
Epidermal growth factor (EGF)-induced up-regulation and agonist- and antagonist-induced...
11
Ž . Brain Research 816 1999 47–57 Research report Ž . Epidermal growth factor EGF -induced up-regulation and agonist- and antagonist-induced desensitization and internalization of A adenosine 1 receptors in a pituitary-derived cell line Angels Navarro, Rafael Zapata, Enric I. Canela, Josefa Mallol, Carmen Lluis, Rafael Franco ) Facultat de Quimica, Departament de Bioquımica i Biologia Molecular, UniÕersitat de Barcelona, Martı i Franques 1, 08028 Barcelona, Catalunya, Spain ´ ´ ` Accepted 6 October 1998 Abstract Ž . This report concerns the study of homologous and heterologous regulation of cell surface A adenosine receptors AR in a 1 1 pituitary-derived cell line. This has been possible by the use of the recently developed anti-A R antibodies in immunocytochemical 1 assays. Functional desensitization and internalization of A R in GH cells occurred after treatment with agonist but also with antagonist. 1 4 Ž . Epidermal growth factor EGF treatment led to the up-regulation of cell surface A R in GH cells. Confocal analysis evidenced an 1 4 EGF-induced increase of A R present in intracellular clathrin-coated vesicles. The up-regulation was blocked by actinomycin D thus 1 suggesting the involvement of protein synthesis in the effect induced by the growth factor. These results constitute the first example of adenosine receptor regulation by EGF and one of the few examples of antagonist-induced desensitization and internalization among G-protein-coupled receptors. q 1999 Elsevier Science B.V. All rights reserved. Keywords: A adenosine receptor; Receptor regulation; Agonist; Antagonist; Epidermal growth factor; Pituitary cell 1 1. Introduction Regulation of receptor expression is a common mecha- nism to control cellular responses to extracellular signals. Ž Thus, the effect of physiological factors hormones, neuro- . transmitters and growth factors is determined by the relative amount of cell surface receptors and the specific signal transduction pathways involved. Like other mem- bers of the G-protein-coupled receptors family, A R are 1 regulated in response to agonist or antagonist stimulation. Agonist-induced desensitization of AR has been de- 1 scribed in intact animals and in cell cultures. Long-term administration of agonists to animals cause functional de- wx sensitization of A R in guinea pig heart 7 , rat adipocytes 1 w x w x w x 22 , rat atrial muscle 23 and rat brain 10,33 . The reduced functional response is due to a net loss of A R 1 Ž . down-regulation and to a decrease in the proportion of A R displaying high-affinity sites for agonists. Growth of 1 Abbreviations: A R: A adenosine receptor; DPCPX: 1,3-dipropyl-8- 1 1 Ž .Ž . cyclopentylxanthine; R-PIA: N6- R - phenylisopropyl adenosine ) Corresponding author. Fax: q 34-934-021219; E-mail: [email protected]; Internet home page: www.bq.ub.esrrecepr franco.html rat adipocytes or smooth muscle DDT MF-2 cells in the 1 presence of R-PIA leads to the desensitization and down- w x regulation of A R 14,15,31 . We have recently demon- 1 strated that treatment of DDT MF-2 cells with the agonist 1 R-PIA induces a rapid clustering and desensitization of the receptor whereas loss of binding sites on the cell mem- brane due to internalization and down-regulation is a slower wx event occurring within hours 3 . A R are also regulated in 1 an heterologous way in response to other hormones. Thus, chronic treatment of rats with benzodiazepines such as diazepam induces the down-regulation of adenosine recep- w x tors in the CNS 17 . In contrast, up-regulation of A R has 1 been demonstrated in rat brain by glucocorticoid treatment w x 35 . On the other hand, morphine or opioids treatment leads to up-regulation or not detectable changes in the w x level of receptor expressed in different brain areas 21 . wx Recently, Arslan et al. 2 have described a down-regu- lation of A adenosine receptors upon NGF-induced dif- 2A ferentiation of PC12 cells. EGF activates, via a tyrosine-kinase receptor, the ex- pression of multiple genes and alters the growth and differentiation of a wide range of cell types. GH pituitary 3 cells are derived from a rat pituitary tumour and secrete prolactin and growth hormone in response to thyrotropin- 0006-8993r99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved. Ž . PII: S0006-8993 98 01062-2
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Epidermal growth factor (EGF)-induced up-regulation and agonist- and antagonist-induced...

Ž .Brain Research 816 1999 47–57
Research report
Ž .Epidermal growth factor EGF -induced up-regulation and agonist- andantagonist-induced desensitization and internalization of A adenosine1
receptors in a pituitary-derived cell line
Angels Navarro, Rafael Zapata, Enric I. Canela, Josefa Mallol, Carmen Lluis, Rafael Franco )
Facultat de Quimica, Departament de Bioquımica i Biologia Molecular, UniÕersitat de Barcelona, Martı i Franques 1, 08028 Barcelona, Catalunya, Spain´ ´ `
Accepted 6 October 1998
Abstract
Ž .This report concerns the study of homologous and heterologous regulation of cell surface A adenosine receptors A R in a1 1
pituitary-derived cell line. This has been possible by the use of the recently developed anti-A R antibodies in immunocytochemical1
assays. Functional desensitization and internalization of A R in GH cells occurred after treatment with agonist but also with antagonist.1 4Ž .Epidermal growth factor EGF treatment led to the up-regulation of cell surface A R in GH cells. Confocal analysis evidenced an1 4
EGF-induced increase of A R present in intracellular clathrin-coated vesicles. The up-regulation was blocked by actinomycin D thus1
suggesting the involvement of protein synthesis in the effect induced by the growth factor. These results constitute the first example ofadenosine receptor regulation by EGF and one of the few examples of antagonist-induced desensitization and internalization amongG-protein-coupled receptors. q 1999 Elsevier Science B.V. All rights reserved.
Keywords: A adenosine receptor; Receptor regulation; Agonist; Antagonist; Epidermal growth factor; Pituitary cell1
1. Introduction
Regulation of receptor expression is a common mecha-nism to control cellular responses to extracellular signals.
