
Determination of Capsaicin and Dihydrocapsaicin in ... - MDPI

Autonomic Neuroscience: Basic and Clinical 86 (2000) 37–46www.elsevier.com/ locate /autneu
Peptide immunoreactivity and ultrastructure of rat urinary bladder nervefibers after topical desensitization by capsaicin or resiniferatoxin
a a,b ,*´Antonio Avelino M.Sc. , Francisco Cruz M.D., Ph.D.aInstitute of Histology and Embryology, Faculty of Medicine of Oporto, IBMC of University of Oporto, 4200 Oporto, Portugal
b ˜ ˜Department of Urology, Hospital Sao Joao, Porto, Portugal
Received 14 April 2000; received in revised form 26 July 2000; accepted 23 August 2000
Abstract
In the present study the decrease of neuropeptide containing nerve fibers and the increase in the volume threshold to reflex micturitionoccurring in the rat bladder after intravesical application of capsaicin or resiniferatoxin were compared. The ultrastructure of bladderterminal axons was evaluated at the moment of maximal peptide depletion and compared to that of nerve fibers after systemic capsaicinapplication. Adult Wistar rats were treated intravesically for 30 min with 0.5 ml of 100 nM RTX, 1 mM capsaicin or 30% ethanol insaline, the vehicle solution. Twenty-four hours and 1, 2, 3, 4 and 8 weeks later the bladders were immunostained for CGRP, SP, VIP andNPY. Cystomanometric studies were performed 24 h and 1, 8, and 12 weeks after vanilloid instillation. Twenty-four hours after systemiccapsaicin or intravesical capsaicin or RTX, bladders were prepared for electron microscopic (EM) observation. Intravesical capsaicin orRTX decreased, in a similar way, the number of CGRP and SP-IR (immunoreactive) fibers coursing in the muscular layer and the mucosa.IR fibers amounted to less than 20% of controls at 24 h and returned to normal levels in the eighth week. At the EM level, bladderstreated with topical vanilloids did not show morphological changes in terminal axons coursing in the mucosa. In contrast, bladders fromanimals treated systemically with capsaicin contained numerous grossly degenerated nerve fibers. VIP and NPY-IR fibers were notaffected by the treatment. Cystometrograms showed an increase of the volume threshold to reflex micturition that started at 24 h anddisappeared at 12 weeks. We conclude that intravesical capsaicin or RTX were equally effective in terms of reducing the number of SPand CGRP-IR fibers and increasing the volume threshold for reflex micturition. Both changes were transient and were not associated withultrastructural changes of the bladder nerve fibers, excluding terminal axon degeneration as the main mechanism of action of intravesicalvanilloids. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Desensitization; Bladder; Rat; Capsaicin; Resiniferatoxin; Neuropeptides; Ultrastructure
1. Introduction (IR) nerve fibers was also found in the human epidermisafter prolonged topical capsaicin application (Nolano et al.,
Topical application of capsaicin to the urinary bladder 1999). In this case the disappearance of the neuropeptideshas been shown to induce a marked sensory neuropeptide would result from the loss of nerve endings and eventualdepletion, namely SP or CGRP (Maggi et al., 1988, 1989). peptide re-accumulation would be dependent upon nerveThe mechanisms involved are, however, still unclear. The sprouting. In contrast, other studies failed to recognizeoccurrence of nerve degeneration after intravesical cap- nerve degeneration after topical capsaicin. Repeated appli-saicin has recently been suggested following the observa- cation of capsaicin to the rat skin did not decrease eithertion that nerve fibers immunoreactive to the pan-neuronal the number of sensory axons coursing in the spinal dorsalmarker protein gene product (PGP 9.5) were drastically roots or the number of small DRG neurons (McMahon etreduced in the subepithelial layer of human bladders al., 1991). In the rabbit cornea, topical capsaicin causedinstilled with the neurotoxin (Dasgupta et al., 2000). A only minor alterations, such as mitochondrial swelling,similar reduction in the number of PGP immunoreactive cristae disruption and some reduction of synaptic vesicles
in axon terminals, but never overt axonal degeneration´(Szolcsanyi et al., 1975).
*Corresponding author. Institute of Histology and Embryology, FacultyIn the present study this problem was re-addressed in theof Medicine of Oporto, IBMC of University of Oporto, 4200 Oporto,
rat bladder with the aid of electron microscopy to de-Portugal. Tel.: 1351-2-509-1468; fax: 1351-2-550-5728.E-mail address: [email protected] (F. Cruz). termine the occurrence of degenerative nerve changes. The
1566-0702/00/$ – see front matter 2000 Elsevier Science B.V. All rights reserved.PI I : S1566-0702( 00 )00204-6

38 A. Avelino, F. Cruz / Autonomic Neuroscience: Basic and Clinical 86 (2000) 37 –46
magnitude and duration of SP or CGRP nerve fiber 0.1 M phosphate buffer, pH 7.4, and cryoprotected for 24 hreduction in the mucosa and the muscular layer was in 30% sucrose in phosphate buffer. Forty micrometerdetermined after a single intravesical application of cap- transverse sections were cut in a freezing microtome andsaicin or RTX. Full desensitizing concentrations of both stored in a cryoprotective solution at 2208C (Lu andvanilloids (Avelino et al., 1999; Cruz et al., 1996), Haber, 1992). When all animal experiments were com-presently under evaluation in clinical trials (Cruz et al., pleted, all the stored sections were washed with PBS and1997a,b; Silva et al., 2000), were used. The ultrastructure incubated overnight at room temperature in the sameof subepithelial unmyelinated terminal axons was ex- batches with polyclonal antisera against CGRP (Amer-amined at the moment of maximal neuropeptide depletion. sham, 1:40 000), SP, VIP and NPY (Peninsula Labs,The changes brought about by the intravesical vanilloid 1:5000). The immune reaction was visualized by thetreatment on the micturition reflex were investigated by avidin–biotin method (Vector Elite ABC-HRP) usingcystometry. Preliminary results have been presented in diaminobenzidine as chromogen.abstract form (Avelino et al., 1998). As immunoreactive fiber density is unequally distributed
throughout the bladder wall, only bladder sections passingthrough the trigone (two per animal) were used to quantify
2. Materials and methods immunoreactivity. This was ascertained by the presence ofthe intramural part of the ureters in the selected sections.
