
EFFECT OF TREE AGE ON VARIATION OF PINUS RADIATA D. DON CHEMICAL COMPOSITION
Upload
independentCategory
view
0download
0
Spatial variation and understorey competition effect of Pinus radiata fineroots in a silvopastoral system in New Zealand
M.K. Gautam1, D.J. Mead2, S.X. Chang3,* and P.W. Clinton4
1School of Resources, Environment and Society, Australian National University, 0200 Canberra, Australia;2Silviculture Advisor, Golden Bay, New Zealand; 3Centre for Enhanced Forest Management, Department ofRenewable Resources, University of Alberta,, 4–42 Earth Sciences Building, T6G 2E3 Edmonton, Alberta,Canada; 4New Zealand Forest Research Institute Ltd, P.O. Box 29237, Christchurch, New Zealand; *Authorfor correspondence (e-mail: [email protected]; phone: (1)(780) 492-6375; fax: (1)(780) 492-1767)
Received 13 March 2001; accepted in revised form 23 April 2002
Key words: Fine root length density (FRLD), Genotype, Microsite, Ripping, Root distribution, Tree-pasture in-teraction
Abstract
In designing agroforestry systems, the combination of tree genotype (or species) and pasture species and thespatial arrangement of trees are important considerations. The spatial variation of fine root length density (FRLD)of three radiata pine (Pinus radiata D. Don) genotypes, referred to here as clone 3, clone 4 and seedlings, wasstudied in a three-year-old temperate silvopastoral experiment. The genotypes were planted with three understo-rey types: ryegrass (Lolium perenne) mixed with clovers (Trifolium spp), lucerne (Medicago sativa), and control(bare ground). Also fine root distribution of both tree and pasture species with soil depth and in relation to treerow (0.9 m north or south of and within the ripped tree row) was studied. Greater FRLD was found in clonalthan in seedling trees in the bare ground treatment but not in the two pasture treatments, and in the 0–0.1 m butnot in the 0.1–0.2 or 0.2–0.3 m soil layers. Clonal trees had a greater ability to develop a more extensive rootsystem, especially in the 0–0.1 m soil layer, but that advantage disappeared when they were planted with pasturespecies since competition from the pasture species was most severe in the 0–10 cm layer. The FRLD of lucernewas greater than that of ryegrass/clovers, consistent with the greater aboveground biomass production of lucerne.Pasture species FRLD was greater on the south (wetter) than on the north side of the ripline or in the ripline. Theinterception of prevailing southerly rain-bearing wind by tree crowns resulted in the south side being wetter thanthe north side. Results indicated that production and distribution of fine roots of both tree and pasture speciesresponded to changes in the microclimate. We suggest that to optimize pasture/tree biomass production plantingtrees in the north-south direction is better than in the east-west direction at the studied site.
Introduction
Knowing the spatial distribution of fine roots of thetrees and crops is important to understand the inter-actions between components in agroforestry systems.The horizontal and vertical growth and distribution oftree fine roots differ between species (Nambiar 1983;Jonsson et al. 1988) and sites (Hendrick and Pregitzer1993; Ares and Peinemann 1992). The rooting abilityand root development of trees are under strong ge-netic control; different clones of a given tree species
are known to respond differently to site factors andmanagement (Stokes et al. 1997). Fine root lengthdensity (FRLD) varied among radiata pine (Pinus ra-diata) families (Theodorou and Bowen 1993) and in-creased with increasing soil moisture availability, andvice versa (Theodorou et al. 1991). Some genotypesof radiata pine, the most popular plantation and agro-forestry tree species in New Zealand, may be moreadaptable than others to dry conditions, and are thusmore suitable in low-rainfall regions, or where under-storey competition for soil moisture is intense. One
89Agroforestry Systems 55: 89–98, 2002.© 2002 Kluwer Academic Publishers. Printed in the Netherlands.
of the objectives of establishing the Lincoln Univer-sity agroforestry experiment was to test if radiata pinegenotypes can be selected to better suit a site wherethere is competition from pasture species and wheremoisture supply is limited (Mead et al. 1993).
Tree spacing and spatial arrangement can alter thedistribution of fine roots in forest plantations (East-ham and Rose 1990). In agroforestry systems, treesare often planted in rectangular patterns to allow un-derstorey crops to be cultivated and utilised betweentree rows. Such arrangements may determine the hor-izontal and vertical distribution of fine roots of bothtree and pasture species (Eastham and Rose 1990),and the mutualistic or competitive relationships be-tween species (Anderson and Sinclair 1993). Treesand understorey species will compete strongly for re-sources if the fine roots of both are concentrated inthe same soil layer; but if the fine roots of the speciesare distributed in different soil layers, there may beminimal or no competition between the species (Ong1991). One of the challenges is to understand rela-tionships between the distribution of fine roots of un-derstorey species and that of trees in agroforestry sys-tems and the implications for tree growth. Suchknowledge may be useful in the selection of tree spe-cies or genotypes, and understorey species to reducebelowground competition and achieve optimal pro-ductivity in agroforestry systems. There is a dearth ofinformation, however, on how tree-understorey con-figuration/competition affects tree fine root distribu-tion in agroforestry systems.
