
Characterization of mammary stromal remodeling during the dry period
Upload
independentCategory
view
1download
0
NATURE MEDICINE • VOLUME 7 • NUMBER 12 • DECEMBER 2001 1339
ARTICLES
Myeloid dendritic cells (MDCs, DC1s) and plasmacytoid DCs(also known as plasmacytoid monocytes1 or DC2s (ref. 2)) are thetwo principal subpopulations of human DCs. Adoptive transferof cultured, antigen-charged DC1s induces anti-tumor immu-nity associated with some clinical responses in humans3–5, al-though efficacy has not been firmly established. Tumor orstroma prevent normal DC1 function and differentiationthrough production of soluble factors such as interleukin-6 (IL-6), IL-10, macrophage-colony stimulating factor (M-CSF) andvascular endothelial growth factor (VEGF)6–9, and by retainingmature DC1s locally10. DC1s in tumor-bearing hosts may be defi-cient in accessory-molecule expression, and inefficient in lym-phoid homing6.
Circulating lineage-negative, CD1a–CD4+CD11c–DR+ cells thatexpress CD123, the IL-3 receptor α-chain2,11,12, or which can beidentified by expression of immunoglobulin-like transcripts1,may be preDC2s (refs. 1,2,13). DC2s and preDC2s are poorlycharacterized, as they are rare and difficult to cultivate invitro1,2,11,12. Virus infection affects preDC2 activation, causing se-cretion of high-level type-I interferons (IFNs) that contribute toIFN-γ induction13–15. IL-3 induces preDC2 differentiation, favor-ing IL-4 induction over IFN-γ (2,14). The role of DC2s orpreDC2s in human tumor immunity is unknown as viabletumor-associated cells have not yet been studied.
Inflammatory cytokines and bacterial products stimulate DCmigration to lymph nodes16. However, factors governing the
trafficking of DCs are incompletely understood. Functional stud-ies have so far looked primarily at DC1s (refs. 17–20) and havenot examined the tumor microenvironment. Here we describecells we characterize as tumor-associated preDC2s that were iso-lated from malignant human ascites in patients with ovarian ep-ithelial carcinomas.
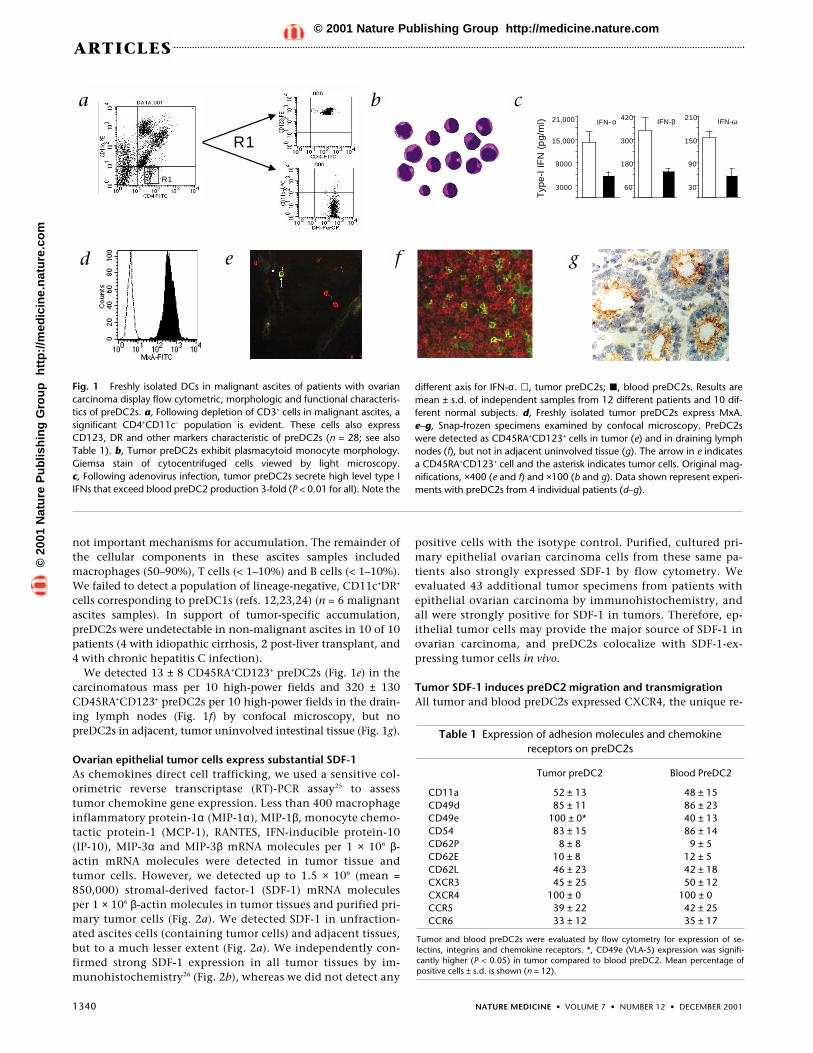
Functional preDC2s accumulate in the tumor microenvironmentWe studied 28 patients with previously untreated ovarian carci-noma and identified a population of CD4+CD11c–CD123+DRhi
cells in tumor ascites (Fig. 1a) that morphologically resembledblood preDC2s (Fig. 1b). These cells secreted high-level type-IIFNs after virus infection (Fig. 1c), confirming their identity asfunctional preDC2s (refs. 1,13,15). Tumor preDC2s also ex-pressed MxA (Fig. 1d), an IFN-α–inducible intracellular proteinwell established as a surrogate marker for local IFN-α produc-tion21,22; this finding was consistent with reports of preDC2s inblood13. PreDC2s comprised 0.83 ± 0.35% (mean ± s.d.) of totalascites cells, which was five-fold higher than in patient blood(0.17 ± 0.08% of total mononuclear cells; P < 0.01). BloodpreDC2s in ovarian cancer patients were also lower than in nor-mal subjects (0.42 ± 0.27%; n = 12; P < 0.01). CD34+ precursorswere undetectable in ascites, and tumor preDC2s did not expressthe Ki-67 antigen (a marker of cell proliferation). These data sug-gest that preDC2s were specifically recruited to the tumor mi-croenvironment, and that local generation or proliferation were
Stromal-derived factor-1 in human tumors recruits and altersthe function of plasmacytoid precursor dendritic cells
WEIPING ZOU1,2, VÉRONIQUE MACHELON3, AURORE COULOMB-L’HERMIN4, JOZEF BORVAK2,FRANÇOISE NOME3, TATYANA ISAEVA2, SHUANG WEI1,2, ROMAN KRZYSIEK5,
INGRID DURAND-GASSELIN5, ALAN GORDON6, TERRI PUSTILNIK7, DAVID T. CURIEL7,PIERRE GALANAUD5, FRÉDÉRIQUE CAPRON4, DOMINIQUE EMILIE5 & TYLER J. CURIEL1
1Tulane Medical School, Section of Hematology and Medical Oncology, New Orleans, Louisiana, USA2Baylor Institute for Immunology Research, Dallas, Texas, USA
3INSERM, U-355, Institut Paris-Sud sur les Cytokines, Clamart, France4Service d’Anatomie Pathologique, Hôpital Antoine Béclère, Clamart, France
5Institut Paris-Sud sur les Cytokines, INSERM U 131, Clamart, France6Baylor University Medical Center, Dallas, Texas, USA
7Wallace Tumor Institute, University of Alabama, Birmingham, Alabama, USACorrespondence should be addressed to T.J.C.; e-mail: [email protected], or W.Z.; email: [email protected]
Dendritic-cell (DC) trafficking and function in tumors is poorly characterized, with studies con-fined to myeloid DCs (DC1s). Tumors inhibit DC1 migration and function, likely hindering spe-cific immunity. The role of plasmacytoid DCs (DC2s) in tumor immunity is unknown. We showhere that malignant human ovarian epithelial tumor cells express very high levels of stromal-de-rived factor-1, which induces DC2 precursor (preDC2) chemotaxis and adhesion/transmigra-tion, upregulates preDC2 very late antigen (VLA)-5, and protects preDC2s from tumormacrophage interleukin-10–induced apoptosis, all through CXC chemokine receptor-4. TheVLA-5 ligand vascular-cell adhesion molecule-1 mediated preDC2 adhesion/transmigration.Tumor preDC2s induced significant T-cell interleukin-10 unrelated to preDC2 differentiation oractivation state, and this contributed to poor T-cell activation. Myeloid precursor DCs (preDC1s)were not detected. Tumors may weaken immunity by attracting preDC2s and protecting themfrom the harsh microenvironment, and by altering preDC1 distribution.
