
The omnivorous collared peccary negates an insectivore-generated trophic cascade in Costa Rican wet...
12
Journal of Tropical Ecology http://journals.cambridge.org/TRO Additional services for Journal of Tropical Ecology: Email alerts: Click here Subscriptions: Click here Commercial reprints: Click here Terms of use : Click here The omnivorous collared peccary negates an insectivore-generated trophic cascade in Costa Rican wet tropical forest understorey Nicole L. Michel, Thomas W. Sherry and Walter P. Carson Journal of Tropical Ecology / Volume 30 / Issue 01 / January 2014, pp 1 - 11 DOI: 10.1017/S0266467413000709, Published online: 11 November 2013 Link to this article: http://journals.cambridge.org/abstract_S0266467413000709 How to cite this article: Nicole L. Michel, Thomas W. Sherry and Walter P. Carson (2014). The omnivorous collared peccary negates an insectivore- generated trophic cascade in Costa Rican wet tropical forest understorey. Journal of Tropical Ecology, 30, pp 1-11 doi:10.1017/S0266467413000709 Request Permissions : Click here Downloaded from http://journals.cambridge.org/TRO, IP address: 71.17.241.253 on 16 Dec 2013
Transcript of The omnivorous collared peccary negates an insectivore-generated trophic cascade in Costa Rican wet...

Journal of Tropical Ecologyhttp://journals.cambridge.org/TRO
Additional services for Journal of Tropical Ecology:
Email alerts: Click hereSubscriptions: Click hereCommercial reprints: Click hereTerms of use : Click here
The omnivorous collared peccary negates an insectivore-generatedtrophic cascade in Costa Rican wet tropical forest understorey
Nicole L. Michel, Thomas W. Sherry and Walter P. Carson
Journal of Tropical Ecology / Volume 30 / Issue 01 / January 2014, pp 1 - 11DOI: 10.1017/S0266467413000709, Published online: 11 November 2013
Link to this article: http://journals.cambridge.org/abstract_S0266467413000709
How to cite this article:Nicole L. Michel, Thomas W. Sherry and Walter P. Carson (2014). The omnivorous collared peccary negates an insectivore-generated trophic cascade in Costa Rican wet tropical forest understorey. Journal of Tropical Ecology, 30, pp 1-11doi:10.1017/S0266467413000709
Request Permissions : Click here
Downloaded from http://journals.cambridge.org/TRO, IP address: 71.17.241.253 on 16 Dec 2013

Journal of Tropical Ecology (2014) 30:1–11. © Cambridge University Press 2013doi:10.1017/S0266467413000709
The omnivorous collared peccary negates an insectivore-generated trophiccascade in Costa Rican wet tropical forest understorey
Nicole L. Michel∗,†,1, Thomas W. Sherry† and Walter P. Carson‡
∗ School of Environment and Sustainability, University of Saskatchewan, 117 Science Place, Saskatoon, SK S7N 5C8, Canada† Department of Ecology and Evolutionary Biology, Tulane University, 400 Boggs Hall, 6823 St. Charles Avenue, New Orleans, LA 70118, USA‡ Department of Biological Sciences, University of Pittsburgh, 154A Crawford Hall, Pittsburgh, PA 15260, USA
(Received 27 May 2013; revised 11 October 2013; accepted 11 October 2013; first published online 11 November 2013)
Abstract: Insectivorous birds and bats often protect plants through density- and trait-mediated cascades, but the degreeto which insectivores reduce herbivorous arthropods and leaf damage varies among systems. Top-down interactionstrength may be influenced by the biotic and abiotic context, including the presence of vegetation-disturbing animals.We tested two hypotheses: (1) insectivorous birds and bats initiate trophic cascades in tropical rain-forest understorey;and (2) the native, omnivorous collared peccary (Pecari tajacu) negates these cascades via non-trophic effects. Westudied the top-down effects of birds and bats on understorey plants in north-eastern Costa Rica using 60 nettedexclosures within and outside existing peccary exclosures. Excluding birds and bats increased total arthropod densitiesby half, both with and without peccaries. Bird/bat exclosures increased Diptera density by 28% and leaf damage by24% without peccaries, consistent with a trophic cascade. However, bird/bat exclosures decreased Diptera density by32% and leaf damage by 34% with peccaries, a negation of the trophic cascade. Excluding peccaries increased leafdamage by 43% on plants without birds and bats. This is the first study, to our knowledge, to demonstrate that thenon-trophic activity of an omnivorous ungulate can reverse a trophic cascade.
Key Words: density-mediated, herbivory, insectivory, La Selva Biological Station, non-trophic effects, Pecari tajacu,trait-mediated, trophic cascade, trophic downgrading, ungulates
INTRODUCTION
Insectivores, particularly birds and bats, in natural(i.e. non-agricultural) tropical forests initiate top-downdensity- and trait-mediated trophic cascades that protectplants by reducing leaf damage from herbivorousarthropods (Dyer et al. 2012, Kalka et al. 2008, Mantylaet al. 2011, Mooney et al. 2010, Schmitz et al. 2004).Trophic cascade strength varies considerably due tovarying biotic and abiotic contexts (Agrawal et al. 2007).Most studies of context-dependence to date have focusedon direct trophic linkages to one or more participants inthe trophic cascade, for example, identity of predators (e.g.bats or birds); intraguild predation; and plant productivity(Borer et al. 2005, Kalka et al. 2008, Mooney et al. 2010,Van Bael & Brawn 2005, Vance-Chalcraft et al. 2007).
However, trophic cascades occur within largerbiological communities, and participants interact with
1 Corresponding author. Email: [email protected]
many other organisms not typically considered inresearch on trophic cascades (Kefi et al. 2012). Forexample, ungulates such as pigs (Suidae) and peccaries(Tayassuidae) may mediate trophic dynamics throughdisturbance to forest understorey (Queenborough et al.2012). While these omnivores do consume some plantmaterial, their greatest effects on plants are non-trophic,via disturbances such as trampling and rooting thatalter microhabitats and microclimates and, potentially,arthropod predator abundance (Beck 2005, Ickeset al. 2001, Queenborough et al. 2012). Consequently,ungulates may reduce the abundance or foragingintensity of arthropod herbivores and leaf damage (Coteet al. 2004, Stewart 2001). Ungulates may also increaseabundance of other vertebrate insectivores through theireffects on litter depth, complexity and nutrient cycling. Forexample, peccaries increased the abundance of leaf-litterherpetofauna relative to experimental peccary exclosuresat La Selva Biological Station, Costa Rica (Reider et al.2013). Consequently, ungulates could weaken, negate oreven reverse the direction of the trophic cascade through

2 NICOLE L. MICHEL, THOMAS W. SHERRY AND WALTER P. CARSON
primarily non-trophic interactions. Thus, weaker trophiccascades in wet tropical forest understorey may be dueto abundant ungulates rather than, or in addition to,lower productivity or a more diverse producer trophiclevel (Boege & Marquis 2006, Van Bael & Brawn 2005,Van Bael et al. 2003).