2.1. Animals and drugs Six images from the mucosa and another six from themuscular layer were digitally acquired with a frame
Adult male and female Wistar rats from the Gulbenkian grabber in each section and laser printed at a finalFoundation Colony weighing 220–250 g were used. Cap- magnification of 3203. The immunoreactive profiles oc-saicin and resiniferatoxin were purchased from Sigma and curring in each print were overwritten with a digital pen indiluted in 30% ethanol in saline. All the non-terminal a digital table and converted into black lines and dots on aanimal manipulations were performed under halothane white background. The number of black pixels wasanaesthesia (4% for induction and 2% for maintenance). counted by a personal computer program. Results areIntraperitoneal 35% chloral hydrate (1 ml /kg weight) was shown as the mean area occupied by black pixels.used as anaesthetic in terminal procedures with the excep-tion of cystometrograms which were performed under 2.3. Cystometrogramsurethane anaesthesia (1.2 g /kg, subcutaneous). All theexperimental manipulations followed the guidelines of the For these experiments only female rats were used. RatsInternational Association for the Study of Pain were anaesthetized with halothane and the bladder(Zimermann, 1983). catheterized through the urethra and completely emptied.
They were thereafter instilled with 1 mM capsaicin or 1002.2. Determination of SP, CGRP VIP and NPY nerve nM RTX solutions in 30% ethanol in saline. Cystometro-density grams were performed under urethane anesthesia 24 h, 1
week, 8 weeks and 12 weeks after treatment using 4Male and female rats were used in these experiments. In animals for each time point. In addition, cystometrograms
male rats, the bladder was exposed through a low midline were obtained from 4 intact rats and 4 rats treated onlyabdominal incision and a 20-gauge needle inserted in the with the vehicle solution 24 hours before. The bladdersbladder dome. In female rats, bladders were catheterized were exposed through a midline abdominal incision and awith a 22-gauge polyethylene catheter inserted through the 20-gauge needle were inserted into the lumen through theurethra. Urine was aspirated and the bladders were filled dome and connected to a pressure transducer (Gouldwith 0.5 ml of a 1 mM capsaicin or 100 nM RTX solution Instruments, USA) and an infusion pump. The rats werein 30% ethanol in saline. The solutions were left in contact placed on a heating pad and body temperature keptwith the mucosa for 30 min. The solutions were then between 36.5 and 378C. After a 30-min equilibrationremoved and the bladders rinsed with saline. Finally, the period, saline was infused at 3 ml /h and bladder pressureanimals were allowed to recover after closure of the recorded until the first voiding contraction leading toabdominal wound in male rats or removal of the urethral micturition occurred. The infused volume was taken as thecatheter in female rats. volume threshold to reflex micturition. Three cystometrog-
Twenty-four hours and 1, 2, 3, 4 and 8 weeks after rams were obtained from each animal.treatment the animals were re-anaesthetized and bladdersremoved. For each time point, two males and three females 2.4. Electron microscopic study of the subepithelialwere used. In addition, the bladders of two males and three terminal axons after systemic capsaicin treatment orfemales instilled 24 h before with 30% ethanol in saline intravesical capsaicin or RTX application
¨(the vehicle solution) and the bladders of two naıve males¨and three naıve females were obtained. Bladders were For systemic capsaicin treatment, two adult female rats
fixed overnight by immersion in 4% paraformaldehyde in received 100 mg/kg body weight of capsaicin as a single

A. Avelino, F. Cruz / Autonomic Neuroscience: Basic and Clinical 86 (2000) 37 –46 39
subcutaneous injection behind the neck under halothane reduced by more than 80% both in the mucosa and theanaesthesia. The rats were pre-medicated with theophylline muscular layer, the difference being highly significant(5 mg/kg, i.p.), atropine (2 mg/kg, i.p.) and respiratory (P , 0.001) in all experiments (see Fig. 2A–D). At 1 and 2nebulization with salbutamol (0.5 ml of salbutamol in 5 ml weeks after capsaicin or RTX instillation the area ofof saline). For intravesical instillation, female rats were CGRP- and SP-IR fibers was still very small, aroundanaesthetized with halothane and the bladder filled with 25–30% of controls (Fig. 2A–D). From this time point on0.5 ml of a 30% ethanol solution (two animals) or with the a gradual increase of CGRP- or SP-IR fibers occurred. At 4same solution containing 1 mM capsaicin (two animals) or weeks, the number of IR profiles was still significantly100 nM RTX (two animals) as described above. Twenty- different from the control values in the mucosa in allfour hours later the rats were re-anaesthetized and the experiments and in the muscular layer for SP in capsaicin-bladders filled in situ with 0.5 ml of 5% glutaraldehyde in and for CGRP in RTX-treated animals. Control valuesphosphate buffer injected through the dome and then were reached by the eighth week after capsaicin (Figs.removed and cut into small longitudinal strips which were 1D,D9 and 2A,B) or RTX (Figs. 1E,E9 and 2C,D) treat-left overnight in the same fixative at 48C. The bladder ment with the exception of SP in the mucosa of RTX-strips were washed with 0.1 M phosphate buffer, post-fixed treated animals.