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://m
edic
ine.
nat
ure
.co
m© 2001 Nature Publishing Group http://medicine.nature.com
1340 NATURE MEDICINE • VOLUME 7 • NUMBER 12 • DECEMBER 2001
ARTICLES
not important mechanisms for accumulation. The remainder ofthe cellular components in these ascites samples includedmacrophages (50–90%), T cells (< 1–10%) and B cells (< 1–10%).We failed to detect a population of lineage-negative, CD11c+DR+
cells corresponding to preDC1s (refs. 12,23,24) (n = 6 malignantascites samples). In support of tumor-specific accumulation,preDC2s were undetectable in non-malignant ascites in 10 of 10patients (4 with idiopathic cirrhosis, 2 post-liver transplant, and4 with chronic hepatitis C infection).
We detected 13 ± 8 CD45RA+CD123+ preDC2s (Fig. 1e) in thecarcinomatous mass per 10 high-power fields and 320 ± 130CD45RA+CD123+ preDC2s per 10 high-power fields in the drain-ing lymph nodes (Fig. 1f) by confocal microscopy, but nopreDC2s in adjacent, tumor uninvolved intestinal tissue (Fig. 1g).
Ovarian epithelial tumor cells express substantial SDF-1As chemokines direct cell trafficking, we used a sensitive col-orimetric reverse transcriptase (RT)-PCR assay25 to assesstumor chemokine gene expression. Less than 400 macrophageinflammatory protein-1α (MIP-1α), MIP-1β, monocyte chemo-tactic protein-1 (MCP-1), RANTES, IFN-inducible protein-10(IP-10), MIP-3α and MIP-3β mRNA molecules per 1 × 106 β-actin mRNA molecules were detected in tumor tissue andtumor cells. However, we detected up to 1.5 × 106 (mean =850,000) stromal-derived factor-1 (SDF-1) mRNA moleculesper 1 × 106 β-actin molecules in tumor tissues and purified pri-mary tumor cells (Fig. 2a). We detected SDF-1 in unfraction-ated ascites cells (containing tumor cells) and adjacent tissues,but to a much lesser extent (Fig. 2a). We independently con-firmed strong SDF-1 expression in all tumor tissues by im-munohistochemistry26 (Fig. 2b), whereas we did not detect any
positive cells with the isotype control. Purified, cultured pri-mary epithelial ovarian carcinoma cells from these same pa-tients also strongly expressed SDF-1 by flow cytometry. Weevaluated 43 additional tumor specimens from patients withepithelial ovarian carcinoma by immunohistochemistry, andall were strongly positive for SDF-1 in tumors. Therefore, ep-ithelial tumor cells may provide the major source of SDF-1 inovarian carcinoma, and preDC2s colocalize with SDF-1-ex-pressing tumor cells in vivo.
Tumor SDF-1 induces preDC2 migration and transmigrationAll tumor and blood preDC2s expressed CXCR4, the unique re-
R1
R1
a b
3000
9000
15,000
IFN- α21,000
60
180
300
420IFN-β
30
90
150
210IFN-ω
Typ
e-I I
FN
(p
g/m
l)
c
d e f g
Fig. 1 Freshly isolated DCs in malignant ascites of patients with ovariancarcinoma display flow cytometric, morphologic and functional characteris-tics of preDC2s. a, Following depletion of CD3+ cells in malignant ascites, asignificant CD4+CD11c– population is evident. These cells also expressCD123, DR and other markers characteristic of preDC2s (n = 28; see alsoTable 1). b, Tumor preDC2s exhibit plasmacytoid monocyte morphology.Giemsa stain of cytocentrifuged cells viewed by light microscopy. c, Following adenovirus infection, tumor preDC2s secrete high level type IIFNs that exceed blood preDC2 production 3-fold (P < 0.01 for all). Note the
different axis for IFN-α. �, tumor preDC2s; �, blood preDC2s. Results aremean ± s.d. of independent samples from 12 different patients and 10 dif-ferent normal subjects. d, Freshly isolated tumor preDC2s express MxA. e–g, Snap-frozen specimens examined by confocal microscopy. PreDC2swere detected as CD45RA+CD123+ cells in tumor (e) and in draining lymphnodes (f), but not in adjacent uninvolved tissue (g). The arrow in e indicatesa CD45RA+CD123+ cell and the asterisk indicates tumor cells. Original mag-nifications, ×400 (e and f) and ×100 (b and g). Data shown represent experi-ments with preDC2s from 4 individual patients (d–g).
Table 1 Expression of adhesion molecules and chemokinereceptors on preDC2s
Tumor preDC2 Blood PreDC2
CD11a 52 ± 13 48 ± 15CD49d 85 ± 11 86 ± 23CD49e 100 ± 0* 40 ± 13CD54 83 ± 15 86 ± 14CD62P 8 ± 8 9 ± 5CD62E 10 ± 8 12 ± 5CD62L 46 ± 23 42 ± 18CXCR3 45 ± 25 50 ± 12CXCR4 100 ± 0 100 ± 0CCR5 39 ± 22 42 ± 25CCR6 33 ± 12 35 ± 17
Tumor and blood preDC2s were evaluated by flow cytometry for expression of se-lectins, integrins and chemokine receptors. *, CD49e (VLA-5) expression was signifi-cantly higher (P < 0.05) in tumor compared to blood preDC2. Mean percentage ofpositive cells ± s.d. is shown (n = 12).
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://m
edic
ine.
nat
ure
.co
m© 2001 Nature Publishing Group http://medicine.nature.com

NATURE MEDICINE • VOLUME 7 • NUMBER 12 • DECEMBER 2001 1341
ARTICLES
ceptor for SDF-1 (refs. 27,28), with moderate and equivalent ex-pression of the chemokine receptors CCR2, CCR5, CCR6 andCXCR3 (Table 1). As ligands for chemokine receptors other thanCXCR4 were undetectable or poorly expressed in tumors, we fur-ther studied SDF-1. Tumor-cell culture supernatants, malignantascites fluid or recombinant SDF-1 each induced significantpreDC2 migration (Fig. 2c) and adhesion/transmigration (Fig.2d), confirming that CXCR4 is functional and that tumor-de-rived SDF-1 induces preDC2 migration and adhesion/transmi-gration. Migration and adhesion/transmigration weresignificantly decreased by incubating preDC2s with anti-CXCR4antibody (Fig. 2c and d), suggesting the presence of SDF-1 intumor culture supernatants and malignant ascites. Non-malig-nant ascites fluid from patients with hepatitis C did not inducepreDC2 migration (data not shown).
