
Apc bridges Wnt/β-catenin and BMP signaling during osteoblast differentiation of KS483 cells
12
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
Transcript of Apc bridges Wnt/β-catenin and BMP signaling during osteoblast differentiation of KS483 cells

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Research Article
Apc bridges Wnt/β-catenin and BMP signaling during osteoblastdifferentiation of KS483 cells☆
Razvan L. Micleaa,1, Geertje van der Horstb, Els C. Robanus-Maandagc, Clemens W.G.M. Löwikd,Wilma Oostdijka, Jan M. Wita, Marcel Karperiene,⁎
aDepartment of Pediatrics, Leiden University Medical Centre (LUMC), Leiden, the NetherlandsbDepartment of Urology, LUMC, Leiden, the NetherlandscDepartment of Human Genetics, LUMC, Leiden, the NetherlandsdDepartment of Endocrinology and Metabolic Diseases, LUMC, Leiden, the NetherlandseMIRA Institute for Biomedical Technology and Technical Medicine, Department of Tissue Regeneration, University of Twente,Zuidhorst Room ZH 144, Drienerlolaan 5, 7522 NB Enschede, the Netherlands
A R T I C L E I N F O R M A T I O N A B S T R A C T
Article Chronology:
Received 19 October 2010Revised version received27 February 2011Accepted 7 March 2011Available online 11 March 2011
The canonical Wnt signaling pathway influences the differentiation of mesenchymal cell lineagesin a quantitative and qualitative fashion depending on the dose of β-catenin signaling.Adenomatous polyposis coli (Apc) is the critical intracellular regulator of β-catenin turnover.To better understand the molecular mechanisms underlying the role of Apc in regulating thedifferentiation capacity of skeletal progenitor cells, we have knocked down Apc in the murinemesenchymal stem cell-like KS483 cells by stable expression of Apc-specific small interfering RNA.In routine culture, KSFrt-Apcsi cells displayed a mesenchymal-like spindle shape morphology,exhibited markedly decreased proliferation and increased apoptosis. Apc knockdown resulted inupregulation of the Wnt/β-catenin and the BMP/Smad signaling pathways, but osteogenic
differentiation was completely inhibited. This effect could be rescued by adding highconcentrations of BMP-7 to the differentiation medium. Furthermore, KSFrt-Apcsi cells showedno potential to differentiate into chondrocytes or adipocytes.These results demonstrate that Apc is essential for the proliferation, survival and differentiation ofKS483 cells. Apc knockdown blocks the osteogenic differentiation of skeletal progenitor cells, aprocess that can be overruled by high BMP signaling.
© 2011 Elsevier Inc. All rights reserved.
Keywords:
Apc
β-CateninWntOsteoblastDifferentiation
Introduction
During endochondral bone formation, skeletal progenitor cells(SPC) arise from mesenchymal cells, transit several differentiation
steps to ultimately develop into bone or cartilage [24]. Theircommitment to one of the two lineages requires a very intricateand tightly controlled crosstalk between transcription factors,cytokines, and growth factors [25]. However, the precisemolecular
E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 1 4 1 1 – 1 4 2 1
☆ Funding resources: This work was financially supported by an unrestricted educational grant from IPSEN FARMACEUTICA BV.⁎ Corresponding author. Fax: +31 53 489 2150.
E-mail addresses: [email protected] (R.L. Miclea), [email protected] (G. van der Horst), [email protected](E.C. Robanus-Maandag), [email protected] (C.W.G.M. Löwik), [email protected] (W. Oostdijk), [email protected] (J.M. Wit),[email protected] (M. Karperien).1 Present address: Department of Radiology, LUMC, Leiden, the Netherlands.
0014-4827/$ – see front matter © 2011 Elsevier Inc. All rights reserved.doi:10.1016/j.yexcr.2011.03.007
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /yexc r

Author's personal copy
interactions that control their lineage commitment and differen-tiation to mature skeletal cells are not fully understood.
Increasing evidence suggests an important role of the canonicalWnt signaling pathway in the regulation of lineage commitment ofSPC [31]. In this pathway, in the absence of the Wnt signal,cytoplasmic β-catenin is degraded in the proteasome upon itsphosphorylation at specific Ser–Thr residues by a destructioncomplex consisting of Axin, adenomatous polyposis coli (APC),glycogen synthase kinase 3β (GSK3β) and casein-kinase 1α (CK1α).Wnt growth factors bind to the receptor Frizzled and low-densitylipoprotein receptor-related protein-5 or 6 (LRP-5/6) to inactivatethis destruction complex, via Disheveled (DVL). This leads toaccumulation of unphosphorylated β-catenin and subsequenttranslocation into the nucleus. Together with members of the Tcell factor/lymphoid enhancer factor (TCF/LEF) family, nuclear β-catenin stimulates transcription of Wnt target genes [8]. Upregula-tion of β-catenin in bi-potential SPC leads to osteoblast formation,whereas down-regulation favors their commitment to the chondro-genic lineage [17].
Another signaling cascade equally important in the differen-tiation of SPC is the bone morphogenetic protein (BMP)/Smadpathway which promotes both osteo- and chondrogenesis [4]. Inthis pathway, BMPs bind to and activate BMP type I or II receptorsthereby initiating phosphorylation of receptor-regulated Smads(R-Smads) 1, 5, and 8. Phosphorylated active R-Smads formheteromeric complexes with common-partner Smad4 that trans-locate to the nucleus to regulate the transcription of target genesin cooperation with other transcription factors [47].
Due to the great importance of the Wnt/β-catenin and BMPpathway during both osteogenic and chondrogenic differentiationof SPC, the interaction between these two powerful regulatorypathways has received much attention. For example, it hasbeen shown that BMP-2 upregulates expression of Wnt-3a andβ-catenin and that β-catenin is crucial for BMP-induced new boneformation [7,11]. However, the BMP signal can also antagonizeWnt in SPC by promoting an interaction between Smad1 and Dvlthat restricts β-catenin accumulation [32]. These and other datasuggest that Wnt and BMP signaling can alternatively synergize orantagonize one another in differentiation of SPC [6,56].
We have recently shown that, by downregulating the canonicalWnt/β-catenin signal, Apc is essential for the commitment of SPC tothe chondrogenic andosteogenic lineage [36].Moreover, distinctApcmutations unevenly affect the differentiation potential of mouseembryonic stem cells (ES): whereas Apc alleles entirely deficient inβ-catenin downregulation domains block the differentiation poten-tial of ES, more hypomorphic alleles which are still able to partiallydownregulateβ-catenin impair thedifferentiation of ES only to sometissues, e.g., bone and cartilage [26]. In cells carrying a hypomorphicApcmutation, the levels ofβ-catenin are upregulated onlywhen Apcactivity levels are below 2% of normal [26].
To further unravel the subtle role of Apc in the regulation of SPCdifferentiation, we have knocked down the mouse Apc gene usingRNA interference (RNAi) in the murine mesenchymal stem cell-likeKS483 cell line. This cell line shows SPC-like characteristics, since itcan form osteoblasts, chondrocytes, and adipocytes [52]. Our datasuggest that Apc knockdown in KS483 cells leads to upregulation notonly of the Wnt/β-catenin, but also of the BMP signaling pathway,further sustaining the interaction of these biological routes duringvarious steps of SPC differentiation. Low levels of Apc inhibitedosteoblast, chondrocyte and adipocyte differentiation. Interestingly,
the inhibitory effects of Apc knockdown on osteogenic differentia-tion could be rescued by high levels of BMP-7.
