
Capsaicin 8% Patch for Alternative Therapy of Painful Diabetic ...
Upload
independentCategory
view
5download
0
THE JOURNAL OF COMPARATIVE NEUROLOGY 308~79-90 (1991)
Prior Optic Nerve Transection Reduces Capsaicin-Induced Degeneration in Rat
Subcortical Visual Structures
SUE RITTER AND THU T. DINH Department of Veterinary and Comparative Anatomy, Pharmacology, and Physiology,
Washington State University, Pullman, Washington 99164-6520
ABSTRACT Capsaicin is a neurotoxin capable of causing degeneration in specific sites throughout the
neuraxis, including the suprachiasmatic nucleus (SCh), the ventrolateral geniculate nucleus (VLG), the intergeniculate leaflet (IGL), and the olivary and medial pretectal nuclei (OPT and MPT). In this experiment, we tested the hypothesis that capsaicin-induced terminal degenera- tion in the SCh, VLG, IGL, OPT, and MPT results from destruction of retinal ganglion cells and their axonal projections to these sites. In the first experiment, silver stains were used to examine degeneration in the retina induced by systemic capsaicin treatment. Capsaicin caused degeneration of ganglion cells, bipolar cells, and nerve terminals in the retina, which could be observed between 2 and 24 hours after treatment. In the second experiment, 15-day-old rat pups were enucleated unilaterally. Five days or 2, 5, or 10 months later, they were injected systemically with capsaicin and killed 6 hours (pups) or 18 hours (adults) later for analysis with a cupric silver stain. In rats of all ages, prior monocular enucleation reduced or eliminated capsaicin-induced degeneration in the contralateral SCh, VLG, IGL, OPT, and MPT. In the third experiment, rat pups were treated systemically with capsaicin or vehicle solution at 12 days of age and given unilateral intravitreal injections of cholera toxin conjugated to horseradish peroxidase (CT-HRP) 3 days prior to sacrifice at 20 days of age. Transport of CT-HRP to the SCh, VLG, IGL, MPT, and OPT was attenuated but not abolished by capsaicin pretreatment. Results suggest that capsaicin causes degeneration in the SCh, VLG, IGL, MPT, and OPT by selective destruction of a subpopulation of retinal ganglion cells with axonal projections to these sites.
Key words: visual pathways, suprachiasmatic nucleus, geniculate bodies, retina
Capsaicin's neurotoxicity for small-diameter unmyeli- nated primary sensory neurons of peripheral origin has been well described (for reviews, Monsureenusorn et al., '82; Russell and Burchiel, '84; Buck and Burks, '86; Maggi and Meli, '88). Until recently, it was believed that capsai- cin's neurotoxicity was limited to this class of neurons (Jancso and Kiraly, '77, '80, '81; Jancso, '78, '81). However, our recent silver stain studies demonstrated that degenera- tion induced by systemic capsaicin treatment of adult or preweanling rats is not limited to areas of the brain known to be innervated by primary sensory neurons but can be observed in discrete areas throughout the neuraxis (Ritter and Dinh, '88, '90). Capsaicin-sensitive sites include areas innervated by small-diameter primary sensory neurons, several areas containing projections of higher order sensory neurons and some areas with no known sensory function. Of particular interest for the present experiment is the fact that capsaicin, when administered systemically to rats prior to 30 days of age, causes terminal degeneration in several
sites innervated by retinal ganglion cells: the suprachias- matic nucleus (SCh), the magnocellular subnucleus of the ventrolateral geniculate nucleus (VLGMC), the intergenic- ulate leaflet (IGL), and the olivary and medial pretectal nuclei (OPT and MPT, respectively). Capsaicin also causes substantial degeneration in these nuclei in adult rats. However, in the VLG, IGL, and OPT, capsaicin-induced degeneration in adult rats is significantly less than in rat pups (Ritter and Dinh, '88, '90; Dinh and Ritter, '89).
The SCh, VLG, IGL, and OPT are known projection sites for retinal ganglion cells (for a general review, see Sefton and Dreher, '85). However, nonretinal projections share these terminal fields. Therefore, capsaicin-sensitive termi- nals in these nuclei could be of either retinal or nonretinal
Accepted February 11,1991. Address reprint requests to Sue Ritter, Dept. of Veterinary and Compara-
tive Anatomy, Pharmacology, and Physiology, Washington State University, Pullman, WA 99164-6520.
O 1991 WILEY-LISS, INC.