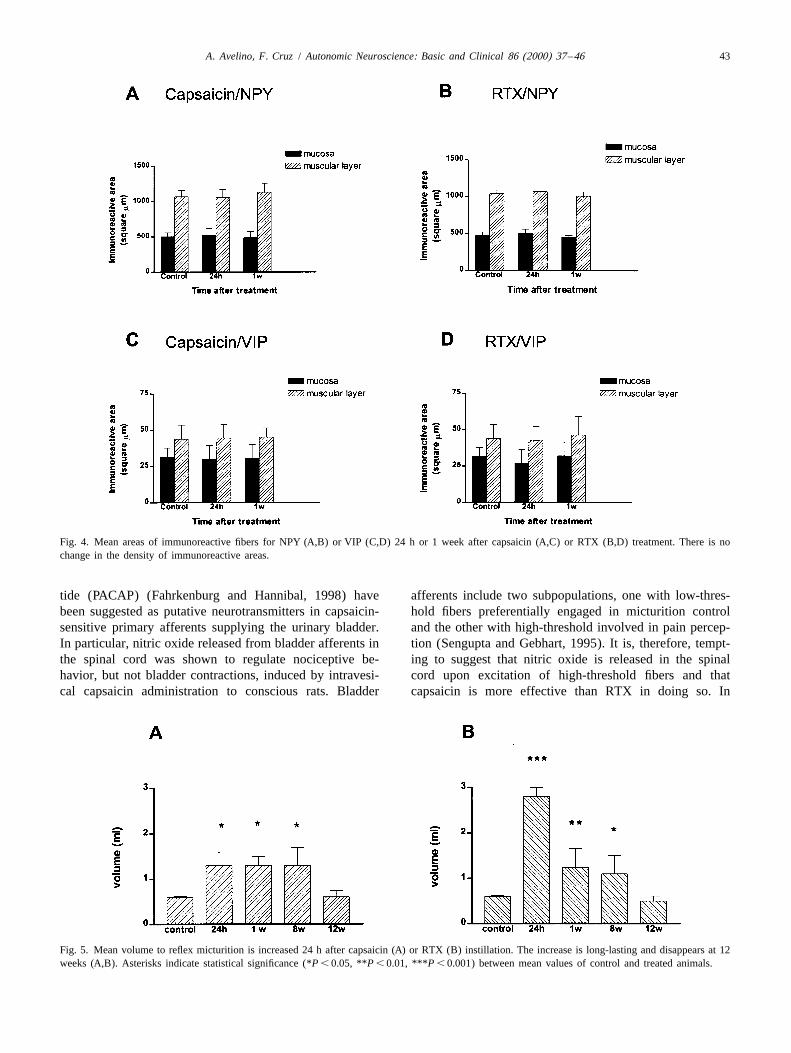
¨for 1 h in 1% osmium tetroxide in phosphate buffer, In naıve or vehicle-treated rats, NPY-IR fibers alsodehydrated in graded ethanol and embedded in Epon formed two dense networks (Fig. 3A). The immuno-
¨(Gabella, 1995). In addition, the bladders of two naıve rats reactive area in the mucosa or in the muscular layerwere similarly fixed and prepared for EM analysis. Three measured 24 h or 1 week after intravesical application ofblocks of the mucosa from the trigonal region were capsaicin or RTX was unchanged (Figs. 3B and 4A,B).prepared from each bladder and cut into ultrathin silver VIP-IR fibers were very scarce when compared to thesections in an LKB ultramicrotome. Sections were stained other IR fibers studied here. The area of VIP-IR fibers in
¨with lead citrate and uranyl acetate and observed in a Jeol controls (naıve and vehicle-treated animals) was alsoelectron microscope. unchanged by intravesical instillation of capsaicin or RTX
(Fig. 4C,D).2.5. Statistical analysis
3.2. CystometrogramsAll numeric data are shown as mean6standard devia-
¨tion. When indicated, data were compared by one-way Cystometrograms obtained from naıve rats or from ratsANOVA followed by the post-hoc Bonferroni test for treated with the vehicle solution were identical and were,multiple comparisons. therefore, included in a control group. Twenty-four hours
after intravesical capsaicin the mean volume threshold toreflex micturition increased from 0.660.02 to 1.360.3 ml
3. Results (P , 0.01, Fig. 5A). Twenty-four hours after RTX theincrement was even more notable, the mean bladder
3.1. Distribution of CGRP, SP, VIP and NPY volume being 2.860.2 ml (P , 0.001, Fig. 5B). The¨immunoreactive fibers in naıve, vehicle-treated and volume threshold to reflex micturition was still above that
vanilloid-treated animals of control animals at the eighth week after capsaicin orRTX treatment (Fig. 5A,B). At the 12th week after
Varicose CGRP- or SP-IR fibers formed distinct net- instillation, it became similar to that of controls (Fig.¨works in the mucosa and the muscular layer of naıve and 5A,B).
vehicle-treated animals, seemingly non-affected by animalgender. In the mucosa, immunoreactive fibers ran beneath 3.3. Electron microscopyand within the epithelium or in close proximity to bloodvessels (Fig. 1A,A9). In the muscular layer, CGRP- and A total of 806 nerve profiles were photomicrographed inSP-IR fibers ran parallel to the muscle fibers (Fig. 1A,A9). the subepithelial layer of intact, vehicle-treated and van-
¨As the nerve density of the fiber networks of naıve or illoid-treated bladders. In the former, numerous unmyeli-vehicle-treated animals was similar, data obtained from nated axons occurred isolated or grouped in thin nerveboth series was included in a single control group (Fig. 2). bundles wrapped in by a single Schwann cell (Fig. 6A).
