
designing silvopastoral systems for the amazon: a framework ...
333
1 DESIGNING SILVOPASTORAL SYSTEMS FOR THE AMAZON: A FRAMEWORK FOR THE EVALUATION OF NATIVE SPECIES Thesis submitted in partial fulfilment of the degree Doctor of Philosophy at UCL by Lucy Jayne Dablin
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of designing silvopastoral systems for the amazon: a framework ...

1
DESIGNING SILVOPASTORAL SYSTEMS FOR THE AMAZON: A
FRAMEWORK FOR THE EVALUATION OF NATIVE SPECIES
Thesis submitted in partial fulfilment of the degree Doctor of Philosophy at UCL by
Lucy Jayne Dablin

2
SIGNED DECLARATION
I, Lucy Jayne Dablin, confirm that the work presented in this thesis is my own. Where
information has been derived from other sources, I confirm that this has been
indicated in the thesis.

3
ACKNOWLEDGEMENTS
This work was possible due to the kind help and support of my supervisors, Dr Mark
Lee, Professor Simon Lewis, Dr William Milliken, Dr Alex Monro and my parents. I
would like to thank the many people who volunteered their efforts for this research.
This work was supported by the Natural Environment Research Council
[NE/L002485/1]; the Emily Holmes Memorial Scholarship; and the Bentham Moxon
Trust.

4
ABSTRACT
The integration of trees into pastures, termed silvopasture systems, could make
livestock farming more economically and environmentally sustainable. A framework
to assess the forage potential of native tree species would assist in developing
regionally appropriate silvopasture for pastures in the Amazon. Five tree species of
with reported silvopastoral potential, Senegalia loretensis, Ceiba pentandra,
Erythrina berteroana, Inga edulis and Leucaena leucocephala, were selected to
investigate their potential use in a silvopastoral system. A 26-month field experiment
planted 900 trees of the five species to assess tree mortality, biomass production
and palatability for cattle. Five monospecific replicates of each species and a control
with no trees planted were installed on a Brachiaria brizantha pasture in Madre de
Dios, Peru. After 18 months, S. loretensis and C. pentandra showed double the rate
of mortality compared to the other species and were deemed inappropriate for
continued consideration. Destructive harvests of the remaining three species showed
that total biomass production and hence carbon storage was greater for I. edulis (7.2
Mg dry mass ha-1) than E. berteroana (4.4 Mg dry mass ha-1) or L. leucocephala (4.3
Mg dry mass ha-1) at 24 months. These biomass gains were not at the expense of
grass: edible biomass within the reach of cattle was 1.5 Mg ha-1 in the no tree
control, whereas E. berteroana produced 2.3 Mg ha-1, I. edulis 2.3 Mg ha-1 and L.
leucocephala 2.2 Mg ha-1. Cattle were introduced into each treatment to assess
palatability. Cattle consumed 99% of E. berteroana, 75% of I. edulis and 80% of L.
leucocephala. E. berteroana had a mortality six times greater than other species, but

5
all species have silvopastoral potential. This evaluation of tree species supports
regionally appropriate silvopastoral systems and shows that silvopasture can
increase productivity on existing cleared land and can contribute to mitigating climate
change.

6
IMPACT STATEMENT
Silvopastoral farming systems, where trees are planted in cattle pasture, could make
livestock farming more economically and environmentally sustainable when
compared with traditional models of cattle farming. In some areas, a barrier to the
adoption of silvopasture is a lack of knowledge of which environmentally adapted
trees have forage potential and whether these trees can meet the needs of animal
production. This thesis describes a framework that can be used to evaluate the
silvopastoral suitability of native and locally adapted tree species. The framework
presents a set of design considerations and recommends that silvopastoral trials be
carried out in a well-replicated scientifically robust design over a minimum of a two-
year timeframe. This approach has been designed for on-farm use in a way that can
be adapted by practitioners in different parts of the world.
The framework was applied in the context of a typical Amazon pasture in Madre de
Dios, Peru. Five locally adapted trees were chosen following a review of scientific
literature that suggested locally adapted species that could survive in the pasture
environment and were palatable for cattle. When compared with conventional
models of farming in the region, the silvopastoral system provided gains in
productivity, accruing as increased total biomass, an increase in available forage for
cattle and the sequestration of carbon. A statistical method to provide non-
destructive assessments of the biomass production and carbon sequestration
potential of the tree components of three species is presented.

7
An accomplishment of this study was the establishment of a 3-hectare experimental
silvopastoral system. This serves as a demonstration of the procedures advocated in
this thesis and can continue inspire those who seek to implement silvopastoral
systems. Site visits by stakeholders may increase the adoption of more sustainable
cattle farming techniques. It is hoped that the dissemination of this research via
scientific journals will contribute to a growing body of evidence that supports the
adoption of more environmentally friendly practices in livestock production, of
interest to extension workers, policy makers, governments and businesses.
This work is an attempt to find sustainable ways to intensify the current extensive
livestock production in the Amazon using natural technology and simultaneously
increase the provision of ecosystem services from these landscapes. It is envisaged
that widespread adoption of silvopastoral systems on existing cleared land could
reduce the need for further deforestation, reverse the trend of pasture degradation
and soil erosion, increase biodiversity in agricultural landscapes and contribute to the
mitigation of climatic change. This work is a preliminary assessment but it could be
of use to future researchers and extension workers who may be able to address the
shortcomings of this study and further improve the recommendations for the design
of silvopastoral systems. Finally, the ultimate impact that this study could hope to
achieve is that the communities that rely on cattle farming in the Amazon adopt
these more profitable and sustainable approaches, and that this results in a decline
in the rate of pasture degradation and a decline in the rate of deforestation of the
Amazon.

8
TABLE OF CONTENTS
DESIGNING SILVOPASTORAL SYSTEMS FOR THE AMAZON: A FRAMEWORK
FOR THE EVALUATION OF NATIVE SPECIES ....................................................... 1
SIGNED DECLARATION ....................................................................................... 2
ACKNOWLEDGEMENTS ....................................................................................... 3
ABSTRACT ............................................................................................................ 4
IMPACT STATEMENT ........................................................................................... 6
TABLE OF CONTENTS .......................................................................................... 8
TABLE OF FIGURES ........................................................................................... 13
TABLE OF TABLES .............................................................................................. 20
TABLE OF APPENDICES .................................................................................... 25
CHAPTER I .......................................................................................................... 35
1. Livestock Production .............................................................................. 35
2. Introduction to Silvopastoral Systems .................................................... 37
3. Selection and Management of Species for Silvopastoral Systems ........ 41
4. Traditional Livestock Production Models And The Potential For
Silvopastoral Systems: Influence On Soil Health ....................................... 51
5. Evaluating the Forage Potential of Tree Species ................................... 55
6. Summary and Rationale......................................................................... 60
References ................................................................................................. 62
CHAPTER II ......................................................................................................... 87
1. Objective ................................................................................................ 87

9
2. The Purpose .......................................................................................... 89
3. The Site.................................................................................................. 89
3.1 Site Description ................................................................................ 90
3.2 Floristic Description of Site ............................................................... 92
3.3 Climatic Description of Site ............................................................... 93
3.4 Edaphic Description of Site .............................................................. 95
3.5 Social and Economic History ............................................................ 96
3.6 Future of the Amazon ....................................................................... 97
3.7 Site Preparation ................................................................................ 98
4. Species ................................................................................................ 102
4.1 Evaluation of Desirable Plant Traits and Candidate Trees in
Silvopastoral Systems .......................................................................... 102
4.2. Species Information ....................................................................... 108
4.2.1 Inga edulis ............................................................................... 108
4.2.2 Ceiba pentandra ...................................................................... 112
4.2.3 Erythrina berteroana ................................................................ 114
4.2.4 Senegalia loretensis ................................................................. 116
4.2.5 Leucaena leucocephala ........................................................... 117
4.2.6 Brachiaria brizantha ................................................................. 118
4.3 Design and Management of Tree Nursery ...................................... 120
4.4 Treatment and Sowing of Seeds .................................................... 124
5. Planting Design .................................................................................... 128
6. Experimental Design ............................................................................ 130
6.1 Planting Sequence ......................................................................... 132
References ............................................................................................... 136

10
CHAPTER III ...................................................................................................... 152
Abstract .................................................................................................... 152
1. Introduction .......................................................................................... 153
2. Methods ............................................................................................... 155
2.1. Censuses of Tree Growth .............................................................. 156
2.2. Soil Chemical Analysis .................................................................. 158
2.3 Pollard Treatment ........................................................................... 160
2.4. Effects of Trees on Rates of Pasture Regrowth ............................ 160
3. Results ................................................................................................. 161
4. Discussion ............................................................................................ 177
References ............................................................................................... 185
CHAPTER IV ...................................................................................................... 193
Abstract .................................................................................................... 193
1. Introduction .......................................................................................... 194
2. Methods ............................................................................................... 199
2.1 Measurements of Tree Biomass ..................................................... 199
2.2 Data Exploration ............................................................................. 202
2.3 Model Comparison and Selection ................................................... 203
2.4 Model Validation ............................................................................. 203
2.5 Allometric Models .......................................................................... 204
2.6 Calculation of Carbon Stocks ......................................................... 205
3. Results ................................................................................................. 206
3.1 Measurements of Tree Biomass ..................................................... 206
3.2 Data Exploration ............................................................................. 210
3.3 Model Selection .............................................................................. 213

11
4. Discussion ............................................................................................ 218
References ............................................................................................... 229
Appendices .............................................................................................. 243
CHAPTER V ....................................................................................................... 265
Abstract .................................................................................................... 265
1. Introduction .......................................................................................... 266
2. Methods ............................................................................................... 270
2.1 Experimental Design and Cattle Herd ............................................ 270
2.2. Browsing Detection Using Destructive Measurements .................. 272
2.3. Browsing Detection Using Non-Destructive Measurements .......... 274
2.4 Browsing Detection Using Leaf Counts .......................................... 276
2.5 Measurement Of Browsing Impact ................................................. 276
2.6 Estimation Of Grass Forage ........................................................... 277
2.7 Effect Of Trees and Cattle on Soil Bulk Density ............................. 278
3. Results ................................................................................................. 280
4. Discussion ............................................................................................ 289
References ............................................................................................... 299
CHAPTER VI ...................................................................................................... 307
1. Summary of Research Findings ........................................................... 307
2. Comparison with Previous Research ................................................... 311
3. Theoretical Implications of Findings ..................................................... 315
4. Limitations of the Study ........................................................................ 317
5. Future Research .................................................................................. 321
5.1 Direct Extensions of the Study ....................................................... 321
5.2 Broader Issues for Future Work ..................................................... 321

12
7. Conclusion ........................................................................................... 323
References ............................................................................................... 324

13
TABLE OF FIGURES
Figure 1. Theoretical framework for the evaluation of candidate species for
agroforestry systems ......................................................................................... 88
Figure 2. Location of the study site in Madre de Dios, Peru ..................................... 91
Figure 3. Experimental site in 2002 with the future location of the plots projected .. 92
Figure 4. Experimental site in 2015 with the future location of the plots projected. .. 92
Figure 5. Average (solid line), maximum (dotted line), and minimum (dashed line)
monthly temperature (°C) from the period 1970-2000 for the study site derived
from the WorldClim dataset (Hijmans et al., 2005). ........................................... 94
Figure 6. Average monthly precipitation (mm) from the period 1970-2000 for the
study site derived from the WorldClim dataset (Hijmans et al., 2005). .............. 94
Figure 7. Average monthly level of solar irradiation (kJ m-2 day-1) from the period
1970-2000 for the study site derived from the WorldClim dataset (Hijmans et al.,
2005). ................................................................................................................ 95
Figure 8. Experimental site in October 2016 with the future plots projected. ........ 100
Figure 9. A monumental task. All site clearance was carried out by hand to minimise
disturbance to the soils. .................................................................................. 100
Figure 10. An international team of volunteers involved helped with the pile and
burn. ................................................................................................................ 101
Figure 11. When the site had been cleared of branches, the weeds at the site had
grown to be unmanageable. A glyphosphate herbicide was applied across the
site and the vegetation had to be cleared again. Clearance of this vegetation

14
was done using a hoe and rake. All residues were taken off site in
wheelbarrows. ................................................................................................. 101
Figure 12. Mind map of desirable plant traits in the context of developing
silvopastoral systems at a site in the Amazon that could be advantageous in
achieving the goal of increasing the productivity per hectare through the use of
silvopastoral systems compared with traditional methods of cattle production in
the region. ....................................................................................................... 104
Figure 13. Resources for identifying potential species for inclusion in agroforestry
systems. .......................................................................................................... 105
Figure 14. Inga edulis growth (Height) over time (Months), from: (1) Sobanski and
Marques (2014), (2) Alegre (2005), (3) Kanmegne et al., (2000), (4) Rezende &
Vieira (2019), (5) Butterfield (1996), (6) Butterfield (1995), (7) Gonzalez and
Fisher (1994) ................................................................................................... 110
Figure 15. Biomass after 11 years (1993 – 2004) in five replications of two
treatments, T1 (control) and T6 (interplanted with Inga edulis), in Mg ha-1.
Biomass of weeds and Inga was measured in 1 x 3 m subplots within treatment
plots and converted to a per hectare basis. Biomass of Terminalia amazonia is
the sum of crown foliage and bole wood biomasses of a sample tree for each
treatment plot. Crown and bole biomasses were estimated from volumes
calculated from diameter and height measurements multiplied by foliage density
and wood density, respectively. Once the tree biomass was obtained, biomass
was converted to a per hectare basis depending on the number of trees in the
plot. Total biomass is the sum of the weed and Inga biomass plus the T.
amazonia biomass on a per hectare basis (from Nichols and Carpenter, 2006).
........................................................................................................................ 111

15
Figure 16. Little is known about Senegalia loretensis, but despite it’s sharp spines
the leaves are highly palatable to cattle. ......................................................... 117
Figure 17. (Clockwise from left to right) The first attempt at digging the well, the base
of the steel poles, which contained a 2.5 cm iron bar, alignment of the steel
poles for the nurseries. .................................................................................... 121
Figure 18. The nursery mesh was elevated upon steel poles above the plants, with
enough room to allow air to circulate and to enable easy access for watering the
plants. ............................................................................................................. 122
Figure 19. Some prepared soil for filling the bags for the plants. .......................... 123
Figure 20. View over the nurseries with 10,000 bags of soil. Borders made from
trees define the beds. The bags of soil are arranged in rows to give easy access
for weeding and watering of the plants. In the background is the water tower that
enabled water to the site. ................................................................................ 124
Figure 21. Seed pods and leaves of C. pentandra. ............................................... 125
Figure 22. I edulis .................................................................................................. 126
Figure 23. Seeds of L. leucocephala ..................................................................... 126
Figure 24. Seedlings of L. leucocephala ............................................................... 127
Figure 25. Beds of S. loretensis (foreground) and C. pentandra (background). .... 127
Figure 26. E. berteroana was planted from stakes, pictured here 2 months post
planting in association with the immature pasture grass. ................................ 128
Figure 27. The randomised replicate plot layout, where EB is Erythrina berteroana,
IE is Inga edulis, LL is Leucaena leucocephala, SL is Senegalia loretensis, CP is
Ceiba pentandra and C is a control of grass. .................................................. 131

16
Figure 28. Map of experimental site. 0.09 ha plots as red rectangles, tree rows as
black dashed lines. Tree rows were continuous between plots. There were no
tree lines on the red lines. ............................................................................... 132
Figure 29. Satellite image of experimental site, 0.09 ha parcels as red rectangles.
Image taken in April 2018. Image by Maxar Technologies. ............................. 132
Figure 30. The plastic tubes used to mark the 30 m intervals of the plot corners and
in the background weeds responding to the glyphosphate applied across the
site. ................................................................................................................. 133
Figure 31.Team planting L. leucocephala in January of 2017. .............................. 135
Figure 32. Planting lines of I. edulis........................................................................ 135
Figure 33. Satellite image from April 2017 with the parcels projected, showing faint
tree lines and the establishing grass. .............................................................. 136
Figure 34. Relative growth rate for changes in height of five tree species after
planting at six (C2), twelve (C3) and eighteen (C4) month intervals post-planting
for tree species Leucaena leucocephala, Senegalia loretensis, Inga edulis,
Ceiba pentandra and Erythrina berteroana. .................................................... 164
Figure 35. Relative growth rate for changes in root collar diameter (RCD) and of five
tree species at between planting and six (C2), twelve (C3) and eighteen (C4)
month intervals post-planting for tree species Leucaena leucocephala,
Senegalia loretensis, Inga edulis, Ceiba pentandra and Erythrina berteroana 165
Figure 36. Height (cm) and standard deviation of five tree species growing in a
Brachiaria brizantha cv. Marandú pasture as of census 1. March 2017, 2. July
2017, 3. March 2018 and 4. July 2018. ........................................................... 166

17
Figure 37. RCD (mm) and standard deviation of five tree species growing in a
Brachiaria brizantha cv. Marandú pasture as of census 1. March 2017, 2. July
2017, 3. March 2018 and 4. July 2018. ........................................................... 167
Figure 38. DBH (mm) of five tree species growing in a Brachiaria brizantha cv.
Marandú pasture as of census 2. July 2017, 3. March 2018 and 4. July 2018.
........................................................................................................................ 168
Figure 39. Rows of I. edulis .................................................................................... 169
Figure 40. Row of Ceiba pentandra ....................................................................... 170
Figure 41. Replicate of Senegalia loretensis .......................................................... 171
Figure 42. Leucaena leucocephala in March 2017 (above) and October 2017 (below)
........................................................................................................................ 172
Figure 43. Proportion of alive, dead and trees classified as regrowth of five tree
species in a Brachiaria brizantha cv. Marandú pasture in the SE Amazon 18
months after planting at 0.5 x 7.5 m ................................................................ 173
Figure 44. Mean dry weight biomass and standard deviation (S.D.) of pollarded
biomass (over 2 m) of 18-month-old trees 0.5 x 7.5 m planting design (2667
trees ha-1) for individual trees, sample size represented in parenthesis. Means
sharing the same superscript are not significantly different from each other
(Tukey's HSD, P<0.05) ................................................................................... 175
Figure 45. Mean total dry matter biomass accumulation (Mg ha-1 year-1) and
standard deviation (S.D.) of 30-day regrowth of Brachiaria brizantha cv.
Marandú collected from different treatments, sample size represented in
parenthesis. Means sharing the same superscript are not significantly different
from each other (Tukey's HSD, P<0.05) ......................................................... 176

18
Figure 46. Mean total dry matter biomass accumulation (Mg ha-1 year-1) and standard
deviation (S.D.) of six months regrowth of Brachiaria brizantha cv. Marandú
following the pollarding of associated tree species to 2 m, sample size
represented in parenthesis. Means sharing the same superscript are not
significantly different from each other (Tukey's HSD, P<0.05) ........................ 177
Figure 47. Diagram representing the location of measurements on tree, where CD is
crown diameter, CDep is crown depth, CH is crown height, H is height, RCD is
root collar diameter, SD is stump diameter and DBH is diameter at breast
height. ............................................................................................................. 200
Figure 48. The log transformation of x (root collar diameter (mm) + height (cm))
derived using a linear model for the log transformation of 1) y as total biomass
(g, R2 = 0.77, P <0.01) and 2) y as trunk biomass (g, R2 = 0.76, P<0.01) for
Erythrina berteroana following the equations presented in Table 13............... 216
Figure 49. The log transformation of crown diameter (cm) derived using a linear
model for the log transformation of branch biomass (g, R2 = 0.73, P<0.01) and
leaf biomass (g, R2 = 0.58, P<0.01) for Erythrina berteroana following the
equations presented in Table 13. .................................................................... 216
Figure 50. The log transformation of root collar diameter (mm) derived using a linear
model for the log transformation of total biomass (g, R2 = 0.70, P<0.01), trunk
biomass (g, R2 = 0.71, P<0.01), branch biomass (g, R2 = 0.48, P<0.01) and leaf
biomass (g, R2 = 0.48, P<0.01) for Inga edulis following the equations presented
in Table 13. ..................................................................................................... 217
Figure 51. The log transformation of x where 1) x is root collar diameter (mm) +
height (cm) derived using a linear model for the log transformation of total
biomass (g, R2 = 0.86, P<0.01), trunk biomass (g, R2 = 0.89, P<0.01) and leaf

19
biomass (g, R2 = 0.61, P<0.01) and 2) x is crown diameter (cm) derived using a
linear model for the log transformation of branch biomass (g, R2 = 0.51, P<0.01)
for Leucaena leucocephala following the equations presented in Table 13. ... 217
Figure 52. Map of the cattle trial from east to west Phase I (red), Phase II (blue) and
Phase III (yellow). ............................................................................................ 272
Figure 53. Cattle browsing Inga edulis .................................................................. 281
Figure 54. Browsing line where cattle consumed all the foliage they could reach. . 282
Figure 55. Post-browsing, an Erythrina berteroana row is missing several trees ... 282
Figure 56. Leucaena leucocephala defoliated on low branches ............................. 283
Figure 57. Percentage change of Erythrina berteroana biomass for four assessment
methodologies used to detect biomass removal following cattle browsing. ..... 285
Figure 58. Percentage change of Inga edulis biomass components between
methodologies used to detect biomass removal following a cattle-browsing
experiment. ..................................................................................................... 286
Figure 59. Percentage change of Leucaena leucocephala biomass components
between methodologies used to detect biomass removal following a cattle-
browsing experiment. ...................................................................................... 286

20
TABLE OF TABLES
Table 1. Plantation tree species killed and damaged by exposure to approximately
80-100 cattle in a 3-hour period. Data provided by Camino Verde (2018). . ... 107
Table 2. Criteria used in the selection of the final five species for the present
experiment. Where ++ is best suited to -- worse suited, + or – are weak
evidence to support either way, and ? indicate there was no information found
linking the species to the criteria. .................................................................... 107
Table 3. Plant height, soil cover and dry matter yield of forage grasses Brachiaria
brizantha cv. Marandú, B. humidicola, Paspalum atratum BRA-9610, P.
guenoarum BRA-3824, P. regnelli BRA-0159, P. plicatulum BRA-9661 and
Hemarthria altissima established under a 12 year old rubber plantation
reproduced from Costa et al. (2001). .............................................................. 119
Table 4. Data for the first day of each month from a site in Madre de Dios, Peru (lat -
12.53945°, long -69.17288°) in 2016, including the altitude and azimuth of the
sun at 12:00, the shadow cast by an object of 2 m height and the daylight
duration. .......................................................................................................... 130
Table 5. Plant growth census dates. ...................................................................... 157
Table 6. Height (cm), root collar diameter (RCD mm), and diameter at breast height
(DBH mm) of five tree species in a Brachiaria brizantha cv. Marandú pasture,
with census number (C), time after planting in months (t), mean average (Mean)
standard deviation (S.D.), annual relative growth rate (RGR), n as number of
individuals sampled and λ as annual mortality (Shiel et al., 1995). Means

21
sharing the same superscript are not significantly different from each other
within a census interval (Tukey's HSD, P<0.05).............................................. 163
Table 7. Descriptive statistics of biomass parameters of individual harvested
Erythrina berteroana used in constructing allometric equations, including sample
size (n), root collar diameter (RCD, 0.05 m), stump diameter (SD, 0.3 m) and
diameter at breast height (DBH, 1.3 m). Mean, standard deviation (SD),
minimum (min), maximum (max), lower 25th percentile (25th perc), upper 75th
percentile (75th perc), as well as coefficients of skewness and kurtosis are
presented. ....................................................................................................... 207
Table 8. Descriptive statistics of biomass parameters of individual harvested Inga
edulis used in constructing allometric equations, including sample size (n), root
collar diameter (RCD, 0.05 m), stump diameter (SD, 0.3 m) and diameter at
breast height (DBH, 1.3 m). Mean, standard deviation (SD), minimum (min),
maximum (max), lower 25th percentile (25th perc), upper 75th percentile (75th
perc), as well as coefficients of skewness and kurtosis are presented. .......... 208
Table 9. Descriptive statistics of biomass parameters of individual harvested
Leucaena leucocephala used in constructing allometric equations, including
sample size (n), root collar diameter (RCD, 0.05 m), stump diameter (SD, 0.3 m)
and diameter at breast height (DBH, 1.3 m). Mean, standard deviation (SD),
minimum (min), maximum (max), lower 25th percentile (25th perc), upper 75th
percentile (75th perc), as well as coefficients of skewness and kurtosis are
presented. ....................................................................................................... 209
Table 10. Spearman’s correlation of E. berteroana biomass components. Total
biomass (Total), trunk biomass (Trunk), branch biomass (Branch), total foliar
biomass (Leaf), root collar diameter (RCD, 0.05 m), stump diameter (SD, 0.3 m),

22
diameter at breast height (DBH, 1.3 m), height, crown volume (CVol) and crown
diameter (CD).................................................................................................. 211
Table 11. Spearman’s correlation of I. edulis biomass components. Total biomass
(Total), trunk biomass (Trunk), branch biomass (Branch), total foliar biomass
(Leaf), root collar diameter (RCD, 0.05 m), stump diameter (SD, 0.3 m),
diameter at breast height (DBH, 1.3 m), height, crown volume (CVol) and crown
diameter (CD).................................................................................................. 211
Table 12. Spearman’s correlation of L. leucocephala biomass components. Total
biomass (Total), trunk biomass (Trunk), branch biomass (Branch), total foliar
biomass (Leaf), root collar diameter (RCD, 0.05 m), stump diameter (SD, 0.3 m),
diameter at breast height (DBH, 1.3 m) height, crown volume (CVol) and crown
diameter (CD).................................................................................................. 212
Table 13. Log-linear allometric equations demonstrated the lowest Furnival’s Index
(F.I.) against linear or quadratic models for biomass components of 0.5 x 7.5 m
alley rows of Erythrina berteroana, Inga edulis and Leucaena leucocephala 6
months post pollarding to a height of 2 m. Where ln(x) is the log transformation
of the following measurements used as predictors of biomass: Root collar
diameter (RCD mm, 0.05 m), height (cm) and crown diameter (CD, cm). ln(y) is
the log transformation of total biomass (g), trunk biomass (g), branch biomass
(g) or leaf biomass (g). Where b0 is the intercept coefficient, b1 and b2 are the
slope coefficients, all of which are reported with standard error (S.E.) and
significance (P). Lambda (λ) and standard deviation (S.D.) as back
transformation according to Marklund (1987), root mean square error of model
(Model rMSE) and results of K-fold model cross validation (k=10) reported as
root mean squared error (CV rMSE). .............................................................. 215

23
Table 14. Total above ground biomass fraction and carbon (C) stock (Mg ha-1) after
24 months, including previously removed biomass > 2 m at 18 months (Chapter
III) of Erythrina berteroana, Inga edulis and Leucaena leucocephala when 2667
trees ha-1 (0.5 x 7.5 m), using the IPCC default C fraction for tropical forests of
0.47. Where n is sample size and MAI is Mean Annual Increment (Mg ha-1 year-
1) (Eggleston et al., 2006)................................................................................ 218
Table 15. Pairing combinations and random rotation of cattle (8 cattle numbered 1-8)
between four treatments in three, eight day phases of a cattle trial. ............... 271
Table 16. Dry foliar biomass for individual trees (dry matter g tree-1), standard
deviation (S.D.) and number of individuals sampled (n) prior to and post cattle
browsing either mean total tree foliar biomass or foliar biomass harvested <1.6
m on potential browse species Erythrina berteroana, Inga edulis and Leucaena
leucocephala. Significant differences (P<0.05) from prior value to post value
following cattle browsing denoted by an asterisk (*). ....................................... 280
Table 17. Mean average (Av) and standard deviation (S.D.) of change per individual
tree in crown volume (M3) and leaf emergence height (LEH, cm) prior to and
post a cattle browsing experiment on individuals (n) of potential browse species,
Erythrina berteroana, Inga edulis and Leucaena leucocephala. Significant
differences between sampling period prior to and post (P<0.05) denoted by an
asterix (*). ........................................................................................................ 284
Table 18. Mean leaf count per tree (number of leaves tree-1) and standard deviation
(S.D.) prior to and post a cattle-browsing trial on individuals (n) of potential
browse species Erythrina berteroana, Inga edulis and Leucaena leucocephala.
Significant differences (P<0.05) between leaves counted prior to and post
browsing are denoted by an asterix (*). ........................................................... 285

24
Table 19. Impacts measured following a cattle trial on three potential browse species
Erythrina berteroana, Inga edulis and Leucaena leucocephala. Impact types
were classified as browsing-type damage (Browsed) or breakage-type damage
(Broken) of two categories, branches (all species) and leaves (I. edulis).
Average diameter (mm), average height (cm) and maximum height (cm) and
respective standard deviations (S.D.) reported on a number (n) of individual
trees. Significant differences between species (P<0.05) denoted by asterisk (*).
........................................................................................................................ 287
Table 20. Edible and non-edible fractions of mean average dry weight grass biomass
(Mg ha-1) produced in four treatments, reported for sampling carried out prior to
and post the introduction of cattle. .................................................................. 289

25
TABLE OF APPENDICES
Appendix 1. Table. The wet to dry relationship for biomass components of
subsamples.taken from Inga edulis, Erythrina berteroana and Leucaena
leucocephala, where R2 is the coefficient of determination, RSE is the residual
square error and n is the sample size. ............................................................ 243
Appendix 2. Figure. Total biomass (g) and root collar diameter (mm) of Inga edulis
plotted using the linear regression line (y = aX + b) showing clockwise from left
(a) scatter plot and linear regression line, (b) departure from normal Q-Q plot
and (c) residuals vs. leverage ......................................................................... 244
Appendix 3. Figure. Relationship between total biomass (g) and root collar diameter
(mm) for Inga edulis plotted using the quadratic regression model (equation y =
ax2 + bx + c) showing clockwise from left (a) scatter plot and polynomial
regression line, (b) departure from normal Q-Q plot and (c) residuals vs.
leverage .......................................................................................................... 245
Appendix 4. Figure. Relationship between total biomass (g) and root collar diameter
(mm) for I. edulis plotted using the log-linear transformation ln(y) = a + b * lnx
showing clockwise from left (a) scatter plot and regression line, (b) departure
from normal Q-Q plot and (c) residuals vs. leverage ....................................... 246
Appendix 5. Table. Allometric equations following the linear model for biomass
components of 0.5 x 7.5 m alley rows of Erythrina berteroana 6 months post
pollarding to a height of 2 m. Where (x) is the measurement used as a predictor
of biomass: Root collar diameter (RCD mm, 0.05 m), diameter at breast height

26
(DBH (mm), 1.3 m), stump diameter (SD mm, 0.3 m), crown volume (CV cm3)
and crown diameter (CD cm), (y) is the log transformation of total biomass(g),
trunk biomass (g), branch biomass (g) or leaf biomass (g). b0 as the intercept
coefficient, b1 as the slope coefficient, both reported with standard error (S.E.)
and significance (P). Coefficient of determination (R2), root mean square error of
model (Model RMSE, which is the same as Furnival’s Index for untransformed
data) and results of K-fold model cross validation (k=10) reported as root mean
squared error (CV RMSE). .............................................................................. 247
Appendix 6. Table. Allometric equations following the linear model for biomass
components of 0.5 x 7.5 m alley rows of Erythrina berteroana 6 months post
pollarding to a height of 2 m. Where (x) is the root collar diameter (RCD mm,
0.05 m), or RCD and Height (cm) as predictors of biomass. (y) is the log
transformation of total biomass (g), trunk biomass (g), branch biomass (g) or
leaf biomass (g), b0 as the intercept coefficient, b1 and b2 as the slope coefficient,
both reported with standard error (S.E.) and significance (P). Coefficient of
determination (R2), root mean square error of model (Model RMSE, which is the
same as Furnival’s Index F.I. for untransformed data) and results of K-fold
model cross validation (k=10) reported as root mean squared error (CV RMSE).
........................................................................................................................ 248
Appendix 7. Table. Allometric equations following the linear model for biomass
components of 0.5 x 7.5 m alley rows of Inga edulis 6 months post pollarding to
a height of 2 m. Where (x) is the measurement used a predictor of biomass:
Root collar diameter (RCD mm, 0.05 m), diameter at breast height (DBH mm,
1.3 m), stump diameter (SD mm, 0.3 m), crown volume (CV cm3) and crown
diameter (CD cm), (y) is the log transformation of total biomass (g), trunk

27
biomass (g), branch biomass (g) or leaf biomass (g), b0 as the intercept
coefficient, b1 as the slope coefficient, both reported with standard error (S.E.)
and significance (P). Coefficient of determination (R2), root mean square error of
model (Model RMSE, which is the same as Furnival’s Index for untransformed
data) and results of K-fold model cross validation (k=10) reported as root mean
squared error (CV RMSE). .............................................................................. 249
Appendix 8. Table. Allometric equations following the linear model for biomass
components of 0.5 x 7.5 m alley rows of Inga edulis 6 months post pollarding to
a height of 2 m. Where (x) is the root collar diameter (RCD mm, 0.05 m), or
RCD and Height (cm) as predictors of biomass, (y) is the log transformation of
total biomass (g), trunk biomass (g), branch biomass (g) or leaf biomass (g). b0
as the intercept coefficient, b1 and b2 as the slope coefficient, both reported with
standard error (S.E.) and significance (P). Coefficient of determination (R2), root
mean square error of model (Model RMSE, which is the same as Furnival’s
Index F.I. for untransformed data) and results of K-fold model cross validation
(k=10) reported as root mean squared error (CV RMSE)................................ 250
Appendix 9. Table. Allometric equations following the linear model for biomass
components of 0.5 x 7.5 m alley rows of Leucaena leucocephala 6 months post
pollarding to a height of 2 m. Where (x) is the measurement used as predictor of
biomass: Root collar diameter (RCD mm, 0.05 m), diameter at breast height
(DBH mm, 1.3 m), stump diameter (SD mm, 0.3 m), crown volume (CV cm3) and
crown diameter (CD cm), (y) is the log transformation of total biomass (g), trunk
biomass (g), branch biomass (g) or leaf biomass (g), b0 as the intercept
coefficient, b1 as the slope coefficient, both reported with standard error (S.E.)
and significance (P). Coefficient of determination (R2), root mean square error of

28
model (Model RMSE, which is the same as Furnival’s Index for untransformed
data) and results of K-fold model cross validation (k=10) reported as root mean
squared error (CV RMSE). .............................................................................. 251
Appendix 10. Table. Allometric equations following the linear model for biomass
components of 0.5 x 7.5 m alley rows of Leucaena leucocephala 6 months post
pollarding to a height of 2 m. Where (x) is the root collar diameter (RCD mm,
0.05 m), or RCD and Height (cm) as predictors of biomass, (y) is the log
transformation of total biomass (g), trunk biomass (g), branch biomass (g) or
leaf biomass (g). b0 as the intercept coefficient, b1 and b2 as the slope coefficient,
both reported with standard error (S.E.) and significance (P). Coefficient of
determination (R2), root mean square error of model (Model RMSE, which is the
same as Furnival’s Index F.I. for untransformed data) and results of K-fold
model cross validation (k=10) reported as root mean squared error (CV RMSE).
........................................................................................................................ 252
Appendix 11. Table. Allometric equations following a quadratic model for biomass
components of 0.5 x 7.5 m alley rows of Erythrina berteroana 6 months post
pollarding to a height of 2 m. Where (x) is the measurement used as a predictor
of biomass: Root collar diameter (RCD mm, 0.05 m), diameter at breast height
(DBH mm, 1.3 m), stump diameter (SD mm, 0.3 m), crown volume (CV, cm3)
and crown diameter (CD cm), (y) is the log transformation of total biomass (g),
trunk biomass (g), branch biomass (g) or leaf biomass (g), b0 as the intercept
coefficient, b1 as the slope coefficient, both reported with standard error (S.E.)
and significance (P). Coefficient of determination (R2), root mean square error of
model (Model RMSE, which is the same as Furnival’s Index for untransformed