80 S. RITTER AND T.T. DINH
Fig. 1. Photomicrographs showing the time course of degeneration in retinas of 20-day-old rat pups treated 2-48 hours earlier with capsaicin (75 mgikg, s.c.). Abbreviations at the top right side of the figure identify retinal layers. Retinas were processed with a cupric silver stain, which produces intense labeling of degenerating cells and terminals against a clear background of unlabeled normal tissue (see also Fig. 2). Degenerating perikarya were observed in the inner nuclear (inl) and ganglion cell layers (gcl). Degenerating nerve terminals were most prevalent in the inner plexiform layer (ipl), but were also present
in the outer plexiform (opl) and gcl. A s depicted, degeneration-induced argyrophilia was maximal for both perikarya and terminals between 2 and 12 hours after capsaicin treatment. Note that perikarya in the outer nuclear layer (on11 were argyrophilic in both control and capsaicin- treated rats using the cupric silver stain. In retinas stained with the Gallyas silver technique (not shown), the pattern of degeneration was similar In the other layers, but no staining was observed in the onl. Bar = 200 pm.
2nR 2nL 3v AF'TD D3V DLG fi fr gcl Hi IGL in1 ipl lo LHh LV MPT onf
optic nerve, right optic nerve, left third ventricle anterior pretectal nucleus, dorsal part dorsal third ventricle dorsolateral geniculate nucleus fimbria of the hippocampus fasciculus retroflexus ganglion cell layer, retina hippocampus intergeniculate leaflet inner nuclear layer, retina inner plexiform layer, retina lateral olfactory tract lateral habenular nucleus lateral ventricle medial pretectal nucleus optic nerve fiber layer, retina
Abbreviations
on1 OPl OPT opt OT
Pel ox
POS PPT SCh SFO sm St
VLG VLGMC VLGPC VMH
outer nuclear layer, retina outer plexiform layer, retina olivary pretectal nucleus optic tract nucleus of the optic tract optic chiasm pigment epithelial layer photoreceptor outer segment posterior pretectal nucleus suprachiasmatic nucleus suhfornical organ stria medullaris stna terminalis ventrolateral geniculate nucleus ventrolateral geniculate nucleus, magnocellular part ventrolateral genlculate nucleus, parvocellular part ventromedial hypothalamic nucleus
CAPSAICIN-SENSITIVE RETINAL PROJECTIONS 81
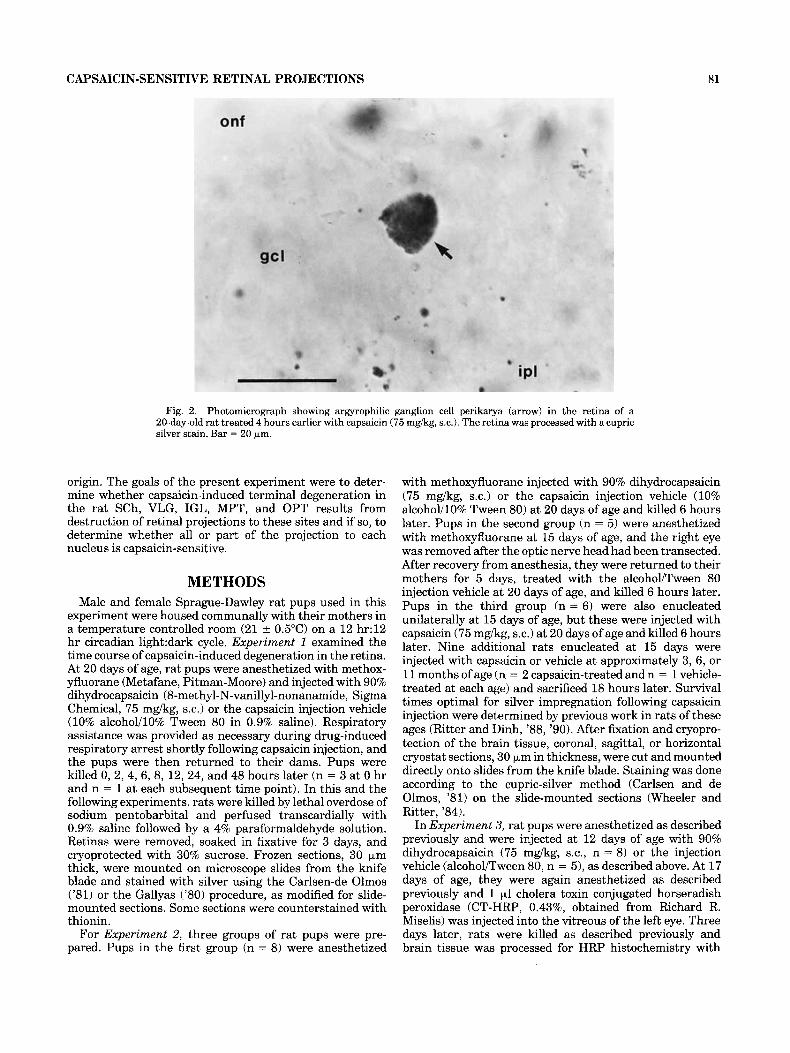
Fig. 2. Photomicrograph showing argyrophilic ganglion cell perikarya (arrow) in the retina of a 20-day-old rat treated 4 hours earlier with capsaicin (75 mgkg, s.c.). The retina was processed with a cupric silver stain. Bar = 20 bm.
