
Structural and Antigenic Properties of Merozoite Surface Protein 4 of Plasmodium falciparum

Esther Lutgens, Martin Zenke, Christoph J. Binder, Christian Weber and Alma ZerneckeSoehnlein, Martin Busch, Marco Manca, Rory R. Koenen, Jaroslav Pelisek, Mat J. Daemen,
Yvonne Döring, Helga D. Manthey, Maik Drechsler, Dirk Lievens, Remco T.A. Megens, OliverPromote Atherosclerosis
Auto-Antigenic Protein-DNA Complexes Stimulate Plasmacytoid Dendritic Cells to
Print ISSN: 0009-7322. Online ISSN: 1524-4539 Copyright © 2012 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation doi: 10.1161/CIRCULATIONAHA.111.046755
2012;125:1673-1683; originally published online March 2, 2012;Circulation.
http://circ.ahajournals.org/content/125/13/1673World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circ.ahajournals.org/content/suppl/2012/03/02/CIRCULATIONAHA.111.046755.DC1.htmlData Supplement (unedited) at:
http://circ.ahajournals.org//subscriptions/
is online at: Circulation Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer this process is available in the
click Request Permissions in the middle column of the Web page under Services. Further information aboutOffice. Once the online version of the published article for which permission is being requested is located,
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the EditorialCirculationin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

Auto-Antigenic Protein-DNA Complexes StimulatePlasmacytoid Dendritic Cells to Promote Atherosclerosis
Yvonne Doring, PhD; Helga D. Manthey, PhD; Maik Drechsler, PhD; Dirk Lievens, PhD;Remco T.A. Megens, PhD; Oliver Soehnlein, MD, PhD; Martin Busch, MSc; Marco Manca, PhD;
Rory R. Koenen, PhD; Jaroslav Pelisek, PhD; Mat J. Daemen, MD, PhD; Esther Lutgens, MD, PhD;Martin Zenke, PhD; Christoph J. Binder, MD, PhD; Christian Weber, MD; Alma Zernecke, MD
Background—Inflammation has been closely linked to auto-immunogenic processes in atherosclerosis. Plasmacytoiddendritic cells (pDCs) are specialized to produce type-I interferons in response to pathogenic single-stranded nucleicacids, but can also sense self-DNA released from dying cells or in neutrophil extracellular traps complexed to theantimicrobial peptide Cramp/LL37 in autoimmune disease. However, the exact role of pDCs in atherosclerosisremains elusive.
Methods and Results—Here we demonstrate that pDCs can be detected in murine and human atherosclerotic lesions. Exposureto oxidatively modified low-density lipoprotein enhanced the capacity of pDCs to phagocytose and prime antigen-specific Tcell responses. Plasmacytoid DCs can be stimulated to produce interferon-� by Cramp/DNA complexes, and we furtheridentified increased expression of Cramp and formation of neutrophil extracellular traps in atherosclerotic arteries.Whereas Cramp/DNA complexes aggravated atherosclerotic lesion formation in apolipoprotein E– deficient mice,pDC depletion and Cramp-deficiency in bone marrow reduced atherosclerosis and anti– double-stranded DNAantibody titers. Moreover, the specific activation of pDCs and interferon-� treatment promoted plaque growth,associated with enhanced anti– double-stranded–DNA antibody titers. Accordingly, anti– double-stranded DNAantibodies were elevated in patients with symptomatic versus asymptomatic carotid artery stenosis.
Conclusions—Self-DNA (eg, released from dying cells or in neutrophil extracellular traps) and an increasedexpression of the antimicrobial peptide Cramp/LL37 in atherosclerotic lesions may thus stimulate a pDC-drivenpathway of autoimmune activation and the generation of anti– double-stranded-DNA antibodies, criticallyaggravating atherosclerosis lesion formation. These key factors may thus represent novel therapeutic targets. (Circulation.2012;125:1673-1683.)
Key Words: atherosclerosis � dendritic cells � immune system � inflammation � leukocytes
Atherosclerosis is a chronic inflammatory disease of thearterial wall, modulated by innate and adaptive immune
responses. Besides monocytes/macrophages, T cells and no-tably also plasmacytoid dendritic cells (pDCs) can be de-tected within human atherosclerotic lesions.1–4 PlasmacytoidDCs uniquely express high levels of Toll-like receptor (TLR)7 and 9 specialized for sensing pathogenic single-strandednucleic acids during viral and microbial infections, resultingin the production of large amounts of type-I interferons(IFN-�/�).5,6 Although pDCs usually do not respond toself-DNA,7 it became evident that cathelicidin antimicrobial
peptide LL37 (known as Cramp in mice) can convert self-DNA into a trigger of pDC activation in human autoimmunediseases (eg, systemic lupus erythematosus or psoriasis).7
Additionally, neutrophil extracellular traps (NETs) and cir-culating immune complexes containing self-DNA and anti-microbial peptides were recently shown to trigger pDCactivation via TLR9.8–11
Clinical Perspective on p 1683Previously, human plaque–residing pDCs were shown to
respond to type-A oligonucleotides (CpGs) with an enhanced
Received June 1, 2011; accepted February 22, 2012.From the Institute for Cardiovascular Prevention, Ludwig-Maximilians-University Munich, Munich, Germany (Y.D., M.D., D.L., R.T.A.M., O.S., R.R.K.,
E.L., C.W.); Institute for Biomedical Engineering, Department of Cell Biology, RWTH Aachen University, Aachen, Germany (Y.D., M.Z.); Rudolf VirchowCenter, DFG Research Center for Experimental Medicine, University of Wurzburg, Wurzburg, Germany (H.D.M., M.B., A.Z.); Cardiovascular ResearchInstitute Maastricht, Maastricht University, Maastricht, the Netherlands (M.M., M.D., C.W.); Department of Vascular Surgery, Klinikum rechtsder Isar der Technischen Universitat Munchen, Munich, Germany (J.P.); Center for Molecular Medicine (CeMM) of the Austrian Academy of Sciences andDepartment of Laboratory Medicine, Medical University of Vienna, Vienna, Austria (C.J.B.); and Munich Heart Alliance, Munich, Germany (C.W.).
The online-only Data Supplement is available with this article at http://circ.ahajournals.org/lookup/suppl/doi:10.1161/CIRCULATIONAHA.111.046755/-/DC1.
Correspondence to Alma Zernecke, MD, Rudolf Virchow Center, DFG Research Center for Experimental Medicine, University of Wurzburg,Josef-Schneider Str. 2, 97080 Wurzburg, Germany. E-mail [email protected]
© 2012 American Heart Association, Inc.
Circulation is available at http://circ.ahajournals.org DOI: 10.1161/CIRCULATIONAHA.111.046755
1673 at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

IFN-� expression, which in turn amplified inflammatorycytokine responses and immune cell activation,3,4 suggestingproatherogenic functions of this DC subset. Conversely,pDCs can also exert tolerogenic functions, as shown in avascularized graft model.12 Thus, the role of pDCs in athero-sclerosis remains to be conclusively explored.
MethodsMouse ModelsApolipoprotein E–deficient (Apoe�/�), low-density lipoprotein–deficient (Ldlr�/�), lymphocyte-deficient (Rag1�/�), and wild-typemice (C57BL/6J) were purchased from the Jackson Laboratory. At 6to 8 weeks, female mice were subjected to a high-fat diet (HFD, 21%fat, 0.15% cholesterol, Altromin). Plasmacytoid DC depletion inApoe�/� mice was performed by i.v. injection of the monoclonalantibody PDCA113,14 (500 �g/mouse, Miltenyi Biotech) or anisotype control antibody (500 �g/mouse, BioXcell) at day 1 and day7 after the initiation of HFD feeding. PDCA1 antibody treatmentefficiently depleted pDCs in lymph nodes, spleen, and bone marrowwhile not altering cDC, T-cell or B-cell frequencies, as assessed byfluorescence-activated cell-sorting (FACS) analysis (online-onlyData Supplement Figure I). Some mice were injected with oligonu-cleotide CpG 1585 (25 �g/mouse, Inovagen) complexed to DOTAP(15 �g/mouse, Roche) or with DOTAP or PBS alone 3 times/weekfor 4 weeks. Some mice were i.p. injected with murine IFN-�(100 000 U, HyCultBiotec) 2 times/week for 4 weeks. SomeApoe�/� mice were i.v. injected with Cramp/DNA complexes15 3times/week for 4 weeks, together with receiving PDCA1 or isotypecontrol antibody. Female Ldlr�/� mice were irradiated and recon-stituted with Cramp�/�16 or Cramp�/� bone marrow17 and afterrecovery were placed on HFD for 4 weeks. Some Apoe�/� micewere treated with Cramp (i.p. 100 �g/mouse), DNase I (i.v., 120U/mouse, Sigma), or both 3 times/week for 4 weeks. No changes incholesterol or triglyceride serum levels, as assessed by EnzyChromhigh-density lipoprotein and LDL/VLDL Assay Kit (BioAssay Sys-tems), were observed between groups of mice (online-only DataSupplement Table I). Rag1�/�Ldlr�/� mice were fed a HFD for 12weeks. High-fat diet–fed lysozyme M (lys)–enhanced green fluores-cent protein (EGFP) Apoe�/� mice18 were i.v. injected with 250 �Lof liposomes containing clodronate19 1 day before and propidiumiodide (2 �g/mL final concentration) immediately before imaging.Experiments were approved by local authorities.
Atherosclerotic Lesion Quantificationand ImmunohistochemistryThe extent of atherosclerosis was assessed in aortic roots and inaortas by staining for lipid depositions with Oil-red-O17. The relativecontent of macrophages and the localization of pDCs, Cramp, andplaque immunoglobulin Gs (IgGs) were determined by immunofluo-rescence staining. Nuclei were counterstained by 4�,6-Diamidino-2-phenylindol (DAPI). Human carotid artery specimens were obtainedfrom autopsy or vascular surgery. The location of blood dendriticcell antigen-2 (BDCA2) and LL37 expression in the plaque wasdetermined using standard immunohistochemistry techniques.20
DC Isolation and Stimulation In VitroPlasmacytoid DCs were isolated from spleens of C57BL/6J miceusing the Plasmacytoid Dendritic Cell Isolation Kit II (MiltenyiBiotec). Alternatively, pDCs were sorted (FACSAria, BD Biosci-ence) from bone marrow of C57BL/6J with an antibody cocktailagainst CD45, SiglecH, and CD45RA (B220). Isolated pDCs werestimulated with oxidatively modified low-density lipoprotein (ox-LDL) or di-oxLDL (50 �g/mL), Cramp (50 �g/mL, Innovagen),DNA (10 �g/mL, mouse C57Bl6/6J female genomic DNA, Zyagen),CpG 1585 (5 �g/mL, Invivogen), or Cramp/DNA complexes15 (10�g Cramp, mixed with 2 �g of DNA), 4 �L of mouse serumcontaining high (36 U/�L) or low (1 U/�L) titers of anti–double-
stranded DNA (antidsDNA) antibodies per well, or serum obtainedfrom Rag1�/�Ldlr�/� mice.
Antigen-Specific T-Cell Proliferation In VivoCD4� T cells were isolated from the spleen of OT-II mice (CD4 Tcell isolation kit, Miltenyi), and labeled with CFSE (Fluka). Plas-macytoid DCs prestimulated with oxLDL (50 �g/mL), CpG1585 (5�g/mL), or a combination of both were pulsed for 45 minutes withOVA323 to 339 peptide (30 �g/mL). Untreated pDCs were used ascontrol. For in vivo studies, 5�106 CD4� CFSE-labeled OT-II cellswere i.v. injected into C57BL/6 recipient mice. One day later, 1�105
untreated or treated OVA-2 peptide-pulsed pDCs were injected intothe footpad. After 3 days, T-cell proliferation was analyzed by CFSEdye dilution in peripheral lymph nodes of recipient mice by flowcytometry.
Flow Cytometry and ELISALymph nodes and spleen were dissociated into single-cell suspen-sions using collagenase D, and aortas digested using Liberase III(Roche). Staining for flow cytometric analysis was conducted usingcombinations of specific antibodies from eBiosciences, Miltenyi,and BD Bioscience. Data were acquired using a FACSCantoII (BDBioscience), and results analyzed by FlowJo software (Tree Star).Cell culture supernatants or sera were analyzed by ELISA detectingIFN� (VeriKine Mouse Interferon-Alpha ELISA Kit, PBL Labora-tories), dsDNA (Mouse antidsDNA Kit, Alpha Diagnostic), andantidsDNA antibodies (Varelisa dsDNA Antibody ELISA, Phadia)according to the manufacturer’s protocol; antidsDNA Ab (�) �55IU/mL. Reciprocal serum dilutions to determine antinuclear antibod-ies in humans were performed by indirect immunofluorescence onHEp-2 substrate according to the manufacturer’s instructions (ANAIFT IgG Kit, Viramed); (�) �1:160. Antibodies to malondialdeyhde-modified (MDA)-LDL and oxLDL were measured by chemilumines-cent ELISA.21
Quantitative Real-Time PolymeraseChain ReactionRNA was isolated from aortas with the ZR RNA MicroPrep RNAisolation kit (Zymo Research), and complementary DNA was reversetranscribed from total RNA (Promega). Real-time polymerase chainreaction analysis was performed using SYBRGreen (Fermentas) andspecific primer pairs. Amplification was performed in duplicateusing a 7900 HT fast real-time polymerase chain reaction system(Applied Biosystems).
Two-Photon MicroscopyIn vivo 2-photon microscopy of exposed carotid arteries wasperformed as described.22 In short, the exposed left carotid arterywas visualized using an upright Olympus FV1000MPE 2-photonmicroscope system equipped with a 1.05 water dipping objective. Apulsed Ti-Sapphire laser (MaiTai DeepSee, Spectra Physics) wastuned to 810 nm for visualization of second harmonic generation(SHG) signal of collagen, green fluorescent protein (GFP), andpropidium iodide. Emitted fluorescent signals were detected with 3photomultiplier tubes. Series of subsequent xy-sections (256�256pixels) were obtained over time at an acquisition rate of 4Hz. Allimage processing was performed using Image-Pro 3D Analyzer 7.0(Media Cybernetics).
Human Microarray Data EvaluationSeventeen atherosclerotic tissue samples from atherosclerotic carotidartery segments (9 from early and 8 from advanced lesions) from theMaastricht Pathology Tissue Collection were processed as de-scribed.23 Total RNA was isolated and purified and RNA qualitydetermined using a 20100 Bioanalyzer (Agilent Technologies).Samples with an RNA integrity number of 5 were further processed(mean RNA integrity number�7.8). Double-stranded complemen-tary DNA was synthesized (One-Cycle Target Labeling Kit, Af-fymetrix) and used as a template for the preparation of biotin-labeledcomplementary RNA using the GeneChip IVT Labeling Kit (Af-
1674 Circulation April 3, 2012
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

