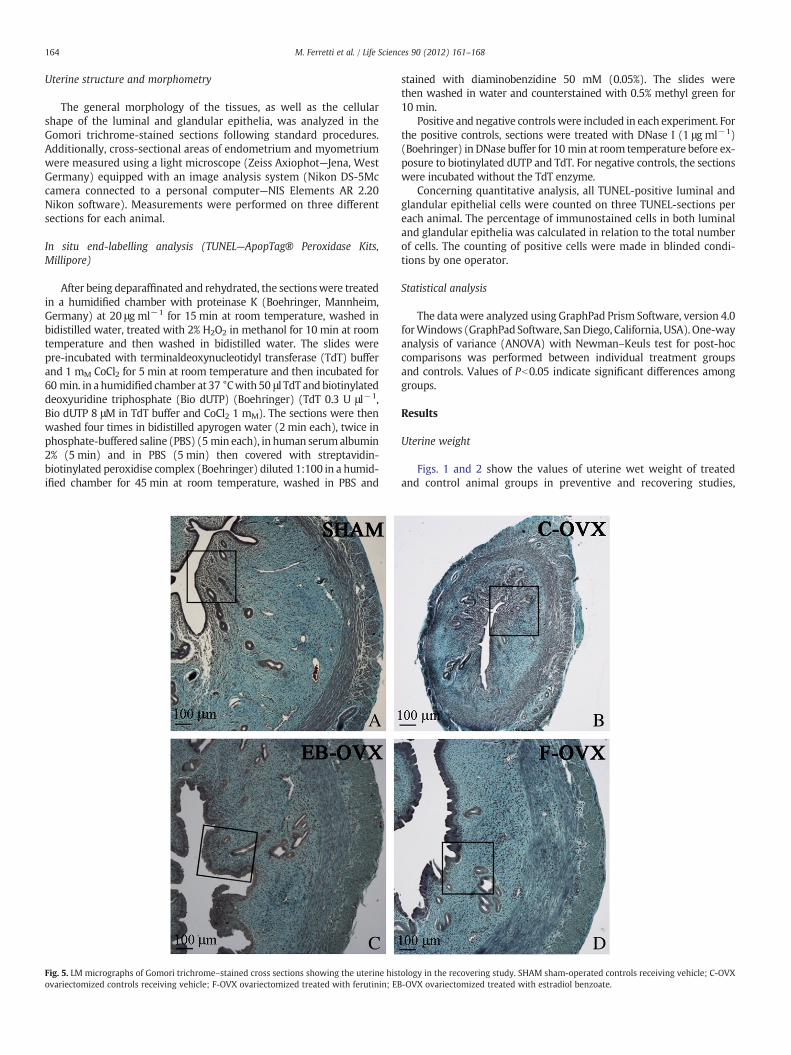
Fig. 5. LM micrographs of Gomori trichrome–stained cross sections showing the uterine histovariectomized controls receiving vehicle; F-OVX ovariectomized treated with ferutinin; EB
stained with diaminobenzidine 50 mM (0.05%). The slides werethen washed in water and counterstained with 0.5% methyl green for10 min.
Positive and negative controlswere included in each experiment. Forthe positive controls, sections were treated with DNase I (1 μg ml−1)(Boehringer) in DNase buffer for 10 min at room temperature before ex-posure to biotinylated dUTP and TdT. For negative controls, the sectionswere incubated without the TdT enzyme.
Concerning quantitative analysis, all TUNEL-positive luminal andglandular epithelial cells were counted on three TUNEL-sections pereach animal. The percentage of immunostained cells in both luminaland glandular epithelia was calculated in relation to the total numberof cells. The counting of positive cells were made in blinded condi-tions by one operator.
Statistical analysis
The data were analyzed using GraphPad Prism Software, version 4.0forWindows (GraphPad Software, SanDiego, California, USA). One-wayanalysis of variance (ANOVA) with Newman–Keuls test for post-hoccomparisons was performed between individual treatment groupsand controls. Values of Pb0.05 indicate significant differences amonggroups.
Results
Uterine weight
Figs. 1 and 2 show the values of uterine wet weight of treatedand control animal groups in preventive and recovering studies,
ology in the recovering study. SHAM sham-operated controls receiving vehicle; C-OVX-OVX ovariectomized treated with estradiol benzoate.