origin. The goals of the present experiment were to deter- mine whether capsaicin-induced terminal degeneration in the rat SCh, VLG, IGL, MPT, and OPT results from destruction of retinal projections to these sites and if so, to determine whether all or part of the projection to each nucleus is capsaicin-sensitive.
METHODS Male and female Sprague-Dawley rat pups used in this
experiment were housed communally with their mothers in a temperature controlled room (21 f 0.5"C) on a 12 hr:12 hr circadian 1ight:dark cycle. Experiment 1 examined the time course of capsaicin-induced degeneration in the retina. At 20 days of age, rat pups were anesthetized with methox- yfluorane (Metafane, Pitman-Moore) and injected with 90% dihydrocapsaicin (8-methyl-N-vanillyl-nonanamide, Sigma Chemical, 75 mgkg, s.c.) or the capsaicin injection vehicle (10% alcohol/lO% Tween 80 in 0.9% saline). Respiratory assistance was provided as necessary during drug-induced respiratory arrest shortly following capsaicin injection, and the pups were then returned to their dams. Pups were killed 0, 2, 4, 6, 8, 12, 24, and 48 hours later (n = 3 at 0 hr and n = 1 at each subsequent time point). In this and the following experiments, rats were killed by lethal overdose of sodium pentobarbital and perfused transcardially with 0.9% saline followed by a 4% paraformaldehyde solution. Retinas were removed, soaked in fixative for 3 days, and cryoprotected with 30% sucrose. Frozen sections, 30 pm thick, were mounted on microscope slides from the knife blade and stained with silver using the Carlsen-de Olmos ('81) or the Gallyas ('80) procedure, as modified for slide- mounted sections. Some sections were counterstained with thionin.
For Experiment 2, three groups of rat pups were pre- pared. Pups in the first group (n = 8) were anesthetized
with methoxyfluorane injected with 90% dihydrocapsaicin (75 mg/kg, s.c.) or the capsaicin injection vehicle (10% alcohol/lO% Tween 80) at 20 days of age and killed 6 hours later. Pups in the second group (n = 5 ) were anesthetized with methoxyfluorane at 15 days of age, and the right eye was removed after the optic nerve head had been transected. After recovery from anesthesia, they were returned to their mothers for 5 days, treated with the alcohol/Tween 80 injection vehicle at 20 days of age, and killed 6 hours later. Pups in the third group (n = 6) were also enucleated unilaterally at 15 days of age, but these were injected with capsaicin (75 mgkg, s.c.) at 20 days of age and killed 6 hours later. Nine additional rats enucleated at 15 days were injected with capsaicin or vehicle at approximately 3, 6 , or 11 months of age (n = 2 capsaicin-treated and n = 1 vehicle- treated at each age) and sacrificed 18 hours later. Survival times optimal for silver impregnation following capsaicin injection were determined by previous work in rats of these ages (Ritter and Dinh, '88, '90). After fixation and cryopro- tection of the brain tissue, coronal, sagittal, or horizontal cryostat sections, 30 p,m in thickness, were cut and mounted directly onto slides from the knife blade. Staining was done according to the cupric-silver method (Carlsen and de Olmos, '81) on the slide-mounted sections (Wheeler and Ritter, '84).
In Experiment 3, rat pups were anesthetized as described previously and were injected at 12 days of age with 90% dihydrocapsaicin (75 mg/kg, s.c., n = 8 ) or the injection vehicle (alcohol/Tween 80, n = 5), as described above. At 17 days of age, they were again anesthetized as described previously and 1 pl cholera toxin conjugated horseradish peroxidase (CT-HRP, 0.43%, obtained from Richard R. Miselis) was injected into the vitreous of the left eye. Three days later, rats were killed as described previously and brain tissue was processed for HRP histochemistry with

82 S. RITTER AND T.T. DINH
Fig. 3. Photomicrographs showing retinas of rat pups treated 4 hrs earlier with capsaicin (75 mg/kg, s.c.). Retinas were stained with cupric silver and counterstained with thionin. Degenerating ganglion cell perikarya can be seen in the ganglion cell layer (gcl) in A and at higher magnification in B and C (arrows). Degenerating bipolar cell perikarya
can be seen in the inner nuclear layer (inl) in A and at higher magnification in D (example indicated by arrow). Degenerating nerve terminals can be seen in the inner pkexiform layer (ipl) in A-D (example indicated by arrow in B). Bar for A = 25 pm; B-D = 20 pm.
tetramethyl benzidine as the chromogen (Mesulam, '78) and examined for anterograde labeling by bright and dark field microscopy.