Changes of CGRP or SP nerve immunoreactivity in- Some axonal profiles contained synaptic vesicles (Fig. 6A).duced by intravesical application of capsaicin or RTX Following instillation of 30% ethanol, most of the nervefollowed a similar pattern. Twenty-four hours after cap- profiles in the lamina propria looked normal. However, insaicin (Fig. 1B,B9) or RTX (Fig. 1C,C9), CGRP- and 10% of the profiles some morphological alterations couldSP-IR fibers were nearly absent from the entire depth of be detected, ranging from mitochondrial disruption to anthe bladder wall. irregular densification of the cytoplasm (Fig. 6B). Follow-
Twenty-four hours after capsaicin or RTX instillation ing the instillation of capsaicin or RTX solutions the largethe area occupied by the immunoreactive profiles was majority of the nerve profiles looked normal (Fig. 7A,B).

40 A. Avelino, F. Cruz / Autonomic Neuroscience: Basic and Clinical 86 (2000) 37 –46
Fig. 1. Photomicrographs of the bladder wall immunostained for CGRP (A–E) and SP (A9–E9). Two distinct plexuses are evident in the mucosa andmuscular layer of vehicle-treated animals (A,A9). Twenty-four hours after capsaicin (B,B9) or RTX (C,C9) treatment a massive disappearance ofimmunoreactivity occurs in both layers. The density of CGRP and SP immunoreactive fibers returned to control levels 8 weeks after capsaicin (D,D9) orRTX (E,E9) treatment. Calibration bar 100 mm.

A. Avelino, F. Cruz / Autonomic Neuroscience: Basic and Clinical 86 (2000) 37 –46 41
Fig. 2. Mean areas of immunoreactive fibers for CGRP (A,C) and SP (B,D). Following capsaicin (A,B) or RTX (C,D) treatment a massive decrease of theimmunoreactivity for the two peptides occurs both in the mucosa and muscular layer. At 8 weeks post-treatment the immunoreactive area for CGRP or SPreturned to control levels. Asterisks indicate statistical significance (*P , 0.05, **P , 0.01, ***P , 0.001) between mean values of control and treatedanimals.
However, the axoplasmic changes recognized in the ve- control values. Intravesical capsaicin or RTX did nothicle-treated bladders occurred similarly after intravesical change the density of VIP- or NPY-IR fibers in the bladdercapsaicin or RTX instillation, affecting 11 and 9% of the wall.profiles, respectively. Systemic capsaicin administrationcaused the most evident morphological changes. About 4.1. Topical desensitization: functional process rather35% of the unmyelinated profiles were shrunken and than selective dennervationdarkened or contained disrupted mitochondria and roun-dish dense structures of different sizes in their cytoplasm In neonatal animals, systemic capsaicin induces a(Fig. 7C). permanent loss of SP or CGRP sensory fibers (Sharkey et
al., 1983; Su et al., 1986) as a consequence of a decreasein the total number of small dorsal root ganglia (DRG)
´4. Discussion neurons (Jancso et al., 1977; Sharkey et al., 1983). Nervelesions can be recognized ultrastructurally as soon as 2 h
The present study shows that the almost complete after neurotoxin administration (Nagy et al., 1980). Indisappearance of SP- and CGRP-IR nerve fibers from the adult rats, systemic capsaicin does not appreciably de-mucosa and muscular layer 24 h after intravesical cap- crease the number of SP- and CGRP-IR DRG neuronssaicin or RTX in full desensitizing concentrations (Avelino (Kashiba et al., 1997). Nevertheless, degenerative changeset al., 1999; Cruz et al., 1996) is not accompanied by nerve could be recognized in the terminal part of primarydegeneration. Neuropeptide immunoreactive nerve fibers afferent axons innervating the ureter 24 h after subcuta-increased gradually thereafter in the two layers and their neous capsaicin administration (Chung et al., 1985).number achieved control values 8 weeks after treatment. Similar neurotoxic effects of systemic capsaicin wereFour weeks later the bladder volume to reflex micturition, observed in the present work. Twenty-four hours afterwhich had increased markedly at 24 h, also returned to subcutaneous administration of the neurotoxin, numerous

42 A. Avelino, F. Cruz / Autonomic Neuroscience: Basic and Clinical 86 (2000) 37 –46
considered. PGP is a neuron-specific hydrolase universallypresent in the axons of normal nerves and involved inneuronal responses to stressful events (Wilkinson et al.,1989). It is produced in cell bodies and transported to theperiphery at very low rates exclusively by the slowcomponent of the anterograde transport system (Bizzi etal., 1991). As desensitization causes axonal transportblockade in sensory fibers (Taylor et al., 1984), this mayfurther slow down the arrival of PGP to peripheral axons.