ŽThus, the effect of physiological factors hormones, neuro-.transmitters and growth factors is determined by the
relative amount of cell surface receptors and the specificsignal transduction pathways involved. Like other mem-bers of the G-protein-coupled receptors family, A R are1
regulated in response to agonist or antagonist stimulation.Agonist-induced desensitization of A R has been de-1
scribed in intact animals and in cell cultures. Long-termadministration of agonists to animals cause functional de-
w xsensitization of A R in guinea pig heart 7 , rat adipocytes1w x w x w x22 , rat atrial muscle 23 and rat brain 10,33 . Thereduced functional response is due to a net loss of A R1Ž .down-regulation and to a decrease in the proportion ofA R displaying high-affinity sites for agonists. Growth of1
Abbreviations: A R: A adenosine receptor; DPCPX: 1,3-dipropyl-8-1 1Ž . Ž .cyclopentylxanthine; R-PIA: N6- R - phenylisopropyl adenosine
) Corresponding author. Fax: q 34-934-021219; E-mail:[email protected]; Internet home page: www.bq.ub.esrreceprfranco.html
rat adipocytes or smooth muscle DDT MF-2 cells in the1
presence of R-PIA leads to the desensitization and down-w xregulation of A R 14,15,31 . We have recently demon-1
strated that treatment of DDT MF-2 cells with the agonist1
R-PIA induces a rapid clustering and desensitization of thereceptor whereas loss of binding sites on the cell mem-brane due to internalization and down-regulation is a slower
w xevent occurring within hours 3 . A R are also regulated in1
an heterologous way in response to other hormones. Thus,chronic treatment of rats with benzodiazepines such asdiazepam induces the down-regulation of adenosine recep-
w xtors in the CNS 17 . In contrast, up-regulation of A R has1
been demonstrated in rat brain by glucocorticoid treatmentw x35 . On the other hand, morphine or opioids treatmentleads to up-regulation or not detectable changes in the
w xlevel of receptor expressed in different brain areas 21 .w xRecently, Arslan et al. 2 have described a down-regu-
lation of A adenosine receptors upon NGF-induced dif-2A
ferentiation of PC12 cells.EGF activates, via a tyrosine-kinase receptor, the ex-
pression of multiple genes and alters the growth anddifferentiation of a wide range of cell types. GH pituitary3
cells are derived from a rat pituitary tumour and secreteprolactin and growth hormone in response to thyrotropin-
0006-8993r99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved.Ž .PII: S0006-8993 98 01062-2
( )A. NaÕarro et al.rBrain Research 816 1999 47–5748
Ž .releasing hormone TRH . GH cells are clonal isolates of4
GH cells, selected for high prolactin production; both cell3
lines are extensively used as a model to study regulation ofw xhormone secretion in the endocrine system. Felix et al. 9
Žhave described that treatment of rat pituitary cells either.GH cultures or pituitary cultures from neonatal rats with3
EGF increased the proportion of classical lactotropes, cellsŽ .that only secrete prolactin PRL , and the amount of
prolactin secretion in these cells. In GH cells, the stimula-4
tion of prolactin synthesis by EGF results from an in-w xcreased PRL mRNA synthesis induction 1,12 . The effect
of dopamine and TRH, two important regulators of pro-lactin secretion on these cells, is also modulated by EGF.In fact, treatment with EGF caused a reduction in TRHreceptor number and a diminished response to TRH as aconsequence of a decrease in the content of cellular TRH
w xreceptor mRNA 18,28 . In contrast, EGF induces thew xexpression of functional D dopamine receptors 11 . Since2
adenosine acting through G-protein-coupled A R has been1
described to regulate PRL and GH secretion in pituitaryw xcells 6,8,27,36 it is possible that EGF affects the expres-
sion of A R in these cells.1
In this paper, the homologous regulation of A R by1
R-PIA, an agonist, and DPCPX, an antagonist, as well asthe heterologous regulation of these receptors by EGF,have been studied in a pituitary-derived cell line. Theresults are relevant since they provide evidence about theup-regulation of the adenosine receptors by EGF and theirdesensitization and internalization by both agonists andantagonists.
2. Materials and methods
2.1. Reagents
Ž . Ž . Ž .N6- R - phenylisopropyl adenosine R-PIA , cyclohex-imide, actinomycin D, calf intestine adenosine-deaminaseŽ .ADA , fura 2AM and EDTA were purchased from SigmaŽ .St. Louis, MO, USA . 1,3-dipropyl-8-cyclopentylxanthineŽ .DPCPX was from Research Biochemicals International
Table 1Ligand-induced desensitization of A R in GH cells1 4
Ž .Calcium response %
1 h 12 h 24 h
Control 100"6 100"7 100"10R-PIA-treated 70"10 50"5 39"5DPCPX-treated 80"10 47"7 42"9
Ž .Untreated cells or cells treated 1, 12 or 24 h with 100 mM R-PIA or 40mM DPCPX were extensively washed prior to the assays. The effect of40 mM R-PIA upon calcium mobilization induced by 10 mM KCl
Ž .depolarization was measured see Section 2 .Data are mean"SD from a representative experiment in triplicates.
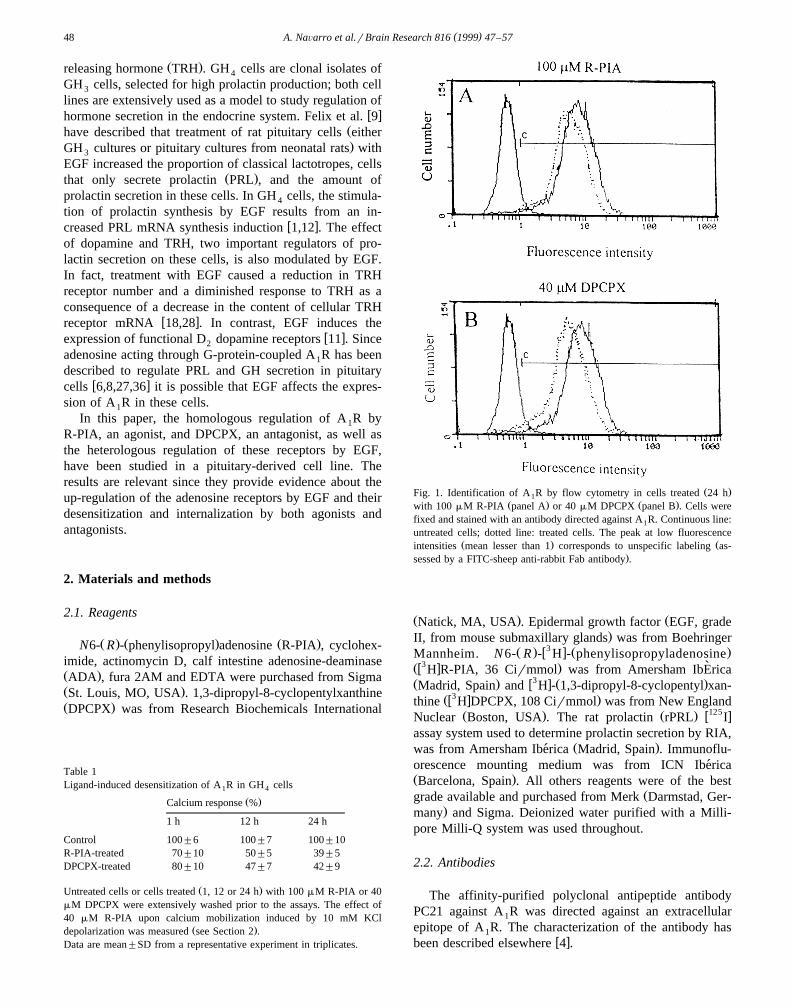
Ž .Fig. 1. Identification of A R by flow cytometry in cells treated 24 h1Ž . Ž .with 100 mM R-PIA panel A or 40 mM DPCPX panel B . Cells were
fixed and stained with an antibody directed against A R. Continuous line:1
untreated cells; dotted line: treated cells. The peak at low fluorescenceŽ . Žintensities mean lesser than 1 corresponds to unspecific labeling as-
.sessed by a FITC-sheep anti-rabbit Fab antibody .
Ž . ŽNatick, MA, USA . Epidermal growth factor EGF, grade.II, from mouse submaxillary glands was from Boehringer
Ž . w3 x Ž .Mannheim. N6- R - H - phenylisopropyladenosine3 `Žw x .H R-PIA, 36 Cirmmol was from Amersham IbErica
Ž . w3 x Ž .Madrid, Spain and H - 1,3-dipropyl-8-cyclopentyl xan-Žw3 x .thine H DPCPX, 108 Cirmmol was from New England
Ž . Ž . w125 xNuclear Boston, USA . The rat prolactin rPRL Iassay system used to determine prolactin secretion by RIA,
Ž .was from Amersham Iberica Madrid, Spain . Immunoflu-´orescence mounting medium was from ICN Iberica´Ž .Barcelona, Spain . All others reagents were of the best
Žgrade available and purchased from Merk Darmstad, Ger-.many and Sigma. Deionized water purified with a Milli-
pore Milli-Q system was used throughout.
2.2. Antibodies
The affinity-purified polyclonal antipeptide antibodyPC21 against A R was directed against an extracellular1
epitope of A R. The characterization of the antibody has1w xbeen described elsewhere 4 .