In Experiments 2 and 3, anatomical structures contain- ing capsaicin-induced argyrophilia or CT-HRP were identi-
fied by their correspondance in all three planes of section with labeled structures in the Paxinos and Watson atlas of the rat brain (Paxinos and Watson, '86). Interpretation of the pretectal neuroanatomy was facilitated by reference to Scalia ('72) and Scalia and Arango ('79).

CAPSAICIN-SENSITIVE RETINAL PROJECTIONS 83
A 0.78 mm
@$Lc VLGPC
b o3
72 mm Fig. 4. Semi-diagrammatic map (horizontal plane of section, rostra1
end downward) showing subcortical visual structures containing degen- erating nerve terminals (shaded areas) 6 hours after capsaicin injection (75 mgikg, s.c.). The suprachiasmatic nucleus (SCh) is shown in A; the
magnocellular part of the ventrolateral geniculate nucleus (VLGMC) and the intergeniculate leaflet (IGL) are shown in B; and the olivary and medial pretectal nuclei (OPT and MPT) are shown in C. Numbers at right indicate approximate distance (mm) dorsal to interaural line.

84 S. RITTER AND T.T. DINH
Fig. 5. Dark field photomicrographs of silver stained brain sections showing the suprachiasmatic nucleus (SCh) from three 20-day-old rat pups. Brain sections are in the horizontal plane in the photomicro- graph. In A, capsaicin (75 mgikg, s.c.1 was injected 6 hours prior to death, causing degeneration of nerve terminals bilaterally in the SCh. In B and C, rats were enucleated unilaterally (right side) at 15 days of age. At 20 days of age, they were injected with the control solution (B) or capsaicin (75 mgikg, s.c.) 6 hours prior to death (C). Degenerating optic nerve fibers, but not terminals, were still argyrophilic in rats enucleated unilaterally 5 days prior to death. Monocular enucleation greatly reduced capsaicin-induced terminal degeneration in the con- tralateral SCh, but not in the ipsilateral SCh. Since the retinofugal innervation of the SCh is largely crossed, the loss of capsaicin-sensitive terminals contralateral to enucleation indicates that capsaicin-induced degeneration in the SCh is due to destruction of retinal ganglion cells. Symbol (*I indicates the side contralateral to enucleation. Bar = 500 Pm.
RESULTS Experiment 1 As shown in Figure 1, systemic capsai-
cin caused pronounced degeneration of terminals and cell bodies in the retina. The time course of the degeneration in the retina was similar to that observed elsewhere in the brain for preweanling rats (Ritter and Dinh, '89, 'go), with argyrophilia reaching a maximum in cell bodies and termi- nals between 4 and 8 hours after capsaicin injection and declining to negligible levels within 24 hours of the treat- ment. Degenerating terminals were abundant in the inner plexiform layer, but were also distributed sparcely through- out the outer plexiform layer. Degenerating cell bodies were numerous in the inner nuclear layer and appeared to be a subpopulation of bipolar cells. Degenerating cell bodies were also present in the ganglion cell layer, as shown in Figures 2 and 3. Degenerating ganglion cells were sparcely distributed throughout the retina, but our impression was that they occurred with a slightly greater frequency in the periphery of the retina. Because silver impregnation meth- ods stain actively degenerating neurons and do not ade- quately delineate dendritic arborizations of such cells, morphological characterization of the capsaicin-sensitive ganglion cells was not possible in this experiment.