165M. Ferretti et al. / Life Sciences 90 (2012) 161–168
respectively. In both studies the mean values of uterine weight of F-OVX animals were similar to those of EB-OVX and SHAM groupsand significantly higher with respect to C-OVX rats.
Uterine structure
Histological observations of uterine cross sections from treatedand control animals are shown in Figs. 3–6. As expected, in both pre-ventive and recovering studies, the thickness of uterine wall appearsto be clearly thinner in C-OVX rats with respect to SHAM and treated(F-OVX and EB-OVX) animals (Figs. 3 and 5). It emerged that no rel-evant differences exist in uterine size between the two treated groups(F-OVX and EB-OVX) in both preventive and recovering studies, asalso confirmed by the histomorphometric analysis (see below). Asfar as the endometrium structure is concerned, the uterus of SHAMgroups displayed in luminal and glandular epithelia monostratifiedcylindrical cells (Figs. 4A and 6A); in C-OVX groups, instead, the epi-thelia presented a mono-layer of more flattened or cuboidal cellswithout mitotic figures (Figs. 4B and 6B). Concerning the groups trea-ted with ferutinin (F-OVX) and estradiol benzoate (EB-OVX), thestructure of luminal epithelium showed some differences comparedwith SHAM groups: the cell size increased and the outline was irreg-ular; in particular, the cells displayed a columnar pseudostratified-like aspect with randomly located nuclei, vacuolar degeneration andpresence of intracellular cysts; no differences in cell height were in-stead observed in glandular epithelia, with respect to SHAM animals,even if intracellular cysts were present (Figs. 4C, 4D, 6C, 6D).
Fig. 6. LM micrographs of endometria in the recovering study. A, B, C, D: enlargements of theectomized controls receiving vehicle; F-OVX ovariectomized treated with ferutinin; EB-OVXstratified epithelial cells; the arrow heads point to intracellular cysts.
Uterine morphometry
The histomorphometric evaluations of endometrial and myome-trial areas in both preventive and recovering studies are shown inFigs. 7 and 8, respectively; it is to be noted that in both studies themean values were always significantly higher in the treated groups(F-OVX and EB-OVX) with respect to C-OVX groups, as also clearlyobserved in cross sections (Figs. 3 and 5—see above); moreover,only in recovering study the endometrial area was significantlylower in treated rats with respect to SHAM ones.
Uterus TUNEL (apoptotic index)
The numbers of TUNEL-positive cells were evaluated in bothluminal and glandular epithelia (Figs. 9 and 10). In the preventivestudy the percentage of apoptotic cells of luminal epithelium oftreated animals (F-OVX and EB-OVX) showed the highest value; onlyin EB-OVX group it reached statistical significance with respect toboth C-OVX and SHAM groups (Fig. 9A). Concerning the glandular epi-thelium, the percentage of TUNEL-positive cells in F-OVX group was al-ways significantly higher with respect to all other groups (Fig. 9B).In the recovering study, the apoptotic cell percentage in both luminaland glandular epithelia in F-OVX groupwas significantly higherwith re-spect to all other groups (Fig. 10). Regarding apoptosis in glandular ep-ithelia, it is to be noted that statistically significant differences existbetween ferutinin treated animals versus estrogen treated ones; suchdifferences reached the highest significance in the preventive protocol
squared areas in Fig. 5. SHAM sham-operated controls receiving vehicle; C-OVX ovari-ovariectomized treated with estradiol benzoate. The arrows point to columnar pseudo-

SHAM C-OVX EB-OVX F-OVX0
1.0×106
2.0×106
3.0×106
4.0×106
5.0×106
###
******
AE
ndom
etri
al a
rea
( µm
2 )
SHAM C-OVX EB-OVX F-OVX0
1.0×106
2.0×106
3.0×106
4.0×106
5.0×106
***
**
##
B
Myo
met
rial
are
a (
µm2 )
Fig. 7. Mean values of endometrial (A) and myometrial (B) areas in the preventivestudy. Values are expressed as mean±SEM. **Pb0.01, ***Pb0.001 versus C-OVX;##Pb0.01, ###Pb0.001 versus SHAM (ANOVA followed by Newman–Keuls test).SHAM sham-operated controls receiving vehicle; C-OVX ovariectomized controls re-ceiving vehicle; F-OVX ovariectomized treated with ferutinin; EB-OVX ovariectomizedtreated with estradiol benzoate.