( )A. NaÕarro et al.rBrain Research 816 1999 47–57 49
Goat anti-clathrin and FITC-conjugated rabbit anti-goatIgG were from Sigma. Sheep anti-rabbit IgG-fluoresceinŽ Ž . .F ab fragment was from Boehringer Mannheim.2
2.3. Culture conditions and cell counting
Ž .GH C GH , a clonal rat pituitary cell line, was4 1 4
grown as a monolayer culture at 378C in a humidifiedatmosphere of 5% CO . Cells were maintained in Dul-2
Ž .becco’s modified eagle medium DMEM supplementedŽwith 15% horse serum Gibco, Life Technologies,
. ŽBarcelona, Spain , 2.5% fetal calf serum Boehringer.Mannheim Biochemical Spain, Barcelona , 1 mM sodium
pyruvate, 2 mM L-glutamine, 1% MEM–nonessentialamino acids, 50 mM 2-mercaptoethanol and antibioticsŽ100 Urml penicillin, 100 mgrml streptomycin and 0.25
.mgrml fungizone; these reagents were from Gibco . Thismedium is referred as complete DMEM medium.
ŽCells detached by addition of EDTA 5 mM final.concentration and further homogenized by repeated pipet-
ting were counted using a Multisizer particle counterŽ .Coulter, Hieleah, USA .
2.4. Ligand binding experiments
GH cells were seeded at 30,000 cellsrwell in 24-well4
plates. Twenty-four hours later, EGF was added in freshmedium at the indicated concentration. Binding was per-formed directly in 24-well plates using serum-free DMEMbuffered with 20 mM HEPES, pH 7.4, containing 2 Urml
w3 xADA. Competition of 4 nM H R-PIA or 3 nMw3 xH DPCPX binding was performed by incubation withdifferent concentrations of the indicated ligands. Nonspe-cific binding was defined as the binding remaining in the
Žpresence of an excess of displacer 200 mM R-PIA or 30.mM DPCPX . After standing enough time to achieve the
Ž . Ž . Ž .Fig. 2. Morphology of GH C pituitary cells. A Untreated cells. B Cells treated with 30 ngrml EGF for 72 h 200= .4 1
( )A. NaÕarro et al.rBrain Research 816 1999 47–5750
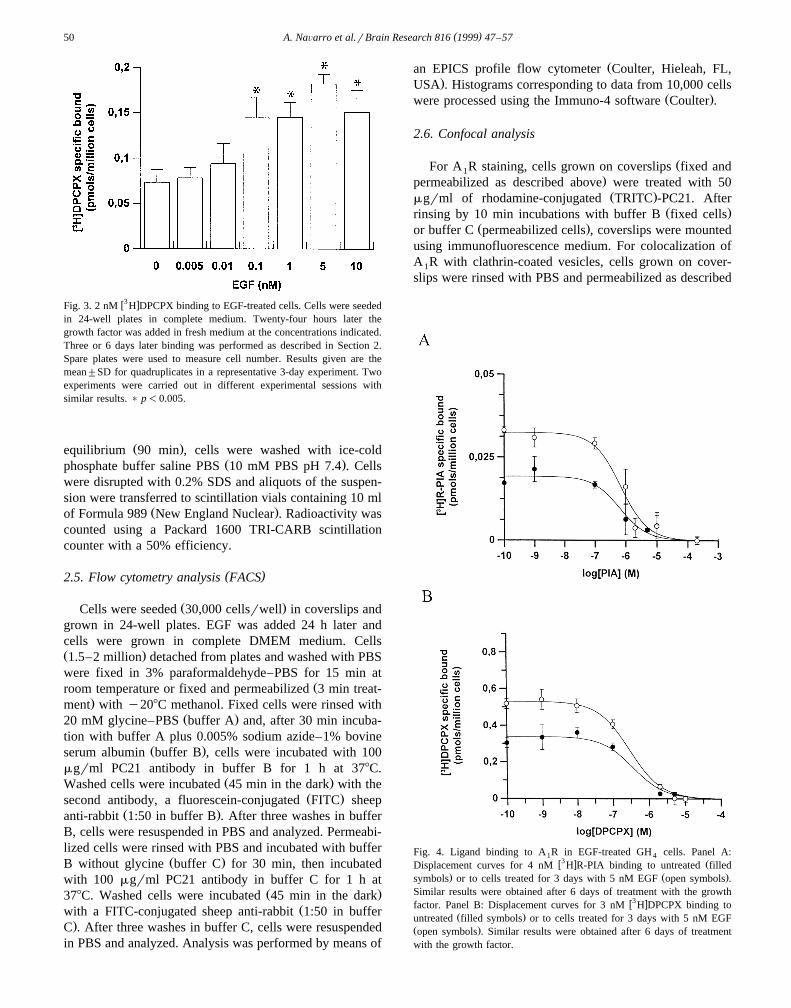
w3 xFig. 3. 2 nM H DPCPX binding to EGF-treated cells. Cells were seededin 24-well plates in complete medium. Twenty-four hours later thegrowth factor was added in fresh medium at the concentrations indicated.Three or 6 days later binding was performed as described in Section 2.Spare plates were used to measure cell number. Results given are themean"SD for quadruplicates in a representative 3-day experiment. Twoexperiments were carried out in different experimental sessions withsimilar results. ) p-0.005.
Ž .equilibrium 90 min , cells were washed with ice-coldŽ .phosphate buffer saline PBS 10 mM PBS pH 7.4 . Cells
were disrupted with 0.2% SDS and aliquots of the suspen-sion were transferred to scintillation vials containing 10 ml
Ž .of Formula 989 New England Nuclear . Radioactivity wascounted using a Packard 1600 TRI-CARB scintillationcounter with a 50% efficiency.
( )2.5. Flow cytometry analysis FACS
Ž .Cells were seeded 30,000 cellsrwell in coverslips andgrown in 24-well plates. EGF was added 24 h later andcells were grown in complete DMEM medium. CellsŽ .1.5–2 million detached from plates and washed with PBSwere fixed in 3% paraformaldehyde–PBS for 15 min at
Žroom temperature or fixed and permeabilized 3 min treat-.ment with y208C methanol. Fixed cells were rinsed with
Ž .20 mM glycine–PBS buffer A and, after 30 min incuba-tion with buffer A plus 0.005% sodium azide–1% bovine
Ž .serum albumin buffer B , cells were incubated with 100mgrml PC21 antibody in buffer B for 1 h at 378C.
Ž .Washed cells were incubated 45 min in the dark with theŽ .second antibody, a fluorescein-conjugated FITC sheep
Ž .anti-rabbit 1:50 in buffer B . After three washes in bufferB, cells were resuspended in PBS and analyzed. Permeabi-lized cells were rinsed with PBS and incubated with buffer
Ž .B without glycine buffer C for 30 min, then incubatedwith 100 mgrml PC21 antibody in buffer C for 1 h at
Ž .378C. Washed cells were incubated 45 min in the darkŽwith a FITC-conjugated sheep anti-rabbit 1:50 in buffer
.C . After three washes in buffer C, cells were resuspendedin PBS and analyzed. Analysis was performed by means of
Žan EPICS profile flow cytometer Coulter, Hieleah, FL,.USA . Histograms corresponding to data from 10,000 cells
Ž .were processed using the Immuno-4 software Coulter .
2.6. Confocal analysis
ŽFor A R staining, cells grown on coverslips fixed and1.permeabilized as described above were treated with 50Ž .mgrml of rhodamine-conjugated TRITC -PC21. After
Ž .rinsing by 10 min incubations with buffer B fixed cellsŽ .or buffer C permeabilized cells , coverslips were mounted
using immunofluorescence medium. For colocalization ofA R with clathrin-coated vesicles, cells grown on cover-1
slips were rinsed with PBS and permeabilized as described
Fig. 4. Ligand binding to A R in EGF-treated GH cells. Panel A:1 4w3 x ŽDisplacement curves for 4 nM H R-PIA binding to untreated filled
. Ž .symbols or to cells treated for 3 days with 5 nM EGF open symbols .Similar results were obtained after 6 days of treatment with the growth
w3 xfactor. Panel B: Displacement curves for 3 nM H DPCPX binding toŽ .untreated filled symbols or to cells treated for 3 days with 5 nM EGF
Ž .open symbols . Similar results were obtained after 6 days of treatmentwith the growth factor.