The current study was carried out to compare fineroot ( � 2 mm diameter) length density for three ra-diata pine genotypes growing under contrasting un-derstorey treatments. Those genotypes are different inaboveground growth rates (Mead et al. 1993). Thespecific objectives of this study were to examine: (1)whether understorey competition altered the distribu-tion of FRLD of radiata pine genotypes; (2) whetherradiata pine genotypes respond to understorey com-petition differently; and (3) to what extent horizontaland vertical distribution of FRLD of both trees andunderstorey changed with the microsite conditions.We hypothesized that (1) the genotypes that havehigher aboveground growth rate also have greater fineroot production and thus have greater ability to com-pete with pasture species for soil resources; and (2)FRLD of pine and pasture species will be in greatercompetition in the north side because of unfavourablemicrosite conditions and the scarcity of resourceavailability that limit plant growth in general.
Materials and methods
The site
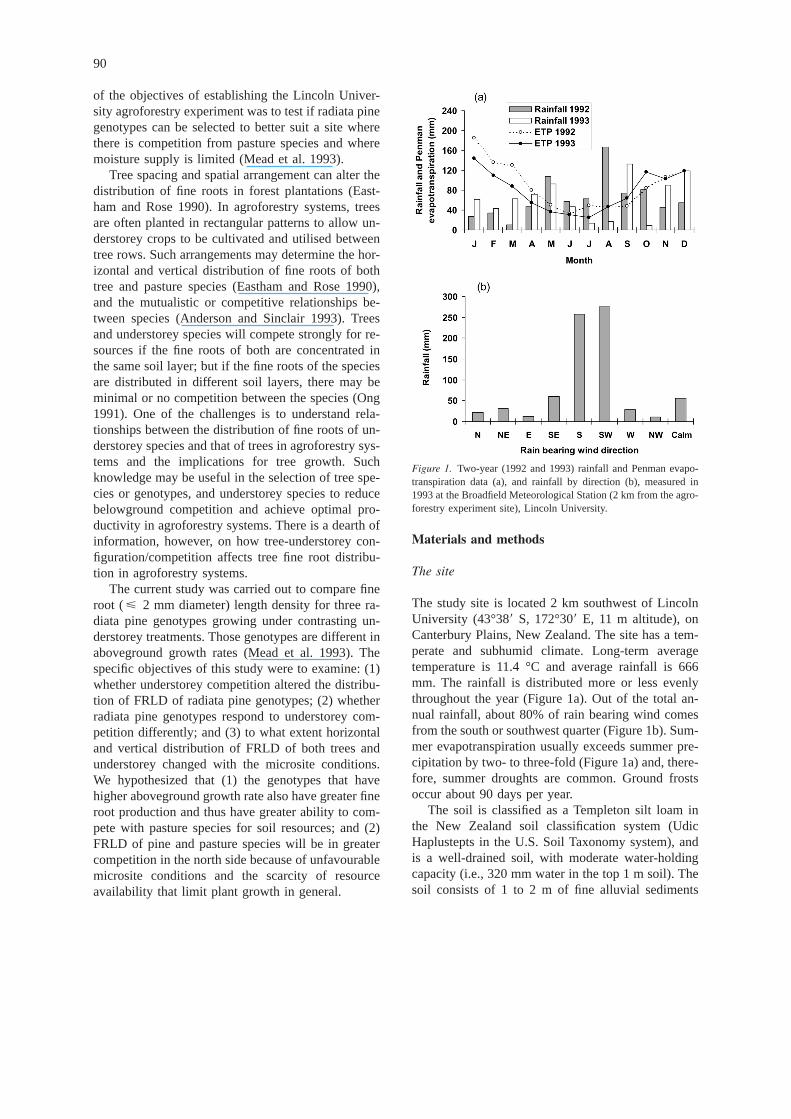
The study site is located 2 km southwest of LincolnUniversity (43°38� S, 172°30� E, 11 m altitude), onCanterbury Plains, New Zealand. The site has a tem-perate and subhumid climate. Long-term averagetemperature is 11.4 °C and average rainfall is 666mm. The rainfall is distributed more or less evenlythroughout the year (Figure 1a). Out of the total an-nual rainfall, about 80% of rain bearing wind comesfrom the south or southwest quarter (Figure 1b). Sum-mer evapotranspiration usually exceeds summer pre-cipitation by two- to three-fold (Figure 1a) and, there-fore, summer droughts are common. Ground frostsoccur about 90 days per year.
The soil is classified as a Templeton silt loam inthe New Zealand soil classification system (UdicHaplustepts in the U.S. Soil Taxonomy system), andis a well-drained soil, with moderate water-holdingcapacity (i.e., 320 mm water in the top 1 m soil). Thesoil consists of 1 to 2 m of fine alluvial sediments
Figure 1. Two-year (1992 and 1993) rainfall and Penman evapo-transpiration data (a), and rainfall by direction (b), measured in1993 at the Broadfield Meteorological Station (2 km from the agro-forestry experiment site), Lincoln University.
90

over gravels. The top 0.2 m contains about 2.8% car-bon and 0.24% nitrogen, and has a pH of 5.8 (Changet al. 2001). The soil is considered one of the mostproductive in the region and peas (Pisum sativa) wereplanted before the experiment was established. Thispractice may have improved the soil N status of thesite but should have not interfered with this study, aspeas were uniformly planted across the site.