Capsaicin caused degeneration of nerve terminals in the SCh (ventroposteriolateral part), VLG (magnocellular part), IGL, OPT, and MPT in the 20-day-old rat pup. Figure 4 summarizes these results semi-diagramat- ically on drawings of horizontal brain sections. Photomicro- graphs of horizontal brain sections from representative capsaicin-treated rats, are shown in Figures 5A (SCh), 6A (OPT and MPT), and 7A (VLG and IGL). Figures 5B, 6B, 7B show the effects of unilateral enucleation in the same structures. Five days after monocular enucleation, fiber degeneration could still be observed in the optic nerve and tract, primarily on the contralateral side. However, nerve terminals degenerating as a result of the enucleation were no longer present. Figures 5C, 6C, 7C show the effect of prior monocular enucleation on capsaicin-induced terminal degeneration ipsilateral and contralateral to enucleation. In the most ventral portions of the SCh, capsaicin-induced degeneration was present across the entire mediolateral extent of the nucleus. In this most ventral aspect of the SCh, monocular enucleation reduced, but did not abolish, capsaicin-induced degeneration on the contralateral side. However, at a slightly more dorsal level in the SCh, as in Figure 5C, capsaicin-induced degeneration was confined to the ventroposterlateral quadrant of the nucleus. At this level, capsaicin-induced degeneration was totally abolished by contralateral enucleation. These findings are consistent with previous reports (e.g., Hendrickson et al., '72; Mason and Lincoln, '76) and with results of Experiment 3 regard- ing the distribution of the retinofugal innervation within the SCh and the significant bilaterality of this projection. In the VLG and IGL (Fig. 6C) and in the OPT and MPT (Fig. 7 0 , capsaicin-induced degeneration appeared to be totally eliminated on the side contralateral to enucleation. This is consistent with the fact that the retinal projections to these nuclei are predominantly crossed (Hayhow et al., '62). Capsaicin-induced degeneration in other brain regions re- ported previously (Ritter and Dinh, '88, '901, including diencephalic and ventral forebrain regions, did not appear to be attenuated by monocular enucleation.
Consistent with our previous reports (Ritter and Dinh, '88, '90; Dinh and Ritter, '891, capsaicin treatment of adult rats (3, 6, and 11 months of age) caused degeneration of
Experiment 2

CAPSAICIN-SENSITIVE RETINAL PROJECTIONS 85
Fig. 6. Dark field photomicrographs of silver stained brain sections showing the OPT and MPT bilaterally from three 20-day-old rat pups. Brain sections are in the horizontal plane in the photomicrograph. In A, capsaicin (75 mg/kg, s.c.) was injected 6 hours prior to death, causing degeneration of nerve terminals bilaterally in the OPT and MPT. Rats shown in B and C were enucleated unilaterally at 15 days of age and injected 5 days later with the control solution (B) or capsaicin (75
mgkg, s.c.) (C) 6 hours prior to death. Degenerating optic tract fibers, but not terminals, were still argyrophilic in rats enucleated unilaterally 5 days prior to death. Enucleation eliminated capsaicin-induced termi- nal degeneration in the contralateral OPT and MPT, but not in the ipsilateral nuclei. Symbol (*) indicates the side contralateral to enucle- ation. Bar = 500 km.
nerve terminals in the SCh, VLG, IGL, OPT, and MPT. In the VLG, IGL, and OPT, however, capsaicin-induced degen- eration in adult rats was much less than in the pup. Prior monocular enueleation had effects in adult rats that were
similar to those observed in rat pups. Enucleation abolished capsaicin-induced argyrophilia in the contralateral VLG, IGL, OPT, and MPT and reduced it significantly in the contralateral SCh.

86 S. RITTER AND T.T. DINH
Figure 7

CAPSAICIN-SENSITIVE RETINAL PROJECTIONS 87
Experiment 3 In both capsaicin- and vehicle-treated rats, CT-HRP was transported bilaterally from the injected retina to the SCh, VLG, IGL, OPT, and MPT, as well as to other subcortical visual areas (Figs. 8,9). However, labeling was more extensive contralateral to the injected eye, as would be expected from previous work (Hayhow et al., '62; Scalia, '72; Scalia and Arango, '79; Sefton and Dreher, '85). As described previously for the cat (Koontz et al., '851, retinorecipient zones in the pretectum formed rostomedial to caudolateral strips spanning the pretectum in the horizon- tal plane. The strips, which included fibers of passage and terminals, were roughly parallel to each other and to the rostral border of the tectum.
Capsaicin pretreatment did not eliminate transport of CT-HRP to the SCh, OPT, and MPT (Fig. 8) or VLG and IGL (Fig. 9), although a slight reduction in the amount of label was perceptible both ipsi- and contralateral to the injected eye. These results indicate that capsaicin destroys only a subpopulation of the ganglion cells which send axonal projections to these nuclei.
DISCUSSION These experiments confirm our previous reports that
capsaicin causes degeneration in several subcortical nuclei associated with visual function: the SCh, VLG, IGL, OPT, and MPT. The major new finding is that capsaicin-induced degeneration in these subcortical nuclei is due to destruc- tion of retinal ganglion cells and their processes. This conclusion is based on two important results: (1) reduction of retinal input to these nuclei by contralateral ocular enucleation reduced or eliminated degeneration induced by subsequent capsaicin treatment in rats of all ages, and (2) degenerating ganglion cell bodies were observed within the retina after capsaicin treatment.
