
Homodimerization of the Wnt Receptor DERAILED Recruits the Src Family Kinase SRC64B

Carcinogenesis vol.31 no.10 pp.1844–1853, 2010doi:10.1093/carcin/bgq169Advance Access publication August 23, 2010
Specific targeting of Wnt/b-catenin signaling in human melanoma cells by a dietarytriterpene lupeol
Rohinton S.Tarapore1,2, Imtiaz A.Siddiqui2,Mohammad Saleem2,3, Vaqar M.Adhami2,Vladimir S.Spiegelman1,2 and Hasan Mukhtar1,2,�
1Molecular and Environmental Toxicology Center, University of Wisconsin,Madison, WI 53706, USA and 2Department of Dermatology, School ofMedicine and Public Health, University of Wisconsin, Madison, WI 53706,USA3Present address: Department of Molecular Chemoprevention andTherapeutics, The Hormel Institute, University of Minnesota, Austin, MN55912, USA
�To whom correspondence should be addressed. Department of Dermatology,University of Wisconsin-Madison, Medical Sciences Center, Room 4385, 1300University Avenue, Madison, WI 53706, USA. Tel: þ1 608 263 3927;Fax: þ1 608 263 5223;Email: [email protected]
Wingless (Wnt) signaling pathway regulates a variety of cellularprocesses including proliferation, differentiation, survival,apoptosis and cell motility. Aberrant activation of Wnt/b-cateninpathway has been observed in approximately one-third of mela-nomas and this subset has very poor prognosis suggesting thattargeting Wnt signaling could be a promising strategy against thissubtype. Mel 928 and Mel 1241 melanoma cells representative ofcells with constitutive activation of Wnt/b-catenin signaling path-way and Mel 1011 representative of cells that lack this pathwaywere treated with a dietary triterpene lupeol and its effects ongrowth, proliferation, b-catenin transcriptional activity and Wnttarget genes were determined both in vitro and in vivo. Lupeoltreatment to Mel 928 andMel 1241 but not Mel 1011 cells resultedin a dose-dependent (i) decrease in cell viability, (ii) induction ofapoptosis, (iii) decrease in colonogenic potential, (iv) decrease inb-catenin transcriptional activity and (v) decrease in the expres-sion of Wnt target genes. Most importantly, lupeol restricted thetranslocation of b-catenin from the cytoplasm to the nucleus.Lupeol also decreased the growth of Mel 928 but not Mel 1011-derived tumors implanted in the athymic nude mice. The decreasein Mel 928-derived tumor growth was associated with a decreasein the expression of Wnt target genes c-myc, cyclin D1, prolifer-ation markers proliferating cell nuclear antigen and Ki-67 andinvasion marker osteopontin. We suggest that lupeol alone or asan adjuvant to current therapies could be developed as an agentfor the management of human melanomas harboring constitutiveWnt/b-catenin signaling.
Introduction
Melanoma, the most serious form of skin cancer, is curable in itsinitial stages but when detected at later stages, is one of the mostlethal malignancies (1,2). The increasing incidence of melanomaand its poor prognosis advocates the need for the development ofnovel preventive approaches such as the use of specific targetingagents derived from natural products (3).
Lupeol [Lup-20(29)-en-3b-ol], a dietary triterpene, found in vari-ous fruits, vegetables and some medicinal herbs (4–8) has been shownto possess strong anti-inflammatory, anticarcinogenic, anti-mutagenicand anti-malarial activity both in vitro and in vivo (5,7,9–15). Our
recent studies have also shown that lupeol possesses chemopreventiveeffects in a two-stage mouse skin carcinogenesis model (16).
Wnt/b-catenin signaling pathway is reported to play an importantrole in embryogenesis, stem cell maintenance and tumorigenesis,including melanoma progression (17–19). b-catenin is a key com-ponent of the Wnt signaling pathway and its translocation to thenucleus initiates transcription of downstream target genes (20).Overexpression of Wnt ligands, mutations in components of Wntpathway and stabilization of b-catenin are the most common alter-ations associated with constitutive activation of Wnt signaling path-way and are observed in a variety of cancers including melanomas.Melanoma cells exhibit differential b-catenin expression with Mel1241 cell line carrying a stabilizing mutation of b-catenin, whereasMel 928 expresses wild-type b-catenin without detectable adeno-matosis polyposis coli (APC). Mel 1011 cells harbor both intactb-catenin and APC. Mel 928 and Mel 1241 cells are also character-ized by nuclear localization of b-catenin and constitutive activationof b-catenin/Tcf-dependent transcription (21,22). It is noteworthythat the hallmark of active Wnt signaling, nuclear localization ofb-catenin, has been observed in about a third of primary and meta-static human melanomas (21,23).
Recently, we demonstrated that lupeol inhibits the growth of met-astatic melanoma cells in vitro and in vivo (24). While defining themechanism of these inhibitory effects of lupeol on melanoma cells,we made a novel observation that it specifically targets melanomacells that harbor constitutive Wnt/b-catenin signaling pathway. Thisinformation coupled with our previously published data (24), whichdemonstrated that lupeol exerts negligible toxicity on normal humanmelanocytes, advocates that lupeol could be developed as a potentialagent for the management of melanomas that harbor constitutive Wnt/b-catenin signaling pathway.
Materials and methods
Materials
The plasmids dominant-negative T-cell factor (TCF) 4, b-cateninS33Y, pcDNAand TOP-Flash reporter plasmid bearing the TCF4-binding sequence wereobtained from Drs K.Kinzler and B.Vogelstein (25–28), whereas pTK-puroplasmid was obtained from Addgene (Cambridge, MA). Renilla luciferase(pRL-TK) plasmid was obtained from Promega (Madison, WI). Precast 12%Tris-glycine polyacrylamide gels were obtained from Invitrogen (Carlsbad,CA). Antibodies against proliferating cell nuclear antigen (PCNA), Ki-67,osteopontin, b-catenin, vascular endothelial growth factor (VEGF) and matrixmetalloproteinase (MMP) 2/9 antibodies that detect activated proteins wereobtained from Santa Cruz Biotechnology (Santa Cruz, CA). Antibodies againstb-actin, cyclin D1, c-myc were procured from Sigma (St Louis, MO), CellSignaling Technology (Danvers, MA) and BD Biosciences Pharmingen (SanJose, CA), respectively. Pathscan� ELISA kit for b-catenin was obtained fromCell Signaling Technology.
Cell lines and cell culture
The melanoma cell lines Mel 928, Mel 1241 and Mel 1011 were kindlyprovided by Dr Paul Robbins (Center of Cancer Research, National CancerInstitute, Bethesda, MD) (22). Cells were maintained inDulbecco’s modifiedEagle’s medium, supplemented with 10% fetal bovine serum and 1%antibiotic–antimycotic solution, containing penicillin, streptomycin and am-photericin B (PSM) under standard growth conditions (5% CO2, 37�C, humid-ified atmosphere). The 293T cells were obtained from the American TypeCulture Collection, Manassas, VA.
Treatment of cells with lupeol
A stock solution of lupeol (10 mM) was prepared by dissolving it in warmethanol and diluting in dimethyl sulfoxide in a 1:1 ratio. For dose-dependentstudies, the cells (50% confluent) were treated with lupeol (20–60 lM) for 48 hin complete cell media. All treatment protocols and controls were conducted asdescribed previously (24).
Abbreviations: APC, adenomatosis polyposis coli; MMP, matrix metallopro-teinase; PCNA, proliferating cell nuclear antigen; TCF, T-cell factor; TIMP,tissue inhibitors of matrix metalloproteinase; VEGF, vascular endothelialgrowth factor.
� The Author 2010. Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected] 1844
Cell proliferation assay
The effect of lupeol on the viability of melanoma cells was determinedby 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay(Sigma) as described earlier (24). A total of 3 � 103 cells per well in200 ll complete medium were seeded in a 96-well plate and treated withlupeol as described earlier (24). All treatment concentrations were repeatedin eight wells.
Caspase 3/7 activity assay
In a 96-well white-walled luminometer plate, 3 � 103 cells per well wereseeded in 200 ll media. The following day, the cells were treated with lupeolas described earlier. Each concentration was repeated in eight wells. Thecaspase 3/7 activity was measured using Caspase-Glo� 3/7 Assay (Promega)as per instructions of the manufacturer. The luminescence of each sample wasread using a luminometer.