The wild pig (Sus scrofa) and peccaries, including theomnivorous collared peccary (Pecari tajacu), have in-creased in abundance in some fragmented tropical forests,due at least in part to apex predator loss and increasedfood availability from nearby farms (Beck 2005, Ickes2001, Michel & Sherry 2012, Terborgh & Estes 2010,Timm 1994). Here, we evaluate the non-trophic effectsof the collared peccary on an insectivorous bird/bat–arthropod–understorey plant trophic cascade in natural(i.e. non-agricultural) tropical forest. We tested twointerdependent hypotheses. First, insectivorous birds andbats cause trophic cascades by reducing the abundanceor changing the behaviour of insect herbivores, therebyindirectly causing a decrease in plant damage. Second,the collared peccary weakens, negates, or reverses thedirection of these cascades. We experimentally testedthese two hypotheses by excluding insectivorous birdsand bats from a diverse group of understorey plant speciesboth within and outside of large peccary exclosures.
METHODS
Study site
We conducted this research at La Selva Biological Station,Costa Rica (10°26ʹN, 83°59ʹW, 35–130 m asl), a 1611-ha lowland rain-forest reserve receiving 4 m of rainannually, with a February–April dry season (Sanfordet al. 1994). The collared peccary is highly abundantat La Selva (Kuprewicz 2013), and most other large,terrestrial vertebrates are extirpated or rare (e.g. white-lipped peccary (Tayassu pecari), Baird’s tapir (Tapirusbairdii); Timm 1994).
Design of mammal and bird/bat exclosures
We established five paired 20 × 50-m fenced exclosureand control plots ( = five blocks) within old-growth forestin 1999. These exclosures allow entry to small and mid-sized mammals (e.g. Central American agouti, Dasyproctapunctata) while only excluding peccaries (Reider et al.2013); hereafter we refer to these as peccary exclosures.We subdivided each peccary exclosure or control plotinto 40, 5 m × 5-m subplots, following a split-split plotrandomized complete block design. In June 2007, threeof these subplots were randomly assigned to contain net-covered exclosures excluding birds and bats, and three as
controls (N = 60 total). We constructed 8-m3 (2 × 2 × 2m) bird/bat exclosures using plastic bird-control netting(3 × 3.5 cm mesh) stretched across four PVC poles tiedback to nearby trees and staked to the ground (followingBoege & Marquis 2006). These 8-m3 bird/bat exclosuresare hereafter referred to as netted exclosures. This meshsize excludes all understorey insectivorous birds (exceptperhaps Myiornis atricapillus, a small canopy-subcanopyforager; T. Sherry unpubl. data), bats, and other mid-sizeand large insectivores (e.g. mouse opossums, Marmosaspp.), while allowing entry by small insectivorous lizards,frogs and all but the largest arthropods (Boege & Marquis2006, Van Bael & Brawn 2005). While the netting couldprovide additional substrate for orb-weaving spiders, weobserved few orb-weavers using the netting, and removedthose we did find. We did not use procedural controls(open-sided exclosures to control for net effects) sinceprior studies found that the chosen mesh type and sizereduces light by <1% and has no impact on microclimaticvariables such as temperature or rainfall (Marquis &Whelan 1994, Van Bael et al. 2003). All leaf litteraccumulated on top of the netted exclosures was removedand placed inside the exclosures, and any tears in thenetting were repaired at each visit. Accumulated leaf littercould reduce light and, consequently, photosynthetic ratebetween monthly removals, and netting tears (thoughrare) could allow birds and bats temporary access, thusweakening trophic cascade strength. However, becausethe netting effects should be consistent across peccaryexclosure treatments, litter accumulation and temporarygaps in the netting should not influence the key questionof how peccaries affect trophic cascades.
Each netted exclosure or control was centred on asingle focal plant 0.5–2 m tall and possessing at least 20leaves. This size range was determined by the size of theexclosures (2 × 2 × 2 m), and the need for a minimumof 20 leaves for leaf area and damage assessments. Weincluded a total of 14 species from 12 families and fiveclades (Angiosperm Phylogeny Group III 2009; Appendix1). Our peccary exclosure and control locations, coupledwith our randomized sampling design in tropical rain-forest understorey, necessitated using more diverse shrubspecies than previous studies (Mantyla et al. 2011), buthad the advantage of allowing us to evaluate whethertrophic cascades occurred across phylogenetically diversetaxa. Focal plants were chosen using a stratified randomsampling design; within each randomly selected 5 × 5-m subplot we chose a plant from one of the focal speciesselected for their relative frequency and appropriate size.
Arthropod surveys
We visually surveyed all leaves and stems of each focalplant for arthropods immediately prior to constructing

Peccaries negate bird/bat–insect–plant cascade 3
netted exclosures (June 2007) and again in July 2007,October 2007 and April 2008. All arthropods wereidentified to order and family (where possible, e.g.Coleoptera, Hemiptera), and assigned to one of fourforaging guilds (predator, phloem-feeding herbivore,chewing herbivore and omnivore; Triplehorn & Johnson2004). We calculated arthropod densities as numberof individuals per unit leaf area for each survey. Westudied arthropod predators because of their potentiallyimportant role in the trophic cascade as intraguildpredators (Mooney et al. 2010, Vance-Chalcraft et al.2007).
Leaf-damage and area assessments
We assessed plant damage by determining missing leafarea using a 0.25-mm2 transparent plastic grid, andmeasuring leaf length and width on 20 randomly selectedleaves per focal plant, including both young and oldleaves (Boege & Marquis 2006). Original, undamaged leafarea was calculated using leaf-area regression equationsderived from leaf width and length for 100 undamagedleaves per species. We then estimated damage as percent area missing using the ratio of total missing areato original, undamaged leaf area. We calculated total leafarea per plant by multiplying mean extant leaf area timesthe number of leaves on the plant. Total leaf area andper cent damage were calculated in June and October2007 and April 2008. Finally, we visually surveyedeach leaf for signs of mammalian browsing, particularlywhether whole or partial leaves had been eaten orremoved.
Data analysis
We separately analysed densities of all arthropods,predatory arthropods, chewing herbivores, phloem-feeding herbivores, and the six most common arthropodorders (Hemiptera, Araneae, Hymenoptera, Diptera,Lepidoptera and Orthoptera), as well as per cent leafdamage, using repeated-measures generalized linearmixed models (Proc GLIMMIX, SAS 9.3, SAS Institute,Cary, NC, USA). Arthropod density and leaf damageestimates were averaged across the three replicatenetted exclosure or control subplots within each peccaryexclosure and control plot, resulting in N = 20.Replicates were averaged to avoid pseudoreplication,but as replicates included multiple plant species, thisprevented the inclusion of plant as a random effect inthe analysis. Peccary exclosure treatment (α), bird/batexclosure treatment (β), time (τ ) and all two- andthree-way interactions were fixed factors. Random effectsincluded an intercept for each block (δ) and slopes
for all two-, three- and four-way interactions betweenblock, peccary exclosure, bird/bat exclosure and time.GLIMMIX calculates a single, composite error term (εijklmn)incorporating all whole plot and subplot error usingrandom effects. Denominator degrees of freedom werecalculated using the between-within method, whichdivides residual degrees of freedom into between-subjectand within-subject components (SAS Institute 2009). Thecomplete model was
y = a + αi + β j + (αβ)i j + τk + (ατ )i k + (βτ ) j k
+ (αβτ )i j k + δl + (aδ)i l + (βδ) j l + (aβδ)i j l
+ (aτδ) j kl + (βτδ)i j k + (αβτδ)i j kl + εi j klmn,
where a represents the fixed-effect intercept.We included initial density and initial leaf area as
covariates for all arthropod density analyses. All two- andthree-way interactions between covariates and bird/batand peccary exclosure treatments were non-significant,and were removed from subsequent analyses. For the leafdamage analysis we included initial leaf area and initialleaf damage as covariates; mean predator and chewingherbivore densities were also included as covariates to testwhether bird/bat- or peccary-exclosure effects persistedafter controlling for arthropod densities (Shipley 2009).Pseudo-information criteria (Pseudo-AICc and Pseudo-BIC) were used to select the optimal distribution and link(Dick 2004). Significant bird/bat × peccary treatmentinteractions were identified using two-tailed tests of leastsquares means with Tukey–Kramer HSD adjustments formultiple comparisons (Littell et al. 2006).
