
Spreading depression features and Iba1 immunoreactivity in the cerebral cortex of developing rats...
8
Int. J. Devl Neuroscience 33 (2014) 98–105 Contents lists available at ScienceDirect International Journal of Developmental Neuroscience j ourna l ho me pa g e: www.elsevier.com/locate/ijdevneu Spreading depression features and Iba1 immunoreactivity in the cerebral cortex of developing rats submitted to treadmill exercise after treatment with monosodium glutamate Cássia Borges Lima a , Georgia de Sousa Ferreira Soares b , Suênia Marcele Vitor b , Belmira Lara da Silveira Andrade-da-Costa a , Bernardo Castellano c , Rubem Carlos Araujo Guedes b,∗ a Department of Physiology and Pharmacology, Universidade Federal de Pernambuco, 50670901 Recife, Brazil b Department of Nutrition, Universidade Federal de Pernambuco, 50670901 Recife, Brazil c Unit of Medical Histology, Institute of Neuroscience and Dept Cell Biology, Physiology and Immunology, Autonomous University of Barcelona, Spain a r t i c l e i n f o Article history: Received 17 July 2013 Received in revised form 9 November 2013 Accepted 18 December 2013 Keywords: Brain development Brain electrophysiology Cortical spreading depression Food flavoring agents Rats Treadmill exercise a b s t r a c t Physical exercise and excessive consumption of monosodium glutamate (MSG) can affect the morpho- logical and electrophysiological organization of the brain during development. However, the interaction of both factors remains unclear. We analyzed the effect of this interaction on the excitability-related phenomenon known as cortical spreading depression (CSD) and the microglial reaction expressed as Iba1-immunolabeled cells in the rat motor cortex. MSG (2 g/kg or 4 g/kg) was administered every other day during the first 14 postnatal days. Treadmill exercise started at 21–23 days of life and lasted 3 weeks, 5 days/week, for 30 min/day. At 45–60 days, CSD was recorded for 4 h at two cortical points and the CSD parameters (velocity, amplitude, and duration of the negative potential change) calculated. Confirming previous observations, exercised rats presented with lower CSD velocities (3.29 ± 0.18 mm/min) than the sedentary group (3.80 ± 0.18 mm/min; P < 0.05). MSG increased CSD velocities in the exercised rats com- pared to saline-treated and exercised animals in a dose-dependent manner (3.49 ± 0.19, 4.05 ± 0.18, and 3.27 ± 0.26 for 2 g/kg MSG, 4 g/kg MSG, and saline, respectively; P < 0.05). The amplitude (ranging from 14.3 ± 5.9 to 18.7 ± 6.2 mV) and duration (46.7 ± 11.1 to 60.5 ± 11.6 s) of the negative slow potential shift of the CSD were similar in all groups. Both exercise and MSG treatment increased Iba1 immunolabeling. The results confirm that physical exercise decelerates CSD propagation. However, it does not impede the CSD-accelerating action of MSG. These effects were accompanied by a cortical microglia reaction. Therefore, the data suggest that treadmill exercise early in life can influence the development of cortical electrical activity. © 2013 ISDN. Published by Elsevier Ltd. All rights reserved. 1. Introduction In addition to the well-known improvement in cardiovascular function (Karacabey, 2005), physical exercise seems to positively influence the nervous system, as evidenced by improved cogni- tive function, general arousal, and well-being (Martinsen, 2008; Anish, 2005; Hillman et al., 2004; Cotman and Berchtold, 2002; Cotman and Engesser-Cesar, 2002). In laboratory animals, exer- cise has been associated with improved learning and memory (Van ∗ Corresponding author at: Department of Nutrition, Universidade Federal de Pernambuco, 50670901 Recife, PE, Brazil. Tel.: +55 81 21268936; fax: +55 81 21268473. E-mail addresses: [email protected], [email protected] (R.C.A. Guedes). Praag et al., 2005), neurogenesis and increased neuron survival in the hippocampus (Brown et al., 2003; Farmer et al., 2004; Van Praag et al., 2005), and capillary growth and increased vascular flow in the cerebellum and motor cortex (Black et al., 1990; Swain et al., 2003). In the hippocampus, the neural effects of exercise also include increased expression of neurotrophins (Gomez-Pinilla et al., 2002; Fang et al., 2013) and signaling molecules (Shen et al., 2001), changes in the expression of genes (Tong et al., 2001), and enhanced hippocampal excitability, as indicated by long-term potentiation (Farmer et al., 2004). Therefore, the effects of physical exercise are thought to be neuroprotective, promoting brain health, preconditioning the brain against ischemic insult, and improv- ing long-term functioning (Zhang et al., 2013). In adult animals, these exercise-mediated alterations influence brain excitability and excitability-dependent neurological diseases (Peixinho-Pena 0736-5748/$36.00 © 2013 ISDN. Published by Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.ijdevneu.2013.12.008
Transcript of Spreading depression features and Iba1 immunoreactivity in the cerebral cortex of developing rats...

Sca
CBRa
b
c
a
ARRA
KBBCFRT
1
fitACc
Pf
0h
Int. J. Devl Neuroscience 33 (2014) 98–105
Contents lists available at ScienceDirect
International Journal of Developmental Neuroscience
j ourna l ho me pa g e: www.elsev ier .com/ locate / i jdevneu
preading depression features and Iba1 immunoreactivity in theerebral cortex of developing rats submitted to treadmill exercisefter treatment with monosodium glutamate
ássia Borges Limaa, Georgia de Sousa Ferreira Soaresb, Suênia Marcele Vitorb,elmira Lara da Silveira Andrade-da-Costaa, Bernardo Castellanoc,ubem Carlos Araujo Guedesb,∗
Department of Physiology and Pharmacology, Universidade Federal de Pernambuco, 50670901 Recife, BrazilDepartment of Nutrition, Universidade Federal de Pernambuco, 50670901 Recife, BrazilUnit of Medical Histology, Institute of Neuroscience and Dept Cell Biology, Physiology and Immunology, Autonomous University of Barcelona, Spain
r t i c l e i n f o
rticle history:eceived 17 July 2013eceived in revised form 9 November 2013ccepted 18 December 2013
eywords:rain developmentrain electrophysiologyortical spreading depressionood flavoring agentsatsreadmill exercise
a b s t r a c t
Physical exercise and excessive consumption of monosodium glutamate (MSG) can affect the morpho-logical and electrophysiological organization of the brain during development. However, the interactionof both factors remains unclear. We analyzed the effect of this interaction on the excitability-relatedphenomenon known as cortical spreading depression (CSD) and the microglial reaction expressed asIba1-immunolabeled cells in the rat motor cortex. MSG (2 g/kg or 4 g/kg) was administered every otherday during the first 14 postnatal days. Treadmill exercise started at 21–23 days of life and lasted 3 weeks,5 days/week, for 30 min/day. At 45–60 days, CSD was recorded for 4 h at two cortical points and the CSDparameters (velocity, amplitude, and duration of the negative potential change) calculated. Confirmingprevious observations, exercised rats presented with lower CSD velocities (3.29 ± 0.18 mm/min) than thesedentary group (3.80 ± 0.18 mm/min; P < 0.05). MSG increased CSD velocities in the exercised rats com-pared to saline-treated and exercised animals in a dose-dependent manner (3.49 ± 0.19, 4.05 ± 0.18, and3.27 ± 0.26 for 2 g/kg MSG, 4 g/kg MSG, and saline, respectively; P < 0.05). The amplitude (ranging from
14.3 ± 5.9 to 18.7 ± 6.2 mV) and duration (46.7 ± 11.1 to 60.5 ± 11.6 s) of the negative slow potential shiftof the CSD were similar in all groups. Both exercise and MSG treatment increased Iba1 immunolabeling.The results confirm that physical exercise decelerates CSD propagation. However, it does not impedethe CSD-accelerating action of MSG. These effects were accompanied by a cortical microglia reaction.Therefore, the data suggest that treadmill exercise early in life can influence the development of corticalelectrical activity.© 2013 ISDN. Published by Elsevier Ltd. All rights reserved.