Colony formation assay
Cells (800 cells per well) were seeded in a six-well plate and treated thefollowing day with lupeol (40 and 60 lM). The media was removed andreplaced with fresh media containing lupeol every 3 days. After 17–20 daysposttreatment, the colonies were fixed with formaldehyde, stained with crystalviolet (Ricca Chemical Company, Arlington, TX) and counted.
To test the efficacy of lupeol upon modulation of Wnt/b-catenin signaling,melanoma cells (Mel 928 and Mel 1241) were co-transfected with pTK-puroplasmid (Addgene) and dominant-negative TCF4 constructs. Mel 1011 cellswere also co-transfected with pTK-puro and stable b-catenin (b-cateninS33Y).Transfections were performed using Lipofectamine-2000 reagent, according tomanufacturer’s recommendations (Invitrogen). Twelve hours post-transfection,
cells were screened against puromycin (1.25 lg/ml). Forty-eight hours aftertransfection, the cells were treated with lupeol as described earlier and thelupeol treatment was repeated every 2 days. Seven to ten days posttreatment,the colonies were fixed with formaldehyde, stained with crystal violet (RiccaChemical Company). The plates were scanned for pictorial representation. Thecrystal violet was dissolved in 50% acetic acid and absorbance at 540 nm wasmeasured for graphical representation of the data.
Annexin V staining
The annexin V-FLUOS staining kit (Roche Applied Biosciences, Indianapolis,IN) was used for the detection of apoptotic cells. Assay was performed asdescribed (29).
Luciferase reporter assay
Using Lipofectamine-2000 reagent, melanoma cells were transfected as pervendor’s protocol (Invitrogen). Fresh media containing lupeol (40 and 60 lM)was added 12 h post-transfection. After 24 h, the cells were harvested andtranscriptional activity was measured in terms of luciferase activity by usingthe dual-luciferase reporter assay system (Promega) as per manufacturer’srecommendations. The transcriptional activity was measured in a dose- andtime-dependent manner.
Preparation of cell lysates for immunoblot analysis
Whole cell, cytosolic and nuclear lysates were prepared and western blottingwas performed as described earlier (24,29).
Enzyme-linked immunosorbent assay
b-catenin ELISA kit was used for the quantitative analysis of b-catenin incytosolic and nuclear lysates as per vendor’s protocol. The absorbance was
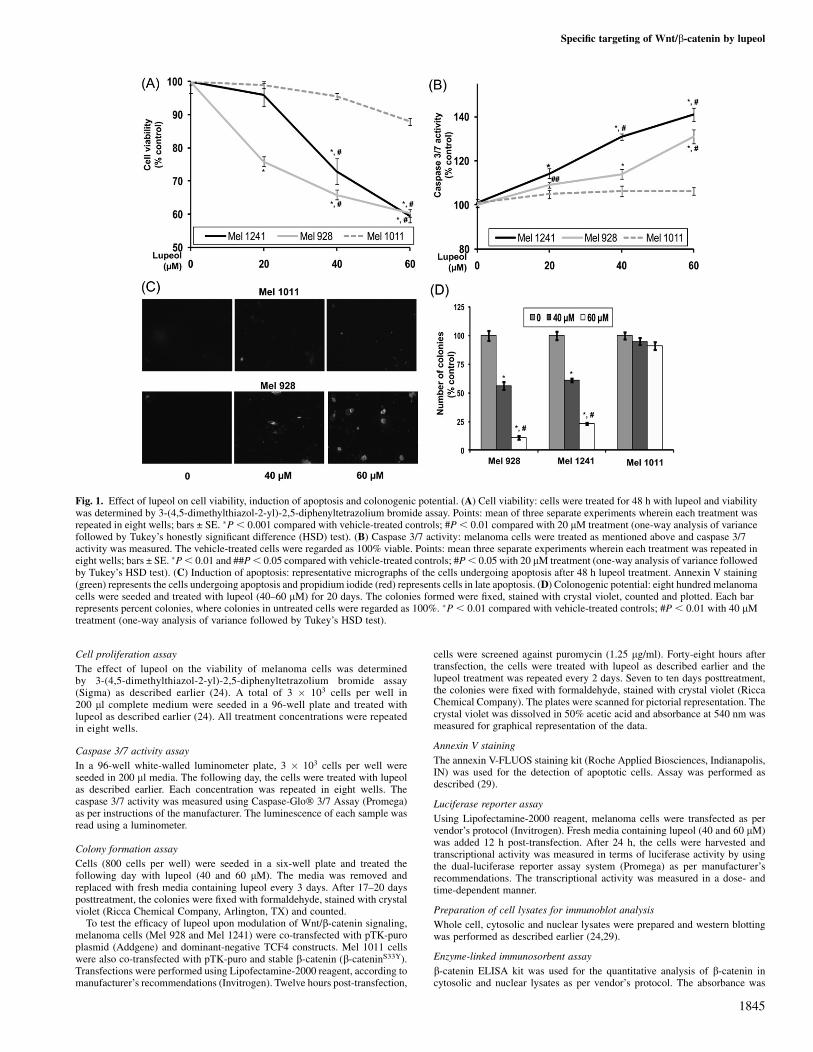
Fig. 1. Effect of lupeol on cell viability, induction of apoptosis and colonogenic potential. (A) Cell viability: cells were treated for 48 h with lupeol and viabilitywas determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay. Points: mean of three separate experiments wherein each treatment wasrepeated in eight wells; bars ± SE. �P, 0.001 compared with vehicle-treated controls; #P, 0.01 compared with 20 lM treatment (one-way analysis of variancefollowed by Tukey’s honestly significant difference (HSD) test). (B) Caspase 3/7 activity: melanoma cells were treated as mentioned above and caspase 3/7activity was measured. The vehicle-treated cells were regarded as 100% viable. Points: mean three separate experiments wherein each treatment was repeated ineight wells; bars ± SE. �P, 0.01 and ##P, 0.05 compared with vehicle-treated controls; #P, 0.05 with 20 lM treatment (one-way analysis of variance followedby Tukey’s HSD test). (C) Induction of apoptosis: representative micrographs of the cells undergoing apoptosis after 48 h lupeol treatment. Annexin V staining(green) represents the cells undergoing apoptosis and propidium iodide (red) represents cells in late apoptosis. (D) Colonogenic potential: eight hundred melanomacells were seeded and treated with lupeol (40–60 lM) for 20 days. The colonies formed were fixed, stained with crystal violet, counted and plotted. Each barrepresents percent colonies, where colonies in untreated cells were regarded as 100%. �P , 0.01 compared with vehicle-treated controls; #P , 0.01 with 40 lMtreatment (one-way analysis of variance followed by Tukey’s HSD test).
Specific targeting of Wnt/b-catenin by lupeol
1845
taken at 450 nm by using ELISA reader (Multiscan MCC/340; FisherScientific, Pittsburgh, PA).
Immunocytostaining
Melanoma cells were cultured on chambered slides and treated with lupeol asindicated previously. The cells were fixed in cold acetone for 5 min andblocked with 2.5% normal serum. Slides were incubated overnight withanti-b-catenin (Santa Cruz Biotechnology, Santa Cruz, CA) followed byfluorescence-tagged secondary antibody, covered with ProLong Gold antifadereagent with 4#,6-diamidino-2-phenylindole (Invitrogen) and visualized usingNikon Eclipse Ti microscope. Images were captured with an attached camera.
Tumorigenicity studies in athymic nude mice
Athymic (nu/nu) nude mice (male; 4–6 weeks old) were obtained from Na-tional Cancer Institute and housed in the animal care facility at The Universityof Wisconsin School of Medicine and Public Health under pathogen-free con-ditions with 12 h light–dark schedule) and fed with autoclaved Harlan Tekladsterilizable rodent diet (Harlan, Madison, WI) and water ad libitum. We de-termined the efficacy of lupeol in two settings reflective of chemopreventiveand chemotherapeutic protocols. Twenty-four athymic nude mice were im-planted with human melanoma cells that harbor (Mel 928) or lack (Mel1011) constitutive Wnt/b-catenin signaling. Melanoma cells (1 � 106) weresuspended in a 50 ll media and 50 ll Matrigel (BD Biosciences Pharmingen)and inoculated subcutaneously on the left and right flanks of athymic nudemice. On the following day, the animals were randomly divided into threegroups reflective of control, chemopreventive and chemotherapeutic protocols.