29
data) and results of K-fold model cross validation (k=10) reported as root mean
squared error (CV RMSE). .............................................................................. 253
Appendix 12. Table. Allometric equations following the quadratic model for biomass
components of 0.5 x 7.5 m alley rows of Erythrina berteroana 6 months post
pollarding to a height of 2 m. Where (x) is the root collar diameter (RCD mm,
0.05 m), or RCD and Height (cm) as predictors of biomass, (y) is the log
transformation of total biomass (g), trunk biomass (g), branch biomass (g) or
leaf biomass (g). b0 as the intercept coefficient, b1 and b2 as the slope coefficient,
both reported with standard error (S.E.) and significance (P). Coefficient of
determination (R2), root mean square error of model (Model RMSE, which is the
same as Furnival’s Index F.I. for untransformed data) and results of K-fold
model cross validation (k=10) reported as root mean squared error (CV RMSE).
........................................................................................................................ 254
Appendix 13. Table. Allometric equations following a quadratic model for biomass
components of 0.5 x 7.5 m alley rows of Inga edulis 6 months post pollarding to
a height of 2 m. Where (x) is the measurement used as a predictor of biomass:
Root collar diameter (RCD mm, 0.05 m), diameter at breast height (DBH mm,
1.3 m), stump diameter (SD mm, 0.3 m), crown volume (CV, cm3) and crown
diameter (CD cm), (y) is the log transformation of total biomass (g), trunk
biomass (g), branch biomass (g) or leaf biomass (g), b0 as the intercept
coefficient, b1 as the slope coefficient, both reported with standard error (S.E.)
and significance (P). Coefficient of determination (R2), root mean square error of
model (Model RMSE, which is the same as Furnival’s Index for untransformed
data) and results of K-fold model cross validation (k=10) reported as root mean
squared error (CV RMSE). .............................................................................. 255

30
Appendix 14. Table. Allometric equations following the quadratic model for biomass
components of 0.5 x 7.5 m alley rows of Inga edulis 6 months post pollarding to
a height of 2 m. Where (x) is the root collar diameter (RCD mm, 0.05 m), or
RCD and Height (cm) as predictors of biomass, (y) is the log transformation of
total biomass (g), trunk biomass (g), branch biomass (g) or leaf biomass (g). b0
as the intercept coefficient, b1 and b2 as the slope coefficient, both reported with
standard error (S.E.) and significance (P). Coefficient of determination (R2), root
mean square error of model (Model RMSE, which is the same as Furnival’s
Index F.I. for untransformed data) and results of K-fold model cross validation
(k=10) reported as root mean squared error (CV RMSE)................................ 256
Appendix 15. Table. Allometric equations following a quadratic model for biomass
components of 0.5 x 7.5 m alley rows of Leucaena leucocephala 6 months post
pollarding to a height of 2 m. Where (x) is the measurement used as a predictor
of biomass: Root collar diameter (RCD mm, 0.05 m), diameter at breast height
(DBH mm, 1.3 m), stump diameter (SD mm, 0.3 m), crown volume (CV cm3) and
crown diameter (CD cm), (y) is the log transformation of total biomass (g), trunk
biomass (g), branch biomass (g) or leaf biomass (g), b0 as the intercept
coefficient, b1 as the slope coefficient, both reported with standard error (S.E.)
and significance (P). Coefficient of determination (R2), root mean square error of
model (Model RMSE, which is the same as Furnival’s Index for untransformed
data) and results of K-fold model cross validation (k=10) reported as root mean
squared error (CV RMSE). .............................................................................. 257
Appendix 16. Table. Allometric equations following the quadratic model for biomass
components of 0.5 x 7.5 m alley rows of Leucaena leucocephala 6 months post
pollarding to a height of 2 m. Where (x) is the root collar diameter (RCD mm,

31
0.05 m), or RCD and Height (cm) as predictors of biomass, (y) is the log
transformation of total biomass (g), trunk biomass (g), branch biomass (g) or
leaf biomass (g). b0 as the intercept coefficient, b1 and b2 as the slope coefficient,
both reported with standard error (S.E.) and significance (P). Coefficient of
determination (R2), root mean square error of model (Model RMSE, which is the
same as Furnival’s Index F.I. for untransformed data) and results of K-fold
model cross validation (k=10) reported as root mean squared error (CV RMSE).
........................................................................................................................ 258
Appendix 17. Table. Allometric equations following the log-linear model for biomass
components of 0.5 x 7.5 m alley rows of Erythrina berteroana 6 months post
pollarding to a height of 2 m. Where ln(x) is the log transformation of the
following measurements used as predictors of biomass: Root collar diameter
(RCD mm, 0.05 m), diameter at breast height (DBH mm, 1.3 m), stump diameter
(SD mm, 0.3 m), crown volume (CV cm3) and crown diameter (CD cm) ln(y) is
the log transformation of total biomass (g), trunk biomass (g), branch biomass
(g) or leaf biomass (g). b0 as the intercept coefficient, b1 as the slope coefficient,
both reported with standard error (S.E.) and significance (P). Coefficient of
determination (R2), lambda (λ) and standard deviation (S.D.) as back
transformation according to Marklund (1987), Furnival’s Index (F.I.) according to
Kershaw et al. (2016), root mean square error of model (Model RMSE) and
results of K-fold model cross validation (k=10) reported as root mean squared
error (CV RMSE). ............................................................................................ 259
Appendix 18. Table. Allometric equations following the log-linear model for biomass
components of 0.5 x 7.5 m alley rows of Erythrina berteroana 6 months post
pollarding to a height of 2 m. Where ln(x) is the log transformation of root collar

32
diameter (RCD mm, 0.05 m), or RCD and Height (cm) as predictors of biomass.
ln(y) is the log transformation of total biomass (g), trunk biomass (g), branch
biomass (g) or leaf biomass (g). b0 as the intercept coefficient, b1 as the slope
coefficient, both reported with standard error (S.E.) and significance (P).
Coefficient of determination (R2), lambda (λ) and standard deviation (S.D.) as
back transformation according to Marklund (1987), Furnival’s Index (F.I.)
according to Kershaw et al. (2016), root mean square error of model (Model
RMSE) and results of K-fold model cross validation (k=10) reported as
root mean squared error (CV RMSE). ............................................................. 260
Appendix 19. Table. Allometric equations following the log-linear model for biomass
components of 0.5 x 7.5 m alley rows of Inga edulis 6 months post pollarding to
a height of 2 m. Where ln(x) is the log transformation of the following
measurements used as predictors of biomass: Root collar diameter (RCD mm,
0.05 m), diameter at breast height (DBH mm, 1.3 m), stump diameter (SD mm,
0.3 m), crown volume (CV cm3) and crown diameter (CD cm) ln(y) is the log
transformation of total biomass(g), trunk biomass (g), branch biomass (g) or leaf
biomass (g). b0 as the intercept coefficient, b1 as the slope coefficient, both
reported with standard error (S.E.) and significance (P). Coefficient of
determination (R2), lambda (λ) and standard deviation (S.D.) as back
transformation according to Marklund (1987), Furnival’s Index (F.I.) according to
Kershaw et al. (2016), root mean square error of model (Model RMSE) and
results of K-fold model cross validation (k=10) reported as root mean squared
error (CV RMSE). ............................................................................................ 261
Appendix 20. Table. Allometric equations following the log-linear model for biomass
components of 0.5 x 7.5 m alley rows of Inga edulis 6 months post pollarding to

33
a height of 2 m. Where ln(x) is the log transformation of root collar diameter
(RCD mm, 0.05 m), or RCD and Height (cm) as predictors of biomass. ln(y) is
the log transformation of total biomass(g), trunk biomass (g), branch biomass (g)
or leaf biomass (g). b0 as the intercept coefficient, b1 as the slope coefficient,
both reported with standard error (S.E.) and significance (P). Coefficient of
determination (R2), lambda (λ) and standard deviation (S.D.) as back
transformation according to Marklund (1987), Furnival’s Index (F.I.) according to
Kershaw et al. (2016), root mean square error of model (Model RMSE) and
results of K-fold model cross validation (k=10) reported as root mean squared
error (CV RMSE). ............................................................................................ 262
Appendix 21. Table. Allometric equations following the log-linear model for biomass
components of 0.5 x 7.5 m alley rows of Leucaena leucocephala 6 months post
pollarding to a height of 2 m. Where ln(x) is the log transformation of the
following measurements used as predictors of biomass: Root collar diameter
(RCD mm, 0.05 m), diameter at breast height (DBH mm, 1.3 m), stump diameter
(SD mm, 0.3 m), crown volume (CV cm3) and crown diameter (CD cm) ln(y) is
the log transformation of total biomass(g), trunk biomass (g), branch biomass (g)
or leaf biomass (g). b0 as the intercept coefficient, b1 as the slope coefficient,
both reported with standard error (S.E.) and significance (P). Coefficient of
determination (R2), lambda (λ) and standard deviation (S.D.) as back
transformation according to Marklund (1987), Furnival’s Index (F.I.) according to
Kershaw et al. (2016), root mean square error of model (Model RMSE) and
results of K-fold model cross validation (k=10) reported as root mean squared
error (CV RMSE). ............................................................................................ 263

34
Appendix 22. Table. Allometric equations following the log-linear model for biomass
components of 0.5 x 7.5 m alley rows of Leucaena leucocephala 6 months post
pollarding to a height of 2 m. Where ln(x) is the log transformation of root collar
diameter (RCD mm, 0.05 m), or RCD and Height (cm) as predictors of biomass.
ln(y) is the log transformation of total biomass(g), trunk biomass (g), branch
biomass (g) or leaf biomass (g). b0 as the intercept coefficient, b1 as the slope
coefficient, both reported with standard error (S.E.) and significance (P).
Coefficient of determination (R2), lambda (λ) and standard deviation (S.D.) as
back transformation according to Marklund (1987), Furnival’s Index (F.I.)
according to Kershaw et al. (2016), root mean square error of model (Model
RMSE) and results of K-fold model cross validation (k=10) reported as
root mean squared error (CV RMSE). ............................................................. 264

35
CHAPTER I
AMAZON PASTURES: THE
POTENTIAL FOR SILVOPASTORAL SYSTEMS
1. Livestock Production
The world’s population is predicted to reach 9.6 billion by the year 2050
(Alexandratos and Bruinsma, 2012). Meat and milk currently provide a third of global
protein intake and demand for livestock products is expected to increase by 70 to
80% by the year 2050 (Rosegrant et al., 2009; Herrero, 2015). Pasturelands cover
35 million km2, or 68% of the total global land area dedicated to agricultural
production (White et al., 2000). The rising demand for livestock products is likely to
be met with a concurrent expansion of livestock production into wilderness areas
(Thornton and Herrero, 2010).
To date, deforestation has resulted in the loss of more than a third of all global forest
(Hansen et al., 2013). An example of this is the 413,506 km2 that have been
deforested since 1988 in the Brazilian Amazon to be replaced by pastures for the
production of cattle (Downing, 2019; PRODES, 2019). Central and Latin America
have one of the highest rates of land conversion for livestock production (Wassenaar
et al., 2007). It is estimated that the livestock sector drives climatic change
contributes 14.5% of the total anthropogenic greenhouse gases (GHG) emissions
(Gerber et al., 2013). It is a sector that may struggle to adjust to climatic change, as

36
livestock producers must contend with predictions of increased pressure on
productive land, water shortages and declining forage production under rising
temperatures (Lee et al., 2017; Fellman et al., 2018).
In order to meet the growing demand for livestock products, efforts should focus on
intensifying land that is already dedicated to livestock production. This would have
the dual benefit of avoiding further emissions of GHGs and ensure the preservation
of biodiversity and wilderness areas (Godde, 2018). Increasing the productivity per
hectare of land in a sustainable way is known as sustainable intensification
(Carvalho et al., 2018). Interest has been growing in the potential of sustainable
intensification to integrate multiple land uses that can result in increases in economic
and environmental benefits from the same unit of land (Schultze-Kraft et al., 2018).
Integrating cattle with crops, rotational stocking systems and the use of improved
forages have led to advances in the intensification of cattle farming (Bogaerts et al.,
2017). Farms that integrate trees and shrubs in a practice known as silvopasture
have been documented to have substantial increases in on-farm productivity and
observed benefits to biodiversity (Murgueitio et al., 2011). A critical challenge facing
humanity is to reconcile the production of food for a growing population with the
protection of natural ecosystems while confronted with uncertain predictions of the
impacts of climatic change (Rojas-Downing et al., 2017).

37
2. Introduction to Silvopastoral Systems
Agroforestry systems, as defined by the Food and Agriculture Organisation, are
“traditional and modern land-use systems where trees are managed together with
crops and/or animal production systems in agricultural settings” (FAO, 2016). Many
terms are used to describe arrangements where trees are incorporated into livestock
production systems. Hecht (1982) makes a distinction between “silvopastoral”
systems, “a situation where animals graze a ground cover crop grown under
plantation trees,” and “agropastoral” systems, which “encompass livestock as well as
trees grown for food (human or animal)”. Both may be viewed as “integrated farming
systems” (Hecht, 1982). For the purposes of this discussion, silvopastoral systems
are defined according to Kaur et al. (2002) as an agroforestry practice that integrates
livestock, forage production, and forestry on the same land-management unit.
The incentive to increase production on existing lands is growing as property rights
in the Amazon are strengthening and landowners reach the boundaries of their
properties. Brazil’s commitment to the 15th United Nations Conference on Climate
Change in 2009 resulted in the Low Carbon Agriculture plan, a national credit
incentive that has supported the conversion to silvopastoral systems of
approximately 2 million hectares (Embrapa, 2016; Santos et al., 2018). Many
systems aim to coproduce timber, and often favor exotics such as Tectona grandis
and Eucalyptus.
Farmers can manage trees to provide a range of products and services.
Management of trees commonly include planting arrangements such as living fence
posts, fodder banks or alley farming, a term defined by Kang et al. (1990) as “the

38
cultivation of food crops between hedgerows of multipurpose trees” (Russo and
Botero, 1994; Sanchez, 2002). Appropriate management of agroforestry systems
can help farmers increase production of crops, fuelwood, timber and fodder (Russo
and Botero, 1994). Additionally, a range of ecosystem services are associated with
the presence of trees in agricultural systems, some of which directly benefit the
productivity of agri-ecosystems, wheras others accrue as less-tangible global good
by, for example, increasing the amount of carbon dioxide sequestered.
The presence of trees helps to maintain and improve soil fertility (Hohnwald et al.,
2005; Murgueitio et al., 2011). Trees can act as ‘nutrient pumps’, as they have the
advantage of accessing nutrients from deep soil horizons (Scholes and Archer 1997;
Sanchez, 2002), that are recycled into the system as leaf or root litter and can
contribute to crop available nutrients (Menezes et al., 2002). Nitrogen fixing trees are
commonly employed in agricultural systems as they can increase the nitrogen inputs
of a system (Dommergues, 1987). Some tree legumes have an advantageous
relationship with rhizobia; bacteria that infect roots and form nodules which fix
nitrogen (Downie, 2014). The extent to which tree legumes can access or contribute
nutrients to a given system is dependent on, amongst other factors, mycorrhizal
associations, soil, climate, species and management (Szott et al., 1991). Trees also
promote complex soil food webs (Montagnini and Ashton, 1999) and increase the
habitat available to ecosystem engineers such as decomposers, predators and
parasitoids that control harmful insects (Giraldo et al., 2011; Murgueitio et al., 2011).
Silvopastoral systems can offer improved production and sustainability over
traditional pasture systems. In temperate climates, some silvopastoral systems have

39
been productive for over 4,500 years (Stevenson and Harrison, 2014). The presence
of shade trees can reduce heat stress in cattle and may increase their weight, milk
production or reproductive success. The inclusion of trees can provide a useful
adaptation among smallholder livestock farmers in response to climatic changes. For
example, in times of drought, fodder production by trees may continue when pasture
productivity decreases due to water stress and therefore the (Altieri et al., 2015).
Silvopastoral systems also generate additional products that can increase a farmer’s
income, such as timber, fuelwood, fruits and fodder for cattle. In the Amazon there
are low stocking densities, typically less than one cattle per hectare (Hohnwald et al.,
2006). A pasture which incorporated tree legumes (Leucaena leucocephala) in
Western Australia reported record liveweight gains for cattle and a stocking rate of 3-
4 animals per hectare (Jones, 1986 in Russo and Botero 1994). In the South Eastern
United States the native and endangered longleaf pine Pinus palustris was used
within a silvopastoral system which subsequently produced more profits than either
traditional ranching or traditional forestry (Stainback and Alavalapati, 2004). In this
approach, a threatened species was incorporated into a silvopastoral system which
resulted in both a profitable farming systems and the continued promotion of the
existence of Pinus palustris.
Silvopastoral systems also have considerable potential as a landscape level tool for
conservation. The increased complexity of silvopastoral systems results in a greater
provision of ecosystem services and increased habitat for wildlife (Mcadam et al.
2007). In areas of fragmented forests, silvopastoral systems can increase
connectivity and act as wildlife corridors (Ibrahim et al., 2006). Silvopastoral systems
may sequester more carbon than traditional pasture (Haile et al. 2010). The inclusion

40
of certain forages containing tannins may reduce methane emissions from livestock
production (Hess et al., 2005) and control livestock parasites (Nguyen et al., 2005).
Silvopastoral systems represent an opportunity for sustainable intensification and
livelihood diversification, while simultaneously providing benefits for the provision of
ecosystem services and biodiversity conservation. Such systems can challenge the
traditional view of livestock production as the antithesis of biodiversity conservation.
Livestock production in the Amazon is constrained by the complex agro-ecological
problem of early pasture degradation. Within ten years pasture may reach an
advanced stage of degradation (Martínez and Zinck, 2004). Some direct causes at
the farm level of this phenomenon include decreasing soil fertility, water stress,
insect pests, inadequate grass species, weed invasion and overstocking, whereas
underlying causes of pasture degradation include low return yields, lack of technical
knowledge, perverse policy environments, and insecure property rights (Hohnwald,
2016). Estimates of the extent of pasture degradation are not consistent, but authors
indicate that up to half of the pastures sown in Brazil’s rain forest may be degraded
to some degree (Serrao et al., 1993). The presence of forage trees can enhance the
productivity of pasture systems compared to grass monocultures, with effects
varying dependent on the arrangement of trees, selection of species and soil
conditions of the site (Szott et al., 1991).
Relatively few studies have attempted to increase the productivity of Amazonian
livestock production systems using silvopastoral techniques. A successful
rehabilitation from bulldozed land to a productive silvopastoral system is reported for
a study in Yurimaguas in 1988. The experiment established Centrosema

41
macrocarpum as a forage ground cover crop between peach palm Bactris gasipaes.
Eighteen months after establishment, the Centrosema macrocarpum was grazed.
Average increase in live-weight gains of the cattle was 445 g-1animal-1 per day, an
improvement over traditional grazing systems in the region. Soil bulk density,
mechanical resistance, acidity and aluminium saturation decreased considerably
(Arevalo et al., 1998). Silvopasture can therefore present a unique opportunity to
rehabilitate degraded pastureland and increase the productivity of marginal land.
Such studies reinforce the potential for silvopastoral systems to outcompete
traditional grazing management in the tropics while increasing the sustainability of
livestock production. A framework to assist in the evaluation of candidate species for
inclusion in silvopastoral systems could help interested stakeholders participate in
trialling species they identify as having potential, while generating data that
documents the success of the species.
3. Selection and Management of Species for Silvopastoral Systems
Certain characteristics are shared by species that perform well in silvopastoral
systems; they fix nitrogen, are fast growing, demonstrate high foliage productivity
and nitrogen content, are palatable and readily eaten by animals when fresh,
nutritious for livestock, exhibit tannin content lower than 5%, can withstand trampling,
coppicing, heavy grazing pressure or even complete defoliation, show rapid rates of
regrowth from numerous growing points on the remaining stems, develop a vigorous
tap root, retain their leaves in the dry season, have small leaflets and open crowns
which allow light to reach the ground (Devendra, 1990; Arevalo et al., 1998;
Sanchez, 2002; Vitti et al., 2005; ; Hohnwald, 2016).

42
Appropriate species for silvopastoral systems may be specific to the local
environmental and climatic variables that can affect production. What is proven to
work in one context can fail in another. For example, Leucaena leucocephala is a
nitrogen fixing tree leguminous native to the Yucutan peninsular of Mexico and used
worldwide as a forage tree. When trialed in the acidic Utisols of Yurimaguas in the
Peruvian Amazon failed to be productive due to high levels of aluminum saturation
(Sanchez, 1987 in Loker, 1994). On acidic, low base status soils species such as
Fleminga macrophylla and Erythrina poeppigiana have been shown to be more
productive than L. leucocephala (Kass et al., 1992).
Physical and chemical soil characteristics effect the plants and species they support.
It is important to understand the effects of soil characteristics on a species success.
Degraded, acidic soils are prevalent across the Americas, yet a paucity of studies
have evaluated the adaptability native tree species to these conditions for
consideration in incentive-supported reforestation schemes (Tilki and Fisher, 1998).
The majority of research has focused on Costa Rica, with results demonstrating the
importance of using locally adapted species for rehabilitation of degraded lands
(Montagnini and Sancho, 1990; Perez et al., 1993; González and Fisher, 1994).
Despite the paucity of studies that consider the Amazon, much of the knowledge that
has been acquired regarding the impact of soils and associated biota on trees is
transferrable. For example, data acquired by Carpenter et al. (2004) suggested
survival and growth of trees in their study were negatively correlated with soil
erosion. This observation may be due to the negative relationship between
arbuscular mycorrhizal fungi and levels of soil erosion (Carpenter et al., 2001). Trees

43
associated with arbuscular mycorrhizal fungi have shown improved utilisation of
phosphorous, a nutrient that is frequently limiting to plant growth in the tropics.
Inoculation with appropriate strains of arbuscular mycorrhizal fungi is important as it
can significantly increase plant survival and productivity.
Characteristics that may increase success of a species in the context of pasture
lands in the Amazon include tolerance to high levels of irradiance and aluminium,
fast growth and biomass production that enables trees to outcompete weeds and
certain species may be able provide an advantageous association with neighbouring
trees or crops through their ability to fix atmospheric nitrogen. Nichols et al. (2001)
studied techniques to enhance the growth of Terminalia amazonia on degraded
Utisols in Costa Rica. Four years after planting, T. amazonia inter-planted with the
leguminous nitrogen fixing tree Inga edulis (with no fertiliser applied) was 15% taller
than all other treatments, including T. amazonia treated with annual applications of
fertiliser. Transfer of nitrogen from nitrogen fixing leguminous trees to pasture
grasses has also been recorded (Sierra and Nygren, 2006; Sierra et al., 2007).
Experimental evaluation of local species led one Colombian farmer to discover the
silvopastoral benefits of an endemic and rare legume, Mimosa trianae, which
outperformed the other species in the trial and shows promise for the development of
silvopastoral systems of the Andean foothills (Calle et al., 2012). Native species
consistently show promising results in trials, but are frequently overlooked.
Butterfield (1995) screened 84 species of hardwoods for forestry development
potential in Costa Rica and found that of the species whose performance was in the
top 25%, 67% were native. In one of the only studies of its kind, research that

44
considered capoeira (secondary vegetation) species as supplementary forages in
Northeastern Brazil highlighted many species that could be of potential use to
silvopastoral arrangements in the Amazon. Hohnwald (2016) used a randomized
block design to assess relative palatability, height, biomass production, and chemical
composition of foliage. Their results highlighted the potential of unexplored species
present in diversity of the capoeira for formal integration into livestock production.
The promising results of this research suggest similarly useful but unexplored plant
capital may exist across Amazonia.
The International Livestock Centre for Africa defines four management strategies for
silvopastoral systems: fodder tree banks, intensive feed gardens, alley grazing via a
rotational system or alley grazing via a permanent system (Reynolds and Cobbina,
1992). ‘Fodder tree banks’ or ‘tree protein banks’ are closely spaced planting
arrangements at high densities. Livestock access to the trees can be controlled by
fencing. Fodder banks and intensive feed gardens incur additional management
inputs such as labour but offer opportunities for greatly increased yields due to
higher planting densities (Reynolds and Cobbina, 1992). More than 10,000 fodder
shrubs or trees per ha-1 may be considered an ‘intensive’ silvopastoral system
(Murgueitio et al., 2011). Hedge (1983) recommended a density of 10 to 15 trees per
m2 for Leucaena leucocephala (Devendra, 1990). Pathak et al. (1980) report higher
yields from trees at a density of 40,000 trees ha-1 than 15,000 trees ha-1 (Sanchez,
2002). Savory and Breen (1979) found a planting density of 60,000 trees ha-1
produced greater yields than 10,000 or 30,000 trees ha-1 (Sanchez, 2002).
Intensive feed gardens are cut and carry systems similar to fodder tree banks.

45
However, in the intensive feed garden system, livestock are kept separate from the
forage plants. Livestock can be housed and fed in stalls, or pasture grazed and
offered supplementary forage. An example of management of this system may
include closely spaced hedgerows of forage plants (2 - 4 m spacing) with grass sown
between rows (Reynolds and Cobbina, 1992).
A rotational system of alley grazing uses a wider spacing between hedgerows (3 - 4
m) that livestock graze directly. Livestock are rotated between plots of forage plants,
allowing plants time to rest. For forage tree Gliricidia sp., a suitable rotation may be 2
weeks of direct grazing followed by 8 to 10 weeks of rest period (Reynolds and
Cobbina, 1992). The alley grazing-rotational system has been used successfully in
El Hatico nature reserve in Colombia, where conventional ranching management of
pasture was replaced with an intensive silvopastoral system with a resultant increase
in milk production of 130% (Murgueitio et al., 2011).
A permanent system of alley grazing may use the widest hedgerow spacing (7-10
m). Free ranging livestock graze the system directly. Defoliation intensity of fodder
trees is offset by the higher palatability of grasses. Knowledge of the effect of trees
on animal nutrition and palatability is critical as care must be taken to manage the
exposure of livestock to excessive intake of secondary compounds (e.g. Mimosine in
the highly palatable Leucaena leucocephala) (Reynolds and Cobbina, 1992).
Of particular relevance to the alley grazing systems are the interactions between
trees and pasture grasses. In tree-pasture systems there may be competition
between the trees and the pasture for light, moisture and nutrients (Sousa et al.,

46
2015). Selecting deep-rooted tree species may reduce this competition. In addition,
shading caused by trees can have negative impacts on the productivity of pasture,
which may reduce potential stocking capacity for livestock production (Parsons et al.,
1983; Jackson and Ash, 1998). For this reason, in alley systems, hedgerows of
forage trees can be planted on east-west lines to minimise pasture shading. Pasture
productivity may also be greater in silvopastoral systems (Sousa et al., 2015). As an
additional benefit, grass species adapted to shade generally have improved nutrient
composition than species tolerant to high levels of irradiance (Russo and Botero,
1994). At a planting density of 6 x 6 m in Costa Rica, the legume tree Erythrina
poeppigana (subfamily Fabeaceae) was shown to positively affect yields of some
adjacent pasture grasses species such as Panicum maximum, Brachiaria brizantha,
B. humidicoia and Cynodon nlemfuensis, but decrease yields of other species such
as B. dictyoneura and Pennisetum purpureum (Russo and Botero, 1994). In practice,
any positive effect of trees on pasture productivity may be negated at high stocking
densities combined with low canopy cover as livestock may seek shade and
therefore inhibit tree growth and degrade shaded grasses and soil.
A range of destructive and non-destructive techniques have been developed to
determine pasture productivity. Non-destructive techniques can also give good
estimates of pasture yield; Mannetje (2000) categorised these indirect methods as
visual estimation (including the pasture plate meter and pasture capacitance meter
techniques) and estimation based on grass height and density measurements.
Destructive techniques are generally accepted to be a more reliable indicator of
pasture productivity, but it is important to include an adequate number of samples
(Wilm et al., 1944; Frame, 1981). Grazed swards display inherent variation, which

47
means a large number of samples are required (Shaw et al., 1976; Vermeire et al.,
2002). Sampling is usually done by quadrats; it has been found that, particularly in
row sown pastures, rectangular quadrats exhibit decreased variation than square
ones (Mannetje, 1978). The cutting height of samples also affects estimates of
pasture yield. Thomas and Laidlaw (1981) recommend that for forage estimates,
cutting height be selected at minimum grazing height used by the type of animals the
experiment considers and the experiment’s objectives. This is in contrast to Hodgson
(1979) who suggests cutting at ground level. Ultimately, the technique used to
determine pasture productivity in a study will be determined by time and cost
constraints.
Cattle browse on trees and shrubs, and trees and shrubs can be managed as
resources for cattle production. There is need for more research into which species
cattle browse, and the amount and type of browse consumed by cattle of each
species. To determine how to best document the interaction of cattle with browse at
the species level it is necessary to review the existing literature on how plants
respond and adapt to the herbivory. Research on herbivory is highly relevant to the
present study, yet such information is scarce, particularly at the species level.
Therefore, the following discussion seeks to understand the underlying processes by
which plants respond to defoliation. Trees and shrubs show varying levels of
adaptive responses to defoliation, which may be a result of herbivory by arboreal or
terrestrial animals, or by humans harvesting foliage or wood for a variety of purposes
(e.g. coppicing) (Hardesty and Box, 1988). Defoliation may affect the productivity of
trees or cause mortality in species that are not adequately adapted to the type of
defoliation they are subjected to. The response to defoliation is species specific, and

48
can also be influenced by the type of defoliation, intensity and/or frequency of
defoliation and environmental factors such as water availability in the period of
defoliation. Management of species for their foliage or wood therefore relies on an
understanding of the implications of different management strategies on productivity.
Erdmann et al. (1993) highlight the difficulty of comparing the biomass productivity of
trees between studies due to inter-study inconsistencies and incomplete information,
as important parameters such as the age of plants, cutting heights, plant densities
and plant component biomass are not always specified (Erdmann et al., 1993).
The majority of work on defoliation has focused on effects of defoliation frequency
and defoliation intensity. However, the ‘cutting’ height has also been shown to be of
importance in the management of some species. For example, Sesbania grandiflora
experiences mortality following repeated cutting of the main stem (Ella et al., 1989),
but has been reported to survive well if only side branches are removed (Stür et al.,
1994). Dry matter productivity for Leucaena leucocephala was shown to vary with
cutting height by Osman (1981). Osman (1981) reported greater yields from a cutting
height of 90 cm compared to heights of 15 cm or 150 cm. Similarly, variations in yield
as a response to cutting height have been shown to exist in other forage trees such
as Calliandra calothyrsus (Catchpoole and Blair, 1990). Other species, such as
Gliricidia sepium, exhibit less response to cutting height. Erdmann et al. (1993)
determined a lower yield at a cutting height of 0 cm but no significant difference in
dry matter yield between a cutting height of 25 cm or 100 cm.
Defoliation intensity can be conceptualized as a measure of the amount of
vegetation removed relative to the total plant biomass in a defoliation event.

49
Defoliation frequency is the time between defoliation events. Intensity and frequency
of defoliation are often combined and compared in studies. Abbott et al. (1993)
developed a ‘defoliation index’, where visual estimates of manual defoliation
intensities were made and combined with defoliation frequency. For example, a
defoliation index of 100 would be derived from four seasonal annual defoliations of
25%, or one single defoliation event of 100%. They conclude, in the case of
Eucalyptus marginata, that repetitive, low-intensity defoliation has a greater impact
on growth (measured by stem diameter) than defoliation events at low frequency but
with high-intensity. Teague (1989) found Acacia karroo trees response to more
frequent defoliations (8 weeks versus 12 weeks) resulted in a higher yield, and that
they adjusted to cope with frequent defoliation. In contrast, Erdmann et al. (1993)
found Gliricidia sepium produced low biomass when cut at 3-week intervals, which is
consistent with the findings of Ella et al. (1989) and Duguma et al. (1988) for
Gliricidia sepium and also those of Guevarra et al. (1978) for Leucaena
leucocephala.
Studies that neglect to compare different defoliation frequencies and intensities may
overlook important inter-species variation important for the appropriate management
of individual species. By comparing growth rates with change in mean growth rates
management strategies can be tailored to optimise the cutting interval and result in
higher productivity and less risk of mortality (Stür et al., 1994). Ella et al. (1991)
demonstrated the benefit to productivity of a long period before the first cut or
harvest of forage trees by showing that age at first harvest was positively related to
yield at subsequent harvests. They found shorter cutting intervals were positively
correlated with an increase in leaf biomass, whereas longer cutting intervals

50
generally stimulated production of woody biomass (Ella et al., 1991). There are
significant differences in the effects of browsing or coppicing on species (Hardesty
and Box, 1988). Browsing is often less severe than coppicing as the ratio of wood to
leaf area removed is generally lower. In turn, this may mean there are more buds
and leaf regrowth area left from which trees may regenerate foliage. As Hardesty
and Box (1988) assert in their study of the effects of coppicing and browsing on
species in North-eastern Brazil, there is weak evidence to support the assumption
that defoliation may stimulate growth which exceeds that of plants which do not
experience defoliation. This response is highly dependent on the species and the
context in which they are found.
Only by measuring these characteristics and adaptations can species and
appropriate management strategies be identified which are likely to result in
increased productivity of browse components for animal consumption, of interest to
the development of silvopastoral systems. Furthermore, context specific factors such
as soil health, climate or presence of pests may result in considerable intra-species
variations and should not be ignored. Trials that evaluate these characteristics of
species must be carried out in a context appropriate to the application of the results.
Methodologies that experimentally evaluate the impact of browsing on foliage are
needed in order to measure the context specific potential production of browse.