The preDC2 chemokine receptors CXCR3 and CCR5 mediatedchemotaxis in response to recombinant forms of their respectiveligands IP-10 and MIP-1α (ref. 27) (Fig. 2c, d and data notshown). However, blockade of CCR5 or CXCR3 with specific an-tibodies affected ascites-fluid–induced preDC2 migration to amuch lesser extent (Fig. 2c, d and data not shown), suggesting alimited role for these receptors in tumor preDC2 migration inmalignant ascites.
Tumor preDC2s express ligands binding tumor endotheliumTumor and blood preDC2s expressed comparable levels of theintegrins CD11a, CD49d and CD54, and the selectin CD62L,
whereas selectins CD62P and CD62E were weakly ex-pressed by preDC2s from either source (Table 1).Significantly more tumor preDC2s expressed CD49ecompared with blood preDC2s (100% versus 40 ± 13%; P< 0.05). P-selectin glycoprotein ligand (PSGL)-1 is a lig-and for selectins, and vascular cell adhesion molecule(VCAM)-1 is the ligand for CD49. Both VCAM-1 andPSGL-1 were expressed on tumor vascular endothelialcells (Fig. 2e and not shown), suggesting that eitherCD49e (VLA-5) or CD62L could participate in recruit-
ment of preDC2s into the tumor microenvironment.
SDF-1 upregulates VLA-5 expression on preDC2sBlood preDC2s incubated in tumor ascites fluid upregulatedVLA-5 expression by approximately 2-fold (Fig. 2f), to levelscomparable to tumor preDC2s (Table 1). Blocking CXCR4 withspecific antibody significantly reduced this effect (Fig. 2f).Recombinant SDF-1 also upregulated blood preDC2 VLA-5 ex-pression, but less potently than tumor ascites fluid. Neither as-cites nor recombinant SDF-1 mediated a significant effect ontumor preDC2 VLA-5 expression as it was likely already maxi-mal, or CD62L and CD49d expression on tumor or bloodpreDC2s (data not shown).
Tumor preDC2 transmigration depends on VLA-5 and VCAM-1To define the roles of VLA-5 and CD62L in preDC2 tumor traf-ficking, we incubated human umbilical vein endothelial cellswith antibodies against VCAM-1 or PSGL-1. Anti–VCAM-1 sig-nificantly inhibited preDC2 transmigration across endothelialcells (Fig. 2d), suggesting that VLA-5 was important for preDC2tumor trafficking into tumor tissue. Although blockade ofCD62L did not reduce preDC2 adhesion/transmigration in thisstatic assay (Fig. 2d), its role in preDC2 recruitment under dy-namic conditions cannot be excluded.
SDF-1 inhibits IL-10–mediated preDC2 apoptosisTumor macrophages produce IL-10, which induces preDC2
a b
Medium
Tumor culture supernatant + anti-CXC R4
Ascites fluid + anti-CXC R4
Tumor culture supernatant
Ascites fluid
Migrated cells (x 103)
SDF-1
IP-10
0 2 4 6 8
Tumor culture supernatant + anti-CXCR3
***
c
Medium
Ascites fluid + anti-CXC R4
Ascites fluid
Transmigrated cells (%
SDF-1
IP-10
Ascites fluid + anti-CXCR3
Ascites fluid+ anti-VCAM-1
Ascites fluid+ anti-PSGL-1
0 25 50 75
**
d
e
0
40
80
120
VLA
-5 e
xpre
ssin
g ce
lls (
% )
f
Fig. 2 Ovarian epithelial carcinoma cells express functionalSDF-1. a, SDF-1 mRNA was detected in tumor and ascites cells,but not isolated macrophages by RT-PCR (n = 12). Lanes: 1,tumor tissue; 2, tumor macrophages; 3, adjacent tissues; 4, totalascites cells; 5, purified tumor cells; 6, DNA ladder. b, High-levelSDF-1 was detected in the tumor mass of every patient studied (n= 43) as demonstrated with anti-SDF-1 antibody. Magnification,×100. c, SDF-1 in malignant ascites and ovarian carcinoma cellsinduces preDC2 chemotaxis. *, P < 0.001 compared withmedium. Blockade of preDC2 CXCR4 with a specific antibodyagainst CXCR4 (100 ng/ml) significantly inhibited preDC2 migra-tion. Results are the mean ± s.d. of 5 independent experimentsusing samples from 5 separate patients. d, SDF-1 in malignant as-cites and ovarian carcinoma cells induces preDC2 adhesion/trans-migration. *, P < 0.001 compared to medium. Adhesion/transmigration was blocked by anti-CXCR4 (100 ng/ml) or anti-VCAM-1 (100 ng/ml) but not anti–PSGL-1 (500 ng/ml) antibody.Results are the mean ± s.d. of 8 independent experiments using 8separate ascites samples and 5 separate tumor supernatants. e,VCAM-1 is highly expressed on tumor endothelial-cell surfaces byimmunohistochemistry (red; n = 4). Magnification, ×100. f,Ascites (�) induces upregulation of the VCAM-1 ligand VLA-5(CD49e) on blood preDC2, which is blocked by antibody againstCXCR4 (�). Control preDC2s (�) maintained in culture an equiv-alent length of time without ascites did not significantly changebaseline VLA-5 expression. 1 representative experiment of 3.
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://m
edic
ine.
nat
ure
.co
m© 2001 Nature Publishing Group http://medicine.nature.com

1342 NATURE MEDICINE • VOLUME 7 • NUMBER 12 • DECEMBER 2001
ARTICLES
apoptosis2,12. We investigated how tumor preDC2s survived inan environment where most cells were macrophages. First, weconfirmed that these tumor macrophages spontaneously pro-duced IL-10 (>10 pg/ml/105 cells/24 h). Thus, we hypothesizedthat a tumor environmental factor(s) protected against IL-10–mediated preDC2 apoptosis. To address this hypothesis, weexposed preDC2s to tumor macrophages in transwells, inducingpreDC2 apoptosis, which was reversed with neutralizing anti-body against IL-10 (Table 2). Remarkably, tumor cells in tran-swells also protected preDC2s against apoptosis (Table 2).Blocking CXCR4 with specific antibodies reversed tumor-medi-ated protection from apoptosis, suggesting that protection wasmediated by a tumor SDF-1/preDC2–CXCR4 interaction. In con-firmation, recombinant SDF-1 protected preDC2s from IL-10–in-duced apoptosis (Table 2).