RESULTS
Peccary exclusion effects
Peccary exclusion effects on all arthropods variedover time (peccary × time interaction), significantlyincreasing total arthropod density only in the first monthpost-exclosure (July 2007; Table 1, Figure 1a). Peccaryexclusion effects on predators and spiders (Araneae) alsovaried over time (peccary × time interactions; Tables 1,2). Peccary exclusion increased densities of arthropodpredators – 94% of which were spiders – twofold in Julyand by half in October, but reduced arthropod predatordensities by 42% in April (Figure 1b). Peccary exclusionincreased mean Hymenoptera densities by 73% on plantsboth with and without birds and bats (mean ± SE,peccary exclosure: 3.7 ± 0.6 m−2, peccary control: 2.1± 0.3 m−2; Table 2). Peccary exclusion reduced Dipteradensities by 31% on plants with birds and bats, but didnot affect Diptera densities on plants without birds andbats (bird/bat × peccary interaction; Table 2, Figure 2).Peccary exclusion reduced per cent leaf damage by 27%

4 NICOLE L. MICHEL, THOMAS W. SHERRY AND WALTER P. CARSON
Table 1. Results of repeated measures generalized linear mixed models used to test responses of total arthropod density, predator density, phloem-feeding and chewing herbivore density, and leaf damage (per cent leaf area missing) in a factorial experiment with two levels of bird/bat treatment(exclosure and control), two levels of peccary treatment (exclosure and control), and four sampling periods over a 10-mo period (June, July, andOctober 2007 and April 2008) in tropical rain-forest understorey at La Selva Biological Station, Costa Rica. ∗ P < 0.05; ∗∗ P < 0.01; ∗∗∗ P <
0.001; ∗∗∗∗ P � 0.0001.
Total arthropods Predators Phloem-feeding herbivores Chewing herbivores Leaf damage
Fixed effects F df F df F df F df F df
Bird/bat exclosure 13.7∗∗ 1,14 0.32 1,14 3.17 1,14 2.83 1,14 0.97 1,14Peccary exclosure 4.42 1,14 5.61∗ 1,14 0.21 1,14 0.14 1,14 0.16 1,14Bird/bat × peccary 0.08 1,14 3.104 1,14 0.09 1,14 1.11 1,14 9.3∗∗ 1,14Time 57.7∗∗∗∗ 2,32 37.0∗∗∗∗ 2,32 33.8∗∗∗∗ 2,32 28.5∗∗∗∗ 2,32 3.88 1,14Bird/bat × time 1.70 2,32 2.25 2,32 3.77∗ 2,32 1.03 2,32 0.02 1,14Peccary × time 4.12∗ 2,32 7.31∗∗∗ 2,32 0.48 2,32 0.49 2,32 0.59 1,14Bird/bat × peccary × time 0.99 2,32 0.40 2,32 1.16 2,32 0.46 2,32 0.17 1,14
CovariatesInitial density (damage) 1.27 1,14 0.78∗ 1,14 1.32 1,14 1.13 1,14 6.48∗ 1,14Initial leaf area 13.27∗∗ 1,14 0.22 1,14 8.73∗ 1,14 7.10∗ 1,14 0.25 1,14Predator density 6.48∗ 1,14Chewing herbivore density 4.31 1,14
Table 2. Results of repeated measures generalized linear mixed models used to test responses of densities of the most common arthropod orders(Araneae, Diptera, Hemiptera, Hymenoptera, Lepidoptera and Orthoptera) in a factorial experiment with two levels of bird/bat treatment (exclosureand control), two levels of peccary treatment (exclosure and control), and four sampling periods over a 10-mo period (June, July and October 2007and April 2008) in rain-forest understorey at La Selva Biological Station, Costa Rica. ∗ P < 0.05; ∗∗ P < 0.01; ∗∗∗ P < 0.001; ∗∗∗∗ P � 0.0001.
Araneae Diptera Hemiptera Hymenoptera Lepidoptera Orthoptera
Fixed effects F df F df F df F df F df F df
Bird/bat exclosure 0.26 1,14 0.35 1,14 1.06 1,14 2.99 1,14 1.22 1,14 0.00 1,14Peccary exclosure 3.67 1,14 0.23 1,14 0.13 1,14 5.48∗ 1,14 1.01 1,14 0.00 1,14Bird/bat × peccary 2.12 1,14 8.43∗ 1,14 0.04 1,14 0.03 1,14 0.11 1,14 0.75 1,14
Time 36∗∗∗∗ 2,32 107∗∗∗∗ 2,32 29∗∗∗∗ 2,32 7.2∗∗ 2,32 0.72 2,32 36∗∗∗∗ 2,32Bird/bat × time 3.05 2,32 0.02 2,32 4.66∗ 2,32 0.43 2,32 1.89 2,32 0.40 2,32Peccary × time 7.27∗∗ 2,32 0.02 2,32 1.02 2,32 0.82 2,32 0.56 2,32 0.62 2,32Bird/bat × peccary × time 0.42 2,32 1.42 2,32 1.22 2,32 0.03 2,32 0.57 2,32 2.91 2,32
CovariatesInitial density 6.95 1,14 0.03 1,14 2.04 1,14 10.03∗∗ 1,14 6.33∗ 1,14 3.96 1,14Initial leaf area 0.42 1,14 1.02 1,14 5.41∗ 1,14 0.48 1,14 1.04 1,14 0.6 1,14
on plants with birds and bats, but increased leaf damage by57% on plants without birds and bats (bird/bat × peccaryinteraction; Table 1, Figure 3).
Bird and bat exclusion effects
Excluding birds and bats, with or without peccaries,increased total arthropod density by 52% (mean ± SE,bird/bat exclosure: 31.9% ± 2.5%, bird/bat control:20.9% ± 1.6%; Table 1, Figure 4). Bird and bat exclusioneffects on phloem-feeding herbivore and Hemipteradensities varied over time, and were unaffected bypeccary presence or absence (bird/bat × time interaction;Tables 1, 2). Bird and bat exclusion doubled phloem-feeding herbivore density – 91% of which were in the orderHemiptera – only in the first month post-exclosure (July;Tables 1, 2; Figure 5). Bird and bat exclusion had no effectson arthropod predators or chewing herbivores (Table 1).
However, bird and bat exclusion increased Diptera densityby 28% without peccaries, but decreased Diptera densityby 32% with peccaries (bird/bat × peccary interaction;Table 2, Figure 2).
Excluding birds and bats increased per cent leaf damageby 35% without peccaries, consistent with a trophiccascade (bird/bat × peccary interaction; Table 1, Figures3a, b). However, bird and bat exclusion reduced leafdamage by 38% with peccaries, contrary to a trophiccascade (Figures 3a, b). Whereas per cent leaf damageincreased by 49% during the experiment when birds andbats were excluded without peccaries, leaf damage onlynominally increased when birds and bats were excludedwith peccaries. Across all treatments, an increase inarthropod predator density led to reduced leaf damage(predators explained a significant proportion of thevariation in leaf damage, F1,14 = 6.5, P = 0.02) whilechewing-herbivore density did not explain any variationin leaf damage (F1,14 = 4.3, P = 0.06).