For the chemopreventive approach, the animals received an intraperito-neal injection of lupeol (40 mg/kg body wt) in 200 ll of corn oil 24 hpostinoculation. This dose was used successfully in our prior studies andis physiologically attainable and is non-toxic (24). For the therapeutic ap-proach, the treatment with lupeol was initiated when the tumor volumereached �150 mm3. This group of animals received intraperitoneal injectionof lupeol (40 mg/kg body wt) in 200 ll of corn oil. The control groupreceived corn oil alone, which was initiated 24 h postinoculation. All treat-ments with lupeol were performed thrice a week until the termination of theexperiment. The animals were monitored closely for tumor growth. Thetreatment schedule was continued until the tumors in the control group
reached a volume of �800 mm3. At this time, the animals were withdrawnfrom the study and euthanized. The tumors were excised, snap frozen inliquid nitrogen and kept at �80�C for further biochemical analysis.Throughout the experiment, the animals were monitored for possibletreatment-associated toxicity and changes in body weight. Tumor sizewas measured by determining two perpendicular dimensions and the heightusing vernier caliper. The tumor volume was calculated using the formulaV 5 1/2(4p/3) (L1/2) (L2/2) (H) 5 0.5238 L1L2H, where L1 is the longdiameter, L2 is short diameter and H is the height (30).
Preparation of tumor lysates
For immunoblotting analysis, a portion of the tumor tissues was thawed on iceand homogenized using a hand held homogenizer in 700 ll ice-cold lysisbuffer as described earlier (24). The tumor lysates were stored at �80�C.The protein concentration was determined by the BCA protein assay kit usingthe manufacturer’s protocol (Pierce, Rockford, IL).
Immunohistochemical analysis
Immunohistochemical staining was performed as described earlier (24,31).Paraffin-embedded sections were de-waxed, rehydrated, blocked and incu-bated overnight with primary antibody followed by subsequent secondaryfor 1 h. All the antibodies were procured from Santa Cruz Biotechnology.The stained slides were dehydrated, mounted in Permount and visualized usingNikon Eclipse Ti microscope. Images were captured with a camera attached tothe microscope.
Densitometry analysis
Immunoblots were scanned by HP Precisionscan Pro 3.13 (Hewlett-PackardCo., Palo Alto, CA). Densitometry measurements of the scanned bands wereperformed using digitalized scientific software program UN-SCAN-IT (SilkScientific Corporation, Orem, UT). Data were normalized to b-actin or suitableloading controls and expressed as mean ± SEM followed by appropriate sta-tistical analysis.
Statistical analysis
Data were analyzed for statistical significance with the use of analysis ofvariance followed by Tukey’s honestly significant difference test. Values of
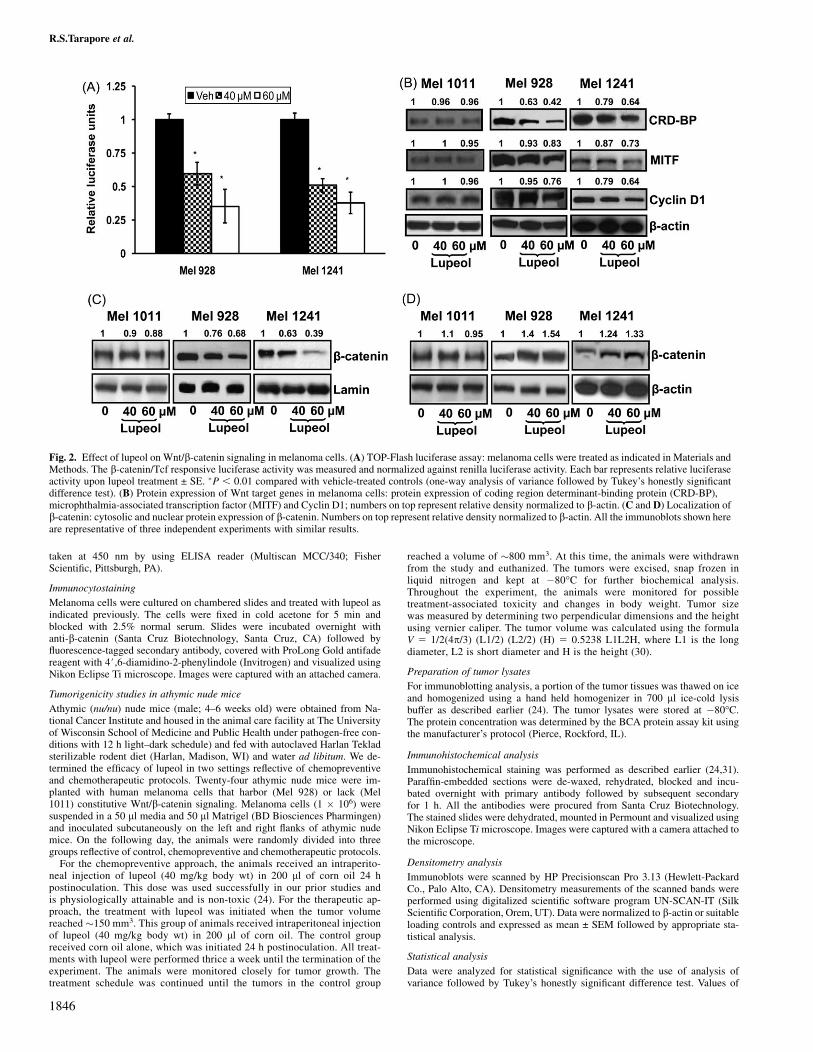
Fig. 2. Effect of lupeol on Wnt/b-catenin signaling in melanoma cells. (A) TOP-Flash luciferase assay: melanoma cells were treated as indicated in Materials andMethods. The b-catenin/Tcf responsive luciferase activity was measured and normalized against renilla luciferase activity. Each bar represents relative luciferaseactivity upon lupeol treatment ± SE. �P , 0.01 compared with vehicle-treated controls (one-way analysis of variance followed by Tukey’s honestly significantdifference test). (B) Protein expression of Wnt target genes in melanoma cells: protein expression of coding region determinant-binding protein (CRD-BP),microphthalmia-associated transcription factor (MITF) and Cyclin D1; numbers on top represent relative density normalized to b-actin. (C and D) Localization ofb-catenin: cytosolic and nuclear protein expression of b-catenin. Numbers on top represent relative density normalized to b-actin. All the immunoblots shown hereare representative of three independent experiments with similar results.
R.S.Tarapore et al.
1846
P , 0.05 were considered to be statistically significant. For in vivoexperiments, survival estimates were performed by ‘R-Project’ for statisticalcomputing.
Results
Effect of lupeol on human melanoma cell viability, induction ofapoptosis and colonogenic potential
Using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromideassay, we first evaluated the effect of lupeol (20–60 lM; 48 h) on cellviability. Lupeol treatment resulted in a significant dose-dependentdecrease in the viability of Mel 928 and Mel 1241 cells that exhibitconstitutively active Wnt/b-catenin signaling. These effects wereabsent in Mel 1011 cells that lack constitutively active Wnt/b-cateninsignaling (Figure 1A). The IC50 values for Mel 928, Mel 1241 andMel 1011 were 75, 72 and 135 lM, respectively.
We next determined the effect of lupeol treatment on capsase 3/7activity in melanoma cells. Lupeol treatment caused a dose-dependentincrease in caspase 3/7 activity in Mel 928 and Mel 1241 cells, whereasno significant change was observed in Mel 1011 cells (Figure 1B).We next determined the effect of lupeol on induction of apoptosis inmelanoma cells. We observed increased staining of annexin V in Mel928 cells, whereas minimal staining was observed in Mel 1011 cellsindicating that lupeol treatment resulted in increased apoptosis in Mel928 cells but not in Mel 1011 cells (Figure 1C).
Next, we asked whether treatment with lupeol could exert greateractivity on the formation of colonies, which allows an investigationover a longer period of time. More than 50% decrease in the number
of colonies was observed in Mel 928 and Mel 1241 cells, whereas nosignificant decrease in the number or size of colonies was observed inMel 1011 cells treated with lupeol (Figure 1D).