( )A. NaÕarro et al.rBrain Research 816 1999 47–57 51
above. After rinsing with PBS and a 15 min incubationwith buffer C, labelling was performed using goat anti-
Ž .clathrin antibody 1:40 dilution in buffer C for 1 h at378C. After four rinses in buffer B, a FITC-conjugated
Ž . Žrabbit anti-goat IgG 1:200 in buffer C was added 45.min, room temperature . Cells washed with buffer C, were
Ž .incubated with PC21-TRITC 50 mgrml in buffer C for 1h at 378C. After four washes with buffer C, coverslips
Žwere mounted in ImmunoFluore Mounting Medium ICN.Biomedicals . Preparations were observed using a confocal
Žscanning laser microscope Leica TCS 4D from Leica.Lasertechnick, Heidelberg, Germany adapted to an in-
verted Leitz DMIRBE microscope. Photographs were takenusing a Kodak Echtachrome Elite 100 film.
2.7. Prolactin secretion
For prolactin secretion experiments, cells seeded in6-well plates were incubated for 45 min at 378C in serum-free medium containing 0.1% bovine serum albumin, 20
mM HEPES pH 7.4 and 2 Urml ADA. Prolactin wasŽ .determined in the supernatants 1500=g, 10 min at 48C
Ž . w125 xusing the rat prolactin rPRL I assay system.
2.8. Calcium measurements
Calcium mobilization was induced by 10 mM KCldepolarization. The level of intracellular calcium was mea-
w xsured as described elsewhere 19 . Briefly, trypsinized andfura 2AM-loaded cells were washed twice in HBSS bufferŽ120 mM NaCl, 5 mM KCl, 0.4 mM MgCl , 0.4 mM2
CaCl , 10 mM HEPES, 10 mM NaHCO , 10 mM glucose2 3.and 5 mM Na HPO pH 7.4. After 30 min of standing,2 4
Žfluorescence corresponding to 4 million cells total vol-.ume: 2 ml was measured in a RF-5000 Shimadzu spectro-Žfluorimeter excitation wavelengths 340 and 380 nm; emis-
.sion wavelength: 505 nm . Calcium levels were estimatedw xas described by Grynkiewicz et al. 16 . Maximum fluores-
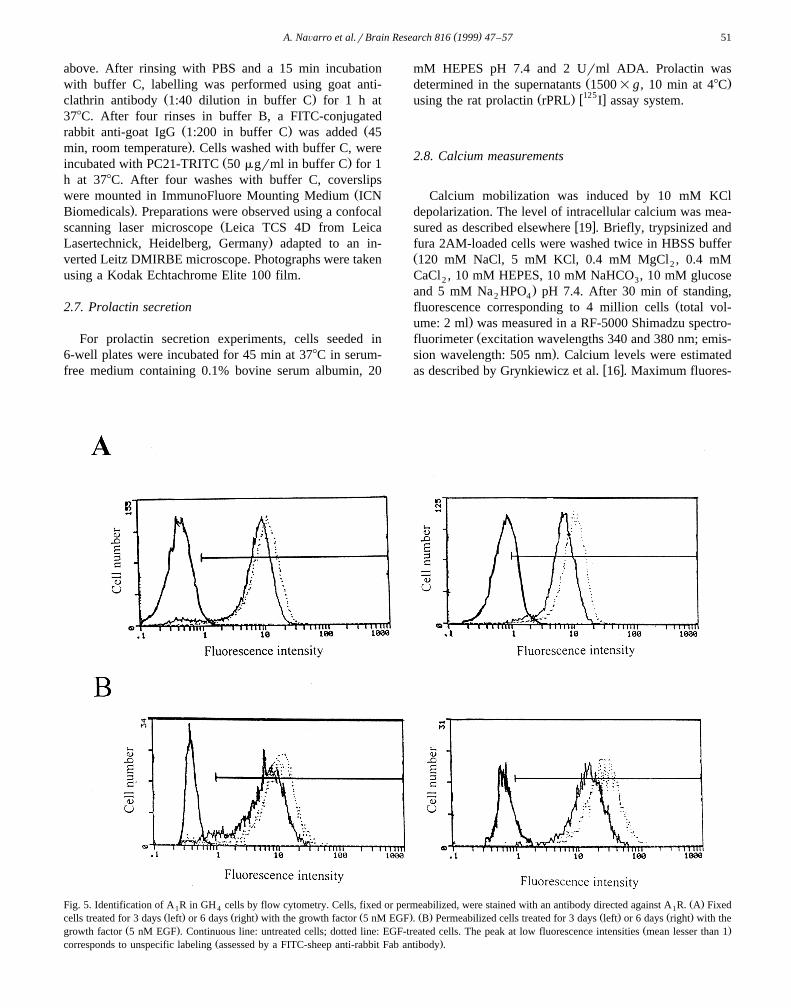
Ž .Fig. 5. Identification of A R in GH cells by flow cytometry. Cells, fixed or permeabilized, were stained with an antibody directed against A R. A Fixed1 4 1Ž . Ž . Ž . Ž . Ž . Ž .cells treated for 3 days left or 6 days right with the growth factor 5 nM EGF . B Permeabilized cells treated for 3 days left or 6 days right with the
Ž . Ž .growth factor 5 nM EGF . Continuous line: untreated cells; dotted line: EGF-treated cells. The peak at low fluorescence intensities mean lesser than 1Ž .corresponds to unspecific labeling assessed by a FITC-sheep anti-rabbit Fab antibody .
( )A. NaÕarro et al.rBrain Research 816 1999 47–5752
cence was determined after cell permeabilization with0.1% Triton and minimum fluorescence was determinedafter 0.1% Triton plus 5 mM EGTA treatment. Drugs wereadded into the cuvette at the time indicated by arrows inthe figures.
2.9. Data analysis
In binding experiments three replicates of each pointwere performed. Data from displacement experiments wereanalyzed by nonlinear regression. Goodness of fit was
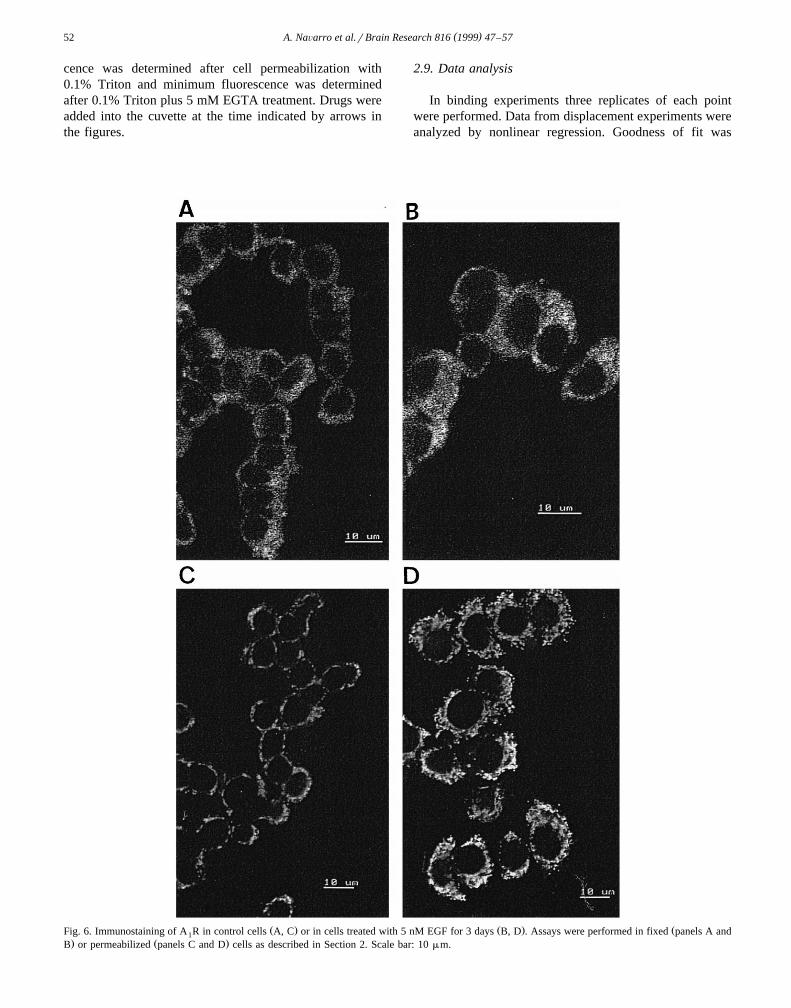
Ž . Ž . ŽFig. 6. Immunostaining of A R in control cells A, C or in cells treated with 5 nM EGF for 3 days B, D . Assays were performed in fixed panels A and1. Ž .B or permeabilized panels C and D cells as described in Section 2. Scale bar: 10 mm.