Direct innervation by retinal ganglion cells has been previously reported in the rat for the SCh (Hendrickson et al., '72; Moore and Lenn, '72), VLG and IGL (Hayhow et al., '62), and OPT (Scalia, '72; Scalia and Arango, '79). More- over, this previous work has shown that these nuclei are innervated predominantly by the contralateral retina. Therefore, our demonstration of capsaicin-induced degener- ation contralateral to ocular enucleation is in accord with the known distribution of retinofugal projections in the rat.
One aspect of our results that may differ from previous anatomical reports should be noted. We found that capsaicin- induced degeneration within the MPT was abolished by contralateral enucleation. In addition, this same site was labeled by vitreal injections of CT-HRP. To our knowledge,
Fig. 7. Dark field photomicrographs of silver stained brain sections (horizontal plane) showing the VLG and IGL from three 20-day-old rat pups. Right (A, B, C) and left (A', B', C') sides are shown. In A and A , capsaicin (75 mgikg, s.c.) was injected 6 hours prior to death, causing degeneration of nerve terminals bilaterally in the VLGMC and IGL. In B, B' and C, C' rats were enucleated unilaterally (right side) at 15 days of age. At 20 days of age, they were injected with the control solution (B, B') or capsaicin (75 mgikg, s.c.) 6 hours prior to death (C, C'). Degenerating optic tract fibers, but not terminals, were still argyro- philic in rats enucleated unilaterally 5 days prior to death, mainly contralateral to the lesion (B' and C'). Enucleation eliminated capsaicin- induced terminal degeneration in the contralateral VLGMC and IGL (C'), but not in the ipsilateral nuclei (C). Symbol (+) indicates the side contralateral to enucleation. Bar = 500 km.
direct retinofugal innervation of the MPT has not been previously reported. The present result may be attributable to the fact that CT-HRP labels neuronal processes more effectively than methods previously used to study the retinal projections to this brain region. We hasten to add, however, that the boundaries of the pretectal nuclei in the rat are indistinct and even the most definitive anatomical descriptions of this brain region (Scalia, '72; Scalia and Arango, '79) are incomplete. Therefore, the capsaicin- sensitive area we describe as being within the MPT bound- aries defined by Paxinos and Watson ('86) conceivably could be a retinorecipient portion of an adjacent nucleus.
Although degenerating bipolar cells in the retina were fairly numerous after systemic capsaicin treatment, the number of degenerating ganglion cell bodies appeared to be quite small. However, previous reports indicate that the retinal innervation of the nuclei under investigation in this study consists of a small percentage of the total retinofugal projection and involves only a small number of ganglion cells. In the golden hamster, only 6-30 ganglion cells were labeled by injections of HRP into the SCh (Pickard, '80). In the rat, the number of retinofugal fibers entering the SCh has been reported to be only 0.1%, the number entering the VLGMC only 8%, and the number entering the pretectum only 13% of the total optic nerve fiber population (Mason and Lincoln, '76; Togaand Collins, '81), The fact that only a small number of ganglion,cells appeared to be destroyed by capsaicin is thus consistent with predictions from published work and also with 6 e small reduction by capsaicin of CT/HRP transport to affected nuclei in the present experi- ment.
The retinofugal innervation of most subcortical areas originates from several distinguishable classes of ganglion cells (Boycott and Wassel, '74; Lennie, '80; Fukuda et al., '84; for review see Rodieck, '79). The restriction ofcapsaicin- induced degeneration to specific subcortical visual nuclei, to subpopulations of terminals within those nuclei, and to a small ganglion cell population suggests that capsaicin's toxicity is selective for a single ganglion cell type.
This experiment did not attempt to characterize the morphological or functional features of capsaicin-sensitive ganglion cells. However, in a number of species, including rats, hamsters, and cats, the SCh, VLGMC, IGL, and pretectum are innervated by small-diameter, slowly conduct- ing retinofugal fibers (Millhouse, '77; Hale and Sefton, '78; Nagata and Hayashi, '84). Therefore, it is possible that capsaicin's toxicity in the visual system, as in the somato- and viscerosensory systems (for review, Buck and Bulks, '86; Maggi and Meli, '881, is limited to neurons with small caliber, slowly conducting axons. However, it is noteworthy that capsaicin does not cause degeneration in all areas innervated by ganglion cells with these morphological characteristics. For example, the nucleus of the optic tract and the rostral colliculus, which also receive small-diameter retinofugal fibers, do not appear to be innervated by capsaicin-sensitive ganglion cells. This suggests that a distinct class of ganglion cells exists in the rat that is distinguishable by its capsaicin-sensitivity and the unique distribution of its projections within the diencephalon and pretectum.