Materials and methods
Generation of the KS483 cell lines with stable expression ofApcsi constructs
To obtain the KSFrt-Apcsi stable cell line, the shRNA plasmid p5H1-Apcsi, designed to express shRNA targeting the mouse Apc gene,was constructed as described previously [52]. To obtain thecontrol, KSFrt-mtApcsi stable cell line, the shRNA plasmid p5H1-mtApcsi was generated by introducing mismatches at position 7and 15 of the Apc target sequence. To demonstrate the biologicalreproducibility of our results, the KSFrt-Apc*si and the KSFrt-mtApc*si cell lines were also generated using the p5H1-Apc*si andthe p5H1-mtApc*si plasmid (carryingmismatches at position 5 and16 of the Apc*si construct), respectively. The target sequences usedto specifically silence Apc (Apcsi, Apc*si) and their correspondingmutant (control) sequences (mtApcsi, mtApc*si) are shown inFig. 1A. Stable transfections of the 4 C3 Frt clone of the KS483murine host cell line were performed as previously described [52].In this clone, a unique Flp recombinase target (FRT) sequence isintroduced in the genome. This site is subsequently used fortargeted insertion of the short hair pin vector using Flp-mediatedhomologous recombination [52].
Cell culture
KS483 cells were cultured as described previously [35]. For theKSFrt 4C3 host cell line the medium was supplemented withblasticidin S HCl (2 μg/ml; Invitrogen). All stably transfected celllines were cultured in the presence of hygromycin B (100 μg/ml;Invitrogen).
Immunofluorescence
Immunofluorescence for Apc and β-catenin was performed asdescribed previously with minor modifications [35]. In brief,cells were seeded on glass slides (BD Falcon) and either leftuntreated or treated with Wnt3a (30 ng/ml; R&D Systems) for3 h. The primary antibodies were rabbit polyclonal anti-Apc(1:500 in NETGEL; Abcam) and rabbit polyclonal anti-β-catenin(1:500 in NETGEL; Abcam). The second antibody used was goatanti-rabbit FITC-conjugate (1:250; Sigma). The F-actin cytoskel-eton was counterstained using Phalloidin-TRITC (0.33 mg/ml;Sigma). Cells were imaged using the 63× objective of an invertedLeica SP2 confocal microscope.
Western blot
Approximately 2×107 cells were either cultured in the controlconditions for 24 h or with 30 ng/ml Wnt3a, rinsed twice with PBSand lysed for 5 min on ice in 400 μl of cell lysis buffer (Cell Signaling)and a cocktail of protease inhibitors (Roche). For detection of Apc andβ-catenin proteins byWestern blot, whole cell lysates were loaded ona 4–20% linear gradient Tris–HCl Gel (BIO-RAD) and transferred ontoPVDF membranes (Millipore) by 1 h electroblotting at 300 mAconstant current at RT inblotting buffer (BIO-RAD). Following transfer,
1412 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 1 4 1 1 – 1 4 2 1

Author's personal copy
themembranes were blockedwith 5% nonfat dry milk in TPBS (0.05%Tween 20 in PBS) for 1 h. Incubation with primary antibodies wasperformed overnight at 4 °C using rabbit polyclonal anti-Apc (1:100;Abcam) or mouse monoclonal anti-β-catenin (1:2000; Epitomics)antibodies. Blots were washed three times with PBS and incubatedwith horseradish peroxidase-conjugated secondary antibodies for 1 hat room temperature. Theperoxidasewas visualized andquantified byenhanced chemiluminescence using the Molecular Imager Gel DocXR+System (BIO-RAD).
Real-time quantitative PCR
Real-time quantitative PCR was performed using QuantiTect real-time PCR primers (Qiagen) for the detection of the mouse Apc,Ctnnb1, Axin2, Smad1, Smad3, Smad4, and Bmp7 genes andanalyzed as described previously [35].
Proliferation assay
For proliferation assays, the CellTiter 96® AQueous Non-Radioactive Cell Proliferation Assay (Promega) was used. Cellswere seeded at a density of 2500 cells/cm2. After 24, 48, 72 and96 h, 20 μl of MTS [3-(4,5-dimethylthiazol-2-yl)-5-(3-carbox-ymethoxyphenyl)-2-(4-sulfophenyl)-2 H-tetrazolium)] wasadded to the medium and the mitochondrial activity wasmeasured at 490 nm after 2 h incubation at 37 °C.
Apoptosis assay
For detection of apoptotic cells, Annexin V staining was performedusing Annexin V-FITC (1:250; Bender Med Systems), whichspecifically binds phosphatidyl serine (PS) residues on the cellmembrane and propidium iodide (PI; Bender Med Systems) at
Fig. 1 – Morphological and structural characterization of the KSFrt-Apcsi cell line. (A) The Apc target sequence of Apcsi, mtApcsi,Apc*si, and mtApc*si RNAi vectors driven by the hRNAseH1 promoter used to obtain the KSFrt-Apcsi, KSFrt-mtApcsi, KSFrt-Apc*si, andKSFrt-mtApc*si cell line, respectively. (B, C) QuantifiedWestern blot analysis, performed on total protein lysates, demonstrated thatKSFrt-Apcsi cells (black bars) express less Apc and β-catenin protein in comparison to control KSFrt-mtApcsi cells (white bars).(D) Bright field (upper row), IF for Apc (green; middle row) and for β-catenin (green; lower row) pictures of the controlKSFrt-mtApcsi and of the KSFrt-Apcsi cells coupledwith Phalloidin staining for the F-actin cytoskeleton (red). Note the spindle shapemesenchymal-likemorphology of the KSFrt-Apcsi cells, expressing less Apc andmostly nuclear β-catenin.Wnt3a (30 ng/ml) inducedneither morphological nor structural changes in the KSFrt-Apcsi cells. Bar represents 50 μm (bright field pictures) or20 μm (IF pictures). *p<0.05.
1413E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 1 4 1 1 – 1 4 2 1

Author's personal copy
1 μg/ml which binds to DNA once the cell membrane has becomepermeable. Cells were analyzed by flow cytometry (FACS-Calibur,Becton Dickinson) using the CellQuest program (FACS-Calibur,Becton Dickinson).
Luciferase transient transfection assays
The KSFrt-Apcsi and KSFrt-mtApcsi stable cells were seeded at adensity of 19,000 cells/cm2 and 9500 cells/cm2, respectively, in 24-well plates, and transiently transfected with 2 μg of the reporterconstruct (pGL3-(BRE)2-Luc, BAT-Luc or pSAR-MT-APC) usingFugene HD transfection reagent (Roche), according to themanufacturer's protocol. To correct for transfection efficiency,25 ng of Renilla luciferase (pGL4-CAAGS; Promega) was co-transfected. Twenty-four hours after transfection, transfectedcells were either left non-stimulated or stimulated for anadditional 24 h. Luciferase assays were performed as describedpreviously [35].