. Introduction
In addition to the well-known improvement in cardiovascularunction (Karacabey, 2005), physical exercise seems to positivelynfluence the nervous system, as evidenced by improved cogni-ive function, general arousal, and well-being (Martinsen, 2008;
nish, 2005; Hillman et al., 2004; Cotman and Berchtold, 2002;otman and Engesser-Cesar, 2002). In laboratory animals, exer-ise has been associated with improved learning and memory (Van∗ Corresponding author at: Department of Nutrition, Universidade Federal deernambuco, 50670901 Recife, PE, Brazil. Tel.: +55 81 21268936;ax: +55 81 21268473.
E-mail addresses: [email protected], [email protected] (R.C.A. Guedes).
736-5748/$36.00 © 2013 ISDN. Published by Elsevier Ltd. All rights reserved.ttp://dx.doi.org/10.1016/j.ijdevneu.2013.12.008
Praag et al., 2005), neurogenesis and increased neuron survival inthe hippocampus (Brown et al., 2003; Farmer et al., 2004; VanPraag et al., 2005), and capillary growth and increased vascularflow in the cerebellum and motor cortex (Black et al., 1990; Swainet al., 2003). In the hippocampus, the neural effects of exercisealso include increased expression of neurotrophins (Gomez-Pinillaet al., 2002; Fang et al., 2013) and signaling molecules (Shen et al.,2001), changes in the expression of genes (Tong et al., 2001),and enhanced hippocampal excitability, as indicated by long-termpotentiation (Farmer et al., 2004). Therefore, the effects of physicalexercise are thought to be neuroprotective, promoting brain health,
preconditioning the brain against ischemic insult, and improv-ing long-term functioning (Zhang et al., 2013). In adult animals,these exercise-mediated alterations influence brain excitabilityand excitability-dependent neurological diseases (Peixinho-Pena
Neuro
ee
tcmapgEaP
astiwoittdarccctvau(ppbCbk
w4oetmostmr
2
2
(CdoB(ap
C.B. Lima et al. / Int. J. Devl
t al., 2012). However, little is known about the electrophysiologicalffects of exercise on the developing cerebral cortex.
MSG is a neuroexcitatory amino acid used primarily to reinforcehe flavor of several foods. When consumed in great amounts, MSGan be harmful to the brain (Nemeroff et al., 1978). In mice andonkeys, administration of MSG results in brain lesions early in life
nd obesity later in life (Abraham et al., 1971). In adult Wistar rats,rolonged administration of high doses of MSG results in neurode-enerative changes in pyramidal cells (Musa and Sunday, 2013).xperimental evidence also indicates that MSG influences electricalctivity in the brain, facilitating the initiation of seizures (Lopez-erez et al., 2010).
We recently demonstrated that treadmill exercise deceler-tes the excitability-related brain phenomenon known as corticalpreading depression (CSD) (Batista-de-Oliveira et al., 2012), andhat MSG treatment early in life increases microglial reactionsn the rat cortex and accelerates CSD (Lima et al., 2013). CSD
as first described as a slowly propagating wave of suppressionf electrical activity in the rabbit cortex (Leão, 1944, 1947), ands characterized by a depolarization of neurons and glial cellshat propagates in the gray matter of the central nervous sys-em (Maia et al., 2009; Marrannes et al., 1988). CSD has beenescribed electrophysiologically in laboratory animals (Gorji, 2001)nd humans (Dohmen et al., 2008; Fabricius et al., 2008) as aeversible and propagating phenomenon that can be elicited byhemical, mechanical, or electrical stimulation of one point of theortical surface. Several studies have suggested that alterations inortical excitability are associated with changes in CSD propaga-ion (Liebetanz et al., 2006; Fregni et al., 2007). Measuring theelocity of CSD propagation along the cortical tissue is a reason-ble and simple method for estimating brain CSD susceptibilitynder clinically relevant conditions that influence brain excitabilityAmaral et al., 2009). Conditions that modify CSD generation andropagation constitute useful opportunities for understanding therocesses underlying neurological diseases that are dependent onrain excitability, such as epilepsy (Leão, 1944, 1972; Guedes andavalheiro, 1997; Guedes et al., 2009). In addition, the role playedy microglial cells in such neurological disorders is not completelynown.
Therefore, we decided to investigate in 45–60-day-old ratshether physical (treadmill) exercise (performed from 21–23 to
2–44 days of life) interacts with MSG in regards to the effectsn CSD and microglial reaction. We characterized CSD param-ters (propagation velocity and the amplitude and duration ofhe negative slow potential shift) and the Iba1-immunolabeled
icroglial reaction in exercised rats, comparing them to thosef sedentary animals. Furthermore, we compared MSG- andaline-treated groups of exercised rats. We hypothesized thatreadmill exercise attenuates the effects of previous MSG treat-
ent on the brain’s ability to propagate CSD and microgliaeaction.
. Materials and methods
.1. Animals
The animals were handled in accordance with the normsApproval Protocol no. 23076.002006/2009-80) of the Ethicsommittee for Animal Research of the Universidade Federale Pernambuco, Brazil, which complies with the “Principlesf Laboratory Animal Care” (National Institutes of Health,
ethesda, USA). The rats were housed in polypropylene cages51 cm × 35.5 cm × 18.5 cm) in a room maintained at 22 ± 1 ◦C with12:12 h light–dark cycle (lights on at 7:00 a.m.) and fed a 23%rotein lab chow diet (Purina do Brazil Ltd., São Paulo, Brazil).
science 33 (2014) 98–105 99
2.2. Treatment with MSG
Male Wistar rat pups (n = 28) were treated with MSG as pre-viously described (Lima et al., 2013). Briefly, the animals receivedsubcutaneous injections of 2 g/kg (n = 13; MSG-2) or 4 g/kg MSG(n = 15; MSG-4) administered every other day during the first 14days of life. Two control groups were used for comparison: oneinjected with saline (n = 12) and the other not injected at all (naïve;n = 6).
2.3. Treadmill exercise
After weaning, from postnatal days 21–23 to 42–44, the ani-mals were submitted to daily sessions of running on a treadmill(Insight EP-131, 0◦ inclination; 30 min/day, 5 days/week, total time3 weeks). As control group, sedentary rats (n = 15) were placed inthe treadmill apparatus daily for a time-period similar to that ofthe exercised group, but the treadmill apparatus remained off.
2.4. Body weights
The rats were weighed at postnatal days 2, 10, and 45–50.
2.5. CSD recording
When the animals were 45–60 days old they were submittedto CSD recording for 4 h as described previously (Accioly et al.,2012). Under anesthesia (1 g/kg urethane plus 40 mg/kg chloralose,ip), the rat’s head was secured in a stereotaxic apparatus (Kopf,USA) and three trephine holes (2–3 mm in diameter) drilled inthe right side of the skull: two holes in the parietal bone and onein the frontal bone, aligned in the anteroposterior direction andparallel to the midline. CSD was elicited at 20-min intervals byapplying a cotton ball (1–2 mm diameter) soaked in 2% KCl solu-tion (approximately 0.27 M) to the anterior (frontal) hole for 1 min.The electrocorticogram (ECoG) and slow direct current (DC) poten-tial change accompanying CSD were recorded simultaneously atthe two parietal points on the cortical surface via a pair of Ag-AgCl agar-Ringer electrodes. These electrodes consisted of plasticconic pipettes (5 cm length, 0.5 mm tip inner diameter) filled withRinger solution solidified by adding 0.5% agar and into which a chlo-rided silver wire was inserted. The pipettes were fixed in pairs withcyanoacrylate glue, maintaining a constant interelectrode distancefor each pair (range: 4–5.5 mm). The electrode pair was fixed tothe electrode holder of the stereotaxic apparatus in a manner thatallowed the electrodes to be gently placed on the intact dura-materunder the control of a low-power microscope without any exces-sive pressure on the cortical surface. A common reference electrodeof the same type was placed on the nasal bones. The velocity ofCSD propagation was calculated based on the time required for aCSD wave to cross the interelectrode distance. When measuringCSD velocities, the initial point of each DC negative rising phasewas used as the reference point. We also calculated the amplitudeand duration of the CSD negative slow potential shifts. During therecording session, rectal temperature was maintained at 37 ± 1 ◦Cusing a heating blanket. At the end of the recording session, theanimal was euthanized by an overdose of anesthetic.