Tumor preDC2s induced poorly proliferating T cellsHaving determined mechanisms for tumor preDC2 ingress andtheir survival in the harsh tumor microenvironment, we focusedon preDC2 function. Freshly isolated tumor and blood preDC2sboth induced similar IFN-γ and IL-4 expression in allogeneic,naive CD4+CD45RA+ T cells (Fig. 3a), consistent with the previ-ously reported Th0-polarizing capacity of blood preDC2s (ref. 1).T-cell cytokine secretions induced by both tumor and bloodpreDC2s were confirmed to be similar by ELISA, but with a ten-dency for tumor preDC2s to elicit slightly less IFN-γ (Fig. 4a). Blood DC2s differentiated from preDC2s with IL-3 reportedly differ from preDC2s in IFN-γ and IL-4 induction13.Tumor and blood preDC2s elicited similar T-cell IFN-γ and IL-4following culture in IL-3 (Figs. 3b and 4a). Tumor preDC2s, how-ever, were significantly less effective in inducing T-cell IL-2 com-pared with blood preDC2s (data not shown), even followingculture in IL-3 (Fig. 3b). Deficient IL-2 induction suggested thattumor preDC2s (and their differentiated progeny) would be de-fective in activating T-cell proliferation. In confirmation, allo-geneic T-cell proliferation induced by tumor preDC2s wassignificantly less efficient than that effected by blood preDC2s,either when freshly isolated (P < 0.05) or after culture in IL-3 (P <
0.01), although IL-3 culture improved the T-cell stimulating ca-pacity of both (Fig. 3c).
We hypothesized that poor T-cell proliferation induced bytumor preDC2s could be due to induced IL-10 production. Totest this, we cocultured freshly isolated tumor and bloodpreDC2s with naive, allogeneic T cells. Blood preDC2s fromboth normal and tumor-bearing patients elicited low and com-parable quantities of T-cell IL-10 (18 ± 6 pg/ml each; n = 6).However, tumor preDC2s elicited significantly more IL-10 asshown by ELISA (P = 0.006; Fig. 4a). Differences in preDC2 dif-ferentiation or activation state can lead to differential IL-10 in-duction13. Thus, we studied IL-10 induction after freshly isolatedpreDC2s were cultured with IL-3, or activated by infection withtwo different adenoviruses (≥ 95% CD83+ cells, data not shown).Tumor preDC2s elicited significantly more IL-10 compared withblood preDC2s after differentiation with IL-3 or activation withadenovirus, as determined by flow cytometry (Fig. 4b) and con-firmed by ELISA (P < 0.05 for all comparisons; Fig. 4a). Thesedata suggest that tumor preDC2 were not fully activated or dif-ferentiated. Freshly isolated or IL-3–differentiated preDC2s in-duced less IFN-γ compared with their blood counterparts, withstatistically significant results (P < 0.05) for preDC2 activatedwith adenovirus alone, or adenovirus expressing an antibody de-livering a CD40 agonist signal29.
IL-10 receptor blockade inhibited tumor preDC2-induced T-cell IL-10 (Fig. 4c) and partially restored defective T-cell prolifer-ation induced by tumor preDC2 (Fig. 4d). Type I IFNs seem tohave no role in IL-10 induction, but contributed significantly toIFN-γ induction (Fig. 4c) as observed in blood preDC2s followinginfluenza virus infection and CD40L-induced maturation13,14.
Finally, we determined whether tumor DC2 could inhibittumor-specific T-cell proliferation. To induce proliferation,CD14+ cells were differentiated into DCs with granulocyte-macrophage colony-stimulating factor (GM-CSF) plus IL-4, andincubated with apoptotic bodies from autologous primarytumor cells as a source of tumor antigen. Tumor ascites T cellsproliferated significantly in response to these antigen-loadedDCs (Fig. 4e). T cells previously incubated with tumor DC2s sig-
50% 46%
43%
Tumor preDC2 Blood preDC2
48%
a
Tumor IL-3-DC2 Blood IL-3-DC2
0.3% 3.3%
52%
0.5% 8.1%
45%
5.1% 3.5%
16%
1.8% 1.8%
13%
b
0
10
20
30
40
50
3000 9000[3H
]th
ymid
ine
inco
rpor
atio
n (c
.p.m
. X
10
Number of DCs per well
3)c
Fig. 3 Tumor and blood preDC2s activate T cells in distinct mannerswhen freshly isolated and following differentiation with IL-3. a, Equivalentinduction of IL-4 and IFN-γ from naive, allogeneic CD4+CD45RA+ T cellswith freshly isolated tumor or blood preDC2s. IL-4 production was con-firmed by ELISA (41 ± 30 versus 46 ± 17 pg/ml, respectively for tumor ver-sus blood preDC2). b, Tumor and blood preDC2s differentiated in IL-3
still effected similar T-cell IFN-γ and IL-4 production, but tumor preDC2swere significantly less able to induce T-cell IL-2 (n = 5; P = 0.04). c, PreDC2s from tumor (�) and blood (�) differ in their capacity to in-duce T-cell proliferation when freshly isolated (P < 0.05), or following IL-3–induced differentiation (P < 0.01) in blood (×) or tumor (�). 1representative experiment of 3–5 for each panel.
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://m
edic
ine.
nat
ure
.co
m© 2001 Nature Publishing Group http://medicine.nature.com

NATURE MEDICINE • VOLUME 7 • NUMBER 12 • DECEMBER 2001 1343
ARTICLES
nificantly inhibited this tumor-specific T-cell proliferationacross a transwell membrane, suggesting a soluble mediator.Antibody against IL-10R significantly recovered T-cell prolifera-tion (Fig. 4e). These data suggest that tumor DC2s inhibittumor-specific immunity, mediated in part through induced T-cell IL-10.
DiscussionThe two known principal subpopulations of human DCs, DC1sand DC2s, may influence immunity distinctly2,16. DC1s augment
anti-tumor immunity in experimental animals30 and humans3,5.PreDC2s may primarily mediate anti-pathogen immunity14.PreDC2s and DC2s have been observed in histologic sections oflymphoid neoplasms and granulomas1, but functional tumor-as-sociated counterparts have not been recovered for studies.Malignant ascites contains tumor-specific immune cells, tumor-derived antigens and soluble factors, and viable tumor cells31,32,providing a relevant model for the tumor microenvironment.Thus, to investigate the role of DC2 in tumor immunity we stud-ied cell populations in malignant ascites of patients with epithe-lial ovarian carcinomas with no specific prior treatments.
We identified cells in malignant ascites lacking lineage mark-ers, expressing CD4, CD45RA, CD80, CD86, CD123, and HLA-DR, and morphologically resembling blood preDC2s (refs.1,11,15). These freshly isolated cells elicited T-cell IFN-γ and IL-4and secreted high-level type I IFNs following adenovirus infec-tion, identifying them as functional preDC2, (refs. 2,14,15).Blood preDC2 concentrations that we observed in normal sub-jects were consistent with earlier reports33, but 2.5-fold higherthan in blood of ovarian cancer patients. The preDC2 concen-tration was five-fold higher in ascites compared with patientblood, consistent with active recruitment. We also detectedpreDC2 in the tumor mass and associated lymph nodes. Thus,preDC2 or their progeny accumulate in the tumor microenvi-ronment of epithelial ovarian carcinomas.