Effect of lupeol on Wnt/b-catenin signaling in melanoma cells
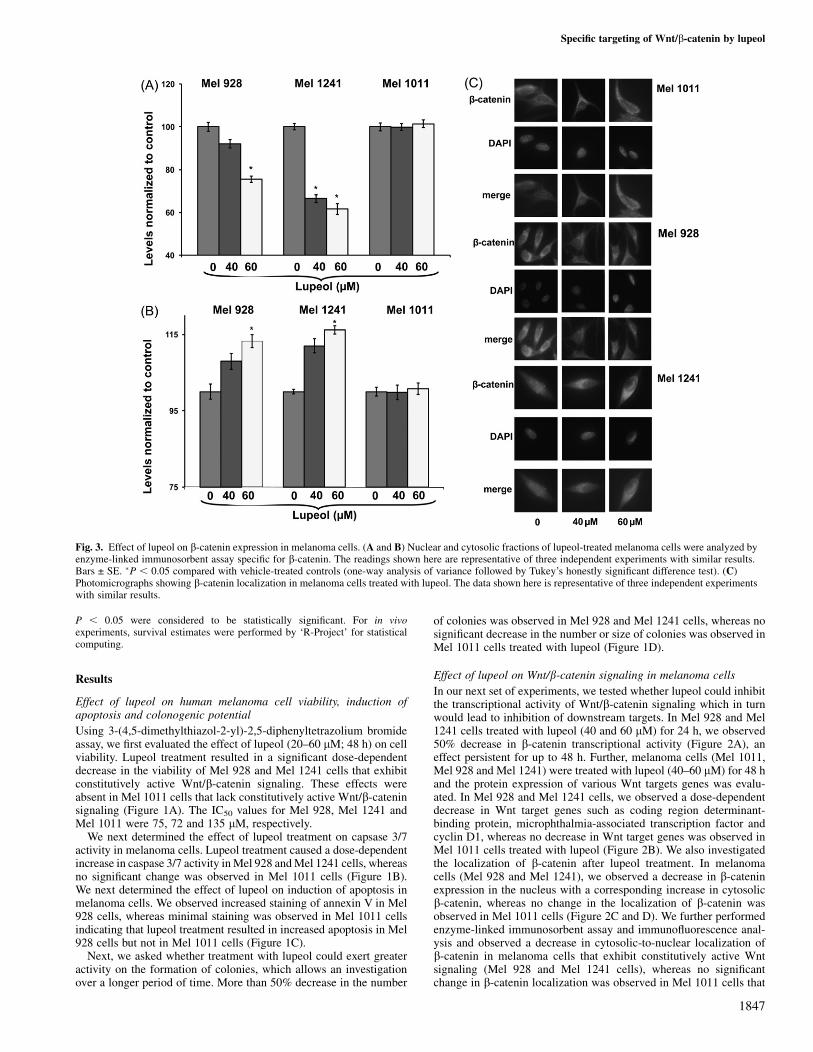
In our next set of experiments, we tested whether lupeol could inhibitthe transcriptional activity of Wnt/b-catenin signaling which in turnwould lead to inhibition of downstream targets. In Mel 928 and Mel1241 cells treated with lupeol (40 and 60 lM) for 24 h, we observed50% decrease in b-catenin transcriptional activity (Figure 2A), aneffect persistent for up to 48 h. Further, melanoma cells (Mel 1011,Mel 928 and Mel 1241) were treated with lupeol (40–60 lM) for 48 hand the protein expression of various Wnt targets genes was evalu-ated. In Mel 928 and Mel 1241 cells, we observed a dose-dependentdecrease in Wnt target genes such as coding region determinant-binding protein, microphthalmia-associated transcription factor andcyclin D1, whereas no decrease in Wnt target genes was observed inMel 1011 cells treated with lupeol (Figure 2B). We also investigatedthe localization of b-catenin after lupeol treatment. In melanomacells (Mel 928 and Mel 1241), we observed a decrease in b-cateninexpression in the nucleus with a corresponding increase in cytosolicb-catenin, whereas no change in the localization of b-catenin wasobserved in Mel 1011 cells (Figure 2C and D). We further performedenzyme-linked immunosorbent assay and immunofluorescence anal-ysis and observed a decrease in cytosolic-to-nuclear localization ofb-catenin in melanoma cells that exhibit constitutively active Wntsignaling (Mel 928 and Mel 1241 cells), whereas no significantchange in b-catenin localization was observed in Mel 1011 cells that
Fig. 3. Effect of lupeol on b-catenin expression in melanoma cells. (A and B) Nuclear and cytosolic fractions of lupeol-treated melanoma cells were analyzed byenzyme-linked immunosorbent assay specific for b-catenin. The readings shown here are representative of three independent experiments with similar results.Bars ± SE. �P , 0.05 compared with vehicle-treated controls (one-way analysis of variance followed by Tukey’s honestly significant difference test). (C)Photomicrographs showing b-catenin localization in melanoma cells treated with lupeol. The data shown here is representative of three independent experimentswith similar results.
Specific targeting of Wnt/b-catenin by lupeol
1847
do not harbor constitutive activation of Wnt/b-catenin signaling(Figure 3).
Effect of lupeol administration on tumor growth in athymic nude mice
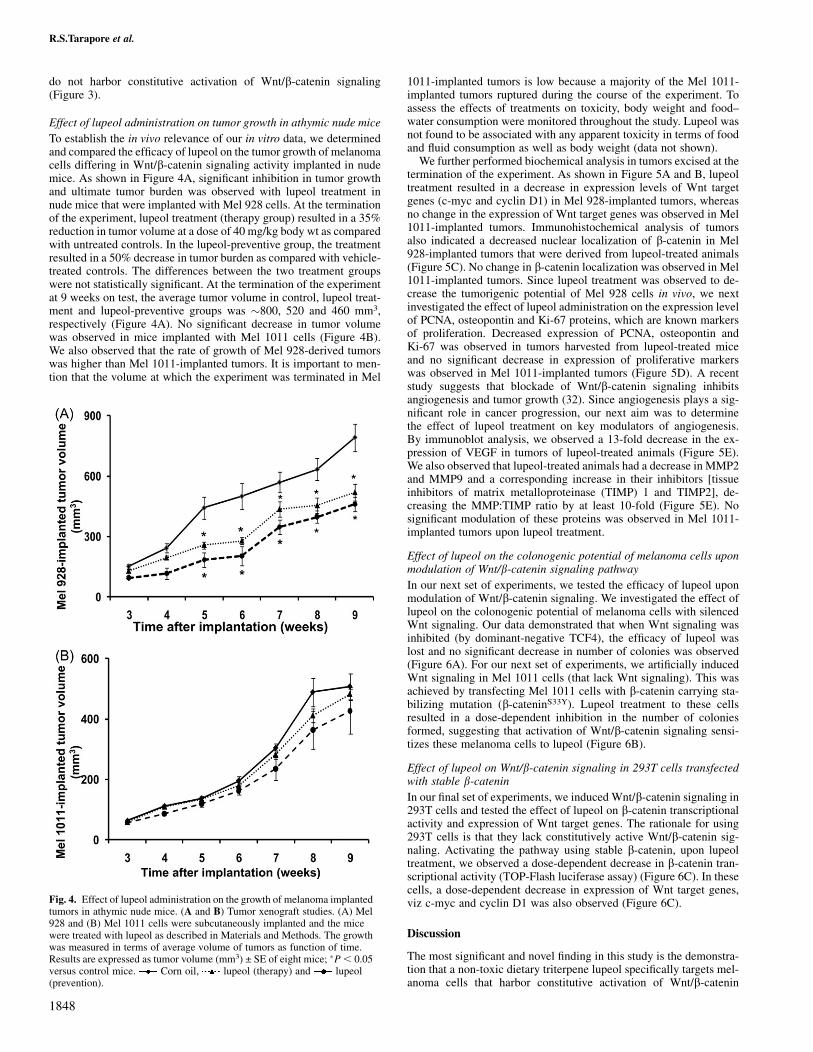
To establish the in vivo relevance of our in vitro data, we determinedand compared the efficacy of lupeol on the tumor growth of melanomacells differing in Wnt/b-catenin signaling activity implanted in nudemice. As shown in Figure 4A, significant inhibition in tumor growthand ultimate tumor burden was observed with lupeol treatment innude mice that were implanted with Mel 928 cells. At the terminationof the experiment, lupeol treatment (therapy group) resulted in a 35%reduction in tumor volume at a dose of 40 mg/kg body wt as comparedwith untreated controls. In the lupeol-preventive group, the treatmentresulted in a 50% decrease in tumor burden as compared with vehicle-treated controls. The differences between the two treatment groupswere not statistically significant. At the termination of the experimentat 9 weeks on test, the average tumor volume in control, lupeol treat-ment and lupeol-preventive groups was �800, 520 and 460 mm3,respectively (Figure 4A). No significant decrease in tumor volumewas observed in mice implanted with Mel 1011 cells (Figure 4B).We also observed that the rate of growth of Mel 928-derived tumorswas higher than Mel 1011-implanted tumors. It is important to men-tion that the volume at which the experiment was terminated in Mel
1011-implanted tumors is low because a majority of the Mel 1011-implanted tumors ruptured during the course of the experiment. Toassess the effects of treatments on toxicity, body weight and food–water consumption were monitored throughout the study. Lupeol wasnot found to be associated with any apparent toxicity in terms of foodand fluid consumption as well as body weight (data not shown).