fymetrix). Biotin-labeled complementary RNA was hybridized induplicates to the Human Genome U133 Plus 2.0 Array (Affymetrix),washed, and stained. Arrays were laser scanned with the GeneChipScanner 3000 (Affymetrix). Data were quantified using GCOS 1.2(Affymetrix), and the resulting CEL files processed in R. Theresulting expression set was analyzed by limma. After Bayesiansmoothing of the SEs of the estimated log-fold changes, thetranscripts of interest were reported as log2 fold changes, averageexpression, and Benjamin-Hochberg false discovery rate adjusted Pvalues.
Study PatientsA total of 30 patients with advanced carotid artery stenosis (�70%,determined by measuring the peak systolic velocity by ultrasonog-raphy) were included in this study. Asymptomatic patients (n�15)without neurological symptoms were diagnosed at the VascularCentre of the Technical University Munich; symptomatic patients(n�15) displaying neurological symptoms (transient ischemic attackor nondisabling stroke) were assigned from the Institute of Neurol-ogy of the hospital Klinikum rechts der Isar, Munich. Patients withchronic kidney disease, diabetes mellitus, autoimmune disease, andongoing steroid or other immunosuppressive medication were ex-cluded from the study. Blood sampling was performed within 2 dayspreceding surgical intervention.
StatisticsUnpaired Student t tests were used to compare Cramp messengerRNA (mRNA) and dsDNA antibody titers between control andHFD-fed Apoe�/� mice, dsDNA antibody titers between IgG- andPDCA1-treated Apoe�/� mice, and dsDNA antibody titers andplaque size between Cramp�/�‹Ldlr�/� and Cramp�/�‹Ldlr�/�
mice. Mann Whitney tests were applied to compare pDC frequenciesand IFN-� levels between control and HFD-fed Apoe�/� mice,Di-oxLDL uptake in vitro, plaque sizes between IgG- and PDCA1-
treated Apoe�/� mice, and dsDNA antibody titers between patientswith asymptomatic versus symptomatic carotid artery stenosis.ANOVA 1-way analysis of variance followed by the Tukey multiplecomparison post tests were used for comparing SigH and IFN�mRNA expression between control and HFD-fed Apoe�/� mice,CD36 expression, OVAII-488 uptake, and IFN-� secretion by pDCsin vitro, plaque size between controls, CpG, and CpG�PDCA1-treated, or Cramp and Cramp�DNAse-treated Apoe�/� mice.Kruskal-Wallis tests were applied to compare T-cell proliferation invivo and plaque size between controls, Cramp/DNA�isotype, andCramp/DNA�PDCA1-treated Apoe�/� mice. Differences where Pwas �0.05 were considered statistically significant.
ResultsPlasmacytoid DCs are not only present in lymph nodes andspleen (online-only Data Supplement Figure II) but couldalso be detected in atherosclerotic lesions. As shown bystaining for Siglec-H�, single pDCs were localized to theplaque shoulders and areas bordering the necrotic core inaortic root plaques of Apoe�/� mice fed a HFD for 12 weeks(Figure 1A). Compared with healthy wild-type Apoe�/� miceor Apoe�/� mice on normal chow, diet-fed Apoe�/� micedisplayed higher frequencies of aortic PDCA1� pDCs (Fig-ure 1B, data not shown). In addition, mRNA transcripts ofSiglecH and Ifn-� were increased in aortic tissue togetherwith elevated serum protein levels of IFN-� in Apoe�/� micefed a HFD compared with Apoe�/� mice on normal chow(Figure 1C and D).
Modified lipoproteins (eg, oxLDL) are deposited in thearterial wall during atherogenesis1,2 and can be taken up by
Figure 1. Plasmacytoid dendritic cells (pDCs) accumulate in atherosclerosis. A, Single SiglecH� pDCs (green, red arrow heads) weredetected by immunofluorescence in the aortic root of apolipoprotein E–deficient (Apoe�/�) mice fed a high-fat diet (HFD) for 12 weeks;cell nuclei are counterstained with DAPI (blue). B, Enzymatic digestion and fluorescence-activated cell-sorting (FACS) analysis of aortaeof healthy 6-week-old Apoe�/� mice on normal chow (NC) and Apoe�/� mice fed a HFD for 12 weeks; representative dot blots andpercentages within quadrants are shown, SSC indicates side scatter (n�5 mice per group). Bars represent median with interquartilerange. C, SiglecH (SigH) and interferon � (Ifn-�) messenger RNA (mRNA) expression in aortae of HFD-fed Apoe�/� mice (n�5) relativeto expression in healthy 6-week-old Apoe�/� mice (control, n�6 mice per group). Bars represent mean�SD. D, Interferon � serum pro-tein levels in HFD-fed Apoe�/� mice (n�9) relative to expression in healthy 6-week-old Apoe�/� mice (n�5) on NC. Bars representmedian with interquartile range. *P�0.05, **P�0.01, ***P�0.001. Apoe, apolipoprotein E; pDCs, plasmacytoid dendritic cells; NC, nor-mal chow; HFD, high-fat diet; mRNA, messenger RNA; ctrl, control; SiglecH, sialic acid binding immunoglobulin-like lectin H; and IFN�,interferon �.
Doring et al pDCs in Atherosclerosis 1675
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

CD11c� DCs in the vessel wall.24 Moreover, oxLDL hasbeen found to induce the upregulation of scavenger receptorson DCs.25 We therefore tested the capacity of pDCs to take upand to respond to oxLDL. Indeed, sorted pDCs were capableof taking up di-labeled oxLDL (Figure 2A). Similar toprototypical TLR7/TLR9 activation with CpGs,6 exposure tooxLDL selectively enhanced surface expression of CD36(Figure 2A), which has been implicated in its phagocytosis.26
However, oxLDL did not alter surface expression of CD68,SR-A, MHC-II, and CD86 nor secretion of IFN-� protein,whereas MHC-II and CD86 expression increased after CpGstimulation (online-only Data Supplement Figure IIIA anddata not shown).
Mature pDCs are capable of initiating T-cell responses.27
Accordingly, we observed that OVA-2–loaded pDCs but notunpulsed pDCs induced antigen-specific proliferation ofOT-II CD4� T cells in vivo. Interestingly, T-cell proliferativeresponses were potentiated by pretreatment of pDCs withoxLDL, amounting to a response resembling that after pre-treatment with CpGs (Figure 2B). Combining oxLDL andCpGs did not yield additive effects (data not shown). A robustIFN-� protein secretion was detectable in CpG-stimulated
pDC–T-cell cocultures in vitro but not in supernatants of Tcells cocultured with pDCs or after addition of oxLDL(online-only Data Supplement Figure IIIB). Notably, oxLDLtreatment enhanced phagocytosis of the fluorescently labeledOVA-2 antigen in a manner similar to that of CpGs (Figure2C), implying that the enhanced phagocytic capacity onexposure to oxLDL may boost pDC-driven immune re-sponses in the context of atherosclerosis.
To scrutinize the role of pDCs in atherosclerotic lesiondevelopment, Apoe�/� mice fed a HFD were injected with apDC-depleting PDCA1 or isotype control antibody. After 4weeks, a substantial reduction in plaque size in the aortic rootand aorta as well as a decrease in lesional macrophagefrequencies was observed in pDC-depleted Apoe�/� micecompared with IgG-injected Apoe�/� controls (Figure 3A,online-only Data Supplement Figure IVA). This decrease inplaque size was associated with a significant reduction inIFN-� serum levels in Apoe�/� mice depleted of pDCscompared with IgG-injected Apoe�/� controls (10.5�4.5versus 44.1�11.7 pg/mL, respectively, P�0.05). Conversely,aggravated lesion formation and macrophage content wasobserved in diet-fed Apoe�/� mice injected with IFN-� for 4
Figure 2. Oxidatively modified low-density lipoprotein (oxLDL) can be taken up by plasmacytoid dendritic cells (pDCs) and enhancespDC-driven T-cell responses. A, The uptake of Di-labeled oxLDL (left) and expression of CD36 (right) after exposure to oxLDL, type-Aoligonucleotides (CpGs), or PBS over 12 hours by sorted plasmacytoid dendritic cells (pDCs) assessed by fluorescence-activated cell-sorting (FACS) analysis; representative histograms with fluorescence minus 1 (FMO) controls and quantifications of mean fluorescenceof 3 independent experiments are shown. Bars represent median with interquartile range. B, Sorted pDCs (unpulsed control), OVA-2-pulsed pDCs pretreated with or without oxLDL or CpGs over 12 hours were transferred into wild-type mice transfused with CFSE�OT-IIT cells. After 3 days, T-cell proliferation was quantified by CSFE dilution and FACS analysis; representative dot plots and percentage ofCFSE�CD4� T cells (red markers) within gates are shown, n�4 independent experiments. Bars represent median with interquartilerange. C, The uptake of 488-labeled OVA peptide pretreated with PBS, oxLDL (left) or CpGs (right) over 12 hours was assessed insorted pDCs by FACS analysis; representative histograms with FMO controls and quantifications of mean fluorescence of 3 indepen-dent experiments are shown. Bars represent mean�SD. *P�0.05, ***P�0.001. oxLDL indicates oxidatively modified low-density lipo-protein; and CpG, type-A oligonucleotide.
1676 Circulation April 3, 2012
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