( )A. NaÕarro et al.rBrain Research 816 1999 47–57 53
tested according to the reduced SD values given by theregression program. If not indicated, each figure is repre-sentative of three experiments independently performed.
2.10. Statistical analysis
Statistical comparisons were made using a two-tailedStudent’s t-test.
3. Results
3.1. Homologous regulation of A R by R-PIA and DPCPX1
Activation of A R with adenosine or analogues leads to1
inhibition of the intracellular calcium flux caused by KClw xdepolarization 37 . Chronic treatment with R-PIA led to a
reduction in this response, i.e., to a functional desensitiza-tion. In fact, the effect of 100 mM R-PIA in cells previ-ously incubated with R-PIA and exhaustively washed,decreased by 30, 50 and 61%, respectively, after 1, 12 or
Ž .24 h of agonist treatment Table 1 . Unexpectedly, whenGH cells were treated with an antagonist such as DPCPX,4
desensitization took place to a similar extent than that
found with the agonist. Thus, the inhibition of the R-PIA-Žinduced functional response inhibition of calcium currents
.in depolarizing conditions in cells treated with the antago-nist was reduced to a similar extent than in the case of
Ž .cells treated with the agonist Table 1 .A R distribution on the cell surface was examined by1
immunocytochemistry and flow cytometry analysis. Un-treated and R-PIA- or DPCPX-treated cells were stainedusing fluorescein-conjugated rabbit anti-A R antibody1Ž .PC21-FITC and analyzed by means of a flow cytometer.Despite the treatment with the agonist did not change thepercentage of cells expressing A R, which approached1
100%, the intensity of the staining was reduced in the caseŽ .of R-PIA-treated cells Fig. 1 . Internalization of A R was1
even more evident in DPCPX-treated cells than in R-PIA-Ž .treated cells Fig. 1 . There was a decrease in the mean
fluorescence intensity of labelling exhibited by the popula-tion of treated cells; the decrease in R-PIA-treated cellswas 17"3% and the reduction in DPCPX-treated cellswas 22"4%. These results show that both agonist andantagonist cause desensitization and internalization of A R1
in pituitary cells.
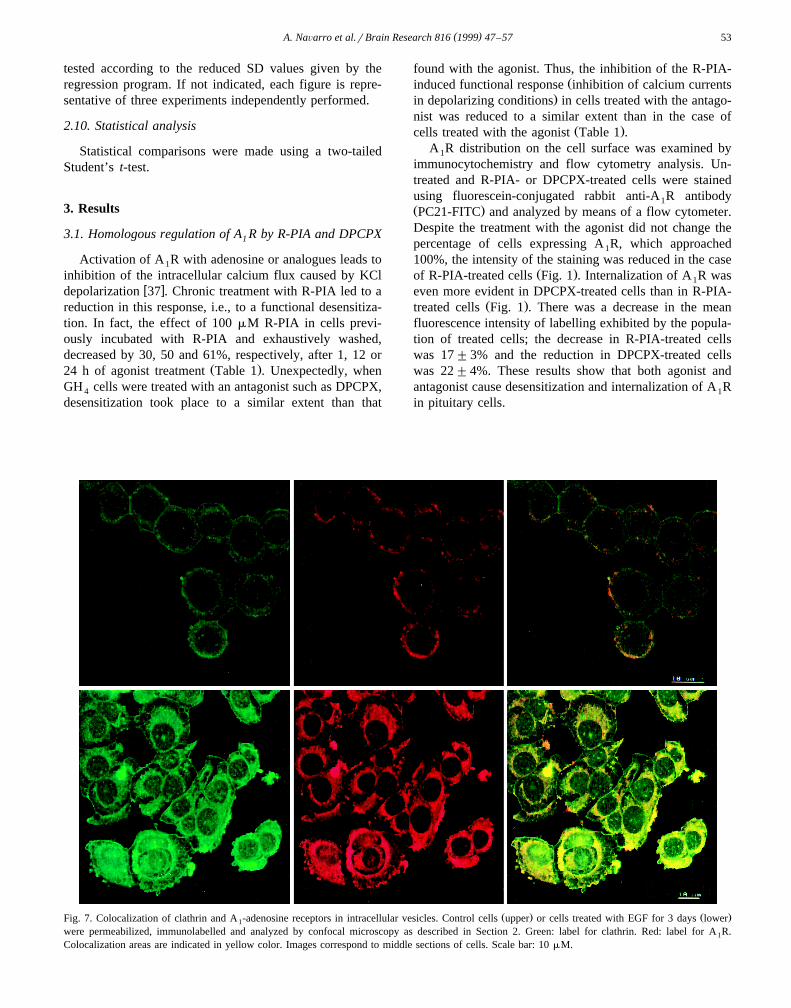
Ž . Ž .Fig. 7. Colocalization of clathrin and A -adenosine receptors in intracellular vesicles. Control cells upper or cells treated with EGF for 3 days lower1
were permeabilized, immunolabelled and analyzed by confocal microscopy as described in Section 2. Green: label for clathrin. Red: label for A R.1
Colocalization areas are indicated in yellow color. Images correspond to middle sections of cells. Scale bar: 10 mM.

( )A. NaÕarro et al.rBrain Research 816 1999 47–5754
3.2. Heterologous regulation of A adenosine receptors by1
EGF
Treatment of GH C pituitary cells with 5 nM EGF for4 1
3 days induced an increase in size and a change in theŽ .shape of cells in culture Fig. 2 . This change in the
morphology from a rounded to an elongated pattern wasdue in part to a higher attachment to the dish and in part todifferentiation. An increase in basal prolactin secretion,from 1.3"0.2 ng PRLr106 cells to 29"1 ng PRLr106
cells was also found after 3 days in culture with EGF. ThisŽ .basal not TRH induced secretion of prolactin measured
after a 45 min incubation in a buffered nonsupplementedmedium was not modified by the presence of R-PIAŽ 6 .26"1 ng PRLr10 cells .