51
4. Traditional Livestock Production Models And The Potential For Silvopastoral
Systems: Influence On Soil Health
Silvopastoral systems have been proposed as a method for rehabilitating degraded
soils as they incorporate trees, which have been shown to maintain and improve soil
characteristics (Hohnwald et al., 2005; Murgueitio et al., 2011). Trees can benefit the
physical properties of soils and can reduce soil erosion events caused by runoff
(Murgueitio et al., 2011) and leaching (Bishop, 1983), enhance water infiltration
(Ilstedt et al., 2007), counteract compaction (Murgueitio et al., 2011) and increase
soil aeration (Bishop, 1983). Silvopastoral systems may therefore present a positive
contribution to soil health compared to current models of livestock production in the
Amazon, which typically result in pasture degradation (Martínez and Zinck, 2004).
The magnitude of the effect of livestock on soils has been related to stocking rate,
grazing system, soil texture and soil moisture content (McCalla et al., 1984; Van
Haveren, 1983; Martínez and Zinck, 2004). The success of post pasture
abandonment recovery of soils has been shown to vary with the prior intensity of
pasture use (Uhl et al., 1988). An important factor to consider when selecting
species for rehabilitation potential is the influence of a species on soil properties
(Tilki and Fisher, 1998).
Attempts to quantify levels of pasture degradation have used soil physical
parameters such as bulk density and infiltration rate as indicators of degradation.
Bulk density is related to soil compaction and measurements are simple and cost-
efficient when compared to other methods developed to measure compaction such
as computerised tomography (Pedrotti et al., 2005) or sensor based technology
(Quraishi and Mouazen, 2013). Bulk density is the weight of dry soil divided by a

52
known given volume; it is typically measured by extracting soil cores in a cylinder of
known volume, Guo et al. (2016) used a cylinder with a volume of 100 cm3 and
Pedrotti et al. (2005) a volume of 90 cm3. It is generally accepted that larger volume
cylinders reduce edge effects of this sampling technique.
Soil with a bulk density greater than 1.6 g cm3 begins to restrict root growth in some
species. An increase of 30% in bulk density compared to forest has been used as a
threshold value corresponding to severe soil degradation of pastures (Hecht, 1982).
Increased bulk density is positively related to increases in the intensity of grazing
regimes. High bulk density has a negative effect on pasture production and has been
shown to reduce the protein content of pasture grasses and is associated with
increased presence of weeds (McCarty and Mazurak 1976; Martínez and Zinck
2004). Reátegui et al. (1990) found a 20% increase in bulk density when
continuously grazed pastures were compared to pastures protected from grazing in
the Peruvian Amazon. In another study into Amazon pasture degradation, Moraes et
al. (1996) found after nine years of grazing a 26% increase in bulk density compared
to forest. Stephenson and Veigel (1987) found it took 16 months to recover initial
bulk density once grazing had ceased, which is consistent with Alegre and Cassel
(1996) who found the effect of grazing on bulk density was reversible once livestock
were removed. McCalla et al. (1984) found bulk density can predict infiltration rate
across a variety of soils and ground cover characteristics. Viana et al. (2014) found
soil bulk density was variable across a gradient of land rehabilitation in the Amazon.
Bulk density was highest in degraded pasture, decreased in two rehabilitation trials
using woody species and was lowest in natural forest. Total soil nitrogen
concentration increased along the same gradient. They proposed degradation and

53
rehabilitation measurements of Amazon pastures can be derived from measures of
soil total nitrogen and bulk density (Viana et al., 2014).
Water infiltration is a measure of the time taken for water to infiltrate a soil. It is an
important factor governing the runoff of water over land. If infiltration rate is low then
the probability of flooding and soil erosion are increased. Measurement of infiltration
rates is commonly carried out using either single or double ring methodologies.
However, results may be affected by soil disturbance from ring installation and an
unrealistic water pressure (Benegas et al., 2014). The presence of livestock in
pastures has been shown to decrease infiltration rate, whereas tree presence has
been shown to increase infiltration rate (Benegas et al., 2014). Basic infiltration has
been shown to decrease 98% when grazed pastures are compared to intact forest in
studies on fine textured soils (Martínez and Zinck, 2004). Martínez and Zinck (2004)
observed that the majority of changes in infiltration rate occur within the first 3 years
of pasture establishment. Alegre and Cassel (1996) found that, in the Amazon,
infiltration rates were decreased after five years of pasture establishment. Benegas
et al. (2014) found infiltration rates in pastures were positively correlated with
proximity to trees.
Soil organic matter (SOM) refers to the fraction of soil that is derived from living or
once living plant or animal organisms. SOM has an influence on the physical,
chemical and biological properties of soils (Stolt and Lindbo, 2010). Soil SOM
contributes to the quantity and diversity of microbes in soils. A relatively high soil
organic matter is considered beneficial for agriculture as the presence of SOM can
increase the physical properties of the soil, such as the aeration and water infiltration

54
rate. SOM can alter the soils chemical properties, a soil with high SOM is more likely
to have a higher cation exchange capacity than a soil with low SOM, and be more
likely to resist changes of pH. Buschbacher et al. (1988) found that biomass was not
correlated with nutrient content in regenerating pastures, but did not consider soil
organic matter in their study. This contrasts with the findings of Montagnini and
Sancho (1990), who studied the impacts of native trees on the abandoned pasture
soils of Costa Rica. They used the Walkley-Black acid digestion method to determine
soil organic matter. After 2.5 years they reported an increase in soil organic matter
from 4.83% in a grass control to 5.31-6.60% in the upper 15 cm of soil beneath
native trees. They found a positive relationship between the soil organic matter
content and sum of bases, which highlights the importance of soil organic matter to
cation retention capacity. They observed that land use practices that negatively
affect the level of organic matter, such as the use of fire, were less sustainable than
practices that have been shown to recover soil organic matter such as planting trees
which can increase soil organic matter through the inputs of leaf litter and root
turnover.
Sustainable systems are often managed with the goal of balancing the nutrient cycle.
It has been proposed that trees can act as ‘nutrient pumps’, accessing reservoirs of
nutrients that may be present in lower soil horizons and distributing them to the
surface as litter. Kumar et al. (1998) present indicative results that when soil is
associated with trees it has a higher organic carbon, nitrogen, phosphorous and
potassium content when compared with soils without trees. Nitrogen fixing species
increased soil nitrogen, whereas levels of soil organic carbon were highest under
Leucaena leucocephala and Acacia auriculiformis. However, as discussed by Wang

55
et al. (1991), there are noticeable differences in the nutrient cycling of systems
where the tree component is produced with the intention of improving soil
characteristics and systems where biomass is produced with the intention of removal
from the site. Nutrient concentrations are not consistently distributed between
different plant components and in addition this distribution may vary considerably
between species (Wang et al., 1991). This assertion is consistent with the findings of
Achat et al. (2015), who found removal of biomass reduced the amount of organic
matter in soils, reduced total and available nutrients in soils and resulted in increased
acidification of the soils. Boddey et al. (2004) studied nitrogen cycling in grazed
Brachiaria pastures in Brazil and found cattle unevenly deposit nitrogen acquisitions,
the highest concentrations were found in rest areas and proximate to drinking
troughs. This means nutrient accounting related to the removal of biomass is a
complex, but important requisite of designing any truly ‘sustainable’ system. In the
context of silvopastoral systems, understanding the nutrient cycle is additionally
complicated by the animal component that may result in nutrient removal or their
unequal deposition (Boddey et al., 2004).
5. Evaluating the Forage Potential of Tree Species
Tree forages are an important source of feed for ruminants around the world (Dynes
et al., 2002). In order to intensify production of livestock systems it is necessary to
understand how the quantity and quality of inputs govern outputs. Therefore, it is
crucial to evaluate forages to understand their value to livestock. Forage value,
sometimes referred to as nutritive value, refers to the relationship between a unit of
forage intake and a unit of animal productivity (Dynes et al., 2002). ‘Feeding value’ is

56
used to describe the amount of voluntary intake of forage by an animal and the
forage nutritive value. Ulyatt (1973) calculated that variation in intake could account
for up to 70% of variation in feeding value. Weston (1982; 1985; 1996) proposed the
“forage consumption constraint” in reference to the phenomena whereby animals’
intake is less than required for optimal output. It was proposed the constraints
include environmental stress, forage components which complicate removal from the
rumen, low palatability and high harvesting expenditure. Understanding constraints
to intake of forages is important to the goal of increasing animal productivity, as
forage of high nutritive value is of limited use if it is not palatable to animals.
Palatability can be conceptualised as an animal’s choice of feed, and the feed intake
displayed by an animal. The amount of forage eaten by animals has not been related
to any specific or fixed pattern of its nutrient composition (Crampton, 1957).
Therefore, palatability can only be evaluated through feeding trials, but this gives no
indication as to the digestibility of the forage. Digestibility is “a measure of the
availability of nutrients” (Khan et al., 2003); yet, digestibility is not always an accurate
predictor of nutritive value, or vice versa (Dynes et al., 2002). In vivo, in vitro and in
sacco techniques are methods that have been used to evaluate the digestibility of
forages. The ‘total collection’ technique is commonly considered the most reliable
measure of digestibility (Khan et al., 2003). This approach uses feeding trials and
records feed intake, refusals and faecal output. Minson et al. (1964) described
digestibility as a function of the percentage of faecal output relative to the weight of
organic matter consumed. Feeding trials are costly, time consuming, and while a 7 to
21 day period is allowed for cows to adjust to a feed, their intake may still be affected
by environmental stress (Khan et al., 2003). This method of estimating digestibility

57
also suffers from an inherent animal-to-animal variation of 1 to 4% (Khan et al.,
2003). Other in vivo approaches include the use of markers by identifying either a
naturally indigestible constituent of the feed or applying external substances. The
presence of markers can then be identified from faecal samples.
In vitro digestibility and in sacco degradability can be used to indicate the nutritional
value of forage (Rittner and Reed, 1992). Tilley and Terry (1963) used a method for
in vitro digestibility by incubating the forage in rumen fluid followed by acid pepsin
and measuring the amount of forage degraded over time. A modification to this
technique is measuring the gas production of this process which has the additional
benefit of accounting for microbial growth and soluble and insoluble fractions in feed,
including tannins (Theodorou et al., 1994). The nylon bag technique is an in sacco
technique which additionally provides some measure of kinetic digestion processes
in ruminants. Both in vitro and in sacco experiments require the use of fistulated
animals and experiments demonstrate poor reproducibility, in part due to variations
in the rumen fluid used. Furthermore, results yielded by this methodological
approach offer no indices of palatability or animal production.
While there are many factors that influence the efficiency of digestion, one of the
most significant is the chemical composition of the feed. This is of particular
relevance when considering forages derived from tree foliage. Many tree species
develop high levels of secondary structures in their foliage to deter herbivores.
Chemical compounds can alter taste and odor which make them unpalatable or limit
their nutritional value (Hess et al., 2005). These compounds make multiple demands
on the animal. For example, they can affect methanogenesis in cattle (Sun et al.,

58
2012). Carulla et al. (2005) found condensed tannins in Acacia species decreased
hydrogen and resulted in a 13-24% drop in methanogenesis. Voluntary intake of a
forage may also change over time, presumably as a response to post-ingestion
feedbacks (Maasdorp et al., 1999).
Hydrolysable tannins and condensed tannins are the two major groups of plant
tannins generally found in tree foliage (Reed, 1995). Hydrolysable tannins are
potentially toxic to ruminants whereas condensed tannins may reduce the
digestibility of a forage (Kumar, 1992). In some cases, the presence of condensed
tannins can have beneficial effect on protein digestion and metabolism. Barry and
Blaney (1987) report a beneficial effect of condensed tannins when foliar
concentration is 2-3% of dry matter. The rumen escape theory suggests this
beneficial effect is due to tannins forming complexes with proteins which escape
ruminal degradation and become available in the lower gastrointestinal tract (Reed,
1995). However, Vitti et al. (2005) assert that tannin concentrations above 5% are
deleterious to metabolism.
One of the most widely planted forage tree legumes, Leucaena leucocephala, offers
an example of the complexity of plant chemistry in relation to animal production. L.
leucocephala contains the toxic secondary compound mimosine in its leaves and
pods, at dry matter concentrations reported as 12% in growing tips and 3-5% in
young leaves (Monoj and Bandyopadhyay, 2007). For this reason, it is generally
recommended that L. leucocephala comprise no greater than 20-30% of ruminant
diet. Ruminant animals can be inoculated with microbes shown to be able to break
down mimosine to non-toxic compounds. This suggests the existence of other rumen

59
organisms capable of degrading other phenolic compounds (Devendra, 1990). Due
to the multiple interactions between plants, chemistry and livestock it is difficult to
ascertain the value of a new species for forage value through chemical analysis
alone (El Hassan et al. 2000). Feeding trials may be considered the most
appropriate way of ascertaining the effect of forage on animal nutrition (Kumar,
1992).
Maasdorp et al. (1999) employed 20 cows in a feeding trial where amount of forage
consumed and milk production offered insights into the forage value of different trees
to ruminants. They found supplementary feeding of Leucaena leucocephala and
Acacia boliviana increased milk yields, but that Calliandra calothyrsus did not have a
significant effect. They reported a strong association between milk yield and the
organic matter degradability of tree forages. Forage content of ytterbium-precipitated
polyphenols and neutral detergent fiber were both negatively correlated with milk
yield. When measured via fodder intake trees exhibited different levels of palatability.
The palatability of Calliandra calothyrsus appeared to improve with time. The study
showed the effect of different tree legumes on cattle milk production. It also
highlighted the divergence between measurable chemical constituents and observed
livestock production.
An ongoing challenge of forage evaluation is to identify plant characteristics that can
be used to predict the value of a forage species for animal feed. Many farmers are
aware of potential forages for cattle, reflected in the common practice of allowing
cattle to graze on fallow lands and roadsides. Farmers may also exhibit preferences
towards some trees and remove others (Fujisaka et al., 2000). An improved

60
understanding of native species management requirements and forage quality with
regard to livestock productivity may provide evidence necessary for practitioners to
demonstrate the efficacy of such systems over conventional management.
6. Summary and Rationale
The Amazon rain forest represents a vast natural capital that has been under-
evaluated and under-utilised in the context of livestock production. Many studies that
aim to develop agroforestry or silvopastoral systems have highlighted the importance
of evaluating native species adapted to the local environmental conditions to
increase chances of success. The current paradigm of livestock production in the
Amazon is essentially a consumptive land use that drives forest loss and reduces the
provision of ecosystem services. Silvopastoral systems represent an opportunity for
sustainable intensification while simultaneously rehabilitating ecosystem functions.
This thesis aims to develop a framework that could be used to assist the decision
making of how to integrate trees into agriculture to meet the needs of a given
landowner. It is envisaged that this framework would be transferrable between
locations and could outline some key questions that arise when attempting to
identify, at a given site, appropriate agroforestry or silvopastoral arrangements. The
framework could describe methods for the appropriate selection of species, planting
design and management of resources. This framework will be introduced in Chapter
II, in which the decision-making process for the design of the present study will be
outlined with a detailed description of the study site.

61
This thesis aims to address the contextual limitations of previous research into
silvopastoral systems by developing a novel silvopastoral system that could be
adopted in the pasturelands of the Amazon. The objective of this silvopastoral
system is to increase the per hectare productivity of edible tree forage for cattle and
carbon sequestration potential per hectare when compared with conventional
pasture models in the region.
In order to achieve this, this thesis will need to address the lack of knowledge of
locally adapted forage plants in the Amazon. It is beyond the scope of this thesis to
provide an overview of all potential forage plants. Therefore, this thesis selects five
tree species that are native or locally adapted and have some positive reference to
palatability in the literature or in local knowledge. These five species will be planted
in an experimental trial in a pasture that is typical of the region. Chapter III will
discuss the growth, biomass production and effects of these species on soil
chemistry and a commonly used pasture grass of the Amazon.
The most comprehensive assessment of carbon stocks is derived from the
destructive sampling and combustion of individual biomass components. A less
labour intensive method is to choose an easily measurable part of a tree that has
some relationship to total biomass (usually the diameter at breast height, measured
at 1.3 m above the ground) and develop allometric equations that relate this
measurement to total biomass, which in turn can be used to estimate carbon
sequestration. This approach can has also been used to estimate the forage
production of trees and shrubs. As both the sequestration of carbon and forage
production are objectives of this thesis, allometric equations are presented for each

62
of the species that showed good adaptations to the pasture environment. The
allometric equations are calculated for each biomass component, and the fit of linear,
quadratic and log linear models for each of these components are compared in
Chapter IV.
Complex methodologies and resources that are beyond the reach of a farmer
support the current research into the forage potential of plants. Less work has
concentrated on how measurements can be taken before a plant is exposed to cattle
and repeated afterwards to measure the removal of biomass by cattle. Such an
approach could be a valuable on-farm tool for a preliminary assessment of the
forage potential of trees. Chapter V describes and compares four such
methodologies for estimating the removal of foliage by cattle and the impact of the
cattle on the trees. The total available biomass of tree and grass foliage per hectare
is estimated for each of the treatments.
Chapter VI concludes the present study, and summarises some of the major findings
and limitations of this study. The present work is considered in the context of the
existing literature. Finally, future research is described as both direct extensions of
the present work and the broader issues that need to be addressed to support the
transition of silvopastoral systems from theory to practice in the Amazon rainforest.
References
Abbott, I., Van Heurck, P., Burbidge, T., 1993. Impact of frequency
and intensity of defoliation on growth of Jarrah (Eucalyptus

63
marginata): an experimental study with saplings. Forest Ecology and
Management 56, 175–183. https://doi.org/10.1016/0378-1127(93)90111-Y
Achat, D.L., Deleuze, C., Landmann, G., Pousse, N., Ranger, J.,
Augusto, L., 2015. Quantifying consequences of removing harvesting
residues on forest soils and tree growth – A meta-analysis. Forest
Ecology and Management 348, 124–141.
https://doi.org/10.1016/j.foreco.2015.03.042
Alegre, J.C., Cassel, D.K., 1996. Dynamics of soil physical properties
under alternative systems to slash-and-burn. Agriculture, Ecosystems &
Environment 58, 39–48. https://doi.org/10.1016/0167-8809(95)00654-0
Alexandratos, N. and Bruinsma, J., 2012. World agriculture towards 2030/2050: the
2012 revision. ESA Working paper No. 12-03. Rome, FAO.
Altieri, M.A., Nicholls, C.I., Henao, A., Lana, M.A., 2015.
Agroecology and the design of climate change-resilient farming
systems. Agronomy for Sustainable Development 869–890.
https://doi.org/10.1007/s13593-015-0285-2
Arevalo, L.A., Alegre, J.C., Bandy, D.E., Szott, L.T., 1998. The
effect of cattle grazing on soil physical and chemical properties in a
silvopastoral system in the Peruvian Amazon. Agroforestry Systems 40,
109–124. https://doi.org/10.1023/a:1006075114659

64
Barry, T.N., Blaney, B.J., 1987. Secondary compounds of forages. In Hacker, J.B.
and Ternouth, J.H. (eds) The Nutrition Of Herbivores, Sydney, Academic Press 91-
119
Benegas, L., Ilstedt, U., Roupsard, O., Jones, J., Malmer, A., 2014.
Effects of trees on infiltrability and preferential flow in two
contrasting agroecosystems in Central America. Agriculture, Ecosystems
& Environment 183, 185–196. https://doi.org/10.1016/j.agee.2013.10.027
Bishop, J.P., 1983. Tropical forest sheep on legume forage/fuelwood
fallows. Agroforestry Systems 1, 79–84.
https://doi.org/10.1007/BF00596350
Boddey, R.M., MacEdo, R., Tarré, R.M., Ferreira, E., De Oliveira,
O.C., De, C., Cantarutti, R.B., Pereira, J.M., Alves, B.J.R.,
Urquiaga, S., 2004. Nitrogen cycling in Brachiaria pastures: The key
to understanding the process of pasture decline. Agriculture,
Ecosystems and Environment 103, 389–403.
https://doi.org/10.1016/j.agee.2003.12.010
Bogaerts, M., Cirhigiri, L., Robinson, I., Rodkin, M., Hajjar, R.,
Costa Junior, C., Newton, P., 2017. Climate change mitigation through
intensified pasture management: Estimating greenhouse gas emissions on
cattle farms in the Brazilian Amazon. Journal of Cleaner Production

65
162, 1539–1550. https://doi.org/10.1016/J.JCLEPRO.2017.06.130
Buschbacher, R., Uhl, C., Serrao, E.A.S., 1988. Abandoned Pastures in
Eastern Amazonia. II. Nutrient Stocks in the Soil and Vegetation.
Journal of Ecology 76, 682–699.
Butterfield, R.P., 1995. Promoting biodiversity: advances in
evaluating native species for reforestation. Forest Ecology and
Management 75, 111–121.
Calle, Z., Murgueitio, E., Chará, J., 2012. Integrating forestry,
sustainable cattle-ranching and landscape restoration. Unasylva 63,
31–40.
Carpenter, F.L., Mayorga, S.P., Quintero, E.G., Schroeder, M., 2001.
Land-use and erosion of a Costa Rican ultisol affect soil chemistry,
mycorrhizal fungi and early regeneration. Forest Ecology and
Management 144, 1–17.
Carpenter, L.F., Nichols, D.J., Eduer, S., 2004. Early growth of
native and exotic trees planted on degraded tropical pasture. Forest
Ecology and Management 196, 367–378.
https://doi.org/10.1016/j.foreco.2004.03.030
Carulla, J.E., Kreuzer, M., Machmüller, A., Hess, H.D., 2005.

66
Supplementation of Acacia mearnsii tannins decreases methanogenesis
and urinary nitrogen in forage-fed sheep. Australian Journal of
Agricultural Research 56, 961. https://doi.org/10.1071/AR05022
Catchpoole, D., Blair, G., 1990. Forage tree legumes. I. Productivity
and N economy of Leucaena, Gliricidia, Calliandra and Sesbania and
tree/green panic mixtures. Australian Journal of Agricultural Research
41, 521. https://doi.org/10.1071/AR9900521
Crampton, E.W., 1957. Interrelations between digestible nutrient and
energy content, coluntary dry matter intake and the overall feeding
value of forages. Journal of Animal Science 16, 546–552.
https://doi.org//1957.163546x
Devendra, C., 1990. Shrubs and tree fodders for farm animals :
proceedings of a workshop in Denpasar, Indonesia, 24-29 July 1989,
International Development Research Centre, Ottawa.
Dommergues, Y.R., 1987. The role of biological nitrogen fixation in
agroforestry, in: Steppler, H.A., Nair, P.K.R. (Eds.), Agroforestry: A
Decade of Development. ICRAF, Nairobi, pp. 245–272.
Downie, J.A., 2014. Legume nodulation. Current Biology 24, R184–R190.
https://doi.org/10.1016/j.cub.2014.01.028

67
Downing, T.E., 2019. Development or destruction: the conversion of tropical forest to
pasture in Latin America. Routledge.
Duguma, B., Kang, B.T., Okali, D.U.U., 1988. Effect of pruning
intensities of three woody leguminous species grown in alley cropping
with maize and cowpea on an alfisol. Agroforestry Systems 6, 19–35.
https://doi.org/10.1007/BF02220106
Dynes, R.A., Henry, D.A., Masters, G., 2002. Characterising Forages
for Ruminant Feeding. Proceedings of the 2002 International Symposium on “Recent
Advances in Animal Nutrition,” New Delhi, India.
El Hassan, S.M., Lahlou Kassi, A., Newbold, C.J., Wallace, R.J., 2000.
Chemical composition and degradation characteristics of foliage of
some African multipurpose trees. Animal Feed Science and Technology
86, 27–37. https://doi.org/10.1016/S0377-8401(00)00158-9
Ella, A., Blair, G.J., Stür, W.W., 1991. Effect of age of forage tree
legumes at the first cutting on subsequent production. Tropical
Grasslands - Forrajes Tropicales 25, 275–280.
Ella, A., Jacobsen, C., Stur, W.W., Blair, G., 1989. Effect of plant
density and cutting frequency on the productivity of four tree
legumes. Tropical Grasslands - Forrajes Tropicales 23, 28–34.

68
Erdmann, T.K., Nair, P.K.R., Kang, B.T., 1993. Effects of cutting
frequency and cutting height on reserve carbohydrates in Gliricidia
sepium (Jacq.) Walp. Forest Ecology and Management 57, 45–60.
https://doi.org/10.1016/0378-1127(93)90161-F
Fellmann, T., Witzke, P., Weiss, F., Van Doorslaer, B., Drabik, D., Huck, I., Salputra,
G., Jansson, T. and Leip, A., 2018. Major challenges of integrating agriculture into
climate change mitigation policy frameworks. Mitigation and adaptation strategies for
global change, 23(3), pp.451-468.
Food and Agriculture Organisation of the United Nations, F., 2016.
Agroforestry [WWW Document]. URL www.fao.org/forestry/agroforestry/en/
(accessed 2.13.16).
Frame, J., 1981. Herbage mass, in: Hodgson, J., Baker, R.D., Davis,
A., Laidlaw, A.S., Leaver, J.. (Eds.), Sward Measurement Handbook. Br.
Grassl. Soc., Berkshire, U.K., pp. 39–67.
Fujisaka, S., Escobar, G., Veneklaas, E.J., 2000. Weedy fields and
forests: interactions between land use and the composition of plant
communities in the Peruvian Amazon. Agriculture, Ecosystems &
Environment 78, 175–186. https://doi.org/10.1016/S0167-8809(99)00122-X
Gerber, P.J., Steinfeld, H., Henderson, B., Mottet, A., Opio, C., Dijkman, J., Falcucci,
A. and Tempio, G., 2013. Tackling climate change through livestock: a global

69
assessment of emissions and mitigation opportunities. Food and Agriculture
Organization of the United Nations (FAO).
Giraldo, C., Escobar, F., Chara, J.D., Calle, Z., 2011. The adoption
of silvopastoral systems promotes the recovery of ecological processes
regulated by dung beetles in the Colombian Andes. Insect Conservation
and Diversity 4, 115–122.
https://doi.org/10.1111/j.1752-4598.2010.00112.x
González, J.E., Fisher, R.F., 1994. Growth of native forest species
planted on abandoned pasture land in Costa Rica. Forest Ecology and
Management 70, 159–167. https://doi.org/10.1016/0378-1127(94)90083-3
Guevarra, A.B., Whitney, A.S., Thompson, J.R., 1978. Influence of
Intra-Row Spacing and Cutting Regimes on the Growth and Yield of
Leucaena1. Agronomy Journal 70, 1033.
https://doi.org/10.2134/agronj1978.00021962007000060034x
Guo, L., Wu, G., Li, Y., Li, C., Liu, W., Meng, J., Liu, H., Yu, X.,
Jiang, G., 2016. Effects of cattle manure compost combined with
chemical fertilizer on topsoil organic matter, bulk density and
earthworm activity in a wheat–maize rotation system in Eastern China.
Soil and Tillage Research 156, 140–147.
https://doi.org/10.1016/j.still.2015.10.010

70
Haile, S.G., Nair, V.D., Nair, P.K.R., 2010. Contribution of trees to
carbon storage in soils of silvopastoral systems in Florida, USA.
Global Change Biology 16, 427–438.
https://doi.org/10.1111/j.1365-2486.2009.01981.x
Hansen, M.C., Potapov, P. V, Moore, R., Hancher, M., Turubanova, S.A.,
Tyukavina, A., Thau, D., Stehman, S. V, Goetz, S.J., Loveland, T.R.,
Kommareddy, A., Egorov, A., Chini, L., Justice, C.O., Townshend,
J.R.G., 2013. High-resolution global maps of 21st-century forest cover
change. Science (New York, N.Y.) 342, 850–3.
https://doi.org/10.1126/science.1244693
Hardesty, L.H., Box, T.W., 1988. Defoliation Impacts on Coppicing
Browse Species in Northeast Brazil. Journal of Range Management 41,
66–70.
Hecht, S., 1982. Amazonia, Agriculture and Land Use Research:
Proceedings of the International Conference. CIAT.
Hedge, N., 1983. Leucaena forage management in India. In Leucaena
research in the Asian--Pacific region, in: Proceedings of a Workshop
Held in Singapore,. International Development Research Centre, Ottawa,
Ont., Canada. IDRC-21 Le. pp. 73–78.

71
Herrero, M., Wirsenius, S., Henderson, B., Rigolot, C., Thornton, P., Havlík, P., de
Boer, I. and Gerber, P.J., 2015. Livestock and the environment: what have we
learned in the past decade?. Annual Review of Environment and Resources, 40,
pp.177-202.
Hess, H., Stuerm, C., Tiemann, T., Lascano, C., Kreuzer, M., 2005. Effects of
mixtures of tropical legumes with contrasting tannin contents as supplements to low-
quality grass diets on ruminal fermentation in vitro, In: Tielkes, E., Hülsebusch, C.,
Häuser, I., Deininger, A., Becker, K., (eds.). The Global food & product chain:
Dynamics, innovations, conflicts, strategies: Tropentag 2005: International research
on food security, natural resource management and rural development: Book of
abstracts, University of Hohenheim, Stuttgart October 11-13, 2005. Universität
Hohenheim, Tropenzentrum, Centre for Agriculture in the Tropics and Subtropics,
Stuttgart, DE. p. 1.
Hodgson, J., 1979. Nomenclature and definitions in grazing studies.
Grass and Forage Science 34, 11–17.
https://doi.org/10.1111/j.1365-2494.1979.tb01442.x
Hohnwald, S., Rischkowsky, B., Schultze-Kraft, R., Rodrigues-Filho, J.
A., Camarão, A.P., 2005. Experiences with establishing legumes as part
of a ley pasture in a low-input farming system of the eastern Amazon,
Brazil. Pasturas Tropicales 27, 2–12.

72
Hohnwald, S., Rischkowsky, B., Camarão, A.P., Schultze-Kraft, R.,
Rodrigues Filho, J.A., King, J.M., 2006. Integrating cattle into the
slash-and-burn cycle on smallholdings in the Eastern Amazon, using
grass-capoeira or grass-legume pastures. Agriculture, Ecosystems &
Environment 117, 266–276. https://doi.org/10.1016/J.AGEE.2006.04.014
Hohnwald, S., 2016. Relative palatability and growth performance of
capoeira species as supplementary forages in the NE-Amazonia.
Agriculture, Ecosystems & Environment 218, 107–115.
https://doi.org/10.1016/j.agee.2015.11.015
Ibrahim, M., Villanueva, C., Casasola, F., Rojas, J., 2006. Sistemas
silvopastoriles como una herramienta para el mejoramiento de la
productividad y restauración de la integridad ecológica de paisajes
ganaderos. Pastos y Forrajes 29, 383–419.
Ilstedt, U., Malmer, A., Verbeeten, E., Murdiyarso, D., 2007. The
effect of afforestation on water infiltration in the tropics: A
systematic review and meta-analysis. Forest Ecology and Management
251, 45–51. https://doi.org/10.1016/j.foreco.2007.06.014
Jackson, J., Ash, A.J., 1998. Tree-grass relationships in open
eucalypt woodlands of Northeastern Australia: influence of trees on
pasture productivity, forage quality and species distribution.
Agroforestry Systems 40, 159–176.

73
https://doi.org/10.1023/A:1006067110870
Kang, B.T., Reynolds, L., Atta-Krah, A.N., 1990. Alley Farming. Advances in
Agronomy Volume 43. Elsevier.
https://doi.org/10.1016/S0065-2113(08)60481-2
Kass, M., Benavides, J., Romero, F., Pezo, D., Speedy, A., Pugliese,
P.L., 1992. Lessons from main feeding experiments conducted at CATIE
using fodder trees as part of the N-ration., in: Legume Trees and
Other Fodder Trees as Protein Sources for Livestock. FAO Animal
Production and Health Paper. Food and Agriculture Organization of the
United Nations, pp. 161–175.
Kaur, B., Gupta, S.R., Singh, G., 2002. Carbon storage and nitrogen
cycling in silvopastoral systems on a sodic soil in northwestern
India. Agroforestry Systems 54, 21–29.
Khan, M.A., Mahr-Un-Nisa, S.M. and Sarwar, M., 2003. Techniques measuring
digestibility for the nutritional evaluation of feeds. International Journal of Agriculture
and Biology, 5(1), pp.91-94.
Kumar, M.B., George, S.J., Jamaludheen, V., Suresh, T.K., 1998.
Comparison of biomass production, tree allometry and nutrient use
efficiency of multipurpose trees grown in woodlot and silvopastoral
experiments in Kerala, India. Forest Ecology and Management 112,

74
145–163. https://doi.org/10.1016/S0378-1127(98)00325-9
Kumar, R., 1992. Anti-nutritional factors, the potential risks of
toxicity and methods to alleviate them, in: Expert Consultation on
Legume Trees and Other Fodder Trees as Protein Sources for Livestock,
Kuala Lumpur (Malaysia), 14-18 Oct 1991. FAO Animal Production and
Health Paper (FAO), 1992.
Lal, R., 2004. Agricultural activities and the global carbon cycle.
Nutrient Cycling in Agroecosystems 70, 103–116.
https://doi.org/10.1023/B:FRES.0000048480.24274.0f
Lee, M.A., Davis, A.P., Mizeck G.G.C., Manning, P., 2017. Forage
quality declines with rising temperatures, with implications for
livestock production and methane emissions. Biogeosciences 14,
1403–1417. https://doi.org/doi:10.5194/bg-14-1403-2017
Loker, W.M., 1994. Where’s the beef?: Incorporating cattle into
sustainable agroforestry systems in the Amazon Basin. Agroforestry
Systems 25, 227–241. https://doi.org/10.1007/BF00707462
Maasdorp, B.V., Muchenje, V., Titterton, M., 1999. Palatability and
effect on dairy cow milk yield of dried fodder from the forage trees
Acacia boliviana, Calliandra calothyrsus and Leucaena leucocephala.
Animal Feed Science and Technology 77, 49–59.

75
https://doi.org/10.1016/S0377-8401(98)00232-6
Mannetje, L. ’t., 2000. Measuring biomass of grassland vegetation, in:
Mannetje, L.’t., Jones, R.M. (Eds.), Field and Laboratory Methods for
Grassland and Animal Production Research. C.A.B.I. Publishing. Univ.
Press, Cambridge, UK, pp. 151–177.
Mannetje, L. ’t., 1978. Measuring quantity of grassland vegetation,
in: L.’t., M. (Ed.), Measurement of Grassland Vegetation and Animal
Production. Commonwealth Agricultural Bureaux, Bucks, England, pp.
63–95.
Martínez, L.J., Zinck, J.A., 2004. Temporal variation of soil
compaction and deterioration of soil quality in pasture areas of
Colombian Amazonia. Soil and Tillage Research 75, 3–17.
https://doi.org/10.1016/j.still.2002.12.001
Mcadam, J.H., Sibbald, A.R., Teklehaimanot, Z., Eason, W.R., 2007.
Developing silvopastoral systems and their effects on diversity of
fauna. Agroforestry Systems 70, 81–89.
https://doi.org/10.1007/s10457-007-9047-8
McCalla, G.R., Blackburn, W.H., Merrill, L.B., 1984. Effects of
Livestock Grazing on Infiltration Rates, Edwards Plateau of Texas.
Journal of Range Management 37, 265–269.

76
McCarty, M.K., Mazurak, A.P., 1976. Soil Compaction in Eastern
Nebraska after 25 Years of Cattle Grazing Management and Weed Control.
Journal of Range Management 29, 384 – 386.
Menezes, R.S.C., Salcedo, I.H., Elliott, E.T., 2002. Microclimate and
nutrient dynamics in a silvopastoral system of semiarid northeastern
Brazil. Agroforestry Systems 56, 27–38.
https://doi.org/10.1023/A:1021172530939
Minson, D.J., Harris, C.E., Raymond, W.F., Milford, R., 1964. The
digestibility and voluntary intake of s22 and h.1 ryegrass, s170 tall
fescue, s48 timothy, s215 meadow fescue and germinal cocksfoot. Grass
and Forage Science 19, 298–305.
https://doi.org/10.1111/j.1365-2494.1964.tb01177.x
Monoj, K.G., Bandyopadhyay, S., 2007. Mimosine Toxicity-A Problem of
Leucaena Feeding in Ruminants. Asian Journal of Animal and Veterinary
Advances 2, 63–73.
Montagnini, F., Sancho, F., 1990. Impacts of Native Trees on Tropical
Soils: A Study in the Atlantic Lowlands of Costa Rica. Ambio 19,
386–390.