weeks compared with vehicle-treated mice (online-only DataSupplement Figure IVB). To further assess sequelae of pDCactivation, diet-fed Apoe�/� mice were repetitively injectedwith CpGs over 4 weeks. Lesion formation and macrophageaccumulation were exacerbated in the aortic root and aorta inCpG-injected Apoe�/� mice compared with control-treatedmice (Figure 3B, online-only Data Supplement Figure IVC).These effects were abrogated in Apoe�/� mice additionallydepleted of pDCs by PDCA1 antibody injections (Figure 3B,online-only Data Supplement Figure IVC). These findingsclearly demonstrate that pDCs and their effector cytokineIFN-� drive early lesion development.
Breakdown of tolerance to otherwise inert self-DNA frag-ments released by dying cells6 (eg, in psoriasis) can betriggered by Cramp, an antimicrobial peptide rapidly releasedfrom neutrophil granules,28,29 which binds self-nucleic acidsto form complexes that trigger TLR7/TLR9 activation andtype-I IFN secretion by pDCs.11,15 Also, NETs containingself-DNA and antimicrobial peptides released by activatedneutrophils during cell death, a process termed “NETosis,”can trigger pDC activation.7,10,30 Like CpGs and dsDNAcontaining CpG motifs15 (online-only Data Supplement Fig-ure VA), wild-type Bl6 DNA complexed with Cramp (but notDNA or Cramp alone) induced IFN-� production by murinepDCs (Figure 4A), without affecting MHC-II expression(online-only Data Supplement Figure VB). Likewise, a pDC-dependent increase in IFN-� serum levels was observed inBl6 mice infused with Cramp/DNA complexes (online-onlyData Supplement Figure VC), confirming pDC responsestoward DNA-containing complexes also in vivo.
An increased expression of Cramp mRNA (Figure 4B) andCramp protein (23.1�6.9 versus 0% staining/plaque area)
was observed in the vessel wall of diet-fed Apoe�/� micecompared with healthy Apoe�/� controls. Cramp was de-tected in the vicinity of segment-nucleated neutrophils withinatherosclerotic lesions (Figure 4C), likely reflecting neutro-phil accumulation during early lesion development,17 and incolocalization with small DNA fragments in necrotic coreareas (Figure 4C). Using 2-photon microscopy of atheroscle-rotic carotid arteries, we furthermore revealed the presence ofNETs in atherosclerotic carotid arteries and within plaques ofmonocyte-depleted LysmEGFP/EGFP Apoe�/� mice carryingonly fluorescent neutrophils (Figure 4D, data not shown).Extracellular NETs were detected by use of propidium iodide,a vital cell-impermeable nuclear marker that stained clouds ofDNA in the vicinity of EGFP� neutrophils at atheroscleroticsites and could be observed as early as 2 weeks after theinitiation of diet (online-only Data Supplement Table II).Notably, the injection of Cramp/DNA complexes intoApoe�/� mice promoted an enhanced atherosclerotic lesionformation in the aortic root when compared with control-injected Apoe�/� mice, which was averted in mice addition-ally depleted of pDCs by PDCA1 antibody treatment (Figure4E). These data clearly indicate that Cramp/self-DNAcomplex-mediated pDC activation drives atheroscleroticplaque development. Thus, Cramp-complexed self-DNA de-rived from neutrophil-expelled NETs and dying plaque cellsmay contribute to breakdown of tolerance to self-DNA.7–10,31
In systemic lupus erythematosus, patients were found todevelop autoantibodies to self-DNA and antimicrobial pep-tides in NETs, indicating that these complexes serve asauto-antigens to trigger B-cell activation and the generationof anti-DNA–specific antibodies.7 We here demonstrate thatserum antidsDNA antibody titers were increased in hyperlip-
Figure 3. Plasmacytoid dendritic cells (pDCs) and their activation drive atherogenesis. A and B, Oil-red� atherosclerotic lesions in theaortic root in apolipoprotein E–deficient (Apoe�/�) mice fed a high fat diet (HFD) and treated with immunoglobulin G (IgG) or PDCA1antibody (A, bars represent median with interquartile range) or DOTAP (control), type-A oligonucleotides (CpGs) or CpGs�PDCA1 (B,bars represent mean�SD) for 4 weeks; representative images of the aortic root are shown. *P�0.05. IgG indicates immunoglobulin G;CpG, type-A oligonucleotide.
Doring et al pDCs in Atherosclerosis 1677
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

idemic Apoe�/� mice compared with healthy controlApoe�/� mice (Figure 5A). Conversely, antidsDNA antibodytiters were reduced in Apoe�/� mice treated with PDCA1antibody to deplete pDCs compared with IgG-injected con-trols. Furthermore, antidsDNA titers were elevated in IFN-�–and CpG-treated Apoe�/� mice but not in CpG-injected micedepleted from pDCs (online-only Data Supplement FigureVD and Figure 5B). Interestingly, humoral responses to otherknown auto-antigens in atherosclerosis, namely MDA-LDLand oxLDL, were not affected by pDC depletion, as shown byunaltered IgG antibody concentrations toward these antigensin PDCA1 versus control-treated Apoe�/� mice (online-onlyData Supplement Figure VI). This indicates that pDCsspecifically promote dsDNA antibody formation in earlyatherosclerosis. However, increased MDA-LDL and oxLDL-specific IgG antibody levels were observed in CpG-treatedApoe�/� mice but not in mice additionally treated withPDCA1 antibody (online-only Data Supplement Figure VI).This suggests that strong pDC activation also promotesunspecific amplification of humoral immune responses toantigens in atherosclerosis, likely because of abundant type-IIFN production.32,33
Notably, diminished antidsDNA antibody titers were ob-served in Ldlr�/� mice reconstituted with Cramp�/� versusCramp�/� bone marrow (Figure 5C) together with a protec-tion from atherosclerotic lesion formation (Figure 5D) and areduction in macrophage content in the aortic root (19.2�2.4versus 30.2�3.5%, respectively, P�0.05). Conversely, treat-ing mice with Cramp promoted atherosclerotic plaque growthcompared with control-treated Apoe�/� mice but was inef-fective in mice additionally treated with DNAse to degradeextracellular DNA (Figure 5E), indicating that proatherogenicfunctions of Cramp require the association with extracellularself-DNA.
Immunogenic DNA-containing immune complexes havebeen shown to contain LL37 and to further enhance pDCactivation.7–10 This concurs with results that serum ofApoe�/� mice containing high levels of antidsDNA antibodytiters but not serum containing low titers significantly in-creased IFN-� production in pDCs (Figure 5F), leavingMHC-II expression unaltered (online-only Data SupplementFigure VE). Conversely, serum derived from atheroscleroticRag1�/�Ldlr�/� mice did not elicit IFN-� responses in pDCs(data not shown). Although we refrained from purifying
Figure 4. Auto-antigenic protein-DNAcomplexes stimulate plasmacytoid den-dritic cells (pDCs), and Cramp and NETsare detectable in atherosclerotic arteries.A, Interferon � (IFN-�) protein secretionby sorted pDCs stimulated overnight asindicated (Bars represent mean�SD,n�3–5 independent experiments). B,Cramp messenger RNA (mRNA) expres-sion in aortas of atherosclerotic high-fatdiet (HFD)–fed apolipoprotein E–deficient(Apoe�/�) mice (n�9) relative to expres-sion in healthy 6-week-old Apoe�/� mice(n�5). Bars represent mean�SD. C,Cramp staining (red) detected by immu-nofluorescence in the aortic root in diet-fed Apoe�/�; cell nuclei and DNA arestained by DAPI (blue); arrowheads indi-cate segment-nucleated neutrophils. D,Luminal presence of propidium iodide–positive extracellular DNA (red) in colo-calization with enhanced green fluores-cent protein–positive (EGFP�)neutrophils (green) in atheroscleroticLysmEGFP/EGFP Apoe�/� mice; collagenof the vessel wall was visualized by sec-ond harmonic generation (SHG, blue);autofluorescence of elastin bands of thevessel wall appears in aquamarine. E,Oil-red� lesions in the aortic root in diet-fed Apoe�/� mice treated with PBS(control), or Cramp/DNA complexestogether with isotype control or PDCA1antibody for 4 weeks; representativeimages of the aortic root are shown.Bars represent median with interquartilerange. *P�0.05, ***P�0.001. IFN-� indi-cates interferon �; mRNA, messengerRNA; HFD, high-fat diet.
1678 Circulation April 3, 2012
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

IgG-containing immune complexes because of the limitedamount of serum available from mice, these data suggest thatIgG complexes containing DNA in serum may contribute topDC activation. Interestingly, abundant IgG deposits could bedetected in atherosclerotic plaques of diet-fed Apoe�/� mice,predominantly in the vicinity of necrotic plaque areas (Figure5G). Thus, auto-immunogenic complexes deposited withinlesions in vivo may contribute to the pathogenic insult inatherosclerosis.
To assess whether these mechanisms may also be relevantto human disease, differential expression analysis was per-formed on gene expression microarrays from human athero-sclerotic lesions. Comparisons of early and advanced carotidartery specimens revealed an increase in expression of pDCmarkers in advanced versus early lesions (online-only Data
Supplement Table III). Accordingly, BDCA2� pDCs andLL37 were detected in advanced human carotid plaques butless frequently in early lesions (Figure 6A). Negative controlsshowed no staining (data not shown). This suggests that thepresence of pDCs correlates with plaque progression inhuman atherosclerosis. Furthermore, reciprocal serum dilu-tions to determine auto-antibodies directed against nuclearantigens revealed average titers of 1:190.0�99.1 in asymp-tomatic patients but 1:871.1�548.4 in symptomatic patients(n�8–9, P�0.1336). Moreover, elevated levels of circulatingantidsDNA antibodies were observed in patients with symp-tomatic compared with asymptomatic carotid artery stenosis(Figure 6B); age, risk factors, and medication were notdifferent between patient groups (online-only Data Supple-ment Table IV).
Figure 5. Anti–double-stranded DNA (antidsDNA) antibody generation correlates with atherosclerotic lesion formation. A and B, antids-DNA antibody titers in serum of apolipoprotein E–deficient (Apoe�/�) mice fed a high-fat diet (HFD) for 12 weeks (n�18) and in healthy6-week-old Apoe�/� mice (n�8 mice, A) and in serum of HFD-fed Apoe�/� mice and treated with immunoglobulin G (IgG), PDCA1,DOTAP (control), type-A oligonucleotides (CpGs), or CpGs�PDCA1 for 4 weeks (n�5–10 mice, B). C, AntidsDNA antibody titers inserum of Apoe�/� mice in low-density lipoprotein receptor–deficient (Ldlr�/�) mice reconstituted with Cramp�/� or Cramp�/� bonemarrow (n�7 each) and fed a HFD for 4 weeks. D, Aortic Oil-red O� atherosclerotic lesions in Ldlr�/� mice reconstituted withCramp�/� or Cramp�/� bone marrow and fed a HFD for 4 weeks. E, Atherosclerotic lesions in the aortic root in diet-fed Apoe�/� micetreated with PBS (control), Cramp, or Cramp together with DNAse for 3 weeks. F, Interferon � (IFN-�) protein secretion by sorted plas-macytoid dendritic cells (pDCs) stimulated overnight with PBS (control) and serum with low (�) or high (Œ) antidsDNA titers (n�5). Barsrepresent mean�SD. *P�0.05, **P�0.01. G, Immunoglobulin-G deposits (red) detected by immunofluorescence staining in the aorticroot in HFD-fed Apoe�/� mice. Cell nuclei are counterstained by hematoxylin (blue). Apoe�/� indicates apolipoprotein E deficient; HFD,high-fat diet; CpG, type-A oligonucleotide; Ldlr�/�, low-density lipoprotein receptor deficient; a-ds-DNA-Ab, anti-double stranded-DNAantibody.
Doring et al pDCs in Atherosclerosis 1679
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