Study design
The main experiment, established to study tree/pas-ture interactions, includes six understorey treatmentsin the main plots and five radiata pine planting stocktypes (genotypes) in the subplots in a split-plot de-sign. Details of the experimental design were de-scribed in Mead et al. (1993). For the current study,we selected three understorey types, i.e., bare ground,perennial ryegrass (Lolium perenne) with clovers(Trifolium spp.), and lucerne (Medicago sativa), as-signed to the main plot level, and three Pinus radiatagenotypes, i.e., tissue-cultured clone 3 and clone 4,and seedlings (raised from open pollinated seed), as-signed to the subplot level.
The main plots were 42 × 46.2 m (0.194 ha). Ineach plot, trees were planted in the winter of 1990 at7 (between rows) × 1.4 m (within a row) spacing,which gives an initial density of 1000 stems/ha. Afterplowing, the rows for tree planting were ripped to adepth of 0.6 m. The planted tree rows were in east-west direction. Proper buffer rows were established.Trees in the plots were thinned in 1992 with 200stems/ha removed. Tree density was 800 stems/ha atthe time of this study (three years after planting), withno tree pruning.
In the spring of 1990, the pasture crops were sownbetween the tree rows, 0.5 m away from the ripline.The pasture was cut and carried for silage four timesin the first two growing seasons, as the trees were toosmall and could be damaged by grazing sheep. Themain plots were fenced and rotational grazing com-menced in spring 1993. Throughout the experiment,no fertilizer or irrigation was used. Weeds in 1 mstrips centred on tree rows were controlled with re-peated application of a herbicide (glyphosate)throughout this study.
The sub-sub-plot and sub-sub-sub-plot factorsstudied in this paper were position in relation to treerows and soil depth, respectively. The positions stud-ied were 0.9 m north and 0.9 m south of tree rows
and in the tree row (ripline). Soil depths studied were0–0.1, 0.1–0.2, and 0.2–0.3 m.
Root sampling
Three trees from each sub-plot were selected for rootsampling, following the stratified sampling proceduredescribed in Avery and Burkhart (1983). The treesselected for sampling covered the range of diametersobserved from the whole treatment plots. At the timeof root sampling in November 1993, mean tree heightwas greater in the bare ground than in the ryegrass/clovers, or lucerne treatments, and was greater inclone 3 than in both clone 4 and seedling trees (Ta-ble 1). Mean basal diameter was greater in the bareground than in the ryegrass/clovers or lucerne treat-ments; however, differences between genotypes weresmall (Table 1).
To ensure a good representation of the fine rootpopulation, four soil cores from both the north andsouth positions and two cores (0.4 m away from treestems) from the ripline were taken following the pro-cedure used by Hughes et al. (1986) and Vogt andPersson (1991). A 45-mm internal diameter steel coresampler was driven into the ground and extractedwith a tripod and tackle. Soil cores were taken to adepth of 0.3 m and divided into three layers by 0.1 mintervals; replicates for each position were pooled toform a composite sample for each depth. Sampleswere kept in airtight polythene bags and stored in acold room at 2 °C until they were washed.
For root washing, the soil cores were soaked inwater for 4–5 hours and then transferred on to fourstacked metal sieves with different sizes (2.5, 2, 1,and 0.5 mm). Once the soil was washed away the re-mainder was transferred from the sieves into a beakerfor decantation. Organic debris was discarded, andthe dead roots were separated from the live onesbased on the visual appearance of root bark (i.e., theblack, soggy and floating nature of dead roots). Deadroots were discarded. Tree roots were separated fromthe pasture roots using characteristics such as roottexture, colour, structure (waviness and branchiness),thickness, and density (suspension). In general, pas-ture roots were smaller in diameter than pine rootsand lighter in colour. Also, pasture roots were rela-tively wavy but straighter than pine roots. Lucerneroots were tangled in appearance and sometimes hadbacterial nodules. Pine roots greater than 2 mm diam-eter were discarded from the sample. There were nopasture roots greater than 1 mm in diameter.
91

Root length was measured using a ‘Comair’ rootlength scanner (Commonwealth Aircraft CorporationLtd, Melbourne, Victoria) (Richards et al. 1979). Thescanner was calibrated to a known length of wire withdiameter similar to that of the fine roots. The regres-sion equation was: Actual Length = 1.0326 × ScannedLength (R2 = 0.96).
Soil moisture content (volumetric) was measuredby time domain reflectometry or TDR (Trase, Model6050X1, Soil Moisture Equipment Corp., Santa Bar-bara, CA). The precipitation and evapotranspirationdata were provided by the Broadfield MeteorologicalStation at Lincoln University. The rainfall by winddirection was partitioned based on the prevailingwind and the precipitation recorded for a particularday or rainfall event. Tree height was measured usinga height pole and basal diameter using a digital cal-liper. Volume index was calculated as diameter2 ×height.
Data analyses
The data was analysed using a split-plot model whereunderstorey was the main plot effect, tree genotypewas the sub-plot effect, position (north or south ofripline or in the ripline) was the sub-sub-plot effect,and soil depth was the sub-sub-sub-plot effect. Allfour factors were treated as having fixed effects.