SHAM C-OVX EB-OVX F-OVX0
1.0×106
2.0×106
3.0×106
4.0×106
###
******##
#
A
End
omet
rial
are
a (
µm2 )
SHAM C-OVX EB-OVX F-OVX0
1.0×106
2.0×106
3.0×106
4.0×106
###
******
B
Myo
met
rial
are
a (
µm2 )
Fig. 8. Mean values of endometrial (A) and myometrial (B) areas in the recoveringstudy. Values are expressed as mean±SEM. ***Pb0.001 versus C-OVX; #Pb0.05,##Pb0.01, ###Pb0.001 versus SHAM (ANOVA followed by Newman–Keuls test).SHAM sham-operated controls receiving vehicle; C-OVX ovariectomized controls re-ceiving vehicle; F-OVX ovariectomized treated with ferutinin; EB-OVX ovariectomizedtreated with estradiol benzoate.
SHAM C-OVX EB-OVX F-OVX0
1
2
3
4
5
*#
A
TU
NE
L-p
ositi
ve c
ells
(%
)SHAM C-OVX EB-OVX F-OVX
0
1
2
3
4
5 ***###+++
B
TU
NE
L-p
ositi
ve c
ells
(%
)Fig. 9. Mean values of TUNEL-positive cell percentage in luminal (A) and glandular (B)epithelia in the preventive study. Values are expressed as mean±SEM. *Pb0.05,***Pb0.001 versus C-OVX; #Pb0.05, ###Pb0.001 versus SHAM; +++Pb0.001 versusEB-OVX (ANOVA followed by Newman-Keuls test). SHAM sham-operated controls re-ceiving vehicle; C-OVX ovariectomized controls receiving vehicle; F-OVX ovariecto-mized treated with ferutinin; EB-OVX ovariectomized treated with estradiol benzoate.
SHAM C-OVX EB-OVX F-OVX0
1
2
3
*+
A
TU
NE
L-p
ositi
ve c
ells
(%
)
SHAM C-OVX EB-OVX F-OVX0
1
2
3
#*
B +
TU
NE
L-p
ositi
ve c
ells
(%
)
Fig. 10. Mean values of TUNEL-positive cell percentage in luminal (A) and glandular(B) epithelia in the recovering study. Values are expressed as mean±SEM. *Pb0.05versus C-OVX; #Pb0.05 versus SHAM; +Pb0.05 versus EB-OVX (ANOVA followed byNewman–Keuls test). SHAM sham-operated controls receiving vehicle; C-OVX ovariec-tomized controls receiving vehicle; F-OVX ovariectomized treated with ferutinin; EB-OVX ovariectomized treated with estradiol benzoate.
166 M. Ferretti et al. / Life Sciences 90 (2012) 161–168

Fig. 11. LM micrographs of endometrial epithelia of EB-OVX and F-OVX animals in the preventive study. The arrows point to positive cells in luminal epithelia; the arrow headspoint to positive cells in glandular epithelia.
167M. Ferretti et al. / Life Sciences 90 (2012) 161–168
(Figs. 9B, 10B). Fig. 11 shows, as example, the aspect of TUNEL positivecells in luminal and glandular epithelia.
Discussion
Phytoestrogens have long been recognized for their uterotropic ac-tivity in a variety of animal species, often showing dose-dependent ef-fects. In the present study, the effects of chronic treatments withferutinin on the uterus of ovariectomized rats, particularly concerningitsweight, size, morphology and structure, have been evaluated in com-parisonwith those elicited by estradiol benzoate treatment, both in pre-ventive and recovering protocols.
Ferutinin seems to exert the same effect as estrogen benzoate in in-creasing uterine weight not only in preventive study but also in recov-ering study, in which 2 months have elapsed between ovariectomyand the onset of the treatment. This effect is in line with observationsrecorded for genistein that, in a dose-dependent manner, binds to theER; indeed, according to Santell et al. (1997) the ligand/receptor com-plexes induce the expression of estrogen-responsive genes which ulti-mately results in increased uterine mass.
Another observation to be discussed, under the morphologicalviewpoint, is that in both preventive and recovering studies the treat-ments with ferutinin and estradiol benzoate would seem to interferedifferently on the structures of luminal and glandular epithelia. Bothferutinin and estradiol benzoate seem to affect, similarly, the histo-logical features of the luminal epithelium, while they do not seemto modify the morphology of the glandular epithelium. These histo-logical aspects were very different from those observed in C-OVXrats, in which ovariectomy induced atrophy of the endometrium.