DiscussionIn recent years, it has become evident that a number ofspecialized immune cell populations contribute to atherogen-esis. Besides monocytes/macrophages, several T-cell subsets,conventional DCs, and neutrophils have emerged as criti-cal players in atherosclerosis.1,17 The role of pDCs, how-ever, has remained elusive to date. Here we provide thefirst evidence that a pDC-dependent pathway of autoim-mune activation critically drives early atherosclerotic le-sion formation.
Plasmacytoid DCs have previously been found in carotidartery plaques, primarily in the plaque shoulder region.4 Inaddition, reduced circulating pDC counts have been describedin patients with coronary artery disease, which may corre-spond to their enhanced accumulation within atheroscleroticlesions.34 Accordingly, we observed an accumulation ofpDCs in murine and human atherosclerotic plaques but not inother secondary lymphoid organs (not shown) and an in-creased expression of pDC markers in advanced versus earlyhuman atherosclerotic carotid artery lesions.
Described as inflammatory amplifiers, plaque-residingpDCs were shown to respond to CpGs (containing motifstypically found in microbial DNA) with an enhanced IFN-�expression, which in turn amplified inflammatory TLR4,TNF- �, IL-12, and matrix metalloproteinase-9 expression bymyeloid DCs and cytolytic T-cell functions, and whichcorrelated with plaque instability.3,4 In order to scrutinize therole of pDCs in atherosclerosis, we employed a specificpDC-depleting antibody. In line with the notion that pDCsexert proinflammatory effects, we here demonstrate thatpDCs and their activation critically drive atheroscleroticlesion formation. Administration of a PDCA1 antibody todeplete pDCs protected from lesion formation. Conversely,stimulation of Apoe�/� mice with type A CpGs promotedplaque growth in a pDC-dependent manner, as revealed byabrogating CpG-induced lesion formation by the additionaldepletion of this cell type. Of note, treatment of Apoe�/�
mice with type-B CpGs, known to activate B cells rather thanpDCs, was previously shown to not affect atheroscleroticlesion formation.35 These findings, furthermore, corroboratethat activating pDCs rather than other cell types by type-ACpGs (used in our study) mediates enhanced plaque growth.Although reduced IFN- � serum levels could be observed inpDC-depleted mice, administration of IFN-� aggravatedlesion formation and macrophage plaque content in Apoe�/�
mice, extending findings that treatment with IFN-� or IFN-�accelerates atherosclerotic lesion formation and that IFN-�promotes macrophage accumulation in plaques.36,37 Plasma-cytoid DC–derived type-I IFNs can likewise also activate otherimmune cells, such as cDCs and B cells, to promote autoimmu-nity,38 which may also be relevant in atherosclerosis.
Plasmacytoid DCs have also been shown to exert tolero-genic functions; in a vascularized graft model, pDC depletionor prevention of pDC lymph-node homing inhibited regula-tory T-cell differentiation and tolerance induction whereasadoptive transfer of tolerized alloantigen-presenting pDCsprolonged graft survival.12 In the present study, we clearlydemonstrate that pDCs exert proatherogenic functions duringearly lesion formation.
The responsiveness of pDCs to viruses or bacterial infec-tions, as epitomized by treatment of Apoe�/� mice withCpGs, may corroborate the link between atherosclerosis andchronic infection burdens and possibly the inflammatoryactivation of vulnerable plaques in response to acute infec-tions.3 However, nucleotides released from necrotic orapoptotic cells may also be prone to binding TLR7/TLR9and to inducing IFN-� production by pDCs in the presenceof antimicrobial peptides released from inflammatorycells.39 For instance, self-DNA fragments released bydying cells (eg, in psoriasis) can be bound by Cramp, anantimicrobial peptide rapidly released from neutrophilgranules,6,28,29 to form complexes that trigger TLR7/TLR9activation and type-I IFN secretion.11,15 Similarly, NETs,
Figure 6. Plasmacytoid dentric cell(pDC) numbers and LL37 expression aswell as circulating anti–double-strandedDNA (antidsDNA) antibody titers increasein human atherosclerosis. A, BDCA2 andLL37 staining (brown) detected by immu-nohistochemistry in carotid arteries withearly (top) versus advanced lesions (bot-tom); cell nuclei are counterstained byhematoxylin (blue). B, Serum concentra-tion of antidsDNA antibodies in patientswith asymptomatic versus symptomaticcarotid artery stenosis (n�15 per group).Bars represent median with interquartilerange. **P�0.01. BDCA2 indicates bloodcell-derived antigen-2; and dsDNA-Ab,double-stranded DNA antibodies.
1680 Circulation April 3, 2012
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

web-like structures containing self-DNA and antimicrobialpeptides, can trigger pDC activation.10,11,30
These factors may also be present in atherosclerosisplaques. Extending findings in humans, where an increasedexpression of LL37 has been described in atheroscleroticcarotid artery plaques compared with normal arteries incolocalization with macrophages and endothelial cells,40 wehere observed increased Cramp expression in atheroscleroticApoe�/� arteries. Cramp protein was detected in colocaliza-tion with DNA fragments in plaque necrotic core areas,suggesting that Cramp/self-DNA complexes may form inatherosclerosis. Notably, injection of Cramp promoted ath-erosclerotic plaque growth dependent on the presence ofextracellular self-DNA. Thus, the formation of Cramp/self-DNA complexes may contribute to pDC activation andenhanced atherosclerosis, as exemplified by their capabilityof triggering IFN-� secretion by pDCs in vitro and theirpDC-dependent acceleration of lesion formation in Apoe�/�
mice in vivo. Cramp may be released by neutrophils recruitedduring early lesion development,17,29,41 as implied by itsdetection in the vicinity of segment-nucleated neutrophils inplaques. In addition, we unprecedentedly observed the for-mation of NETs in atherosclerotic carotid arteries. TheseCramp/self-DNA complexes contained in neutrophil-expelled NETs may further trigger pDC activation.10,11
Cytokines including IFN-� may reciprocally prime neutro-phils to allow the formation of NETs,42 thus drivingpositive feedback amplification.
Interestingly, oxLDL enhanced phagocytosis of pDCs andtheir capacity to prime antigen-specific T-cell responseswhile not directly contributing to IFN-� responses or co-stimulatory molecule upregulation. Exposure to oxLDLwithin plaques1,2 may thus boost uptake of antigenic com-plexes by pDCs and their activation in atherosclerosis.
Interestingly, diminished antidsDNA antibody titers wereobserved in Ldlr�/� mice reconstituted with Cramp�/� ver-sus Cramp�/� bone marrow, together with a protection fromatherosclerotic lesion formation. Moreover, antidsDNA anti-body titers but not antibodies toward MDA- or oxLDL-derived antigens were found to be decreased in pDC-depletedmice whereas antidsDNA antibody titers were enhanced inApoe�/� mice stimulated with CpGs or treated with IFN-�.These data intriguingly suggest that Cramp-dependentbreakdown of tolerance to self-DNA may stimulate pDCsand IFN-� production, contributing to the formation ofantidsDNA antibodies. This hypothesis may also be in linewith findings that IFN-� supports humoral immunity (eg,by enabling B cells to undergo isotype switching and tomature into antigen-secreting cells).32 The comparativelylow levels of such autoimmune activity may be related toa locally confined pattern of this mechanism. PrototypicalpDC stimulation and large amounts of IFN-� in CpG-treated Apoe�/� mice may in turn explain increasedantibodies also toward other known antigens in atheroscle-rosis, namely MDA or oxLDL. It remains to be determinedif self-DNA complexed to Cramp may in addition alsodirectly active B cells via TLR9, which can be similarlyenhanced by IFN-�.38
In line with previous work, we have detected the presenceof IgG deposits within atherosclerotic arteries.43 Likewise,antidsDNA antibodies may also be deposited within plaques.It was recently shown that these immune complexes containself-DNA as well as antimicrobial peptides and can effec-tively trigger pDC activation and IFN-� production.7–11,31 Inaccordance with this, serum of Apoe�/� mice containing highlevels of antidsDNA antibody titers but not serum containinglow titers significantly increased IFN-� production in pDCsin vitro whereas serum derived from atherosclerotic Rag1�/�
Ldlr�/� mice did not elicit IFN-� responses in pDCs. Thus,antidsDNA antibodies, generated as a consequence of pDCactivation, may contribute to the pathogenic insult inatherosclerosis.
Although values may be of limited clinical relevance in arheumatological context, increased levels of circulating an-tidsDNA antibodies in atherosclerotic Apoe�/� mice, as wellas in patients with symptomatic and more advanced athero-sclerosis, may support a pathogenic role of antidsDNAantibodies. Importantly, elevated anti-nuclear antibody titerswere associated with decreased carotid elasticity in youngFinns and were postulated to participate in the developmentof early atherosclerosis.44 Moreover, in an animal model ofsystemic lupus erythematosus with enhanced antibody titersagainst dsDNA, atherosclerotic lesion formation wasaccelerated.45
Chronically increased IFN-� levels and circulating antids-DNA antibody titers in patients with psoriasis and systemiclupus erythematosus6 may likewise predispose for an in-creased risk to develop hyperlipidemic atherosclerosis.46
Collectively, our data provide evidence for a hithertounrecognized pathway of autoimmune activation in athero-sclerosis, driven by pDCs that sense self-DNA fragmentscomplexed to Cramp and contribute to the formation of anti-nuclear antibodies. Exposure to oxLDL during atherogenesismay amplify the immunogenicity of pDCs by enhancing phago-cytosis and priming of antigen-specific T-cell responses. Target-ing pDC activation may thus constitute a promising approach tolimit atherosclerotic lesion development.
AcknowledgmentsWe thank Melanie Garbe, Yvonne Jansen, Theresa Moritz, SilviaRoubrocks, Susanne Schmitz, Melanie Schott, Roya Soltan, andStephanie Wilbertz for excellent technical assistance.
Sources of FundingThis work was supported by the Deutsche Forschungsgemein-schaft (FOR809, ZE827/1-2 to Dr Zernecke and Dr Zenke,SFB688 TPA12 and ZE827/4-1 to Dr Zernecke), the LeducqTransatlantic Network of Excellence CVGeneF(x) to Dr Weber,FöFoLe program of the Medical Faculty, LMU Munich to Dr.Soehnlein, and the Interdisciplinary Centre for Clinical ResearchBIOMAT within the Faculty of Medicine at the RWTH AachenUniversity (VV-B112c) to Dr Zernecke.
DisclosuresNone.
References1. Weber C, Zernecke A, Libby P. The multifaceted contributions of leu-
kocyte subsets to atherosclerosis: lessons from mouse models. Nat RevImmunol. 2008;8:802–815.
Doring et al pDCs in Atherosclerosis 1681
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