Pine and pasture FRLD was calculated for a par-ticular position (e.g., north or south side), and forthree depths (0–0.1, 0.1–0.2, and 0.2–0.3 m), and ex-
pressed as cm/100 cm3 after Bi et al. (1992). Theproportional pine FRLD (pine FRLD as a percentageof the total FRLD, i.e., tree plus pasture FRLD) wascalculated for the lucerne and ryegrass/clovers treat-ments. The proportional pine FRLD is an indicator ofthe magnitude of rooting competition. This was notcalculated for the bare ground, as there were no un-derstorey roots in those plots.
Data were sparsely distributed and skewed andwere natural log transformed before further analysis.Arc sine transformation was performed for the pro-portional data. Means reported in the Results sectionwere back transformed. The GLM procedure of theSYSTAT (5th edition, Evienston, Illinois, USA) soft-ware was used for the analysis of variance. Multiplecomparison (LSD) was performed to test the differ-ences between means. The aboveground tree size wasanalysed using the initial size as covariate and thecovariate was not significant at the � = 0.05 level.
Results
Precipitation and soil moisture content
Total precipitation in 1992 and 1993 was very similar(769 and 760 mm, respectively) and was greater thanthe long-term average of 666 (Mead et al. 1993).However, the evapotranspiration potential was muchgreater than the actual precipitation in both years,with 1069 and 941 mm/year for 1992 and 1993, re-
Table 1. Basal diameter, height, and volume index of three-year-old Pinus radiata trees sampled for FRLD (fine root length density) studyin the Lincoln University agroforestry experiment, New Zealand*
Treatment Basal diameter (mean ± SD, mm) Height (mean ± SD, m) Volume index (mean ± SD, m3 × 10−5)
Bare ground
Clone 3 118 (25) 3.34 (0.30) 9.9 (3.93)
Clone 4 132 (18) 2.99 (0.53) 10.0 (4.96)
Seedlings 116 (31) 2.41 (0.60) 5.5 (3.89)
Ryegrass/clovers
Clone 3 96 (15) 3.21 (0.31) 5.5 (2.69)
Clone 4 90 (13) 2.89 (0.48) 4.8 (2.55)
Seedlings 94 (16) 2.54 (0.67) 3.3 (2.59)
Lucerne
Clone 3 82 (19) 2.85 (0.53) 7.9 (3.88)
Clone 4 75 (14) 2.51 (0.44) 7.2 (3.31)
Seedlings 81 (14) 2.58 (0.35) 5.1 (3.50)
* n = 9.
92

spectively (Figure 1a). There was considerably morerainfall in the summer period (November to Januaryin New Zealand) in 1993 than in 1992, resulting inthe smaller evapotranspiration potential and a smallersummer soil water deficit in 1993. Most of the totalrainfall comes from the south and southwest direc-tions (Figure 1b). The north side was in the rainshadow. As the trees had branches to ground levelwhen this study was initiated, the soil in the north sideof the ripline (tree row) was drier than that in thesouth side of the ripline (Figure 2), particularly in thewinter when precipitation was being stored in the soil.
Pine root length density (FRLD)
Genotype, sampling position, and depth affected pineFRLD (P < 0.01, ANOVA not shown). Understorey
did not affect FRLD (P = 0.219), however, the inter-actions between understorey and genotype (P =0.046), understorey and position (P = 0.007), geno-type and depth (P = 0.007), and position and depth(P = 0.013) were significant.
The interaction between understorey and genotypewas caused by a similar FRLD among clone 3, clone4 and seedling trees in the ryegrass/clovers and luc-erne treatments, whereas the FRLD of the clonal treeswas twice that of the seedling trees in the bare groundtreatment (Figure 3a). The interaction between geno-type and depth was due to a significantly greaterFRLD in clone 3 and clone 4 than in the seedlingtrees in the 0–0.1 m layer, whereas the FRLD of thegenotypes was similar in the 0.1–0.2 and 0.2–0.3 mlayers (Figure 3b). In addition, pine FRLD was lowerin the 0–0.1 m than in the 0.1–0.2 or 0.2–0.3 m lay-ers, regardless of genotype (Figure 3b).
Figure 2. Mean soil moisture content in the 0–0.4 m layer at 0.90m north or south of the ripline (tree row) under (a) ryegrass/clo-vers, (b) lucerne, and (c) bare ground treatments, in the LincolnUniversity agroforestry experiment, New Zealand. The arrow indi-cates the time of soil coring for fine root study.
Figure 3. Genotype × understorey (P = 0.046, Figure a) and geno-type × depth (P = 0.007, Figure b) interactions on pine FRLD inthe Lincoln University agroforestry experiment, New Zealand.Means with the same lowercase letters were not significantly dif-ferent between the genotypes within an understorey treatment (a),or depth (b). Means with the same uppercase letters were not sig-nificantly different between understorey treatments (a), or depth(b), within a genotype (n = 27).
93

Pine FRLD was greater in the ripline than in ei-ther side of the ripline (Figure 4a), indicating that lesspine roots grow to a distance 0.9 m away from thetree row because of root system structure and/or com-petition induced reduction in fine root growth. Theinteraction between understorey and position wascaused by a much smaller pine FRLD in the bareground than in the ryegrass/clovers treatment in theripline, whereas pine FRLD was not different amongthe pasture treatments on the south side (Figure 4a).On the north side, understorey competition signifi-cantly reduced pine FRLD.