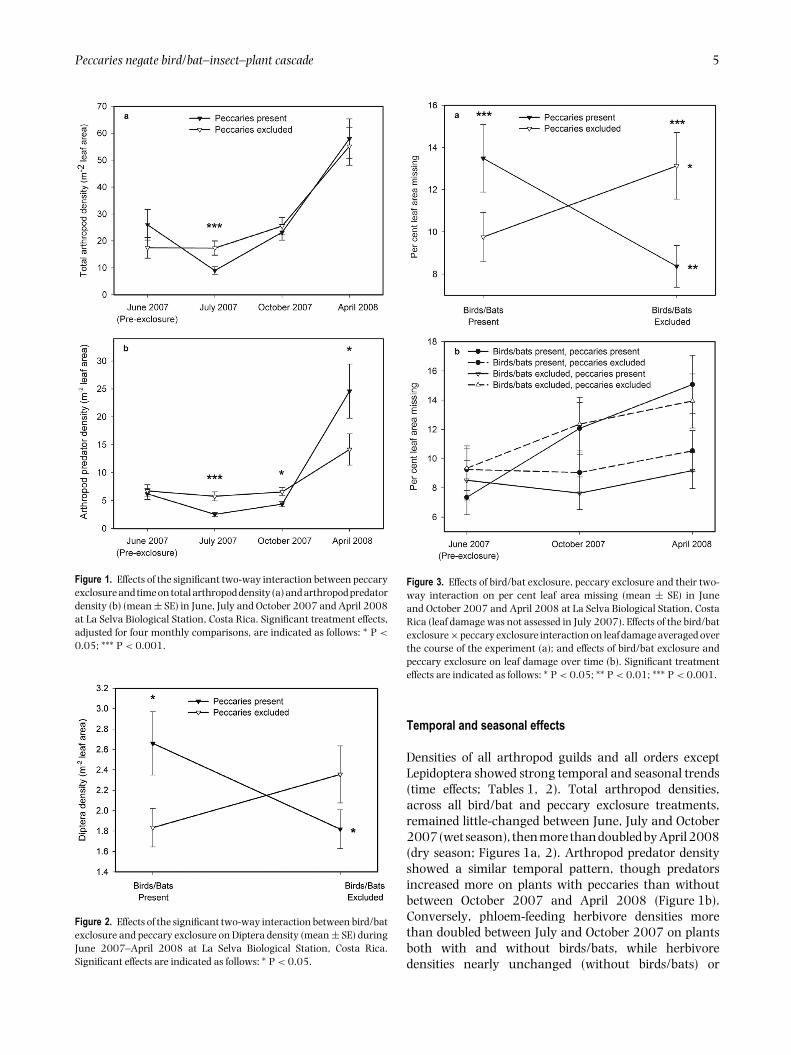
Peccaries negate bird/bat–insect–plant cascade 5
Figure 1. Effects of the significant two-way interaction between peccaryexclosure and time on total arthropod density (a) and arthropod predatordensity (b) (mean ± SE) in June, July and October 2007 and April 2008at La Selva Biological Station, Costa Rica. Significant treatment effects,adjusted for four monthly comparisons, are indicated as follows: ∗ P <
0.05; ∗∗∗ P < 0.001.
Figure 2. Effects of the significant two-way interaction between bird/batexclosure and peccary exclosure on Diptera density (mean ± SE) duringJune 2007–April 2008 at La Selva Biological Station, Costa Rica.Significant effects are indicated as follows: ∗ P < 0.05.
Figure 3. Effects of bird/bat exclosure, peccary exclosure and their two-way interaction on per cent leaf area missing (mean ± SE) in Juneand October 2007 and April 2008 at La Selva Biological Station, CostaRica (leaf damage was not assessed in July 2007). Effects of the bird/batexclosure × peccary exclosure interaction on leaf damage averaged overthe course of the experiment (a); and effects of bird/bat exclosure andpeccary exclosure on leaf damage over time (b). Significant treatmenteffects are indicated as follows: ∗ P < 0.05; ∗∗ P < 0.01; ∗∗∗ P < 0.001.
Temporal and seasonal effects
Densities of all arthropod guilds and all orders exceptLepidoptera showed strong temporal and seasonal trends(time effects; Tables 1, 2). Total arthropod densities,across all bird/bat and peccary exclosure treatments,remained little-changed between June, July and October2007 (wet season), then more than doubled by April 2008(dry season; Figures 1a, 2). Arthropod predator densityshowed a similar temporal pattern, though predatorsincreased more on plants with peccaries than withoutbetween October 2007 and April 2008 (Figure 1b).Conversely, phloem-feeding herbivore densities morethan doubled between July and October 2007 on plantsboth with and without birds/bats, while herbivoredensities nearly unchanged (without birds/bats) or
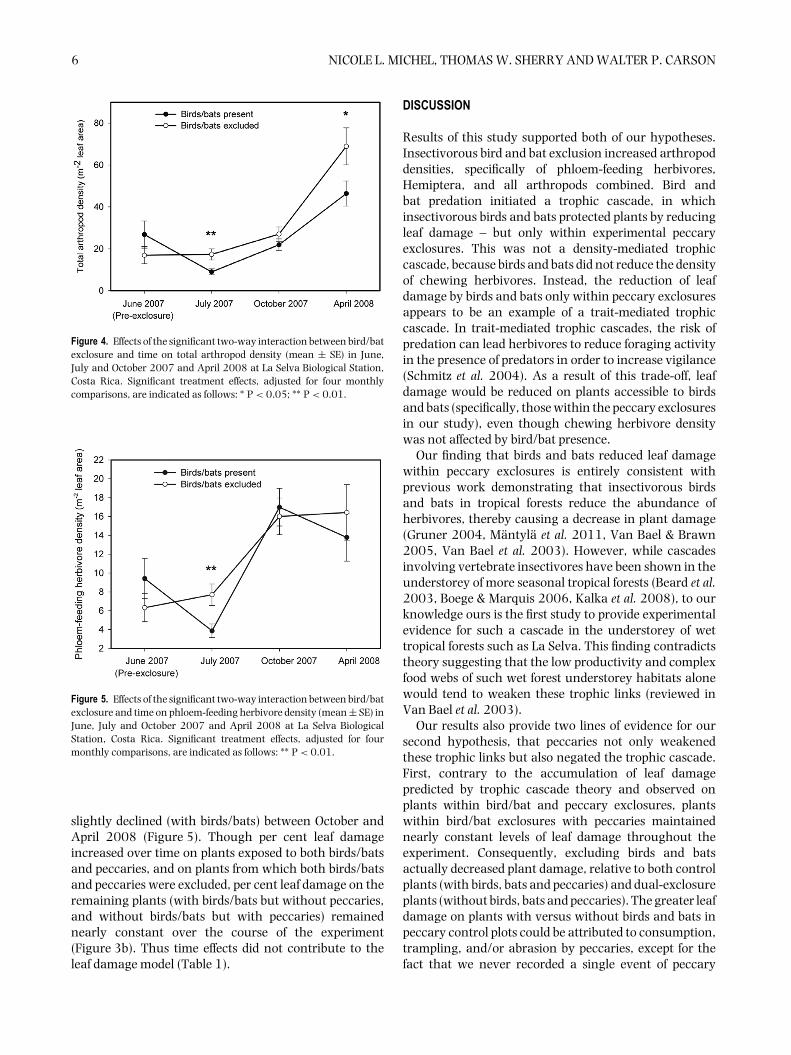
6 NICOLE L. MICHEL, THOMAS W. SHERRY AND WALTER P. CARSON
Figure 4. Effects of the significant two-way interaction between bird/batexclosure and time on total arthropod density (mean ± SE) in June,July and October 2007 and April 2008 at La Selva Biological Station,Costa Rica. Significant treatment effects, adjusted for four monthlycomparisons, are indicated as follows: ∗ P < 0.05; ∗∗ P < 0.01.