Distinct chemokine receptors and adhesion molecules directDC trafficking17,20,34,35, although the process is not well under-stood. Moreover, these studies all looked at DC1 (refs. 17,20)and did not examine the tumor microenvironment. CXCR3and CD62L have been proposed as mediators of preDC2 lym-
0 100 200 300 4000 200 400 600 800 1000
IFN-γ (pg/ml)
PreDC2
IL-3-DC2
Ad-DC2
Ad/CD40-DC2
IL-10 (pg/ml)
500
PMA/Ionomycin
**
**
**
a
Tumor Ad-DC2 Blood Ad-DC2
27% 13% 9.2% 10%
17% 34%
b
IL-10 (pg/ml)IFN- γ (pg/ml)
0 100 200 300 4000 250 500 750 1000 1250 1500
**
*
PMA + lonomycin
Control Ab
Anti-IL-10R
Anti-IFNαRβ
c
[3H ]thymidine
incorporation
(c.p.m . x 10 )
0 20 40 60
*
3
[3H ]thymidine incorporation
(c.p.m. x 10 3)
0
10 20 30 40 50 60
* * *
d e
Fig. 4 Tumor preDC2s induce significant T-cell IL-10 induction indepen-dent of differentiation or activation status. a and b, ELISA (a) showing cy-tokine production from allogeneic, naive T cells was compared using freshlyisolated preDC2s from blood (�) or tumor (�), or the same cells followingculture for 4 d in IL-3 or 4 d after infection with adenovirus (Ad) or aden-ovirus expressing a CD40-agonist antibody29 (Ad/CD40). *, P < 0.05. b, Intracellular cytokine production (flow cytometry). c, Regulation of T cellIL-10 and IFN-γ induction. Allogeneic, naive T cells were induced to secretecytokines using preDC2s activated with Ad/CD40. Blockade of the IL-10 re-ceptor (anti–IL-10R) significantly recovered IFN-γ secretion and inhibited Tcell IL-10 secretion, whereas blockade of the type I IFN receptor (anti-
IFNαRβ) significantly reduced IFN-γ secretion and had no effect on IL-10 se-cretion. d, Blockade of the IL-10R (�) significantly recovered T-cell prolifer-ation induced by Ad/CD40-activated tumor preDC2s versus control (�).Data shown represent experiments using preDC2s from 4 individual pa-tients. Error bars indicate s.d. *, P < 0.05 e, Tumor DC2s induce T cells to se-crete IL-10 that suppresses tumor-specific immunity. DCs derived frompatient CD14+ cells were loaded with autologous, apoptotic tumor bodiesand used to induce proliferation of autologous tumor ascites T cells (�).Allogeneic T cells were incubated separately with tumor DC2s and added tothe upper wells of the transwell (�). Blockade of the IL-10R (�) significantlyrecovered T-cell proliferation. �, control. *, P < 0.01; **, P < 0.05.
Table 2 SDF-1 protects preDC2 from IL-10–induced apoptosis
PercentCell ratio apoptotic cells
preDC2 25 ± 11preDC2/MΦ 1/1 28 ± 9preDC2/MΦ 1/10 52 ± 15preDC2/MΦ 1/100 80 ± 13 preDC2/MΦ 1/100 32 ± 10
+ anti–IL-10preDC2/MΦ/Oc 1/100/10 30 ± 10preDC2/MΦ/Oc 1/100/10 75 ± 18
+ anti-CXCR4preDC2/MΦ 1/100 35 ± 8
+ SDF-1
Incubation of tumor preDC2 with IL-10–secreting tumor macrophages (MΦ) inducedsignificant (P < 0.01) apoptosis, which was blocked by neutralizing IL-10. RecombinantSDF-1, or tumor (Oc) in transwells mediated significant protection against apoptosis (P< 0.01), which was abrogated by blocking the CXCR4 receptor. Mean ± s.d. of 3 inde-pendent experiments using materials from 3 separate subjects is shown.
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://m
edic
ine.
nat
ure
.co
m© 2001 Nature Publishing Group http://medicine.nature.com

1344 NATURE MEDICINE • VOLUME 7 • NUMBER 12 • DECEMBER 2001
ARTICLES
phoid trafficking based on immunohistochemical and cytomet-ric studies, but not functional studies1,21. To identify moleculesresponsible for preDC2 recruitment into the tumor microenvi-ronment, we examined expression and function of adhesionmolecules and chemokine receptors on tumor preDC2s, and ex-pression and function of their ligands on tumor and associatedvascular endothelium.
PreDC2s expressed functional CXCR3, but we did not identifycorresponding ligands in tumor. This receptor did not mediatesignificant migration of preDC2s by ascites or tumor. Thus,whereas CXCR3 may mediate preDC2 recruitment into lymphnodes, different factors appear to mediate preDC2 migrationinto tumor. Both tumor and blood preDC2s expressed CD62L,and vascular endothelial cells from ovarian cancer constitutivelyexpressed its ligand, PSGL-1. Nonetheless, functional experi-ments did not confirm that CD62L was important in preDC2tumor trafficking in this static model. However, as CD62L is im-portant in adhesion under flow conditions36, we cannot excludeits role in preDC2 trafficking into the tumor microenvironment.
We showed through several lines of work that VLA-5 (CD49e)was important in preDC2 migration into the tumor microenvi-ronment. Tumor-derived SDF-1 upregulated preDC2 VLA-5 ex-pression, helping mediate adhesion/transmigration on vascularendothelial cells through its ligand, VCAM-1.Immunohistochemical analysis confirmed that tumor endothe-lium expressed VCAM-1. SDF-1 augmented VLA-5 expression inan effect similar to that on CD34+CXCR4+ hematopoietic precur-sor cells37. There are no prior reports in this regard for DCs.
Blockade of preDC2 CXCR4 inhibited ascites or tumor-in-duced chemotaxis and adhesion/transmigration. Analysis of 43consecutive ovarian epithelial carcinomas confirmed uniform,high-level tumor SDF-1 expression in all. Thus, CXCR4–SDF-1interactions are important for preDC2 migration into the tumormicroenvironment. Notably, tumor SDF-1 also protectedpreDC2 from IL-10–induced apoptosis, in contrast to reportssuggesting that tumors inhibit DC1 differentiation6–9. Type IIFNs are autocrine preDC2 survival factors14. However, our datademonstrated that type I IFNs did not mediate preDC2 survivalinduced by SDF-1.
We did not detect a significant population of lineage-negative,CD11c+DR+ DC1 precursor cells in malignant ascites, nor lin-eage-negative CD1a+CD11c+CD14–DR+ DC1s. Thus, tumor fac-tors may not only attract and protect preDC2s, but may alsohinder DC1 trafficking or differentiation.
In vitro and flow cytometric studies demonstrate that thetumor microenvironment influences DC differentiation andfunction6–9,38,39. Thus, having established a mechanism forpreDC2 ingress, we next studied the function of tumor-associ-ated preDC2s. Although blood preDC2s induced IL-10 express-ing T cells, freshly isolated tumor preDC2s elicited significantlymore T-cell IL-10. Further, sufficient IL-10 was elicited by tumorpreDC2s to inhibit T-cell proliferation and IFN-γ production,which was not the case for the IL-10 induced by blood preDC2s.Virus activation13 and preDC2 differentiation13 alter induced IL-10. However, even after adenovirus activation or IL-3–mediateddifferentiation, tumor preDC2s induced significantly more IL-10compared with blood preDC2s, which inhibited tumor-specificT-cell proliferation. Thus, differences in IL-10 induction do notappear to relate to the activation or differentiation state oftumor compared with blood preDC2s. Moreover, these data con-firm that freshly isolated tumor preDC2s are not fully differenti-ated or activated.
PreDC2s secrete biologically active type I IFNs following spe-cific stimuli1,14,15, which may mediate IFN-γ induction13,14, en-hance survival of activated T-cells40 or hinder DC1development41. We detected little type I IFN in ascites althoughtumor preDC2s expressed MxA, as described for blood preDC2s(ref. 13), suggesting type I IFN production22. Type I IFNs pro-duced by both tumor and blood preDC2s induced significant T-cell IFN-γ secretion in vitro, and did not contribute to IL-10induction. Contributions of endogenous type I IFNs to anti-tumor immunity merit further investigation.