The pine FRLD was influenced by sampling posi-tion in relation to tree row (P = 0.0003, ANOVA notshown), and was in the following order: ripline >south > north (Figure 4b). Pine FRLD in the 0.1–0.2or 0.2–0.3 m layers was much greater than that in the0–0.1 m layer in the ripline and on the south side, andgreater in the 0.1–0.2 m than in the 0–0.1 m layer on
the north side, giving rise to a significant depth × po-sition interaction (P = 0.013, ANOVA not shown).
Proportional pine root length density
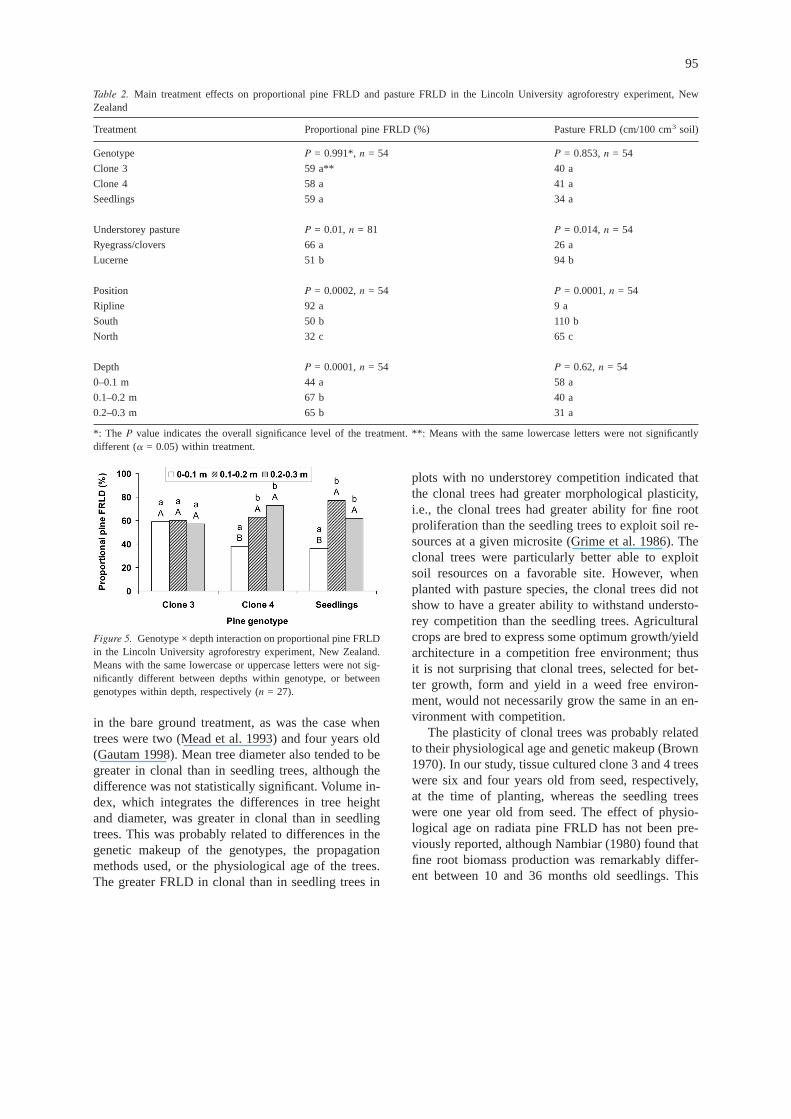
Understorey, sampling position, and depth affectedproportional pine FRLD (P = 0.01, 0.0002, and0.0001, respectively, Table 2). The genotype effectwas not significant but the genotype by depth inter-action was significant (P = 0.028, ANOVA notshown). The interaction between genotype and depthwas due to the similar proportional pine FRLD in allthree layers for clone 3 trees, whereas for clone 4 andseedling trees the proportional pine FRLD wasgreater in the 0.1–0.2 and 0.2–0.3 m than in the 0–0.1m soil layer (Figure 5).
The proportional pine FRLD was about 30%greater in the ryegrass/clovers than in the lucernetreatments (Table 2). Proportional pine FRLD wasgreater on the south side than on the north side, andwas greater in the ripline than in either the south ornorth sides.
Pasture root length density (FRLD)
Pasture FRLD was different between the two species(P = 0.014) and was affected by sampling position (P= 0.0001, Table 2), however, there was no significantinteraction between any of the factors studied. Pas-ture FRLD was greater in the lucerne than in theryegrass/clovers treatment, lucerne being a more pro-lific root producer and a deep rooting plant species.Pasture FRLD was greater on the south side than onthe north side of the trees, and was minimal in theripline where pasture growth had been controlled withherbicides. Pasture FRLD was not significantly af-fected by depth, although it tended to decrease withincreasing depth.
Discussion
Clonal trees have greater morphological plasticity
Fine root growth and distribution of trees have beenfound to be related directly to the amount of resourcesavailable to them, especially in competing situations(Nambiar 1984). This could result in differences inthe above ground growth of trees (Mead et al. 1993).At the time of this study, the clonal trees were tallerthan the seedling trees at the age of three, particularly
Figure 4. Understorey × position (a) and depth × position (b) in-teractions on pine FRLD. Means with the same lowercase letterswere not significantly different between the understorey (a), ordepths (b), within a position, in the Lincoln University agroforestryexperiment, New Zealand. Means with the same uppercase letterswere not significantly different between the positions within an un-derstorey treatment (a), or depth (b) (n = 27).