2.6. Iba1 immunolabeling of microglial cells
Thirteen rats treated with saline (n = 4), MSG-2 (n = 4), or MSG-4 (n = 5) were perfused with 0.9% saline solution followed by 4%
paraformaldehyde diluted in 0.1 M phosphate-buffered saline (pH7.4). After being immersed in the fixative for 4 h, the brains weretransferred to a 30% (w/v) sucrose solution for cryoprotection. Lon-gitudinal serial sections (40-�m thickness) were obtained at −20 ◦C
1 Neuroscience 33 (2014) 98–105
uamWsH1saatroaTt(dxpa(Uhdsstoa
2
cweti
3
3
csehaws
3
DtsarfFsKb
Fig. 1. (A) Body weights of 45–50-day-old sedentary and exercised rats, show-ing a significant weight reduction in the exercised group. (B) Body weights of 2,
3.3. Measurement of CSD parameters
We present the mean CSD propagation velocities in seden-tary and exercised rats that were not treated with MSG in
Fig. 2. Electrocorticogram (E) and slow potential change (P) recorded during corticalspreading depression (CSD) in three 45–60-day-old rats from the naïve (Nv) group,saline (Sal) group, and the group treated with 4 g/kg monosodium glutamate (MSG-4). The horizontal bars in P1 show the period (1 min) of stimulation with 2% KCl
00 C.B. Lima et al. / Int. J. Devl
sing a cryoslicer (Leica 1850). Sections were immunolabeled with polyclonal antibody against ionized calcium-binding adapterolecule 1 (Iba-1) to detect microglia (anti-Iba-1, #019-19741;ako Pure Chemical Industries Ltd., Osaka, Japan). Free-floating
ections were submitted to endogenous peroxidase blocking (2%2O2 in 70% methanol for 10 min) and the sections incubated for
h in blocking buffer (BB) solution containing 0.05 M Tris-bufferedaline (TBS; pH 7.4), 10% fetal calf serum, 3% bovine serum albumin,nd 1% Triton X-100. The sections were then incubated overnightt 4 ◦C with rabbit anti-Iba-1 (1:1500 diluted in BB solution). Afterhree washes with TBS + 1% Triton, sections were incubated atoom temperature for 1 h with biotinylated anti-rabbit (1:500) sec-ndary antibodies. Sections were then rinsed in TBS + 1% Tritonnd incubated with horseradish peroxidase streptavidin (1:500).he peroxidase reaction was visualized by incubating the sec-ions in Tris buffer containing 0.5 mg/ml 3,3′-diaminobenzidineDAB) and 0.33 �l/ml H2O2. Finally, the sections were mounted,ehydrated in graded alcohols, and coverslipped in Entellan® afterylene treatment. Densitometric analysis was performed on fourarallel longitudinal sections for each animal. In each section, wenalyzed photomicrographs of four fields within the motor cortexlayer 5) using the Image J software (National Institutes of Health,SA, version 1.46r). A Leica DMLS microscope coupled to a Samsungigh-level color camera (model SHC-410NAD) was used to obtainigital images from brain sections. Images of the motor cortextained for Iba1 were obtained using a 20×, 40× and 100× micro-cope objective. Care was taken to obtain the digital images usinghe same light intensity. We analyzed the percentage of the areaccupied by the Iba1-labeled cells, as well as their total immunore-ctivity expressed as arbitrary units.
.7. Statistical analysis
Statistical analyses were performed using Sigmastat® statisti-al software (version 3.5, San Jose, CA, USA). Intergroup differencesere compared using ANOVA, including exercise status (Treadmill
xercised or sedentary) and drug treatment (saline or MSG) as fac-ors. ANOVA was followed by a post hoc (Holm–Sidak) test whenndicated. Differences were considered significant when P ≤ 0.05.
. Results
.1. Body weight
On the day of the CSD recording (45–60 days of age) the exer-ised rats had lower body weights (168.1 ± 22.7 g; n = 15) than theedentary animals (214.0 ± 32.5 g; n = 10; P < 0.05; Fig. 1A). The twoxercised groups that were previously treated with MSG (Fig. 1B)ad lower body weights (140.0 ± 25.1 g and 141.9 ± 34.7 g for MSG2nd MSG4, respectively) compared to the exercised group treatedith saline (165.1 ± 21.1 g; P < 0.05). This group had a body weight
imilar to non-treated (“naïve”) rats (172.7 ± 26.3 g).
.2. CSD recording
Fig. 2 shows the typical electrophysiological recordings (slowC potential change and ECoG) in three exercised rats, represen-
ative of the naïve, saline, and MSG-4 groups. As a rule, the 1-mintimulation with 2% KCl at one point of the frontal cortex elicited
single CSD wave that propagated without interruption and wasecorded by the two electrodes located more posterior on the sur-ace of the parietal cortex (see stimulation and recording points in
ig. 2, lower left panel). In each recording point, the ECoG depres-ion and slow potential change confirmed the presence of CSD afterCl application. As a rule, the electrophysiological changes causedy CSD always recovered after a few minutes, and we maintained a10, and 45–50-day-old exercised rats. Sal, saline; Nv, naïve, no treatment; MSG-2,2 g/kg MSG; MSG-4, 4 g/kg MSG (subcutaneous injections). Data are expressed asmean ± S.E.M. *P < 0.05 (ANOVA plus Holm–Sidak test).
20-min interval between subsequent KCl stimulations. The patternof CSD recording in the MSG-treated and sedentary rats (not shownin Fig. 2) was similar to that of the exercised animals.
necessary to elicit CSD. The vertical bars are equivalent to −10 mV and −1 mV forthe P and E recordings, respectively (negativity is upwards). The locations of KClapplication and the reference electrode are indicated in the skull diagram, whichalso shows recording points 1 and 2 from which the traces marked at the centerwith the same numbers were recorded.
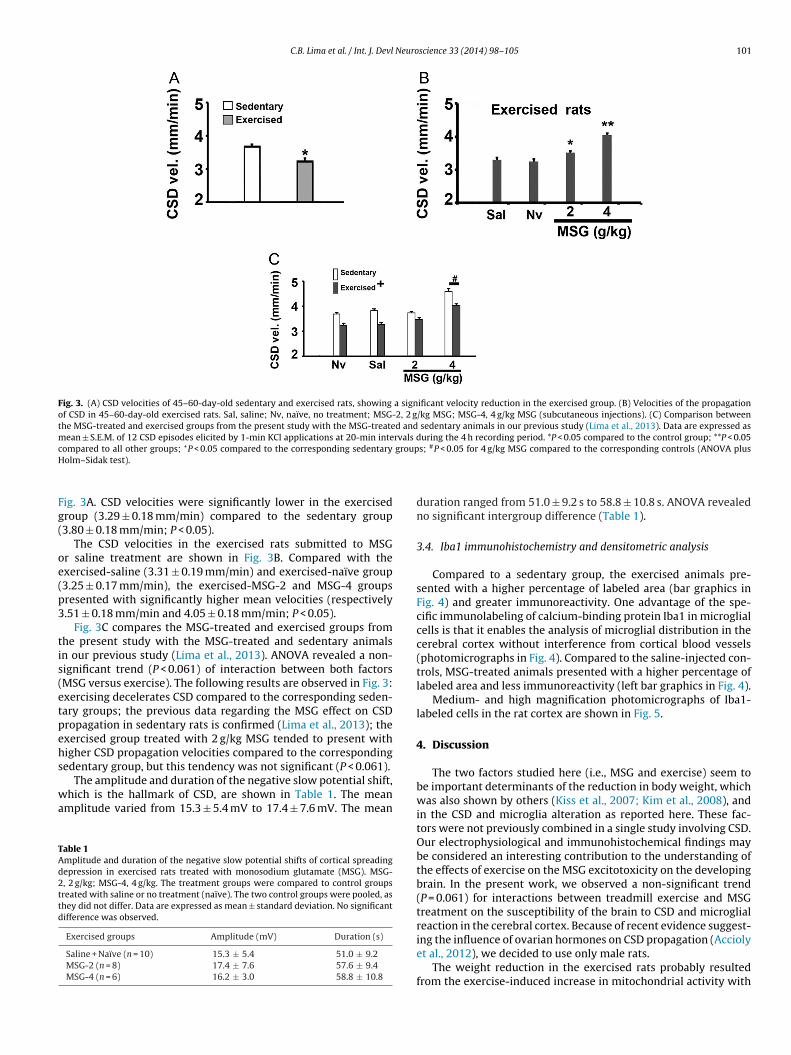
C.B. Lima et al. / Int. J. Devl Neuroscience 33 (2014) 98–105 101
Fig. 3. (A) CSD velocities of 45–60-day-old sedentary and exercised rats, showing a significant velocity reduction in the exercised group. (B) Velocities of the propagationof CSD in 45–60-day-old exercised rats. Sal, saline; Nv, naïve, no treatment; MSG-2, 2 g/kg MSG; MSG-4, 4 g/kg MSG (subcutaneous injections). (C) Comparison betweenthe MSG-treated and exercised groups from the present study with the MSG-treated and sedentary animals in our previous study (Lima et al., 2013). Data are expressed asmean ± S.E.M. of 12 CSD episodes elicited by 1-min KCl applications at 20-min intervals during the 4 h recording period. *P < 0.05 compared to the control group; **P < 0.05c groupH
Fg(
oe(p3
tis(etpehs
wa
TAd2ttd
ompared to all other groups; +P < 0.05 compared to the corresponding sedentaryolm–Sidak test).