Tumor DC2s express low levels of T-cell costimulatory mole-cules, are poor in activating naive T cells and induce IL-10 se-creting T cells: all characteristics of anergizing or tolerizingcells42. Whether tumor DC2s induce anergy or tolerance requiresfurther investigation. The tumor-derived SDF-1 recruitspreDC2s, and protects them from harmful effects of the mi-croenvironment. We speculate that tumor-derived SDF-1 couldalso enhance the survival and function of detrimental T cells bydelivering proliferative or survival signals28,43,44.
In summary, ovarian epithelial tumors produce high-level SDF-1. This SDF-1 is a chemoattractant for preDC2s, and inducestheir migration into the tumor microenvironment in conjunc-tion with VLA-5. SDF-1 protects tumor preDC2s from IL-10–in-duced apoptosis mediated by local macrophages. These tumorpreDC2s do not appear to mediate effective anti-tumor immu-nity, and appear detrimental to such immunity through IL-10induction. Moreover, neither DC1s nor their precursors were de-tected in the tumor microenvironment. Thus, ovarian epithelialtumors may dysregulate immunity by attracting and protectingpreDC2s, and by hindering accumulation or differentiation ofpreDC1s. Interference with trafficking of specific DC subsetsthrough distinct signaling pathways may be a common theme intumor immunopathogenesis. If CXCR4–SDF-1 interactions in-duce ingress of counterproductive cells such as preDC2s, agentsthat block CXCR4 may be useful therapeutically in addition totheir potential applications as anti-metastatic agents44.
MethodsHuman subjects. Subjects gave written, informed consent. The study wasapproved by the local Institutional Review Board. No cancer patients re-ceived prior specific treatments.
Ascites, ascites cells and tissue biopsies. We collected ascites aseptically,and harvested cells by centrifugation over a Ficoll–Hypaque density gradi-ent (Amersham, Uppsala, Sweden), and cryopreserved them at –86° C untiluse. CD3-, CD14-, CD16-, CD19- and CD56-expressing cells were depletedusing paramagnetic beads (Miltenyi, Auburn, California), and preDC2swere obtained by flow cytometry gating on CD4+CD11c– cells. Cell popula-tions were ≥ 99% pure by flow cytometry. Tissue biopsies were snap-frozenin OCT in liquid nitrogen and kept at –80° C until use. Tumor epithelial cellswere obtained by depleting CD3-, CD14-, CD16-, CD19- and CD56-ex-pressing cells using microbeads (Miltenyi), and sorting CD45– cells by flowcytometry, and maintained in medium with no growth factors. Cellular pu-rity was ≥ 98% as determined by FACS staining with antibody against HER-2/neu.
Peripheral blood cells. Peripheral blood was collected from normal volun-teers aseptically by venipuncture or by cytopheresis, and mononuclear cellswere then obtained by Ficoll–Hypaque density centrifugation. BloodpreDC2s were obtained by flow cytometry after depleting CD3-, CD14-,CD16-, CD19- and CD56-expressing cells as described for ascites cells. Cellswere ≥ 99% pure as determined by flow cytometry.
Reagents. Cytokines and chemokines were recombinant, human proteinspurchased from R&D Systems, Minneapolis, MN, except IP-10 and MIG
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://m
edic
ine.
nat
ure
.co
m© 2001 Nature Publishing Group http://medicine.nature.com

NATURE MEDICINE • VOLUME 7 • NUMBER 12 • DECEMBER 2001 1345
ARTICLES
(Biosource, Camarillo, California). Antibodies for flow cytometry: antibod-ies against (CD3, CD4, CD8, CD11c, CD16, CD19, CD45RA, CD54, CD56,CD62P, CD62E, CD62L, CD80, CD123, CXCR3, CXCR4, CCR5, CCR6,HLA-DR, Ki-67, HER-2/neu and goat anti-mouse immunoglobulin) werepurchased from BD PharMingen (San Jose, California); antibody againstCD14 was from Caltag (Burlingame, California). Antibodies for neutralizingexperiments were from BD PharMingen (anti-CXCR4, clone 12G5, IgG2a;anti-CXCR3, clone 1C6, IgG1; anti-CCR5, clone 2D7, IgG2a; anti-CCR6,clone 11A9, IgG1; anti-VCAM-1, clone 51-10C9, IgG1; anti-PSGL-1, cloneKPL-1, IgG1), from R&D (Minneapolis, Minnesota) (anti-IL-10, clone23738.111, IgG2b; anti-IL-10 receptor, clone 37607.11, IgG1), and fromPBL (New Brunswick, New Jersey) (anti–IFN-α receptor β-chain, cloneMMHAR-2, IgG2a).
Chemotaxis assay. PreDC2s (1 × 105) were transferred into the upperchambers of 5-µm pore-size transwell plates (Neuro Probe, Gaithersburg,Maryland). Chemokines (SDF-1, IP-10, MIG, MIP-1α, MIP-3α and MIP-3β)(100 ng/ml), cell-free ascites or the supernatants from primary ovarian can-cer epithelial cell culture were added to the lower chamber. After 1 h at 37 °C, migration was quantified by counting cells in the lower chamberand cells adhering to the bottom of the membrane. PreDC2 identity wasconfirmed by flow cytometry for CD123 expression. Efficiency was deter-mined by dividing recovered cells by input cells.
Adhesion/transmigration. 24-well plates with 8-µm pore-size polycarbon-ate membranes (Costar, Cambridge, Massachusetts) were coated with fi-bronectin (20 µg/ml). CRL-2480 cells (4 × 105) (ATCC, Manassas, Virginia)were seeded on the inserts 24 h previously. PreDC2s (1 × 105) were addedto the monolayer directly, or 4 h after incubation of the monolayer with an-tibodies. After 20 h, adhering/transmigrating preDC2s were counted as forchemotaxis. Identity of migrating cells was determined by flow cytometryfor CD4-FITC, CD123-PE, HLA-DR PerCP and CD11c-APC.
Immunohistochemistry and confocal analysis. Immunohistochemistrywas performed on 8-µm cryosections of acetone-fixed tissues as described26.Tumor tissues were incubated for 2 h at room temperature with anti–SDF-1antibody26 (clone K15C, IgG2a, 10 µg/ml), anti–PSGL-1 antibody (10 µg/ml)or anti–VCAM-1 antibody (10 µg/ml). Antibody binding was detected withbiotinylated anti-mouse antibodies and streptavidin conjugated to alkalinephosphatase (Biogenex, San Ramon, California) using fast red substrate(Sigma). Sections were counterstained with Mayler hematoxylin. PreDC2s intumors and lymph nodes were detected by staining with paired fluorescentantibodies as described21. Tissue sections were incubated with anti–SDF-1 oranti–CD45RA antibody (clone HI100, IgG2b, 0.8 µg/ml, BD PharMingen)for 1 h at room temperature, followed by Texas Red-conjugated donkeyanti-mouse IgG (1/400 dilution, Jackson Immunoresearch, West Grove,Pennsylvania) for 40 min, and saturated with 10% mouse serum, then incu-bated with biotin-labeled anti-CD123 antibody (clone 9F5, IgG1, 2 µg/ml,PharMingen) and revealed with FITC-streptavidin (1/50 dilution, Caltag,Burlingame, California). In all staining experiments, negative controls wereobtained both by omission of the primary antibodies and by incubation withirrelevant and concentration-matched isotype antibodies. Cells were blindlyexamined by one pathologist (A.C.H.) and confirmed by another investiga-tor (W.Z.) at ×400 magnification using a fluorescence microscope (OlympusBmax 60, Tokyo Japan). The number of positive cells was expressed as themean ± s.d. in 10 different ×400 fields. Confocal microscopy was performedusing a Leica TCS-NT SP equipped with argon, krypton and helium/neonlasers. A spectrophotometer was used to separate the detection channels ofFITC (510–550 nm) and Texas Red (580–660 nm). RT-PCR was performed asdescribed25.