94
in the bare ground treatment, as was the case whentrees were two (Mead et al. 1993) and four years old(Gautam 1998). Mean tree diameter also tended to begreater in clonal than in seedling trees, although thedifference was not statistically significant. Volume in-dex, which integrates the differences in tree heightand diameter, was greater in clonal than in seedlingtrees. This was probably related to differences in thegenetic makeup of the genotypes, the propagationmethods used, or the physiological age of the trees.The greater FRLD in clonal than in seedling trees in
plots with no understorey competition indicated thatthe clonal trees had greater morphological plasticity,i.e., the clonal trees had greater ability for fine rootproliferation than the seedling trees to exploit soil re-sources at a given microsite (Grime et al. 1986). Theclonal trees were particularly better able to exploitsoil resources on a favorable site. However, whenplanted with pasture species, the clonal trees did notshow to have a greater ability to withstand understo-rey competition than the seedling trees. Agriculturalcrops are bred to express some optimum growth/yieldarchitecture in a competition free environment; thusit is not surprising that clonal trees, selected for bet-ter growth, form and yield in a weed free environ-ment, would not necessarily grow the same in an en-vironment with competition.
The plasticity of clonal trees was probably relatedto their physiological age and genetic makeup (Brown1970). In our study, tissue cultured clone 3 and 4 treeswere six and four years old from seed, respectively,at the time of planting, whereas the seedling treeswere one year old from seed. The effect of physio-logical age on radiata pine FRLD has not been pre-viously reported, although Nambiar (1980) found thatfine root biomass production was remarkably differ-ent between 10 and 36 months old seedlings. This
Table 2. Main treatment effects on proportional pine FRLD and pasture FRLD in the Lincoln University agroforestry experiment, NewZealand
Treatment Proportional pine FRLD (%) Pasture FRLD (cm/100 cm3 soil)
Genotype P = 0.991*, n = 54 P = 0.853, n = 54
Clone 3 59 a** 40 a
Clone 4 58 a 41 a
Seedlings 59 a 34 a
Understorey pasture P = 0.01, n = 81 P = 0.014, n = 54
Ryegrass/clovers 66 a 26 a
Lucerne 51 b 94 b
Position P = 0.0002, n = 54 P = 0.0001, n = 54
Ripline 92 a 9 a
South 50 b 110 b
North 32 c 65 c
Depth P = 0.0001, n = 54 P = 0.62, n = 54
0–0.1 m 44 a 58 a
0.1–0.2 m 67 b 40 a
0.2–0.3 m 65 b 31 a
*: The P value indicates the overall significance level of the treatment. **: Means with the same lowercase letters were not significantlydifferent (� = 0.05) within treatment.
Figure 5. Genotype × depth interaction on proportional pine FRLDin the Lincoln University agroforestry experiment, New Zealand.Means with the same lowercase or uppercase letters were not sig-nificantly different between depths within genotype, or betweengenotypes within depth, respectively (n = 27).
95

physiological age effect could have been confoundedby the genetic improvement of the genotypes (The-odorou et al. 1991). The growth and form (GF) rat-ings for clone 3 and 4 were 17 and 15, respectively,whereas the seedling trees were open pollinated witha GF rating of 14 (Mead et al. 1993). The geneticmakeup was found to have a positive effect on treeroot growth in both Scots and radiata pine, especiallyon sites with relatively high fertility and good mois-ture supply (Brown 1970; Theodorou et al. 1991).Difference in propagation methods could also haveresulted in differences in tree rooting behaviour(Sasse 1994 (unpublished); Ritchie et al. 1992). How-ever, we are unable to separate the effect of propaga-tion methods from the genetic effect in this study; thedifferences among the genotypes are the sum of alleffects.
The greater plasticity of clonal trees in fine rootproduction seemed to be an integral part of the growthpattern. In another study on those trees that examinedthe seasonal dynamics of root growth, the rate ofmaximum growth of fine roots was found to be higherin the clonal than in the seedling trees in the bareground treatment (Gautam 1998). Fine root growthrate of the seedling trees declined more rapidly thanthat of the clonal trees after reaching the point ofmaximum growth in a growing season, which resultedin 65% more fine root biomass in clone 3 than inseedling trees in the bare ground treatment at the endof the growing season (Gautam 1998). This showsthat results from this one time sampling of fine rootswere consistent with those obtained from a yearlongstudy of fine root dynamics.
Growth habit and competition may explain fine rootdistribution
Foliage density was significantly lower in clone 3than in seedling trees, and clone 3 trees had longerinternodes than seedling trees in the bare ground plots(Bandara 1997 (unpublished)). This crown architec-ture of the clonal trees allowed more solar radiationto penetrate through the tree crown to reach tree fo-liage than that of the seedling trees (Yunusa et al.1995a), which resulted in higher foliage efficiency inthe clonal trees (Bandara 1997 (unpublished)). There-fore, it could be hypothesized that there may be lesscarbon allocated to foliage in the clonal than in theseedling trees, and as a consequence the supply ofcarbohydrate from the source to root may be higherin the clonal than in the seedling trees. Such an ex-
planation may account for the higher FRLD in clonalthan in seedling trees under reduced competition lev-els. The higher FRLD in clonal trees allowed a greatervolume of soil to be exploited for water and nutrients.The higher FRLD in clonal than in seedling trees inthe 0–0.1 m layer, a layer subjected to greater evapo-ration, further demonstrated the vigorous growthhabit of roots of the clonal trees.