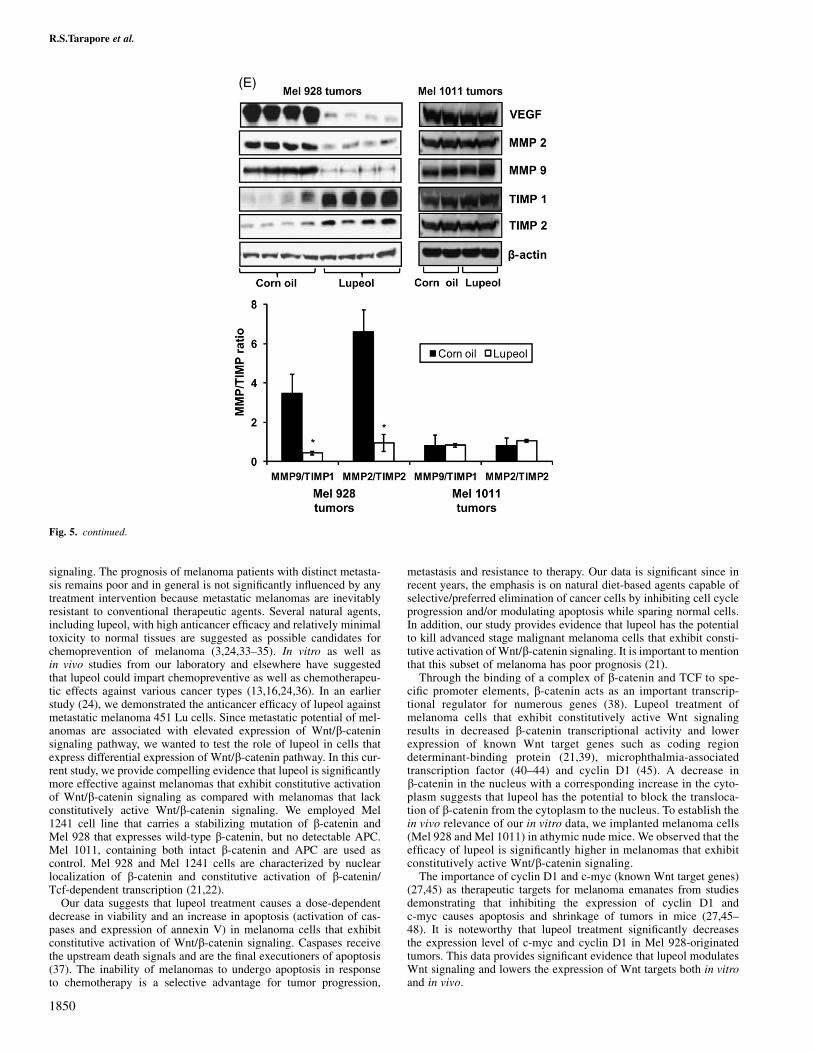
We further performed biochemical analysis in tumors excised at thetermination of the experiment. As shown in Figure 5A and B, lupeoltreatment resulted in a decrease in expression levels of Wnt targetgenes (c-myc and cyclin D1) in Mel 928-implanted tumors, whereasno change in the expression of Wnt target genes was observed in Mel1011-implanted tumors. Immunohistochemical analysis of tumorsalso indicated a decreased nuclear localization of b-catenin in Mel928-implanted tumors that were derived from lupeol-treated animals(Figure 5C). No change in b-catenin localization was observed in Mel1011-implanted tumors. Since lupeol treatment was observed to de-crease the tumorigenic potential of Mel 928 cells in vivo, we nextinvestigated the effect of lupeol administration on the expression levelof PCNA, osteopontin and Ki-67 proteins, which are known markersof proliferation. Decreased expression of PCNA, osteopontin andKi-67 was observed in tumors harvested from lupeol-treated miceand no significant decrease in expression of proliferative markerswas observed in Mel 1011-implanted tumors (Figure 5D). A recentstudy suggests that blockade of Wnt/b-catenin signaling inhibitsangiogenesis and tumor growth (32). Since angiogenesis plays a sig-nificant role in cancer progression, our next aim was to determinethe effect of lupeol treatment on key modulators of angiogenesis.By immunoblot analysis, we observed a 13-fold decrease in the ex-pression of VEGF in tumors of lupeol-treated animals (Figure 5E).We also observed that lupeol-treated animals had a decrease in MMP2and MMP9 and a corresponding increase in their inhibitors [tissueinhibitors of matrix metalloproteinase (TIMP) 1 and TIMP2], de-creasing the MMP:TIMP ratio by at least 10-fold (Figure 5E). Nosignificant modulation of these proteins was observed in Mel 1011-implanted tumors upon lupeol treatment.
Effect of lupeol on the colonogenic potential of melanoma cells uponmodulation of Wnt/b-catenin signaling pathway
In our next set of experiments, we tested the efficacy of lupeol uponmodulation of Wnt/b-catenin signaling. We investigated the effect oflupeol on the colonogenic potential of melanoma cells with silencedWnt signaling. Our data demonstrated that when Wnt signaling wasinhibited (by dominant-negative TCF4), the efficacy of lupeol waslost and no significant decrease in number of colonies was observed(Figure 6A). For our next set of experiments, we artificially inducedWnt signaling in Mel 1011 cells (that lack Wnt signaling). This wasachieved by transfecting Mel 1011 cells with b-catenin carrying sta-bilizing mutation (b-cateninS33Y). Lupeol treatment to these cellsresulted in a dose-dependent inhibition in the number of coloniesformed, suggesting that activation of Wnt/b-catenin signaling sensi-tizes these melanoma cells to lupeol (Figure 6B).
Effect of lupeol on Wnt/b-catenin signaling in 293T cells transfectedwith stable b-cateninIn our final set of experiments, we induced Wnt/b-catenin signaling in293T cells and tested the effect of lupeol on b-catenin transcriptionalactivity and expression of Wnt target genes. The rationale for using293T cells is that they lack constitutively active Wnt/b-catenin sig-naling. Activating the pathway using stable b-catenin, upon lupeoltreatment, we observed a dose-dependent decrease in b-catenin tran-scriptional activity (TOP-Flash luciferase assay) (Figure 6C). In thesecells, a dose-dependent decrease in expression of Wnt target genes,viz c-myc and cyclin D1 was also observed (Figure 6C).
Discussion
The most significant and novel finding in this study is the demonstra-tion that a non-toxic dietary triterpene lupeol specifically targets mel-anoma cells that harbor constitutive activation of Wnt/b-catenin
Fig. 4. Effect of lupeol administration on the growth of melanoma implantedtumors in athymic nude mice. (A and B) Tumor xenograft studies. (A) Mel928 and (B) Mel 1011 cells were subcutaneously implanted and the micewere treated with lupeol as described in Materials and Methods. The growthwas measured in terms of average volume of tumors as function of time.Results are expressed as tumor volume (mm3) ± SE of eight mice; �P, 0.05versus control mice. Corn oil, lupeol (therapy) and lupeol(prevention).
R.S.Tarapore et al.