77
Montagnini, F., Ashton, M., 1999. The Silvicultural Basis For
Agroforestry Systems. CRC Press.
Moraes, J.F.L. de, Volkoff, B., Cerri, C.C., Bernoux, M., 1996. Soil
properties under Amazon forest and changes due to pasture installation
in Rondônia, Brazil. Geoderma 70, 63–81.
Murgueitio, E., Calle, Z., Uribe, F., Calle, A., Solorio, B., 2011.
Native trees and shrubs for the productive rehabilitation of tropical
cattle ranching lands. Forest Ecology and Management 261, 1654–1663.
https://doi.org/10.1016/j.foreco.2010.09.027
Nguyen, T.M., Binh, D. Van, Ørskov, E.R., 2005. Effect of foliages
containing condensed tannins and on gastrointestinal parasites. Animal
Feed Science and Technology 121, 77–87.
https://doi.org/10.1016/j.anifeedsci.2005.02.013
Nichols, J.D., Rosemeyer, M.E., Carpenter, F.L., Kettler, J., 2001.
Intercropping legume trees with native timber trees rapidly restores
cover to eroded tropical pasture without fertilization. Forest Ecology
and Management 152, 195–209.
https://doi.org/10.1016/S0378-1127(00)00603-4
Osman, A.M., 1981. Effects of cutting interval on the relative dry
matter production of four cultivars of leucaena. Leucaena Res. Rep. 2,

78
33–38.
Parsons, A.J., Leafe, E.L., Collett B., Penning, P.D., Lewis, J.,
1983. The Physiology of Grass Production Under Grazing. II.
Photosynthesis, Crop Growth and Animal Intake of Continuously-Grazed
Swards. Journal of Applied Ecology 20, 127–139.
Pathak, P.S., Rai, P., Roy, R.D., 1980. Forage production from koo-bab
ool (Leucaena leucocephala (Lam.) de Wit). 1. Effect of plant density,
cutting intensity and interval. Forage Research 6, 83–90.
Pedrotti, A., Pauletto, E.A., Crestana, S., Holanda, F.S.R., Cruvinel,
P.E., Vaz, C.M.P., 2005. Evaluation of bulk density of Albaqualf soil
under different tillage systems using the volumetric ring and
computerized tomography methods. Soil and Tillage Research 80,
115–123. https://doi.org/10.1016/j.still.2004.03.003
Perez, J.B., Boraemisza, E., Sollins, P., 1993. Identificacion de
especies forestales acumuladoras de aluminio en una plantación
forestal experimental ubicada en Sarapiquı, Costa Rica. Agronomıa
Costarricense 17, 99–103.
PRODES 2019 Programa de Cálculo do Desflorestamento da Amazônia, Amazon
Deforestation Calculation Program [WWW]
URL: http://www.obt.inpe.br/prodes/index.php [accessed September 25 2019]

79
Quraishi, M.Z., Mouazen, A.M., 2013. A prototype sensor for the
assessment of soil bulk density. Soil and Tillage Research 134,
97–110. https://doi.org/10.1016/j.still.2013.07.011
Reátegui, K., Ruiz, R., Cantera, G., Lascano, C., 1990. Persistencia
de pasturas asociadas con diferentes manejos del pastoreo en un
Ultisol arcilloso de Puerto Bermúdez, Perú. Tropical Grasslands -
Forrajes Tropicales 12, 16–24.
Reed, J.D., 1995. Nutritional toxicology of tannins and related
polyphenols in forage legumes. Journal Of Animal Science 73, 1516–28.
Reynolds, L., Cobbina, J., 1992. Unit 4: The Integration of Livestock
Production in Alley Farming, in: Tripathi, B.R., Psychas, P.J. (Eds.),
The AFNETA Alley Farming Training Manual - Volume 1: Core Course in
Alley Farming. International Institute of Tropical Agriculture,
Ibadan.
Rittner, U., Reed, J.D., 1992. Phenolics and in-vitro degradability of
protein and fibre in West African Browse. Journal of the Science of
Food and Agriculture 58, 21–28. https://doi.org/10.1002/jsfa.2740580105
Rojas-Downing, M.M., Nejadhashemi, A.P., Harrigan, T., Woznicki, S.A.,
2017. Climate change and livestock: Impacts, adaptation, and

80
mitigation. Climate Risk Management 16, 145–163.
https://doi.org/10.1016/J.CRM.2017.02.001
Rosegrant, M. W., Fernandez, M., Sinha, A., Alder, J., Ahammad, H., de Fraiture,
Charlotte, Eickhour, B., Fonseca, J., Huang, J., Koyama, O., Omezzine, A. M.,
Pingali, P., Ramirez, R., Ringler, C., Robinson, S., Thornton, P., van Vuuren, D.,
Yana-Shapiro, H. 2009. Looking into the future for agriculture and AKST. In
McIntyre, B. D., Herren, H. R., Wakhungu, J., Watson, R. T. (Eds.). International
Assessment of Agricultural Knowledge, Science and Technology for Development
(IAASTD): Agriculture at a Crossroads, global report. Washington, DC, USA: Island
Press. pp.307-376.
Russo, R., Botero, R.B., 1994. Nitrogen fixing trees for animal
production on acid soils. Powell, Mark H.(ed.). Workshop on Nitrogen
Fixing Trees for Acid Soils (1994, Turrialba, Costa Rica). Nitrogen
fixing trees for acid soils: A field manual. 1994.
Sanchez, F.J.S., 2002. Integrating fodder trees into animal production
systems in the tropics. Tropical and Subtropical Agroecosystems 1,
1–11.
Sanchez, P., 1987. Soil productivity and sustainability in
agroforestry systems, in: Agroforestry: A Decade of Development.
ICRAF, Nairobi, pp. 205–223.

81
Savory, R., Breen, J., 1979. The production of Leucaena leucocephala
in Malawi. (iii) Plant population studies. United Nations Development
Programme/Food and Agriculture Organization Project MLW /757020. (No.
17).
Scholes, S.J., Archer, S.R., 1997. Tree-grass Interactions in
Savannas. Annual Review of Ecology and Systematics 28, 517–544.
Serrao, E.D., Uhl, C., Nepstad, D., 1993. Deforestation for pasture in
the humid tropics: Is it economically and environmentally sound in the
longterm?, in: Proceedings of the XVII International Grassland
Congress, Rockhampton, Australia and Palmerston North, New Zealand. p.
2215−2221.
Shaw, N.H., Mannetje, L., Jones, R.M., 1976. Pasture Measurements, in:
Shaw, N.H., Bryan, W.. (Eds.), Tropical Pasture Research. Principles
and Methods. Commonwealth Agricultural Bureaux, Bucks, England, pp.
235–262.
Sierra, J., Daudin, D., Domenach, A.M., Nygren, P., Desfontaines, L.,
2007. Nitrogen transfer from a legume tree to the associated grass
estimated by the isotopic signature of tree root exudates: A
comparison of the 15N leaf feeding and natural 15N abundance methods.
European Journal of Agronomy 27, 178–186.
https://doi.org/10.1016/j.eja.2007.03.003

82
Sousa, L.F., Mauricio, R.M., Paciullo, D.S.C., Silveria, S.R.,
Riberio, R.S., Calsavara, L.H., Moreira, G.R., 2015. Forage intake,
feeding behavior and bio-climatological indices of pasture grass,
under the influence of trees, in a silvopastoral system. Tropical
Grasslands - Forrajes Tropicales 3, 129.
https://doi.org/10.17138/TGFT(3)129-141
Stainback, A.G., Alavalapati, J.R.R., 2004. Restoring longleaf pine
through silvopasture practices: an economic analysis. Forest Policy
and Economics 6, 371–378. https://doi.org/10.1016/j.forpol.2004.03.012
Stephenson, G.R., Veigel, A., 1987. Recovery of Compacted Soil on
Pastures Used for Winter Cattle Feeding. Journal of Range Management
1, 46–48.
Stevenson, A.C., Harrison, R.J., 2014. Ancient Forests in Spain: A
Model for Land-use and Dry Forest Management in South-west Spain from
4000 BC to 1900 AD. Proceedings of the Prehistoric Society 58,
227–247. https://doi.org/10.1017/S0079497X00004175
Stolt, M.H., Lindbo, D.L., 2010. Interpretation of Micromorphological
Features of Soils and Regoliths, Interpretation of Micromorphological
Features of Soils and Regoliths. Elsevier.
https://doi.org/10.1016/B978-0-444-53156-8.00017-9

83
Stür, W.W., Shelton, H.M., Gutteridge, R.C., 1994. Defoliation
management of forage tree legumes in Forage tree legumes in tropical
agriculture. Cab International.
Sun, X.Z., Waghorn, G.C., Hoskin, S.O., Harrison, S.J., Muetzel, S.,
Pacheco, D., 2012. Methane emissions from sheep fed fresh brassicas
(Brassica spp.) compared to perennial ryegrass (Lolium perenne).
Animal Feed Science and Technology 176, 107–116.
https://doi.org/10.1016/j.anifeedsci.2012.07.013
Szott, L.T., Fernandes, E.C.M., Sanchez, P.A., 1991. Soil-plant
interactions in agroforestry systems. Forest Ecology and Management
45, 127–152. https://doi.org/10.1016/0378-1127(91)90212-E
Teague, W.R., 1989. Effect of intensity and frequency of defoliation
on aerial growth and carbohydrate reserve levels in Acacia karroo
plants. African Journal of Range and Forage Science.
Theodorou, M.K., Williams, B.A., Dhanoa, M.S., McAllan, A.B., France,
J., 1994. A simple gas production method using a pressure transducer
to determine the fermentation kinetics of ruminant feeds. Animal Feed
Science and Technology 48, 185–197.
https://doi.org/10.1016/0377-8401(94)90171-6

84
Thomas, H., Laidlaw, A.S., 1981. Planning, design, and establishment
of experiments, in: Hodgson, J., Baker, R.D., Davis, A., Laidlaw,
A.S., Leaver, J.D. (Eds.), Sward Measurement Handbook. Br. Grassl.
Soc, Berkshire, U.K., pp. 15–38.
Thornton, P.K. and Herrero, M., 2010. The inter-linkages between rapid growth in
livestock production, climate change, and the impacts on water resources, land use,
and deforestation. The World Bank.
Tilki, F., Fisher, R.F., 1998. Tropical leguminous species for acid
soils: Studies on plant form and growth in Costa Rica. Forest Ecology
and Management 108, 175–192.
https://doi.org/10.1016/S0378-1127(98)00225-4
Tilley, J.M.A., Terry, R.A., 1963. A two-stage technique for the in
vitro digestion of forage crops. Grass and Forage Science 18, 104–111.
https://doi.org/10.1111/j.1365-2494.1963.tb00335.x
Uhl, C., Buschbacher, R., Serrao, E.A.., 1988. Abandoned Pastures in
Eastern Amazonia. I. Patterns of Plant Succession. Journal of Ecology
76, 663–681.
Ulyatt, M.J., 1973. The feeding value of herbage, in: Butler, G.W.,
Bailey, R.W. (Eds.), Chemistry and Biochemistry of Herbage. Academic
Press, London, pp. 131–178.

85
Van Haveren, B.P., 1983. Soil Bulk Density as Influenced by Grazing
Intensity and Soil Type on a Shortgrass Prairie Site. Journal of Range
Management 5, 586–588. https://doi.org/http://doi.org/10.2307/3898346
Vermeire, L.T., Ganguli, A.C., Gillen, R.L., 2002. A Robust Model for
Estimating Standing Crop across Vegetation Types. Journal of Range
Management 55, 494–497.
Viana, R.M., Ferraz, J.B.S., Neves, A.F., Vieira, G., Pereira, B.F.F.,
2014. Soil quality indicators for different restoration stages on
Amazon rainforest. Soil and Tillage Research 140, 1–7.
https://doi.org/10.1016/j.still.2014.01.005
Vitti, D.M.S.S., Abdalla, A.L., Bueno, I.C.S., Filho, J.C.S., Costa,
C., Bueno, M.S., Nozella, E.F., Longo, C., Vieira, E.Q., Filho,
S.L.S.C., Godoy, P.B., Mueller-Harvey, I., 2005. Do all tannins have
similar nutritional effects? A comparison of three Brazilian fodder
legumes. Animal Feed Science and Technology 119, 345–361.
https://doi.org/10.1016/j.anifeedsci.2004.06.004
Wassenaar, T., Gerber, P., Verburg, P.H., Rosales, M., Ibrahim, M. and Steinfeld,
H., 2007. Projecting land use changes in the Neotropics: The geography of pasture
expansion into forest. Global Environmental Change, 17(1), pp.86-104.

86
Wang, D., Bormann, F.H., Lugo, A.E., Bowden, R.D., 1991. Comparison of
nutrient-use efficiency and biomass production in five tropical tree
taxa. Forest Ecology and Management 46, 1–21.
https://doi.org/10.1016/0378-1127(91)90241-M
Weston, R.H., 1996. Some aspects of constraint to forage consumption
by ruminants. Australian Journal of Agricultural Research. 47, 175–198.
Weston, R.H., 1985. The regulation of feed intake in herbage-fed
ruminants. Proceedings of the Nutrition Society of Australia. 10, 55–62.
Weston, R.H., 1982. Animal factors affecting voluntary feed intake,
in: Hacker, J.B. (Ed.), Nutritional Limits to Animal Production from
Pasture. Commonwealth Agricultural Bureaux, Slough, pp. 183–198.
White, R.P., Murray, S., Rohweder, M., Prince, S.D. and Thompson, K.M.,
2000. Grassland ecosystems (p. 81). Washington, DC: World Resources Institute.
Wilm, H.G., Costello, D.F., Klipple, G.E., 1944. Estimating forage
yield by the double-sampling method. Journal of the American Society
of Agronomy.

87
CHAPTER II
FRAMEWORK AND METHODOLOGY
1. Objective
The objective of this chapter is to outline a theoretical framework that can be used to
evaluate the potential of tree species for a purpose driven agroforestry system and
demonstrate how this framework can be employed in a practical setting. The chapter
will describe the establishment of a silvopastoral system based on the principles
presented in the framework and outline a practical example of how the decision
making process has been used in the design of a field experiment. The framework
consists of four interacting components: the purpose, the species, the site and the
planting design (Figure 1).

88
Figure 1. Theoretical framework for the evaluation of candidate species for agroforestry systems
The purpose can be described as the reason that necessitates the implementation of
an agroforestry system. The subsequent components of species, site and planting
design are independent and complex issues that must be overcome in order to
achieve the purpose. Sanchez (1995) assert that the biophysical determinant of
successful agroforestry systems is the appropriate management between trees and
crops for light, water and nutrient competition. Even if the purpose is the plantation of
a single tree species, consideration can be given to additional species that can
increase the benefits provided per hectare, such as ground cover leguminous plants
that may increase soil fertility or mychorrizae that may benefit the species of interest.
The next section aims to put these components into the context of the purpose of
designing a silvopastoral system for the increase in productivity per hectare of edible
biomass production for cattle in the southwestern Amazon.

89
2. The Purpose
The purpose of this study was to identify native potential forage trees in the Amazon
that could increase the production per hectare of edible biomass for cattle. Systems
that manage tree or shrub and animal components are known as silvopastoral
systems. There is no minimum tree or shrub component of a silvopastoral systems,
which may be almost all pasture with a few trees for shade or timber and little
management, it may be a eucalyptus plantation where the manager sees cattle as a
useful byproduct that controls grass, or an intensive silvopastoral system that aims to
provide balanced nutrients to cattle using with strips of forage legumes and highly
productive African grasses, but without a major canopy element. In general, while
silvopastoral systems include both animal and plant elements, they may be managed
predominantly for the production of timber or livestock. The purpose of the current
study is an empirical evaluation of the potential of tree species to increase per
hectare productivity of edible biomass when compared to the status quo (no trees),
under the assumption that an increase in edible biomass can result in an increase in
stocking density and therefore an increase in livestock production per hectare.
3. The Site
In Figure 1, the ‘site’ refers to the biotic and abiotic resources available, and can also
be described as the balance of inputs and available to the system and the
constraints placed upon the system. The conditions of the site will be either
advantageous or disadvantageous to the set of species of under consideration. In
order to design an optimal system it is important to understand how the site, the
species and the management of the species interact. This understanding can be

90
gained via observation of landscapes, consultation with local ecological knowledge,
a review of the literature and the use of pilot studies. For the best chances of
success, agroforestry systems could include diverse species and arrangements and
be subject to adaptive management.
3.1 Site Description
The present study is situated in Madre de Dios, Peru (Figure 2). Madre de Dios is a
part of the Tropical Andes Biodiversity Hotspot and in the Southwestern Amazonian
Moist Forests Global 200 Ecoregion (Olson and Dinerstein, 2002; Myers, 2001). The
site is located 8 km from the region’s capital of Puerto Maldonado in an area known
as the Cachuela.

91
Figure 2. Location of the study site in Madre de Dios, Peru
The site was a cattle pasture from 1970-2000 and in 2000 the site was abandoned.
The Cachuela is an agricultural region prone to seasonal flooding, with a mosaic of
secondary forests, farms and cattle pastures. A total of 4 hectares of land was
available of which 2.7 was to establish an experimental silvopastoral system and 1.3
was to develop the tree seedling nurseries, cattle corrals and associated
infrastructure. In its entirety the plot was 100 x 400 m (Figure 3, Figure 4).

92
Figure 3. Experimental site in 2002 with the future location of the plots projected
Figure 4. Experimental site in 2015 with the future location of the plots projected.
3.2 Floristic Description of Site
The site was historically a cattle pasture that had been given to 15 years of natural
regeneration and species that had been propagated by the cattle dominated, notably
Psidium guajava and Genipa americana. Additional species at the site included
Ormosia coccinea, Cecropia spp., Attalea butyracea, Ficus spp., Moena alcanforada,
Apeiba membranacea, Ceiba spp. and Ochroma pyramidale.
The most eastern point of the site began approximately 150 m west of the river
Madre de Dios and extended 400 m away from the river. The site was on a mid to
high floodplain that may have formed during the Younger Dryas which continued
until approximately 3.7 ka BP (Nikitina et al., 2011). The site did not flood from the

93
river but was prone to inundation from rainfall events. Floods from the adjacent river
were experienced less than 100 m from the site. On the northern edge of the site
there was a seasonal lake, fed by rainfall and seasonally connected to a network of
swamps and clear water streams.
3.3 Climatic Description of Site
From 1970-2000 the annual mean temperate at the site was 25.3 °C with a mean
diurnal range of 10.3 °C (Figure 5). The maximum temperature recorded in the
hottest month during this time period was 31.6 °C and the minimum temperature
recorded in the coldest month was 18.2 °C. On average, annual precipitation was
2122 mm, the highest level of precipitation in the wettest month was 319 mm and
that of the driest month was 55 mm (Figure 6). Solar irradiation on average was
14400 kJ m-2 day-1, with an annual maximum of 16081 kJ m-2 day-1 and a
minimum of 12946 kJ m-2 day-1 (Figure 7). All climate data was derived for the
years 1970-2000 for a 1 km2 resolution from the WorldClim dataset (Hijmans et al.,
2005).

94
Figure 5. Average (solid line), maximum (dotted line), and minimum (dashed line) monthly temperature (°C) from the period 1970-2000 for the study site derived from the WorldClim dataset (Hijmans et al., 2005).
Figure 6. Average monthly precipitation (mm) from the period 1970-2000 for the study site derived from the WorldClim dataset (Hijmans et al., 2005).
15
17
19
21
23
25
27
29
31
33D
egre
es
Cel
siu
s
0
50
100
150
200
250
300
350
(mm
)

95
Figure 7. Average monthly level of solar irradiation (kJ m-2 day-1) from the period 1970-2000 for the study site derived from the WorldClim dataset (Hijmans et al., 2005).
3.4 Edaphic Description of Site
The sites physiography is a recent alluvium (Osher and Buol, 1998). The depth of
the water table varied from 0.5 - 1 m in the rainy season, to 7 - 10 m in the dry
season. The soil at the site could be described as a Gleysol or Fluvisol (FAO, 2015)
or Alfizol or Entisol (Eswaran et al., 2002). There were no clear distinctions of the soil
horizon, which was grey or red brown and dominated by clay particulates. This
resulted in low soil porosity and a slow infiltration rate. Frequent and high intensity
precipitation events combined with a history of cultivation could have contributed to
the highly weathered soils (Moreira et al., 2006). The pH of the site was acidic with a
site average of pH 5.5.
12000
12500
13000
13500
14000
14500
15000
15500
16000
16500(k
J m
-2d
ay-1
)

96
3.5 Social and Economic History
Archeological evidence points to human habitation in the Madre de Dios region for
over 1,000 years (Mann, 2008). Petroglyphs found along the Palotoa, Shinkebenia
and Urubamba and in the Pongo de Mainique suggest these areas were important
for early trade. The Inca’s were well aware of the populations in the forested regions
of Amarumayo, as it was then known. Missionaries, botanists and those in search of
Paititi were the first ‘explorers’ of the region of Madre de Dios, which was inhabited
at the time. The first successful colonization occurred in 1902 as a result of the
rubber boom, when the first government of Madre de Dios was formed in the newly
founded town of Puerto Maldonado. The memory of the rubber trade continues in
modern indigenous memory, and the tribes who overthrew the slave masters and
fled to the forests continue to live in voluntary isolation in the headwaters of the Las
Piedras, Manu and Alto Purus rivers in Madre de Dios.
The first road connecting Cusco to Puerto Maldonado, the capital of Madre de Dios,
was a horse path in use since the 1930’s, and the first road that was constructed to
the region was in the 1960’s. In the period of 1980-1990 the Peruvian government,
led by Alan Garcia, introduced policy incentives to develop agricultural expansion in
the Amazon basin (Chávez Michaelsen et al., 2013). In the same period, the forests
of Madre de Dios became major sources of tropical timber. The practice of selective
logging for high value timber species continues to be a major driver of economic
activity. Other major non-timber economies in the region are the production of
approximately 10 percent of the world’s Brazil nuts and as a popular destination for
tourists who visit the rainforest (Duchelle et al., 2012; Kirkby et al., 2011). The road
that connects Puerto Maldonado to Cusco was continued on to the Brazilian and

97
Bolivian frontiers and was paved in the early 2000’s and converted into the major
Initiative for the Integration of the Regional Infrastructure of South America (IIRSA)
project, also known as the Interoceanic highway, that was completed in 2011. Since
the early 2000’s, Puerto Maldonado has become a popular destination for the
profitable and often illegal activity of the extraction of alluvial gold deposits (Scullion
et al., 2014). Property rights in the region continue to undergo reform (Perz et al.,
2016). This process is slow due to changing responsibility between different
government departments. An arson attack on the Ministry of Agriculture in 2002 and
on the Regional Government in 2008 destroyed many records of land ownership and
was a setback to the formalisation of property rights in the region.
3.6 Future of the Amazon
The climate in the Amazon is changing. 2016 was the hottest year since 1950 (0.9
°C + 0.3 °C) and this has been related to the burning and clearing of the forest and
pastures (Marengo et al., 2018; van der Werf et al., 2010). Soares et al. (2006) used
deforestation rates prior to 2004 to produce a business‐as‐usual scenario that
estimated that 47% of the Brazilian Amazon would be deforested by 2050 (Soares-
Filho et al., 2006). Deforestation is known to reduce evapotranspiration and
precipitation. Consequently the frequency and duration of drought is increasing
(Panisset et al., 2017). Spracklen and Garcia-Carreras (2015) estimated that using
the pre-2004 rates of deforestation, as a business-as-usual scenario would lead to
an 8.1 ± 1.4% reduction in annual mean rainfall in the Amazon basin by 2050. Lima
et al. (2014) found that regional deforestation increases the length of the dry season
and the seasonal amplitude of water flow in the southwestern Amazon. Lawrence

98
and Vandecar (2015) found that tropical deforestation results in a warmer and drier
climate at the local scale and predict that negative impacts could extend beyond the
tropics. Scientists have considered the possibility of a ‘tipping point’, which is
described as a critical threshold at which an irreversible drought occurs as a result
either of global warming or a 25% or 40% loss of the total forest area of the Amazon
(Lenton et al., 2008; Nobre et al., 2016; Lovejoy and Nobre, 2018). Authors who
have modelled climate scenarios for the Amazon using the CPTEC model suggest it
becomes drier as a result of reduced precipitation and that in place of tropical
rainforest there will be an increase in savannah ( Sampaio et al., 2007; Sampaio et
al., 2019). While the future of the Amazon’s response to global warming and
deforestation is unknown, it is known that this is a high-risk scenario. Authors have
attempted to calculate the value of the ecosystem services the Amazon contains and
the financial ramifications of losing the provision of these services. Rammig et al.
(2018) predict that the loss of the Amazon in Brasil will result in short term gains of
USD 576 to 2,880 billion and a long-term loss of USD 1,367 to 6,928 billion. It is not
possible to predict how future governments of Amazon nations will respond to what
is now viewed as a global environmental crisis. However, the demands of a
globalised economy increasingly transcend governments and can place unforeseen
demands on natural resources, such as a US-China trade war resulting in increased
demand for Brazilian soy that may result in a surge in deforestation (Fuchs et al.,
2019).
3.7 Site Preparation
The decision was taken to clear the site by hand to simulate a traditional ‘slash and
burn’. The alternative would have been the use of heavy machinery such as tractors.

99
However, this would have disturbed the soils. Site clearing began in May 2016. All
undergrowth and small trees were removed by machete, and then the trees left
standing were felled with a chainsaw. All fallen organic material was dried in the sun
for three months. Prior to burning a 10 m fire break was cut around the perimeter of
the site and cylinders of water were positioned every 100 m or so to control any
unintended spread of fire. The site was burned in October 2016 (Figure 8). The fire
did not spread beyond the site. However, two weeks later an uncontrolled fire from a
farm 400 m away reached the edge of the site. Remaining fallen trunks were cut with
a chainsaw, piled and burned (Figure 9, Figure 10). A glyphosate herbicide was
applied across the site in December 2016 (Figure 11). Parcels were cleared of all
vegetation by raking and hoeing. Trunk stumps were cut to ground level with a
chainsaw prior to planting.

100
Figure 8. Experimental site in October 2016 with the future plots projected.
Figure 9. A monumental task. All site clearance was carried out by hand to minimise disturbance to the soils.

101
Figure 10. An international team of volunteers involved helped with the pile and burn.
Figure 11. When the site had been cleared of branches, the weeds at the site had grown to be unmanageable. A glyphosphate herbicide was applied across the site and the vegetation had to be cleared again. Clearance of this vegetation was done using a hoe and rake. All residues were taken off site in wheelbarrows.

102
4. Species
4.1 Evaluation of Desirable Plant Traits and Candidate Trees in Silvopastoral
Systems
Kattge et al. (2011) define Plant traits as “the morphological, anatomical,
physiological, biochemical and phenological characteristics of plants and their
organs”. Westoby and Wright (2006) explains their importance as “control ecosystem
processes and define habitat and resources for other taxa; thus, they are a high
priority for understanding the ecosystem at a site”. The identification and selection of
plants for a specific purpose is a critical evaluation of plant traits that determine their
suitability for the intended purpose.
In this study we use the example of developing a silvopastoral system that, in the
context of our study site in the Amazon, can increase the productivity of cattle
production systems over monoculture approaches. In this study, desirable plant traits
are those that can be identified as helping to achieve this goal (Figure 12). In the
Bragantina region of the Brazilian Amazon, Hohnwald et al. (2016) considered the
following criteria of decreasing importance to select species of potential forage value
for an experiment in north eastern Pará, Brazil. “good adaptations to environmental
conditions (poor, acid, and compacted soils), tropical climate (with up to 3 months
long dry seasons), periodical slashings and fires, high leafy biomass production,
highly preferred by cattle (Hohnwald 2002; Hohnwald et al., 2015), fast recuperation
abilities after defoliation, free accessibility on smallholdings, and high nutrient
contents in leaves.” Promising species could be identified by crude protein and fiber
values similar to that of forage grasses, and species to be avoided included those

103
with high amounts of condensed tannins. Pennington (1998) selected species from
the genus Inga with growth chatacteristics suitable for forestry or agroforestry use,
such as land reclamation, alley-cropping systems and fuelwood. The characteristics
they selected for were fast growth rates, high production of leafy and woody
biomass, ability to establish and flourish on marginal sites, and ability to compete
with fast growing secondary vegetation. Within the genus they distinguish between
species that are non-pioneer light demanding gap species and shade tolerant plants
from the forest understory, the latter being less desirable due to their intolerance to
the sun and slow establishment. Román-Dañobeytia et al., (2015) used the following
criteria to select species for the rehabilitation of sites degraded by alluvial mining:
seed availability, ease of propagation, rapid growth in open areas, and wide
geographical distribution across tropical America (Román et al., 2012). There has
been a call for an exploration of the agroforestry potential of native species as they
can provide unique ecosystem services and increase diversity (Butterfield, 1995;
Fisher, 1995; Murgueitio et al., 2011).

104
Figure 12. Mind map of desirable plant traits in the context of developing silvopastoral systems at a site in the Amazon that could be advantageous in achieving the goal of increasing the productivity per hectare through the use of silvopastoral systems compared with traditional methods of cattle production in the region.
The list of desirable traits in Figure 12 is presented as an example of how species
selection was informed in this study. Information was gathered from three categories
of resources: Published knowledge, unpublished knowledge and knowledge gained
by observation (Figure 13). Benavides (1995) outlined three steps for the
identification and characterization of potential forage species (1) surveys of cattle
workers to find out if animals prefer certain woody species (2) direct observation of
animals browsing preference (3) secondary information from existing literature
(Benavides, 1995).

105
Figure 13. Resources for identifying potential species for inclusion in agroforestry systems.
Local ecological and environmental knowledge (LEK) is a valuable resource for the
selection of species. LEK is a resource recognised within the academic realm and
has been used to manage wild plants in Albania (Tomasini and Theilade, 2019),
develop knowledge of plant phenology in the Brazilian Cerrado (Campos et al.,
2018), and surveys with farmers have highlighted important local forage species in
West Africa (Naah and Guuroh, 2017). Linstädter et al. (2013) held a group interview
with three herdsmen of sheep and goats to estimate palatability and nutritive value
for 62 species on a pasture. Informants classified species into “refused, little or
occasionally eaten, regularly eaten, highly favoured” and were asked to rank their
perception of the species’ nutritive value as “bad, medium, good”. Bahru et al. (2014)
carried out an ethnobotanic study of forage plants in Ethiopia with 96 informants.
One hundred twenty-six forage species of 90 genera and 43 families were collected
in the study area. Naah and Guuroh (2017) used a technique known as free listing to
elicit local environmental knowledge of fodder plants for goats and sheep in Ghana
and Burkina Faso and found local reports of fodder potential for 95 genera and 52
plant families. Dechnik-Vázquez et al. (2019) search for potential new useful species

106
for silvopastoral purposes in Chiapas, Mexico. They used field observation by
searching for plants with browsing marks, interviews with stakeholders,
bromatological analysis and vegetative propagation tests. They describe cattle as
generalist browsers.
In some landscapes local people have decades or even centuries of experience
managing ecosystems for the production of resources that are of interest to humans.
The management of cattle in Madre de Dios is a recent occurrence, itself being only
recently colonized, and is predominantly practiced by migrant farmers from the
Andes. The present study was time limited, and doubtless would have benefitted
from a structured and formal review of LEK in the region. Nine informal and
unstructured discussions with cattle farmers were held in preparation for the species
selection in this study. A systematic review of LEK of forage plants in the Amazon is
needed to understand the knowledge farmers may have regarding potential browse
species. Such a review is currently lacking in the published literature.
Expert input for this study came from the author’s supervisory team who first
witnessed cattle browsing on Inga edulis. On a separate occasion the author
observed a plantation of three species of Inga that had been destroyed by cattle.
Additionally, information was requested from a local agroforestry practitioner who
reported the unintended access of 80-100 cattle to their plantation site for
approximately a three-hour period. The data they collected for their records is
presented below (Table 1). However, it should be noted that this was not a replicated
browsing trial by cattle, and some areas were more accessible than others.
Furthermore, no distinction was made on the type of damage sustained by the

107
plants, which may have included trampling or browsing. However, it was reported
that the I. edulis was palatable as cattle were observed to browse on this species.
Direct and indirect observation of browsing preference can be elicited using means
such as species identification and frequency of use as an indication of palatability.
Indirect observations of browsing habits can include monitoring of browsing damage,
and types of damage, on trees and shrubs. The final criteria used in the evaluation
and selection of species for the present study is synthesised in Table 1.
Table 1. Plantation tree species killed and damaged by exposure to approximately 80-100 cattle in a 3-hour period. Data provided by Camino Verde (2018). .
Species # of plants % plants
killed % plants damage
Inga edulis 12719 14 16
Musa sp. 1748 13 2
Theobroma grandiflora 268 6 4
Cocos nucifera 171 9 3
Euterpe oleracea 100 13 6
Hymenaea courbaril 50 14 4
Dipteryx odorata 100 6 6
Morus nigra 728 1 2
Calliandra angustifolia 875 1 1
Myrciaria dubia 100 4 1
Cedrela odorata 97 3 5
Carapa guianensis 152 1 3
Astrocaryum murumuru 48 2 0
Myroxylon balsamum 52 0 0
Genipa americana 115 0 0
Table 2. Criteria used in the selection of the final five species for the present experiment. Where ++ is best suited to -- worse suited, + or – are weak evidence to

108
support either way, and ? indicate there was no information found linking the species to the criteria.
Criteria Leucaena
leucocephala
Inga edulis
Senegalia loretensis
Ceiba pentandra
Erythrina berteroana
Palatable to Cattle ++ ++ ++ ++ ++
Nutritious to Cattle ++ ? ? ? ?
Non-toxic to Cattle - ? ? ? ?
Resilient to browsing ++ + ? ++ ?
Fast Growing ++ ++ ? ++ ?
High proportion of edible biomass - ++ ? -- ?
Can withstand high planting density ++ ++ ? - ++
Can colonise degraded soils ? ++ ? + ?
Can withstand full sun ++ ++ + ++ ?
Resilient to fire ? ? ? ? ? Adapted to site conditions ? ++ ? ++ ++
Fix atmospheric Nitrogen ++ ++ + -- ++ Low susceptibility to pests ++ ++ ? ? ?
Accessible stock ++ ++ ++ ++ ++
Easy to Grow ++ ++ ++ ++ ++
4.2. Species Information
4.2.1 Inga edulis
Species of the genus Inga are nitrogen fixing leguminous trees (subfamily
Mimosoideae) with a range from Mexico to Uruguay (Pennington, 1998). Authors
report they are fast growing (Figure 14), demonstrate excellent biomass production,
coppicing ability, produce copious amounts of leafy biomass which helps control
weed species and are well adapted to acid soil conditions (Szott et al., 1991; Leblanc
et al., 2006). Inga species are depicted in the Mochica pottery of Peru, dated from
100-800AD, and some species have been domesticated and are now widely
distributed (Leon, 1998). One domesticated species of Inga is Inga edulis, cultivated

109
for their edible fruit. Farmers identified I. edulis as “useful” in previous studies in the
Peruvian Amazon (Fujisaka et al., 2000). I. edulis has been used widely in the
recovery of degraded and abandoned lands in the Chanchamayo regions of Peru,
where it has been used on floodplain land, shade for coffee, recovery of nutrient poor
soils following demanding crops such as pineapple or ginger, areas with compacted
soils and scarce organic matter, recuperation of areas dominated by weed growth
(including various aggressive grasses) and as a pioneer species on land abandoned
by landslides (Leon et al., 2016)
Dechnik-Vázquez et al. (2019) found I. edulis was ‘eagerly eaten’ by cattle as
reported by cattle herders in La Sepultura Biosphere Reserve in Chiapas, Mexico.
They found 78% neutral detergent fiber, 64.6% acid detergent fiber, 57.6% non-
structural carbohydrates and 0.31 g/100g of condensable tannins in the foliage of I.
edulis. Staver (1989) considered I. edulis in association with Desmodium ovalifolium
as a “planted fallow suitable for grazing”, and suggested I. edulis ‘grew more
vigorously’ than Leucaena sp., Gliricidia sepium, and Calliandra calothyrsus when
planted in observation plots. Hohnwald (2010) reported that I. edulis was palatable
on a scale of very palatable, palatable or medium palatability. They report that in a
browsing trial by cattle I. edulis plants may have been too small to be detected by the
cattle among the tall adjacent grass. Hohnwald (2016) reported I. edulis as having
‘poor palatability’ for cattle.