ig. 3A. CSD velocities were significantly lower in the exercisedroup (3.29 ± 0.18 mm/min) compared to the sedentary group3.80 ± 0.18 mm/min; P < 0.05).
The CSD velocities in the exercised rats submitted to MSGr saline treatment are shown in Fig. 3B. Compared with thexercised-saline (3.31 ± 0.19 mm/min) and exercised-naïve group3.25 ± 0.17 mm/min), the exercised-MSG-2 and MSG-4 groupsresented with significantly higher mean velocities (respectively.51 ± 0.18 mm/min and 4.05 ± 0.18 mm/min; P < 0.05).
Fig. 3C compares the MSG-treated and exercised groups fromhe present study with the MSG-treated and sedentary animalsn our previous study (Lima et al., 2013). ANOVA revealed a non-ignificant trend (P < 0.061) of interaction between both factorsMSG versus exercise). The following results are observed in Fig. 3:xercising decelerates CSD compared to the corresponding seden-ary groups; the previous data regarding the MSG effect on CSDropagation in sedentary rats is confirmed (Lima et al., 2013); thexercised group treated with 2 g/kg MSG tended to present withigher CSD propagation velocities compared to the correspondingedentary group, but this tendency was not significant (P < 0.061).
The amplitude and duration of the negative slow potential shift,hich is the hallmark of CSD, are shown in Table 1. The mean
mplitude varied from 15.3 ± 5.4 mV to 17.4 ± 7.6 mV. The mean
able 1mplitude and duration of the negative slow potential shifts of cortical spreadingepression in exercised rats treated with monosodium glutamate (MSG). MSG-, 2 g/kg; MSG-4, 4 g/kg. The treatment groups were compared to control groupsreated with saline or no treatment (naïve). The two control groups were pooled, ashey did not differ. Data are expressed as mean ± standard deviation. No significantifference was observed.
Exercised groups Amplitude (mV) Duration (s)
Saline + Naïve (n = 10) 15.3 ± 5.4 51.0 ± 9.2MSG-2 (n = 8) 17.4 ± 7.6 57.6 ± 9.4MSG-4 (n = 6) 16.2 ± 3.0 58.8 ± 10.8
s; #P < 0.05 for 4 g/kg MSG compared to the corresponding controls (ANOVA plus
duration ranged from 51.0 ± 9.2 s to 58.8 ± 10.8 s. ANOVA revealedno significant intergroup difference (Table 1).
3.4. Iba1 immunohistochemistry and densitometric analysis
Compared to a sedentary group, the exercised animals pre-sented with a higher percentage of labeled area (bar graphics inFig. 4) and greater immunoreactivity. One advantage of the spe-cific immunolabeling of calcium-binding protein Iba1 in microglialcells is that it enables the analysis of microglial distribution in thecerebral cortex without interference from cortical blood vessels(photomicrographs in Fig. 4). Compared to the saline-injected con-trols, MSG-treated animals presented with a higher percentage oflabeled area and less immunoreactivity (left bar graphics in Fig. 4).
Medium- and high magnification photomicrographs of Iba1-labeled cells in the rat cortex are shown in Fig. 5.
4. Discussion
The two factors studied here (i.e., MSG and exercise) seem tobe important determinants of the reduction in body weight, whichwas also shown by others (Kiss et al., 2007; Kim et al., 2008), andin the CSD and microglia alteration as reported here. These fac-tors were not previously combined in a single study involving CSD.Our electrophysiological and immunohistochemical findings maybe considered an interesting contribution to the understanding ofthe effects of exercise on the MSG excitotoxicity on the developingbrain. In the present work, we observed a non-significant trend(P = 0.061) for interactions between treadmill exercise and MSGtreatment on the susceptibility of the brain to CSD and microglialreaction in the cerebral cortex. Because of recent evidence suggest-
ing the influence of ovarian hormones on CSD propagation (Acciolyet al., 2012), we decided to use only male rats.The weight reduction in the exercised rats probably resultedfrom the exercise-induced increase in mitochondrial activity with

102 C.B. Lima et al. / Int. J. Devl Neuroscience 33 (2014) 98–105
Fig. 4. Low magnification photomicrographs of Iba1-labeled cells in longitudinal sections of the motor cortex of three exercised rats treated with saline, 2 g/kg MSG, and4 g/kg MSG, respectively. Scale bars = 20 �m. The lower bar graphics show the percent area occupied by the Iba1-labeled cells and immunoreactivity expressed as arbitraryu Data ac
cdpitscMa(
eer2(mmg2mtlrdpcvOC
nits (a.u.) of 50–55-day-old sedentary (right) and treadmill-exercised rats (left).
ompared to the saline and MSG-2 groups (ANOVA plus Holm–Sidak test).
onsequent mobilization of fat and more fatty acid oxidationuring exercise (Moura et al., 2013). This recent evidence sup-orts the postulation that the practice of physical exercise early
n life can contribute to the prevention of obesity and insulin resis-ance (Montesi et al., 2013; Moraes-Silva et al., 2013). The presenttudy also showed that MSG treatment can potentiate the defi-iency in body weight in rats subjected to treadmill exercise; theSG-exercise association may cause a series of metabolic effects,
s demonstrated by others using swimming-exercised animalsAndreazzi et al., 2009; Miranda et al., 2013).
Information on the long-lasting electrophysiological effects ofarly physical exercise on the cerebral cortex is scarce. One rel-vant effect of physical exercise is neural protection, which waseported in a chronic mouse model of Parkinson’s disease (Lau et al.,011). Also, exercise has been shown to promote brain angiogenesisIsaacs et al., 1992) and to increase the levels of NMDA-type gluta-
ate receptor mRNA (Farmer et al., 2004). Exercise also increasesRNA levels of brain-derived neurotrophic factor (BDNF) and nerve
rowth factor in rat brain (Gomez-Pinilla et al., 2002; Soya et al.,007). This finding is of particular importance during develop-ent, as BDNF is the prominent regulator of synaptic plasticity in
he brain (Rothman & Mattson, 2013). Treadmill exercise early inife can modulate the functional maturation of the brain via neu-oplastic processes (Almeida et al., 2013). Environmental stimuliuring early postnatal brain development may result in more com-lex neural circuitry (Linkenhoker et al., 2005). In this study, we
onfirmed that physical exercise per se reduced CSD propagationelocity, which is in agreement with a recent report (Batista-de-liveira et al., 2012), demonstrating a similar effect of exercise onSD in both well-nourished and malnourished, young and agedre expressed as mean ± S.E.M. *P < 0.05 compared to the control group. **P < 0.05
rats. Concerning the cellular and molecular mechanisms under-lying exercise-induced effects on CSD, we speculate that factorssuch as the more effective removal of extracellular K+ (Guedesand Cavalheiro, 1997) and/or glutamate (Swanson et al., 2005;Moskowitz et al., 2004) by astrocytes during cortical neuronal activ-ity may play a relevant role. However, other metabolic factorsshould be taken into consideration in future experiments consid-ering that physical activity considerably impacts on the hepaticmetabolism (Ascensão et al., 2013) with possible repercussions inthe brain.