Differentiation assays
To induceosteogenicdifferentiation, theKSFrt-Apcsi andKSFrt-mtApcsistable cells were seeded at a density of 24,000 cells/cm2 and12,000 cells/cm2, respectively, and cultured in thepresenceor absenceof BMP-7 at the concentrations indicated. The medium was changedevery 3–4 days. At confluence (from Day 4 of culture onward),ascorbic acid (50 μg/ml; Merck) and, when nodules appeared (fromDay 11 of culture onward), β-glycerol-phosphate (5 mM; Sigma)were added to the culture medium. Analysis of the AlkalinePhosphatase activity (at Day 11) and the degree of mineralization(at Day 21) was performed as previously described [50].
To induce chondrogenic differentiation, 300,000 cells werepelleted by centrifugation in a round-bottom-well of a 96-well-plate (Corning) and cultured in 250 μl high-glucose DMEM (Gibco),supplemented with 100 U/ml Pen/Strep, 50 μg/ml ascorbic acid(Merck), 40 μg/ml proline (Sigma), 1 mM Pyruvate, 1:100 ITS+Premix (BD Biosciences). During the first 2 weeks of culture,mediumwas further enriched with 10 ng/ml TGFβ3 (R&D Systems)and 10−7 M dexamethasone (Sigma), while beginning with week 3,500 ng/ml BMP-6 and 5 mM β-glycerol-phosphate (Merck) wasadded to the medium. The medium was replaced every 3–4 days.After 6 weeks of culture, pellets were fixed, embedded in paraffinand sectioned. Sections were stained with Toluidine Blue orimmunostained for collagen II as previously described [36].Glycosaminoglycan quantification corrected for DNA after 2, 4 and6 weeks of culture was performed as previously described [35].
To induce adipogenic differentiation, the KSFrt-Apcsi and KSFrt-mtApcsi stable cells were seeded at a density of 24,000 cells/cm2
and 12,000 cells/cm2, respectively, and cultured in the presence of25 μM indomethacin after confluence (from Day 4 of cultureonward). After 3 weeks of culture, cells were stained with Oil RedO as described previously [50]. Quantification of adipocytes wasperformed by counting adipocytes, defined by the presence of atleast three lipid droplets per cell from nine randomly selectedfields (three fields per well) for each group.
Statistics
All values represent mean±SEM of two or three independenttriplicate experiments. Differences were examined by one way
analysis of variance (ANOVA). Results were considered significantat p<0.05.
Results
The KSFrt-Apcsi cell line is a valid model for studying the roleof Apc in SPC differentiation
To study the role of the Apc gene in regulating lineage commitmentand differentiation of SPC, we generated a cell line with decreasedApc expression by RNA interference (RNAi) using the 4C3 Frt cloneof the KS483 murine host cell line [49].
Overexpression of Apcsi but not of mtApcsi decreased wild-typeApc protein levels with approximately 50%, suggesting an efficientgene knockdown at the protein level (Fig. 1B). KSFrt-Apcsi cells alsoshowed less total β-catenin protein expression in comparison tocontrol mtApcsi cells in whole cell extracts (Fig. 1C). Nevertheless,total β-catenin levels were reduced in both cytoplasmic andnuclear cell fractions (data not shown). Treatment withWnt3a didnot affect the Apc expression, but upregulated β-catenin in bothKSFrt-Apcsi and KSFrt-mtApcsi cells.
The morphology of the KSFrt-Apcsi cells was considerablychanged into thin, elongated, spindle shape mesenchymal-likecells in contrast to control cells that maintained the polygonal,cuboidal shape of the parental 4C3 cell line (Fig. 1D, upper row).Morphologywas not influenced by treatmentwithWnt3a in neitherof the cell lines. To investigate the cellular level and distribution ofApc and β-catenin in the KSFrt-Apcsi cells, we next performedimmunofluorescence (IF) analysis coupled with Phalloidin stainingfor visualizing the F-actin cytoskeleton in non-confluent cultures. IFfor Apc confirmed the WB results, indicating overall less Apcexpression in KSFrt-Apcsi cells in comparison to control cells(Fig. 1D, middle row). Wnt3a affected neither the level of Apc norits cellular distribution in both cell lines. In control cells, β-cateninwas mainly membrane-bound and cytoplasmic, while stimulationwithWnt3a induced β-catenin nuclear translocation (Fig. 1D, lowerrow). In contrast, in the KSFrt-Apcsi cells, β-catenin was mainlypresent in the nucleus in both non- and Wnt3a-stimulatedconditions. Similar results were obtained on confluent cultures ofboth cell lines (data not shown).
Functional characterization of the KSFrt-Apcsi cell line
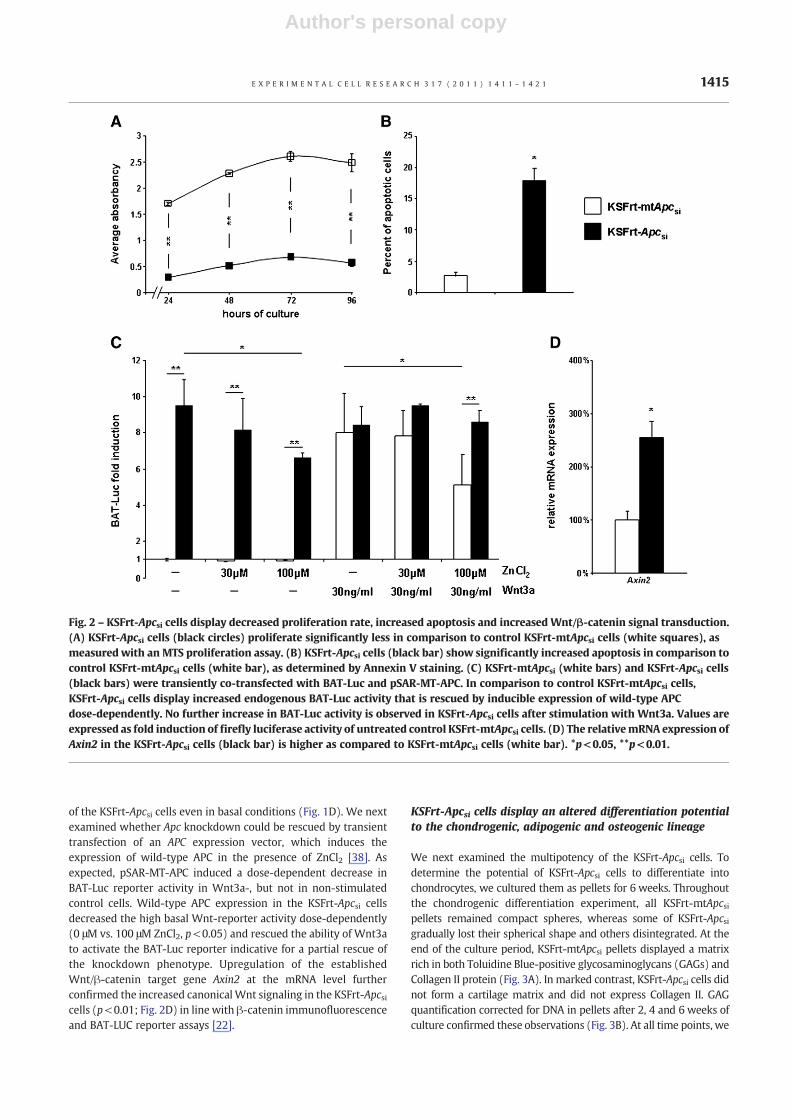
Proliferation of both KSFrt-Apcsi and KSFrt-Apc*si cells wassignificantly reduced after 24, 48, 72 and 96 h of culture incomparison to control cells, as confirmed by MTS proliferationassay (p<0.01 at all time points; Fig. 2A and data not shown). Thepercentage of apoptotic cells detected by Annexin V staining wassignificantly increased in the KSFrt-Apcsi cells as compared tocontrol cells (18.02% and 2.73%, respectively, p<0.05; Fig. 2B).