110
Figure 14. Inga edulis growth (Height) over time (Months), from: (1) Sobanski and Marques (2014), (2) Alegre (2005), (3) Kanmegne et al., (2000), (4) Rezende & Vieira (2019), (5) Butterfield (1996), (6) Butterfield (1995), (7) Gonzalez and Fisher (1994) .
Authors have described plantations of I. edulis in planting arrangements of 3 x 3 m
(Gonzalez and Fisher, 1994; Butterfield, 1995; Haggar et al., 1997; Carpenter, 2006;
Sobanski and Marques, 2014), 3 x 1 m (Kanmegne, 2000), and 1.5 x 1.5 m (Alegre
et al., 2005). I. edulis has also been described as a contour hedgerow planted at a
spacing of 0.5 m (Alegre and Rao, 1996) . It is widely reported to be fast growing, in
the Ecuadorian amazon the trunk diameter of I. edulis grew a mean of 5 cm year-1
(Bishop, 1983). Kanmegne (2000) reports that in Southern Cameroon on a Ferric
Acrisol soil type, I. edulis grew to 3.7 m in 12 months and by 20 months had reached
a height of 7.7 m. The same study coppiced trees to 5 cm from ground level. After 9
months of regrowth, I. edulis produced 26.8 Mg ha-1 of leafy biomass. Alegre (2005)
found that I. edulis, grown in association with Centrosema macrocarpum, had
0
1
2
3
4
5
6
7
8
9
12(1
)
12,(
1)
12,(
2)
12,(
2)
12,(
3)
18,(
4)
20,(
3)
24,(
2)
24,(
2)
30,(
4)
36,(
4)
36,(
5)
36,(
6)
36,(
5)
36,(
2)
36,(
2)
36,(
7)
36,(
5)
42,(
4)
60,(
4)
Heig
ht
(m)
Months After Planting, (Reference)

111
increased height compared with monocultures of I. edulis. Sobanski and Marques
(2014) in Southern Brazil’s Atlantic Forest associated I. edulis with Urochloa cf.
humidicola or Urochloa arrecta on either Cambisol or Greysol soil, and plots were
either tilled or untilled. They found soil conditions were the principal predictor of
seedling performance. Fisher (1995) found I. edulis increased soil Nitrogen from 3.0
g kg-1 in 1987 to 3.4 g kg-1 by 1992 and extractable Phosphorous from 5.5 mg kg-1 to
7.2 mg kg-1 in the same 7-year period.
Figure 15. Biomass after 11 years (1993 – 2004) in five replications of two treatments, T1 (control) and T6 (interplanted with Inga edulis), in Mg ha-1. Biomass of weeds and Inga was measured in 1 x 3 m subplots within treatment plots and converted to a per hectare basis. Biomass of Terminalia amazonia is the sum of crown foliage and bole wood biomasses of a sample tree for each treatment plot. Crown and bole biomasses were estimated from volumes calculated from diameter and height measurements multiplied by foliage density and wood density, respectively. Once the tree biomass was obtained, biomass was converted to a per hectare basis depending on the number of trees in the plot. Total biomass is the sum of the weed and Inga biomass plus the T. amazonia biomass on a per hectare basis (from Nichols and Carpenter, 2006).

112
Nichols and Carpenter (2006) found that Terminalia amazonia showed higher rates
of biomass accumulation when interplanted with I. edulis than in a monoculture
control in 4 of 5 replicates (Figure 15). ‘Pioneer’ or ‘colonising’ tree species may
open up a niche in Brachiaria brizantha pastures which may result in soil moisture,
shade and lower levels of soil compaction (Khurana and Singh, 2001; Garen et al.,
2011). Otsamo (1998) found that Anisoptera marginata grew fastest when
interplanted with the nurse tree Paraserianthes falcataria on an Imperata cylindrica
grassland site in Indonesia. Similarly, the dense shade and ability to suppress weeds
may give I. edulis a unique advantage in the pasture environment and
simultaneously open a niche for additional biodiversity.
4.2.2 Ceiba pentandra
Ceiba pentandra (L.) Gaertn, also known as the kapok tree, is an anemochorous
Bombacaceae (Wikander, 1984). It is a deciduous broadleaved tree with alternate
palmate leaves composed of 5 to 9 leaflets each up to 15 cm long. It is a species
known to associate with arbuscular mychorrizae (Iversen et al., 2007). Román-
Dañobeytia et al. (2015) used C. pentandra in a study of rehabilitation of sites
degraded by alluvial mining in Madre de Dios. They considered C. pentandra to be a
mid successional species as they find they are capable of colonizing open areas but
are generally long‐ lived and grow taller than short-lived pioneer species (Román-
Dañobeytia et al., 2012). It is a species that has been reported to grow rapidly
(Richards, 1952 in Condit et al., 1993). Fertilizer application at an experimental site
in Kerala, India, was shown to “increase” seedling root collar diameter but have “little
effect” on height (Dileep et al., 1994). C. pentandra is a traditionally used forage in
Cambodian goat production. In one feeding trial in Cambodia the inclusion of C.

113
pentandra foliage increased live weight gains of goats by 26% over a 32-week period
(Kouch, Preston and Hieak, 2006). In a study by Kongmanila and Ledin (2009) in
Laos, C. pentandra foliage had a condensed tannin (CT) content in the leaves and
petioles (as dry matter) of 300 g/kg, and 231 g/kg of dry matter in the stem. When
offered to goats, the percentage intake of C. pentandra biomass was 3.8% of the
animal’s body weight, which was less (P<0.05) than Artocarpus heterophyllus (4.4%)
or Erythrina variegata (4.0%) but more (P<0.05) than Ficus racemosa (3.4%),
Mangifera indica (2.7%) or Ziziphus jujuba Mill (2.5%). Thanu, Kadirvel and
Ayyaluswami (1983) fed C. pentandra seeds to day-old New Hampshire chicks. A
control group where no seeds were fed gained 305 g, whereas birds on the raw seed
diet gained 243 g and those on seeds that had been oven dried gained 127 g. In
India, C. pentandra seeds were ground and mixed into linseed oilcakes fed to
Kumauni bullocks with no observed adverse effects (Sahai and Kehar, 1968). In
Ghana, Ansah and Nagbila (2011) report farmers use C. pentandra as fodder, and
also use the fresh pounded leaves medicinally to expel retained placentas in cattle.
There is a precedent for this species in plantations. Majid et al. (1998) grew C.
pentandra in Kampung Pasir, Malaysia, and found that after 23 months trees grown
in association with Calopogonium muconoides were significantly taller (380 cm) and
had a larger diameter (72 mm) than those grown in association with Centrosema
pubescens (height 280 cm / RCD 68 mm) or Puereria phaseoloides (height 220 cm /
RCD 55 cm). At Indira Gandhi Agricultural University, C. pentandra was harvested 9
years after planting at 4 x 4 m, 4 x 6 m and 4 x 8 m. The 4 x 4 m spacing had
reached a DBH of 16.56 cm and a height of 8.29 m, the 4 x 6 m had reached a DBH
of 16.28 cm and a height of 7.49 m and the 4 x 8 m had reached a DBH of 17.04 and

114
a height of 7.55 m (Gawali et al., 2015). This species was documented by Toledo-
Aceves and Swaine (2007) in the Bobiri Forest Reserve in Ghana to be susceptible
to fungal infections. They planted C. pentandra at 0.7 x 0.7 m and after 12 months
found that 39% of the C. pentandra seedlings had died. Another mortality rate,
reported by d’Oliveira (2000) for saplings planted in primary forest, was 10% after 12
months, which rose to an accumulated mortality of 25% after 24 months, which
d’Oliveira (2000) speculated was due to canopy closure.
4.2.3 Erythrina berteroana
Erythrina is a pantropical genus of flowering plants in the family Leguminosae
(subfamily Faboideae). Some species are used as living fences and they have
gained popularity as a species for alley cropping. They are easily propagated.
Camacho et al. (1993) reports that in Costa Rica, production can be 12-16 Mg ha
year-1 with pH of 4.0 and 50% aluminium saturation. Haque et al. (1986) report that
Erythrina are adapted to acid utisols. This is confirmed by a study by Salazar and
Palm (1987) that demonstrated that a species of Erythrina tolerated 75% aluminium
saturation.
Muschler et al. (1993) compared E. berteroana Urban, E. fusca Loureiro and
Gliricidia sepium (Jacq.) Steud in Talamanca, southern Costa Rica. E. berteroana
was found to have the greatest crown diameter (CDiam 2.18
m), crown depth (CDep 2.43 m) than E. fusca (CDiam 1.52 m, CDep 1.68 m) or G.
sepium (CDiam 130 m, CDep 2.65 m). E. berteroana was also reported to produce
the most foliar biomass (1100 g) when compared with E. fusca (800 g) or G. sepium

115
(500 g). Szott et al. (1991) reported that Erythrina sp. have “excellent biomass
production”, coppicing ability and are “well adapted” to acid soils. They report from
Yurimagas, in the northern Peruvian Amazon, that the leaves of Erythrina
mineralized 40% of their initial phosphorus and calcium contents and 75% of their
magnesium and potassium contents within 4 weeks. After 20 weeks there was 25%
or less of initial mineral levels. In the same experiment, after 4 weeks, Inga edulis
had 80% remaining of fallen leaf mass, by 20 weeks this had decreased to 70%.
Salazar et al. (1993) found that grain and stover (above ground biomass) of rice
production in the first cropping phase was greater under Erythrina sp.(4861 kg ha-1)
and Leucaena leucocephala (5289 kg ha-1) than that from Inga edulis (4171 kg ha-1).
By the third cropping phase there were (400 kg ha-1) of weed production in the I.
edulis plots, compared to 1200 kg ha-1 in the Erythrina sp. or 1300 kg ha-1 in the L.
leucocephala plots. Salazar et al. (1993) speculate that the greater biomass
increases in the first crop were due to rapid decomposition of Erythrina sp. and L.
leucocephala leaves and rapid transfer of the nutrients to the associated crops,
whereas I. edulis decomposed more slowly as evidenced by it’s greater weed
suppression, and may therefore have resulted in a delayed nutrient transfer to the
associated crops
Standley et al. (1946) describe how in some species of Erythrina, young leaves are
cooked and eaten like a vegetable. In Guatemala and El Salvador the young shoots
of E. berteroana are popular and their consumption is said to induce a relaxing sleep
(Morton, 1994). Standley et al. (1946) describe how other species of Erythrina have
toxic alkaloids in concentrations that enable the use of crushed branches as a fish

116
poison. In seeds the effects of the toxins are reported to be paralysis or death by
respiratory failure (Cowan et al., 1982). Leaves have been reported to be eaten by
cattle, goats and rabbits without ill effect (Cordero et al., 2003). Payne (1991) found
that b –erythroidine from Erythrina leaves were hydrogenated in the rumen of goats
and detectable in the milk. The same study found partial genetic control of the
alkaloid content.
In Costa Rica, Ibrahim and Mannetje (1998) found that E. berteroana could produce
an annual ‘edible’ yield of more than 11 Mg ha–1 dry matter when pruned every four
months. Experiments carried out at CATIE field sites in Costa Rica found that a
pruning interval of four months maintained the proportion of E. berteroana edible to
non edible biomass, and that when planted ‘similarly to sugarcane’ dry matter yields
of 16.4 Mg ha-1 were obtained after the first year (Pezo et al., 1990; Kass et al.,
1992). Pezo et al. (1990) suggest that E. berteroana had ‘higher’ nutritional quality
than E. fusca or E. costarricenses and that there is the potential for generic
improvement of the forage quality of E. berteroana.
4.2.4 Senegalia loretensis
Senegalia loretensis (J.F.Macbr) is synonymous with Acacia loretensis (J.F. Macbr.)
and Acacia riparia var. angustifoliola Kuntze. It is a tree in the Fabaceae family
(subfamily Mimosoideae). It is an evergreen tree with oblong unequal and small
bipinnate leaflets arranged on a secondary axis. It has scattered prickles. It is native
to Bolivia, Brazil North, Brazil Northeast, Ecuador and Peru. Little literature exists

117
with regard to this species. However, the author observed that it is a common tree
proximate to the study site and is highly palatable to cattle (Figure 16).
Figure 16. Little is known about Senegalia loretensis, but despite it’s sharp spines the leaves are highly palatable to cattle.
4.2.5 Leucaena leucocephala
Leucaena leucocephala (Lam.) de Wit (leucaena) is a shrub or tree in the
Leguminosae (subfamily Mimosoideae) family. It is native to Central America and is
used across the tropics as a multipurpose tree legume. It is long-lived and able to
withstand pruning, pollarding, coppicing and direct browsing. L. leucocephala is used
as a shade tree for plantation crops, and is also well known for its forage value,
despite containing the toxic compound mimosine (Dalzell et al., 2012). L.
leucocephala is promoted as a tree of high potential for agroforestry and
silvopastoral systems (Mauricio et al., 2019). In 1990, L. leucocephala was estimated
to cover 2-5 million hectares worldwide (Brewbaker and Sorensson, 1990). Szott et
al. (1991) conducted 6 years of experimental trials in Yurimaguas, Peru, and found

118
that L. leucocephala and its more acid-tolerant relative, L. diversifolia, failed to
develop adequately and speculate this may have been due to due to aluminum
toxicity.
4.2.6 Brachiaria brizantha
Brachiaria brizantha (Hochst. ex A. Rich.) Stapf. is in the family Poaceae. It is native
to Botswana, Cameroon, Côte D'Ivoire, Ethiopia, Ghana, Guinea, Kenya, Malawi,
Mozambique, Namibia, Nigeria, Sierra Leone, South Africa, Tanzania, Uganda,
Zaire, Zambia and Zimbabwe. Brachiaria species were first introduced to South
America during the 18th century when B. mutica was used as bedding on ships.
Subsequently the Food and Agriculture Organization introduced species of
Brachiaria in 1952 (Serrão et al., 1971). B. brizantha became a popular species due
to its antibiotic resistance to the spittlebug, which feeds on the xylem of Brachiaria
species causing chlorosis and necrosis (CIAT, 1992).
‘Marandú’ (CIAT 6294, BRA000591, CPI 81408, ILCA 16550) is a hairy Kenyan
accession that has good spittlebug resistance. An experiment to see which grass
would be the most appropriate for a silvopastoral system under a 12 year old rubber
plantation in Porto Velho, Rondônia, Brazil recommended B. brizantha cv. Marandú
(Costa et al., 2001). A random block design was employed with three replicates.
Grasses were sown in four rows of 4 m length with a spacing of 0.5 m. Harvest was
performed at 12 week intervals in the wet season and 16 week intervals in the dry
season. In total six evaluations were realized, four during the rainy season and two
in the dry season see Table 3 (Costa et al., 2001)

119
.
Table 3. Plant height, soil cover and dry matter yield of forage grasses Brachiaria brizantha cv. Marandú, B. humidicola, Paspalum atratum BRA-9610, P. guenoarum BRA-3824, P. regnelli BRA-0159, P. plicatulum BRA-9661 and Hemarthria altissima established under a 12 year old rubber plantation reproduced from Costa et al. (2001).
At low N inputs, dry matter yields have been higher in shade than in full sunlight
(Dias-Filho and De Carvalho, 2000). Marandú is intolerant to waterlogging and
flooding (Beloni et al., 2017; Dias-Filho and De Carvalho, 2000). Many species of
Brachiaria are reported to be allelopathic and the Marandú cultivar has been
associated with negative effects on associated Desmodium adscendens, Sida
rhombifolia and Vernonia polyanthes (Favaretto et al., 2018; Souza Filho et al.,
1997). Dupas et al. (2010) found that production of B. brizantha cv. Marandú
improves with N input and fertilization that resulted in an average production of 24.9
Mg ha-1 in the wet season and 8.1 Mg ha-1 in the dry season. Perondi and Oliveira
(2007) found that the annual production was of 23.7 Mg ha-1 of which 8.5 Mg ha-1
was produced in the dry season and 15.1 Mg ha-1 was produced in the wet season

120
4.3 Design and Management of Tree Nursery
A nursery where plants can be grown and monitored is essential prior to the
collection of seeds, as often seeds must be planted immediately. For this study, the
key elements of the nursery included access to water, control of sun exposure, and
an appropriate medium in which to grow the plants. The present site had no existing
well and therefore it was necessary to dig a well (Figure 17). It was also necessary to
build a structure upon which to house a water tank as this facilitated water pressure
with which to water the plants. Plants were watered in the evening due to the high
levels of solar irradiation at the site. The plants in this project were to be housed in
two separate structures (nurseries) of approximately 10 x 20 m each. Due to the
complete site clearance, there was no natural shade. Therefore, a shade nursery
netting of 80% was used to shade the plants. In order to erect this hollow galvanized
steel poles were cut to size and holes was drilled 2.5 cm from one end of each pole.
To stabilize the pole the hollow base of the pole was then fitted in a coaxial fashion
over an iron bar that had been set proud of a concrete base (Figure 17). Further
cement was added to fill the hole to ground level. Through the holes in the poles a
thick galvanized wire was run around the entire perimeter of each nursery, and
tightened (Figure 18). A polystyrene ball was placed on the top of each pole to
protect the mesh from tearing on the pole during extreme weather events.

121
Figure 17. (Clockwise from left to right) The first attempt at digging the well, the base of the steel poles, which contained a 2.5 cm iron bar, alignment of the steel poles for the nurseries.

122
Figure 18. The nursery mesh was elevated upon steel poles above the plants, with enough room to allow air to circulate and to enable easy access for watering the plants. Finally, the nursery mesh was sewn around the galvanized wire and pulled taught at
either end with guide ropes. The beds in which the plants would be accommodated
were on the ground, and defined as 1.20 in width and the length of the nursery. The
beds were lined with perforated plastic. This was to reduce the likelihood of the
plants setting root into the ground, and also to reduce the incidence of weeds
growing up into the bags. A more preferable technique is that of growing plants in
root trainers off the ground. However, such resources were not available for the
current project.
The next task involved preparing the potting medium (Figure 19). Topsoil was
harvested from areas of the site outside of the experimental zone that were identified
as having preferable topsoil. There was a notable lack of good quality soil at the site,
which may be been due to the site history, which was of a cattle pasture and

123
regeneration of secondary forest. The topsoil was very clay heavy and was mixed
with sand to decrease the compaction of the bags that occurs with repeated
watering. The eventual mix did not include any resources that were not found at the
site. This was because this stage was also intended to stimulate how the plants may
establish at the site. The use of fertilisers or enriched soils might have had a
confounding effect on the experiment. A total of 10,000 bags of soil were filled, with
the intention of producing 4,500 plants of five species (Figure 20). As 900 plants of
each species were required of the same age and treatment, an excess of 1,100 trees
were planted per species for a total of 2,000 per species.
Figure 19. Some prepared soil for filling the bags for the plants.

124
Figure 20. View over the nurseries with 10,000 bags of soil. Borders made from trees define the beds. The bags of soil are arranged in rows to give easy access for weeding and watering of the plants. In the background is the water tower that enabled water to the site.
4.4 Treatment and Sowing of Seeds
In September 2016, seed pods of C. pentandra were collected (Figure 21). The
seeds were separated from the fiber and soaked in water for 24 hours. Seeds were
then placed individually into the bags of soil. Seeds of I. edulis were collected,
soaked in a mash of roots from the parent tree for 24 hours, drained and individually
placed into bags of soil (Figure 22). Seeds of L. leucocephala were scarified, soaked
for 24 hours and individually placed into bags of soil (Figure 23, Figure 24). In
October, seeds of S. loretensis were collected, soaked in water for 24 hours and
sown individually into bags of soil (Figure 25). In February 2017, cuttings of E.
berteroana were harvested using a machete and cut into 1 m lengths; these were
planted directly into the planting lines at a depth of 30-40 cm (Figure 26). With the

125
exception of E. berteroana, all trees were grown from seed, kept in 80% shade and
watered daily. The moisture in the bags of soil determined the watering regime.
Plants were removed from the shade one month prior to planting and the watering
regime reduced.
Figure 21. Seed pods and leaves of C. pentandra.

126
Figure 22. I edulis
Figure 23. Seeds of L. leucocephala

127
Figure 24. Seedlings of L. leucocephala
Figure 25. Beds of S. loretensis (foreground) and C. pentandra (background).

128
Figure 26. E. berteroana was planted from stakes, pictured here 2 months post planting in association with the immature pasture grass.
5. Planting Design
The present study envisaged a silvopastoral system in which a tree and shrub
element would be directly browsed by cattle. Tree and shrub elements can be
thought of as ‘vertical’ forage, whereas grasses and ground cover can be described
as a as ‘horizontal’ forage. There is competition between proximate plants for water,
nutrients and sun. Therefore, it can be conceptualised that the production of foliar
biomass and the leaf area of a vertical forage element will reduce the sun exposure
of adjacent horizontal forage. To minimise the shading effect of the vertical forage
the design of agroforestry may incorporate an east to west planting line, or ‘alley
rows’. Species can respond to different planting densities and arrangements by
adapting their ratio of wood to leaf production. To optimise production, trees in rows

129
should be planted as densely as possible without compromising the production of
foliar biomass. The aim is to compensate for the effect of the reduction of horizontal
foliage by producing more vertical foliage. Furthermore, if dense initial planting
proves problematic this can be amended by management techniques such as
thinning, pollarding or the application of fertilisers. The altitude of the sun relative to
the site can be considered when designing appropriate planting arrangements. In
this study where the aim was to maximise available forage to cattle, allowing the
trees to grow taller than the cattle could reach would reduce the light available for the
associated grass. If the maximum height of browsing cattle is 2 meters height, then
this value of 2 m can be used in conjunction with the altitude angle of the sun at the
site to determine the optimum width of the alley row (Leroy et al., 2009; Reid and
Ferguson, 1992). This can be calculated by:
(Equation 1)
Length of shadow = Height of vertical forage Tan(altitude angle of sun)
The above equation can be expanded to include, for example, the light interception
of the trees, and the response to shade of the associated horizontal forage element
(Dupraz et al., 2019; Zhang et al., 2019).
At the site in the present study, trees of all species were planted at a spacing of 0.5 x
7.5 m in east to west lines. The planting of 0.5 m in rows was chosen based on the
experience of previous authors with I. edulis (Alegre and Rao, 1996; Villachica,
1996; Leblanc et al., 2006). A distance of 7.5 m was left between the three rows of
each parcel. A planting arrangement of 0.5 x 7.5 m is 2,667 trees per hectare.
Daylight duration was at a maximum in January and minimum in July (Table 4). The

130
length of a shadow of an object of 2 meters height was at its shortest in November
and its longest in July.
Table 4. Data for the first day of each month from a site in Madre de Dios, Peru (lat -12.53945°, long -69.17288°) in 2016, including the altitude and azimuth of the sun at 12:00, the shadow cast by an object of 2 m height and the daylight duration.
First day of Altitude (°) Azimuth (°)
Shadow of object at 2 m
height (m) Daylight Duration
January 78.58 203.47 0.40 12h51m29s
February 84.99 207.63 0.18 12h38m52s
March 84.21 331.84 0.20 12h20m34s
April 72.09 344.03 0.65 11h58m49s
May 61.51 346.65 1.09 11h39m49s
June 54.81 349.75 1.41 11h26m33s
July 54.09 352.38 1.45 11h24m36s
August 59.29 352.08 1.19 11h34m43s
September 68.55 344.02 0.79 11h52m41s
October 77.6 317.07 0.44 12h13m8s
November 80.14 257.01 0.35 12h34m3s
December 77.61 220.04 0.44 12h49m5s
6. Experimental Design
The experiment used a randomised block design with five replicates. There were six
treatments in the experiment, with each treatment associated with a pasture of
Brachiaria brizantha cv. Marandú. The treatments were (1) Leucaena leucocephala,
(2) Inga edulis, (3) Ceiba pentandra, (4) Senegalia loretensis, (5) Erythrina
berteroana, (6) Control of Brachiaria brizantha cv. Marandú with no tree association.
Ideally the experiment would have been arranged as a 6 x 5 plot rectangle to
minimise edge effects. However, the site area dedicated to the system was 300 x
100 m so the parcel layout was 3 x 10 plots. A moisture gradient was suspected so
the site was divided into 5 blocks of 6. Thirty plots of 30 x 30 m (0.09 ha) were
installed in January 2017 and randomly allocated between the 6 treatments species

131
using the statistics program R (version 3.2.2 for OS X)(Figure 27). Each plot
therefore encompassed 180 trees, distributed in three alley rows of 0.5 x 7.5 m, and
each row of trees was 30 m long and contained 60 trees. The rows between each
replicate were separated by 15 m on the north and south side. The rows were
continuous between plots and species (Figure 28). In other words, there was a 0.5 m
break between one species and the next in a row (Figure 29). To account for the
potential edge effects of this design, the last 5 trees in each row were excluded from
subsequent censuses of plant growth.
Figure 27. The randomised replicate plot layout, where EB is Erythrina berteroana, IE is Inga edulis, LL is Leucaena leucocephala, SL is Senegalia loretensis, CP is Ceiba pentandra and C is a control of grass.

132
Figure 28. Map of experimental site. 0.09 ha plots as red rectangles, tree rows as black dashed lines. Tree rows were continuous between plots. There were no tree lines on the red lines.
Figure 29. Satellite image of experimental site, 0.09 ha parcels as red rectangles. Image taken in April 2018. Image by Maxar Technologies.
6.1 Planting Sequence
A map of the planting area was made in ArcGIS (ArcGIS Developer V.1.2). Due to
the shape of the site it was necessary to stagger the parcels in order to
accommodate them. The coordinates of the corners of each of the parcels were then
transferred onto a Trimble Geo 7X handheld GPS and the 60 points were located to
1-2 m accuracy. A 100 m tape measure was then used to corroborate the spacing of

133
the points. All points were marked plastic tubes that were 4 meters long and buried 1
m into the ground at each point (Figure 30). Using a 30 m tape measure three
additional points were located in each parcel at 7.5 m. These 7.5 m points were the
planting lines within each plot and these points were marked with wooden stakes. A
plastic twine was then held taught between the two stakes and any remaining
vegetation was cleared from the planting lines. Small, flat, wooden splints were
placed in the ground at 0.5 m increments using a 30 m tape measure on the ground.
Manual post diggers were used to dig holes, aiming to keep the wooden splint in the
middle of the hole to be dug. The top 5 cm of soil was returned to the base of each
hole at the time of planting.
Figure 30. The plastic tubes used to mark the 30 m intervals of the plot corners and in the background weeds responding to the glyphosphate applied across the site.
Planting of L. leucocephala began on the 20th of January 2017 and was completed
on the 26th January 2017 (Figure 31). Planting of I. edulis began on the 1st February

134
2017 and was completed on February 6th 2017 (Figure 32). Planting of C. pentandra
began on the 6th February 2017 and finished on the 11th February 2017. Planting of
S. loretensis began on the 11th February 2017 and was completed on the 18th
February 2017. E. berteroana were harvested and planted between the 20th
February 2017 and the 1st March 2017. From April – May 2017 the grass Brachiaria
brizantha cv. Marandú was sown by hand from west to east across the entire
experiment. Working in pairs, one person would drive a 2 m wooden stake with a
pointed end approximately 2.5 cm into the ground at a spacing of 25 x 25 cm. A
person would follow behind dropping approximately 0.05 g of seed in each hole. This
procedure was followed across all 30 parcels (Figure 33).

135
Figure 31.Team planting L. leucocephala in January of 2017.
Figure 32. Planting lines of I. edulis.

136
Figure 33. Satellite image from April 2017 with the parcels projected, showing faint tree lines and the establishing grass.
References
Alegre, J.C., Rao, M.R., 1996. Soil and water conservation by contour hedging in the
humid tropics of Peru. Agriculture, Ecosystems & Environment 57, 17–
25. https://doi.org/10.1016/0167-8809(95)01012-2
Beloni, T., Pezzopane, C.G., Rovadoscki, G.A., Fávero, A.P., Dias-Filho, M.B.,
Santos, P.M., 2017. Morphological and physiological responses and the recovery
ability of Paspalum accessions to water deficit and waterlogging. Grass and Forage
Science 72, 840–850. https://doi.org/10.1111/gfs.12281
Benavides, J.E., 1995. Tropical Feeds and Feeding Systems Research on Forage
Trees, in: FAO (Ed.), Electronic Conference on Tropical Feeds and Feeding
Systems.