Because the capacity for neuroplasticity decreases with increas-ing age (Akopian and Walsh, 2006; Lynch et al., 2006), assessinghow exercise may beneficially regulate neuroplasticity during earlylife is important, as well as determining the basic mechanismof such effects in the normal brain. Taken together, our find-ings support the suggestion that physical exercise early in lifedecreases neuronal excitability in the young rat cortex. Further-more, the exercise-induced enhancement in the cortical microglialimmunoreactivity raises the question of whether this cell reac-tion is an epiphenomenon for the effects of exercise on CSD. Inorder to clarify this point, further experiments should examine themicroglial reaction as a required process for exercise-induced CSDdeceleration.
We previously reported that MSG treatment dose-dependentlyincreases microglial Iba1 immunolabeling in the motor cortex ofsedentary rats (Lima et al., 2013). In the present study, the exercise
in MSG treated animals decreased Iba1 immunoreactivity intensityand increased the immunolabeled area. Based on the qualitativedata shown in Figs. 4 and 5, one possibility for this discrepancy isthat exercise can modify microglia phenotypic changes previously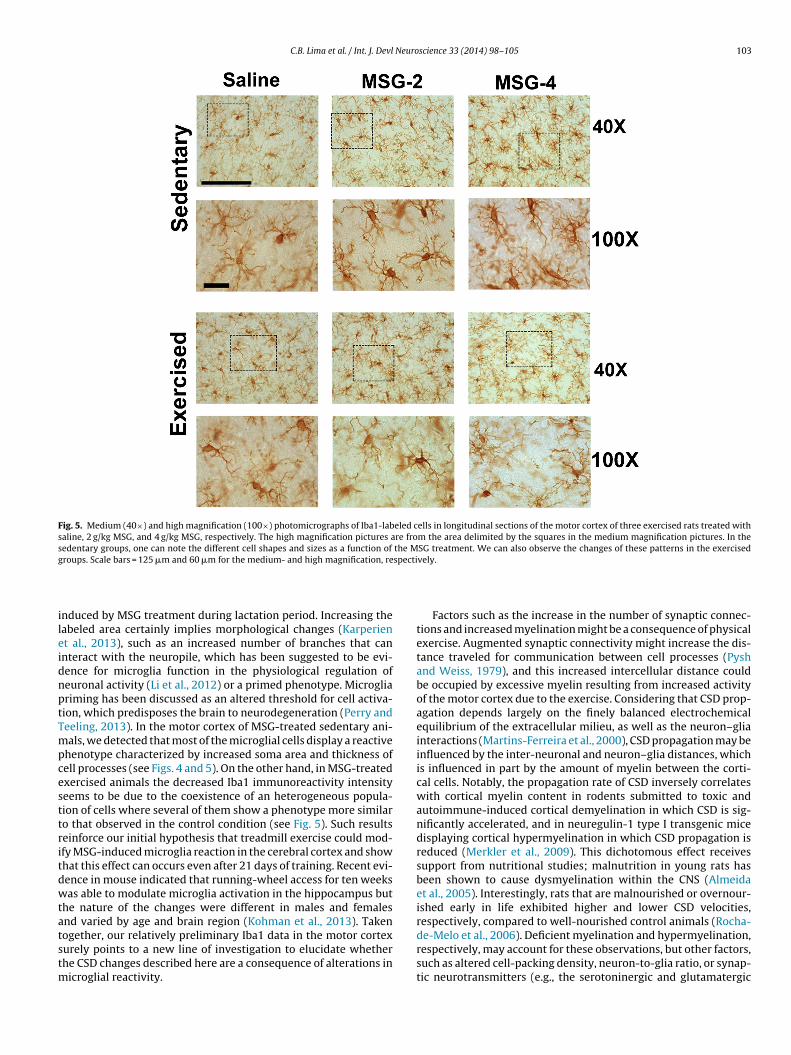
C.B. Lima et al. / Int. J. Devl Neuroscience 33 (2014) 98–105 103
Fig. 5. Medium (40×) and high magnification (100×) photomicrographs of Iba1-labeled cells in longitudinal sections of the motor cortex of three exercised rats treated withsaline, 2 g/kg MSG, and 4 g/kg MSG, respectively. The high magnification pictures are from the area delimited by the squares in the medium magnification pictures. In thesedentary groups, one can note the different cell shapes and sizes as a function of the MSG treatment. We can also observe the changes of these patterns in the exercisedg specti
ileidnptTmpcesttritdwtatstm
roups. Scale bars = 125 �m and 60 �m for the medium- and high magnification, re
nduced by MSG treatment during lactation period. Increasing theabeled area certainly implies morphological changes (Karperient al., 2013), such as an increased number of branches that cannteract with the neuropile, which has been suggested to be evi-ence for microglia function in the physiological regulation ofeuronal activity (Li et al., 2012) or a primed phenotype. Microgliariming has been discussed as an altered threshold for cell activa-ion, which predisposes the brain to neurodegeneration (Perry andeeling, 2013). In the motor cortex of MSG-treated sedentary ani-als, we detected that most of the microglial cells display a reactive
henotype characterized by increased soma area and thickness ofell processes (see Figs. 4 and 5). On the other hand, in MSG-treatedxercised animals the decreased Iba1 immunoreactivity intensityeems to be due to the coexistence of an heterogeneous popula-ion of cells where several of them show a phenotype more similaro that observed in the control condition (see Fig. 5). Such resultseinforce our initial hypothesis that treadmill exercise could mod-fy MSG-induced microglia reaction in the cerebral cortex and showhat this effect can occurs even after 21 days of training. Recent evi-ence in mouse indicated that running-wheel access for ten weeksas able to modulate microglia activation in the hippocampus but
he nature of the changes were different in males and femalesnd varied by age and brain region (Kohman et al., 2013). Taken
ogether, our relatively preliminary Iba1 data in the motor cortexurely points to a new line of investigation to elucidate whetherhe CSD changes described here are a consequence of alterations inicroglial reactivity.
vely.
Factors such as the increase in the number of synaptic connec-tions and increased myelination might be a consequence of physicalexercise. Augmented synaptic connectivity might increase the dis-tance traveled for communication between cell processes (Pyshand Weiss, 1979), and this increased intercellular distance couldbe occupied by excessive myelin resulting from increased activityof the motor cortex due to the exercise. Considering that CSD prop-agation depends largely on the finely balanced electrochemicalequilibrium of the extracellular milieu, as well as the neuron–gliainteractions (Martins-Ferreira et al., 2000), CSD propagation may beinfluenced by the inter-neuronal and neuron–glia distances, whichis influenced in part by the amount of myelin between the corti-cal cells. Notably, the propagation rate of CSD inversely correlateswith cortical myelin content in rodents submitted to toxic andautoimmune-induced cortical demyelination in which CSD is sig-nificantly accelerated, and in neuregulin-1 type I transgenic micedisplaying cortical hypermyelination in which CSD propagation isreduced (Merkler et al., 2009). This dichotomous effect receivessupport from nutritional studies; malnutrition in young rats hasbeen shown to cause dysmyelination within the CNS (Almeidaet al., 2005). Interestingly, rats that are malnourished or overnour-ished early in life exhibited higher and lower CSD velocities,respectively, compared to well-nourished control animals (Rocha-
de-Melo et al., 2006). Deficient myelination and hypermyelination,respectively, may account for these observations, but other factors,such as altered cell-packing density, neuron-to-glia ratio, or synap-tic neurotransmitters (e.g., the serotoninergic and glutamatergic
1 Neuro
scteeattwi(de
aEa(anenoeiGooieccgal(
ymrptctimondhtbb
C
A
cNdA
04 C.B. Lima et al. / Int. J. Devl
ystems), also have to be considered as possible causal agents. Con-erning the serotoninergic system, compelling evidence indicateshat changes in serotoninergic activity inversely influence brainxcitability (Igelström and Heyward, 2012). We know from the lit-rature that physical exercise can alter the branched chain aminocid/tryptophan ratio in the circulating blood, and this increaseshe tryptophan crossing the blood–brain barrier and increase sero-onin production in the neurons (Blomstrand, 2001). In agreementith this idea, some studies have unequivocally demonstrated that
ncreased brain serotonin availability antagonizes CSD propagationGuedes et al., 2002; Amâncio-dos-Santos et al., 2006), whereasecreasing serotonin activity accelerates CSD (Amancio-dos-Santost al., 2013).