2. Galkina E, Ley K. Immune and inflammatory mechanisms of atheroscle-rosis. Annu Rev Immunol. 2009;27:165–197.
3. Niessner A, Sato K, Chaikof EL, Colmegna I, Goronzy JJ, Weyand CM.Pathogen-sensing plasmacytoid dendritic cells stimulate cytotoxic T-cellfunction in the atherosclerotic plaque through interferon-alpha. Circulation.2006;114:2482–2489.
4. Niessner A, Shin MS, Pryshchep O, Goronzy JJ, Chaikof EL, WeyandCM. Synergistic proinflammatory effects of the antiviral cytokineinterferon-alpha and Toll-like receptor 4 ligands in the atheroscleroticplaque. Circulation. 2007;116:2043–2052.
5. Colonna M, Trinchieri G, Liu YJ. Plasmacytoid dendritic cells inimmunity. Nat Immunol. 2004;5:1219–1226.
6. Gilliet M, Cao W, Liu YJ. Plasmacytoid dendritic cells: sensing nucleicacids in viral infection and autoimmune diseases. Nat Rev Immunol.2008;8:594–606.
7. Lande R, Gregorio J, Facchinetti V, Chatterjee B, Wang YH, Homey B,Cao W, Wang YH, Su B, Nestle FO, Zal T, Mellman I, Schroder JM, LiuYJ, Gilliet M. Plasmacytoid dendritic cells sense self-DNA coupled withantimicrobial peptide. Nature. 2007;449:564–569.
8. Lovgren T, Eloranta ML, Bave U, Alm GV, Ronnblom L. Induction ofinterferon-alpha production in plasmacytoid dendritic cells by immunecomplexes containing nucleic acid released by necrotic or late apoptoticcells and lupus IgG. Arthritis Rheum. 2004;50:1861–1872.
9. Means TK, Latz E, Hayashi F, Murali MR, Golenbock DT, Luster AD.Human lupus autoantibody-DNA complexes activate DCs through coop-eration of CD32 and TLR9. J Clin Invest. 2005;115:407–417.
10. Garcia-Romo GS, Caielli S, Vega B, Connolly J, Allantaz F, Xu Z,Punaro M, Baisch J, Guiducci C, Coffman RL, Barrat FJ, Banchereau J,Pascual V. Netting neutrophils are major inducers of type I IFN produc-tion in pediatric systemic lupus erythematosus. Sci Transl Med. 2011;3:73ra20.
11. Lande R, Ganguly D, Facchinetti V, Frasca L, Conrad C, Gregorio J,Meller S, Chamilos G, Sebasigari R, Riccieri V, Bassett R, Amuro H,Fukuhara S, Ito T, Liu YJ, Gilliet M. Neutrophils Activate PlasmacytoidDendritic Cells by Releasing Self-DNA-Peptide Complexes in SystemicLupus Erythematosus. Sci Transl Med. 2011;3:73ra19.
12. Ochando JC, Homma C, Yang Y, Hidalgo A, Garin A, Tacke F, AngeliV, Li Y, Boros P, Ding Y, Jessberger R, Trinchieri G, Lira SA, RandolphGJ, Bromberg JS. Alloantigen-presenting plasmacytoid dendritic cellsmediate tolerance to vascularized grafts. Nat Immunol. 2006;7:652–662.
13. Shen H, Iwasaki A. A crucial role for plasmacytoid dendritic cells inantiviral protection by CpG ODN-based vaginal microbicide. J ClinInvest. 2006;116:2237–2243.
14. Krug A, French AR, Barchet W, Fischer JA, Dzionek A, Pingel JT,Orihuela MM, Akira S, Yokoyama WM, Colonna M. TLR9-dependentrecognition of MCMV by IPC and DC generates coordinated cytokineresponses that activate antiviral NK cell function. Immunity. 2004;21:107–119.
15. Gregorio J, Meller S, Conrad C, Di Nardo A, Homey B, Lauerma A, AraiN, Gallo RL, Digiovanni J, Gilliet M. Plasmacytoid dendritic cells senseskin injury and promote wound healing through type I interferons. J ExpMed. 2010;207:2921–2923.
16. Nizet V, Ohtake T, Lauth X, Trowbridge J, Rudisill J, Dorschner RA,Pestonjamasp V, Piraino J, Huttner K, Gallo RL. Innate antimicrobialpeptide protects the skin from invasive bacterial infection. Nature. 2001;414:454–457.
17. Zernecke A, Bot I, Djalali-Talab Y, Shagdarsuren E, Bidzhekov K,Meiler S, Krohn R, Schober A, Sperandio M, Soehnlein O, Bornemann J,Tacke F, Biessen EA, Weber C. Protective role of CXC receptor 4/CXCligand 12 unveils the importance of neutrophils in atherosclerosis. CircRes. 2008;102:209–217.
18. Faust N, Varas F, Kelly LM, Heck S, Graf T. Insertion of enhanced greenfluorescent protein into the lysozyme gene creates mice with greenfluorescent granulocytes and macrophages. Blood. 2000;96:719–726.
19. Van Rooijen N, Sanders A. Liposome mediated depletion of macro-phages: mechanism of action, preparation of liposomes and applications.J Immunol Methods. 1994;174:83–93.
20. Lutgens E, Lievens D, Beckers L, Wijnands E, Soehnlein O, Zernecke A,Seijkens T, Engel D, Cleutjens J, Keller AM, Naik SH, Boon L, OufellaHA, Mallat Z, Ahonen CL, Noelle RJ, de Winther MP, Daemen MJ,Biessen EA, Weber C. Deficient CD40-TRAF6 signaling in leukocytesprevents atherosclerosis by skewing the immune response toward anantiinflammatory profile. J Exp Med. 2010;207:391–404.
21. Binder CJ, Hartvigsen K, Chang MK, Miller M, Broide D, Palinski W,Curtiss LK, Corr M, Witztum JL. IL-5 links adaptive and natural
immunity specific for epitopes of oxidized LDL and protects from ath-erosclerosis. J Clin Invest. 2004;114:427–437.
22. Weber C, Meiler S, Doring Y, Koch M, Drechsler M, Megens RT,Rowinska Z, Bidzhekov K, Fecher C, Ribechini E, van Zandvoort MA,Binder CJ, Jelinek I, Hristov M, Boon L, Jung S, Korn T, Lutz MB,Forster I, Zenke M, Hieronymus T, Junt T, Zernecke A. CCL17-expressing dendritic cells drive atherosclerosis by restraining regulatory Tcell homeostasis in mice. J Clin Invest. 2011;121:2898–2910.
23. Faber BC, Cleutjens KB, Niessen RL, Aarts PL, Boon W, Greenberg AS,Kitslaar PJ, Tordoir JH, Daemen MJ. Identification of genes potentiallyinvolved in rupture of human atherosclerotic plaques. Circ Res. 2001;89:547–554.
24. Paulson KE, Zhu SN, Chen M, Nurmohamed S, Jongstra-Bilen J,Cybulsky MI. Resident intimal dendritic cells accumulate lipid andcontribute to the initiation of atherosclerosis. Circ Res. 2010;106:383–390.
25. Nickel T, Schmauss D, Hanssen H, Sicic Z, Krebs B, Jankl S, Summo C,Fraunberger P, Walli AK, Pfeiler S, Weis M. oxLDL uptake by dendriticcells induces upregulation of scavenger-receptors, maturation and differ-entiation. Atherosclerosis. 2009;205:442–450.
26. Kunjathoor VV, Febbraio M, Podrez EA, Moore KJ, Andersson L, KoehnS, Rhee JS, Silverstein R, Hoff HF, Freeman MW. Scavenger receptorsclass A-I/II and CD36 are the principal receptors responsible for theuptake of modified low density lipoprotein leading to lipid loading inmacrophages. J Biol Chem. 2002;277:49982–49988.
27. Barchet W, Cella M, Colonna M. Plasmacytoid dendritic cells: virusexperts of innate immunity. Semin Immunol. 2005;17:253–261.
28. Sorensen O, Arnljots K, Cowland JB, Bainton DF, Borregaard N. Thehuman antibacterial cathelicidin, hCAP-18, is synthesized in myelocytesand metamyelocytes and localized to specific granules in neutrophils.Blood. 1997;90:2796–2803.
29. Soehnlein O, Zernecke A, Weber C. Neutrophils launch monocyte extrav-asation by release of granule proteins. Thromb Haemost. 2009;102:198–205.
30. Brinkmann V, Reichard U, Goosmann C, Fauler B, Uhlemann Y, WeissDS, Weinrauch Y, Zychlinsky A. Neutrophil extracellular traps killbacteria. Science. 2004;303:1532–1535.
31. Munoz LE, Lauber K, Schiller M, Manfredi AA, Herrmann M. The roleof defective clearance of apoptotic cells in systemic autoimmunity. NatRev Rheumatol. 2010;6:280–289.
32. Le Bon A, Schiavoni G, D’Agostino G, Gresser I, Belardelli F, ToughDF. Type i interferons potently enhance humoral immunity and canpromote isotype switching by stimulating dendritic cells in vivo.Immunity. 2001;14:461–470.
33. Giordani L, Sanchez M, Libri I, Quaranta MG, Mattioli B, Viora M.IFN-alpha amplifies human naive B cell TLR-9-mediated activation andIg production. J Leukoc Biol. 2009;86:261–271.
34. Van Vre EA, Hoymans VY, Bult H, Lenjou M, Van Bockstaele DR,Vrints CJ, Bosmans JM. Decreased number of circulating plasmacytoiddendritic cells in patients with atherosclerotic coronary artery disease.Coron Artery Dis. 2006;17:243–248.
35. Khallou-Laschet J, Tupin E, Caligiuri G, Poirier B, Thieblemont N,Gaston AT, Vandaele M, Bleton J, Tchapla A, Kaveri SV, Rudling M,Nicoletti A. Atheroprotective effect of adjuvants in apolipoprotein Eknockout mice. Atherosclerosis. 2006;184:330–341.
36. Levy Z, Rachmani R, Trestman S, Dvir A, Shaish A, Ravid M, Harats D.Low-dose interferon-alpha accelerates atherosclerosis in an LDLreceptor-deficient mouse model. Eur J Intern Med. 2003;14:479–483.
37. Goossens P, Gijbels MJ, Zernecke A, Eijgelaar W, Vergouwe MN, vander Made I, Vanderlocht J, Beckers L, Buurman WA, Daemen MJ,Kalinke U, Weber C, Lutgens E, de Winther MP. Myeloid type Iinterferon signaling promotes atherosclerosis by stimulating macrophagerecruitment to lesions. Cell Metab. 2010;12:142–153.
38. Banchereau J, Pascual V. Type I interferon in systemic lupus erythema-tosus and other autoimmune diseases. Immunity. 2006;25:383–392.
39. Marshak-Rothstein A, Busconi L, Rifkin IR, Viglianti GA. The stimu-lation of Toll-like receptors by nuclear antigens: a link between apoptosisand autoimmunity. Rheum Dis Clin North Am. 2004;30:559–574.
40. Edfeldt K, Agerberth B, Rottenberg ME, Gudmundsson GH, Wang XB,Mandal K, Xu Q, Yan ZQ. Involvement of the antimicrobial peptideLL-37 in human atherosclerosis. Arterioscler Thromb Vasc Biol. 2006;26:1551–1557.
41. Drechsler M, Megens RT, van Zandvoort M, Weber C, Soehnlein O.Hyperlipidemia-triggered neutrophilia promotes early atherosclerosis.Circulation. 2010;122:1837–1845.
1682 Circulation April 3, 2012
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