137
Bishop, J.P., 1983. Tropical forest sheep on legume forage/fuelwood fallows.
Agroforestry Systems 1, 79–84. https://doi.org/10.1007/BF00596350
Brewbaker, J.L., Sorensson, C.T., 1990. New tree crops from interspecific leucaena
hybrids, In: Janick, J., Simon, J.E. (Eds) Timber Press, Portland, OR.
Butterfield, R.P., 1995. Promoting biodiversity: advances in evaluating native species
for reforestation. Forest Ecology and Management 75, 111–
121. https://doi.org/10.1016/0378-1127(95)03535-I
Campos, L.Z., Nascimento, A.L.B., Albuquerque, U.P., Araújo, E.L., 2018. Use of
local ecological knowledge as phenology indicator in native food species in the
semiarid region of Northeast Brazil. Ecological Indicators 95, 75–
84. https://doi.org/10.1016/J.ECOLIND.2018.07.016
Chávez Michaelsen, A., Huamani Briceño, L., Fernandez Menis, R., Bejar Chura, N.,
Valera Tito, F., Perz, S., Brown, I., Domínguez Del Aguila, S., Pinedo Mora, R.,
Alarcón Aguirre, G., 2013. Regional Deforestation Trends within Local Realities:
Land-Cover Change in Southeastern Peru 1996–2011. Land 2, 131–
157. https://doi.org/10.3390/land2020131
CIAT, 1992. Pastures for the Tropical Lowlands, CIAT’s Contribution. Centro
Internacional de Agricultura Tropical, Cali, Colombia.
Condit, R., Hubbell, S.P., Foster, R.B., 1993. Identifying fast-growing native trees

138
from the neotropics using data from a large, permanent census plot. Forest Ecology
and Management 62, 123–143. https://doi.org/10.1016/0378-1127(93)90046-P
Cordero, J., Boshier, D., Barrance, A., Centro Agronomico Tropical de Investigacion
y Ensenanza., Oxford Forestry Institute., Great Britain. Forestry Research
Programme., 2003. Arboles de Centroamerica : un manual para extensionistas.
Centro Agronomico Tropical de Investigacion y Ensenanza.
Costa, N.L., Paulino, V.T., Igreja, A.C.M., Townsend, C.R., Magalhães, J.A., Pereira,
R.G., Paulino, T.S., 2001. Agronomic evaluation of forage grasses under mature
rubber plantation. EMBRAPA, Roraima.
Cowan, R.S., Allen, O.N., Allen, E.K., 1982. The Leguminosae: A Source Book of
Characteristics, Uses, and Nodulation. Taxon 31,
133. https://doi.org/10.2307/1220602
Dalzell, S.A., Burnett, D.J., Dowsett, J.E., Forbes, V.E., Shelton, H.M., 2012.
Prevalence of mimosine and DHP toxicity in cattle grazing Leucaena leucocephala
pastures in Queensland, Australia. Animal Production Science 52,
365. https://doi.org/10.1071/AN11236
Dechnik-Vázquez, Y.A., García-Barrios, L., Ramírez-Marcial, N., van Noordwijk, M.,
Alayón-Gamboa, A., 2019. Assessment of browsed plants in a sub-tropical forest
frontier by means of fuzzy inference. Journal of Environmental Management 236,
163–181. https://doi.org/10.1016/J.JENVMAN.2019.01.071

139
Dias-Filho, M.B., De Carvalho, C.J.R., 2000. Physiological and morphological
responses of Brachiaria spp. to flooding. Pesquisa Agropecuaria Brasileira 35,
1959–1966. https://doi.org/10.1590/S0100-204X2000001000006
Dileep, M., Sudhakara, K., Santhoshkumar, A. V., Nazeema, K.K., Ashokan, P.K.,
1994. Effect of seed size, rooting medium and fertilizers on the growth of seedlings
of Ceiba pentandra (Linn.) Gaertn., Indian Journal of Forestry. Bishen Singh
Mahendra Pal Singh.
D’Oliveira, M.V.N., 2000. Artificial regeneration in gaps and skidding trails after
mechanised forest exploitation in Acre, Brazil. Forest Ecology and Management 127,
67–76. https://doi.org/10.1016/S0378-1127(99)00117-6
Duchelle, A.E., Guariguata, M.R., Less, G., Albornoz, M.A., Chavez, A., Melo, T.,
2012. Evaluating the opportunities and limitations to multiple use of Brazil nuts and
timber in Western Amazonia. Forest Ecology and Management 268, 39–
48. https://doi.org/10.1016/J.FORECO.2011.05.023
Dupraz, C., Wolz, K.J., Lecomte, I., Talbot, G., Vincent, G., Mulia, R., Bussière, F.,
Ozier-Lafontaine, H., Andrianarisoa, S., Jackson, N., Lawson, G., Dones, N.,
Sinoquet, H., Lusiana, B., Harja, D., Domenicano, S., Reyes, F., Gosme, M., Van
Noordwijk, M., 2019. Hi-sAFe: A 3D Agroforestry Model for Integrating Dynamic
Tree–Crop Interactions. Sustainability 11, 2293. https://doi.org/10.3390/su11082293

140
Eswaran, H., Rice, T., Ahrens, R., Stewart, B.A. (Eds.), 2002. Soil Classification.
CRC Press. https://doi.org/10.1201/9781420040364
FAO, 2015. World reference base for soil resources, International soil classification
system for naming soils and creating legends for soil maps.
Favaretto, A., Scheffer-Basso, S.M., Perez, N.B., 2018. Allelopathy in Poaceae
species present in Brazil. A review. Agronomy for Sustainable Development 38,
22. https://doi.org/10.1007/s13593-018-0495-5
Fisher, R.F., 1995. Amelioration of degraded rain forest soils by plantations of native
trees. Soil Science Society of America Journal 59, 544–
549. https://doi.org/10.2136/sssaj1995.03615995005900020039x
Fuchs, R., Alexander, P., Brown, C., Cossar, F., Henry, R.C., Rounsevell, M., 2019.
Why the US–China trade war spells disaster for the Amazon. Nature 567, 451–
454. https://doi.org/10.1038/d41586-019-00896-2
Fujisaka, S., Escobar, G., Veneklaas, E.J., 2000. Weedy fields and forests:
interactions between land use and the composition of plant communities in the
Peruvian Amazon. Agriculture, Ecosystems & Environment 78, 175–
186. https://doi.org/10.1016/S0167-8809(99)00122-X
Garen, E.J., Saltonstall, K., Ashton, M.S., Slusser, J.L., Mathias, S., Hall, J.S., 2011.
The tree planting and protecting culture of cattle ranchers and small-scale

141
agriculturalists in rural Panama: Opportunities for reforestation and land restoration.
Forest Ecology and Management 261, 1684–
1695. https://doi.org/10.1016/j.foreco.2010.10.011
Hijmans, R.J., Cameron, S.E., Parra, J.L., Jones, P.G., Jarvis, A., 2005. Very high
resolution interpolated climate surfaces for global land areas. International Journal of
Climatology 25, 1965–1978. https://doi.org/10.1002/joc.1276
Ibrahim, M., Mannetje, L.T., 1998. Compatibility, persistence and productivity of
grass-legume mixtures in the humid tropics of Costa Rica. 1. Dry matter yield,
nitrogen yield and botanical composition. Tropical Grasslands. 2, 96–104.
Iversen C.M., McCormack M.L., Powell A.S., Blackwood C.B., Freschet G.T., Kattge
J., Roumet C., Stover D.B., Soudzilovskaia N.A., Valverde-Barrantes O.J., van
Bodegom P.M., Violle C. (2017) A global Fine-Root Ecology Database to address
belowground challenges in plant ecology. New Phytologist. doi:10.1111/nph.14486.
Kass, M., Benavides, J., Romero, F., Pezo, D., Speedy, A., Pugliese, P.L., 1992.
Lessons from main feeding experiments conducted at CATIE using fodder trees as
part of the N-ration. In: Legume Trees and Other Fodder Trees as Protein Sources
for Livestock. FAO Animal Production and Health Paper. Food and Agriculture
Organization of the United Nations, pp. 161–175.
Kattge, J., Díaz, S., Lavorel, S., Prentice, I.C., Leadley, P., Bönisch, G., Garnier, E.,
Westoby, M., Reich, P.B., Wright, I.J., Cornelissen, J.H.C., Violle, C., Harrison, S.P.,

142
Van Bodegom, P.M., Reichstein, M., Enquist, B.J., Soudzilovskaia, N.A., Ackerly,
D.D., Anand, M., Atkin, O., Bahn, M., Baker, T.R., Baldocchi, D., Bekker, R., Blanco,
C.C., Blonder, B., Bond, W.J., Bradstock, R., Bunker, D.E., Casanoves, F.,
Cavender-Bares, J., Chambers, J.Q., Chapin, F.S., Chave, J., Coomes, D.,
Cornwell, W.K., Craine, J.M., Dobrin, B.H., Duarte, L., Durka, W., Elser, J., Esser,
G., Estiarte, M., Fagan, W.F., Fang, J., Fernández-Méndez, F., Fidelis, A., Finegan,
B., Flores, O., Ford, H., Frank, D., Freschet, G.T., Fyllas, N.M., Gallagher, R. V.,
Green, W.A., Gutierrez, A.G., Hickler, T., Higgins, S.I., Hodgson, J.G., Jalili, A.,
Jansen, S., Joly, C.A., Kerkhoff, A.J., Kirkup, D., Kitajima, K., Kleyer, M., Klotz, S.,
Knops, J.M.H., Kramer, K., Kühn, I., Kurokawa, H., Laughlin, D., Lee, T.D.,
Leishman, M., Lens, F., Lenz, T., Lewis, S.L., Lloyd, J., Llusià, J., Louault, F., Ma, S.,
Mahecha, M.D., Manning, P., Massad, T., Medlyn, B.E., Messier, J., Moles, A.T.,
Müller, S.C., Nadrowski, K., Naeem, S., Niinemets, Ü., Nöllert, S., Nüske, A., Ogaya,
R., Oleksyn, J., Onipchenko, V.G., Onoda, Y., Ordoñez, J., Overbeck, G., Ozinga,
W.A., Patiño, S., Paula, S., Pausas, J.G., Peñuelas, J., Phillips, O.L., Pillar, V.,
Poorter, H., Poorter, L., Poschlod, P., Prinzing, A., Proulx, R., Rammig, A., Reinsch,
S., Reu, B., Sack, L., Salgado-Negret, B., Sardans, J., Shiodera, S., Shipley, B.,
Siefert, A., Sosinski, E., Soussana, J.F., Swaine, E., Swenson, N., Thompson, K.,
Thornton, P., Waldram, M., Weiher, E., White, M., White, S., Wright, S.J., Yguel, B.,
Zaehle, S., Zanne, A.E., Wirth, C., 2011. TRY - a global database of plant traits.
Global Change Biology 17, 2905–2935. https://doi.org/10.1111/j.1365-
2486.2011.02451.x
Khurana, E., Singh, J.S., 2001. Ecology of seed and seedling growth for
conservation and restoration of tropical dry forest : a review. Environmental

143
Conservation 28, 39–52. https://doi.org/10.1017/S0376892901000042
Kirkby, C.A., Giudice, R., Day, B., Turner, K., Soares-Filho, B.S., Oliveira-Rodrigues,
H., Yu, D.W., 2011. Closing the ecotourism-conservation loop in the Peruvian
Amazon. Environmental Conservation 38, 6–
17. https://doi.org/10.1017/S0376892911000099
Kongmanila, D., Ledin, I., 2009. Chemical Composition of Some Tropical Foliage
Species and Their Intake and Digestibility by Goats. Asian-Australasian Journal of
Animal Sciences 22, 803–811. https://doi.org/10.5713/ajas.2009.80589
Lawrence, D., Vandecar, K., 2015. Effects of tropical deforestation on climate and
agriculture. Nature Climate Change 5, 27–36. https://doi.org/10.1038/nclimate2430
Leblanc, H.A., Nygren, P., McGraw, R.L., 2006. Green mulch decomposition and
nitrogen release from leaves of two Inga spp. in an organic alley-cropping practice in
the humid tropics. Soil Biology and Biochemistry 38, 349–
358. https://doi.org/10.1016/J.SOILBIO.2005.05.012
Leon, J.G., Reynel, C.R., Pennington, T.D., 1996. Agroforesteria y recuperacion de
areas degradadas con arboles de Inga (Pacae, Guaba) en el valle de
Chanchamayo, Departamento de Junin (Peru), Universidad Nacional Agraria La
Molina, Lima, Peru

144
Leon, J., 1998. History of the utilization of INGA as fruit trees in Mesoamerica and
Peru. Chapter 1 in The Genus Inga, Utilization, T.D. Pennington and E.C.M.
Fernandez (editors). The Royal Botanic Gardens, Kew, Continental Printing,
Belgium.
Leroy, C., Sabatier, S., Wahyuni, N.S., Barczi, J.-F., Dauzat, J., Laurans, M., Auclair,
D., 2009. Virtual trees and light capture: a method for optimizing agroforestry stand
design. Agroforestry Systems 77, 37–47. https://doi.org/10.1007/s10457-009-9232-z
Lima, L.S., Coe, M.T., Soares Filho, B.S., Cuadra, S. V., Dias, L.C.P., Costa, M.H.,
Lima, L.S., Rodrigues, H.O., 2014. Feedbacks between deforestation, climate, and
hydrology in the Southwestern Amazon: implications for the provision of ecosystem
services. Landscape Ecology 29, 261–274. https://doi.org/10.1007/s10980-013-
9962-1
Mann, C.C., 2008. Ancient Earthmovers of the Amazon. Science 321, 1148–1152.
Marengo, J.., Souza, C.., Thonicke, K., Burton, C., Halladay, K., Betts, R.A., Alves,
L.., Soares, W.B., 2018. Changes in Climate and Land Use Over the Amazon
Region: Current and Future Variability and Trends. Frontiers in Earth Science 6, 1–
21. https://doi.org/10.3389/feart.2018.00228
Mauricio, R.M., Ribeiro, R.S., Paciullo, D.S.C., Cangussú, M.A., Murgueitio, E.,
Chará, J., Estrada, M.X.F., 2019. Silvopastoral Systems in Latin America for
Biodiversity, Environmental, and Socioeconomic Improvements, in: Agroecosystem

145
Diversity. Elsevier, pp. 287–297. https://doi.org/10.1016/B978-0-12-811050-8.00018-
2
Moreira, F.M.S., Siqueira, J.O., Brussard, L., 2006. Soil biodiversity in Amazonian
and other Brazilian ecosystems, Soil Biodiversity in Amazonian and other Brazilian
Ecosystems. CABI Pub.
Morton, J.F., 1994. Pito (Erythrina berteroana) and chipilin (Crotalaria longirostrata),
(fabaceae) two soporific vegetables of Central America. Economic Botany 48, 130–
138. https://doi.org/10.1007/BF02908199
Murgueitio, E., Calle, Z., Uribe, F., Calle, A., Solorio, B., 2011. Native trees and
shrubs for the productive rehabilitation of tropical cattle ranching lands. Forest
Ecology and Management 261, 1654–
1663. https://doi.org/10.1016/j.foreco.2010.09.027
Muschler, R.G., Nair, P.K.R., Meléndez, L., 1993. Crown development and biomass
production of pollarded Erythrina berteroana, E. fusca and Gliricidia sepium in the
humid tropical lowlands of Costa Rica. Agroforestry Systems 24, 123–
143. https://doi.org/10.1007/BF00706887
Naah, J.B.S.N., Guuroh, R.T., 2017. Factors influencing local ecological knowledge
of forage resources: Ethnobotanical evidence from West Africa’s savannas. Journal
of Environmental Management 188, 297–
307. https://doi.org/10.1016/J.JENVMAN.2016.11.064

146
Nichols, J.D., Carpenter, F.L., 2006. Interplanting Inga edulis yields nitrogen benefits
to Terminalia amazonia. Forest Ecology and Management 233, 344–
351. https://doi.org/10.1016/j.foreco.2006.05.031
Nikitina, D., Remizova, L., Losco, R.L., 2011. A Preliminary Investigation of the Soils
and Geomorphology of a Portion of the Madre de Dios Region, Peru. Soil Horizons
52, 40. https://doi.org/10.2136/sh2011.2.0040
Panisset, J.S., Gouveia, C.M., Libonati, R., Peres, L., Machado-Silva, F., França,
D.A., França, J.R.A., 2017. Evaluation of last extreme drought events in Amazon
basin using remotely sensing data. 19th EGU General Assembly, EGU2017,
proceedings from the conference held 23-28 April, 2017 in Vienna, Austria., p.16144
19, 16144.
Payne, L., 1991. The Alkaloids of Erythrina: Clonal Evaluation and Metabolic
Fate. Ph.D. Dissertation. Department of Chemistry. Louisiana State Univérsity. 160
p.
Pennington, T.D., 1998. Chapter 2. Growth and Biomass Production of INGA
Species in The Genus Inga, Utilization, T.D. Pennington and E.C.M. Fernandez
(editors). The Royal Botanic Gardens, Kew, Continental Printing, Belgium.
Perz, S.G., Espin, J., Castillo, J., Chavez, A., Rojas, R.O., Barnes, G., 2016. Ideal
type theories and concrete cases in land science: A multi-step appraisal of the

147
evolutionary theory of land rights in Madre de Dios, Peru. Land Use Policy 58, 9–
20. https://doi.org/10.1016/J.LANDUSEPOL.2016.07.008
Pezo, D., Kass, M., Benavides, J., Romero, F. and Chaves, C., 1990. Potential of
legume tree fodders as animal feed in Central America. In Shrubs and tree fodders
for farm animals: proceedings of a workshop in Denpasar, Indonesia, 24-29 July
1989. IDRC, Ottawa, ON, CA.
Rammig, A., Lapola, D.M., Pinho, P., Quesada, C.A.N., Brown, I.F., Kruijt, B.,
Premebida, A., Ometto, J.P.H.B., Marengo, J.A., Vergara, W., Nobre, C.A., 2018.
Estimating the likelihood of an Amazon forest dieback and potential socio-economic
impacts. 20th EGU General Assembly, EGU2018, Proceedings from the conference
held 4-13 April, 2018 in Vienna, Austria
Reid, R., Ferguson, I.S., 1992. Development and validation of a simple approach to
modelling tree shading in agroforestry systems. Agroforestry Systems 20, 243–
252. https://doi.org/10.1007/BF00053142
Román-Dañobeytia, F., Huayllani, M., Michi, A., Ibarra, F., Loayza-Muro, R.,
Vázquez, T., Rodríguez, L., García, M., 2015. Reforestation with four native tree
species after abandoned gold mining in the Peruvian Amazon. Ecological
Engineering 85, 39–46. https://doi.org/10.1016/j.ecoleng.2015.09.075
Román-Dañobeytia, F.J., Levy-Tacher, S.I., Aronson, J., Rodrigues, R.R.,
Castellanos-Albores, J., 2012. Testing the Performance of Fourteen Native Tropical

148
Tree Species in Two Abandoned Pastures of the Lacandon Rainforest Region of
Chiapas, Mexico. Restoration Ecology 20, 378–386. https://doi.org/10.1111/j.1526-
100X.2011.00779.x
Sahai, B., Kehar, N.D., 1968. Investigations on subsidiary feeds. Kapok (Ceiba
pentandra) seed as a feed for livestock. Indian Journal of Veterinary Science and
Animal Husbandry 38, 670–673.
Salazar, A., Szott, L.T., Palm, C.A., 1993. Crop-tree interactions in alley cropping
systems on alluvial soils of the Upper Amazon Basin. Agroforestry Systems 22, 67–
82. https://doi.org/10.1007/BF00707472
Sampaio, G., Nobre, C., Heil Costa, M., Satyamurty, P., Silveira Soares-Filho, B.,
Cardoso, M., 2007. Regional climate change over eastern Amazonia caused by
pasture and soybean cropland expansion. Geophysical Research Letters 34,
17709. https://doi.org/10.1029/2007GL030612
Sampaio, G., Borma, L.S., Cardoso, M., Alves, L.M., von Randow, C., Rodriguez,
D.A., Nobre, C.A., Alexandre, F.F., 2019. Assessing the Possible Impacts of a 4 °C
or Higher Warming in Amazonia, in: Climate Change Risks in Brazil. Springer
International Publishing, Cham, pp. 201–218. https://doi.org/10.1007/978-3-319-
92881-4_8
Sanchez, P.A., 1995. Science in agroforestry. Agroforestry Systems 30, 5–
55. https://doi.org/10.1007/BF00708912

149
Scullion, J.J., Vogt, K.A., Sienkiewicz, A., Gmur, S.J., Trujillo, C., 2014. Assessing
the influence of land-cover change and conflicting land-use authorizations on
ecosystem conversion on the forest frontier of Madre de Dios, Peru. Biological
Conservation 171, 247–258. https://doi.org/10.1016/J.BIOCON.2014.01.036
Serrão, E.A.D., Simão Neto, M., 1, 1971. Informações sobre duas espécies de
gramíneas forrageiras do gênero Brachiaria na Amazônia : B.decumbens Stapf e
B.ruziziensis Germain et Evrard, in: Estudos Sobre Forrageiras Na Amazônia. p. 3.
Soares-Filho, B.S., Nepstad, D.C., Curran, L.M., Cerqueira, G.C., Garcia, R.A.,
Ramos, C.A., Voll, E., McDonald, A., Lefebvre, P., Schlesinger, P., 2006. Modelling
conservation in the Amazon basin. Nature 440, 520–
523. https://doi.org/10.1038/nature04389
Souza Filho, A.P.S., Rodrigues, L.R., Rodrigues, T.D., 1997. Inibição da germinação
e alongamento da radícula de invasoras de pastagens pelos extratos aquosos de
gramineas forrageiras tropicais. Pasturas Tropicales 19, 45–50.
Spracklen, D. V., Garcia-Carreras, L., 2015. The impact of Amazonian deforestation
on Amazon basin rainfall. Geophysical Research Letters 42, 9546–
9552. https://doi.org/10.1002/2015GL066063
Staver, C., 1989. Shortened bush fallow rotations with relay-cropped Inga edulis and
Desmodium ovalifolium in wet central Amazonian Peru. Agroforestry Systems 8,

150
173–196. https://doi.org/10.1007/BF00123120
Szott, L.T., Palm, C.A., Sanchez, P.A., 1991. Agroforestry In Acid Soils Of The
Humid Tropics. Advances in Agronomy 45, 275–301. https://doi.org/10.1016/S0065-
2113(08)60043-7
Toledo-Aceves, T., Swaine, M.D., 2007. Effect of three species of climber on the
performance of Ceiba pentandra seedlings in gaps in a tropical forest in Ghana.
Journal of Tropical Ecology 23, 45–52. https://doi.org/10.1017/S0266467406003725
Tomasini, S., Theilade, I., 2019. Local ecological knowledge indicators for wild plant
management: Autonomous local monitoring in Prespa, Albania. Ecological Indicators
101, 1064–1076. https://doi.org/10.1016/J.ECOLIND.2019.01.076
Torres-Sovero, C., González, J.A., Martín-López, B., Kirkby, C.A., 2012. Social–
ecological factors influencing tourist satisfaction in three ecotourism lodges in the
southeastern Peruvian Amazon. Tourism Management 33, 545–
552. https://doi.org/10.1016/J.TOURMAN.2011.06.008
Villachica, H., de Carvalho, J.E.U., Mercado, R.S., Izquierdo, J. and Egg, A.B.,
1996. Frutales y hortalizas promisorios de la Amazonia (Vol. 44). Lima: Tratado de
Cooperacion Amazonica, Secretaria Pro-Tempore.
Westoby, M., Wright, I.J., 2006. Land-plant ecology on the basis of functional traits.
Trends in Ecology & Evolution 21, 261 268.

151
https://doi.org/10.1016/J.TREE.2006.02.004
Wikander, T., 1984. Mecanismos de Dispersion de Diasporas de una Selva Decidua
en Venezuela. Biotropica 16, 276. https://doi.org/10.2307/2387936
Zhang, W., Wang, B.J., Gan, Y.W., Duan, Z.P., Hao, X.D., Xu, W.L., Li, L.H., 2019.
Different tree age affects light competition and yield in wheat grown as a companion
crop in jujube-wheat agroforestry. Agroforestry Systems 93, 653–
664. https://doi.org/10.1007/s10457-017-0160-z

-152-
CHAPTER III
TREE GROWTH IMPACTS ON GRASS AND SOILS IN FIVE MONOSPECIFIC
SILVOPASTORAL SYSTEMS IN AMAZONIA
Abstract
The integration of trees in pastures, known as silvopastoral systems, relies on
selection of species that are appropriate for a given site. This study seeks to support
the development of silvopastoral systems by critically evaluating the growth of trees
and pasture in a Brachiaria brizantha cv. Marandú pasture in the southwestern
Amazon. Leucaena leucocephala, Inga edulis, Erythrina berteroana, Ceiba
pentandra, Senegalia loretensis and a no-tree control treatment were planted in
monospecific alleys of 0.5 x 7.5 m in five replicates of 0.9 ha random block plots.
Three censuses were carried out at 6-month intervals to evaluate the relative growth
rates of tree species to 18 months. I. edulis and L. leucocephala were the tallest
species after 18 months (443 cm, P<0.05) and I. edulis had a increased root collar
diameter (6.9 cm, P<0.05) and diameter at breast height (3.6 cm, P<0.05) when
compared to other species. 1m2 quadrats were used to evaluate the rate of grass
regrowth between treatments. Prior to pollarding the grass regrowth in I. edulis plots
was 50% less than that of any other treatment. Subsequently, all tree species were
pollarded to 2 m at 18 months. Post-pollard, all tree species treatments except C.
pentandra had a negative impact on the accumulation of grass biomass when

-153-
compared to the control treatment. I. edulis produced more pollard biomass than
other tree species (0.75 Mg ha-1year-1, P<0.05). Soil nitrogen, phosphorous and
organic matter measured at 18 months showed no effect of treatment. I. edulis, L.
leucocephala and E. berteroana are recommended for further investigation at the
present site.
1. Introduction
Livestock production currently contributes around 14.5% to total global greenhouse
gas emissions and is a leading driver of global land cover change (Hardin, 1968;
Steinfeld et al., 2006; McMichael et al., 2007; Lambin and Meyfroidt, 2011; Gerber,
2013; Grossi et al., 2019). Global meat production is projected increase 15% by
2027 to meet growing demand (OECD‐ FAO, 2018). Rising temperatures are
predicted to decrease the forage quality of grasses and this may lead to increased
methane outputs, thereby generating a positive feedback effect that could
exacerbate climatic change (Lee et al., 2017). The use of livestock production
systems that incorporate trees or shrubs, also known as silvopastoral systems, are
associated with increased environmental benefits than conventional livestock
production (Nepstad et al., 2006; Schiermeier, 2019). The presence of trees in
pastures helps maintain soil functioning, hosts biodiversity and is increasingly being
recognized as providing an important function in resilience to climate change in rural
farming communities (Reed, 2013; Altieri et al., 2015). Silvopastoral systems have
been used to implement landscape scale conservation interventions, to increase
livestock or timber productivity per hectare of land and have been found to sequester

-154-
more carbon than the traditional grass monoculture approach to livestock production
(Pagiola et al., 2004; Haile et al., 2010; Tonucci et al., 2011; Broom et al., 2013).
In some regions of the Amazon, the current paradigm of livestock production is of a
cattle stocking density of less than 1 ha-1 ( Valentim and Andrade, 2009; Bowman et
al., 2012; Reis, 2017). Cattle contribute to unequal nutrient deposits and pastures
can reach an advanced stage of degradation in ten years (Boddey et al., 2004;
Martínez and Zinck, 2004). Cattle pastures are viewed as the antithesis of the
ecosystem services provided by intact forests, yet these degraded pastures
represent an excellent opportunity for rehabilitation using silvopastoral systems.
There are a paucity of studies that attempt to explore potential native forages that
would be suitable for silvopastoral systems in the Amazon biome (Hohnwald et al.,
2016). To enhance the success of a plantation requires careful selection of the tree
species to match the environmental conditions of a potential site (Butterfield, 1995).
In many cases, the forage potential of locally adapted species is unknown and there
is a need to identify native trees and shrubs for forage production (Murgueitio et al.,
2011). The pasture environment in the Amazon presents a unique challenge as the
environmental conditions include poor, compacted soils with low pH, high levels of
exchangeable aluminum, high levels of irradiation and predominantly monocultures
of exotic African grasses such as Brachiaria sp. that may have deleterious effects on
adjacent vegetation ( Moraes et al., 1996; Loker et al., 1997; Nesper et al., 2015).
Authors have called for the exploration of the agroforestry potential of native species
as they may be well adapted to their environment and may be able to increase the

-155-
provision of ecosystem services (Butterfield, 1995; Fisher, 1995; Murgueitio et al.,
2011). The present study seeks to test the potential of candidate species for
silvopastoral systems in the SW Amazon via a 21-month field experiment in a
Brachiaria brizantha cv. Marandú pasture. This experiment compares the
establishment success and growth of five tree species and their impact on the
production of pasture grass and soil chemical parameters nitrogen (N), phosphorous
(P) and organic matter (OM). It is hypothesized that:
(a) I. edulis will demonstrate the fastest relative growth rate
(b) L. leucocephala will demonstrate the highest mortality rate
(c) The presence of trees will positively effect soil N and OM and negatively effect
soil P.
(d) I. edulis will produce the greatest quantity of biomass when harvested by a
pollard > 2 m.
(e) The rate of grass growth will be negatively influenced by the presence of all
tree species.
2. Methods
A detailed description of experimental design and planting arrangement for this
experiment can be found in Chapter II and briefly described here. The experiment
used a randomised block design with five replicates. There were six treatments in
the experiment, with each treatment involving a pasture dominated by Brachiaria
brizantha cv. Marandú. The treatments were (1) Leucaena leucocephala, (2) Inga
edulis, (3) Ceiba pentandra, (4) Senegalia loretensis, (5) Erythrina berteroana, (6)
Control of Brachiaria brizantha cv. Marandú with no tree association. These

-156-
treatments were randomly allocated between thirty plots of 30 x 30 m (0.09 ha). Tree
treatments were planted in three rows of 0.5 x 7.5 m. There were a total of 180 trees
per plot.
2.1. Censuses of Tree Growth
All trees were individually labelled. The first census of plant growth was carried out
immediately after the trees had been planted in March 2017, and represents the
initial conditions (C1). The census was subsequently repeated in August 2017 (C2),
March 2018 (C3) and August 2018 (C4), for a total of three, six-month intervals of
plant growth between the censuses (Table 5). A 2.5 m strip around each plot was
excluded from all censuses to reduce edge effects. All trees were monitored during
the first census in March 2017 and the last census in August 2018. During the
census of August 2017 and March 2018, only the 63 trees in the centre of each plot
were monitored (21 from each of the three rows). Fewer trees were monitored during
the August 2017 and March 2018 census due to a lack of resources.

-157-
Table 5. Plant growth census dates.
Census Year Month
Months post-
planting
Interval between census
C1 2017 March 0 - C2 2017 August 6 6 C3 2018 March 12 6 C4 2018 August 18 6
Measurements were taken with a digital Vernier calliper and a 5 m measuring tape.
The following measurements were taken: height from ground to apical meristem of
tallest branch (height, measured in cm), root collar diameter (RCD, measured in mm)
and diameter at 130 cm from the ground (DBH, measured in mm). The tree was
reported as ‘alive’ (i.e. healthy), ‘regrowth’ or ‘dying’. The regrowth category was
employed when the majority (50-100% of the branch and stem material) of the tree
had died and then had begun to regrow. Trees in the regrowth or dying category
were excluded from subsequent analysis. The cause of tree mortality was reported if
known. A replicate of I. edulis was excluded from the analysis due to heavy and
persistent flooding of the plot at the time of experimental establishment. A replicate
of C. pentandra was also excluded from the analysis due to frequent and complete
defoliation by leafcutter ants.
Statistical Analysis
An annual mean relative growth rate was calculated for each census interval to
facilitate comparison between species according to South (1995):
(Equation 2)

-158-
MRGR = (ln(X)2 – ln(X)1)/(t2 – t1)
where X1 and X2 is the size at the beginning t1 and end t2 of the census interval.
Annual mortality (λ) was calculated for each census according to equation 7 in Shiel
et al. (1995):
(Equation 3)
λ = (logeN0 – logeN1)/t
Where N0 is the population count at the beginning of a census and N1 is the
population count at the end of the census and t is the measurement interval.
Statistical analysis of growth rates of trees excluded trees that had been placed into
the regrowth or dying categories. All analyses were carried out using the program R
(3.2.2). Shapiro-Wilk tests were used to test the normality of the distribution of all
continuous measurement variables (height, RCD, and DBH). For each census, a
one-way ANOVA was used to test for differences between each of the
measurements and differences between species (categorical). A Tukey’s Honest
Significant Differences test was used post-hoc to compare the differences between
mean measurements for all species in each census.
2.2. Soil Chemical Analysis
The three tree species that had exhibited the lowest mortality to the time of January
2019 were I. edulis, E. berteroana and L. leucocephala. These species were
therefore chosen as the most likely to have had an effect on the soil chemistry. Soil
chemical analysis was carried out on a total of 16 plots composed of four plots of the

-159-
control treatment of no trees and four plots of each tree treatment. Total nitrogen (N),
phosphorous (P) and organic matter (OM) samples were taken from 0-10 cm of the
soil. Total N, P and soil organic matter were available at the laboratory and due to
limited resources these were chosen as priorities, as the nitrogen fixing species may
increase N, P was expected to decrease under plantation conditions and OM to
increase as a result of input via litterfall. However, due to unforeseen conditions the
samples were taken sooner than originally envisaged (samples were taken in
January 2019, at 24 months post establishment of the experiment, but had originally
been planned for 31 months). For the 20 samples that were taken from within the
tree line the sample was taken from between adjacent two trees (0.25 m from the
base of the two neighboring trees). The samples were mixed and three subsamples
were taken. The same procedure was used between the rows of trees where
samples were taken at a distance of 3.75 m from the tree line. Samples were sent for
analysis in the Laboratorio de Analysis de Suelos de la facultad de Agronomia of the
La Molina University in Lima.
Statistical Analysis
The response variables of N, P and OM were tested for normality using Shapiro-Wilk
test. Differences in N, P, and OM (continuous) between treatments (categorical)
were tested by an ANOVA. To test for an effect of distance from the tree line on N, P
and OM, ANOVAs were run for each tree species with the distance from the tree line
(1 m or 3.75 m) as a categorical predictor variable.

-160-
2.3 Pollard Treatment
Trees of all species were pollarded at 18 months post planting, immediately following
the fourth census of plant growth. This was due to the observation that the grass
production near the base of some species of trees was being negatively effected,
presumably by the competition for light. In order to reduce competition all trees were
pollarded to 2 m. The pollarded biomass of approximately 20 trees per replicate was
collected and weighed. After each sampling event, three subsamples were taken
from a composite of plant material from each species. Subsamples were air dried for
48 hours then oven dried at 65 °C until a constant dry weight was reached. All
harvested plant material was removed from the experimental site. Differences in the
amount of pollarded biomass harvested (continuous variable) were tested between
treatments (categorical variable) using an ANOVA.
2.4. Effects of Trees on Rates of Pasture Regrowth
Quadrats (1 m2) were situated at random either at 1-2 m or 3.25-3.75 m from the
middle row (of three rows of trees within each parcel) within each plot and no less
than 7.5 m from the plot edge to reduce edge effects. The distances of 1-2 m or
3.25-3.75 m were chosen to test for the effect of the proximity of trees on the grass.
The quadrats were located at random in control parcels with no trees. Prior to the
pollarding of all trees to 2 m the rate of pasture regrowth was monitored for a 30-day
period in the first week of July 2018. In all plots, the grass in the eight quadrats was
cut to 10 cm before and after a 30-day period. The accumulated biomass was
weighed fresh, air-dried for 48 hours then oven dried at 65 °C until a constant dry
weight was reached.

-161-
To test an effect of pollarding on grass production, the clipped parcels were left to
regrow for six months until the first week of January 2019, when the accumulated
biomass was clipped to 10 cm and weighed. All biomass was weighed fresh, air-
dried for 48 hours then oven dried at 65 °C until a constant dry weight was reached.
Statistical Analysis
The two monitoring periods of grass biomass were treated separately. The dry
weight of the grass was inspected for outliers and normality using density and scatter
plots. The Shapiro-Wilk test suggested that the dry weights of the grass biomass
were normally distributed. One-way ANOVAs were used to test for the effect of the
categorical predictor variable treatment on the continuous response of grass dry
weight. Where significant effects were found, these were explored with the post-hoc
analysis Tukey’s Honest Significant Differences test. The effect on grass dry weight
of distance from the tree line was tested within treatments by following the same
procedure.
3. Results
After 18 months, I. edulis and L. leucocephala were the tallest species and both
reaching similar mean heights, for I. edulis this was 442.8 ± 103.5 (S.D.) and for L
leucocephala this was 443.1 ± 111 (S.D.) (Table 6). When compared to that of L.
leucocephala, I. edulis had a diameter at breast height (DBH) that was 44%
increased in size and a root collar diameter (RCD) that was 20% increased in size
(Table 6). I. edulis, L. leucocephala, and E. berteroana were the three of the five

-162-
species that consistently showed growth in all parameters over the 18 months of the
experiment (Table 6). All species demonstrated the highest growth rates in 6 months
following planting followed by a decline in the growth rate with the exception of E.
berteroana. E. berteroana showed the slowest relative growth rate of height (0.8)
and RCD (0.2) between the first and second census, and subsequently was the only
species to demonstrate increased growth in the interval between the second and
third census (Figure 34, Figure 35, Figure 36). Between the first and second census,
C. pentandra showed the greatest increase in RCD (324%) followed by A.
loretenesis (266%) (Figure 37). Between the third and fourth census these species
had the lowest increase of any species (1 and 2% respectively). In the same inter-
census interval C. pentandra and A. loretenesis displayed a decline in average
height as a result of dieback, but not a decline in average RCD or DBH (Figure 36,
Figure 37, Figure 38). Pictures of the plantation give an idea of the scale and layout
of the experiment (Figures 39-42).

-163-
Table 6. Height (cm), root collar diameter (RCD mm), and diameter at breast height (DBH mm) of five tree species in a Brachiaria brizantha cv. Marandú pasture, with census number (C), time after planting in months (t), mean average (Mean) standard deviation (S.D.), annual relative growth rate (RGR), n as number of individuals sampled and λ as annual mortality (Shiel et al., 1995). Means sharing the same superscript are not significantly different from each other within a census interval (Tukey's HSD, P<0.05)
Species C t Height (cm) RCD (mm) DBH (mm)
λ Mean S.D. RGR n Mean S.D. RGR n Mean S.D. RGR n
S. loretensis 1 1 73.3c 23.5 715 6.9c 2.2 715 0 2 6 197.9b 59 2 261 25.0a 7.6 2.6 262 9.9d 4 222 0.008 3 12 239.4c 86.4 0.4 189 31.5a 9.8 0.5 189 13.4d 5.2 0.6 170 0.089 4 18 113.6c 114 -1.5 371 32.0d 10.4 0 372 14.8d 7 0.2 137 0.601
C. pentandra 1 1 77.5b 16.4 709 10.9a 2.8 709 0 2 6 202.9b 30.4 1.9 252 46.1c 8 2.9 252 19.2c 6.7 247 0 3 12 227.7c 41.7 0.2 252 53.2c 13.3 0.3 252 22.5a 9.9 0.3 250 0 4 18 209.1b 67.1 -0.2 619 53.0c 13.8 0 619 23.7a 10 0.1 540 0.549
E. berteroana 1 1 69.0d 5.9 891 36.4d 14.8 872 0
2 6 101.1c 25 0.8 258 40.9d 16 0.2 258 10.0bd 1.9 36 0.274
3 12 202.4d 73 1.4 263 50.1c 16.7 0.4 262 19.7b 7.9 1.4 209 0.244
4 18 224.7d 87.6 0.2 509 59.2e 20.2 0.3 510 23.1b 9.5 0.3 413 0.48
I. edulis 1 1 79.3b 17.2 699 10.1b 2.7 700 0.025
2 6 194.3b 41.7 1.8 254 30.2b 8.3 2.2 254 11.2b 3.3 240 0.05
3 12 373.3b 68.2 1.3 230 57.7b 17.7 1.3 231 30.2c 9 2 230 0.148
4 18 442.8a 103.5 0.3 553 68.9b 22.2 0.4 553 36.2c 13.1 0.4 551 0.205
L. leucocephala 1 1 112.0a 25 887 8.4a 2.2 887 0
2 6 281.4a 64.3 1.8 429 24.5a 6.1 2.1 429 13.3a 5 424 0.005
3 12 409.7a 103 0.8 225 34.4a 11.1 0.7 226 20.7ab 6.8 0.9 222 0.079
4 18 443.1a 111 0.2 483 44.3a 18.4 0.5 507 23.1ab 8.7 0.2 503 0.281

-164-
Figure 34. Relative growth rate for changes in height of five tree species after planting at six (C2), twelve (C3) and eighteen (C4) month intervals post-planting for tree species Leucaena leucocephala, Senegalia loretensis, Inga edulis, Ceiba pentandra and Erythrina berteroana.
-2
-1
0
1
2
3
C2 C3 C4
Rela
tive
Gro
wth
Rate
of
Heig
ht
Census
S. loretensis C. pentandra
E. berteroana I. edulis

-165-
Figure 35. Relative growth rate for changes in root collar diameter (RCD) and of five tree species at between planting and six (C2), twelve (C3) and eighteen (C4) month intervals post-planting for tree species Leucaena leucocephala, Senegalia loretensis, Inga edulis, Ceiba pentandra and Erythrina berteroana
0
1
2
3
C2 C3 C4
Rela
tive
Gro
wth
Rate
of
RC
D
Census
L. leucocephala S. loretensis
C. pentandra E. berteroana
I. edulis

-166-
Figure 36. Height (cm) and standard deviation of five tree species growing in a Brachiaria brizantha cv. Marandú pasture as of census 1. March 2017, 2. July 2017, 3. March 2018 and 4. July 2018.