If intravesical capsaicin or RTX does not degenerateterminal axons, excluding denervation as the origin ofpeptide decrease and nerve sprouting as the mechanism forpeptide reappearance, it remains unexplained why the SPand CGRP decrease lasted for 8 weeks. The abruptdisappearance of almost all immunoreactive fibers at 24 his better explained by the massive release of SP or CGRPfrom nerve endings. Peptide release from bladder nerveswas shown to occur following vanilloid application (Maggiet al., 1988, 1990), possibly due to calcium inflow into theneurons (Maggi, 1991). However, a prolonged effect callsfor another explanation. The blockade of axonal transport(Taylor et al., 1984) may provide an important contribu-tion by hampering the orthograde transport of neuro-peptides. In addition, it reduces the retrograde transport ofnerve growth factor required for maintenance of neuro-Fig. 3. Photomicrographs of the bladder wall immunostained for NPY 24peptide levels in some sensory neurons (Miller et al.,h after vehicle (A) or RTX (B) instillation. There is no change in the
density of immunoreactive fibers. Calibration bar 100 mm. 1982). The recent observation that systemic capsaicin andRTX reduce the expression of preprotachykinin and CGRPmessenger RNAs in sensory neurons in adult animals
terminal axons darkened and devoid of organelles could be further supports this hypothesis (Farkas-Szallasi et al.,seen in the bladder wall. 1995; Kashiba et al., 1997).
In contrast, such ultrastructural changes were absentfrom terminal axons 24 h after a single intravesical 4.2. Peptide release and bladder irritationapplication of capsaicin or RTX. This must, therefore, betaken as strong evidence against nerve degeneration. We The acute release of tachykinins contained in peripheraldid not investigate the occurrence of ultrastructural sensory endings was shown to contribute to bladderchanges at a later time point. However, this possibility hyperactivity and pain occurring immediately after intra-seems unlikely for two reasons. First, a 24 h interval was vesical application of vanilloids. Bladder contractionsused in systemic capsaicin experiments which was suffi- could be blocked by the intra-arterial injection near thecient for degeneration to show up using the electron bladder of the selective NK2 tachykinin receptor antago-microscope (Chung et al., 1985). Second, it was the time at nists, SR48968 (Ishisuka et al., 1994, 1995) or MENwhich neuropeptide immunoreactivity loss was more in- 11,420 (Lecci et al., 1997). Similarly, the number of timestense. that conscious rats licked their lower abdomen after
The lack of degenerative nerve changes after intravesical intravesical 100 nM RTX could also be reduced by thevanilloids agrees with previous findings in the rabbit same intra-arterial injection of SR 48968 (Ishisuka et al.,cornea where topical capsaicin caused mitochondrial swell- 1995). Therefore, the similar decrease of SP- or CGRP-IRing, cristae disruption and some reduction of synaptic fibers in the mucosa and the muscular layer 24 h aftervesicles in axon terminals but no axoplasmic densification intravesical capsaicin or RTX application, exceeding 80%
´(Szolcsanyi et al., 1975). However, recent studies have of control values, was surprising. In fact, we have shownsuggested that topical capsaicin could induce terminal axon that intravesical 1 mM capsaicin was extremely painful todegeneration based on the temporary loss of PGP-IR fibers humans (Cruz et al., 1997b) and rats (Cruz et al., 1996),in the human bladder (Dasgupta et al., 2000) or skin while 100 nM RTX was only moderately irritant (Avelino(Nolano et al., 1999). It should, however, be remembered et al., 1999; Cruz et al., 1997a; Silva et al., 2000).that no ultrastructural confirmation of axon degeneration Recent studies offer alternative explanations for thewas obtained in those studies. The possibility that PGP-IR extreme pungency of capsaicin. Nitric oxide (Pandita et al.,was lost without axon degeneration should, therefore, be 1997) and pituitary adenylate cyclase activating polypep-
A. Avelino, F. Cruz / Autonomic Neuroscience: Basic and Clinical 86 (2000) 37 –46 43
Fig. 4. Mean areas of immunoreactive fibers for NPY (A,B) or VIP (C,D) 24 h or 1 week after capsaicin (A,C) or RTX (B,D) treatment. There is nochange in the density of immunoreactive areas.
tide (PACAP) (Fahrkenburg and Hannibal, 1998) have afferents include two subpopulations, one with low-thres-been suggested as putative neurotransmitters in capsaicin- hold fibers preferentially engaged in micturition controlsensitive primary afferents supplying the urinary bladder. and the other with high-threshold involved in pain percep-In particular, nitric oxide released from bladder afferents in tion (Sengupta and Gebhart, 1995). It is, therefore, tempt-the spinal cord was shown to regulate nociceptive be- ing to suggest that nitric oxide is released in the spinalhavior, but not bladder contractions, induced by intravesi- cord upon excitation of high-threshold fibers and thatcal capsaicin administration to conscious rats. Bladder capsaicin is more effective than RTX in doing so. In
Fig. 5. Mean volume to reflex micturition is increased 24 h after capsaicin (A) or RTX (B) instillation. The increase is long-lasting and disappears at 12weeks (A,B). Asterisks indicate statistical significance (*P , 0.05, **P , 0.01, ***P , 0.001) between mean values of control and treated animals.