42. Martinelli S, Urosevic M, Daryadel A, Oberholzer PA, Baumann C, FeyMF, Dummer R, Simon HU, Yousefi S. Induction of genes mediatinginterferon-dependent extracellular trap formation during neutrophil dif-ferentiation. J Biol Chem. 2004;279:44123–44132.
43. Vlaicu R, Niculescu F, Rus HG, Cristea A. Immunohistochemical local-ization of the terminal C5b-9 complement complex in human aorticfibrous plaque. Atherosclerosis. 1985;57:163–177.
44. Pertovaara M, Kahonen M, Juonala M, Laitinen T, Taittonen L, Leh-timaki T, Viikari JS, Raitakari OT, Hurme M. Autoimmunity and ath-
erosclerosis: the presence of antinuclear antibodies is associated withdecreased carotid elasticity in young women: the Cardiovascular Risk inYoung Finns Study. Rheumatology. 2009;48:1553–1556.
45. Braun N, Wade NS, Wakeland EK, Major AS. Accelerated athero-sclerosis is independent of feeding high fat diet in systemic lupuserythematosus-susceptible LDLr(�/�) mice. Lupus. 2008;17:1070 –1078.
46. Frostegard J. Systemic lupus erythematosus and cardiovascular disease.Lupus. 2008;17:364–367.
CLINICAL PERSPECTIVEAltherosclerosis remains the number 1 cause of death in the Western world, and the therapeutic options currently availableare limited. Chronic inflammation of the vessel wall has been closely linked to autoimmunogenic processes inatherosclerosis. The role of the specialized subset of plasmacytoid dendritic cells present in human atherosclerotic plaques,however, has not been addressed previously. We provide evidence for a hitherto unrecognized plasmacytoid dendriticcell–driven pathway of autoimmune activation in atherosclerosis that critically amplifies early atherosclerotic lesionformation. Self-DNA (eg, released from dying cells or in neutrophil extracellular traps) and an increased expression of theantimicrobial peptide Cramp/LL37 in atherosclerotic lesions was shown to stimulate breakdown of tolerance to self-DNAand promote interferon-� production by plasmacytoid dendritic cells, aggravating early atherosclerosis and the formationof anti–double-stranded DNA antibodies in apolipoprotein E–deficient mice. Notably, anti–double-stranded DNAantibodies were also found to be elevated in patients with symptomatic versus asymptomatic carotid artery stenosis.Although possibly of limited clinical relevance in a rheumatological context, moderately increased levels of circulatinganti–double-stranded DNA antibodies may predispose to atherosclerosis, in line with findings showing elevatedanti-nuclear antibody titers to be associated with decreased carotid elasticity and early atherosclerosis. More importantly,chronically increased interferon-� levels and circulating anti–double-stranded DNA antibody titers may provide anexplanation for the increased risk of atherosclerosis well recognized in patients with psoriasis and systemic lupuserythematosus.
Doring et al pDCs in Atherosclerosis 1683
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

SUPPLEMETAL MATERIAL
Supplemental Methods
Mouse Models
Apoe-/-, Ldlr-/- and Rag1-/- and C57BL/6J wild type mice were purchased from the
Jackson Laboratory. Six to eight weeks old female Apoe-/- mice were fed an atherogenic
diet (21% fat, 0.15% cholesterol, Altromin). For in vivo stimulation of pDCs, mice were
injected with the oligonucleotide (ODN) CpG 1585 (25 µg/mouse, Inovagen) complexed
to DOTAP (15 µg/mouse, Roche) in 200 µl PBS, or with DOTAP or PBS (Gibco) alone
3 times/week for 4 weeks. Efficiency of pDC stimulation was confirmed by determining
IFNα concentrations in serum (data not shown). pDC depletion in Apoe-/- mice was
performed by intravenous injection of the monoclonal antibody PDCA-11, 2 (500
µg/mouse, Miltenyi Biotech) or an isotype control antibody (500 µg/mouse, BioXcell) at
day 1 and day 7 after the initiation of high fat diet feeding. Some Apoe-/- mice were
injected with murine IFNα (100,000 U, HyCult Biotec) twice a week i.p. for 4 weeks.
Some Apoe-/- mice were i.v. injected with Cramp/DNA complexes* (100 μg Cramp,
Innovagen, mixed with 20 µg of DNA, ODNs, Integrated DNA technologies, in 200 μl of
PBS/mouse) 3 times/week for 4 weeks, together with receiving PDCA1 or isotype control
antibody (see injection schema below).
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

2
Six to eight weeks old female Ldlr-/- mice (Jackson laboratory) were irradiated and
reconstituted with Cramp-/-3 or Cramp+/+ bone marrow as described4. Briefly, bone
marrow cells (5×106 in PBS) were administered to Ldlr-/- mice by intravenous tail vein
injection 24 h after an ablative dose of whole-body irradiation (2×6.5 Gy). After
recovery, transplanted Ldlr-/- mice were placed on high fat diet for 4 weeks. Another
group of female Apoe-/- mice was treated with Cramp (i.p., 100 µg/mouse) with or
without DNase I (i.v., 120 U/mouse, Sigma Aldrich) 3 times/week for 3 weeks. No
changes in cholesterol or triglyceride serum levels, as assessed by EnzyChromTM HDL
and LDL/VLDL Assay Kit (BioAssay Systems), were observed between groups of mice
(Supplemental Table 3). Rag1-/- mice were crossed with Ldlr-/- mice (Rag1-/-Ldlr-/- mice)
and fed an atherogenic diet for 12 weeks. Lysozyme M (lys)-enhanced green fluorescent
protein (EGFP) mice5, kindly provided by Dr. T. Graf (Albert Einstein College, New
York), were i.v. injected with 250 μl of liposomes containing clodronate6 one day and PI
(2µg/ml final concntration) immediately before imaging. Efficient monocyte depletion
and the identification of remaining circulating EGFP+ cells as SSChi CD115- CD11b+ Gr-
1+ neutrophils was confirmed by FACS analysis (data not shown). Experiments were
approved by local authorities (Landesamt für Natur, Umwelt und Verbraucherschutz
Nordrhein-Westfalen, Recklinghausen, Germany) to comply with German animal
protection law.
Atherosclerotic lesion quantification and immunohistochemistry
The extent of atherosclerosis was assessed in aortic roots and in aortas by staining for
lipid depositions with oil-red-O 4 and quantified by computerized image analysis (Diskus
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

3
Software, Hilgers, Königswinter) and Leica Qwin Imaging software (Leica Ltd,
Cambridge, UK). Briefly, atherosclerotic lesions were quantified in 5 μm transverse
sections through the heart and the aortic roots. The thoraco-abdominal aorta was opened
longitudinally, and the percentage of lipid deposition was calculated by dividing the
stained area by total aortic surface. The relative content of macrophages was determined
by mAb staining for MOMA-2 (MCA519, AbD Serotec) or Mac-2 (M3/38, Cedarlane).
pDCs were detected by staining for Siglec-H (440c, HyCult Biotech). A rabbit anti-
Cramp antibody (Innovagen) was used to detect Cramp on acetone fixed fresh frozen
sections. Plaque IgG was detected using a biotinylated rabbit anti-mouse IgG (Vector
Laboratories) following by a streptavidin-conjugated Alexafluor-555 (Molecular Probes)
on 4% paraformaldehyde-fixed frozen sections. Nuclei were counter-stained by 4',6-
Diamidino-2-phenylindol (DAPI). Images were recorded with a Leica DMLB
fluorescence microscope and CCD camera. Human carotid artery specimens containing
all stages of atherosclerosis (from intimal xanthoma to fibrous cap atheroma with plaque
rupture) were obtained from autopsy or vascular surgery (IMCAR tissue collection; n=3-
6 per plaque type). Atherosclerotic plaques were fixed o/n in 4% paraformaldehyde,
processed, embedded in paraffin and sectioned (4 μm). The location and intensity of
BDCA2 (mAB, Clone AC144, Miltenyi Biotec) and LL37 (mAB, Clone 3D11, Hycult
Biotech) expression in the plaque was determined using standard immunohistochemistry
techniques7.
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

4
pDC isolation and stimulation in vitro
pDCs were isolated from spleens of C57BL/6J mice using the Plasmacytoid Dendritic
Cell Isolation Kit II (Miltenyi Biotec) according to the manufactures’ protocol.
Alternatively pDCs were sorted (FACSAria, BD Bioscience) from bone marrow of
C57BL/6J with an antibody cocktail against CD45, SiglecH and CD45RA (B220). Purity
of isolated pDCs was consistently above >90-95% (Miltenyi) and >90% (sorting), as
assessed by FACS analysis (data not shown). Isolated pDCs were seeded at 1 x 105
cells/200µl into flat bottom 96-well plates in RPMI-1640 medium supplemented with
10% FCS, L-glutamine (2 mM), penicillin/streptomycin (100 U/ml), β-mercaptoethanol
(50 µM, Sigma) and 25 ng/ml FLT3-L (Peprotech) and were stimulated with Cramp (50
µg/ml, Innovagen), DNA (10 µg/ml µg, mouse C57Bl6/6J female genomic DNA,
Zyagen, or ODNs8, Integrated DNA technologies), CpG 1585 (5µg/ml, Invivogen),
oxLDL or di-oxLDL (50 µg/ml), and 4 µl of mouse serum containing high (36 Units/µl)
or low (1 Unit/µl) titers of anti-dsDNA antibodies per well, and serum obtained from
Rag1-/-Ldlr-/- mice. For the generation of complexes, 10 μg of Cramp was mixed with 2
µg self-DNA in 20 μl of PBS and incubated for 15 min at room temperature9.
Antigen-specific T-cell proliferation in vivo
CD4+ T cells were isolated from the spleen of OT-II mice by negative selection of CD4+
T cells (CD4 T cell isolation kit, Miltenyi), and labeled with CFSE (Fluka). 1x105 pDCs
were pre-stimulated with oxLDL (50 µg/ml), CpG 1585 (5 mg/ml) or in combination
with both and were pulsed for 45 min with OVA323-339 peptide (30ug/ml)
(ISQAVHAAHAEINEAGR, OVA-2, ANASPEC) for MHC class II-restricted T-cell
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

5
stimulation. Untreated pDCs were used as control. For in vivo studies, 5×106 CD4+
CFSE-labeled OT-II cells were injected into the tail vein of C57BL/6 recipient mice. One
day later, 1×105 untreated or treated OVA-2 peptide-pulsed pDCs were injected into the
footpad. After 3 days, single-cell suspensions were prepared from peripheral LNs of
recipient mice to analyze T-cell proliferation by CFSE dye dilution by flow cytometry.
Enzymatic tissue digestion and flow cytometry
For FACS analysis of tissues: lymph nodes and spleen were dissociated into single-cell
suspensions using collagenase D (Roche) for 30 min at 37°C. Aortas were excised,
flushed in PBS and digested using Liberase III (Roche). Staining for flow cytometric
analysis was conducted using combinations of antibodies from eBiosciences (CD11c,
CD11b, CD115, B220, CD8a, 440c, CD36, CD68, SR-A, CD80, CD86, CD40, CD3,
CD4, CD45), Miltenyi (PDCA-1) and BD Bioscience (MHC II, Gr-1) in HBSS with 0.3
mM EDTA and 0.1% BSA. Data were acquired using a FACSCanto II (BD Bioscience),
and results analyzed by FlowJo software (Tree Star).
Quantitative real time-polymerase chain reaction (PCR)
RNA was isolated from aortas with the ZR RNA MicroPrep RNA isolation kit (Zymo
Research) according to manufacturer’s protocols. cDNA was reverse transcribed from 2
µg of DNAse-treated total RNA (Promega). Real time-PCR analysis was performed
using SYBRGreen (Fermentas) and specific primer pairs (Sigma) according to
manufacturer’s protocols:
Ifn-α:5'- TCTGATGCAGCAGGTGGG -3', 5'-AGGGCTCTCCAGACTTCTGCTCTG-3';
SiglecH:5'- GCTGGGATGCTGCTGTCCCG-3',5'- GGTGGCAGGGCACAAGGACA -3';
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