-167-
Figure 37. RCD (mm) and standard deviation of five tree species growing in a Brachiaria brizantha cv. Marandú pasture as of census 1. March 2017, 2. July 2017, 3. March 2018 and 4. July 2018.

-168-
Figure 38. DBH (mm) of five tree species growing in a Brachiaria brizantha cv. Marandú pasture as of census 2. July 2017, 3. March 2018 and 4. July 2018.

-169-
Figure 39. Rows of I. edulis

-170-
Figure 40. Row of Ceiba pentandra

-171-
Figure 41. Replicate of Senegalia loretensis

172
Figure 42. Leucaena leucocephala in March 2017 (above) and October 2017 (below)

173
Figure 43. Proportion of alive, dead and trees classified as regrowth of five tree species in a Brachiaria brizantha cv. Marandú pasture in the SE Amazon 18 months after planting at 0.5 x 7.5 m
After 18 months, I. edulis was the species with the least number of individuals placed
in the regrowth category (Figure 43). C. pentandra had the lowest rate of mortality.
There was considerable damage to all species by rodents and in many cases this
contributed to the mortality of the trees. The rodents stripped away the outer
cambium in a ring around the bottom of the plant, restricting the plant’s access to
water. Where this occurred, the species would attempt to regrow from the base.
C. pentandra reached a maximum average height of 228 cm after 12 months and
subsequently declined in height between 12 to 18 months as a result of dieback. By
0%
100%
E. berteroana I. edulis S. loretensis L.leucocephala
C. pentandra
% o
f c
ate
tgo
ry
Alive Regrowth Dead

174
the fourth census, it was observed that the majority of C. pentandra had dropped
their leaves and dieback had occurred from the apical meristem towards the ground.
When C. pentandra is healthy it exhibits a green trunk, yet by the fourth census, the
trees exhibited a brown or black dieback, followed by a vivid red then yellow
discolouration. This phenomenon was seen in all replicates of C. pentandra. There
were no visible above ground pests and no common leaf discolouration. There was
no local plant pathologist to consult. One replicate of C. pentandra had been
excluded at the start of the experiment due to repeated total defoliation by ants of the
genus Atta that resulted in the mortality of all trees in the plot. S. loretensis survived
well until the 12 month census, at which point the trees began to die. It was observed
that S. loretensis demonstrated increased height and survival when adjacent to I.
edulis.
When the levels of harvested tree biomass were compared, I. edulis produced 1.1
Mg ha-1 of biomass (Figure 44). This was significantly more than L. leucocephala
(0.7 Mg ha-1, P<0.05), and more than double the biomass production of E.
berteroana, C. pentandra or S. loretensis. It was observed at I. edulis produced
substantial foliar and woody biomass, whereas the majority of L. leucocephala
biomass was woody.

175
Figure 44. Mean dry weight biomass and standard deviation (S.D.) of pollarded biomass (over 2 m) of 18-month-old trees 0.5 x 7.5 m planting design (2667 trees ha-1) for individual trees, sample size represented in parenthesis. Means sharing the same superscript are not significantly different from each other (Tukey's HSD, P<0.05)
Prior to the pollarding event the rate of grass regrowth was measured for 30 days. I.
edulis was found to be the only treatment that negatively impacted the rate of grass
regrowth (Figure 45, P<0.01). Grass regrowth in the I. edulis treatment showed less
than half the growth of any other treatment. There was no effect of the distance from
the tree line on the rate of grass regrowth in any species. Six months following the
following the pollard the grass regrowth was reassessed. The control treatment (no
trees) and C. pentandra had the greatest accumulation of grass biomass of all the
tree treatments (Figure 46). No effect of distance from the tree line was found for any
of the treatments.

176
Figure 45. Mean total dry matter biomass accumulation (Mg ha-1 year-1) and standard deviation (S.D.) of 30-day regrowth of Brachiaria brizantha cv. Marandú collected from different treatments, sample size represented in parenthesis. Means sharing the same superscript are not significantly different from each other (Tukey's HSD, P<0.05)

177
Figure 46. Mean total dry matter biomass accumulation (Mg ha-1 year-1) and standard deviation (S.D.) of six months regrowth of Brachiaria brizantha cv. Marandú following the pollarding of associated tree species to 2 m, sample size represented in parenthesis. Means sharing the same superscript are not significantly different from each other (Tukey's HSD, P<0.05)
There was no effect of treatment on the soil chemical parameters nitrogen (N),
phosphorous (P) or organic matter (OM). There was no effect of the distance from
the tree line on N, P or OM for E. berteroana, I. edulis or L. leucocephala.
4. Discussion
Trees and grass compete for nutrients, light, and water. In this experiment, all
planted trees were competing with a pasture grass, Brachiaria brizantha cv. Marandu
for resources. Inga edulis was the species of tree that was expected to perform well
in the pasture environment and therefore it was hypothesised that I. edulis would

178
have the fastest relative growth rate. I. edulis produces a high proportion of foliar to
total biomass. The foliage of I. edulis has the highest leaf area and light interception
of the tree species in this experiment. In a B. brizantha pasture environment this
adaptation favours I. edulis, as the Brachiaria genus is averse to shade and
therefore the ability of a tree to generate enough shade to protect its roots from
competition with B. brizantha could be strategically important to the practice of
pasture reclamation. B. brizantha is highly invasive and can spread and rapidly
dominate landscapes. Multiple studies report that pasture grasses have a negative
effect on growth and survival of trees in pastures (Ferguson et al., 2003; Hooper et
al., 2002; Hooper et al., 2005). African grasses of the Brachiaria genus contain
phytotoxic substances that produce an allelopathic effect that inhibits the growth of
adjacent plants (Kobayashi and Kato-Noguchi, 2015; Sesto Cabral et al., 2011;
Pivello et al., 1999; Ganade and Brown, 2002).
It is possible that I. edulis was able to avoid the effect of the allelopathic chemicals in
the B. brizantha grass by shading out the proximate grasses and opening up a niche
within the grass. ‘Pioneer’ or ‘colonising’ tree species open a niche which in a
silvopastoral system may include relief from the grass, increased shade, increased
soil moisture and decreased soil compaction (Khurana and Singh, 2001; Garen et
al., 2011). Otsamo (1998) found that Anisoptera marginata grew fastest using the
nurse tree Paraserianthes falcataria on an Imperata cylindrica grassland site in
Indonesia. Similarly, the dense shade and ability to suppress weeds may give I.
edulis a unique advantage in the pasture environment and simultaneously open a
niche for additional biodiversity. Nichols and Carpenter (2006) found that Terminalia
amazonia showed improved growth when interplanted with I. edulis. A positive effect

179
of adjacent I.edulis was observed on the survival of S. loretensis. However, the
present study only contained three incidences where rows of S. loretensis and I.
edulis were adjacent, so it was not possible to quantify this interaction. However, this
observation highlights the importance of accounting for edge effects of adjacent
treatments in sampling design. A future study could attempt to thin an existing I.
edulis plantation to introduce potential tree or shrub species such as C. pentandra
and S. loretensis that may benefit from being interplanted with I. edulis.
Prior to the pollard, I. edulis was the most effective tree species at inhibiting the
growth of the grass. However, the pollard reduced this species’ total leaf area and
subsequently I. edulis was only as effective than any other tree species at controlling
the production of grass biomass. In this experiment, the trees were pollarded as a
strategy to optimise the production of available tree foliage as fodder for cattle.
However, the results suggest that an important ecosystem function of I. edulis in
pasture reclamation is the creation of a niche via the production of shade. Further
work is needed to understand the interaction between the optimal proportion of trees
that can be pollarded for potential forage resources, and the optimal proportion of
trees that should be left without the pollard for the production of shade and
subsequent control of the B. brizantha grass.
In the present study the aim was to maximise the production of biomass for cattle
production. By 12 months all species were over 2 m tall and therefore beyond the
reach of cattle. Pollarding is therefore a useful management strategy for managers
as removal of biomass above a certain point has the dual benefit that it can be used
to increase the availability of biomass by bringing forage into the reach of animals,

180
and it can also reduce the light competition by the trees on adjacent plants. Some
evidence exists to indicate which species can survive pruning and pollarding, but
there are also species that cannot (Lang et al., 2015; Muschler et al., 1993; Garcia et
al., 2001). Care must be taken when choosing the best management approach for
each species. Separating the foliar and the branch biomass at the 18-month pollard
harvest would have improved the experiment, as it is the foliar biomass that is of
interest. In future studies, it would be of interest to measure the survival and
regrowth of trees following pollarding. This was beyond the scope of the present
study.
The failure of C. pentandra and S. loretensis to survive in the experiment was due to
an unknown cause. It is speculated the cause may have been a plant pathogen or
the decline may have resulted from competition with the pasture grass. It was
hypothesised that Leucaena leucocephala would have the greatest mortality rate of
any of the tree species, but this was not the case. This hypothesis was chosen
because L. leucocephala was reported to have failed to establish in a previous trial in
the Amazon, believed to be due to a negative effect of acidic soils and high levels of
exchangeable aluminium on this species (Salazar et al., 1993). However, L.
leucocephala has been observed to occur as a weed in the urban areas of the
Peruvian jungle cities of Puerto Maldonado and Pucallpa (personal observation). L.
leucocephala is a widely used forage tree in silvopastoral systems in tropical regions.
In this experiment, the decision to include L. leucocephala was taken because it is a
species that has existing reports of growth and biomass production and therefore it
was considered appropriate to use as a reference species by which the native
species could be compared. Yet it is also recognised as an invasive species that can

181
quickly colonise a site to the exclusion of native species. Managers of natural
systems may choose different priorities for the landscapes they devise. This study
considers the business as usual scenario in the region where cattle are produced in
association with invasive grass. The objective was to increase the agricultural
productivity and flow of ecosystem services from an already modified agricultural
landscape. While native species may be preferable, at the outset of this study it was
not known which species would be able to survive in the pasture environment, or if
any species would be able to naturally regenerate within the pasture environment.
The potential for natural regeneration of these species within pastures is an area for
future research, and of particular interest where there is a risk of invasion by L.
leucocephala. The growth and biomass production of L. leucocephala has been
shown to be effected by provenance (Prasad et al., 2011). A limitation of the present
study is it only considers a single provenance of any of the species.
It was hypothesised that the trees would positively effect soil nitrogen (N) and soil
organic matter (OM) and negatively effect soil phosphorous (P). This experiment had
limited resources and therefore it is not possible to prove or disprove this hypothesis.
There was no effect detected either between treatments or at the two distances from
the tree line for any of the soil chemical parameters considered. While no treatment
effect was detected, it is possible that there was a treatment effect that was not
detected in the present sampling regime, or that it is too soon to see measurable
effect. Therefore, the results should be treated with caution. It was not possible in
this study to conduct soil chemical analysis prior to the establishment of the
experiment. However, where resources permit this is a useful exercise. This
experiment was designed to contain five replications of each treatment with the

182
intention of minimising any differences across the site, such as soil chemistry, soil
moisture or soil compaction. Szott and Palm (1996) found that soil phosphorous
decreased following establishment of I. edulis due to the uptake of phosphorous in
the living biomass. They theorised that the phosphorous would eventually return to
the soil via leaf litter. Fisher (1995) found I. edulis increased soil nitrogen and
extractable phosphorous after 7 years. However, if the objective is production for
forage then the continuous removal of foliar biomass for fodder may result in the
exportation of nutrients held within the biomass that is removed. This could result in
a net loss of nutrients over time.
It was hypothesised that I. edulis would produce the greatest quantity of biomass
when harvested by a pollard that removed all biomass over 2 m height from the
ground. The findings of this study support this hypothesis. While this is a seminal
result for this management approach, biomass production is closely related to growth
parameters. It is possible to contextualise these findings by reviewing the results of
other studies that have experimentally grown I. edulis. I. edulis individuals in this
study were taller and thinner at breast height (DBH) than in Alegre’s (2005)
plantation in Yurimaguas, Peru. Alegre (2005) planted I. edulis at 4,444 trees ha-1 in
a 1.5 m by 1.5 m arrangement, as both a monoculture and also grown in association
with Centrosema macrocarpum. After the first year, the trees that had been grown as
a monoculture treatment reached a height of 2.5 m and a DBH of 4.4 cm. When I.
edulis was grown with C. macrocarpum, the height in the first year was 3.0 m and
the DBH 3.8 cm. The present study is not directly comparable with other studies due
to differences in planting arrangements that could be responsible for differences in
the tree morphology. The growth of I. edulis in the present study is in the same range

183
of that reported by Kanmegne (2000) in Southern Cameroon on a Ferric Acrisol soil
type. Kanmegne (2000) planted I. edulis at 3,333 trees ha-1, in 3 x 1 m rows. I. edulis
grew to 3.7 m in 12 months and by 20 months had reached a height of 7.7 m
(Kanmegne, 2000). Tilki and Fisher (1998) planted I. edulis at 1,111 trees ha-1 in a 3
x 3 m arrangement in three sites in Sarapiqui, Costa Rica. After three years, I. edulis
displayed an average DBH of 10.5 cm. Shimamoto et al. (2014) measured the
growth of naturally regenerating I. edulis in the Parana State in Southern Brazil. By
the age of 18 – 41 years they report that the average DBH of I. edulis was 24.4 cm
(± 11 SD) and an average height of 16.2 m (± 4 SD). These varied results show that
I. edulis is a species with fast growth in a number of different plantation trials.
The limitations of this experiment include the short time frame, a monospecific
planting arrangement of the five species may have favoured or prejudiced certain
species, and that only pollarded tree biomass was reported when a measure of total
tree biomass would have enabled a comparison of whether tree and grass
arrangements are more productive than grass only arrangements. It may also be of
interest to experiment with the addition of fertiliser, changing the associated grass or
tree species, and attempting the experiment in a location with a different set of
edaphic and climatic conditions. Future studies may also consider the reproduction
and establishment success for all species from seed in the invasive and allelopathic
Brachiaria brizantha cv. Marandú or how establishment is effected by exposure to
browsing. Further work would be required to understand the additional benefits of
silvopastoral systems, in particular, marketable benefits that could be used to
promote their uptake, such as the potential for carbon sequestration.

184
In the present study the aim was to maximise the production of biomass for cattle
production. By 12 months all species were over 2 m tall and therefore beyond the
reach of cattle. Pollarding is therefore a useful management strategy for managers
as removal of biomass above a certain point has the dual benefit that it can be used
to increase the availability of biomass by bringing forage into the reach of animals,
and it can also reduce the light competition by the trees on adjacent plants. Some
evidence exists to indicate which species can survive pruning and pollarding, but
there are also species that cannot (Lang et al., 2015; Muschler et al., 1993; Garcia et
al., 2001). Care must be taken when choosing the best management approach for
each species. Separating the foliar and the branch biomass at the 18-month pollard
harvest would have improved the experiment, as it was the foliar biomass that was of
interest. In future studies, it would be of interest to measure the survival and
regrowth of trees following pollarding. This was beyond the scope of the present
study.
Inga edulis, Leucaena leucocephala and Erythrina berteroana were the three
species that may be recommended as species for further assessment of their
silvopastoral potential at the study site. C. pentandra and S. loretensis may be
suitable candidates for silvopastoral systems but their relatively higher levels of
mortality shown in this study suggest they are species that are not well adapted to
the conditions of this study site. If the pasture grass inhibits the development of tree
species then planting dense lines of I. edulis and subsequent thinning may create a
niche in which other trees and shrubs can be planted. The limitations of this
experiment include the short time frame, a monospecific planting arrangement of the
five species may have favoured or prejudiced certain species, and that only

185
pollarded tree biomass was reported when a measure of total tree biomass would
have enabled a comparison of whether tree and grass arrangements are more
productive than grass only arrangements. It may also be of interest to experiment by
changes to the associated grass or tree species, and attempting the experiment in a
location with a different set of edaphic and climatic conditions. Future studies may
also consider the reproduction and establishment success for all species from seed
in the invasive and allelopathic Brachiaria brizantha cv. Marandú or how
establishment is effected by exposure to browsing. Further work would be required
to understand the additional benefits of silvopastoral systems, in particular,
marketable benefits that could be used to promote their uptake, such as the potential
for carbon sequestration.
References
Altieri, M.A., Nicholls, C.I., Henao, A., Lana, M.A., 2015. Agroecology and the design
of climate change-resilient farming systems. Agronomy for Sustainable Development
869–890. https://doi.org/10.1007/s13593-015-0285-2
Bowman, M.S., Soares-Filho, B.S., Merry, F.D., Nepstad, D.C., Rodrigues, H.,
Almeida, O.T., 2012. Persistence of cattle ranching in the Brazilian Amazon: A
spatial analysis of the rationale for beef production. Land Use Policy 29, 558–
568. https://doi.org/10.1016/j.landusepol.2011.09.009
Butterfield, R.P., 1995. Promoting biodiversity: advances in evaluating native species
for reforestation. Forest Ecology and Management 75, 111–121.

186
Fisher, R.F., 1995. Amelioration of degraded rain forest soils by plantations of native
trees. Soil Science Society of America Journal 59, 544–
549. https://doi.org/10.2136/sssaj1995.03615995005900020039x
Ganade, G., Brown, V.K., 2002. Succession in old pastures of central Amazonia:
Role of soil fertility and plant litter. Ecology 83, 743–
754. https://doi.org/10.1890/0012-9658(2002)083[0743:SIOPOC]2.0.CO;2
Garcia, H., Nygren, P., Desfontaines, L., 2001. Dynamics of nonstructural
carbohydrates and biomass yield in a fodder legume tree at different harvest
intensities. Tree Physiology 21, 523–531. https://doi.org/10.1093/treephys/21.8.523
Garen, E.J., Saltonstall, K., Ashton, M.S., Slusser, J.L., Mathias, S., Hall, J.S., 2011.
The tree planting and protecting culture of cattle ranchers and small-scale
agriculturalists in rural Panama: Opportunities for reforestation and land restoration.
Forest Ecology and Management 261, 1684–
1695. https://doi.org/10.1016/j.foreco.2010.10.011
Gawali, A., Puri, S., Swamy, S.L., 2015. Evaluation Growth and Yield of Wheat
Varieties under Ceiba pentandra (L) Based Agrisilviculture System. Universal
Journal of Agricultural Research 3, 173–181.
Gerber, P.J., 2013. Tackling climate change through livestock : a global assessment
of emissions and mitigation opportunities. Food and Agriculture Organization of the

187
United Nations.
Grossi, G., Goglio, P., Vitali, A., Williams, A.G., 2019. Livestock and climate change:
impact of livestock on climate and mitigation strategies. Animal Frontiers 9, 69–
76. https://doi.org/10.1093/af/vfy034
Haile, S.G., Nair, V.D., Nair, P.K.R., 2010. Contribution of trees to carbon storage in
soils of silvopastoral systems in Florida, USA. Global Change Biology 16, 427–
438. https://doi.org/10.1111/j.1365-2486.2009.01981.x
Hardin, G., 1968. The tragedy of the commons. Science (New York, N.Y.) 162,
1243–8. https://doi.org/10.1126/science.162.3859.1243
Hohnwald, S., Trautwein, J., Camarão, A.P., Wollny, C.B., 2016. Relative palatability
and growth performance of capoeira species as supplementary forages in the NE-
Amazon. Agriculture, Ecosystems & Environment 218, 107–
115. https://doi.org/10.1016/J.AGEE.2015.11.015
Khurana, E., Singh, J.S., 2001. Ecology of seed and seedling growth for
conservation and restoration of tropical dry forest : a review. Environmental
Conservation 28, 39–52. https://doi.org/10.1017/S0376892901000042
Kobayashi, A., Kato-Noguchi, H., 2015. The seasonal variations of allelopathic
activity and allelopathic substances in Brachiaria brizantha. Botanical studies 56,
25. https://doi.org/10.1186/s40529-015-0105-7

188
Lambin, E.F., Meyfroidt, P., 2011. Global land use change, economic globalization,
and the looming land scarcity. Proceedings of the National Academy of Sciences of
the United States of America 108, 3465–
72. https://doi.org/10.1073/pnas.1100480108
Lang, P., Jeschke, M., Wommelsdorf, T., Backes, T., Lv, C., Zhang, X., Thomas,
F.M., 2015. Wood harvest by pollarding exerts long-term effects on Populus
euphratica stands in riparian forests at the Tarim River, NW China. Forest Ecology
and Management 353, 87–96. https://doi.org/10.1016/J.FORECO.2015.05.011
Lee, M.A., Davis, A.P., Mizeck G.G.C., Manning, P., 2017. Forage quality declines
with rising temperatures, with implications for livestock production and methane
emissions. Biogeosciences 14, 1403–1417. https://doi.org/doi:10.5194/bg-14-1403-
2017
Loker, W., Vera, R., Reátegui, K., 1997. Pasture performance and sustainability in
the Peruvian Amazon: Results of long-term on-farm research. Agricultural Systems
55, 385–408. https://doi.org/10.1016/S0308-521X(96)00102-3
Martínez, L.J., Zinck, J.A., 2004. Temporal variation of soil compaction and
deterioration of soil quality in pasture areas of Colombian Amazonia. Soil and Tillage
Research 75, 3–17. https://doi.org/10.1016/j.still.2002.12.001
McMichael, A.J., Powles, J.W., Butler, C.D., Uauy, R., 2007. Food, livestock

189
production, energy, climate change, and health. The Lancet 370, 1253–
1263. https://doi.org/10.1016/S0140-6736(07)61256-2
Moraes, J.F.L. de, Volkoff, B., Cerri, C.C., Bernoux, M., 1996. Soil properties under
Amazon forest and changes due to pasture installation in Rondônia, Brazil.
Geoderma 70, 63–81.
Murgueitio, E., Calle, Z., Uribe, F., Calle, A., Solorio, B., 2011. Native trees and
shrubs for the productive rehabilitation of tropical cattle ranching lands. Forest
Ecology and Management 261, 1654–
1663. https://doi.org/10.1016/j.foreco.2010.09.027
Muschler, R.G., Nair, P.K.R., Meléndez, L., 1993. Crown development and biomass
production of pollarded Erythrina berteroana, E. fusca and Gliricidia sepium in the
humid tropical lowlands of Costa Rica. Agroforestry Systems 24, 123–
143. https://doi.org/10.1007/BF00706887
Nepstad, D.C., Stickler, C.M., Almeida, O.T., 2006. Globalization of the Amazon soy
and beef industries: Opportunities for conservation. Conservation
Biology. https://doi.org/10.1111/j.1523-1739.2006.00510.x
Nesper, M., Bünemann, E.K., Fonte, S.J., Rao, I.M., Velásquez, J.E., Ramirez, B.,
Hegglin, D., Frossard, E., Oberson, A., 2015. Pasture degradation decreases
organic P content of tropical soils due to soil structural decline. Geoderma 257-258,

190
123–133. https://doi.org/10.1016/j.geoderma.2014.10.010
Majid, N., Paudyal, B., Shebli, Z., 1998. Survival and early growth of Acacia
mangium, Ceiba pentandra and Casuarina equisetifolia on sandy tin tailings.
Pertanika Journal of Tropical Agricultural Science 21, 59–65.
OECD-FAO (Ed.), 2018. Chapter 6. Meat Market situation, in: Agricultural Outlook
2018-2027. p. 150. https://doi.org/10.1787/agr-outl-data-en
Parks, N., 2007. Livestock’s long shadow. Frontiers in Ecology and the
Environment. https://doi.org/10.1890/1540-9295(2007)5[4:D]2.0.CO;2
Pivello, V.R., Shida, C.N., Meirelles, S.T., 1999. Alien grasses in Brazilian savannas:
a threat to the biodiversity. Biodiversity and Conservation 8, 1281–
1294. https://doi.org/10.1023/A:1008933305857
Prasad, J.V.N.S., Korwar, G.R., Rao, K.V., Mandal, U.K., G.R.Rao, Srinivas, I.,
Venkateswarlu, B., Rao, S.N., Kulkarni, H.D., 2011. Optimum stand density of
Leucaena leucocephala for wood production in Andhra Pradesh, Southern India.
Biomass and Bioenergy 35, 227–
235. https://doi.org/10.1016/J.BIOMBIOE.2010.08.012
Reis, E., 2017. Opportunities and challenges to the sustainable development of
cattle raising in Brazil, 1970–2005. Economia 18, 18–
39. https://doi.org/10.1016/J.ECON.2016.07.004

191
Salazar, A., Szott, L.T., Palm, C.A., 1993. Crop-tree interactions in alley cropping
systems on alluvial soils of the Upper Amazon Basin. Agroforestry Systems 22, 67–
82. https://doi.org/10.1007/BF00707472
Schiermeier, Q., 2019. Eat less meat: UN climate-change report calls for change to
human diet. Nature 2019 572:7769.
Sesto Cabral, M.E., Schabes, F.I., Sigstad, E.E., 2011. A calorimetric study of the
interaction between Brachiaria platyphylla and soil microbial activity. Thermochimica
Acta 526, 157–162. https://doi.org/10.1016/J.TCA.2011.09.015
Shimamoto, C.Y., Botosso, P.C., Marques, M.C.M., 2014. How much carbon is
sequestered during the restoration of tropical forests? Estimates from tree species in
the Brazilian Atlantic forest. Forest Ecology and Management 329, 1–
9. https://doi.org/10.1016/J.FORECO.2014.06.002
Sheil, D., Burslem, D.F. and Alder, D., 1995. The interpretation and misinterpretation
of mortality rate measures. Journal of Ecology, pp.331-333.
South, D.B., 1995. Relative Growth Rates: a Critique. South African Forestry Journal
173, 43–48. https://doi.org/10.1080/00382167.1995.9629690
Szott, L.T., Palm, C.A., 1996. Nutrient stocks in managed and natural humid tropical
fallows. Plant and Soil 186, 293–309. https://doi.org/10.1007/BF02415525

192
Tilki, F., Fisher, R.F., 1998. Tropical leguminous species for acid soils: studies on
plant form and growth in Costa Rica. Forest Ecology and Management 108, 175–
192. https://doi.org/10.1016/S0378-1127(98)00225-4
Tonucci, R.G., Nair, P.K.R., Nair, V.D., Garcia, R., Bernardino, F.S., 2011. Soil
Carbon Storage in Silvopasture and Related Land-Use Systems in the Brazilian
Cerrado. Journal of Environment Quality 40,
833. https://doi.org/10.2134/jeq2010.0162
Valentim, J.F., Andrade, C.M.S. de, 2009. Tendências e perspectivas da pecuária
bovina na amazônia brasileira. Amazônia: Ciência & Desenvolvimento 4, 7–30.

193
CHAPTER IV
ALLOMETRY AND PRODUCTION OF THREE
MONOSPECIFIC SILVOPASTORAL SYSTEMS IN AMAZONIA
Abstract
The integration of trees could increase biomass production and carbon sequestration
in the cattle pastures of the Amazon rainforest. This paper evaluates the biomass
production at 24 months of three silvopastoral systems following a pollarding event
at 18 months. This study measured the root collar diameter, stump diameter,
diameter at breast height, height, crown volume, and crown diameter to predict the
biomass of total, trunk, branch and foliage components. Species and component
specific allometric equations were developed by comparison of linear, quadratic and
log-linear models using Furnival’s Index. Log-linear models were found to be
superior to linear or quadratic models for the prediction of biomass components in all
species. The results showed that a single parameter model of RCD was the best
predictor of all biomass components of Inga edulis, whereas for Erythrina berteroana
and Leucaena leucocephala a multiple model of RCD and height improved the
predictive measurements of total and trunk biomass. There was reliable prediction of
total biomass for I. edulis (R2= 0.70), E. berteroana (R2= 0.77) and L. leucocephala
(R2= 0.86). After 24 months, I. edulis had accumulated the most biomass (7.2 Mg ha-
1) when compared to E. berteroana (4.4 Mg ha-1) or L. leucocephala (4.3 Mg ha-1).
The predicted carbon sequestration of these species was greatest for I. edulis (3.4

194
Mg ha-1) when compared to E. berteroana (2.1 Mg ha-1) or L. leucocephala (2.0 Mg
ha-1). Silvopastoral systems have the potential to increase the carbon sequestration
of pastures in the Amazon rainforest.
1. Introduction
Livestock production is a major driver of global land cover change (Steinfeld et al.,
2006). The pasturelands once occupied by Amazon Rainforest are typically
unsustainable, and pasture degradation may occur within 10 years (Martínez and
Zinck, 2004). There is a need to develop a new paradigm that can reverse this trend
and enhance the provision of ecosystem services from these agricultural landscapes
(Serrão et al., 1996). It has been shown that silvopastoral systems, the inclusion of
trees and shrubs into pastures, can result in increased livestock production and also
be instrumental in landscape level sustainability (Mauricio et al., 2019). However,
during the establishment phase of silvopastoral systems, cattle may be detrimental
to the establishment of plants and for that reason may need to be excluded from
young plantations (Smit et al., 2015). Therefore, farmers that wish to transform their
land to silvopastoral systems may experience the cost of a temporary loss of land.
Farmers could be compensated for this short-term loss of land and receive payments
for the conversion of land to silvopastoral systems under schemes known as
payments for ecosystem services (Pagiola et al., 2007). Such schemes rely on an
understanding of the costs and benefits of providing an ecosystem service (Claassen
et al., 2008; Tzilivakis et al., 2019).

195
Sharrow and Ismail (2004) hypothesised that as pastures store the majority of their
carbon (C) below ground in soil organic matter whereas forests store the majority of
their C above ground that silvopasture should accumulate more C via increased
biomass production and considerable below and above ground C sequestration.
They found that after 11 years, silvopasture accumulated more C than forest (0.74
Mg ha-1 year-1) or pasture (0.52 ha-1 year-1). C sequestration per hectare is a
measurable ecosystem service for which several markets exist, and payments to
farmers for sequestered C may incentivise them to integrate trees into their pastures
(Corbera et al., 2009; Hissa et al., 2019; Blum and Lövbrand, 2019).
The International Panel on Climate Change produced guidelines in 2003 for the
estimation of C stocks via allometric equations (Penman et al., 2003). These were
intended for the measurement of C for trade mechanisms that arose from the Kyoto
protocol (for example, Reduced Emissions from Deforestation and Degradation).
Allometric equations seek to determine the potential of one measured variable to
predict another variable. A critical application of this theory is that of quantifying the
volume of tree biomass to predict C stock. For example, the diameter of a tree can
be used to predict the total biomass of that tree. A benefit of allometric equations is
that if a strong relationship can be quantified, usually via the destructive
measurement of a subsample of trees, the equation can be used to quantify the
biomass of other trees that do not have to be destroyed. Allometric relationships are
governed by site-specific conditions and management impacts. The coarse
resolution studies that attempt large scale estimates of C stocks can result in high
levels of uncertainty (Chave et al., 2014). Site and species-specific models
demonstrate improved accuracy but are of limited transferability. For example,

196
Huang et al. (2000) found that, when applied to alternative ecoregions from the one
in which they were developed, height-diameter models of white spruce Picea
glauca (Moench) Voss resulted in maximum overestimations of 29% and
underestimations of 22%.
The relationship between variables of tree growth can take many forms. The most
common models in the literature are the power model and the polynomial model
(Picard et al., 2018). When a power relationship is suspected a widely used practice
in the literature is the use of the natural log transformation on the predictor and
response variable data to achieve a linear fit (Kitteredge, 1944; Mascaro et al.,
2014). This relationship can then be described using an ordinary least squares linear
regression and then back-transformed to the arithmetic scale using a conversion
factor (e.g. Marklund, 1987).
While the power function occurs frequently in the field of tree allometry, the
predominance of authors following this approach, with little regard for alternative
methods, has been criticised as failing to detect the most appropriate model for the
distribution of the data (Packard, 2017a; Packard, 2017b). Equations can include
multiple variables but care should be taken that separate variables are not highly
correlated, as this can result in multicollinearity which is known to have a
confounding effect on the model (Sileshi, 2014). Similarly, high order polynomials
begin to result in over-fitting of the model that can result in biologically implausible
models. Sileshi (2014) consider that for each additional predictor the sample size
should be doubled, and noted that ordinary least squares regression can be useful
when few non-collinear predictors are used that have a well-understood relationship

197
to the response variable. Ultimately, the error distribution will determine which
method performs best and therefore a model cannot be selected a priori (Packard,
2014; Xiao et al., 2011; Sileshi, 2014; Mascaro et al., 2014).
Xiao et al. (2011) demonstrated that ordinary least squares linear regression was a
suitable approach for characterising data with multiplicative, heteroscedastic,
lognormal error, whereas data with additive, homoscedastic, normal error was better
suited to non-linear regression approaches. To determine the appropriate error
structure, biological reasoning or likelihood analysis by comparing the Akaike
Information Criteria (AIC) or the coefficient of determination r-squared (R2) is
commonly used (Akaike, 1974; Xiao et al., 2011). However, AIC and R2 are not
appropriate to compare between models where different transformations have been
applied to the data. In such instances, interpretation of the model residuals can be
used in to determine the most appropriate modelling approach. The Furnival’s Index
(1961) is a method that can be used to compare a variety of modelling approaches
to the same dataset, where some models use transformed data and others do not.
The theoretical basis upon which allometric equations are built has also been used
by managers of natural ecosystems who seek to predict the amount of browse
available to herbivores or the amount of browse utilized by herbivores (MacCracken
and Ballenberghe, 1993). A small body of work has investigated the experimental
removal of biomass and subsequent regrowth with the intention of maximising
production of a specific element of biomass such as coppiced sticks or livestock
forage (Harrington and Fownes, 1993; Oppong et al., 2002). There is increasing
demand for landscape level management that optimises the production of multiple

198
ecosystem service benefits, such as forage production, the formation and
maintenance of soil and the sequestration of C (MEA, 2005).
This study seeks to quantify the amount of aboveground total and foliar biomass
produced and C sequestered in the first two years of establishment of three potential
silvopastoral tree species, Inga edulis, Erythrina berteroana and Leucaena
leucocephala. It is assumed that the production of biomass by these tree species
results in additional C sequestration in the pasture when compared to a pasture with
no trees, as previous chapters have shown a negligible impact of these trees on
grass production when managed by pollarding (See Chapter III). The trees of all
three species were harvested 24 months post planting and had previously been
pollarded to a height of 2 m at 18 months. Existing allometric models for E.
berteroana, I. edulis and L. leucocephala were sought on the Internet database
Globallometree (Globallometree.org accessed July 2019). 478 existing equations
were found for L. leucocephala, but no allometric equations existed for I. edulis or E.
berteroana. As allometric equations are specific to the site in which species are
grown and the management regime, it was considered necessary to develop specific
allometric equations for these three species that considered the impact of a
management regime of pollarding at 2 m after 18 months. It was hypothesised that:
(a) When three models were compared, log-linear, linear and quadratic, the log-
linear model would display the best fit.
(b) The greatest single predictor of total biomass will be stem diameter
measurements.
(c) Multiple predictor models will improve performance over single predictor models
for the prediction of biomass components.