When administered to adult rats 3, 6, or 11 months of age, systemic capsaicin treatment caused significant degen- eration in the SCh and MPT but, as reported previously,

88 S. RITTER AND T.T. DINH
Fig. 8. Bright field photomicrographs of the SCh, MPT, and OPT from control (A, C, E) and capsaicin-pretreated (B, D, F) rats given intravitreal injections (left side) of CT-HRP 3 days prior to death. Brain sections are in the horizontal plane in the figure. For the SCh and MPT, nuclei are shown bilaterally. For the OPT, the nucleus contralateral to
the injected eye is shown. Nuclei were labeled bilaterally, but the most intense labeling was contralateral to the injected eye. The amount of CT-HRP transported to these nuclei from the retina appeared to be reduced only slightly by capsaicin pretreatment. Symbol (*I indicates the side contralateral to CT-HRP injection. Bar = 500 pm.
only very small amounts of degeneration in the VLG, IGL, and OPT (Ritter and Dinh, '88, '90; Dinh and Ritter, '89). The reduction of capsaicin-induced degeneration in some, but not all, retinal projections during adulthood (Ritter and Dinh, '88, '90; Dinh and Ritter, '89) is an interesting finding. The altered response to capsaicin may reflect an age-related decline in capsaicin-sensitivity of retinofugal projections to the VLG, IGL, and OPT, a spontaneous attrition of the capsaicin-sensitive ganglion cell population
projecting to these sites, or a spontaneous regression of ganglion cell collateral innervation to these sites.
In summary, results of this study indicate that a subpop- ulation of retinal ganglion cells and their projections to the SCh, VLG, IGL, OPT, and MPT are destroyed by systemic treatment with capsaicin. The highly selective nature of capsaicin's neurotoxicity within the visual system suggests that capsaicin may be a valuable tool for studying the function and circuitry of specific retinal projections.

CAPSAICIN-SENSITIVE RETINAL PROJECTIONS 89
Fig. 9. Bright field photomicrographs of the right and left VLG from control (A and A', respectively) and capsaicin-pretreated (B and B', respectively) rats given intravitreal injections (left eye) of CT-HRP 3 days prior to death. Brain sections are in the horizontal plane in the figure. Nuclei were labeled bilaterally, but the most intense labeling
was contralateral to the injected eye. The amount of CT-HRP trans- ported to these nuclei from the retina appeared to be reduced only slightly by capsaicin pretreatment. Symbol (*) indicates the side contralateral to CT-HRP injection. Bar = 500 bm.

90 S. RITTER AND T.T. DINH
ACKNOWLEDGMENTS We thank Dr. Richard Miselis for his gift of CT-HRP.
LITERATURE CITED Boycott, B.B., and H. Wassle (1974) The morphological types of ganglion
cells of the domestic cat’s retina. J. Physiol. London 240:397419. Buck, S.H., and B.T. Burks (1986) The neuropharmacology of capsaicin:
Review of some recent observations. Pharmacol. Rev. 38:179-226. Carlsen, J., and J.S. de Olmos (1981) Silver impregnation of degenerating
neurons and their processes: A modified cupric silver technique. Brain Res. 208r426-431.
Dinh, T., and S. Ritter (1989) Age-related changes in capsaicin-induced degeneration in rat brain. Soc. Neurosci. Abstr. 15:1022.
Fukuda, Y., and J. Stone (1974) Retinal distribution and central projections ofY-, X- and W-cells of the cat’s retina. J. Neurophysiol. 37:749-772.
Gallyas, F., J.R. Wolff, H. Bottcher, and L. Zaborszky (1980b) A reliable method to localize terminal degeneration and lysosomes in the central nervous system. Stain Technol. 55:299-306.
Hale, P.T., and A.J. Sefton (1978) A comparison o f the visual and electrical response properties of cells in the dorsal and ventral lateral geniculate nuclei. Brain Res. 153:591-595.
Hayhow, W.R., A. Sefton, and C. Webb (1962) Primary optic centers of the rat in relation to the terminal distribution of the crossed and uncrossed optic nerve fibers. J. Comp. Neurol. 118:295-322.
Hendrickson, A.E., N. Wagoner, and W.M. Cowan (1972) An autoradio- graphic and electron microscopic study of retino-hypothalamic connec- tions. Z. Zellforsch. 1 3 5 - 2 6 .