1848
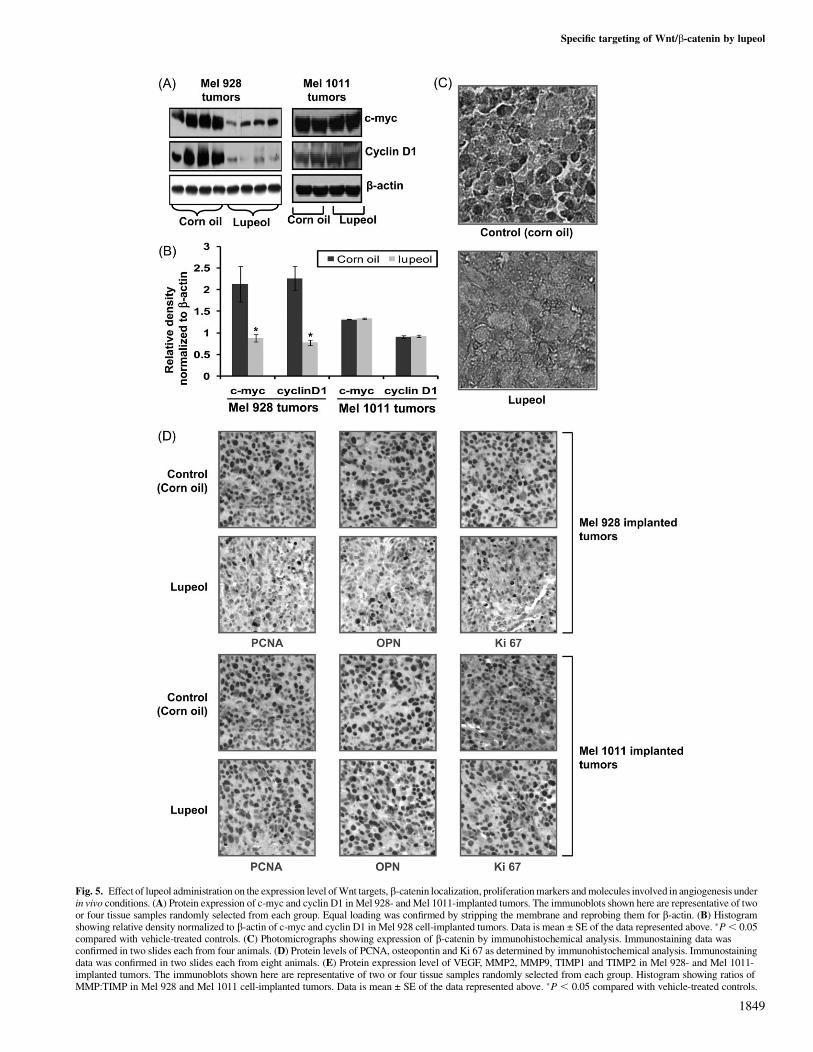
Fig. 5. Effect of lupeol administration on the expression level of Wnt targets, b-catenin localization, proliferation markers and molecules involved in angiogenesis underin vivo conditions. (A) Protein expression of c-myc and cyclin D1 in Mel 928- and Mel 1011-implanted tumors. The immunoblots shown here are representative of twoor four tissue samples randomly selected from each group. Equal loading was confirmed by stripping the membrane and reprobing them for b-actin. (B) Histogramshowing relative density normalized to b-actin of c-myc and cyclin D1 in Mel 928 cell-implanted tumors. Data is mean ± SE of the data represented above. �P, 0.05compared with vehicle-treated controls. (C) Photomicrographs showing expression of b-catenin by immunohistochemical analysis. Immunostaining data wasconfirmed in two slides each from four animals. (D) Protein levels of PCNA, osteopontin and Ki 67 as determined by immunohistochemical analysis. Immunostainingdata was confirmed in two slides each from eight animals. (E) Protein expression level of VEGF, MMP2, MMP9, TIMP1 and TIMP2 in Mel 928- and Mel 1011-implanted tumors. The immunoblots shown here are representative of two or four tissue samples randomly selected from each group. Histogram showing ratios ofMMP:TIMP in Mel 928 and Mel 1011 cell-implanted tumors. Data is mean ± SE of the data represented above. �P , 0.05 compared with vehicle-treated controls.
Specific targeting of Wnt/b-catenin by lupeol
1849
signaling. The prognosis of melanoma patients with distinct metasta-sis remains poor and in general is not significantly influenced by anytreatment intervention because metastatic melanomas are inevitablyresistant to conventional therapeutic agents. Several natural agents,including lupeol, with high anticancer efficacy and relatively minimaltoxicity to normal tissues are suggested as possible candidates forchemoprevention of melanoma (3,24,33–35). In vitro as well asin vivo studies from our laboratory and elsewhere have suggestedthat lupeol could impart chemopreventive as well as chemotherapeu-tic effects against various cancer types (13,16,24,36). In an earlierstudy (24), we demonstrated the anticancer efficacy of lupeol againstmetastatic melanoma 451 Lu cells. Since metastatic potential of mel-anomas are associated with elevated expression of Wnt/b-cateninsignaling pathway, we wanted to test the role of lupeol in cells thatexpress differential expression of Wnt/b-catenin pathway. In this cur-rent study, we provide compelling evidence that lupeol is significantlymore effective against melanomas that exhibit constitutive activationof Wnt/b-catenin signaling as compared with melanomas that lackconstitutively active Wnt/b-catenin signaling. We employed Mel1241 cell line that carries a stabilizing mutation of b-catenin andMel 928 that expresses wild-type b-catenin, but no detectable APC.Mel 1011, containing both intact b-catenin and APC are used ascontrol. Mel 928 and Mel 1241 cells are characterized by nuclearlocalization of b-catenin and constitutive activation of b-catenin/Tcf-dependent transcription (21,22).
Our data suggests that lupeol treatment causes a dose-dependentdecrease in viability and an increase in apoptosis (activation of cas-pases and expression of annexin V) in melanoma cells that exhibitconstitutive activation of Wnt/b-catenin signaling. Caspases receivethe upstream death signals and are the final executioners of apoptosis(37). The inability of melanomas to undergo apoptosis in responseto chemotherapy is a selective advantage for tumor progression,
metastasis and resistance to therapy. Our data is significant since inrecent years, the emphasis is on natural diet-based agents capable ofselective/preferred elimination of cancer cells by inhibiting cell cycleprogression and/or modulating apoptosis while sparing normal cells.In addition, our study provides evidence that lupeol has the potentialto kill advanced stage malignant melanoma cells that exhibit consti-tutive activation of Wnt/b-catenin signaling. It is important to mentionthat this subset of melanoma has poor prognosis (21).
Through the binding of a complex of b-catenin and TCF to spe-cific promoter elements, b-catenin acts as an important transcrip-tional regulator for numerous genes (38). Lupeol treatment ofmelanoma cells that exhibit constitutively active Wnt signalingresults in decreased b-catenin transcriptional activity and lowerexpression of known Wnt target genes such as coding regiondeterminant-binding protein (21,39), microphthalmia-associatedtranscription factor (40–44) and cyclin D1 (45). A decrease inb-catenin in the nucleus with a corresponding increase in the cyto-plasm suggests that lupeol has the potential to block the transloca-tion of b-catenin from the cytoplasm to the nucleus. To establish thein vivo relevance of our in vitro data, we implanted melanoma cells(Mel 928 and Mel 1011) in athymic nude mice. We observed that theefficacy of lupeol is significantly higher in melanomas that exhibitconstitutively active Wnt/b-catenin signaling.
The importance of cyclin D1 and c-myc (known Wnt target genes)(27,45) as therapeutic targets for melanoma emanates from studiesdemonstrating that inhibiting the expression of cyclin D1 andc-myc causes apoptosis and shrinkage of tumors in mice (27,45–48). It is noteworthy that lupeol treatment significantly decreasesthe expression level of c-myc and cyclin D1 in Mel 928-originatedtumors. This data provides significant evidence that lupeol modulatesWnt signaling and lowers the expression of Wnt targets both in vitroand in vivo.
Fig. 5. continued.
R.S.Tarapore et al.