We next used the Wnt responsive BAT-Luc reporter constructto evaluate the effect of Apc knockdown on Wnt responsiveness[33]. In basal conditions, the reporter activity was significantlyincreased in the KSFrt-Apcsi cells in comparison to control cells(p<0.01; Fig. 2C), suggestive for increased endogenous canonicalWnt signaling. Remarkably, the response to Wnt3a was blunted inthe KSFrt-Apcsi cell line. This could be due to the lower totalβ-catenin levels (Fig. 1C) and relatively higher percentage of activeβ-catenin over total β-cateninwhich already resides in the nucleus
1414 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 1 4 1 1 – 1 4 2 1
Author's personal copy
of the KSFrt-Apcsi cells even in basal conditions (Fig. 1D). We nextexamined whether Apc knockdown could be rescued by transienttransfection of an APC expression vector, which induces theexpression of wild-type APC in the presence of ZnCl2 [38]. Asexpected, pSAR-MT-APC induced a dose-dependent decrease inBAT-Luc reporter activity in Wnt3a-, but not in non-stimulatedcontrol cells. Wild-type APC expression in the KSFrt-Apcsi cellsdecreased the high basal Wnt-reporter activity dose-dependently(0 μM vs. 100 μM ZnCl2, p<0.05) and rescued the ability of Wnt3ato activate the BAT-Luc reporter indicative for a partial rescue ofthe knockdown phenotype. Upregulation of the establishedWnt/β-catenin target gene Axin2 at the mRNA level furtherconfirmed the increased canonicalWnt signaling in the KSFrt-Apcsicells (p<0.01; Fig. 2D) in line with β-catenin immunofluorescenceand BAT-LUC reporter assays [22].
KSFrt-Apcsi cells display an altered differentiation potentialto the chondrogenic, adipogenic and osteogenic lineage
We next examined the multipotency of the KSFrt-Apcsi cells. Todetermine the potential of KSFrt-Apcsi cells to differentiate intochondrocytes, we cultured them as pellets for 6 weeks. Throughoutthe chondrogenic differentiation experiment, all KSFrt-mtApcsipellets remained compact spheres, whereas some of KSFrt-Apcsigradually lost their spherical shape and others disintegrated. At theend of the culture period, KSFrt-mtApcsi pellets displayed a matrixrich in both Toluidine Blue-positive glycosaminoglycans (GAGs) andCollagen II protein (Fig. 3A). In marked contrast, KSFrt-Apcsi cells didnot form a cartilage matrix and did not express Collagen II. GAGquantification corrected for DNA in pellets after 2, 4 and 6 weeks ofculture confirmed these observations (Fig. 3B). At all time points, we
Fig. 2 – KSFrt-Apcsi cells display decreased proliferation rate, increased apoptosis and increased Wnt/β-catenin signal transduction.(A) KSFrt-Apcsi cells (black circles) proliferate significantly less in comparison to control KSFrt-mtApcsi cells (white squares), asmeasured with an MTS proliferation assay. (B) KSFrt-Apcsi cells (black bar) show significantly increased apoptosis in comparison tocontrol KSFrt-mtApcsi cells (white bar), as determined by Annexin V staining. (C) KSFrt-mtApcsi (white bars) and KSFrt-Apcsi cells(black bars) were transiently co-transfected with BAT-Luc and pSAR-MT-APC. In comparison to control KSFrt-mtApcsi cells,KSFrt-Apcsi cells display increased endogenous BAT-Luc activity that is rescued by inducible expression of wild-type APCdose-dependently. No further increase in BAT-Luc activity is observed in KSFrt-Apcsi cells after stimulation with Wnt3a. Values areexpressed as fold induction of firefly luciferase activity of untreated control KSFrt-mtApcsi cells. (D) The relativemRNA expression ofAxin2 in the KSFrt-Apcsi cells (black bar) is higher as compared to KSFrt-mtApcsi cells (white bar). *p<0.05, **p<0.01.
1415E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 1 4 1 1 – 1 4 2 1

Author's personal copy
detected significantly lowerGAGcontents in theKSFrt-Apcsi pellets incomparison to controls (p<0.05 at 2 weeks, p<0.01 at 4 and6 weeks).
The adipogenic differentiation potential of the KSFrt-Apcsi cellswas investigated by performing Oil Red O staining on cells culturedfor 1, 2 and 3 weeks in adipogenic medium. After 3 weeks of culture,many of the KSFrt-mtApcsi cells differentiated into adipocytescontaining lipid droplets that positively stained with Oil Red O(Fig. 3C). In contrast, differentiation of KSFrt-Apcsi cells intoadipocytes was severely impaired. Quantification of the number ofadipocytes indicated that after 1, 2 and 3 weeks the number of OilRed O-positive cells was significantly lower in the KSFrt-Apcsi cells incomparison to controls (p<0.05 at 2 weeks, p<0.01 at 3 weeks;Fig. 3D).
To determine the osteogenic potential of KSFrt-Apcsi cells, weperformed short-term osteoblast differentiation experiments.Alkaline phosphatase (ALP) staining and its consequent quanti-fication indicated that, in comparison to control cells, bothKSFrt-Apcsi and KSFrt-Apc*si cells display a significantly de-
creased potential to differentiate into osteoblasts (Fig. 4A anddata not shown). We next tested whether the inhibition ofosteoblastogenesis in the KSFrt-Apcsi cells could be rescued by theaddition of pro-osteogenic growth factors like basic fibroblastgrowth factor (bFGF), transforming growth factor beta 3(TGF-β3), parathyroid hormone-related peptide (PTHrP), insu-lin-like growth factor 1 (IGF-1), and two members of the BMPfamily, BMP-6 and BMP-7. Of these, only BMP-7 (and, to a lesserextent, BMP-6) could rescue the Apcsi-mediated inhibition ofosteogenic differentiation (Fig. 4A and data not shown). Osteo-blast maturation of KSFrt-Apcsi cells was investigated by alizarinRed S staining after long-term cultures to depictmineralization ofthe osteoblast nodules. Similar to their controls, neither KSFrt-Apcsi nor KSFrt-Apc*si cells displayed mineralized nodules in theabsence of BMP-7 (Fig. 4B and data not shown). In contrast toKSFrt-Apcsi cells, low concentrations of BMP-7 (10 and 30 ng/ml)were sufficient to induce matrix mineralization in control cells.Interestingly, high concentrations of BMP-7 (100 ng/ml) efficientlyinduced the formation of alizarin Red S-positive nodules in the
Fig. 3 – KSFrt-Apcsi cells display neither chondrogenic nor adipogenic differentiation potential. (A) KSFrt-Apcsi cells do notdifferentiate into chondrocytes as demonstrated by the absence of Toluidine Blue staining and Collagen II immunostainingperformed after the 6-week-long chondrogenic differentiation protocol. The boxed regions in the pictures of the upper row aremagnified in the lower row. (B) Quantification of GAGs corrected for DNA validates the microscopical findings. The GAG content inthe KSFrt-mtApcsi pellets (white bars) increases time dependently, whereas it remains significantly lower in the KSFrt-Apcsi pellets(black bars) at all time points. (C) KSFrt-Apcsi cells do not differentiate into adipocytes as demonstrated by Oil Red O stainingperformed after the 3-week-long adipogenic differentiation protocol. (D) Adipocyte counting indicates a significantly lowernumber of adipocytes among the KSFrt-Apcsi cells per surface area (black bars) when compared to KSFrt-mtApcsi cells. *p<0.05,**p<0.01.