Figure 5. Effects of the significant two-way interaction between bird/batexclosure and time on phloem-feeding herbivore density (mean ± SE) inJune, July and October 2007 and April 2008 at La Selva BiologicalStation, Costa Rica. Significant treatment effects, adjusted for fourmonthly comparisons, are indicated as follows: ∗∗ P < 0.01.
slightly declined (with birds/bats) between October andApril 2008 (Figure 5). Though per cent leaf damageincreased over time on plants exposed to both birds/batsand peccaries, and on plants from which both birds/batsand peccaries were excluded, per cent leaf damage on theremaining plants (with birds/bats but without peccaries,and without birds/bats but with peccaries) remainednearly constant over the course of the experiment(Figure 3b). Thus time effects did not contribute to theleaf damage model (Table 1).
DISCUSSION
Results of this study supported both of our hypotheses.Insectivorous bird and bat exclusion increased arthropoddensities, specifically of phloem-feeding herbivores,Hemiptera, and all arthropods combined. Bird andbat predation initiated a trophic cascade, in whichinsectivorous birds and bats protected plants by reducingleaf damage – but only within experimental peccaryexclosures. This was not a density-mediated trophiccascade, because birds and bats did not reduce the densityof chewing herbivores. Instead, the reduction of leafdamage by birds and bats only within peccary exclosuresappears to be an example of a trait-mediated trophiccascade. In trait-mediated trophic cascades, the risk ofpredation can lead herbivores to reduce foraging activityin the presence of predators in order to increase vigilance(Schmitz et al. 2004). As a result of this trade-off, leafdamage would be reduced on plants accessible to birdsand bats (specifically, those within the peccary exclosuresin our study), even though chewing herbivore densitywas not affected by bird/bat presence.
Our finding that birds and bats reduced leaf damagewithin peccary exclosures is entirely consistent withprevious work demonstrating that insectivorous birdsand bats in tropical forests reduce the abundance ofherbivores, thereby causing a decrease in plant damage(Gruner 2004, Mantyla et al. 2011, Van Bael & Brawn2005, Van Bael et al. 2003). However, while cascadesinvolving vertebrate insectivores have been shown in theunderstorey of more seasonal tropical forests (Beard et al.2003, Boege & Marquis 2006, Kalka et al. 2008), to ourknowledge ours is the first study to provide experimentalevidence for such a cascade in the understorey of wettropical forests such as La Selva. This finding contradictstheory suggesting that the low productivity and complexfood webs of such wet forest understorey habitats alonewould tend to weaken these trophic links (reviewed inVan Bael et al. 2003).
Our results also provide two lines of evidence for oursecond hypothesis, that peccaries not only weakenedthese trophic links but also negated the trophic cascade.First, contrary to the accumulation of leaf damagepredicted by trophic cascade theory and observed onplants within bird/bat and peccary exclosures, plantswithin bird/bat exclosures with peccaries maintainednearly constant levels of leaf damage throughout theexperiment. Consequently, excluding birds and batsactually decreased plant damage, relative to both controlplants (with birds, bats and peccaries) and dual-exclosureplants (without birds, bats and peccaries). The greater leafdamage on plants with versus without birds and bats inpeccary control plots could be attributed to consumption,trampling, and/or abrasion by peccaries, except for thefact that we never recorded a single event of peccary

Peccaries negate bird/bat–insect–plant cascade 7
browsing on any of our focal plant species. However, directeffects of peccaries cannot explain the dramatically lowerleaf damage on plants without birds and bats in peccarycontrols relative to exclosures, as peccaries were unableto access either set of plants.
Second, bird/bat exclosure significantly reducedDiptera density on plants with peccaries, contrary to theincrease predicted by trophic cascade theory. Peccariessignificantly increased Diptera density on plants withbirds/bats, potentially due to elevated nutrient inputsfrom urine and faeces (Stewart 2001). However, Dipteradensity on plants with peccaries but without birds/batswas significantly lower than both nearby plants withbirds/bats, and plants within dual-exclosures (withoutpeccaries, birds or bats). This also cannot easily beexplained by direct effects of peccaries, and furtherindicates peccary disruption of the bird/bat–arthropod–plant trophic cascade.
The finding that peccaries negate an insectivorousbird/bat-generated trophic cascade was clear andhighly significant. To our knowledge, this is the firstdemonstration that the presence of an omnivorousungulate can mediate a trophic cascade via strictly non-trophic means. Collared peccaries are highly abundantat La Selva (Kuprewicz 2013), thus their effects on thetrophic cascade may be particularly pronounced at thissite. However, collared peccaries are fairly common atother protected Neotropical sites (e.g. Barro ColoradoIsland; Wright et al. 2000), and other ungulates suchas wild pig (S. scrofa) and deer (Odocoileus spp.) are highlyabundant in many tropical and temperate forests (Coteet al. 2004, Ickes 2001). Further studies are needed toinvestigate the effects of peccaries and other ungulates ata range of population densities and across sites.
Our results raise the question of what caused thedeclines in Diptera density and leaf damage inside thesenetted exclosures, but only in the presence of peccaries(i.e. outside peccary exclosures). It could not havebeen due to intraguild predation as typically defined inthese types of trophic cascades, i.e. release of arthropodpredators from predation and competition, as we explainhere. Many similar studies have found that arthropodpredators increase when birds and bats are excluded,and consequently suppress arthropod herbivores andreduce leaf damage (Vance-Chalcraft et al. 2007; butsee Mooney et al. 2010). Peccaries affected arthropodpredator densities, but in order for intraguild predationto explain the reduced leaf damage on plants withinbird/bat exclosures with peccaries, arthropod predatordensities would have to be both (1) consistently greaterwith peccaries, and (2) greater within bird/bat exclosuresthan controls. Instead, we found that peccaries actuallyreduced arthropod predator densities in July and October2007, and birds and bats had no effects on arthropodpredator densities. While we cannot rule out intraguild
predation altogether, the very small F value for the effectof bird/bat exclosure on arthropod predator density (F1,14
= 0.32) makes a Type II error unlikely in this case.While speculative, we suggest instead that the bird/bat
exclosures provided both a disturbance- and predator-free refuge for small insectivorous herpetofauna (e.g.anoles and leaf-litter frogs) that were too small to beexcluded by the netting. Leaf-litter amphibians andreptiles, including insectivorous Anolis (Norops) spp.anoles and Craugastor spp. frogs, are significantly moreabundant with peccaries on the same peccary exclosureand control plots at La Selva as were used in our study(Reider et al. 2013). Moreover, we occasionally observedthese herpetofauna within our bird/bat exclosures, andon our focal plants, during the study (N. Michel, pers.obs.). The bird/bat exclosures also excluded peccaries,with very few exceptions. The bird/bat exclosures mayhave served as a shelter that provided all the proposedbenefits of recent peccary visitation (increased nutrientavailability, litter mixing and rapid decomposition; Reideret al. 2013), without the risk of being trampled or evenconsumed by peccaries. Indeed, removing leaf litter fromthe top of the bird/bat exclosure netting and dispersing itwithin the cage monthly mixed the litter and may haveincreased decomposition rates, mimicking some peccaryeffects associated with greater herpetofaunal abundance(Reider et al. 2013). Furthermore, our netted exclosureswould have also excluded many other ground-dwellingand avian predators of small amphibians and reptiles(e.g. raptors, coati Nasua narica, tinamous (Tinamidae)).Unfortunately, we have no quantitative data onherpetofauna in our study because we were not awareof how important this might be at the time we designed it.