44 A. Avelino, F. Cruz / Autonomic Neuroscience: Basic and Clinical 86 (2000) 37 –46
¨Fig. 6. Electron micrographs of naıve (A) and vehicle-treated animals¨(B). In naıve animals, most terminal axons had clear axoplasm containing
mitochondria, microtubules and microfilaments. Some were full ofspherical synaptic vesicles. Twenty-four hours after instillation of vehicle(B) a few terminal axons presented signs of injury, with darkenedcytoplasm and disrupted organelles. Calibration bar 0.28 mm.
addition, capsaicin, but not RTX, may activate non-neuro-nal inflammatory mechanisms directly, as suggested by its
Fig. 7. Electron micrographs of capsaicin (A) and RTX (B) topicallycapacity to release interleukin-4, a potent pro-inflammatorytreated bladders. Following both vanilloids the large majority of terminal´ ´product, from mast cells (Bıro et al., 1998). axons continued to present a normal ultrastructure. After systemicadministration of capsaicin (C), degenerating nerve profiles were abun-
4.3. Decrease of SP and CGRP containing sensory dant. Calibration bar 0.28 mm.
fibers and increment of the volume threshold to reflexmicturition
Intravesical application of capsaicin or RTX induced, in does not affect the volume threshold to micturition in thethe tested concentrations, a similar increase of the volume normal bladder (Lecci et al., 1993b).threshold to reflex micturition which started at 24 h and In spite of this, a strict inverse correlation between thedisappeared at 12 weeks. As is it known from previous amount of SP in the bladder wall and the volume thresholdstudies that the urinary bladder capacity of adult rats does to reflex micturition was found in adult rats 7 days afternot increase with age (Chun et al., 1989; Lluel et al., subcutaneous capsaicin administration (Maggi et al.,2000), the increment observed here was certainly due to 1989). A similar correlation was also apparent in the firstthe vanilloid action on bladder sensory afferents. part of our study. Since peptides are produced in DRG cell
This long-lasting increment has been attributed to the bodies from which they migrate to both central anddecreased availability of SP in central primary afferent peripheral endings, it is possible that bladder levels may, atterminations, which would impair bladder sensory trans- least partially, reflect those in spinal endings. However,mission at the spinal cord level. In fact, intrathecal during the last 4 weeks of our study, the volume thresholdadministration of NK1 receptor antagonists inhibited the to reflex micturition remained elevated, although thedistension-induced bladder contractions in a way indis- number of SP-IR fibers in the bladder was already normal.tinguishable from that induced by capsaicin (Lecci et al., This may be explained by a four times faster axonal1993a). The disappearance of SP or CGRP from the transport of neuropeptides towards the periphery than tobladder wall seems a less plausible explanation for the the central endings (Harmar and Keen, 1982). Interesting-observed urodynamic changes. Previous studies have ly, previous studies have already suggested that, in bladdershown that the blockade of peripheral tachykinin receptors primary afferents, the recovery of the sensory function

A. Avelino, F. Cruz / Autonomic Neuroscience: Basic and Clinical 86 (2000) 37 –46 45
Avelino, A., Cruz, F., Coimbra, A., 1999. Intravesical resiniferatoxinafter systemic capsaicin was slower than that of thedesensitizes rat bladder sensory fibres without causing intense noxiousefferent function (Maggi et al., 1987).excitation. A c-fos study. Eur. J. Pharmacol. 378, 17–22.
Bizzi, A., Schaetzle, B., Patton, A., Gambetti, P., Autilio-Gambetti, L.,4.4. VIP and NPY invulnerability 1991. Axonal transport of two major components of the ubiquitin
system: free ubiquitin and ubiquitin carboxyl-terminal hydrolase PGP9.5. Brain Res. 548, 292–299.VIP-IR fibers were reported in postganglionic para-
´´ ´Bıro, T., Maurer, M., Modarres, S., Lewin, N.E., Brodie, C., Acs, G.,sympathetic neurons (Alm et al., 1977) and in capsaicin-Acs, P., Paus, R., Blumberg, P.M., 1998. Characterization of func-´sensitive primary afferents (Jancso et al., 1981) coursing intional vanilloid receptors expressed by mast cells. Blood 91, 1332–
the hypogastric and pelvic nerves (Keast and DeGroat, 1340.1992). VIP sensory fibers are, however, so scarce that Chun, A.L., Wallace, U., Gerald, M.C., Wein, A.J., Levin, R.M., 1989.
Effects of age on urinary bladder function in the male rat. J. Urol.capsaicin administration could not significantly reduce the141, 170–173.total number of VIP-IR fibers in the bladder of newborn
Chung, K., Schewn, R.J., Coggeshall, R.E., 1985. Ureteral axon damagerats (Mattiasson et al., 1985). Accordingly, we also did notfollowing subcutaneous administration of capsaicin in adult rats.
find any decrease in VIP-IR fibers after the intravesical Neurosci. Lett. 53, 221–226.administration of capsaicin or RTX. Cruz, F., Avelino, A., Coimbra, A., 1996. Desensitization follows
excitation of bladder primary afferents by intravesical capsaicin, asBladder NPY immunoreactivity can be depleted by 6-shown by c-fos activation in the rat spinal cord. Pain 64, 553–557.hydroxydopamine or postganglionic sympathetic denerva-
˜Cruz, F., Guimaraes, M., Silva, C., Reis, M., 1997a. Suppression oftion, but not by capsaicin treatment of newborn ratsbladder hyperreflexia by intravesical resiniferatoxin. Lancet 350, 640–
(Mattiasson et al., 1985), indicating the exclusive presence 641.of this peptide in postganglionic sympathetic fibers, where ˜Cruz, F., Guimaraes, M., Silva, C., Rio, M.E., Coimbra, A., Reis, M.,
1997b. Desensitization of bladder sensory fibers by intravesicalit colocalizes with noradrenaline (Lundberg and Hokfelt,capsaicin has long lasting clinical and urodynamic effects in patients1986). Therefore, it was not surprising that intravesicalwith hyperactive or hypersensitive bladder dysfunction. J. Urol. 157,capsaicin or RTX did not decrease bladder NPY immuno-585–589.