1416 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 1 4 1 1 – 1 4 2 1

Author's personal copy
Fig. 4 – KSFrt-Apcsi cells display an impaired osteogenic differentiation potential that can be counteracted by high concentrations ofBMP-7. (A) Alp activity after 11-day-long osteoblast differentiation protocol. In the absence and at low concentrations of BMP-7,KSFrt-Apcsi cells (black bars) display significantly less Alp activity in comparison to control KSFrt-mtApcsi cells (white bars). Whenstimulated with 100 ng/ml BMP-7, KSFrt-Apcsi cells show significantly increased osteogenic differentiation potential in comparisonto control KSFrt-mtApcsi cells. Values are expressed as fold induction of Alp activity corrected for DNA of untreated controlKSFrt-mtApcsi cells. (B) Representative images of Alizarin Red S-stained cultures after 21-day-long osteoblast differentiationprotocol. BMP-7 stimulates the formation of mineralized nodules dose-dependently in control KSFrt-mtApcsi cells, whereasKSFrt-Apcsi cells showmineralized nodules only in the presence of high BMP-7 concentrations. (C) Quantification of the amount ofAlizarin Red corrected for DNA. When treated with 100 ng/ml BMP-7, KSFrt-Apcsi cells (black bars) show significantly increasedmineral deposition in comparison to control condition. Values are expressed as fold induction of Alizarin Red S content correctedfor DNA of untreated control KSFrt-mtApcsi cells (white bars). *p<0.05, **p<0.01.
1417E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 1 4 1 1 – 1 4 2 1

Author's personal copy
KSFrt-Apcsi cells. No statistically significant difference was foundwhen the alizarin Red S stainingwas quantified between KSFrt-Apcsiand control cells cultured in the presence of 100 ng/ml BMP-7(Fig. 4C). However, the osteoblast nodules formed by the KSFrt-Apcsicells were bigger in comparison to those formed by control cells.
Increased BMP signaling in the KSFrt-Apcsi cells
We next assessed the level of BMP signaling in the KSFrt-Apcsi cells byperforming transient transfection assays using the BMP-responsivepGL3-(BRE)2-Luc reporter construct [27]. KSFrt-Apcsi cells displayedsignificantly increased endogenous levels of BMP signaling incomparison to control KSFrt-mtApcsi cells (p<0.01; Fig. 5A). BMP-7activated the (BRE)2-Luc reporter dose-dependently in control cells incontrast to KSFrt-Apcsi cells. In these latter cells, only a high BMP-7concentration activated the reporter compared to the controlcondition. The responsewas blunted in the KSFrt-Apcsi cells comparedto KSFrt-mtApcsi cells (0.5-fold vs. 4-fold increase, respectively).Noggin, apotent inhibitorof theBMPsignalingpathway [47],managedto decrease both the endogenous and the BMP-7-induced activity ofthe (BRE)2-Luc reporter in the KSFrt-Apcsi cells, suggestive for
autocrine stimulation of the BMP signaling pathway for example byincreased expression of BMPs. Upregulation of the BMP signalingpathway in the KSFrt-Apcsi cells was further confirmed at the mRNAlevel by quantitative RT-PCR. Smad1, Smad3, and Smad4 weresignificantly increased in the KSFrt-Apcsi cells (p<0.05; Fig. 5B).Interestingly, Bmp7 showed a 4.4-fold higher expression at themRNAlevel in the KSFrt-Apcsi cells in comparison to KSFrt-mtApcsi cells(p<0.01).
Discussion
APC is a multifunctional protein involved in cell adhesion, mitosis,apoptosis, cytoskeletal organization, microtubule assembly, cellfate determination and chromosomal stability, yet it remainsmostly investigated as the key intracellular gate-keeper of thecanonical Wnt/β-catenin signaling pathway [12,16,53]. In ourpresent study, we demonstrate that Apc is required for prolifer-ation, suppression of apoptosis and differentiation of murinemesenchymal stem cell-like KS483 cells into the osteogenic,chondrogenic and adipogenic lineage. We obtained similar results
Fig. 5 – Increased BMP signaling in the KSFrt-Apcsi cells. (A) KSFrt-mtApcsi (white bars) and KSFrt-Apcsi cells (black bars) weretransiently transfected with pGL3-(BRE)2-Luc. KSFrt-Apcsi cells display increased endogenous (BRE)2-Luc activity that is enhancedby treatment with only high concentrations of BMP-7. Noggin inhibits both the endogenous and the BMP-7-induced (BRE)2-Lucactivity in the KSFrt-Apcsi cells. Values are expressed as fold induction of firefly luciferase activity of untreated control KSFrt-mtApcsicells. (B) Significantly increased relative mRNA expression of Smad1, Smad3, Smad4, and Bmp7 in KSFrt-Apcsi cells (black bars) incomparison to KSFrt-mtApcsi cells (white bars). *p<0.05, **p<0.01.
1418 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 1 4 1 1 – 1 4 2 1

Author's personal copy
by using 2 different shRNA sequences targeting Apc, while stabletransfection of the respective control mutant shRNA plasmids(containing two nucleotide mismatches) did not alter theproliferation, survival and differentiation capacity of KS483 cells.This clearly indicates that our results were the consequence of abona-fide and specific siRNA effect lowering wild-type Apcexpression. This was further confirmed by the partial rescue ofBAT-Luc reporter activity by transient transfection of a human APCexpression vector. Interestingly, KSFrt-Apcsi cells displayed notonly high levels of the canonical Wnt/β-catenin pathway, but alsoaugmented BMP signaling, further sustaining the multifacetedinteraction between these two signaling pathways during thedifferentiation of SPC.
RNAi is a complex biological mechanism during which shRNAsact either by cleavage (“slicing”) or by translational repression oftheir target mRNA [42]. KSFrt-Apcsi cells showed decreased Apcexpression at the protein level, thereby documenting an efficientApc knockdown by RNAi [9,43]. B-catenin protein expression wasalso lower in comparison to control cells, suggesting, as has beenreported in other cell lines, that low levels of Apc are sufficient todownregulate β-catenin [45]. Lower β-catenin expression due toApc knockdown contrasts observations in tumors, in which Apcinactivation due to deletion or mutation is linked to increased β-catenin expression [12]. In contrast to these models, KSFrt-Apcsistill expresses wild-type Apc albeit at lower levels. Furthermore,cells carrying hypomorphic Apcmutations show upregulation of β-catenin levels only when the Apc activity is reduced below 2% ofthe normal levels [26]. Interestingly, the increased activity of theBAT-LucWnt responsive construct in the KSFrt-Apcsi cells implies ashift of the inactive/active β-catenin balance in favor of the activefraction. The partial rescue of the Apcsi-induced Wnt activationafter transfection with an APC expression vector demonstratesthat the upregulation of the Wnt signal in the KSFrt-Apcsi cells isdue to Apc knockdown. We recently described that the 4C3 Frtclone of the parental KS483 murine mesenchymal progenitor linecan differentiate into osteoblasts, chondrocytes and adipocytes,when cultured in the appropriate conditions and represents avaluable biological tool for the evaluation of gene function both invivo and in vitro [49,51,52]. Thus, the KSFrt-Apcsi cell line is areliable model to study the role of Apc in regulating differentiationof SPC [49]. It is well established that APC modulates cell shape byorganizing the cytoskeleton in particular through stabilization ofmicrotubules [39]. The KSFrt-Apcsi cell line developed elongatedcellular protrusions, thereby displaying a clearly distinct morphol-ogy from the control cells. In agreement with this, upregulation ofthe canonical Wnt signal has been shown to promote a spindle-like cell morphology [15,36,54].