If true, congregation of insectivorous amphibians andreptiles within bird/bat exclosures in peccary controlscould result in a trait-mediated trophic cascade similarto that observed in the peccary exclosures due to theactions of birds and bats (Schmitz et al. 2004). If peccariesdrove herpetofauna to accumulate within the nettedexclosures, they could account for the observed declinesin plant damage because both Anolis lizards and leaf-litterfrogs are known to cause declines in insect herbivoreabundance and leaf damage (Beard et al. 2003, Dial& Roughgarden 1995). Although we also found nodifferences in chewing herbivore density between plantswith versus without birds and bats in peccary controls,a foraging-predation risk trade-off, as described above,may have occurred here as well. If the predation riskimposed by abundant insectivorous herpetofauna withinthe bird/bat exclosures exceeded the predation risk outsidethe exclosures, leaf damage would be reduced within thenetted exclosures – as we observed. Thus the peccaries,which are highly abundant at La Selva (Kuprewicz 2013)and have relatively small home ranges (38–305 hain tropical forest; Keuroghlian et al. 2004), may have

8 NICOLE L. MICHEL, THOMAS W. SHERRY AND WALTER P. CARSON
repeatedly herded vulnerable small herpetofauna insideour netted exclosures, initiating a trait-mediated trophiccascade that reduced Diptera density and leaf damage.Further studies are needed to investigate herpetofaunadensities within and outside bird/bat exclosures with andwithout peccaries.
Alternatively, it is possible that the netted bird/batexclosures in the peccary control plots (but not withinthe peccary exclosures) attracted highly motile arthropodpredators that were not detected during arthropodsurveys. These arthropods may have sought shelter fromdisturbance within the exclosures as speculated abovefor herpetofauna, and in turn suppressed leaf damage.While this seems unlikely for volant arthropod predators(e.g. wasps, lady beetles) that can easily evade peccarydisturbance, terrestrial arthropod predators (e.g. wolfspiders) could have sought refuge within the exclosures.Moreover, spiders that returned to the leaf litter duringthe arthropod surveys would not have been recorded.Other possible explanations, including a trait-mediatedtrophic cascade altering arthropod predator or herbivorebehaviour or rapid leaf regrowth in the presence ofpeccaries – but only within netted exclosures – seem lesslikely. Further studies testing these or other hypotheticalmechanisms should be conducted. But ultimately, thefinding that peccaries indirectly negate a bird/bat–arthropod–plant trophic cascade is novel in and of itself,whatever the mechanism.
Effects of seasonality
Arthropod densities were highly seasonal, peaking inthe late dry season. Arthropod densities peak in humidrain-forest understorey during the late dry season asarthropods seek moist refugia (Richards & Windsor2007). Moreover, as leaf-flushing peaks in February atthe study site, the availability of young leaves preferredby arthropod herbivores – and, consequently, densities ofherbivores and their predators – also peak in the late dryseason (Coley & Barone 1996).
The direction and strength of bird/bat- and peccary-exclosure treatment effects on arthropods also changedover time. Interestingly, bird/bat exclosure effects on totalarthropod densities were strongest in July, a period oflow plant productivity and low predator diversity (due toabsence of Nearctic-Neotropical migrant birds), contraryto trophic cascade theory as well as previous studies(Philpott et al. 2009, Williams-Guillen et al. 2008; butsee Borer et al. 2005). Instead, we suggest that bird/battop-down effects may be strongest shortly after exclosureconstruction, as herbivorous and omnivorous arthropodsseek refuge from predators within the novel structures.This top-down effect may dampen over time as arthropodand small vertebrate predators are attracted to the highprey densities within the exclosures, or as palatable
leaf material declines in availability due to consumptionor induced defences (Karban & Baldwin 1997, Vance-Chalcraft et al. 2007). Though rarely discussed, top-downeffect strength peaks within a few weeks or months post-exclosure construction are apparent in many previoustropical vertebrate predator exclosure studies (Gruner2004, Johnson et al. 2010, Morrison & Lindell 2012,Williams-Guillen et al. 2008).
Peccary exclosure treatment effects also variedseasonally. Peccaries reduced total arthropod densitiesin July and predatory arthropod densities in July andOctober, but increased predatory arthropod densities inApril. We suggest that arthropods may be attracted to thepeccary exclosures with their greater vegetation density(i.e. food, shelter; Michel 2012) in the low-productivitywet season, whereas plant productivity may be greaterin the peccary controls in the dry season due to nutrientinputs (i.e. urine, dung; Reider et al. 2013, Stewart 2001).However, further studies of the mechanism behind theseasonal reversal of peccary exclosure effects are needed.
Ultimately, seasonal fluctuations in arthropod densitiesdid not cascade to the plant level: per cent leaf damageeither consistently increased (bird/bat exclosure withoutpeccaries, bird/bat control with peccaries) or remainedmostly unchanged over the course of the experiment.Consequently, the trophic cascade observed on plantsin bird/bat exclosures without peccaries, and its notableabsence with peccaries, was unaffected by seasonality inarthropod densities.
Conclusions
Our conclusions strike a cautionary note regarding theuse of bird netting to exclude insectivorous birds and bats.These exclosures do more than just exclude birds and bats;rather they exclude any vertebrate that cannot breachthe bird-netting barrier as well as larger disturbanceagents such as peccaries. The potential for insectivoresother than birds, bats and herpetofauna to cause trophiccascades should be further explored in subsequentexclosure studies. We also suggest that there are naturalanalogues to our netted exclosures. Lianas can formdense impenetrable tangles (Schnitzer et al. 2000) thatmay impede passage by peccaries and other vertebrates(e.g. raptors), thus potentially providing natural shelterfrom trampling and predation (Lambert et al. 2006).Finally, our results show that ungulates can interact withinsectivorous birds and bats in unexpected ways, withcascading consequences for arthropods and plants.
We stress that, regardless of the specific mechanism bywhich peccaries reduced leaf damage in the absence ofbirds and bats, the finding that an omnivorous ungulatecan reverse a bird/bat–arthropod–plant trophic cascadeis novel, and has important theoretical and conservation

Peccaries negate bird/bat–insect–plant cascade 9
implications. Our work adds to the expanding literatureshowing that the magnitude – and even the direction –of species interactions such as trophic cascades dependon the biotic context (Agrawal et al. 2007). In ourstudy, birds and bats protected plants from leaf damageonly in the absence of peccaries. However, understoreyinsectivorous birds are experiencing population declinesin many tropical forests, while at the same time largeungulates like peccaries are being hunted to extirpation(Harrison 2011, Michel & Sherry 2012, Sodhi et al. 2011).Unless gleaning bats are able to compensate for declininginsectivorous bird populations, these concurrent trendsare likely to expose understorey plants to increasedarthropod herbivory pressure, as observed on plantswithin bird/bat and peccary exclosures in this study.Given the on-going changes in large ungulate, bird, batand herpetofaunal populations in tropical forests (Peterset al. 2006, Sodhi et al. 2011, Terborgh & Estes 2010,Whitfield et al. 2007), we urgently need to improve ourunderstanding of the potentially complex interactionsinvolving these important species and the potentialconsequences for tropical biodiversity.