reactivity. Dasgupta, P., Chandiramani, V.A., Beckett, A., Scaravilli, F., Fowler, C.J.,2000. The effect of intravesical capsaicin on the suburothelialinnervation in patients with detrusor hyperreflexia. BJU Int. 85,238–245.5. Conclusions
Fahrkenburg, J., Hannibal, J., 1998. Pituitary cyclase activating poly-peptide immunoreactivity in capsaicin-sensitive nerve fibers supplying
Intravesical capsaicin or RTX decreased the number of the rat urinary tract. Neuroscience 83, 1261–1272.SP or CGRP immunoreactive fibers in the rat urinary Farkas-Szallasi, T., Lundberg, J., Wiesenfeld-Hallin, Z., Hokfelt, T.,bladder similarly without causing significant nerve changes Szallasi, A., 1995. Increased levels of GMAP, VIP and nitric oxid
synthase, and their mRNAs, in lumbar dorsal root ganglia of the ratunder the electron microscope. The number of immuno-following systemic resiniferatoxin treatment. Neuroreport 6, 2230–reactive nerve fibers gradually returned to control levels2234.
within 8 weeks. In addition, both vanilloids increased the Gabella, G., 1995. The structural relations between nerve fibres andvolume threshold to reflex micturition during 12 weeks. muscle cells in the urinary bladder of the rat. J. Neurocytol. 24,We conclude that capsaicin or RTX applied intravesically 159–187.
Harmar, A., Keen, P., 1982. Synthesis and central and peripheral axonalin full desensitizing concentrations exert their effects ontransport of substance P in a dorsal root ganglion–nerve preparationthe bladder without nerve fiber degeneration.in vitro. Brain Res. 231, 379–385.
Ishisuka, O., Mattiasson, A., Andersson, K.-E., 1995. Urodynamic effectsof intravesical resiniferatoxin and capsaicin in conscious rats with and
Acknowledgements without outflow obstruction. J. Urol. 154, 611–616.Ishisuka, O., Igawa, Y., Mattiasson, A., Andersson, K.-E., 1994. Cap-
saicin-induced bladder hyperactivity in normal conscious rats. J. Urol.The authors would like to thank Prof. Antonio Coimbra152, 525–530.
for critical reading of the manuscript and helpful discus- ´ ´ ´Jancso, G., Kiraly, E., Jancso-Gabor, A., 1977. Pharmacologicallysions, and Dr. Vasco Galhardo for programming in Visual induced selective degeneration of chemosensitive primary sensoryBasic the software used for pixel counts. neurons. Nature 270, 741–743.
´Jancso, G., Hokfelt, T., Lundberg, J.M., Kiraly, E., Halasz, N., Nilsson,G., Terenius, L., Rehfeld, J., Steinbuch, H., Verhofstad, A., Elde, R.,Said, S., Brown, M., 1981. Immunohistochemical studies on the effect
References of capsaicin on spinal and medullary peptide and monoamine neuronsusing antisera to substance P, gastrin /CCK, somastatin, VIP, en-
Alm, P., Alumets, J., Hakanson, R., Sundler, F., 1977. Peptidergic kephalin, neurotensin and 5-hydroxytryptamine. J. Neurocytol. 10,(vasoactive intestinal peptide) nerves in the genito-urinary tract. 963–980.Neuroscience 2, 751–754. Kashiba, H., Ueda, Y., Senba, E., 1997. Systemic capsaicin in the adult rat
Avelino, A., Cruz, F., Coimbra, A., 1998. Intravesical capsaicin or differentially affects gene expression for neuropeptides and neuro-resiniferatoxin temporarily decrease sensory peptides in the mucosa trophin receptors in primary sensory neurons. Neuroscience 76, 299–and muscular layer of the rat urinary bladder. Soc. Neurosci. Abstr. 312.24, 1618. Keast, R., DeGroat, W.C., 1992. Segmental distribution and peptide

46 A. Avelino, F. Cruz / Autonomic Neuroscience: Basic and Clinical 86 (2000) 37 –46
content of primary afferent neurons innervating the urogenital organs Mattiasson, A., Ekblad, E., Sundler, F., Uvelius, B., 1985. Origin andand colon of male rats. J. Comp. Neurol. 319, 615–623. distribution of neuropeptide Y-, vasoactive intestinal polypeptide and
Lecci, A., Giuliani, S., Garret, C., Maggi, C.A., 1993a. Evidence for a substance P-containing nerve fibers in the urinary bladder of the rat.role of tachykinins as sensory transmitters in the activation of Cell Tissue Res. 239, 141–146.micturition reflex. Neuroscience 54, 827–837. McMahon, S.B., Lewin, G., Bloom, S.R., 1991. The consequences of
Lecci, A., Giuliani, S., Patacchini, R., Maggi, C.A., 1993b. Evidence long-term topical capsaicin application in the rat. Pain 44, 301–310.against a peripheral role of tachykinins in the initiation of micturition Miller, M., Buck, S., Sipes, G., Yamamura, H., Burks, T., 1982.reflex in rats. J. Pharmacol. Exp. Ther. 264, 1327–1332. Regulation of substance P by nerve growth factor: disruption by