With the seedling trees, fine roots were mostlyfound between the 0.1 and 0.3 m depth. Similar re-sults were reported for young radiata pine stands inSouth Australia (Nambiar 1983) and in an agrofor-estry experiment on Canterbury Plains, New Zealand(Clinton 1990 (unpublished)). There are two possiblebroad reasons for this trend: (a) pasture rooting com-petition is higher in the 0–0.1 m soil layer as indi-cated by a higher pasture FRLD in that layer (Ta-ble 2); or (b) soil moisture content was lower in the0–0.1 m than in the other layers (Gautam 1998).Competition for the often limited soil moisture sup-ply played an important role in this distribution pat-tern. For example, understorey competition reducedpine FRLD on the north side of tree rows (in the rainshadow) more so than on the south side (Figure 4a).
A lower proportional pine FRLD in the 0–0.1 mlayer (Figure 5) indicated that the magnitude of pas-ture root competition with pine roots (clone 4 andseedling trees) was higher in the 0–0.1 m than in the0.1–0.2 and 0.2–0.3 m soil layers. The proportionalpine FRLD for clone 3 was similar in all layers, sug-gesting that clone 3 was less affected by competitionfrom pasture species than clone 4 or the seedling trees(Figure 5). Furthermore, the lower proportional pineFRLD on the north than on the south side (Table 2)and the greater proportional pasture root on the northside, indicated that the relative pasture rooting com-petition for trees was higher on the drier north thanon the wetter south side. This strongly supports thesecond hypothesis that under limited resource avail-ability there will be greater underground competitionbetween the components in agroforestry systems. Thegreater competition might have resulted in lowercoarse root biomass production and less fine rootlength elongation of radiata pine on the north than onthe south side (Gautam 1998). This correlates with thelower soil moisture content on the north than on thesouth side (Figure 2), resulting mostly from southerlyrain-bearing winds (Figure 1b) and the greater solarradiation on the north side in the southern hemi-sphere.
96

Microsite conditions affect root distribution
Plant root systems proliferate in favorable micrositessuch as when they encounter localized water pockets(Grime et al. 1986). Similarly, this study showed thatthe effect of understorey pasture competition on treeFRLD was related to microsite conditions (i.e., posi-tion in relation to tree rows). Tree FRLD was greaterin the ripline than on either side of the ripline, par-ticularly in the ryegrass/clovers and lucerne plots.This greater FRLD resulted from the absence of un-derstorey competition in the ripline as compared witheither side of the ripline. In addition to the lack ofcompetition, soil cultivation in the ripline may havealso enhanced the growth of pine fine roots (Figure4a). Understorey competition with trees was greaterin the lucerne than in the ryegrass/clovers treatmentsas indicated by a lower proportional tree FRLD in thelucerne than in the ryegrass/clovers treatment (Ta-ble 2) and a reversed relationship for FRLD of pas-ture species (Table 2). This indicated that there was ahigher competition for soil moisture in the lucernethan in the ryegrass/clovers treatment (Yunusa et al.1995a).
The wetter and cooler soil condition on the southside provided a more favourable growth media forradiata pine root production in the summer (Heth1980) and thus greater tree rooting density on thesouth side. Similarly, the pasture rooting density onthe north side was lower than that on the south side(Table 2) which agrees with Yunusa et al. (1995b),who found less pasture production on the north thanon the south side of tree rows. This lower pasturerooting density was also related to the lower soilmoisture content (Figure 2) on the north than on thesouth side (Yunusa et al. 1995a).
Conclusions
This study demonstrated that, in a site with substan-tial soil moisture deficit in the summer, understoreycompetition belowground could significantly affecttree root growth. Compared to the ryegrass/cloversunderstorey, lucerne was more competitive for soilresources to affect radiata pine fine root production.As a result, greater reduction was found in tree above-ground growth in the lucerne than in the ryegrass/clovers plots. Hence, selection of understorey typeshould be an important consideration when the goalis to optimise tree growth under agroforestry condi-
tions. Certain radiata genotypes have greater morpho-logical plasticity and are better able to exploit soilresources particularly under more favourable condi-tions. Radiata pine genotypes with superior rootinghabit seem to be better able to compete with under-storey for below ground resources, and this will min-imise the effect of understorey competition on treegrowth in agroforestry systems.
Acknowledgements
The authors would like to thank Prof Richard Row,Dr Joanna Sasse, and Dr Isa Yunusa, of Lincoln Uni-versity, New Zealand, and Prof N. B. Comerford, ofthe University of Florida, USA, for their input intothis work. We appreciate the comments from the twoanonymous reviewers.
References
Anderson L.S. and Sinclair F.L. 1993. Ecological interactions inagroforestry systems. Agrofor Abst 6: 57–91.
Ares A. and Peinemann N. 1992. Fine-root distribution of conifer-ous plantations in relation to site in southern Buenos Aires, Ar-gentina. Can J For Res 22: 1575–1582.
Avery T.E. and Burkhart H.E. 1983. Forest Measurements. 3rd edn.McGraw Hill Publishing Company, New York, 331 pp.