ACKNOWLEDGEMENTS
This work was supported by an Organization forTropical Studies Research Fellowship to N. Michel, aNational Science Foundation (NSF) Doctoral DissertationImprovement Grant to N. Michel and T. Sherry (DEB-1010952), NSF grant DEB-071743 to T. Sherry, anda LA Board of Regents Graduate Fellowship fromTulane University. We thank Rebecca Forkner forassistance with research design, Deedra McClearn andthe staff of La Selva Biological Station for their supportand assistance, Orlando Vargas Ramırez for plantidentification assistance, and Julie Jackson Lewis andJames Lewis for assistance in the field. Lee Dyer, BenHirsch, Egbert Leigh, Tim Nuttle, Sunshine Van Baeland several anonymous reviewers provided constructivecomments that greatly improved this manuscript.
LITERATURE CITED
AGRAWAL, A. A., ACKERLY, D. D., ADLER, F., ARNOLD, A. E.,
CACERES, C., DOAK, D. F., POST, E., HUDSON, P. J., MARON, J.,
MOONEY, K. A., POWER, M., SCHEMSKE, D., STACHOWICZ, J.,
STRAUSS, S., TURNER, M. G. & WERNER, E. 2007. Filling key gaps
in population and community ecology. Frontiers in Ecology and the
Environment 5:145–152.
ANGIOSPERM PHYLOGENY GROUP. 2009. An update of the
Angiosperm Phylogeny Group classification for the orders and
families of flowering plants: APG III. Botanical Journal of the Linnean
Society 161:105–121.
BEARD, K. H., ESCHTRUTH, A. K., VOGT, K. A., VOGT, D. J. &
SCATENA, F. N. 2003. The effects of the frog Eleutherodactylus coqui
on invertebrates and ecosystem processes at two scales in the Luquillo
Experimental Forest, Puerto Rico. Journal of Tropical Ecology 19:607–
617.
BECK, H. 2005. Seed predation and dispersal by peccaries throughout
the Neotropics and its consequences: a review and synthesis. Pp. 77–
115 in Forget, P.-M., Lambert, J. E., Hulme, P. E. & Vander Wall, S. B.
(eds.). Seed fate: predation, dispersal and seedling establishment. CABI,
Wallingford.
BOEGE, K. & MARQUIS, R. J. 2006. Plant quality and predation risk
mediated by plant ontogeny: consequences for herbivores and plants.
Oikos 115:559–572.
BORER, E. T., SEABLOOM, J. B., SHURNIN, K. E., ANDERSON, C. A.,
BLANCHETTER, B. & HALPERN, B. S. 2005. What determines the
strength of a trophic cascade? Ecology 86:528–537.
COLEY, P. D. & BARONE, J. A. 1996. Herbivory and plant defenses in
tropical forests. Annual Review of Ecology and Systematics 27:305–
335.
COTE, S. D., ROONEY, T. P., TREMBLAY, J.-P., DUSSAULT, C. &
WALLER, D. M. 2004. Ecological impacts of deer overabundance.
Annual Review of Ecology Evolution and Systematics 35:113–147.
DIAL, R. & ROUGHGARDEN, J. 1995. Experimental removal of
insectivores from rain forest canopy: direct and indirect effects.
Ecology 76:1821–1834.
DICK, E. J. 2004. Beyond ‘lognormal versus gamma’: discrimination
among error distributions for generalized linear models. Fisheries
Research 70:351–366.
DYER, L. A., CARSON, W. P. & LEIGH, E. G. 2012. Insect outbreaks in
tropical forests: patterns, mechanisms, and consequences. Pp. 219–
245 in Barbosa, P., Letourneau, D. K. & Agrawal, A. A. (eds.). Insect
outbreaks revisited. Wiley-Blackwell, Hoboken, NJ.
GRUNER, D. S. 2004. Attenuation of top-down and bottom-up forces in
a complex terrestrial community. Ecology 85:3010–3022.
HARRISON, R. D. 2011. Emptying the forest: hunting and the
extirpation of wildlife from tropical nature reserves. BioScience
61:919–924.
ICKES, K. 2001. Hyper-abundance of native wild pigs (Sus scrofa) in
a lowland dipterocarp rain forest of peninsular Malaysia. Biotropica
33:682–690.
ICKES, K., DEWALT, S. J. & APPANAH, S. 2001. Effects of native pigs
(Sus scrofa) on woody understorey vegetation in a Malaysian lowland
rain forest. Journal of Tropical Ecology 17:191–206.
JOHNSON, M. D., KELLERMANN, J. L. & STERCHO, A. M. 2010. Pest
reduction services by birds in shade and sun coffee Jamaica. Animal
Conservation 13:140–147.
KALKA, M. B., SMITH, A. R. & KALKO, E. K. V. 2008. Bats limit
arthropods and herbivory in a tropical forest. Science 320:71.
KARBAN, R. & BALDWIN, I. T. 1997. Induced responses to herbivory.
University of Chicago Press, Chicago, IL. 330 pp.
KEFI, S., BERLOW, E. L., WIETERS, E. A., NAVARRETE, S. A., PETCHEY,
O. L., WOOD, S. A., BOIT, A., JOPPA, L. N., LAFFERTY, K. D.,
WILLIAMS, R. J., MARTINEZ, N. D., MENGE, B. A., BLANCHETTE,
C. A., ILES, A. C. & BROSE, U. 2012. More than a meal . . . integrating
non-feeding interactions into food webs. Ecology Letters 15:291–300.

10 NICOLE L. MICHEL, THOMAS W. SHERRY AND WALTER P. CARSON
KEUROGHLIAN, A., EATON, D. P. & LONGLAND, W. S. 2004. Area use
by white-lipped and collared peccaries (Tayassu pecari and Tayassu
tajacu) in a tropical forest fragment. Biological Conservation 120:411–
425.
KUPREWICZ, E. K. 2013. Mammal abundances and seed traits control
the seed dispersal and predation roles of terrestrial mammals in a
Costa Rican forest. Biotropica 45:333–342.
LAMBERT, T. D., MALCOLM, J. R. & ZIMMERMAN, B. L. 2006.
Amazonian small mammal abundances in relation to habitat
structure and resource abundance. Journal of Mammalogy 87:766–
776.
LITTELL, R. C., MILLIKEN, G. A., STROUP, W. W., WOLFINGER, R. D. &
SCHABENBERGER, O. 2006. SAS for mixed models. (Second edition).
SAS Institute, Cary, NC. 840 pp.
MANTYLA, E., KLEMOLA, T. & LAAKSONEN, T. 2011. Birds help
plants: a meta-analysis of top-down trophic cascades caused by avian
predators. Oecologia 165:143–151.
MARQUIS, R. J. & WHELAN, C. J. 1994. Insectivorous birds increase
growth of white oak through consumption of leaf-chewing insects.
Ecology 75:2007–2014.
MICHEL, N. L. 2012. Mechanisms and consequences of avian understory
insectivore population decline in fragmented Neotropical rainforest. Ph.D.
thesis, Tulane University, New Orleans.