6
Cramp:5'-TCCCAAGTCTGTGAGGTTCC-3', 5'- CCCATACACTGCTTCACCAC -3',
Gapdh:5'-CCATCACCATCTTCCAGGAG-3',5'-GTGGTTCACACCCATCACAA-3'.
Amplification (45 cycles, annealing at 58°C) was performed in duplicate using a 7900
HT fast real time PCR system (Applied Biosystems, USA).
Two-photon microscopy
In vivo two-photon microscopy of exposed carotid arteries was performed as described10.
In short, the exposed left carotid artery was visualized using an upright Olympus
FV1000MPE two-photon microscope system equipped with a 1.05; water dipping
objective (Olympus GMBH). A pulsed Ti-Sapphire laser (MaiTai DeepSee, Spectra
Physics) was tuned to 810 nm for visualization of second Harmonic Generation (SHG)
signal of collagen, GFP, and PI. Emitted fluorescent signals were detected with three
photomultiplier tubes (PMTs) tuned to corresponding parts of the emission spectra: SHG,
400-470 nm (PMT 1); GFP, 500-550 nm (PMT 2); PI, 560-660 nm (PMT 3);
autofluorescent signals from elastin bands was detected mainly in PMT 2 and PMT 3.
Series of subsequent xy-sections (256x256 pixels) were obtained over time at an
acquisition rate of 4Hz. Only comparable xy-sections without severe motion artefacts
(due to heart- and respiration cycle11, were selected for further analyses. Overall image
quality was improved by 2D Gaussian weigthed filtering11 by resizing the pixel matrix
two times (bilinear scaling). All image processing was performed using Image-Pro 3D
analyzer 7.0 (Media Cybernetics).
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

7
ELISA
Cell culture supernatants or mouse sera were analyzed by ELISA detecting IFNα
(VeriKineTM Mouse Interferon-Alpha ELISA Kit, PBL laboratories), and double stranded
DNA (Mouse anti-dsDNA Kit, Alpha Diagnostic) according to the manufacture’s
protocol. Anti-dsDNA antibodies in human serum were analyzed by Varelisa dsDNA
Antibody ELISA (Phadia); anti-dsDNA Ab (+) >55 IU/mL. Reciprocal serum dilutions to
determine autoantibodies directed against nuclear antigens (ANAs) in humans were
performed by indirect immunofluorescence on HEp-2 substrate according to the
manufacturer’s instructions (ANA IFT IgG Kit, Viramed). Positive ANA titers (>1:160)
were revealed to be present in 25% of asymptomatic subjects, but 56% of symptomatic
patients. In all cases of positive titers, either homogeneous or speckled staining patterns
could be observed, in line with the recognition of nuclear antigens12. Antibodies to
malondialdeyhde-modified (MDA)-LDL and oxLDL were measured by
chemiluminescent ELISA13. Briefly, sera were diluted 1:100 and binding of IgG to
MDA-LDL and oxLDL were measured using AP-conjugated rat-anti-mouse IgG,
followed by detection with LumiPhos.
Human microarray data evaluation
17 atherosclerotic tissue samples from atherosclerotic carotid artery segments, 9 from
early and 8 from advanced lesions, have been retrieved from the Maastricht Pathology
Tissue Collection (MPTC). Tissue was obtained during autopsy (Dept of Pathology,
Maastricht University Medical Centre) and extracted from carotid arteries, as during
carotid endarterectomy procedures. Collection, storage in the Maastricht Pathology
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

8
Tissue Collection (MPTC) and use of tissue and patient data were performed in
agreement with the "Code for Proper Secondary Use of Human Tissue"
(http://www.federa.org/?s=1&m=99). Samples were processed as described previously14.
In brief, carotid artery specimens were divided into parallel sections for RNA isolation
and histological analysis immediately after resection. Carotid artery segments were snap
frozen for RNA isolation and adjacent sections were fixed and embedded in paraffin.
Based on 4 µm haematoxylin-eosin stained sections, segments were classified according
to Virmani et al.15. Total RNA was isolated using the guanidine isothiocyanate/CsCl16
method followed by further purification and concentration using RNease mini columns
(Qiagen). RNA quantity and quality were determined using a nanodrop spectrometer
(Witec AG) and a 20100 Bioanalyzer (Agilent Technologies) respectively. Only samples
with a RNA Integrity Number of at least 5 were further processed (mean RIN=7.8).
Double stranded cDNA was synthesized from about 2 µg of total RNA using the One-
Cycle Target Labeling Kit (Affymetrix), and used as a template for the preparation of
biotin-labeled cRNA using the GeneChip IVT Labeling Kit (Affymetrix). Biotin-labeled
cRNA was hybridized in duplicate (15 hybrids from early, 14 hybrids from advanced
samples) to the Human Genome U133 Plus 2.0 Array (Affymetrix), washed, stained with
phycoerythrin-streptavidin conjugate (Molecular Probes) and the signals were amplified
by staining with biotin-labeled anti-streptavidin antibody (Vector Laboratories) followed
by phycoerythrin-streptavidin. Arrays were laser scanned with the GeneChip Scanner
3000 (Affymetrix) according to the manufacturer's instructions. Data were saved as raw
images and quantified using GCOS 1.2 (Affymetrix). The resulting .CEL files have been
processed in R (R Development Core17. Uploading, background adjustment and quantile
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

9
normalization have been performed by affy18 according to the vignette's instruction. The
resulting expression set has been analyzed by limma19, combining the technical replicates
and treating patients IDs as random factors. After Bayesian smoothing of the standard
errors of the estimated log-fold changes, the transcripts of interest have been identified
within the top list and are reported as reported as log2 fold changes, average expression,
and Benjamin-Hochberg false discovery rate adjusted p-values. Housekeeping genes
(HPS90 and ACTB) were unaltered in advanced versus early plaques (HSP90AB6P, fold
change 0.990109516457994, adj.p.val 0,150944030384547; HSP90B1, fold change
0.97436370698835, adj.p.val 0,627452445273284; HSP90AA1, fold change
1.02995157375258, adj.p.val 0,724731894920322; ACTB, fold change
1.00904889957475, adj.p.val 0,915087042539356). Data have been uploaded to the GEO
database (accession: http://www.ncbi.nlm.nih.gov/geo, GSE28829).
Study patients
A total of 30 patients with advanced carotid artery stenosis (>70%, determined by
measuring the peak systolic velocity by ultrasonography) were included in this study. All
subjects gave informed consent and the study was approved by an institutional review
committee. Asymptomatic patients (n=15) without neurological symptoms were
diagnosed within the scope of a clinical work-up in consequence of arterial hypertension
and/or diabetes mellitus type 2 or by random screenings at the Vascular Centre of the
Technical University Munich; Symptomatic patients (n=15) displaying neurological
symptoms (transient ischaemic attack or non-disabling stroke) were assigned from the
Institute of Neurology of the hospital Klinikum rechts der Isar, Munich. Patients with
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

10
chronic kidney disease (eGFR < 60 ml min−1), diabetes mellitus, autoimmune disease,
and ongoing steroid or other immunosuppressive medication were excluded from the
study. Blood sampling was performed within two days prior to surgical intervention by
vein puncture. Venous blood was centrifuged at 2,000 x g for 10 minutes at 20°C and
serum stored at -70°C until analysis.
Statistics of Supplemental Data
Unpaired Student t tests were used to compare dsDNA-antibody titers between control
and IFN-α-treated Apoe-/- mice. Mann Whitney tests were applied to compare pDC
frequencies between isotype and PDCA1-treated mice, plaque size and macrophage
content between IgG and PDCA1, and PBS and IFN-α-treated Apoe-/- mice. ANOVA 1-
way analysis of variance followed by Tukey's multiple comparison post tests were used
for comparing plaque size between control, CpG and CpG+PDCA1-treated Apoe-/- mice,
IFN-α secretion in vitro and in vivo, and IgG antibody titers to MDA- and CuoxLDL.
Kruskal-Wallis tests were applied to compare IFN-α secretion by pDCs co-cultured with
T cells in vitro, macrophage content between controls, CpG and CpG+PDCA1-treated
Apoe-/- mice. Differences where P<0.05 were considered to be statistically significant
(Prism 4.0 software, GraphPad).
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

11
Supplemental Table 1
Serum lipid levels in experimental groups of mice.
cholesterol (mg/dL) triglycerides (mg/dL) n
Apoe-/-
IgG 1145.0±233.6 335.4±93.5 4
PDCA1 1047.0±299.9 275.9±71.1 5
Control 1157.0±108.1 311.3±51.9 3
CpG 1110.0±69.3 338.0±82.1 4
PDCA1 + CpG 1025.0±119.5 342.6±77.8 4
Control 1173.0±141.9 285.0±52.4 6
IgG + Cramp/DNA 1060.0±124.6 334.1±124.8 10
PDCA1+ Cramp/DNA 1187.0 ±160.7 364.0±65.0 10
Control 794.7±102.8 208.3±56.3 4
Cramp 718.6±118.2 181.3±41.2 6
Cramp+DNase 709.7±94.2 196.8±37.9 10
Cramp+/+►Ldlr-/- 259.3±95.6 49.7±15.6 4
Cramp-/-►Ldlr-/- 266.0±31.5 36.2±28.5 4
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

12
Supplemental Table 2
Quantification of luminal PI+ extracellular DNA (NETs) in atherosclerotic Apoe-/- mice.
Western Diet 2 wks 4 wks 6 wks normal chow
n = mice 5 4 3 2
# mice with NETs (%) 1 (20) 2 (50) 1 (33) 0 (0)
# luminal NETs in group 4 15 1 0
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from
13
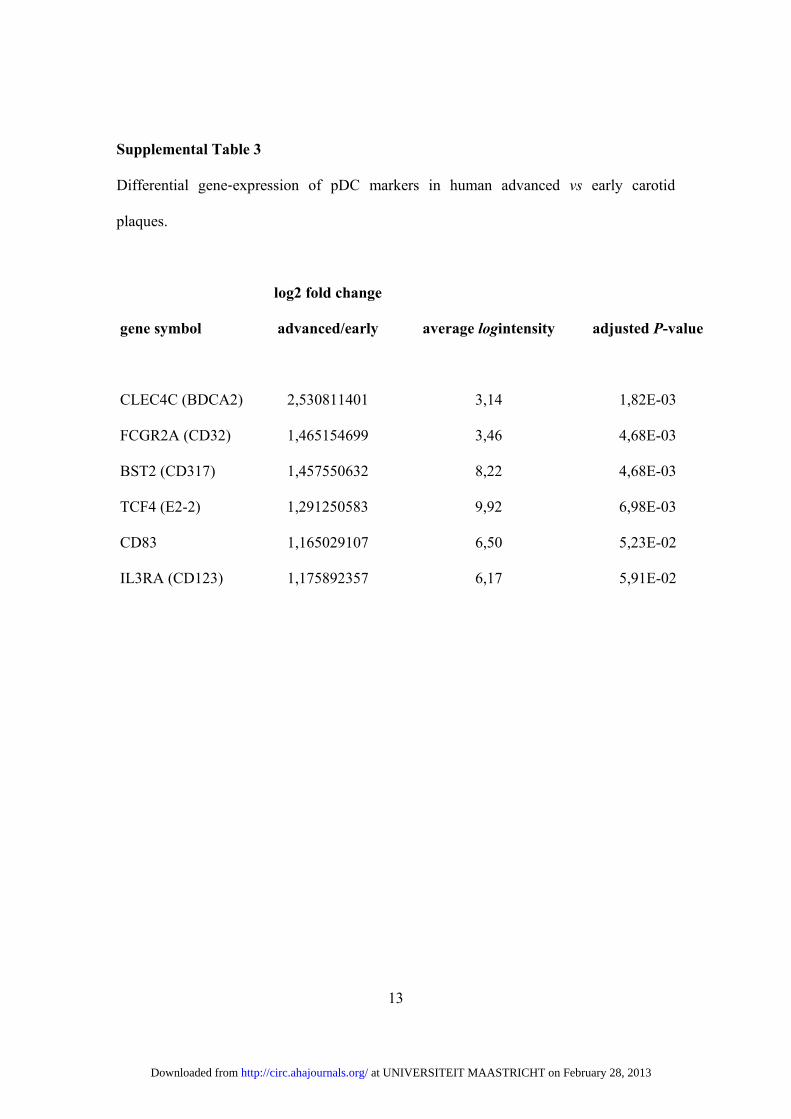
Supplemental Table 3
Differential gene‐expression of pDC markers in human advanced vs early carotid
plaques.
gene symbol
log2 fold change
advanced/early average logintensity adjusted P-value
CLEC4C (BDCA2) 2,530811401 3,14 1,82E-03
FCGR2A (CD32) 1,465154699 3,46 4,68E-03
BST2 (CD317) 1,457550632 8,22 4,68E-03
TCF4 (E2-2) 1,291250583 9,92 6,98E-03
CD83 1,165029107 6,50 5,23E-02
IL3RA (CD123) 1,175892357 6,17 5,91E-02
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