Bandara G. 1997. Ecophysiology of Clonal and Seedling Trees ofPinus radiata D. Don in an Agroforestry System. PhD Disser-tation, Lincoln University, New Zeland (unpublished).
Bi H., Turvey N.D. and Heinrich P. 1992. Rooting density and treesize of Pinus radiata (D. Don) in response to competition fromEucalyptus obliqua (L’ Herit). For Ecol Manage 49: 31–42.
Brown J.H. 1970. Seedling growth of three Scotch pine prove-nances with varying moisture and fertility treatments. For Sci16: 43–45.
Chang S.X., Amatya G., Beare M.H. and Mead D.J. 2001. Soilproperties under a Pinus radiata-ryegrass silvopastoral systemin New Zealand. Part 1. Soil N and moisture availability, soilC, and tree growth. Agrofor Syst 54: 137–147.
Clinton P.W. 1990. Competition for nitrogen and moisture in a Pi-nus radiata-pasture agroforestry system. PhD Dissertation,University of Canterbury, New Zealand (unpublished).
Eastham J. and Rose C.W. 1990. Tree/pasture interactions at arange of tree densities in an agroforestry experiment. I. Root-ing patterns. Australian J Agri Res 41: 683–695.
Gautam M.K. 1998. Rooting characteristics of Pinus radiata D.Don as influenced by understorey conmpetition in an agrofor-estry system. PhD Dissertation, Soil, Plant and Ecological Sci-ences Division, Lincoln University.
97

Grime J.P., Crick J.C. and Rincon J.E. 1986. The ecological sig-nificance of plasticity. In: Jennings D.H. and Trewavas A.J.(eds), Plasticity in Plants. Cambridge University Press, pp.5–29.
Hendrick R.L. and Pregitzer K.S. 1993. Patterns of fine root mor-tality in two sugar maple forests. Nature 361: 59–61.
Heth D. 1980. Root and shoot water potential in stressed pineseedlings. NZ J For Sci 10: 142–147.
Hughes K.A., Gandar P.W., Menalda P.H. and Snow V.O. 1986. ASurvey of Kiwi Fruit Root Systems. Technical Report No. 22.Plant Physiology Division, DSIR, Palmerston North, NewZealand.
Jonsson K., Fidjeland L., Maghenbe J.A. and Hogberg P. 1988. Thevertical distribution of five tree species and maize in Morogoro,Tanzania. Agrofor Syst 6: 63–69.
Mead D.J., Lucus R.J. and Mason E.G. 1993. Studying interactionsbetween pastures and Pinus radiata in Canterbury’s sub-humidtemperate environment – the first two years. NZ For 38: 26–31.
Nambiar E.K.S. 1980. Root configuration and root regeneration inPinus radiata seedlings. NZ J For Sci 10: 249–263.
Nambiar E.K.S. 1983. Root development and configuration in in-tensively managed radiata pine plantations. Plant Soil 71: 37–47.
Nambiar E.K.S. 1984. Significance of first order lateral roots onthe growth of young radiata pine under environmental stress.Australian For Res 14: 187–199.
Ong C.K. 1991. Interaction of light, water and nutrients in agro-forestry systems. In: Avery M.E., Cannell M.G.R. and OngC.K. (eds), Biophysical Research for Asian Agroforestry. Win-rock International, USA, pp. 107–124.
Richards D., Goubran F.H., Garwoli W. and Daly M.W. 1979. Amachine for determining root length. Plant Soil 52: 69–76.
Ritchie G.A., Tanaka Y. and Duke S.D. 1992. Physiology and mor-phology of Douglas-fir rooted cuttings compared to seedlingsand transplants. Tree Physiol 10: 179–194.
Sasse J. 1994. Comparisons of the morphology and physiology ofcuttings and seedlings of Eucalyptus globulus. PhD Disserta-tion, University of Melbourne, Australia (unpublished).
Stokes A., Nicoll B.C., Coutts M.P. and Fitter A.H. 1997. Responseof young Sitka spruce clones to mechanical perturbation andnutrition: effects on biomass allocation, root development, andresistance to bending. Can J For Res 27: 1049–1057.
Theodorou C. and Bowen G.D. 1993. Root morphology, growth,and uptake of phosphorous and nitrogen of Pinus radiata fami-lies in different soils. For Ecol Manage 56: 43–56.
Theodorou C., Cameron J.N. and Bowen G.D. 1991. Growth ofroots of different Pinus radiata genotypes in soil at differentstrength and aeration. Australian For 54: 52–59.
Vogt K.A. and Persson H. 1991. Measuring growth and develop-ment of roots. In: Lassoie J.P. and Hinckley T.M. (eds), Tech-niques and Approaches in Forest Tree Ecophysiology. CRCPress, USA, pp. 477–502.
Yunusa I.A.M., Mead D.J., Pollock K.M. and Lucas R.J. 1995a.Process studies in a Pinus radiata-pasture agroforestry systemin a subhumid temperate environment. I. Water use and lightinterception in the third year. Agrofor Syst 32: 163–183.
Yunusa I.A.M., Mead D.J., Pollock K.M. and Lucas R.J. 1995b.Process studies in a Pinus radiata-pasture agroforestry systemin a subhumid temperate environment. II. Analysis of dry mat-ter yield in the third year. Agrofor Syst 32: 185–204.
98
Copyright © 2022 FDOKUMEN




