It is generally accepted that Apc inhibits cell proliferation viaβ-catenin-dependent and -independent actions and that inactiva-tion of APC represents the early, initiating event in severalmalignant diseases [5,10,41]. However, evidence is also availablesuggesting that APC is essential for cell proliferation [45]. Likewise,no consensus regarding the effect of APC on apoptosis has beenreached since both stimulation and inhibition of apoptosis by APChave been described [1,3,13,38]. The role of APC in apoptosis, suchas observed in the KSFrt-Apcsi can be either β-catenin-dependentor independent [3,16]. Based on these results, we currently favorthe hypothesis that Apc plays opposing roles during developmentand malignant transformation, by modulating cell shape, prolifer-ation, and survival in a context-dependent manner, with distinct
consequences in different cell types and at different developmen-tal stages.
The canonical Wnt/β-catenin signaling pathway governs thelineage commitment of bi-potential SPC into osteoblasts orchondrocytes [18]. Roughly, it is proposed that upregulation ofthis pathway induces the differentiation of SPC into precursors ofthe osteogenic lineage, whereas its downregulation is needed forchondrogenic differentiation [17]. Data available from in vivo andex vivo studies indicate that the osteogenic differentiationpotential is altered when Apc is lacking or mutated, even if theresulting levels of β-catenin are high [26,36]. Although beingexposed to higher levels of transcriptionally active Wnt and BMPsignaling, KSFrt-Apcsi cells display a diminished osteogenicdifferentiation potential. Similar findings were made in condition-al Apc knockout mice, in which inactivation of Apc in SPCscompletely blocked osteoblast and chondrocyte differentiationparticular in early stages of skeletogenesis [36]. The latter studyhas also shown that the inhibitory phase in some skeletal elementsis followed by accelerated osteoblast formation in later develop-mental stages [36]. Complete inhibition of osteogenesis byknockdown of Apc appears in contrast with increased BMD andhigh incidence of osteoma in FAP patients carrying a heterozygousinactivating mutation of APC [37]. In addition, conditional Apcknockout using Cre expression under the influence of theOsteocalcin promoter, a late marker of osteoblast differentiation,results in increased bone formation and lack of osteoclastformation [20]. Therefore we hypothesized that the inhibitoryeffect on osteoblast differentiation in the KSFrt-Apcsi cells is celltype-dependent and may be reversed by environmental factorslike exposure to exogenous growth factors.
Interestingly, when the KSFrt-Apcsi cells were exposed toadditional high concentrations of BMP-7 and to a lesser extentBMP-6, both potent stimulators of osteogenesis [29], they displayedan increased potential to form osteoblasts in comparison to controlcells. Such rescue effect was not observed when using other pro-osteogenic growth factors like bFGF, TGF-β3, PTHrP, IGF-1.Oneof thepotential interpretations is that BMP signaling further activatescanonical Wnt signaling, thus it synergistically induces the osteo-blast differentiation in KSFrt-Apcsi cells. Our results indicate that Apcis essential for the osteogenic differentiation of the KS483 cell lineand that thenoxious effect ofApc knockdownon osteogenesis can beoverruled by high BMP signaling induced by BMP-7. Consistently, invitro observations made in C3H10T1/2 cells demonstrate thatcanonical Wnt signaling itself is not sufficient, but in synergy withBMP signaling it can promote osteoblast differentiation [34].
Both the canonical Wnt and the BMP signaling pathway havebeen shown to promote osteoblast differentiation, maturationand mineralization [30]. However, the complexity of the interac-tions between these regulatory pathways and the abundance of invitro reports investigating this interrelation in different osteo-genic experimental setups, complicate its understanding[2,11,14,23,32,34]. The most probable explanation for the widevariety of effects arising upon this interaction is that theyrepresent different aspects of Wnt and BMP functions that areonly visible in certain cell types, at specific developmental stagesand under particular experimental conditions. Our results addinsight to the complexity of interactions between Wnt/β-cateninand BMP signaling during the differentiation of SPC. In vitro, BMPsinduce Wnt expression [7,44], whereas Wnt signaling inducesBMP expression [2,55], suggesting that both Wnt and BMP
1419E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 1 4 1 1 – 1 4 2 1

Author's personal copy
signaling may jointly regulate each other in osteoblasts. In theKS483 cells, Apc knockdown upregulated not only transduction ofthe Wnt signal, but also the BMP signaling pathway, most likelyvia upregulation of Bmp7 expression. APC can shuttle into and outof the nucleus [19,40], and thus a possible Apc-mediatedinteraction between Wnt and BMP may occur in any of thesetwo subcellular locations. While in the nucleus the Smad/β-catenin/Lef protein complex regulates many shared target genes[21,28,48], in the cytoplasm, BMP can either impede or stimulatethe canonical Wnt signal via Axin [32,46]. Since Apc comprisesboth Axin and β-catenin binding domains, we speculate that Apcmight link the Wnt/β-catenin to BMP signaling pathways duringosteoblast differentiation of KS483 cells.
Our present results indicate that Apc is essential for osteogenic,chondrogenic and adipogenic differentiation of the murinemesenchymal-like KS483 cell line which has SPC-like character-istics. Our approach has provided a valuable model in which wedemonstrate that levels of functional Apc must be tightlycontrolled for proper modulation of the transcriptionally activeβ-catenin and BMP signaling dosage required for multilineage SPCdifferentiation in vitro.
Conflict of interest
All authors have no conflicts of interest.
Acknowledgments
We are grateful to Prof. Dr. Peter ten Dijke (Department ofMolecular Cell Biology, LUMC, Leiden, the Netherlands) for thepGL3-(BRE)2-Luc reporter construct; Dr. Stefano Piccolo (De-partment of Medical Biotechnologies, University of Padua, Italy)for the BAT-Luc reporter construct; and Prof. Dr. Bert Vogelstein(The Ludwig Center for Cancer Genetics and Therapeutics,Howard Hughes Medical Institute and Sidney Kimmel CancerCenter at the Johns HopkinsMedical Institutions, Baltimore, MD,USA) for the pSAR-MT-APC construct. We thank Prof. Dr.Slobodan Vukicevic, Department of Anatomy, School of Medi-cine, Zagreb, Croatia, for kindly providing BMP-6 and BMP-7 forour study.
This work was financially supported by an unrestrictededucational grant from IPSEN FARMACEUTICA BV to RLM.