To analyze whether the growth factor has any effect onadenosine receptor expression, cells were treated withdifferent concentrations of the agent for 3 to 6 days andthen ligand binding experiments were performed. Theresults show an increase in the antagonist binding ex-
w3 x Žpressed as pmol H DPCPX bound per million cells Fig..3 . To better analyze if EGF was enhancing cell surface
expression of the receptor or the change in binding wasdue to an increased receptor affinity, we carried out dis-
w3 xplacement experiments of either H R-PIA binding byw3 x Ž .R-PIA or H DPCPX binding by DPCPX Fig. 4 . The
IC values for R-PIA and DPCPX were similar in the50
case of control cells or in cells treated with the growthfactor for 3 or 6 days: after 3 days of treatment, the IC50
for R-PIA was 0.7"0.2 mM vs. a value of 0.5"0.2 mM
ŽFig. 8. Immunostaining of A R in cells treated with EGF and actinomycin D. Cells were permeabilized prior to the staining procedure. Control cells panel1. Ž . Ž . Ž . Ž .A and cells treated 24 h with either 2 mgrml actinomycin D panel B , 5 nM EGF panel C or with the two reagents panel D were permeabilized and
stained as described in Section 2.
( )A. NaÕarro et al.rBrain Research 816 1999 47–57 55
for untreated cells. For DPCPX the values were 0.4"0.2Ž .mM and 0.5"0.3 mM in control and treated 3 days
cells, respectively.We next studied the presence of A R using specific1
polyclonal antibodies developed in our laboratory andw xwhose characterization has been described elsewhere 4 .
ŽImmunolabelling of cell surface receptors, on fixed Fig.. Ž .5A or permeabilized cells Fig. 5B was analyzed by flow
cytometry. There was an increase in the mean fluorescenceintensity of labelling exhibited by the population of treatedcells. At 3 days of treatment with EGF, the increase in cellsurface labelling was 18"2% and reached a 38"4%after 6 days. The increase in the cellular content of A R1
measured in permeabilized cells was more marked, goingfrom 39"10% at 3 days to 93"14% at 6 days oftreatment.
The analysis of fixed cells by confocal microscopy,shows an increase of the label on the plasma membrane of
Ž .treated cells Fig. 6B referred to that found in untreatedŽ .cells Fig. 6A . These results are in agreement with the
flow cytometry analysis shown in Fig. 5. The analysis ofimmunolabelled EGF-treated cells by confocal microscopyallowed us to identify a high amount of adenosine receptorin intracellular vesicles, which present a punctate labelling
Ž .pattern Fig. 6D . This pattern is different from that foundŽ .in control cells Fig. 6C . Confocal microscopy using
anti-clathrin antibodies led to identify the A R-containing1
intracellular vesicles as clathrin-coated vesicles. This wasevident by the high level of colocalization between A R1
Žand clathrin found in both untreated and treated cells Fig..7 .
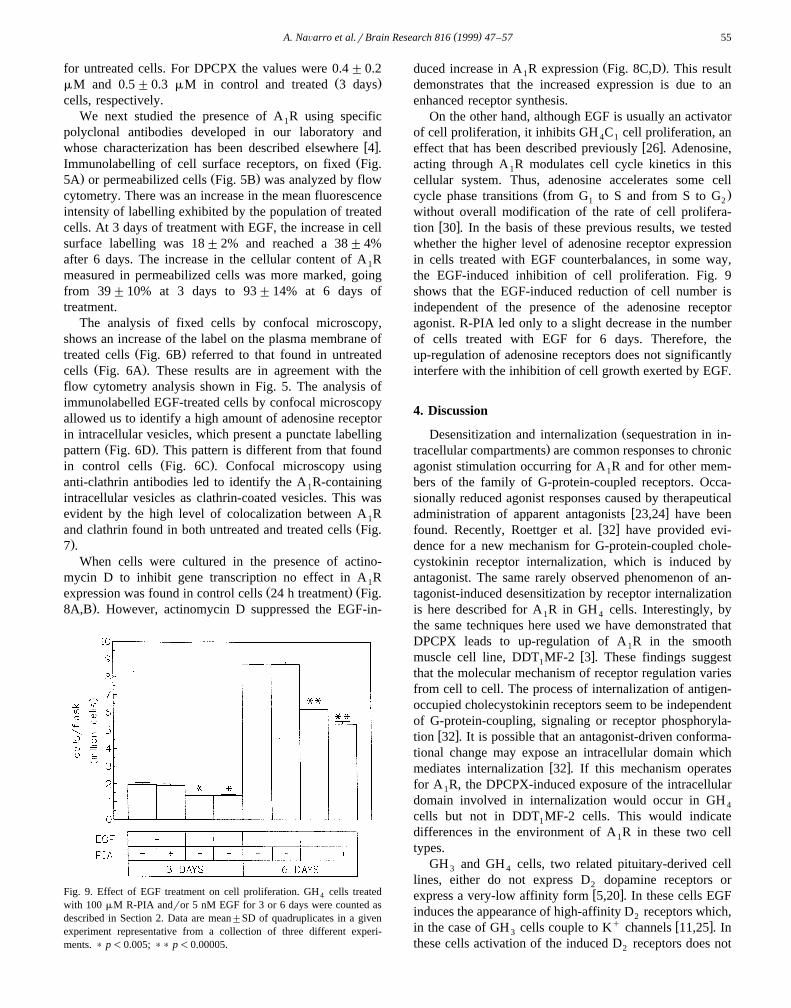
When cells were cultured in the presence of actino-mycin D to inhibit gene transcription no effect in A R1
Ž . Žexpression was found in control cells 24 h treatment Fig..8A,B . However, actinomycin D suppressed the EGF-in-
Fig. 9. Effect of EGF treatment on cell proliferation. GH cells treated4
with 100 mM R-PIA andror 5 nM EGF for 3 or 6 days were counted asdescribed in Section 2. Data are mean"SD of quadruplicates in a givenexperiment representative from a collection of three different experi-ments. ) p-0.005; )) p-0.00005.
Ž .duced increase in A R expression Fig. 8C,D . This result1
demonstrates that the increased expression is due to anenhanced receptor synthesis.
On the other hand, although EGF is usually an activatorof cell proliferation, it inhibits GH C cell proliferation, an4 1
w xeffect that has been described previously 26 . Adenosine,acting through A R modulates cell cycle kinetics in this1
cellular system. Thus, adenosine accelerates some cellŽ .cycle phase transitions from G to S and from S to G1 2
without overall modification of the rate of cell prolifera-w xtion 30 . In the basis of these previous results, we tested
whether the higher level of adenosine receptor expressionin cells treated with EGF counterbalances, in some way,the EGF-induced inhibition of cell proliferation. Fig. 9shows that the EGF-induced reduction of cell number isindependent of the presence of the adenosine receptoragonist. R-PIA led only to a slight decrease in the numberof cells treated with EGF for 6 days. Therefore, theup-regulation of adenosine receptors does not significantlyinterfere with the inhibition of cell growth exerted by EGF.