MICHEL, N. L. & SHERRY, T. W. 2012. Human-altered mesoherbivore
densities and cascading effects on plant and animal communities
in fragmented tropical forests. Pp. 177–202 in Sudarshana, P.,
Nageswara-Rao, M. & Sonegi, J. R. (eds.). Tropical forests. InTech
Open Access Publisher, New York, NY.
MOONEY, K. A., GRUNER, D. S., BARBER, N. A., VAN BAEL, S. A.,
PHILPOTT, S. M. & GREENBERG, R. 2010. Interactions among
predators and the cascading effects of vertebrate insectivores on
arthropod communities and plants. Proceedings of the National
Academy of Sciences USA 107:7335–7340.
MORRISON, E. B. & LINDELL, C. A. 2012. Birds and bats reduce insect
biomass and leaf damage in tropical forest restoration sites. Ecological
Applications 22:1526–1534.
PETERS, S. L., MALCOLM, J. R. & ZIMMERMAN, B. L. 2006. Effects of
selective logging on bat communities in the southeastern Amazon.
Conservation Biology 20:1410–1421.
PHILPOTT, S. M., SOONG, O., LOWENSTEIN, J. H., PULIDO, A. L.,
LOPEZ, D. T., FLYNN, D. F. B. & DECLERCK, F. 2009. Functional
richness and ecosystem services: bird predation on arthropods in
tropical agroecosystems. Ecological Applications 19:1858–1867.
QUEENBOROUGH, S. A., METZ, M. R., WIEGAND, T. & VALENCIA,
R. 2012. Palms, peccaries and perturbations: widespread effects of
small-scale disturbance in tropical forest. BMC Ecology 12:3.
REIDER, K. E., CARSON, W. P. & DONNELLY, M. A. 2013. Effects of
collared peccary (Pecari tajacu) exclusion on leaf litter amphibians and
reptiles in a Neotropical wet forest, Costa Rica. Biological Conservation
163:90–98.
RICHARDS, L. A. & WINDSOR, D. M. 2007. Seasonal variation of
arthropod abundance in gaps and the understorey of a lowland moist
forest in Panama. Journal of Tropical Ecology 23:169–176.
SANFORD, R. L., PAABY, P., LUVALL, J. C. & PHILLIPS, E. 1994.
Climate, geomorphology, and aquatic systems. Pp. 19–33 in McDade,
L. A., Bawa, K. S., Hespenheide, H. A. & Hartshorn, G. S. (eds.). La
Selva: ecology and natural history of a Neotropical rainforest. University
of Chicago Press, Chicago, IL. 493 pp.
SAS INSTITUTE. 2009. SAS/STAT 9.2 user’s guide. SAS Institute, Inc.,
Cary, NC. 7869 pp.
SCHMITZ, O. S., KRIVAN, V. & OVADIA, O. 2004. Trophic cascades:
the primacy of trait-mediated indirect interactions. Ecology Letters
7:153–163.
SCHNITZER, S. A., DALLING, J. W. & CARSON, W. P. 2000. The impact
of lianas on tree regeneration in tropical forest canopy gaps: evidence
for an alternative pathway of gap-phase regeneration. Journal of
Ecology 8:655–666.
SHIPLEY, B. 2009. Confirmatory path analysis in a generalized
multilevel context. Ecology 90:363–368.
SODHI, N. S., SEKERCIOGLU, C. H., BARLOW, J. & ROBINSON, S. K.
2011. Conservation of tropical birds. Wiley-Blackwell, Hoboken, NJ.
300 pp.
STEWART, A. J. A. 2001. The impact of deer on lowland woodland
invertebrates: a review of the evidence and priorities for future
research. Forestry 74:259–270.
TERBORGH, J. W. & ESTES, J. A. 2010. Trophic cascades: predators, prey,
and the changing dynamics of nature. Island Press, Washington, DC.
488 pp.
TIMM, R. M. 1994. The mammal fauna. Pp. 229–237 in McDade, L. A.,
Bawa, K. S., Hespenheide, H. A. & Hartshorn, G. S. (eds.). La Selva:
ecology and natural history of a Neotropical rainforest. University of
Chicago Press, Chicago, IL.
TRIPLEHORN, C. A. & JOHNSON, N. F. 2004. Borror and DeLong’s
introduction to the study of insects. (Seventh edition). Brooks Cole,
Pacific Grove, CA. 888 pp.
VAN BAEL, S. A. & BRAWN, J. D. 2005. The direct and indirect effects of
insectivory by birds in two contrasting Neotropical forests. Oecologia
143:106–116.
VAN BAEL, S. A., BRAWN, J. D. & ROBINSON, S. K. 2003. Birds defend
trees from herbivores in a Neotropical forest canopy. Proceedings of
the National Academy of Sciences USA 100:8304–8307.
VANCE-CHALCRAFT, H. D., ROSENHEIM, J. A., VONESH, J. R.,
OSENBERG, C. W. & SIH, A. 2007. The influence of intraguild
predation on prey suppression and prey release: a meta-analysis.
Ecology 88:2689–2696.
WHITFIELD, S. M., BELL, K. E., PHILIPPI, T., SASA, M., BOLANOS, F.,
CHAVES, G., SAVAGE, J. M. & DONNELLY, M. A. 2007. Amphibian
and reptile declines over 35 years at La Selva, Costa Rica. Proceedings
of the National Academy of Sciences USA 104:8352–8356.
WILLIAMS-GUILLEN, K., PERFECTO, I. & VANDERMEER, J. 2008.
Bats limit insects in a Neotropical agroforestry system. Science 320:
70.
WRIGHT, S. J., ZEBALLOS, H., DOMINGUEZ, I., GALLARDO, M. M.,
MORENO, M. C. & IBANEZ, R. 2000. Poachers alter mammal
abundance, seed dispersal, and seed predation in a Neotropical forest.
Conservation Biology 14:227–239.

Peccaries negate bird/bat–insect–plant cascade 11
Appendix 1. List of numbers of individuals of each plant species used in the experiment, groupedby order (per APG III, 2009) for analysis. The number of plants of each species used in bird andbat exclosures and controls are listed in parentheses in the form (exclosure/control).
Order Family Species Number used
Piperales Piperaceae Piper trigonum C.DC. 5 (1/4)Magnoliales Annonaceae Anaxagorea crassipetala Hemsl. 10 (5/5)Santalales Olacaceae Minquartia guianensis Aubl. 6 (3/3)Fabales Polygalaceae Moutabea gentryi T.Wendt 4 (0/4)Fabales Fabaceae Swartzia sp. 1 (1/0)Rosales Moraceae Brosimum lactescens (S. Moore) C.C.Berg 2 (2/0)Rosales Moraceae Naucleopsis naga Pittier 4 (2/2)Malphigiales Violaceae Gloeospermum boreale Morton 5 (3/2)Malphigiales Humiriaceae Sacoglottis trichogyna Cuatrec. 6 (3/3)Malphigiales Dichapetalaceae Dichapetalum nervatum Cuatrec. 3 (3/0)Myrtales Melastomataceae Clidemia densiflora (Standl.) Gleason 6 (3/3)Sapindales Sapindaceae Paullinia ingifolia Rusby 1 (1/0)Gentianales Rubiaceae Psychotria buchtienii Standl. 4 (3/1)Gentianales Rubiaceae Coussarea talamancana Standl. 3 (0/3)




