R E F E R E N C E S
[1] Y. Ahmed, S. Hayashi, A. Levine, E. Wieschaus, Regulation ofarmadillo by a Drosophila APC inhibits neuronal apoptosis duringretinal development, Cell 93 (1998) 1171–1182.
[2] G. Bain, T. Muller, X. Wang, J. Papkoff, Activated beta-catenininduces osteoblast differentiation of C3H10T1/2 cells andparticipates in BMP2 mediated signal transduction, Biochem.Biophys. Res. Commun. 301 (2003) 84–91.
[3] H. Benchabane, Y. Ahmed, The adenomatous polyposis coli tumorsuppressor and Wnt signaling in the regulation of apoptosis, Adv.Exp. Med. Biol. 656 (2009) 75–84.
[4] P.C. Bessa, M. Casal, R.L. Reis, Bone morphogenetic proteins intissue engineering: the road from the laboratory to the clinic, partI (basic concepts), J. Tissue Eng. Regen. Med. 2 (2008) 1–13.
[5] D.J. Carson, I.M. Santoro, J. Groden, Isoforms of the APC tumorsuppressor and their ability to inhibit cell growth andtumorigenicity, Oncogene 23 (2004) 7144–7148.
[6] M. Chen, M. Zhu, H. Awad, T.F. Li, T.J. Sheu, B.F. Boyce, D. Chen, R.J.O'Keefe, Inhibition of beta-catenin signaling causes defects inpostnatal cartilage development, J. Cell Sci. 121 (2008)1455–1465.
[7] Y. Chen, H.C. Whetstone, A. Youn, P. Nadesan, E.C. Chow, A.C. Lin,B.A. Alman, Beta-catenin signaling pathway is crucial for bonemorphogenetic protein 2 to induce new bone formation, J. Biol.Chem. 282 (2007) 526–533.
[8] H. Clevers, Wnt/beta-catenin signaling in development anddisease, Cell 127 (2006) 469–480.
[9] J.G. Doench, C.P. Petersen, P.A. Sharp, siRNAs can function asmiRNAs, Genes Dev. 17 (2003) 438–442.
[10] M.C. Faux, J.L. Ross, C. Meeker, T. Johns, H. Ji, R.J. Simpson, M.J.Layton, A.W. Burgess, Restoration of full-length adenomatouspolyposis coli (APC) protein in a colon cancer cell line enhancescell adhesion, J. Cell Sci. 117 (2004) 427–439.
[11] L. Fischer, G. Boland, R.S. Tuan, Wnt signaling during BMP-2stimulation of mesenchymal chondrogenesis, J. Cell. Biochem. 84(2002) 816–831.
[12] R. Fodde, The multiple functions of tumour suppressors: it's all inAPC, Nat. Cell Biol. 5 (2003) 190–192.
[13] C. Gaspar, R. Fodde, APC dosage effects in tumorigenesis and stemcell differentiation, Int. J. Dev. Biol. 48 (2004) 377–386.
[14] X. Guo, X.F. Wang, Signaling cross-talk between TGF-beta/BMPand other pathways, Cell Res. 19 (2009) 71–88.
[15] M. Haertel-Wiesmann, Y. Liang, W.J. Fantl, L.T. Williams,Regulation of cyclooxygenase-2 and periostin by Wnt-3 inmouse mammary epithelial cells, J. Biol. Chem. 275 (2000)32046–32051.
[16] C.A. Hanson, J.R. Miller, Non-traditional roles for the adenomatouspolyposis coli (APC) tumor suppressor protein, Gene 361 (2005)1–12.
[17] C. Hartmann, A Wnt canon orchestrating osteoblastogenesis,Trends Cell Biol. 16 (2006) 151–158.
[18] C. Hartmann, Transcriptional networks controlling skeletaldevelopment, Curr. Opin. Genet. Dev. 19 (2009) 437–443.
[19] B.R. Henderson, Nuclear-cytoplasmic shuttling of APC regulatesbeta-catenin subcellular localization and turnover, Nat. Cell Biol. 2(2000) 653–660.
[20] S.L. Holmen, C.R. Zylstra, A. Mukherjee, R.E. Sigler, M.C. Faugere,M.L. Bouxsein, L. Deng, T.L. Clemens, B.O. Williams, Essential roleof beta-catenin in postnatal bone acquisition, J. Biol. Chem. 280(2005) 21162–21168.
[21] S.M. Hussein, E.K. Duff, C. Sirard, Smad4 and beta-cateninco-activators functionally interact with lymphoid-enhancingfactor to regulate graded expression of Msx2, J. Biol. Chem.278 (2003) 48805–48814.
[22] E.H. Jho, T. Zhang, C. Domon, C.K. Joo, J.N. Freund, F. Costantini,Wnt/beta-catenin/Tcf signaling induces the transcription ofAxin2, a negative regulator of the signaling pathway, Mol. Cell.Biol. 22 (2002) 1172–1183.
[23] N. Kamiya, T. Kobayashi, Y. Mochida, P.B. Yu, M. Yamauchi, H.M.Kronenberg, Y. Mishina, Wnt inhibitors Dkk1 and sost aredownstream targets of BMP signaling through the type IAreceptor (BMPRIA) in osteoblasts, J. Bone Miner. Res. (2009).
[24] G. Karsenty, The complexities of skeletal biology, Nature 423(2003) 316–318.
[25] G. Karsenty, H.M. Kronenberg, C. Settembre, Genetic control ofbone formation, Annu. Rev. Cell Dev. Biol. 25 (2009) 629–648.
[26] M.F. Kielman, M. Rindapaa, C. Gaspar, N. van Poppel, C. Breukel,S. van Leeuwen, M.M. Taketo, S. Roberts, R. Smits, R. Fodde, Apcmodulates embryonic stem-cell differentiation by controllingthe dosage of beta-catenin signaling, Nat. Genet. 32 (2002)594–605.
[27] O. Korchynskyi, D.P. ten, Identification and functionalcharacterization of distinct critically important bone
1420 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 1 4 1 1 – 1 4 2 1

Author's personal copy
morphogenetic protein-specific response elements in the Id1promoter, J. Biol. Chem. 277 (2002) 4883–4891.
[28] E. Labbe, A. Letamendia, L. Attisano, Association of Smads withlymphoid enhancer binding factor 1/T cell-specific factormediates cooperative signaling by the transforming growthfactor-beta and wnt pathways, Proc. Natl. Acad. Sci. USA97 (2000) 8358–8363.
[29] K. Lavery, S. Hawley, P. Swain, R. Rooney, D. Falb, M.H.Aoui-Ismaili, New insights into BMP-7 mediated osteoblasticdifferentiation of primary human mesenchymal stem cells, Bone45 (2009) 27–41.
[30] J.B. Lian, G.S. Stein, A. Javed, A.J. van Wijnen, J.L. Stein, M.Montecino, M.Q. Hassan, T. Gaur, C.J. Lengner, D.W. Young,Networks and hubs for the transcriptional control ofosteoblastogenesis, Rev. Endocr. Metab. Disord. 7 (2006) 1–16.
[31] F. Liu, S. Kohlmeier, C.Y. Wang, Wnt signaling and skeletaldevelopment, Cell. Signal. 20 (2008) 999–1009.