14
Supplemental Table 4
Characteristics of asymptomatic and symptomatic patients
asymptomatic symptomatic
(n=15) (n=15)
Age, years 69.7±8.1 69.9±10.4
Sex, females/males 5/10 6/9
Smokers (%) 47 47
Diabetes mellitus, % 0 0
Hypertension, % 67 93
Hyperlipidemia, % 47 40
Medication, %
β-blockers 40 33
Aspirin/Clopidogrel 93 80
Statins 60 60
Thrombosis prophylaxis 53 33
ACE-inhibitor 40 47
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

15
Supplemental Figure 1
Supplemental Figure 1
Efficiency and specificity of pDC depletion. Frequencies of CD3+ T cells, B220+ B cells,
CD11chigh MHCII+ cDCs, and PDCA1+ B220+ pDC were as assessed among CD45+
leukocytes by FACS analysis in Apoe-/- mice 24 hours after i.v. injection of PDCA1 or
isotype control antibody in lymph node , spleen and bone marrow. Bars represent median
with interquartile range (n=4 each). Representative dot plots of cell distributions in bone
marrow are shown; boxed regions indicate pDCs.
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

16
Supplemental Figure 2
Supplemental Figure 2
Localization of pDCs in LN and spleen. SiglecH+ pDCs (green) can be detected by
immunofluorescence and FACS analysis in lymph nodes and spleens of wild type mice.
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

17
Supplemental Figure 3
Supplemental Figure 3
Expression of surface markers and IFN-α by pDCs in response to different stimuli.
(A) Expression of CD68, SR-A, MHCII and CD86 in sorted pDCs stimulated
with/without oxLDL overnight, assessed by FACS analysis; representative histograms
with FMO/fluorescence minus one controls of 3 independent experiments are shown. (B)
IFNα secretion in supernatants of OT-II T cells left untreated or co-cultured with
unpulsed pDCs (control), OVA-2-pulsed pDCs overnight pretreated with or without
oxLDL or CpG over 3 days. Bars represent median with interquartile range, n=4
independent experiments. *P<0.05.
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

18
Supplemental Figure 4
Supplemental Figure 4
Atherosclerotic lesion formation in the aorta and plaque macrophage content in Apoe-/-
mice treated with PDCA1 antibody, recombinant IFN-α or CpG. (A) Atherosclerotic
lesions were quantified in the aorta after staining with oil-red O in Apoe-/- mice fed a high
fat diet and treated with IgG or PDCA1 antibody. The relative content of MOMA-2+
macrophages per total plaque cells was analyzed by quantitative immunofluorescence.
Bars represent median with interquartile range. (B) Quantification of atherosclerotic
lesions in the aorta after staining with oil-red O and of the relative frequencies of
MOMA-2+ macrophages per total plaque cells in the aortic root in Apoe-/- mice fed a high
fat diet and treated with PBS (control) or IFNα. Bars represent median with interquartile
range. (C) Quantification of atherosclerotic lesions in the aorta after staining with oil-red
O and of the relative frequencies of MOMA-2+ macrophages per total plaque cells in the
aortic root of Apoe-/- mice fed a high fat diet and treated with DOTAP (control), CpG or
CpG and PDCA1 antibody for 4 weeks. Bars represent mean ± SD (plaque area) and
median with interquartile range (macrophage content). *P<0.05. **P<0.01
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

19
Supplemental Figure 5
Supplemental Figure 5
pDC stimulation with Cramp/DNA complexes or serum, and anti-ds-DNA antibody titers
in IFN-α-treated Apoe-/- mice. (A) IFN-α protein secretion by sorted pDCs stimulated
overnight as indicated (n= 3-4). *P<0.05, ***P<0.001 vs. untreated, #P<0.05 vs. Cramp;
†P<0.05 vs. DNA. (B) Expression of MHC-II in sorted pDCs stimulated overnight with
Cramp, Cramp+DNA or DNA alone, assessed by FACS analysis; representative
histograms with FMO/fluorescence minus one controls of 3 independent experiments are
shown. (C) Bl6 mice were injected as indicated and IFN-α serum concentrations were
assessed after 4 hours (n= 9-10). *P<0.05 vs. IgG controls, #P<0.05 vs. Cramp/DNA +
IgG. (D) Anti-ds-DNA antibody titers were quantified in serum of Apoe-/- mice fed a
high-fat diet for 4 weeks and injected with PBS (control) or IFNα. n=6 mice per group.
*P<0.05. (E) Expression of MHC-II in sorted pDCs stimulated overnight with serum
with low (▼) or high (▲) anti-dsDNA antibody titers, assessed by FACS analysis;
representative histograms with FMO/fluorescence minus one control of 5 independent
experiments are shown. Bars represent mean ± SD.
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

20
Supplemental Figure 6
Supplemental Figure 6
Quantification of IgG autoantibodies directed against MDA-modified (left panel) and
oxidized LDL (right panel) in serum of Apoe-/- mice fed a high-fat diet for 4 weeks and
treated with IgG or PDCA1 antibody, or DOTAP (control), CpGs or CpGs+PDCA1 for 4
weeks. Bars represent mean ± SD, n=5-12 mice per group, *P<0.05. **P<0.01.
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

21
Supplemental References
1. Shen H, Iwasaki A. A crucial role for plasmacytoid dendritic cells in antiviral protection by CpG ODN-based vaginal microbicide. J Clin Invest. 2006; 116:2237-2243.
2. Krug A, French AR, Barchet W, Fischer JA, Dzionek A, Pingel JT, Orihuela MM, Akira S, Yokoyama WM, Colonna M. TLR9-dependent recognition of MCMV by IPC and DC generates coordinated cytokine responses that activate antiviral NK cell function. Immunity. 2004; 21:107-119.
3. Nizet V, Ohtake T, Lauth X, Trowbridge J, Rudisill J, Dorschner RA, Pestonjamasp V, Piraino J, Huttner K, Gallo RL. Innate antimicrobial peptide protects the skin from invasive bacterial infection. Nature. 2001; 414:454-457.
4. Zernecke A, Bot I, Djalali-Talab Y, Shagdarsuren E, Bidzhekov K, Meiler S, Krohn R, Schober A, Sperandio M, Soehnlein O, Bornemann J, Tacke F, Biessen EA, Weber C. Protective role of CXC receptor 4/CXC ligand 12 unveils the importance of neutrophils in atherosclerosis. Circ Res. 2008; 102:209-217.
5. Faust N, Varas F, Kelly LM, Heck S, Graf T. Insertion of enhanced green fluorescent protein into the lysozyme gene creates mice with green fluorescent granulocytes and macrophages. Blood. 2000; 96:719-726.
6. Van Rooijen N, Sanders A. Liposome mediated depletion of macrophages: mechanism of action, preparation of liposomes and applications. J Immunol Methods. 1994; 174:83-93.
7. Lutgens E, Lievens D, Beckers L, Wijnands E, Soehnlein O, Zernecke A, Seijkens T, Engel D, Cleutjens J, Keller AM, Naik SH, Boon L, Oufella HA, Mallat Z, Ahonen CL, Noelle RJ, de Winther MP, Daemen MJ, Biessen EA, Weber C. Deficient CD40-TRAF6 signaling in leukocytes prevents atherosclerosis by skewing the immune response toward an antiinflammatory profile. J Exp Med. 2010; 207:391-404.
8. Gregorio J, Meller S, Conrad C, Di Nardo A, Homey B, Lauerma A, Arai N, Gallo RL, Digiovanni J, Gilliet M. Plasmacytoid dendritic cells sense skin injury and promote wound healing through type I interferons. J Exp Med. 2010.
9. Ganguly D, Chamilos G, Lande R, Gregorio J, Meller S, Facchinetti V, Homey B, Barrat FJ, Zal T, Gilliet M. Self-RNA-antimicrobial peptide complexes activate human dendritic cells through TLR7 and TLR8. J Exp Med. 2009; 206:1983-1994.
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from

22
10. Drechsler M, Megens RT, van Zandvoort M, Weber C, Soehnlein O. Hyperlipidemia-triggered neutrophilia promotes early atherosclerosis. Circulation. 2010; 122:1837-1845.
11. Megens RT, Reitsma S, Prinzen L, oude Egbrink MG, Engels W, Leenders PJ, Brunenberg EJ, Reesink KD, Janssen BJ, ter Haar Romeny BM, Slaaf DW, van Zandvoort MA. In vivo high-resolution structural imaging of large arteries in small rodents using two-photon laser scanning microscopy. J Biomed Opt. 2010; 15:011108.
12. Tozzoli R, Bizzaro N, Tonutti E, Villalta D, Bassetti D, Manoni F, Piazza A, Pradella M, Rizzotti P. Guidelines for the laboratory use of autoantibody tests in the diagnosis and monitoring of autoimmune rheumatic diseases. Am J Clin Pathol. 2002; 117:316-324.
13. Binder CJ, Hartvigsen K, Chang MK, Miller M, Broide D, Palinski W, Curtiss LK, Corr M, Witztum JL. IL-5 links adaptive and natural immunity specific for epitopes of oxidized LDL and protects from atherosclerosis. J Clin Invest. 2004; 114:427-437.
14. Faber BC, Cleutjens KB, Niessen RL, Aarts PL, Boon W, Greenberg AS, Kitslaar PJ, Tordoir JH, Daemen MJ. Identification of genes potentially involved in rupture of human atherosclerotic plaques. Circ Res. 2001; 89:547-554.
15. Virmani R, Kolodgie FD, Burke AP, Farb A, Schwartz SM. Lessons from sudden coronary death: a comprehensive morphological classification scheme for atherosclerotic lesions. Arterioscler Thromb Vasc Biol. 2000; 20:1262-1275.
16. Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem. 1987; 162:156-159.
17. Team RDC. R: A language and environment for statistical computing. R Foundation for Statistical Computing, V., Austria. 2010; ISBN 3-900051-07-0:URL http://www.R-project.org/.
18. Gautier L, Cope L, Bolstad BM, Irizarry RA. affy--analysis of Affymetrix GeneChip data at the probe level. Bioinformatics. 2004; 20:307-315.
19. Smyth GK. Limma: linear models for microarray data. Bioinformatics and Computational Biology Solutions using R and Bioconductor. Springer. 2005:397-420.
at UNIVERSITEIT MAASTRICHT on February 28, 2013http://circ.ahajournals.org/Downloaded from
Copyright © 2022 FDOKUMEN




