The cerebral cortex constitutes an important target area for thections of physical exercise and MSG during brain development.arly administration of MSG to rat pups can result in excitotoxicitynd neuronal degeneration by over-activating glutamate receptorsBeas-Zárate et al., 2001), which can disrupt the physiological bal-nce between glutamatergic and GABAergic influences in the celletworks of the cerebral cortex and hippocampus (Urena-Guerrerot al., 2009), predisposing the organism to excitability-dependenteurological diseases, such as epilepsy. Also, excessive activationf glutamate receptors is thought to contribute to neurodegen-ration following a wide range of neurological insults, includingschemia, trauma, hypoglycemia, and epileptic seizures (Pellegrini-iampietro et al., 1997). According to these previous studies,ne possible underlying mechanism that contributes to the lossf neurons associated with such brain disorders is an abnormalnflux of Ca2+ through glutamate receptor channels. In this context,nhanced immunolabeling of the microglial cells with a mono-lonal antibody against Iba1 could be interpreted as a specificell reaction against imbalanced calcium influx. Interestingly, bothlutamate receptor activation and transmembrane calcium fluxlterations are known to influence CSD, as demonstrated by ouraboratory (Guedes et al., 1988; Torrente et al., 2013) and othersMarrannes et al., 1988; Siesjö and Bengtsson, 1989).
In conclusion, we demonstrated that the cerebral cortex ofoung rats submitted to treadmill exercise and MSG treat-ent present with reduced and increased susceptibility to CSD,
espectively, as indicated by the corresponding changes in CSDropagation velocities and increased microglial reaction comparedo control animals. Both electrophysiological and immunohisto-hemical data suggested interactions between exercise and MSGreatment. We suggest that this interaction reflects the excitabil-ty effects of both treatments involving glutamate/GABA-based
odulation of cortical excitability. Considering that the rats fromur study were healthy (i.e., they had no demyelination or othereurological disorders), our observations of an exercise-associatedeceleration in CSD propagation suggest that physical exercise canelp prevent neurological insults such as MSG neurotoxicity, buthey can also have a beneficial action on the normal developingrain, which could be long-lasting with a positive effect on the agedrain (Batista-de-Oliveira et al., 2012).
onflicts of interest
None.
cknowledgements
The authors thank the following Brazilian agencies for finan-
ial support: Coordenac ão de Aperfeic oamento de Pessoal deível Superior (CAPES-Procad no. 131/2007), Conselho Nacionale Desenvolvimento Científico e Tecnológico (CNPq), Fundac ão demparo à Ciência e Tecnologia do Estado de Pernambuco (Facepescience 33 (2014) 98–105
– IBPG no. 0085-2.07/09), Instituto Brasileiro de Neurociências(IBN-Net/Finep no. 4191), and Instituto Nacional de Neurociên-cia Translacional (INCT no. 573604/2008-8). R.C.A. Guedes is aResearch Fellow from CNPq (no. 301190/2010-0).
References
Abraham, R., Dougherty, W., Golberg, L., Coulston, F., 1971. The response of thehypothalamus to high doses of monosodium glutamate in mice and monkeys.Cytochemistry and ultrastructural study of lysosomal changes. Exp. Mol. Pathol.15, 43–60.
Accioly, N.E., Benevides, R.D.L., Costa, B.L.S.A., Guedes, R.C.A., 2012. Ovariectomy inthe developing rat decelerates cortical spreading depression in adult brain. Int.J. Dev. Neurosci. 30, 405–410.
Akopian, G., Walsh, J.P., 2006. Pre- and postsynaptic contributions to age-relatedalterations in corticostriatal synaptic plasticity. Synapse 60, 223–238.
Almeida, M.F., Silveira, A.C., Guedes, R.C.A., 2005. Quantitative ultrastructuralevidence of myelin malformation in optic nerves of rats submitted to a mul-tideficient diet. Nutr. Neurosci. 8, 91–99.
Almeida, A.A., Gomes da Silva, S., Fernandes, J., Peixinho-Pena, L.F., Scorza, F.A.,Cavalheiro, E.A., Arida, R.M., 2013. Differential effects of exercise intensities inhippocampal BDNF, inflammatory cytokines and cell proliferation in rats duringthe postnatal brain development. Neurosci. Lett. 553, 1–6.
Amâncio-dos-Santos, A., Pinheiro, P.C.F., Lima, D.S.C., Ozias, M.G., Oliveira, M.B.,Guimarães, N.X., Guedes, R.C.A., 2006. Fluoxetine inhibits cortical spreadingdepression in weaned and adult rats suckled under favorable and unfavorablelactation conditions. Exp. Neurol. 200, 275–282.
Amancio-dos-Santos, A., Maia, L.M.S.S., Germano, P.C.P.S., Negrão, Y.D.S., Guedes,R.C.A., 2013. Tianeptine facilitates spreading depression in well-nourished andearly-malnourished adult rats. Eur. J. Pharmacol. 706, 70–75.
Amaral, A.P., Barbosa, M.S., Souza, V.C., Ramos, I.L., Guedes, R.C.A., 2009.Drug/nutrition interaction in the developing brain: dipyrone enhances spread-ing depression in rats. Exp. Neurol. 219, 492–498.
Andreazzi, A.E., Scomparin, D.X., Mesquita, F.P., Balbo, S.L., Gravena, C., De Oliveira,J.C., Rinaldi, W., Garcia, R.M.G., Grassiolli, S., Mathias, P.C.F., 2009. Swim-ming exercise at weaning improves glycemic control and inhibits the onset ofmonosodium l-glutamate-obesity in mice. J. Endocrinol. 201, 351–359.
Anish, E.J., 2005. Exercise and its effects on the central nervous system. Curr. SportsMed. Rep. 4, 18–23.
Ascensão, A., Martins, M.J., Santos-Alves, E., Gonc alves, I.O., Portincasa, P., Paulo,J., Oliveira, P.J., Magalhães, J., 2013. Modulation of hepatic redox status andmitochondrial metabolism by exercise: therapeutic strategy for liver diseases.Mitochondrion 13 (6), 862–870, http://dx.doi.org/10.1016/j.mito.2013.07.002.
Batista-de-Oliveira, M., Lopes, A.A.C., Mendes-da-Silva, R.F., Guedes, R.C.A., 2012.Aging-dependent brain electrophysiological effects in rats after distinct lacta-tion conditions, and treadmill exercise: a spreading depression analysis. Exp.Gerontol. 47, 452–457.
Beas-Zárate, C., Rivera-Huizar, S.V., Martinez-Contreras, A., Feria-Velasco, A.,Armendariz-Borunda, J., 2001. Changes in NMDA-receptor gene expression areassociated with neurotoxicity induced neonatally by glutamate in the rat brain.Neurochem. Int. 39, 1–10.
Black, J.E., Isaacs, K.R., Anderson, B.J., Alcantara, A.A., Greenough, W.T., 1990. Learningcauses synaptogenesis, whereas motor activity causes angiogenesis, in cerebel-lar cortex of adult rats. Proc. Natl. Acad. Sci. U. S. A. 87, 5568–5572.