Lecci, A., Giuliani, S., Tramontana, M., Criscuoli, M., Maggi, C.A., capsaicin. Brain Res. 250, 193–196.1997. MEN 11,420, a peptide tachykinin NK-2 receptor antagonist, Nagy, J.I., Vincent, S.R., Staines, W.A., Fibiger, H.C., Reisine, T.D.,reduces motor responses induced by the intravesical administration of Yamamura, H.I., 1980. Neurotoxic action of capsaicin on spinalcapsaicin in vivo. Naunyn-Schmiedeberg’s Arch. Pharmacol. 356, substance P neurons. Brain Res. 186, 435–444.182–188. Nolano, M., Simone, D.A., Wendelschafer-Crabb, G., Johnson, T., Hazen,
Lluel, P., Palea, S., Barras, M., Grandadam, F., Heudes, D., Bruneval, P., E., Kennedy, W.R., 1999. Topical capsaicin in humans: parallel loss ofCorman, B., Martin, D.J., 2000. Functional and morphological epidermal nerve fibers and pain sensation. Pain 81, 135–145.modifications of the urinary bladder in aging female rats. Am. J. Pandita, R.K., Persson, K., Andersson, K.E., 1997. Capsaicin-inducedPhysiol. Regul. Integr. Comp. Physiol. 278, 964–972. bladder overactivity in conscious rats: involvement of spinal nitric
Lu, W.X., Haber, S.N., 1992. In situ hybridization histochemistry — a oxide. J. Auton. Nerv. Syst. 11, 184–191.new method for processing material stored for several years. Brain Sengupta, J.N., Gebhart, G.F., 1995. Mechanosensitive afferent fibers inRes. 578, 155–160. the gastrointestinal and lower urinary tracts. In: Gebhart, G.F. (Ed.),
Lundberg, J.M., Hokfelt, T., 1986. Multiple co-existence of peptides and Visceral Pain. Progress in Pain Research and Management, Vol. 5.classical transmitters in peripheral autonomic and sensory neurons — IASP Press, Seattle, pp. 75–98.functional and pharmacological implications. Prog. Brain Res. 68, Sharkey, K.A., Williams, R.G., Schultzberg, M., Dockray, G.J., 1983.241–262. Sensory substance P innervation of the urinary bladder: possible site
Maggi, C.A., 1991. Capsaicin and primary afferent neurons: from basic of action of capsaicin in causing urine retention in rats. Neurosciencescience to human therapy? J. Auton. Nerv. Syst. 33, 1–14. 3, 861–868.
Maggi, C.A., Patacchini, R., Tramontana, M., Amann, R., Giuliani, S., Silva, C., Rio, M.E., Cruz, F., 2000. Desensitization of bladder sensorySanticioli, P., 1990. Similarities and differences in the action of fibres by intravesical resiniferatoxin, a capsaicin analogue: long-termresiniferatoxin and capsaicin on central and peripheral endings of results for the treatment of detrusor hyperreflexia. Eur. Urol., in press.primary sensory neurons. Neuroscience 37, 531–539. Su, H.C., Wharton, J., Polak, J.M., Muldery, P.K., Ghatei, M.A., Gibson,
Maggi, C.A., Santicioli, P., Geppetti, P., Patacchini, R., Frilli, S., Astolfi, S.J., Terenghi, G., Morrison, J.F.B., Ballesta, J., Bloom, S.R., 1986.M., Fusco, B., Meli, A., 1988. Simultaneous release of substance P Calcitonin gene-related peptide immunoreactivity in afferent neuronsand calcitonin gene-related peptide (CGRP)-like immunoreactivity supplying the urinary tract: combined retrograde tracing and immuno-from isolated muscle of the guinea pig urinary bladder. Neurosci. Lett. histochemistry. Neuroscience 18, 727–747.
´ ´87, 163–167. Szolcsanyi, J., Jancso-Gabor, A., Joo, F., 1975. Functional and fineMaggi, C.A., Santicioli, P., Geppeti, P., Furio, M., Frilli, S., Conte, B., structural characteristics of the sensory neuron blocking effect of
Fanciullaci, M., Giuliani, S., Meli, A., 1987. The contribution of capsaicin. Naunyn-Schmiedeberg’s Arch. Pharmacol. 287, 157–161.´capsaicin sensitive innervation to activation of the spinal vesico- Taylor, D.C.M., Pierau, F.-K., Szolcsanyi, J., 1984. Long lasting inhibi-
vesical reflex in rats: relationship between substance P levels in the tion of horseradish peroxidase (HRP) transport in sensory nervesurinary bladder and the sensory-efferent function of capsaicin-sensi- induced by capsaicin treatment of the receptive field. Brain Res. 298,tive sensory neurons. Brain Res. 415, 1–13. 45–49.
Maggi, C.A., Lippe, T., Giuliani, S., Abelli, L., Somma, V., Geppetti, P., Wilkinson, K.D., Lee, K., Deshpande, S., Duerksen-Hughes, P., Boss,´Jancso, G., Santicioli, P., Meli, A., 1989. Topical versus systemic J.M., Pohl, J., 1989. The neuron specific protein PGP 9.5 is a
capsaicin desensitization: specific and unspecific effects as indicated ubiquitin carboxyl terminal hydrolase. Science 246, 670–673.by modification of reflex micturition in rats. Neuroscience 31, 745– Zimermann, M., 1983. Ethical guidelines for investigation of experimen-756. tal pain in conscious animals. Pain 16, 109–110.
Copyright © 2022 FDOKUMEN



![Role of desensitization of AMPA receptors on the neuronal viability and on the [Ca2+]i changes in cultured rat hippocampal neurons](https://static.fdokumen.com/doc/165x107/63230bd664690856e1096f90/role-of-desensitization-of-ampa-receptors-on-the-neuronal-viability-and-on-the-ca2i.jpg)
