1850
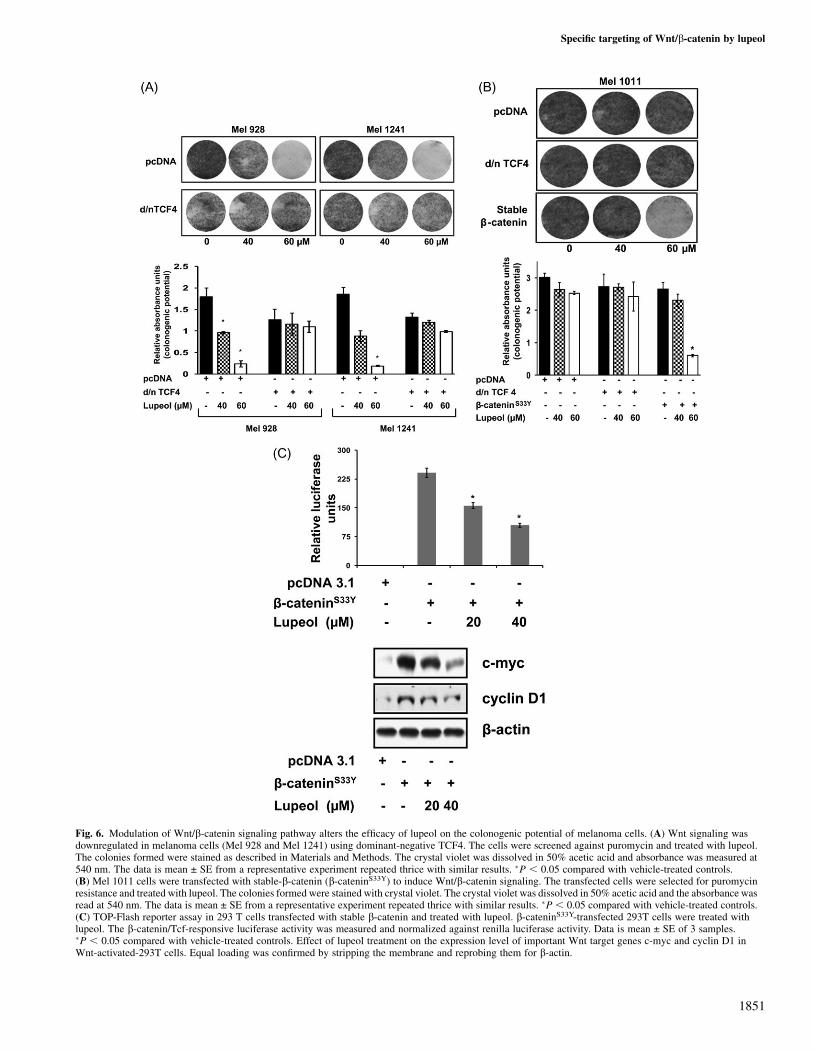
Fig. 6. Modulation of Wnt/b-catenin signaling pathway alters the efficacy of lupeol on the colonogenic potential of melanoma cells. (A) Wnt signaling wasdownregulated in melanoma cells (Mel 928 and Mel 1241) using dominant-negative TCF4. The cells were screened against puromycin and treated with lupeol.The colonies formed were stained as described in Materials and Methods. The crystal violet was dissolved in 50% acetic acid and absorbance was measured at540 nm. The data is mean ± SE from a representative experiment repeated thrice with similar results. �P , 0.05 compared with vehicle-treated controls.(B) Mel 1011 cells were transfected with stable-b-catenin (b-cateninS33Y) to induce Wnt/b-catenin signaling. The transfected cells were selected for puromycinresistance and treated with lupeol. The colonies formed were stained with crystal violet. The crystal violet was dissolved in 50% acetic acid and the absorbance wasread at 540 nm. The data is mean ± SE from a representative experiment repeated thrice with similar results. �P , 0.05 compared with vehicle-treated controls.(C) TOP-Flash reporter assay in 293 T cells transfected with stable b-catenin and treated with lupeol. b-cateninS33Y-transfected 293T cells were treated withlupeol. The b-catenin/Tcf-responsive luciferase activity was measured and normalized against renilla luciferase activity. Data is mean ± SE of 3 samples.�P , 0.05 compared with vehicle-treated controls. Effect of lupeol treatment on the expression level of important Wnt target genes c-myc and cyclin D1 inWnt-activated-293T cells. Equal loading was confirmed by stripping the membrane and reprobing them for b-actin.
Specific targeting of Wnt/b-catenin by lupeol
1851

A discernable decrease in expression of proliferation markers suchas PCNA, osteopontin and Ki-67 was also observed in lupeol-treatedMel 928-implanted tumors suggesting descreased proliferative activ-ity upon lupeol treatment. Recently, there has been an increased in-terest in understanding the angiogenesis machinery that causeschemoresistance in melanoma cells (49,50). VEGF is an importantsignaling protein and plays an essential role in angiogenesis (49). Therole of MMPs in invasion and metastasis in melanomas and othermalignancies has been extensively studied (51,52). MMP activity ismodulated by a family of naturally occurring TIMPs. In Mel 928-implanted tumors, we observed a 13-fold decrease in expression lev-els of VEGF in lupeol-treated animals versus those treated with cornoil only. A 10-fold decrease in ratios of MMP9/TIMP1 and MMP2/TIMP2 was observed in Mel 928-implanted lupeol-treated tumors.Thus, our data suggest that the effect of lupeol on key regulators ofangiogenesis (MMPs and TIMPs) is very significant.
Furthermore, we wanted to test the efficacy of lupeol when Wnt/b-catenin signaling is silenced. Colony formation assay reveals thatupon silencing Wnt/b-catenin signaling, the efficacy of lupeol waslost suggesting that Wnt signaling is the key player for lupeol-inducedeffects. On the other hand, induction of Wnt signaling in Mel 1011cells results in reduced colonogenic potential upon lupeol treatment,further verifying the Wnt-specific role of lupeol. In our conclusiveexperiments, we induced Wnt/b-catenin signaling in non-tumorigenic293T cells. We have demonstrated that lupeol treatment resulted ina dose-dependent decrease in b-catenin transcriptional activity andprotein expression of Wnt target genes (27,45).
In summary, we have demonstrated the effect of lupeol againstWnt/b-catenin signaling network resulting in decrease in prolifera-tion, viability and growth of melanoma cells and tumors in mice.Taken together, our present findings demonstrate the anticancer effi-cacy of lupeol against melanomas that exhibit constitutively activeWnt/b-catenin signaling. These observations coupled with our earlierpublished work showing lack of effect of lupeol on normal humanmelanocytes (24) suggest further in depth studies to understand themechanism behind the modulation of Wnt/b-catenin signaling bylupeol. The positive outcomes of such studies could form a strongbasis for the development of lupeol as a novel agent for malignanciesthat exhibit constitutive Wnt signaling that occurs in a third of mel-anomas that have poor prognosis.
Funding
National Institutes of Health (R01-120451).
Acknowledgements
We would like to acknowledge Drs K.Kinzler and B.Vogelstein for theirgenerous gifts of d/n TCF4, b-cateninS33Y, pcDNA and TOP-Flash reporterplasmids.
Conflict of Interest Statement: None declared.
References
1. Jemal,A. et al. (2009) Cancer statistics, 2009. CA Cancer J. Clin., 59,225–249.
2.Miller,A.J. et al. (2006) Melanoma. N. Engl. J. Med., 355, 51–65.3.Khan,N. et al. (2008) Cancer chemoprevention through dietary antioxi-
dants: progress and promise. Antioxid. Redox Signal., 10, 475–510.4.Beveridge,T.H. et al. (2002) Phytosterol content in American ginseng seed
oil. J. Agric. Food Chem., 50, 744–750.5.Fernandez,M.A. et al. (2001) New insights into the mechanism of action
of the anti-inflammatory triterpene lupeol. J. Pharm. Pharmacol., 53,1533–1539.
6. Imam,S. et al. (2007) Two triterpenes lupanone and lupeol isolated andidentified from Tamarindus indica linn. Pak. J. Pharm. Sci., 20, 125–127.
7.Saleem,M. (2009) Lupeol, a novel anti-inflammatory and anti-cancerdietary triterpene. Cancer Lett., 285, 109–115.
8.You,Y.J. et al. (2003) Antiangiogenic activity of lupeol from Bombax ceiba.Phytother. Res., 17, 341–344.
9.Fernandez,A. et al. (2001) Anti-inflammatory effect of Pimenta racemosavar. ozua and isolation of the triterpene lupeol. Farmaco, 56, 335–338.