Blomstrand, E., 2001. Amino acids and central fatigue. Amino Acids 20, 25–34.Brown, J., Cooper-Kuhn, C.M., Kempermann, G., Van, P.H., Winkler, J., Gage, F.H.,
Kuhn, H.G., 2003. Enriched environment and physical activity stimulate hip-pocampal but not olfactory bulb neurogenesis. Eur. J. Neurosci. 17, 2042–2046.
Cotman, C.W., Berchtold, N.C., 2002. Exercise: a behavioral intervention to enhancebrain health and plasticity. Trends Neurosci. 25, 295–301.
Cotman, C.W., Engesser-Cesar, C., 2002. Exercise enhances and protects brain func-tion. Exerc. Sport Sci. Rev. 30, 75–79.
Dohmen, C., Sakowitz, O.W., Fabricius, M., Bosche, B., Reithmeier, T., Ernestus, R.I.,Brinker, G., Dreier, J.P., Woitzik, J., Strong, A.J., Graf, R., 2008. Spreading depolar-izations occur in human ischemic stroke with high incidence. Ann. Neurol. 63,720–728.
Fabricius, M., Fuhr, S., Willumsen, L., Dreier, J.P., Bhatia, R., Boutelle, M.G., Hartings,J.A., Bullock, R., Strong, A.J., Lauritzen, M., 2008. Association of seizures with cor-tical spreading depression and peri infarct depolarization in the acutely injuredhuman brain. Clin. Neurophysiol. 119, 1973–1984.
Fang, Z.H., Lee, C.H., Seo, M.K., Cho, H., Lee, J.G., Lee, B.J., Park, S.W., Kim, Y.H., 2013.Effect of treadmill exercise on the BDNF-mediated pathway in the hippocampusof stressed rats. Neurosci. Res., http://dx.doi.org/10.1016/j.neures.2013.04.005.
Farmer, J., Zhao, X., Van, P.H., Wodtke, K., Gage, F.H., Christie, B.R., 2004. Effects ofvoluntary exercise on synaptic plasticity and gene expression in the dentategyrus of adult male Sprague-Dawley rats in vivo. Neuroscience 124, 71–79.
Fregni, F., Liebetanz, D., Monte-Silva, K.K., Oliveira, M.B., Santos, A.A., Nitsche, M.A.,Pascual-Leone, A., Guedes, R.C., 2007. Effects of transcranial direct current stim-
ulation coupled with repetitive electrical stimulation on cortical spreadingdepression. Exp. Neurol. 204, 462–466.Gomez-Pinilla, F., Ying, Z., Roy, R.R., Molteni, R., Edgerton, V.R., 2002. Voluntaryexercise induces a BDNF-mediated mechanism that promotes neuroplasticity.J. Neurophysiol. 88, 2187–2195.

Neuro
G
G
G
G
G
H
I
I
K
K
K
K
K
L
L
L
L
L
L
L
L
L
L
M
M
M
M
M
hippocampal neurogenesis in aged mice. J. Neurosci. 25, 8680–8685.
C.B. Lima et al. / Int. J. Devl
orji, A., 2001. Spreading depression: a review of the clinical relevance. Brain Res.Rev. 38, 33–60.
uedes, R.C.A., Cavalheiro, E.A., 1997. Blockade of spreading depression in chronicepileptic rats: reversion by diazepam. Epilepsy Res. 27, 33–40.
uedes, R.C.A., Andrade, A.F.D., Cavalheiro, E.A., 1988. Excitatory amino acids andcortical spreading depression. In: Cavalheiro, E.A., Lehman, J., Turski, L. (Eds.),Frontiers in Excitatory Amino Acid Research. Alan R. Liss. Inc., New York, NY, pp.667–670.
uedes, R.C.A., Santos, A.A., Manhães-de-Castro, R., Costa-Cruz, R.R.G., 2002.Citalopram has an antagonistic action on cortical spreading depression in well-nourished and early-malnourished adult rats. Nutr. Neurosci. 5, 115–123.
uedes, R.C.A., Oliveira, J.A., Amancio-dos-Santos, A., Garcia-Cairasco, N., 2009. Sex-ual differentiation of cortical spreading depression propagation after acute andkindled audiogenic seizures in the Wistar audiogenic rat (WAR). Epilepsy Res.83, 207–214.
illman, C.H., Belopolsky, A.V., Snook, E.M., Kramer, A.F., McAuley, E., 2004. Phys-ical activity and executive control: implications for increased cognitive healthduring older adulthood. Res. Q. Exerc. Sport 75, 176–185.
gelström, K.M., Heyward, P.M., 2012. The antidepressant drug fluoxetine inhibitspersistent sodium currents and seizure-like events. Epilepsy Res. 101, 174–181.
saacs, K.R., Anderson, B.J., Alcantara, A.A., Black, J.E., Greenough, W.T., 1992. Exerciseand the brain: angiogenesis in the adult rat cerebellum after vigorous physicalactivity and motor skill learning. J. Cereb. Blood Flow Metab. 12, 110–119.
aracabey, K., 2005. Effect of regular exercise on health and disease. Neuroen-docrinol. Lett. 26, 617–623.
arperien, A., Ahammer, H., Jelinek, H.F., 2013. Quantitating the subtleties ofmicroglial morphology with fractal analysis. Front. Cell. Neurosci. 7, 1–18.
im, J.-H., Kwak, H.-B., Leeuwenburgh, C., Lawler, J.M., 2008. Lifelong exercise andmild (8%) caloric restriction attenuate age-induced alterations in plantaris mus-cle morphology, oxidative stress and IGF-1 in the Fischer-344 rat. Exp. Gerontol.43, 317–329.
iss, P., Hauser, D., Tamas, A., Lubics, A., Racz, B., Horvath, Z., Farkas, J., Zimmer-mann, F., Stepien, A., Lengvari, I., Reglodi, D., 2007. Changes in open-field activityand novelty-seeking behavior in periadolescent rats neonatally treated withmonosodium glutamate. Neurotox. Res. 12, 85–93.
ohman, R.A., Bhattacharya, T.K., Wojcik, E., Rhodes, J.S., 2013. Exercise reduces acti-vation of microglia isolated from hippocampus and brain of aged mice. J. Neu-roinflammation. 2013; 10, 114, http://dx.doi.org/10.1186/1742-2094-10-114.
au, Y.S., Patki, G., Das-Panja, K., Le, W.D., Ahmad, S.O., 2011. Neuroprotective effectsand mechanisms of exercise in a chronic mouse model of Parkinson’s diseasewith moderate neurodegeneration. Eur. J. Neurosci. 33, 1264–1274.
eão, A.A.P., 1944. Spreading depression of activity in the cerebral cortex. J. Neuro-physiol. 7, 359–390.
eão, A.A.P., 1947. Further observations on the spreading depression of activity inthe cerebral cortex. J. Neurophysiol. 10, 409–414.
eão, A.A.P., 1972. Spreading depression. In: Purpura, D.P., Penry, K., Tower, D.B.,Woodbury, D.M., Walter, R.D. (Eds.), Experimental Models of Epilepsy. RavenPress, New York, pp. 173–195.
i, Y., Du, X.-F., Liu, C.-S., Wen, Z.-L., Jiu-lin Du, J.-L., 2012. Reciprocal regulationbetween resting microglial dynamics and neuronal activity in vivo. Dev. Cell 23,1189–1202.
iebetanz, D., Fregni, F., Monte-Silva, K.K., Oliveira, M.B., Amancio-dos-Santos, A.,Nitsche, M.A., Guedes, R.C.A., 2006. After-effects of transcranial direct currentstimulation (tDCS) on cortical spreading depression. Neurosci. Lett. 398, 85–90.
ima, C.B., Soares, G.S.F., Vitor, S.M., Castellano, B., Andrade-da-Costa, B.L.S., Guedes,R.C.A., 2013. Neonatal treatment with monosodium glutamate lastingly facili-tates spreading depression in the rat cortex. Life Sci. 93, 388–392.
inkenhoker, B.A., von der Ohe, C.G., Knudsen, E.I., 2005. Anatomical traces of juve-nile learning in the auditory system of adult barn owls. Nat. Neurosci. 8, 93–98.