Flow cytometry. Cells were stained with commercial murine monoclonalantibodies and analyzed on a FACSCalibur (Becton–Dickinson, MountainView, California). Intracellular cytokine staining was performed using acommercial kit (BD PharMingen) according to the manufacturer’s direc-tions. PreDC2 and naive T cells were cocultured for 5 d, followed by activa-tion with 25 ng/ml phorbol myristate acetate and 250 ng/ml ionomycin for4 h, followed by monensin treatment for an additional 2 h. Data was ana-lyzed with CellQuest software (Becton–Dickinson). At least 5,000 gatedevents per condition were analyzed.
PreDC2 apoptosis. 12-mm diameter transwells with 0.4-µm pore-size wereused (Costar). 25,000 to 50,000 preDC2 were added to the upper cham-ber. Tumor macrophages with or without tumor epithelial cells were addedto the lower chamber. PreDC2 were maintained in the presence of 10ng/ml IL-3 to support viability. Antibodies against IL-10 (500 ng/ml),CXCR4 (100 ng/ml) or human recombinant SDF-1 (100 ng/ml) was addedas indicated. PreDC2s were recovered from transwells for further study afterincubation at 37 °C in a 5% CO2, humidified atmosphere for 40 h. PreDC2viability and apoptosis were determined by Trypan Blue exclusion, andstaining with 7-AAD plus annexin V antibody (flow cytometry).
Allogeneic mixed lymphocyte reaction. Naive, allogeneic naive T lym-phocytes were purified from peripheral blood using paramagnetic beads(Miltenyi) and cultured with preDC2s as described2. Briefly, the indicatednumber of preDC2s was added to 5 × 105 naive T cells. Proliferation was as-sessed in triplicate by [3H]methylthymidine (NEN) incorporation after 5 d.Antibodies were specific for IL-10 receptor (500 ng/ml) or type I IFN recep-tor β2 chain (1 µg/ml), and added to culture at the outset. Where indicated,preDC2s were differentiated by culture in 10 ng/ml IL-3 (R&D Systems) for4 d, or activated by infection with replication defective adenovirus or thesame adenovirus physically coupled to an agonist anti-CD40 antibody29 at amultiplicity of infection of 100 for 4 d before addition to T cells.
T-cell suppression assay. DC were differentiated from CD14+ tumor ascitescells with GM-CSF plus IL-4 as described46,47. Primary ovarian cancer celllines were induced to become apoptotic by culture in 40 µM beauvericin(Sigma), confirmed by flow cytometry for 7-AAD and annexin V expression.These DCs were cultured for 1 h at 37 °C with tumor bodies, sorted on aCD1a gate (purity > 95%) as described46, and graded numbers plated in thelower chamber of a 12-well transwell plate with purified, autologous tumorCD3+ T lymphocytes (5 × 105/ml). Allogeneic, naive T cells were incubatedwith tumor DC2 for 5 d, and then added to the upper transwell chamber (5× 105/ml). Tumor-specific T-cell proliferation was detected in the lowerchamber as described for the allogeneic mixed lymphocyte reaction.
ELISA. T-cell cytokines (IL-4, IL-10 and IFN-γ) in supernatants of preDC2sand T-cell cocultures were detected on day 5 with commercial kits (R&D).PreDC2 type I IFNs were detected using commercial kits from PBL (IFN-αand -β) and MedSystem (IFN-ω; Vienna, Austria) after 24-h activation withreplication-defective adenovirus.
Statistical analysis. Differences in cell-surface molecule expression weredetermined by χ2 test, and in other variables by unpaired t-test, with P <0.05 considered significant.
AcknowledgmentsWe thank O. Haller for antibody against MxA; A. Amara and F. Arenzana-Seisdedos for monoclonal antibody K15C against SDF-1; and S. Clayton, E.Kraus, S. Coquery, C. Chalouni and D. Olivares for technical assistance. T.J.C.was supported by grants from the NIH, the Baylor endowment and Golfersagainst Cancer. D.E., V.M. and P.G. were supported by the Association deRecherche sur le Cancer (ARC), France.
RECEIVED 18 JUNE; ACCEPTED 23 OCTOBER 2001
1. Cella, M. et al. Plasmacytoid monocytes migrate to inflamed lymph nodes and pro-duce large amounts of type I interferon. Nature Med. 5, 919–923 (1999).
2. Rissoan, M.C. et al. Reciprocal control of T helper cell and dendritic cell differentia-tion. Science 283, 1183–1186 (1999).
3. Nestle, F.O. et al. Vaccination of melanoma patients with peptide- or tumor lysate-pulsed dendritic cells. Nature Med 4, 328–332 (1998).
4. Holtl, L. et al. CD83+ blood dendritic cells as a vaccine for immunotherapy ofmetastatic renal-cell cancer. Lancet 352, 1358 (1998).
5. Hsu, F.J. et al. Vaccination of patients with B-cell lymphoma using autologous anti-gen- pulsed dendritic cells. Nature Med 2, 52–58 (1996).
6. Gabrilovich, D.I., Corak, J., Ciernik, I.F., Kavanaugh, D. & Carbone, D.P. Decreasedantigen presentation by dendritic cells in patients with breast cancer. Clin. CancerRes. 3, 483–490 (1997).
7. Gabrilovich, D. et al. Vascular endothelial growth factor inhibits the developmentof dendritic cells and dramatically affects the differentiation of multiple hematopoi-etic lineages in vivo. Blood 92, 4150–4166 (1998).
8. Menetrier-Caux, C. et al. Inhibition of the differentiation of dendritic cells from
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://m
edic
ine.
nat
ure
.co
m© 2001 Nature Publishing Group http://medicine.nature.com

1346 NATURE MEDICINE • VOLUME 7 • NUMBER 12 • DECEMBER 2001
ARTICLES
CD34+ progenitors by tumor cells: role of interleukin-6 and macrophage colony-stim-ulating factor. Blood 92, 4778–4791 (1998).
9. Chomarat, P., Banchereau, J., Davoust, J. & Palucka, A. K. IL-6 switches the differenti-ation of monocytes from dendritic cells to macrophages. Nature Immunol. 1,510–514 (2000).
10. Bell, D. et al. In breast carcinoma tissue, immature dendritic cells reside within thetumor, whereas mature dendritic cells are located in peritumoral areas. J. Exp. Med.190, 1417–1426 (1999).
11. Grouard, G. et al. The enigmatic plasmacytoid T cells develop into dendritic cells withinterleukin (IL)-3 and CD40-ligand. J. Exp. Med. 185, 1101–1111 (1997).
12. Kohrgruber, N. et al. Survival, maturation, and function of CD11c- and CD11c+ pe-ripheral blood dendritic cells are differentially regulated by cytokines. J. Immunol. 163,3250–3259 (1999).