Histomorphometric analysis performed on cross sections confirmedthat ferutinin, like estradiol benzoate, stimulates the growth of uterinewall acting on both endometrium and myometrium; in the recoveringstudy only the endometrial areas of the two treated groups were signif-icantly lower with respect to SHAM ones, as a consequence of the timeelapsed between ovariectomy and the onset of the treatment. Our find-ings appear to be in line with a previous report (Rimoldi et al., 2006) onthe morphologic changes induced by oral long-term treatment withother phytoestrogens (i.e. 8 prenylnaringenin—8-PN) in the uterus ofcastrated rats. 8-PNwas shown to stimulate a dose-dependent increasein thickness of uterine endometrium: low8-PNdoses seem to have triv-ial effect on uterine structure, whereas estradiol benzoate and high 8-PN doses induce hyperplasia and hypertrophy in endometrial glands,as well as the increase of myometrial thickness.
The uterus is to be considered an extremely complex organ be-cause it shows enormous morphofunctional variations throughoutthe woman lifetime (as well as in animal estrus) and in particular inreproductive cycle, during which maximal proliferation occurs inthe epithelia especially in the late pregnancy. On the basis of theaforesaid results, the morphological and morphometrical data of the
present study suggest that ferutinin acts on the uterus in a mannersimilar to that of estradiol benzoate, stimulating endometrial hyper-trophy. The treatment with ferutinin is of particular interest becausethe apoptotic index in both preventive and recovering studies is al-most always significantly higher in luminal and glandular epitheliawith respect to groups treated with estradiol benzoate. Since an in-creased risk of endometrial cancer due to excessive hypertrophy isone of the recognized prejudicial effects of estrogen, the phytoestro-gen ferutinin increases the percentage of apoptotic epithelial cells,particularly the glandular ones. This effect might exert a protectiverole against uterine carcinoma. The present data on increased apopto-sis processes are in line with previous observations by other authorsusing another phytoestrogen (i.e. genistein) in ovariectomized mice(García-Pérez et al., 2006). Moreover, Eason et al. (2005) suggestedthat lifetime dietary exposure to soy food does not alter uterine cellphenotype in young adult rats, while isoflavone genistein, by enhanc-ing uterine endometrial glandular apoptosis in vivo and in vitro, mayconfer protection against uterine carcinoma. In any case, under a ge-netic point of view, the possible effects of isoflavones on the humanendometrium are not yet totally clear. Some randomized studies per-formed in women treated with isoflavones have been unable to findproliferative changes (Kaari et al., 2006) or increased marker of pro-liferation (Crisafulli et al., 2004); moreover, biochemical data suggestan antiproliferative effect of isoflavones on human endometrium(Inoue et al., 2005).
Notwithstanding the ferutinin effects shown in the present paper,further investigations should be performed to better establish themechanism by which ferutinin acts on organs normally targeted byestrogens. For example, PCNA (proliferative cell nuclear antigen), aprocessivity factor of DNA synthesis, should be detected as a markerthat reveals proliferating cells; it also should be associated with uter-us apoptotic index, in order to evaluate whether a relationship existsbetween the expression of PCNA and apoptosis in the uterine tissues.
Conclusions
In conclusion, Ferutinin could be an interesting alternative goodnew candidate for HRT in treatment of post-menopausal symptoms,although the putative undesired estrogenic-like side effects on uterusof such phytoestrogen have not yet been fully investigated. It seemsto mime the ovarian endocrine function during menopause protect-ing from bone loss induced by ovariectomy (Palumbo et al., 2009;Ferretti et al., 2010) but reducing the risk of uterine cancer due tothe increase of the endometrial epithelium apoptotic index.
Conflict of interest statement
None.

168 M. Ferretti et al. / Life Sciences 90 (2012) 161–168
Acknowledgements
This study was supported by funds of Fondazione of Vignola 2010and Banca Popolare of Emilia Romagna.
References
Abourashed EA, Galal AM, El-Feraly FS, Khan IA. Separation and quantification of themajor daucane esters of Ferula hermonis by HPLC. Planta Med 2001;67:681–2.
Appendino G, Spagliardi P, Cravotto G, Pocock V, Milligan S. Daucane phytoestrogens: astructure–activity study. J Nat Prod 2002;65:1612–5.