opez-Perez, S.J., Urena-Guerrero, M.E., Morales-Villagran, A., 2010. Monosodiumglutamate neonatal treatment as a seizure and excitotoxic model. Brain Res.1317, 246–256.
ynch, G., Rex, C.S., Gall, C.M., 2006. Synaptic plasticity in early aging. Ageing Res.Rev. 5, 255–280.
aia, L.M.S.S., Amancio-dos-Santos, A., Duda-de-Oliveira, D., Angelim, M.K., Ger-mano, P.C.P.S., Santos, S.F., Guedes, R.C.A., 2009. l-Arginine administrationduring rat brain development facilitates spreading depression propagation: evi-dence for a dose- and nutrition-dependent effect. Nutr. Neurosci. 12, 73–80.
arrannes, R., Willems, R., DePrins, E., 1988. Evidence for a role of the NMDA recep-tor in cortical spreading depression in the rat. Brain Res. 457, 226–240.
artins-Ferreira, H., Nedergaard, M., Nicholson, C., 2000. Perspectives on spreadingdepression. Brain Res. Rev. 32, 215–234.
artinsen, E.W., 2008. Physical activity in the prevention and treatment of anxiety
and depression. Nord. J. Psychiatry Suppl. 47, 25–29.erkler, D., Klinker, F., Jurgens, T., Glaser, R., Paulus, W., Brinkmann, B.G., Sereda,M.W., Stadelmann-Nessler, C., Guedes, R.C.A., Bruck, W., Liebetanz, D., 2009.Propagation of spreading depression inversely correlates with cortical myelincontent. Ann. Neurol. 66, 355–365.
science 33 (2014) 98–105 105
Miranda, R.A., Branco, R.C., Gravena, C., Barella, L.F., da Silva Franco, C.C., Andreazzi,A.E., de Oliveira, J.C., Picinato, M.C., de Freitas Mathias, P.C., 2013. Swimtraining of monosodium l-glutamate-obese mice improves the impairedinsulin receptor tyrosine phosphorylation in pancreatic islets. Endocrine 43,571–578.
Montesi, L., Moscatiello, S., Malavolti, M., Marzocchi, R., Marchesini, G., 2013. Phys-ical activity for the prevention and treatment of metabolic disorders. Intern.Emerg. Med., 1–12, http://dx.doi.org/10.1007/s11739-013-0953-7.
Moraes-Silva, I.C., Mostarda, C., Moreira, E.D., Silva, K.A., dos Santos, F., de Ange-lis, K., Farah, V.M., Irigoyen, M.C., 2013. Preventive role of exercise training inautonomic, hemodynamic, and metabolic parameters in rats under high risk ofmetabolic syndrome development. J. Appl. Physiol. 114, 786–791.
Moskowitz, M.A., Bolay, H., Turgay Dalkara, T., 2004. Deciphering migraine mech-anisms: clues from familial hemiplegic migraine genotypes. Ann. Neurol. 55,276–280.
Moura, L.P., Sponton, A.C., de Araújo, M.B., Dalia, R.A., Pauli, J.R., Rostom de Mello,M.A., 2013. Moderate physical activity from childhood contributes to metabolichealth and reduces hepatic fat accumulation in adult rats. Lipids Health Dis. 6,12–29.
Musa, M., Sunday, A.M., 2013. Clumping of the nuclei material of pyramidal cells ofadult Wistar rats following oral administration of monosodium glutamate. Res.Rev. J. Med. Health Sci. 2, 10–15.
Nemeroff, C.B., Lipton, M.A., Kizer, J.S., 1978. Models of neuroendocrine regula-tion: use of monosodium glutamate as an investigational tool. Dev. Neurosci.1, 102–109.
Peixinho-Pena, L.F., Fernandes, J., de Almeida, A.A., Novaes Gomes, F.G., Cassilhas, R.,Venancio, D.P., de Mello, M.T., Scorza, F.A., Cavalheiro, E.A., Arida, R.M., 2012. Astrength exercise program in rats with epilepsy is protective against seizures.Epilepsy Behav. 25, 323–328.
Pellegrini-Giampietro, D.E., Gorter, J.A., Bennett, M.V., Zukin, R.S., 1997. The GluR2(GluR-B) hypothesis: Ca(2+)-permeable AMPA receptors in neurological disor-ders. Trends Neurosci. 20, 464–470.
Perry, V.H., Teeling, J., 2013. Microglia and macrophages of the central nervoussystem: the contribution of microglia priming and systemic inflammation tochronic neurodegeneration. Semin. Immunopathol. 35, 601–612.
Pysh, J.J., Weiss, G.M., 1979. Exercise during development induces an increase inPurkinje cell dendritic tree size. Science 206, 230–232.
Rocha-de-Melo, A.P., Cavalcanti, J.B., Barros, A.S., Guedes, R.C.A., 2006. Manipulationof rat litter size during suckling influences cortical spreading depression afterweaning and at adulthood. Nutr. Neurosci. 9, 155–160.
Rothman, S.M., Mattson, M.P., 2013. Activity-dependent, stress-responsive BDNFsignaling and the quest for optimal brain health and resilience throughout thelifespan. Neuroscience 239, 228–240.
Shen, H., Tong, L., Balazs, R., Cotman, C.W., 2001. Physical activity elicits sus-tained activation of the cyclic AMP response element-binding protein andmitogen-activated protein kinase in the rat hippocampus. Neuroscience 107,219–229.
Siesjö, B.K., Bengtsson, F., 1989. Calcium fluxes, calcium antagonists, and calcium-related pathology in brain ischemia, hypoglycemia, and spreading depression:a unifying hypothesis. J. Cereb. Blood Flow Metab. 9, 127–140.
Soya, H., Nakamura, T., Deocaris, C.C., Kimpara, A., Iimura, M., Fujikawa, T., Chang,H., McEwen, B.S., Nishijima, T., 2007. BDNF induction with mild exercise in therat hippocampus. Biochem. Biophys. Res. Commun. 358, 961–967.
Swain, R.A., Harris, A.B., Wiener, E.C., Dutka, M.V., Morris, H.D., Theien, B.E., Konda, S.,Engberg, K., Lauterbur, P.C., Greenough, W.T., 2003. Prolonged exercise inducesangiogenesis and increases cerebral blood volume in primary motor cortex ofthe rat. Neuroscience 117, 1037–1046.
Swanson, C.J., Bures, M., Johnson, M.P., Linden, A.M., James, A., Monn, J.A., Schoepp,D.D., 2005. Metabotropic glutamate receptors as novel targets for anxiety andstress disorders. Nat. Rev. Drug Discov. 4, 131–144.
Tong, L., Shen, H., Perreau, V.M., Balazs, R., Cotman, C.W., 2001. Effects of exer-cise on gene-expression profile in the rat hippocampus. Neurobiol. Dis. 6,1046–1056.
Torrente, D., Mendes-da-Silva, R.F., Lopes, A.A.C., González, J., Barreto, G.E.,Guedes, R.C.A., 2013. Increased Calcium influx triggers and accelerates corti-cal spreading depression in vivo in male adult rats. Neurosci. Lett. 558, 87–90,http://dx.doi.org/10.1016/j.neulet.2013.11.004.
Urena-Guerrero, M.E., Orozco-Suárez, S., López-Pérez, S.J., Flores-Soto, M.E., Beas-Zárate, C., 2009. Excitotoxic neonatal damage induced by monosodiumglutamate reduces several GABAergic markers in the cerebral cortex and hip-pocampus in adulthood. Int. J. Dev. Neurosci. 27, 845–855.
Van Praag,.H., Shubert, T., Zhao, C., Gage, F.H., 2005. Exercise enhances learning and
Zhang, Y., Zhang, P., Shen, X., Tian, S., Wu, Y., Zhu, Y., Jia, J., Wu, J., Hu, H.,2013. Early exercise protects the blood–brain barrier from ischemic braininjury via the regulation of MMP-9 and occludin in rats. Int. J. Mol. Sci. 14,11096–11112.




