13. Cella, M., Facchetti, F., Lanzavecchia, A. & Colonna, M. Plasmacytoid dendritic cellsactivated by influenza virus and CD40L drive a potent Th1 polarization. NatureImmunol. 1, 305–310 (2000).
14. Kadowaki, N., Antonenko, S., Lau, J. Y.-N. & Liu, Y.-J. Natural interferon α/β-produc-ing cells link innate and adaptive immunity. J. Exp. Med. 192, 219–225 (2000).
15. Siegal, F.P. et al. The nature of the principal type 1 interferon-producing cells inhuman blood. Science 284, 1835–1837 (1999).
16. Banchereau, J. & Steinman, R.M. Dendritic cells and the control of immunity. Nature392, 245–252 (1998).
17. Rubbert, A. et al. Dendritic cells express multiple chemokine receptors used as core-ceptors for HIV entry. J. Immunol. 160, 3933–3941 (1998).
18. Kellermann, S.A., Hudak, S., Oldham, E.R., Liu, Y.J. & McEvoy, L.M. The CCchemokine receptor-7 ligands 6Ckine and macrophage inflammatory protein-3 β arepotent chemoattractants for in vitro- and in vivo-derived dendritic cells. J. Immunol.162, 3859–3864 (1999).
19. Foti, M. et al. Upon dendritic cell (DC) activation chemokines and chemokine recep-tor expression are rapidly regulated for recruitment and maintenance of DC at the in-flammatory site. Int. Immunol. 11, 979–986 (1999).
20. Dieu, M.C. et al. Selective recruitment of immature and mature dendritic cells by dis-tinct chemokines expressed in different anatomic sites. J. Exp. Med. 188, 373–386(1998).
21. Jahnsen, F.L. et al. Experimentally induced recruitment of plasmacytoid (CD123high)dendritic cells in human nasal allergy. J. Immunol. 165, 4062–4068 (2000).
22. Roers, A., Hochkeppel, H.K., Horisberger, M.A., Hovanessian, A. & Haller, O. MxAgene expression after live virus vaccination: a sensitive marker for endogenous type Iinterferon. J. Infect. Dis. 169, 807–813 (1994).
23. Ito, T. et al. A CD1a+/CD11c+ subset of human blood dendritic cells is a direct precur-sor of Langerhans cells. J. Immunol. 163, 1409–1419 (1999).
24. Pulendran, B. et al. Flt3-ligand and granulocyte colony-stimulating factor mobilizedistinct human dendritic cell subsets in vivo. J. Immunol. 165, 566–572 (2000).
25. Zou, W. et al. Quantification of cytokine gene expression by competitive PCR using acolorimetric assay. Eur. Cytokine Netw. 6, 257–264 (1995).
26. Coulomb-L’Hermin, A. et al. Stromal cell-derived factor 1 (SDF-1) and antenatalhuman B cell lymphopoiesis: expression of SDF-1 by mesothelial cells and biliary duc-tal plate epithelial cells. Proc. Natl. Acad. Sci. USA 96, 8585–90 (1999).
27. Rollins, B. J. Chemokines. Blood 90, 909–28 (1997).28. Tilton, B. et al. Signal transduction by CXC chemokine receptor 4. Stromal cell-de-
rived factor 1 stimulates prolonged protein kinase b and extracellular signal-regu-lated kinase 2 activation in T lymphocytes. J. Exp. Med. 192, 313–324 (2000).
29. Tillman, B.W. et al. Maturation of dendritic cells accompanies high-efficiency genetransfer by a CD40-targeted adenoviral vector. J. Immunol. 162, 6378–6383(1999).
30. Mayordomo, J.I. et al. Bone marrow-derived dendritic cells pulsed with synthetictumour peptides elicit protective and therapeutic antitumour immunity. NatureMed. 1, 1297–1302 (1995).
31. Rabinowich, H. et al. Expression of cytokine genes or proteins and signaling mole-cules in lymphocytes associated with human ovarian carcinoma. Int. J. Cancer 68,276–284 (1996).
32. Beatty, P., Hanisch, F.G., Stolz, D.B., Finn, O.J. & Ciborowski, P. Biochemical char-acterization of the soluble form of tumor antigen MUC1 isolated from sera and as-cites fluid of breast and pancreatic cancer patients. Clin. Cancer Res. 7, 781s–787s(2001).
33. Olweus, J. et al. Dendritic cell ontogeny: a human dendritic cell lineage of myeloidorigin. Proc. Natl. Acad. Sci. USA 94, 12551–1256 (1997).
34. Sallusto, F. et al. Distinct patterns and kinetics of chemokine production regulatedendritic cell function. Eur J. Immunol. 29, 1617–1625 (1999).
35. Geijtenbeek, B.H.T. DC-SIGN-ICAM-2 interaction mediates dendritic cell traffick-ing. Nature Immunol. 1, 353–357 (2000).
36. Campbell, J. J. et al. Chemokines and the arrest of lymphocytes rolling under flowconditions. Science 279, 381–4 (1998).
37. Peled, A. et al. The chemokine SDF-1 activates the integrins LFA-1, VLA-4, and VLA-5 on immature human CD34+ cells: role in transendothelial/stromal migration andengraftment of NOD/SCID mice. Blood 95, 3289–3296 (2000).
38. Enk, A. H., Jonuleit, H., Saloga, J. & Knop, J. Dendritic cells as mediators of tumor-induced tolerance in metastatic melanoma. Int. J. Cancer 73, 309–316 (1997).
39. Kiertscher, S.M., Luo, J., Dubinett, S.M. & Roth, M.D. Tumors promote alteredmaturation and early apoptosis of monocyte-derived dendritic cells. J. Immunol.164, 1269–1276 (2000).
40. Marrack, P., Kappler, J. & Mitchell, T. Type I interferons keep activated T cells alive.J. Exp. Med. 189, 521–530 (1999).
41. Zou, W. et al. Reciprical regulation of plasmacytoid dendritic cells and monocytesduring viral infection. Eur.J. Immunol. (in the press).
42. Groux, H., Bigler, M., de Vries, J.E. & Roncarolo, M.G. Inhibitory and stimulatory ef-fects of IL-10 on human CD8+ T cells. J. Immunol. 160, 3188–3193 (1998).
43. Nanki, T. & Lipsky, P.E. Cutting edge: stromal cell-derived factor-1 is a costimulatorfor CD4+ T cell activation. J. Immunol. 164, 5010–5014 (2000).
44. Muller, A. et al. Involvement of chemokine receptors in breast cancer metastasis.Nature 410, 50–56 (2001).
45. Zou, W. et al. Macrophage-derived dendritic cells have strong Th1-polarizing po-tential mediated by β-chemokines rather than IL-12. J. Immunol. 165, 4388–4396(2000).
46. Berard, F. et al. Cross-priming of naive CD8 T cells against melanoma antigensusing dendritic cells loaded with killed allogeneic melanoma cells. J. Exp. Med. 192,1535–1544 (2000).
47. Zou, W. et al. A Guide to isolation, culture and propagation of dendritic cells. inDendritic Cells: Biology and Clinical Applications, 2nd edn. (ed. Lotze, M.) 77(Academic Press, London, 2001).
©20
01 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://m
edic
ine.
nat
ure
.co
m© 2001 Nature Publishing Group http://medicine.nature.com
Copyright © 2022 FDOKUMEN




