4. Discussion
ŽDesensitization and internalization sequestration in in-.tracellular compartments are common responses to chronic
agonist stimulation occurring for A R and for other mem-1
bers of the family of G-protein-coupled receptors. Occa-sionally reduced agonist responses caused by therapeutical
w xadministration of apparent antagonists 23,24 have beenw xfound. Recently, Roettger et al. 32 have provided evi-
dence for a new mechanism for G-protein-coupled chole-cystokinin receptor internalization, which is induced byantagonist. The same rarely observed phenomenon of an-tagonist-induced desensitization by receptor internalizationis here described for A R in GH cells. Interestingly, by1 4
the same techniques here used we have demonstrated thatDPCPX leads to up-regulation of A R in the smooth1
w xmuscle cell line, DDT MF-2 3 . These findings suggest1
that the molecular mechanism of receptor regulation variesfrom cell to cell. The process of internalization of antigen-occupied cholecystokinin receptors seem to be independentof G-protein-coupling, signaling or receptor phosphoryla-
w xtion 32 . It is possible that an antagonist-driven conforma-tional change may expose an intracellular domain which
w xmediates internalization 32 . If this mechanism operatesfor A R, the DPCPX-induced exposure of the intracellular1
domain involved in internalization would occur in GH 4
cells but not in DDT MF-2 cells. This would indicate1
differences in the environment of A R in these two cell1
types.GH and GH cells, two related pituitary-derived cell3 4
lines, either do not express D dopamine receptors or2w xexpress a very-low affinity form 5,20 . In these cells EGF
induces the appearance of high-affinity D receptors which,2q w xin the case of GH cells couple to K channels 11,25 . In3
these cells activation of the induced D receptors does not2

( )A. NaÕarro et al.rBrain Research 816 1999 47–5756
lead to the classical response defined for this receptorw xsubtype, i.e., does not inhibit cAMP formation 25,26 . As
w xwe have identified low affinity A R on GH cells 30 and1 4
due to the fact that the A receptors couple to the same G1 iw xprotein than D receptors do 34 , it was possible that EGF2
would induce the expression of high affinity adenosinereceptors, thus leading to a common mechanism for het-erologous receptor regulation via EGF receptors. The datapresented here demonstrate that despite up-regulation ofthe A R by EGF, the growth factor does not promote the1
presence of high affinity adenosine receptors. In fact, theinduced A R present the same low affinity than A R from1 1
untreated GH cells. Binding analysis shows clearly a4
higher amount of receptor present at the cell surface butdisplaying the same IC value for the agonist, R-PIA, or50
for the antagonist, DPCPX, than in untreated cells. Thesefindings constitute a direct evidence of the multiple path-ways taking place in heterologous receptor regulation.
By immunocytochemistry it was found that the intra-cellular traffick of the adenosine receptor goes mainly viaclathrin-coated vesicles since both A R and clathrin shows1
a high degree of colocalization in these vesicles. Thiscolocalization occurs in both untreated or treated cells,thus suggesting that coated vesicles are important contribu-tors to the adenosine receptor trafficking. We have foundthat EGF effect is sensitive to actinomycin D treatment,suggesting that up-regulation of A R by EGF is due to an1
increased protein synthesis. This is consistent with theincreased amount of intracellular A R-containing vesicles1
induced by EGF. Other reports have suggested a role forclathrin-containing coated vesicles in the intracellular traf-
w xficking of A R expressed in rat brain cells 13,33 .1
Despite the increased expression of adenosine receptorswe have not found a strong evidence for a regulatory effectof adenosine on the inhibition of cell proliferation inducedby EGF. Hence we believe that the effect of EGF onadenosine receptor expression is not important for cellproliferation and may have other purposes. As EGF in-duces the differentiation of the cell line into a lactotrophcell type it is tempting to speculate that the higher level ofA receptors is convenient for a more efficient feed-back1
control of secretion. Data reported by Dorflinger andw x w xSchonbrunn 8 and Delahunty et al. 6 indicate that 100
mM R-PIA reduces by a 37% the prolactin secreted bycontrol cells: from 1.3"0.2 ng PRLr106 cells to 0.8"0.1ng PRLr106 cells. Furthermore, adenosine acceleratesprogression towards the cell cycle, leading the cells to staylonger in the G rG phase, in which they are less suscep-0 1
w xtible to respond to secretory signals 30 . A similar feed-back control can be hypothesized for D dopamine recep-2
tors induced by EGF. Whereas EGF promotes prolactinw x w xmRNA formation 12 and secretion 9,1 , activation of the
induced D receptors leads to inhibition of prolactin re-2w xlease in neuroendocrine cells 26 . It should however noted
that the release of prolactin in EGF-treated cells is veryhigh even in absence of exogenous TRH. In these circum-
stances, activation of up-regulated A receptors in EGF-1
treated cells did not lead to any effect upon prolactinrelease. These results and the A -mediated stimulation of1
w xprolactin release from anterior-pituitaries in vitro 36 indi-cate that the actions of adenosine are multiple and dependupon the cell phenotype and, likely, the hormonal status. Inwhat concerns the mechanism of stimulation of prolactin
w xsynthesis and secretion Aanestad et al. 1 have reportedthat EGF increases the intracellular concentration of freecalcium. The concentration of intracellular calcium can becontrolled by extracellular adenosine, which, acting via
w xA R, inhibits L-type calcium channels in GH cells 37 .1 4
The autocrine and paracrine action of EGF in rat ante-rior pituitary cell cultures has been described by Mouihate
w xand Lestage 29 . From the overall cell population, a 19%of cells are able of secreting EGF and a 48% of cellspresent EGF receptors. Therefore, it is likely that theresults described here are of physiological relevance. A R1
can now be added to the list of the known targets for thegrowth factor in cells from pituitary origin. Since the wayEGF modulates D dopamine and TRH receptors is differ-2
ent from that here described for A R, pituitary cells prove1
to be a good but complex model to study heterologousregulation of G-protein-coupled receptors. On the otherhand, the DPCPX-induced desensitization of A R should1
be taken into account in the use of antagonists of adeno-sine receptors as therapeutic drugs.
Acknowledgements
ŽSupported by a joint Echevarne Foundation and Span-. Ž .ish Ministry of Education PETRI Grant PTR92r0047
administered by Fundacio Bosch i Gimpera and by grants´Ž .from CIRIT-CICYT QFN93r4423 and from CICYT
Ž .PB94r0941 . We are indebted to the Fundacion Marıa´ ´Francisca de RoÕiralta for financial support in the acquisi-tion of the spectrofluorimeter needed for the measurementof intracellular calcium levels. We acknowledge the helpreceived from Drs. Jaume Comas and Susana Castel, re-sponsible of the flow cytometry section and confocalmicroscopy, respectively, in the SerÕeis Cientifico Tecnics`de la UniÕersitat de Barcelona. We thank Robin Rycroftfrom the SerÕei d’Assessorament Linguistic de la UniÕersi-¨tat de Barcelona for the excellent technical assistance inthe preparation of the manuscript. A. Navarro is recipientof a fellowship of Formacion de Personal InÕestigador´from the Spanish Ministerio de Educacion y Ciencia.´
References
w x1 M. Aanestad, J.S. Rotnes, P.A. Torjesen, E. Haug, O. Sand, T.Bjoro, Epidermal growth factor stimulates the prolactin synthesis
Ž .and secretion in rat pituitary cells in culture GH C cells by4 1
increasing the intracellular concentration of free calcium, Acta En-Ž .docrinol. 128 1993 361–366.

( )A. NaÕarro et al.rBrain Research 816 1999 47–57 57
w x2 G. Arslan, E. Konty, B.B. Fredholm, Down-regulation of adenosineA receptors upon NGF-induced differentiation of PC12 cells,2A
Ž .Neuropharmacology 36 1997 1319–1326.w x3 F. Ciruela, C. Saura, E.I. Canela, J. Mallol, C. Lluis, R. Franco,
Ligand-induced phosphorylation, clustering and desensitization ofŽ .A adenosine receptors, Mol. Pharmacol. 52 1997 788–797.1
w x4 F. Ciruela, V. Casado, J. Mallol, E.I. Canela, C. Lluis, R. Franco,´Immunological identification of A adenosine receptors in brain1
Ž .cortex, J. Neurosci. Res. 42 1995 818–828.w x5 M.J. Cronin, N. Faure, J.A. Martial, R.I. Weiner, Absence of
high-affinity dopamine receptors in GH cells: a prolactin-secreting3
clone resistant to the inhibitory action of dopamine, EndocrinologyŽ .106 1980 718–723.
w x6 T.M. Delahunty, M.J. Cronin, J. Linden, Regulation of GH -cell3Ž .function via adenosine A receptors, Biochem. J. 255 1988 69–77.1
w x7 D.M. Dennis, J.C. Shryock, L. Belardinelli, Homologous desensiti-zation of the A adenosine receptor system in the guinea-pig atri-1
Ž .oventricular node, J. Pharmacol. Exp. Ther. 272 1995 1024–1035.w x8 L.J. Dorflinger, A. Schonbrunn, Adenosine inhibits prolactin and
growth hormone secretion in a clonal pituitary cell line, Endocrinol-Ž .ogy 117 1985 2330–2337.