Crisafulli A, Marini H, Bitto A, Altavilla D, Squadrito G, Romeo A, et al. Effects of genis-tein on hot flushes in early postmenopausal women: a randomized, double-blindEPT- and placebo-controlled study. Menopause 2004;11:400–4.
Eason RR, Till SR, Velarde MC, Geng Y, Chatman Jr L, Gu L, et al. Uterine phenotype ofyoung adult rats exposed to dietary soy or genistein during development. J NutrBiochem 2005;16:625–32.
Fanti P, Monier-Faugere MC, Geng Z, Schmidt J, Morris PE, Cohen D, et al. The phytoes-trogen genistein reduces bone loss in short-term ovariectomized rats. OsteoporosisInt 1998;8:274–81.
Ferretti M, Bertoni L, Cavani F, Zavatti M, Resca E, Carnevale G, et al. Influence of ferutininon bonemetabolism in ovariectomized rats. II: role in recovering osteoporosis. J Anat2010;217:48–56.
García-Pérez MA, Noguera R, del Val R, Noguera I, Hermenegildo C, Cano A. Compara-tive effects of estradiol, raloxifene, and genistein on the uterus of ovariectomizedmice. Fertil Steril 2006;86:1003–5.
Ikeda K, Arao Y, Otsuka H, Nomoto S, Horiguchi H, Kato S, et al. Terpenoids found in theUmbelliferae family act as agonists/antagonists for ERα and ERβ: differential tran-scription activity between ferutinine-liganded ERα and ERβ. Biochem Biophys ResCommun 2002;291:354–60.
Inoue A, Takeuchi S, Takahashi S. Insulin-like growth factor-I stimulated DNA replicationin mouse endometrial stromal cells. J Reprod Dev 2005;51:305–13.
Johnson EB, Muto MG, Yanushpolsky EH, Mutter GL. Phytoestrogen supplementationand endometrial cancer. Obstet Gynecol 2001;98:947–50.
Kaari C, Haidar MA, Junior JM, Nunes MG, Quadros LG, Kemp C, et al. Randomized clin-ical trial comparing conjugated equine estrogens and isoflavones in postmeno-pausal women: a pilot study. Maturitas 2006;53:49–58.
Kalu DN. The ovariectomized rat model of postmenopausal bone loss. Bone Miner1991;15:175–91.
Kuiper GG, Lemmen JG, Carlsson B, Corton JC, Safe SH, van der Saag PT, et al. Interactionof estrogenic chemicals and phytoestrogens with estrogen receptor β. Endocrinol-ogy 1998;139:4252–63.
Palumbo C, Ferretti M, Bertoni L, Cavani F, Resca E, Casolari B, et al. Influence of bone me-tabolism in ovariectomized rats. I: role in preventing osteoporosis. J Bone MinerMetab 2009;27:538–45.
Patisaul HB, Melby M, Whitten PL, Young LJ. Genistein affects ERβ- but not ERα-dependent gene expression in the hypothalamus. Endocrinology 2005;143:2189–97.
Rimoldi G, Christoffel J, Wuttke W. Morphologic changes induced by oral long-termtreatment with 8-prenylnaringenin in the uterus, vagina, and mammary gland ofcastrated rats. Menopause 2006;13:669–77.
Santell RC, Chang YC, Nair MG, Helferich WG. Dietary genistein exerts estrogenic ef-fects upon the uterus, mammary gland and the hypothalamic/pituitary axis inrats. J Nutr 1997;127:263–9.
Whitten PL, Patisaul HB. Cross-species and interassay comparisons of phytoestrogenaction. Environ Health Perspect 2001;109:5-20.
Zanoli P, Rivasi M, Zavatti M, Brusiani F, Vezzalini F, Baraldi M. Activity of single com-ponents of Ferula hermonis on male rat sexual behavior. Int J Impot Res 2005;17:513–8.
Zanoli P, Zavatti M, Geminiani E, Corsi L, Baraldi M. The phytoestrogen ferutinin affectsfemale sexual behavior modulating ERα expression in the hypothalamus. BehavBrain Res 2009;199:283–7.
Zavatti M, Montanari C, Zanoli P. Role of ferutinin in the impairment of female sexualfunction induced by Ferula hermonis. Physiol Behav 2006;89:656–61.
Zavatti M, Benelli A, Montanari C, Zanoli P. The phytoestrogen ferutinin improves sex-ual behavior in ovariectomized rats. Phytomedicine 2009;16:547–54.




