10.Lima,L.M. et al. (2007) Anti-inflammatory and analgesic activities of theethanolic extracts from Zanthoxylum riedelianum (Rutaceae) leaves andstem bark. J. Pharm. Pharmacol., 59, 1151–1158.
11.Vasconcelos,J.F. et al. (2008) The triterpenoid lupeol attenuates allergicairway inflammation in a murine model. Int. Immunopharmacol., 8,1216–1221.
12.Latha,R.M. et al. (2001) A novel derivative pentacyclic triterpene andomega 3 fatty acid. Prostaglandins Leukot. Essent. Fatty Acids, 64, 81–85.
13.Sultana,S. et al. (2003) Lupeol, a triterpene, prevents free radical mediatedmacromolecular damage and alleviates benzoyl peroxide induced biochem-ical alterations in murine skin. Indian J. Exp. Biol., 41, 827–831.
14.Lira Wde,M. et al. (2008) Modulatory effect of Byrsonima basilobaextracts on the mutagenicity of certain direct and indirect-acting mutagensin Salmonella typhimurium assays. J. Med. Food, 11, 111–119.
15.Cmoch,P. et al. (2008) Synthesis of lupane-type saponins bearing mannosyland 3,6-branched trimannosyl residues and their evaluation as anticanceragents. Carbohydr. Res., 343, 995–1003.
16.Saleem,M. et al. (2004) Lupeol modulates NF-kappaB and PI3K/Akt path-ways and inhibits skin cancer in CD-1 mice. Oncogene, 23, 5203–5214.
17.Chin,L. et al. (2006) Malignant melanoma: genetics and therapeutics in thegenomic era. Genes Dev., 20, 2149–2182.
18.Giles,R.H. et al. (2003) Caught up in a Wnt storm: wnt signaling in cancer.Biochim. Biophys. Acta, 1653, 1–24.
19.Weeraratna,A.T. (2005) A Wnt-er wonderland—the complexity of Wntsignaling in melanoma. Cancer Metastasis Rev., 24, 237–250.
20.Moon,R.T. (2005) Wnt/beta-catenin pathway. Sci. STKE, 2005, cm1.21.Elcheva,I. et al. (2008) Overexpression of mRNA-binding protein CRD-BP
in malignant melanomas. Oncogene, 27, 5069–5074.22.Rubinfeld,B. et al. (1997) Stabilization of beta-catenin by genetic defects in
melanoma cell lines. Science, 275, 1790–1792.23.Rimm,D.L. et al. (1999) Frequent nuclear/cytoplasmic localization of
b-catenin without exon 3 mutations in malignant melanoma. Am. J. Pathol.,154, 325–329.
24.Saleem,M. et al. (2008) Lupeol inhibits growth of highly aggressive humanmetastatic melanoma cells in vitro and in vivo by inducing apoptosis. Clin.Cancer Res., 14, 2119–2127.
25.Biechele,T.L. et al. (2008) Assaying-catenin/TCF transcription with b-catenin/TCF transcription-based reporter constructs. Methods Mol. Biol., 468, 99–110.
26.Korinek,V. et al. (1997) Constitutive transcriptional activation by a beta-catenin-Tcf complex in APC-/- colon carcinoma. Science, 275, 1784–1787.
27.He,T.C. et al. (1998) Identification of c-MYC as a target of the APCpathway. Science, 281, 1509–1512.
28.Morin,P.J. et al. (1997) Activation of beta-catenin-Tcf signaling in coloncancer by mutations in beta-catenin or APC. Science, 275, 1787–1790.
29.Siddiqui,I.A. et al. (2008) Green tea polyphenol EGCG sensitizes humanprostate carcinoma LNCaP cells to TRAIL-mediated apoptosis and syner-gistically inhibits biomarkers associated with angiogenesis and metastasis.Oncogene, 27, 2055–2063.
30.Siddiqui,I.A. et al. (2006) Inhibition of CWR22Rnu1 tumor growth andPSA secretion in athymic nude mice by green and black teas. Carcinogen-esis, 27, 833–839.
31.Adhami,V.M. et al. (2004) Oral consumption of green tea polyphenolsinhibits insulin-like growth factor-I-induced signaling in an autochthonousmouse model of prostate cancer. Cancer Res., 64, 8715–8722.
32.Hu,J. et al. (2009) Blockade of Wnt signaling inhibits angiogenesisand tumor growth in hepatocellular carcinoma. Cancer Res., 69,6951–6959.
33.Afaq,F. et al. (2002) Botanical antioxidants for chemoprevention of photo-carcinogenesis. Front. Biosci., 7, d784–d792.
34.Becker,J.C. et al. (2006) Molecularly targeted therapy for melanoma:current reality and future options. Cancer, 107, 2317–2327.
35.Nichenametla,S.N. et al. (2006) A review of the effects and mechanisms ofpolyphenolics in cancer. Crit. Rev. Food Sci. Nutr., 46, 161–183.
36.Nigam,N. et al. (2007) Preventive effects of lupeol on DMBA inducedDNA alkylation damage in mouse skin. Food Chem. Toxicol., 45,2331–2335.
37.Philchenkov,A. (2004) Caspases: potential targets for regulating cell death.J. Cell. Mol. Med., 8, 432–444.
38.Zhang,Z. et al. (2009) Secreted frizzled related protein 2 protects cells fromapoptosis by blocking the effect of canonical Wnt3a. J. Mol. Cell. Cardiol.,46, 370–377.
R.S.Tarapore et al.
1852

39.Noubissi,F.K. et al. (2006) CRD-BP mediates stabilization of betaTrCP1 and
c-myc mRNA in response to beta-catenin signalling. Nature, 441, 898–901.40.Dorsky,R.I. et al. (2000) Direct regulation of nacre, a zebrafish MITF
homolog required for pigment cell formation, by the Wnt pathway. Genes
Dev., 14, 158–162.41.Saito,H. et al. (2002) Melanocyte-specific microphthalmia-associated tran-
scription factor isoform activates its own gene promoter through physical
interaction with lymphoid-enhancing factor 1. J. Biol. Chem., 277,
28787–28794.42.Saito,H. et al. (2003) Microphthalmia-associated transcription factor in the
Wnt signaling pathway. Pigment Cell Res., 16, 261–265.43.Yasumoto,K. et al. (2002) Microphthalmia-associated transcription factor
interacts with LEF-1, a mediator of Wnt signaling. EMBO J., 21, 2703–2714.44.Vachtenheim,J. et al. (2004) Microphthalmia transcription factor: a specific
marker for malignant melanoma. Prague Med. Rep., 105, 318–324.45.Shtutman,M. et al. (1999) The cyclin D1 gene is a target of the beta-
catenin/LEF-1 pathway. Proc. Natl Acad. Sci. USA, 96, 5522–5527.
46.Sauter,E.R. et al. (2002) p53 alone or in combination with antisense cyclinD1 induces apoptosis and reduces tumor size in human melanoma. CancerGene Ther., 9, 807–812.
47.Florenes,V.A. et al. (2000) Levels of cyclin D1 and D3 in malignant mel-anoma: deregulated cyclin D3 expression is associated with poor clinicaloutcome in superficial melanoma. Clin. Cancer Res., 6, 3614–3620.
48.Sauter,E.R. et al. (2002) Cyclin D1 is a candidate oncogene in cutaneousmelanoma. Cancer Res., 62, 3200–3206.
49.Marneros,A.G. (2009) Tumor angiogenesis in melanoma. Hematol. Oncol.Clin. North Am., 23, 431–446., vii–viii.
50.Fecher,L.A. et al. (2009) Drug targeting of oncogenic pathways in mela-noma. Hematol. Oncol. Clin. North Am., 23, 599–618., x.
51.Lopez-Otin,C. et al. (2009) Protective roles of matrix metalloproteinases:from mouse models to human cancer. Cell Cycle, 8, 3657–3662.
52.Hofmann,U.B. et al. (2005) Role of matrix metalloproteinases in melanomacell invasion. Biochimie, 87, 307–314.
Received June 29, 2010; revised August 4, 2010; accepted August 8, 2010
Specific targeting of Wnt/b-catenin by lupeol
1853
Copyright © 2022 FDOKUMEN




