w x9 R. Felix, U. Meza, G. Cota, Induction of classical lactotropes byepidermal growth factor in rat pituitary cell cultures, Endocrinology
Ž .136 1995 939–946.w x10 M. Fernandez, P. Svenningsson, B.B. Fredholm, Adaptative changes´
in adenosine receptors following long-term treatment with theadenosine receptor agonist R-phenylisopropyladenosine, Life Sci. 9Ž .1996 769–776.
w x11 R. Gardette, R. Rasolonjanahary, C. Kordon, A. Enjalbert, Epider-mal growth factor treatment induces D dopamine receptors func-2
Ž .tionally coupled to delayed outward potassium current I inKŽ .GH C clonal anterior pituitary cells, Neuroendocrinology 59 19944 1
10–19.w x12 C.A. Gilchrist, J.D. Shull, Epidermal growth factor induces prolactin
mRNA in GH C cells via a protein synthesis-dependent pathway,4 1Ž .Mol. Cell. Endocrinol. 92 1993 201–206.
w x13 G. Gonzalez-Calero, M. Martın, A. Cubero, A. Andres, Bovine brain´ ´coated-vesicles contain adenosine A receptors. Presence of adeny-1
Ž .late cyclase coupled to the receptor, J. Neurochem. 67 19901469–1477.
w x14 A. Green, J.L. Johnson, G. Milligan, Down-regulation of G sub-i
types by prolonged incubation of adipocytes with an A adenosine1Ž .receptor agonist, J. Biol. Chem. 265 1990 5206–5210.
w x15 A. Green, G. Milligan, S.B. Dobias, G down-regulation as ai
mechanism for heterologous desensitization in adipocytes, J. Biol.Ž .Chem. 267 1992 3223–3229.
w x 2q16 G. Grynkiewicz, M. Poenie, R.Y. Tsien, A new generation of Caindicators with greatly improved fluorescence properties, J. Biol.
Ž .Chem. 260 1985 3440–3450.w x17 M. Hawkins, M. Pravika, M. Radulovacki, Chronic administration of
diazepam down-regulates adenosine receptors in the rat brain, Phar-Ž .macol. Biochem. Behav. 30 1988 303–308.
w x18 P.M. Hinkle, E.D. Shanshala II, Z. Yan, Epidermal growth factorŽ .decreases the concentration of thyrotropin-releasing hormone TRH
receptors and TRH responses in pituitary GH C cells, Endocrinol-4 1Ž .ogy 129 1991 1283–1288.
w x19 M.A. Javors, M. Liu, B.S. Cuvelier, C.L. Bowden, Characterizationof the effect of the adenosine agonist cyclohexyladenosine on platelet
w 2q xactivating factor-induced increases in Ca in human platelets iniŽ .vitro, Cell Calcium 10 1990 647–653.
w x20 J.M. Johnston, D.F. Wood, E.A. Bolaji, D.G. Johnston, The
dopamine D receptor is expressed in GH cells, J. Mol. En-2 3Ž .docrinol. 7 1991 131–136.
w x21 G.B. Kaplan, K.A. Leite-Morris, M.T. Sears, Alterations of adeno-Ž .sine A receptors in morphine dependence, Brain Res. 657 19941
347–350.w x22 J.P. Longabaugh, J. Didsbury, A. Spiegel, G.L. Stiles, Modification
of the rat adipocyte A adenosine receptor–adenylate cyclase system1
during chronic exposure to an A adenosine receptor agonist: alter-1
ations in the quantity of G and G are not associated withsa ia
Ž .changes in their mRNAs, Mol. Pharmacol. 36 1989 681–688.w x23 H.T. Lee, C.I. Thompson, A. Hernandez, J.L. Lewy, F.L. Belloni,´
Cardiac desensitization to adenosine analogues after prolonged R-PIAŽ .infusion in vivo, Am. J. Physiol. 265 1993 H1916–H1927.
w x24 H.Y. Meltzer, J.F. Nash, Effect of antipsychotic drugs on serotoninŽ .receptors, Pharmacol. Rev. 43 1991 588–600.
w x25 C. Missale, L. Castelletti, F. Boroni, M. Memo, P. Spano, Epidermalgrowth factor induces the functional expression of dopamine recep-
Ž .tors in the GH cell line, Endocrinology 128 1991 13–20.3w x26 C. Missale, F. Boroni, L. Castelletti, R.D. Toso, N. Gabellini, S.
Sigala, P.F. Spano, Lack of coupling of D receptors to adenylate2
cyclase in GH cells exposed to epidermal growth factor, J. Biol.3Ž .Chem. 266 1991 23392–23398.
w x27 P. Mollard, N. Guerineau, C. Chiavaroli, W. Schlegel, D.M.F.´Cooper, Adenosine A receptor-induced inhibition of Ca2q tran-1
sients linked to action potentials in clonal pituitary cells, Eur. J.Ž .Pharmacol. 206 1991 271–277.
w x28 T. Monden, M. Yamada, S. Konaka, T. Satoh, H. Ezawa, T.Iwasaki, Mori, Transcriptional down-regulation by epidermal growthfactor of TRH receptor mRNA in rat pituitary cells, J. Mol. En-
Ž .docrinol. 15 1995 73–79.w x29 A. Mouihate, J. Lestage, Epidermal growth factor: a potential
paracrine and autocrine system within the pituitary, Neuroen-Ž .docrinology 6 1995 1273–1276.
w x30 A. Navarro, R. Zapata, E.I. Canela, J. Mallol, C. Lluis, R. Franco,Modulation of GH cell cycle via A adenosine receptors, J. Neu-4 1
Ž .rochem. 69 1997 2145–2154.w x31 V. Ramkumar, M.E. Olah, K.A. Jacobson, G.L. Stiles, Distinct
pathways of desensitization of A - and A - adenosine receptors in1 2Ž .DDT MF-2 cells, Mol. Pharmacol. 40 1991 639–647.1
w x32 B.F. Roettger, D. Ghanekat, R. Rao, C. Toledo, J. Yingling, D.Pinon, L.J. Miller, Antagonist-stimulated internalization of the G-
Ž .protein-coupled cholecystokinin receptor, Mol. Pharmacol. 51 1997357–362.
w x33 A. Ruiz, J.M. Sanz, G. Gonzalez-Calero, M. Fernandez, A. Cubero,´M. Ros, Desensitization and internalization of adenosine A recep-1
tors in rat brain by in vivo treatment with R-PIA: involvement ofŽ .coated vesicles, Biochim. Biophys. Acta 1310 1996 168–174.
w x34 S.E. Senogles, A.M. Spiegel, E. Padrell, R. Iyengar, M.G. Caron,Specificity of receptor–G-protein interactions. Discrimination of Gi
subtypes by the D dopamine receptor in a reconstituted system, J.2Ž .Biol. Chem. 265 1990 4507–4514.
w x35 P. Svenningsson, B.B. Fredholm, Glucocorticoids regulate the ex-pression of adenosine A but not A receptors in rat brain, J.1 2A
Ž .Pharmacol. Exp. Ther. 280 1997 1094–1101.w x36 W.H. Yu, M. Kimura, A. Walczewska, J.C. Porter, S.M. McCann,
Adenosine acts by A receptors to stimulate release of prolactin1
from anterior-pituitaries in vitro, Proc. Natl. Acad. Sci. USA 95Ž .1998 7795–7798.
w x37 R. Zapata, A. Navarro, E.I. Canela, R. Franco, C. Lluis, J. Mallol,Regulation of L-type calcium channels via A adenosine receptors,1
Ž .J. Neurochem. 69 1997 2546–2554.




