Jancso, G. (1978) Selective degeneration of chemosensitive primary sensory neurones induced by capsaicin. Cell Tiss. Res. 195t145-152.
Jancso, G. (1981) Intracisternal capsaicin: Selective degeneration of chemosensitive primary sensory afFerents in the adult rat. Neurosci. Lett. 27:41-45.
Jancso, G., and E. Kiraly (1980) Distribution of chemosensitive primary sensory afferents in the central nervous system of the rat. J. Comp. Neurol. 190: 78 1-792.
Jancso, G., and E. Kiraly (1981) Sensory neurotoxins: Chemically induced selective destruction of primary sensory neurons. Brain Res. 210:83-89.
Jancso, G., E. Kiraly, and A. Jancso-Gabor (1977) Pharmacologically in- duced selective degeneration of chemosensitive primary sensory neu- rons. Nature (Lond.) 270r741-743.
Koontz, M.A., R.W. Rodieck and S.G. Farmer (1985) The retinal projection to the cat pretectum. J. Comp. Neurol. 236:42-59.
Lennie, P. (1980) Parallel visual pathways: A review. Vision Res. 20:561- 594.
Maggi, C.A., and A. Meli (1988) The sensowefferent function of capsaicin-
Mason. C.A., and D.W. Lincoln (1976) Visualization of the retinohvpotha- sensitive sensory neurons. Gen. Pharmac. 19:143.
lamic projection in the rat by cobalt precipitation. Cell Tiss;; Res. 168:117-132.
Mesulam, M.-M. (1978) Tetramethyl benzidine for horseradish peroxidase neurohistochemistry: A noncarcinogenic blue reaction-product with superior sensitivity for visualizing neural afferents and efferents. J. Histochem. Cytochem. 26:106-117.
Millhouse, O.E. (1977) Optic chiasm collaterals afferent to the suprachias- matic nucleus. Brain Res. 137:351-355.
Monsereenusorn, Y., K. Sathapana, and P.D. Pezalla (1982) Capsaicin-a literature survey. CRC Critical Reviews in Toxicology 10t321-339.
Moore, R.Y., and N.J. Lenn (1972) A retinohypothalamic projection in the rat. J. Comp. Neurol. 146:l-14.
Nagata, T., and Y. Hayashi (1984) The visual field representation of the rat ventral geniculate nucleus. 3. Comp. Neurol. 227:582-588.
Paxinos, G., and C. Watson (1986) The Rat Brain in Stereotaxic Coordinates. New York: Academic Press.
Pickard, G.E. (1980) Morphological characteristics of retinal ganglion cells projecting to the suprachiasmatic nucleus: A horseradish peroxidase study. Brain Res. I83:458-465.
Ritter, S., and T.T. Dinh (1988) Capsaicin-induced neuronal degeneration: Silver impregnation of cell bodies, axons and terminals in the central nervous system of the adult rat. J. Comp. Neurol. 271:79-90.
Ritter, S., and T.T. Dinh (1989) Prior unilateral optic nerve transection abolishes capsaicin-induced degeneration in diencephalic and pretectal visual structures. SOC. Neurosci. Abstr. 15461.
Ritter, S., and T.T. Dinh (1990) Capsaicin-induced neuronal degeneration in the brain and retina of preweanlingrats. J. Comp. Neurol. 296:447461.
Rodieck, R.W. (1979) Visual pathways. Ann. Rev. Neurosci. 2193-225. Russell, L.C., and K.J. Burchiel (1984) Neurophysiological effects of capsai-
cin. Brain Res. Rev. 8r165-176. Scalia, F. (1972) The termination of retinal axons in the pretectal region of
mammals. J. Comp. Neurol. 145t223-58. Scalia, F., and V. Arango (1979) Topographic organization of the projections
of the retina to the pretectal region in the rat. J. Comp. Neurol. 186:271-292.
Sefton, A.J., and B. Dreher (1985) Visual System. In: G. Paxinos (ed) The Rat Nervous System. Vol. 1. Australia: Academic Press, pp 169-221.
Toga, A.W., and R.C. Collins 11981) Metabolic response of optic centers to visual stimuli in albino rat: Anatomical and physiological consideration. J. Comp. Neurol. 199:442-464.
Wheeler, D.A., and S. Ritter (1984) A modification of the Carlsen-de Olmos cupric-silver impregnation method for use on mounted cryostat sections. Soc. Neurosci. Abstr. 10:424.
Copyright © 2022 FDOKUMEN



