[32] Z. Liu, Y. Tang, T. Qiu, X. Cao, T.L. Clemens, A dishevelled-1/Smad1interaction couples WNT and bone morphogenetic proteinsignaling pathways in uncommitted bone marrow stromal cells, J.Biol. Chem. 281 (2006) 17156–17163.
[33] S. Maretto, M. Cordenonsi, S. Dupont, P. Braghetta, V. Broccoli, A.B.Hassan, D. Volpin, G.M. Bressan, S. Piccolo, MappingWnt/beta-catenin signaling during mouse development andin colorectal tumors, Proc. Natl. Acad. Sci. USA 100 (2003)3299–3304.
[34] G. Mbalaviele, S. Sheikh, J.P. Stains, V.S. Salazar, S.L. Cheng, D.Chen, R. Civitelli, Beta-catenin and BMP-2 synergize to promoteosteoblast differentiation and new bone formation, J. Cell.Biochem. 94 (2005) 403–418.
[35] R.L. Miclea, E.C. Robanus-Maandag, J.J. Goeman, L. Finos, H. Bloys,C.W.G.M. Löwik, J.M. Wit, M. Karperien. Inhibition of GSK3-betainduces loss of the chondrocytic phenotype and cartilage matrixdegradation through activation of canonical Wnt signaling.submitted. 2010.
[36] R.L. Miclea, M. Karperien, C.A. Bosch, H.G. van der, V. d. van, T.Kobayashi, H.M. Kronenberg, G. Rawadi, P. Akcakaya, C.W. Lowik,R. Fodde, J.M. Wit, E.C. Robanus-Maandag, Adenomatouspolyposis coli-mediated control of beta-catenin is essential forboth chondrogenic and osteogenic differentiation of skeletalprecursors, BMC Dev. Biol. 9 (2009) 26.
[37] R.L. Miclea, M. Karperien, A.M. Langers, E.C. Robanus-Maandag, L.A.van, B. van der Hiel, M.P. Stokkel, B.E. Ballieux,W.Oostdijk, J.M.Wit,H.F. Vasen, N.A. Hamdy, APC mutations are associated withincreased bone mineral density in patients with familialadenomatous polyposis, J. Bone Miner. Res. (2010).
[38] P.J. Morin, B. Vogelstein, K.W. Kinzler, Apoptosis and APC incolorectal tumorigenesis, Proc. Natl. Acad. Sci. USA 93 (1996)7950–7954.
[39] I.S. Nathke, The adenomatous polyposis coli protein: the Achillesheel of the gut epithelium, Annu. Rev. Cell Dev. Biol. 20 (2004)337–366.
[40] K.L. Neufeld, R.L. White, Nuclear and cytoplasmic localizations ofthe adenomatous polyposis coli protein, Proc. Natl. Acad. Sci. USA94 (1997) 3034–3039.
[41] K.S. Park, S.H. Jeon, S.E. Kim, Y.Y. Bahk, S.L. Holmen, B.O. Williams,K.C. Chung, Y.J. Surh, K.Y. Choi, APC inhibits ERK pathway
activation and cellular proliferation induced by RAS, J. Cell Sci.119 (2006) 819–827.
[42] C.P. Petersen, M.E. Bordeleau, J. Pelletier, P.A. Sharp, Short RNAsrepress translation after initiation in mammalian cells, Mol. Cell21 (2006) 533–542.
[43] M.K. Rao, J. Pham, J.S. Imam, J.A. MacLean, D. Murali, Y. Furuta, A.P.Sinha-Hikim, M.F. Wilkinson, Tissue-specific RNAi reveals thatWT1 expression in nurse cells controls germ cell survival andspermatogenesis, Genes Dev. 20 (2006) 147–152.
[44] G. Rawadi, B. Vayssiere, F. Dunn, R. Baron, S. Roman-Roman,BMP-2 controls alkaline phosphatase expression and osteoblastmineralization by a Wnt autocrine loop, J. Bone Miner. Res. 18(2003) 1842–1853.
[45] J. Schneikert, J. Behrens, Truncated APC is required for cellproliferation and DNA replication. Int.J, Cancer 119 (2006)74–79.
[46] Y. Tang, Z. Liu, L. Zhao, T.L. Clemens, X. Cao, Smad7 stabilizesbeta-catenin binding to E-cadherin complex and promotescell-cell adhesion, J. Biol. Chem. 283 (2008) 23956–23963.
[47] P. ten Dijke, Bone morphogenetic protein signal transduction inbone, Curr. Med. Res. Opin. 22 (Suppl 1) (2006) S7–11.
[48] T. Theil, S. Aydin, S. Koch, L. Grotewold, U. Ruther,Wnt and Bmpsignalling cooperatively regulate graded Emx2 expression inthe dorsal telencephalon, Development 129 (2002)3045–3054.
[49] G. van der Horst, K.E. de Rooij, J. Hoogendam, H.C.M. Sips, A.L.Feitsma, I. Que, L. van der Wee-Pals, C.W.G.M. Löwik, M.Karperien. Functional genomics, drug screening and biomaterialevaluation using mesenchymal progenitor cells. 2010.
[50] G. van der Horst, H. Farih-Sips, C.W. Lowik, M. Karperien,Hedgehog stimulates only osteoblastic differentiation ofundifferentiated KS483 cells, Bone 33 (2003) 899–910.
[51] G. van der Horst, H. Farih-Sips, C.W. Lowik, M. Karperien, Multiplemechanisms are involved in inhibition of osteoblastdifferentiation by PTHrP and PTH in KS483 Cells, J. Bone Miner.Res. 20 (2005) 2233–2244.
[52] G. van der Horst, S.M. van der Werf, H. Farih-Sips, R.L. vanBezooijen, C.W. Lowik, M. Karperien, Downregulation of Wntsignaling by increased expression of Dickkopf-1 and −2 is aprerequisite for late-stage osteoblast differentiation of KS483cells, J. Bone Miner. Res. 20 (2005) 1867–1877.
[53] J.H. van Es, R.H. Giles, H.C. Clevers, The many faces of the tumorsuppressor gene APC, Exp. Cell Res. 264 (2001) 126–134.
[54] A.T. Weeraratna, Y. Jiang, G. Hostetter, K. Rosenblatt, P. Duray, M.Bittner, J.M. Trent, Wnt5a signaling directly affects cell motilityand invasion of metastatic melanoma, Cancer Cell 1 (2002)279–288.
[55] D.G. Winkler, M.S. Sutherland, E. Ojala, E. Turcott, J.C. Geoghegan,D. Shpektor, J.E. Skonier, C. Yu, J.A. Latham, Sclerostin inhibition ofWnt-3a-induced C3H10T1/2 cell differentiation is indirect andmediated by bone morphogenetic proteins, J. Biol. Chem. 280(2005) 2498–2502.
[56] Y. Yan, D. Tang, M. Chen, J. Huang, R. Xie, J.H. Jonason, X. Tan, W.Hou, D. Reynolds, W. Hsu, S.E. Harris, J.E. Puzas, H. Awad, R.J.O'Keefe, B.F. Boyce, D. Chen, Axin2 controls bone remodelingthrough the beta-catenin-BMP signaling pathway in adult mice, J.Cell Sci. 122 (2009) 3566–3578.
1421E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 1 4 1 1 – 1 4 2 1




