199
(d) Measurements of the crown will be the best predictor of total foliar biomass.
2. Methods
2.1 Measurements of Tree Biomass
Three tree species were chosen for the development of allometric models, Inga
edulis, Erythrina berteroana and Leucaena leucocephala. These species had been
grown in a Brachiaria brizantha cv. Marandú pasture in a planting arrangement of 0.5
x 7.5 m for a density of 2,667 trees ha-1 (see Chapter II for further information on the
experimental design). Five replicates of each species were available at the beginning
of the study. This experiment chose three replicates at random from each species.
Each replicate had 180 trees, for a total of 540 trees per species. All trees of all three
species had been pollarded to 2 m at 18 months. The present study occurs after 24
months of growth, and 6 months after the pollard event. Approximately 40% of the
trees in the three replicates were harvested for the development of allometric
equations. A further 30% of trees were harvested to develop estimates of C
sequestration.
In allometric studies, there is a requirement that samples should represent a range of
at least one measurement to develop models that are representative of a range of
size classes. This is because the equations can only describe the relationship
between the variables they are given. In other words, a model that was developed
based on trees of up to 2 m height is likely to display considerable error if applied to
a tree of 20 m height. Therefore, where possible, the sample should include
representatives including the maximum and minimum sizes in a range to increase

200
the accuracy of scale-dependent equations (LaBarbera, 1989). Height, age and
diameter at breast height (DBH, 1.3 m) are commonly used in selecting size classes.
However, this study considered a plantation of trees that were of the same age and
had been pollarded to 2 m. Some trees were shorter than the commonly used
parameter of DBH. Therefore, RCD was used to select size classes. For each tree
species, the RCD data were divided into 10 size classes and nine trees were
randomly drawn from each size class, to represent three trees per replicate in each
size class.
Figure 47. Diagram representing the location of measurements on tree, where CD is crown diameter, CDep is crown depth, CH is crown height, H is height, RCD is root collar diameter, SD is stump diameter and DBH is diameter at breast height.
.
Height (cm) was measured from the ground to the top of the highest meristem
(Figure 47). Crown height (cm) was measured from the ground to where foliage
volume assumes 5% of the total. Callipers were used to measure, at the widest
point, the RCD (mm), diameter at stump height (mm, SD, measured 0.3 m above

201
ground) and DBH (mm). Where trees presented branches or other features that may
have changed the diameter at this point, the decision was taken to measure at the
nearest regular point below the irregularity. In the case of the RCD, if the base of the
tree was irregularly damaged, this measurement was excluded. In trees where
multiple stems were present, each stem was measured and the mean average was
reported (Parent, 2000). A 5 m measuring tape was used to measure crown
diameter (CD, mm), taken as the mean average of three horizontal measurements of
the crown perpendicular to the ground. The crown radius (r) was calculated as the
crown diameter divided by two. Crown depth was derived using equation 4. Crown
area (CA) was assumed to be a circle through observation and was derived using
equation 5. Crown volume (CV) was assumed to equate to a cylinder, the best fit for
the pollarded trees, and was calculated using equation 6:
(Equation 4) Crown depth = height – crown height
(Equation 5) CA = πr2
(Equation 6) CV =πr2CD
Trees were felled at ground level with a chainsaw. For each tree, biomass was
divided into stem, branch and foliage. Due to resource constraints, only aboveground
biomass was considered in the present study. Each biomass component was
weighed separately using an appropriate spring-loaded scale. All biomass was
grouped according to its component and three subsamples drawn from each at the
end of every sampling period. Subsamples were dried at 80 °C until a constant dry
weight was reached. Stem samples were divided into 1 m sections and their fresh
weight volume calculated according to Newton’s equation:

202
(Equation 7) 𝑉 =𝐿(𝐴𝑏+4𝐴𝑚+𝐴𝑠)
6
Where b is diameter at the base, m is diameter at the middle and s is diameter at the
endpoint of the stem section.
2.2 Data Exploration
All analyses were carried out using the statistical program R (3.2.2, Mac OS X).
Variables were visually assessed for relationships and outliers using scatter plots.
Spearman’s correlation tests were used to determine the level of correlation between
variables as Spearman’s is a ranked approach and is therefore not sensitive to data
transformations.
The present study hypothesises that the growth presented by the species in this
study is multiplicative and therefore the best-fit models will be log-linear. This
assumption must be tested because the decision to log transform the data should
not be arbitrary (Sileshi, 2014). Three modelling approaches were chosen to test the
error distributions and fit for the formulation of allometric equations, linear, log linear
and quadratic. The residual plots and error distributions were reviewed for these
three modelling approaches. In all models both normal and kernel density distribution
models were used to plot the residuals against the predicted values to test for
normality and homogeneity. Outliers were detected by using Cook’s Distance.

203
2.3 Model Comparison and Selection
Linear, log-linear and quadratic equations were developed and compared using
Furnival’s Index for the selection of the most appropriate model (Furnival, 1961):
(Equation 8) F. I. = rMSE
GM(d(𝑓(Y))/dY)
Where rMSE is the root mean square error of the fitted equations, f(Y) is the
transformation function on the dependent variable d(f(Y))/dY is the derivative of the
transformation fitted to Y, and GM denotes the geometric mean of 1/Y (Kershaw et
al., 2016).
Furnival’s Index (F.I.) can be compared between models with transformed and
untransformed variables. A lower F.I. results from a lower error associated with the
model and therefore an improved fit. The R2 or coefficient of determination is
typically used to compare models and is interpreted as a percentage explanation of
the model fit of the data where a higher R2 represents an improved fit. The R2 is
derived from the sum of squares residual divided by the sum of squares total (the R2
can also be ‘adjusted’ for increased model parameters). However, it is inappropriate
to compare the R2 of a model with the R2 of a model that uses a transformed
predictor variable.
2.4 Model Validation
The models were cross-validated by the repeated cross validation technique. The
dataset was divided into ‘k groups’ also known as ‘k-folds’ where the training data
was defined as k = 10, which results in a manageable bias-variance trade-off, and

204
the test data was k-1 (James et al., 2013; Arlot and Celisse, 2010). This can also be
described as a random 10% subset of the data being used to test 90% of the data.
This process was repeated three times. The mean average of the root of the residual
mean squared error for three repeats is reported (rMSE). The data were
subsequently combined and the full dataset was used to generate the allometric
models.
2.5 Allometric Models
The following linear equation was used to test allometric models:
(Equation 9) y = a + b * X
Where a is the intercept coefficient, b is slope coefficient, y is a biomass component
either total biomass, trunk biomass, branch biomass, leaf biomass and X is Root
Collar Diameter (RCD, mm), Diameter at Breast Height (DBH, mm), Stump Diameter
(SD, mm), Crown Volume (CV, cm3), Crown Diameter (CD, cm) or as a function of
both RCD and Height (cm). These parameters were also modeled in the quadratic
form, where c is an additional model coefficient:
(Equation 10) y = aX2 + bX + c
Kitteredge (1944) found that Huxley's (1932) equation could be used to derive the
biomass of several species of spruce and pine trees. This is a common allometric
equation for the estimation of tree biomass:
(Equation 11) y = aXb

205
This can also be used in the logarithmic form to produce a log-linear model. The use
of the logarithmic form requires back transformation of the dependent variable. This
paper calculates the error associated with log transformation as lambda (λ) and
associated standard deviation according to the equation proposed by Marklund
(1987), where n is sample size:
(Equation 12) 𝜆 = ∑ 𝑌𝑖
𝑛𝑖=1
∑ 𝑒ln ��𝑖𝑛𝑖=1
The following log-linear allometric model was tested:
(Equation 13) ln(y) = a + b * lnX
Where ln(y) is the natural logarithmic transformation of the biomass components or
expansion factors listed above, ln(X) is the log transformation of the measurements
listed above. Where multiple predictors were present in the model the adjusted
coefficient of determination (R2) was reported.
2.6 Calculation of Carbon Stocks
The C content was estimated by multiplying the individual tree biomass (Mg ha-1) by
the IPCC default C value for trees in tropical forests of 0.47 (Eggleston et al., 2006).
The value for the C sequestration of each tree was then multiplied by the planting
density (2,667 trees ha-1). The average and standard deviation was reported. To
calculate mean annual increment (MAI) of biomass produced and C sequestered the
results of the total biomass at the 24-month destructive harvest and the pollard of
biomass at 18 months over 2 m were combined and divided by the number of years
of data. No estimate was made of the carbon present in belowground biomass as
this data was not collected in the present study due to resource constraints.

206
3. Results
3.1 Measurements of Tree Biomass
Subsamples of all biomass components showed strong relationships between wet
and dry weight (Appendix 1). The contribution of foliar biomass to total biomass was
3% in E. berteroana (Table 7), 15% in I. edulis (Table 8) and 2.4% in L. leucocephala
(Table 9). The sample size (n) of 68 for DBH of E. berteroana in Table 7 is due to not
all trees exceeding the 1.3 m at which the measurement was taken. I. edulis
produced over 60% more total biomass and 660% more foliar biomass than either E.
berteroana or L. leucocephala. I. edulis had a 223% greater crown volume than that
of E. berteroana or and a 163% greater crown volume than that of L. leucocephala.
I. edulis had 111% more branch biomass than E. berteroana and 220% more branch
biomass than L. leucocephala. Average trunk biomass of all three species were
similar 1.3 ± 0.8 kg for I. edulis, 1.0 ± 0.8 kg for E. berteroana and 1.0 ± 0.5 kg for L.
leucocephala. 6 months post pollard to 2 m, L. leucocephala had grown by 1 m to a
height of 298 cm, which represented an increase on the 242 cm height of I. edulis
and 219 cm height of E. berteroana. L. leucocephala was the species with the lowest
average diameter measurements for RCD (44 mm, compared to 67 mm of I. edulis
and 63 mm of E. berteroana) SD (35 mm, compared to 50 mm and 48 mm of I.
edulis and E. berteroana respectively) and had a DBH (22 mm) similar to that of E.
berteroana (21 mm) but lower than that of I. edulis (34 mm).

207
Table 7. Descriptive statistics of biomass parameters of individual harvested Erythrina berteroana used in constructing allometric equations, including sample size (n), root collar diameter (RCD, 0.05 m), stump diameter (SD, 0.3 m) and diameter at breast height (DBH, 1.3 m). Mean, standard deviation (SD), minimum (min), maximum (max), lower 25th percentile (25th perc), upper 75th percentile (75th perc), as well as coefficients of skewness and kurtosis are presented.
Variable n Mean SD Min Max 25th perc 75th perc Skewness Kurtosis
Total Biomass (g) 73 1316.3 1005.1 36.4 4517 505.3 1935 1.1 0.8
Trunk Biomass (g) 73 1015.4 792.9 35.2 4087 417.9 1378 1.3 2.1
Branch Biomass (g) 72 262.6 315.8 0.6 1942 61.3 360.7 2.7 9.8
Leaf Biomass (g) 73 42 51.5 0.4 231.3 9.2 67 1.9 3.5
RCD (mm) 73 62.6 21.5 21.6 123.1 50.7 74.8 0.4 0.4
SD (mm) 73 48.2 18.3 17 105.4 33.6 61.6 0.5 0.1
DBH (mm) 68 21.4 7.6 8.4 47.7 15.8 25.4 0.8 1.0
Height (cm) 73 218.8 53.2 72 200 218.8 314.0 -0.74 0.1
Crown Volume (cm3) 73 939219.3 1028901 1341.8 4125900 156800 1390000 1.5 1.5
Crown Diameter (cm) 73 88.5 42.9 10.3 191.7 53.3 122.3 0.4 -0.7

208
Table 8. Descriptive statistics of biomass parameters of individual harvested Inga edulis used in constructing allometric equations, including sample size (n), root collar diameter (RCD, 0.05 m), stump diameter (SD, 0.3 m) and diameter at breast height (DBH, 1.3 m). Mean, standard deviation (SD), minimum (min), maximum (max), lower 25th percentile (25th perc), upper 75th percentile (75th perc), as well as coefficients of skewness and kurtosis are presented.
Variable n Mean SD Min Max 25th perc 75th perc Skewness Kurtosis
Total Biomass (g) 78 2191.2 1402.6 177 6692 1269 2943 0.9 0.3
Trunk Biomass (g) 78 1327 804.7 162.5 4229 721.5 1795 1.0 1.3
Branch Biomass (g) 78 554.6 579.2 0.3 2190 71.4 835.8 1.1 0.4
Leaf Biomass (g) 78 323.1 332.7 1.6 1290 66.7 448.4 1.3 1.1
RCD (mm) 78 65.6 22.5 12.8 127.2 52.4 80.8 0.1 -0.3
SD (mm) 78 50.3 16.4 16.1 87.2 39.2 60.3 0.1 -0.5
DBH (mm) 78 34.4 14.5 12.9 89.7 25.5 39.6 1.6 3.3
Height (cm) 78 242.1 45.9 108 358 220.0 262 -0.3 2.1
Crown Volume (cm3) 78 3039987 3226258 148100 16300000 1013000 3855000 2.3 5.0
Crown Diameter (cm) 78 149.3 64.8 52.3 485.7 107.3 179.5 1.8 6.7

209
Table 9. Descriptive statistics of biomass parameters of individual harvested Leucaena leucocephala used in constructing allometric equations, including sample size (n), root collar diameter (RCD, 0.05 m), stump diameter (SD, 0.3 m) and diameter at breast height (DBH, 1.3 m). Mean, standard deviation (SD), minimum (min), maximum (max), lower 25th percentile (25th perc), upper 75th percentile (75th perc), as well as coefficients of skewness and kurtosis are presented.
Variable n Mean SD Min Max 25th perc 75th perc Skewness Kurtosis
Total Biomass (g) 63 1259.5 670.1 178 2941.9 721.3 1665 0.39 -0.5
Trunk Biomass (g) 63 1056.5 546.8 174.3 2661 643.6 1378 0.44 -0.2
Branch Biomass (g) 63 172.6 215.9 0.4 1300 24.8 215.4 2.7 9.9
Leaf Biomass (g) 63 30.4 36.4 0.4 197.3 5.1 38.7 2.09 5.5
RCD (mm) 63 44 12.8 20.2 2661.3 643.6 1378 0.13 -0.5
SD (mm) 63 35 10.8 9.5 61.3 27.3 42.8 -0.1 -0.4
DBH (mm) 63 22.2 8.6 5.6 43.5 16.3 28.9 0.45 -0.5
Height (cm) 63 293.7 63.7 144 420 250 343 0.1 -0.8
Crown Volume (cm3) 63 1158571 1349790 15400 6355000 186100 1798000 1.65 2.6
Crown Diameter (cm) 63 97.8 45.5 30 220 63.5 120 0.72 -0.1

210
3.2 Data Exploration
When Spearman’s correlations were compared between biomass parameters and
tree measurements for E. berteroana, the total biomass showed the strongest
correlation with the measurement of crown volume (0.75), the trunk biomass equally
by RCD (0.71) and SD (0.71), the branch biomass by crown diameter (0.83) and the
leaf biomass by crown diameter (0.77) (Table 10). RCD and height did not have
strong correlation (0.39). For I. edulis, RCD was the strongest predictor of total
biomass (0.82), trunk biomass (0.81), and branch biomass (0.66). Crown volume
was found to have the strongest correlation with leaf biomass (0.68) (Table 11).
Height was found to have an association (0.57) with total biomass and showed a
weak correlation (0.45) with RCD. For L. leucocephala RCD was the strongest
predictor of total biomass (0.87) and trunk biomass (0.91). Branch biomass was
equally predicted by crown volume (0.67) and crown diameter (0.67). Crown
diameter was also the most correlated measurement with leaf biomass (0.78) (Table
12). In L. leucocephala, RCD had a weak correlation with height (0.45).

211
Table 10. Spearman’s correlation of E. berteroana biomass components. Total biomass (Total), trunk biomass (Trunk), branch biomass (Branch), total foliar biomass (Leaf), root collar diameter (RCD, 0.05 m), stump diameter (SD, 0.3 m), diameter at breast height (DBH, 1.3 m), height, crown volume (CVol) and crown diameter (CD).
Total Trunk Branch Leaf RCD SD DBH Height CVol CD
Total 1 0.97 0.7 0.78 0.73 0.68 0.68 0.67 0.75 0.73
Trunk 0.97 1 0.55 0.72 0.71 0.71 0.68 0.67 0.69 0.64
Branch 0.7 0.55 1 0.7 0.63 0.43 0.51 0.43 0.72 0.83
Leaf 0.78 0.72 0.7 1 0.55 0.53 0.58 0.64 0.71 0.77
RCD 0.73 0.71 0.63 0.55 1 0.7 0.6 0.39 0.55 0.59
SD 0.68 0.71 0.43 0.53 0.7 1 0.63 0.41 0.46 0.54
DBH 0.68 0.68 0.51 0.58 0.6 0.63 1 0.55 0.56 0.58
Height 0.67 0.67 0.43 0.64 0.39 0.41 0.55 1 0.81 0.56
CVol 0.75 0.69 0.72 0.71 0.55 0.46 0.56 0.81 1 0.85
CD 0.73 0.64 0.83 0.77 0.59 0.54 0.58 0.56 0.85 1
Table 11. Spearman’s correlation of I. edulis biomass components. Total biomass (Total), trunk biomass (Trunk), branch biomass (Branch), total foliar biomass (Leaf), root collar diameter (RCD, 0.05 m), stump diameter (SD, 0.3 m), diameter at breast height (DBH, 1.3 m), height, crown volume (CVol) and crown diameter (CD).
Total Trunk Branch Leaf RCD SD DBH Height CVol CD
Total 1 0.88 0.86 0.8 0.82 0.7 0.49 0.49 0.66 0.67
Trunk 0.88 1 0.6 0.58 0.81 0.8 0.57 0.44 0.58 0.58
Branch 0.86 0.6 1 0.71 0.66 0.55 0.38 0.38 0.57 0.63
Leaf 0.8 0.58 0.71 1 0.65 0.5 0.32 0.53 0.68 0.67
RCD 0.82 0.81 0.66 0.65 1 0.78 0.62 0.47 0.67 0.66
SD 0.7 0.8 0.55 0.5 0.78 1 0.69 0.4 0.55 0.54
DBH 0.49 0.57 0.38 0.32 0.62 0.69 1 0.24 0.36 0.36
Height 0.49 0.44 0.38 0.53 0.47 0.4 0.24 1 0.63 0.4
CVol 0.66 0.58 0.57 0.68 0.67 0.55 0.36 0.63 1 0.88
CD 0.67 0.58 0.63 0.67 0.66 0.54 0.36 0.4 0.88 1
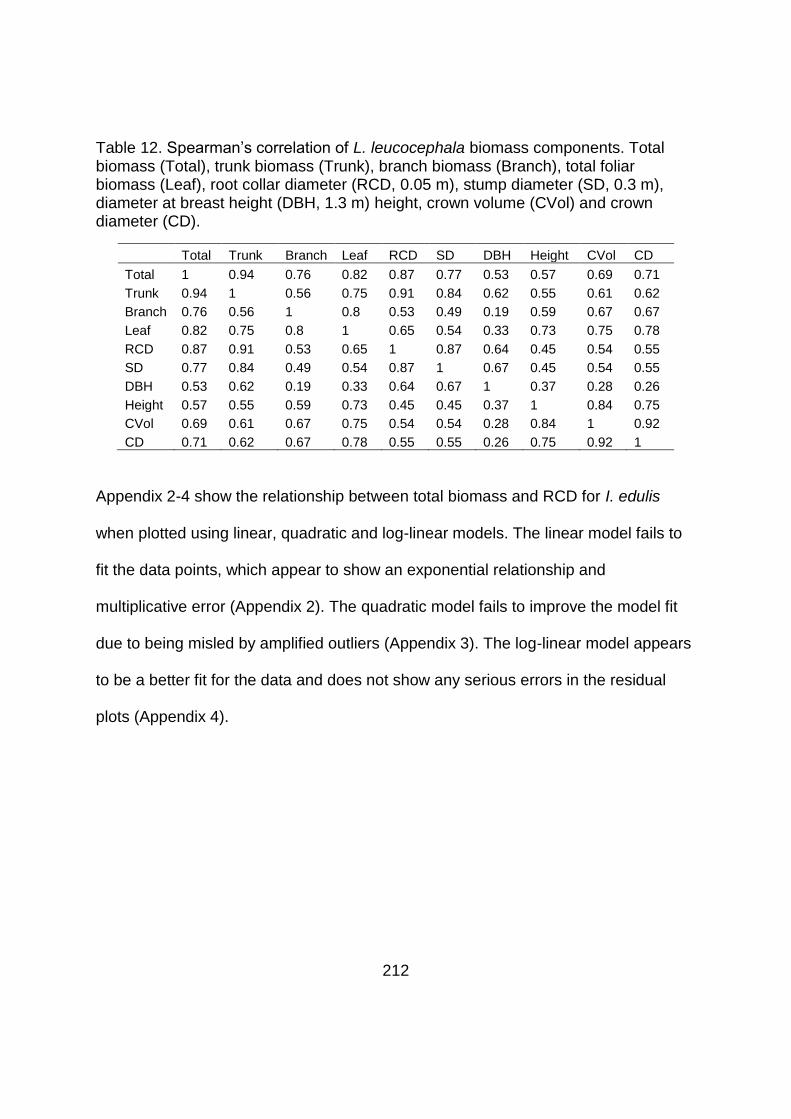
212
Table 12. Spearman’s correlation of L. leucocephala biomass components. Total biomass (Total), trunk biomass (Trunk), branch biomass (Branch), total foliar biomass (Leaf), root collar diameter (RCD, 0.05 m), stump diameter (SD, 0.3 m), diameter at breast height (DBH, 1.3 m) height, crown volume (CVol) and crown diameter (CD).
Total Trunk Branch Leaf RCD SD DBH Height CVol CD
Total 1 0.94 0.76 0.82 0.87 0.77 0.53 0.57 0.69 0.71
Trunk 0.94 1 0.56 0.75 0.91 0.84 0.62 0.55 0.61 0.62
Branch 0.76 0.56 1 0.8 0.53 0.49 0.19 0.59 0.67 0.67
Leaf 0.82 0.75 0.8 1 0.65 0.54 0.33 0.73 0.75 0.78
RCD 0.87 0.91 0.53 0.65 1 0.87 0.64 0.45 0.54 0.55
SD 0.77 0.84 0.49 0.54 0.87 1 0.67 0.45 0.54 0.55
DBH 0.53 0.62 0.19 0.33 0.64 0.67 1 0.37 0.28 0.26
Height 0.57 0.55 0.59 0.73 0.45 0.45 0.37 1 0.84 0.75
CVol 0.69 0.61 0.67 0.75 0.54 0.54 0.28 0.84 1 0.92
CD 0.71 0.62 0.67 0.78 0.55 0.55 0.26 0.75 0.92 1
Appendix 2-4 show the relationship between total biomass and RCD for I. edulis
when plotted using linear, quadratic and log-linear models. The linear model fails to
fit the data points, which appear to show an exponential relationship and
multiplicative error (Appendix 2). The quadratic model fails to improve the model fit
due to being misled by amplified outliers (Appendix 3). The log-linear model appears
to be a better fit for the data and does not show any serious errors in the residual
plots (Appendix 4).

213
3.3 Model Selection
When the Furnival’s Index of all modelled approaches was compared, it was
revealed that the log-linear models are always superior to the quadratic or linear
models for all biomass parameters in all species in this study (Appendix 5 - 22). The
models with the lowest Furnival’s Index (F.I.) for each biomass parameter are
presented in Table 13. The best-fit models are plotted in Figures 48 - 51. It was only
possible to graphs with the data points for linear models (Fig 49 and Fig 50). For
species where quadratic models offered best fit the regression line has been plotted
without the data points (Fig 48 and Fig 51).
In this study, the species differ in which measurements can be used to best predict
components of biomass. The inclusion of height with RCD as a multiple predictor
resulted in the lowest F.I. for total and trunk biomass in E. berteroana and L.
leucocephala. For I. edulis the inclusion of height did not improve the prediction of
biomass and the single predictor of RCD had the lowest F.I. for all of the biomass
measurements. Predictors for the total biomass of L. leucocephala (R2=0.86) were
more effective then predictors of total biomass of either E. berteroana (R2=0.77) or I.
edulis (R2=0.70). The hypothesis that measurements of the crown would be the best
predictor of total foliar biomass was upheld for E. berteroana by the measurement of
crown diameter (R2=0.58) but rejected for L. leucocephala (RCD and height as
combined predictors with lowest F.I.) and I. edulis (RCD as single predictor with
lowest F.I.).

214
When the root mean square error (rMSE) was compared within species between
biomass parameters, the rMSE in I. edulis branch biomass was increased 282%
when compared to the rMSE present in the prediction of trunk biomass and leaf
biomass showed a 205% increase in rMSE when compared to trunk biomass. For E.
berteroana the branch biomass rMSE was increased 42% compared to trunk
biomass, and leaf biomass rMSE increased 96% compared to trunk biomass. For L.
leucocephala the branch biomass had a 495% increased rMSE than trunk biomass,
the leaf biomass rMSE showed a 320% increase when compared with the rMSE of
trunk biomass.

215
Table 13. Log-linear allometric equations demonstrated the lowest Furnival’s Index (F.I.) against linear or quadratic models for biomass components of 0.5 x 7.5 m alley rows of Erythrina berteroana, Inga edulis and Leucaena leucocephala 6 months post pollarding to a height of 2 m. Where ln(x) is the log transformation of the following measurements used as predictors of biomass: Root collar diameter (RCD mm, 0.05 m), height (cm) and crown diameter (CD, cm). ln(y) is the log transformation of total biomass (g), trunk biomass (g), branch biomass (g) or leaf biomass (g). Where b0 is the intercept coefficient, b1 and b2 are the slope coefficients, all of which are reported with standard error (S.E.) and significance (P). Lambda (λ) and standard deviation (S.D.) as back transformation according to Marklund (1987), root mean square error of model (Model rMSE) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV rMSE).
Species ln(y)
(Biomass) ln(x) b0 (S.E.) P b1 (S.E.) P b2 (S.E.) P R2 λ S.D. F.I. Model rMSE
CV rMSE
E. berteroana Total RCD + Height -6.41 (1.02) <0.001 1.53 (0.2) <0.001 1.31 (0.25) <0.001 0.77 1.12 0.83 523.2 0.47 0.49
Trunk RCD + Height -6.91 (1.03) <0.001 1.38 (1.38) <0.001 1.47 (0.25) <0.001 0.76 1.13 0.94 409.5 0.48 0.50
Branch CD -3.99 (0.65) <0.001 2.04 (0.15) <0.001 0.73 0.25 0.19 104.8 0.68 0.74
Leaf CD -7.06 (1.07) <0.001 2.28 (0.24) <0.001 0.58 1.49 1.36 20.1 0.94 0.93
I. edulis Total RCD 0.56 (0.51) 0.28 1.68 (0.12) <0.001 0.70 1.07 0.54 759.2 0.44 0.46
Trunk RCD 0.95 (0.43) 0.03 1.47 (0.10) <0.001 0.71 1.07 0.56 409.6 0.38 0.39
Branch RCD -8.89 (1.67) <0.001 3.45 (0.40) <0.001 0.48 2.37 4.64 287.9 1.45 1.51
Leaf RCD -6.47 (1.33) <0.001 2.78 (0.32) <0.001 0.48 2.19 6.52 169.1 1.16 1.20
L. leucocephala Total RCD + Height -3.04 (0.79) <0.001 1.69 (0.12) <0.001 0.66 (0.16) <0.001 0.86 0.98 0.26 64.33 0.24 0.25
Trunk RCD + Height -2.26 (0.67) 0.001 1.71 ( 0.1) <0.001 0.47 (0.13) <0.001 0.89 1.03 0.21 39.02 0.20 0.21
Branch CD -7.16 (1.43) <0.001 2.55 (0.32) <0.001 0.51 1.86 2.33 83.7 1.19 1.23
Leaf RCD + Height -21.53 (2.75) <0.001 1.92 (0.40) <0.001 3.01 (0.55) <0.001 0.61 1.34 1.05 10.62 0.84 0.89

216
Figure 48. The log transformation of x (root collar diameter (mm) + height (cm)) derived using a linear model for the log transformation of 1) y as total biomass (g, R2 = 0.77, P <0.01) and 2) y as trunk biomass (g, R2 = 0.76, P<0.01) for Erythrina berteroana following the equations presented in Table 13.
Figure 49. The log transformation of crown diameter (cm) derived using a linear model for the log transformation of branch biomass (g, R2 = 0.73, P<0.01) and leaf
0 1 2 3 4 5 6 7
−6
−4
−2
02
4
ln(x)
ln(y
)Biomass
Total
Trunk
3.5 4.0 4.5 5.0
23
45
67
ln(CROWN DIAMETER)
ln(B
IOM
AS
S)
Biomass
Branch
Leaf

217
biomass (g, R2 = 0.58, P<0.01) for Erythrina berteroana following the equations presented in Table 13.
Figure 50. The log transformation of root collar diameter (mm) derived using a linear model for the log transformation of total biomass (g, R2 = 0.70, P<0.01), trunk biomass (g, R2 = 0.71, P<0.01), branch biomass (g, R2 = 0.48, P<0.01) and leaf biomass (g, R2 = 0.48, P<0.01) for Inga edulis following the equations presented in Table 13.
Figure 51. The log transformation of x where 1) x is root collar diameter (mm) + height (cm) derived using a linear model for the log transformation of total biomass
2.5 3.0 3.5 4.0 4.5
67
8
ln(RCD)
ln(B
IOM
AS
S)
Biomass
Total
Trunk
Branch
Leaf
0 1 2 3 4
−2
02
46
8
ln(x)
ln(y
)
Biomass
Total
Trunk
Branch
Leaf

218
(g, R2 = 0.86, P<0.01), trunk biomass (g, R2 = 0.89, P<0.01) and leaf biomass (g, R2 = 0.61, P<0.01) and 2) x is crown diameter (cm) derived using a linear model for the log transformation of branch biomass (g, R2 = 0.51, P<0.01) for Leucaena leucocephala following the equations presented in Table 13.
Total above ground biomass fraction and C stock after 24 months is presented in
Table 14. When the biomass from the destructive harvest at 24 months was
combined with the biomass from the pollard exercise at 18 months to represent total
accumulated biomass over the course of the 24 months of this study, I. edulis had
accumulated the most biomass and therefore more C (3.4 Mg C ha-1) than E.
berteroana (2.2 Mg C ha-1) and L. leucocephala (2.0 Mg C ha-1) (Table 14).
Table 14. Total above ground biomass fraction and carbon (C) stock (Mg ha-1) after 24 months, including previously removed biomass > 2 m at 18 months (Chapter III) of Erythrina berteroana, Inga edulis and Leucaena leucocephala when 2667 trees ha-1 (0.5 x 7.5 m), using the IPCC default C fraction for tropical forests of 0.47. Where n is sample size and MAI is Mean Annual Increment (Mg ha-1 year-1) (Eggleston et al., 2006).
Species n Tree Biomass
Mg ha-1
MAI Tree Biomass C Mg ha-1 MAI C
E. berterona 106 4.4 2.2 2.1 1.0
I. edulis 112 7.2 3.6 3.4 1.7
L. leucocephala 96 4.3 2.2 2.0 1.0
4. Discussion
The results of this study show that Erythrina berteroana, Inga edulis and Leucaena
leucocephala can produce substantial amounts of biomass after 24 months and
show good potential for the rapid sequestration of C in above ground biomass. This
study shows a considerable increase in C sequestration via the addition of these
trees when the tree biomass is assumed to be additional to the business-as-usual

219
carbon sequestration of monoculture pastures in the region. This assumption is
based on the trees being managed in such a way so as to have negligible impact on
grass production (see Chapter III). Further work may be carried out that examines
the interaction of C sequestration by grassland or silvopasture based on the
interaction of the tree and grass elements in such systems.
This paper alludes to an assumption in the literature that the ratio of dry weight of tree
biomass to carbon content averages around 0.47-0.49. The estimates of C in this study
were drawn from the IPCC report that reports the average value for tropical forests
(Eggleston et al., 2006). This assumption was challenged by the results of a study in
Panama that showed the ratio of biomass dry weight taken from tree cores to carbon to
vary from a proportion of 0.42–0.52 when 59 rainforest trees were sampled (Martin
and Thomas, 2011). The same study did not find any correlation between C stocks with
relative growth rate or wood density. The reliability of the estimates of C
sequestration in the present study could be improved by specific analysis of C
content in the plant material between species and biomass components and the
inclusion of sampling that considers the production of below ground biomass to
enable the C sequestered belowground to be estimated.
Site level differences such as planting arrangement, water availability, nutrient
availability, soil compaction, temperature, solar irradiation and species associations
may also limit the transferability of one study’s findings for a species biomass

220
accumulation to other regions (Sharma and Ambasht, 1991; Prasad et al., 2011).
Montagnini and Nair (2004) found that agroforestry practiced by small holders in the
tropics had a C sequestration potential of 1.5 to 3.5 Mg C ha-1 year-1 (Montagnini and
Nair, 2004). Less is known about the effects of pollarding or biomass removal on C
sequestration potential of these systems. A fodder bank in Mali, west Africa of Gliricidia
sepium, Pterocarpus lucens and P. erinaceus at 7.5 years was found to sequester
0.29 Mg C ha-1 year-1 (Takimoto et al., 2008).
The findings of the present study can be viewed in light of what other authors have
found to be true of the biomass production in different planting arrangements and
site conditions. Szott et al. (1994) planted I. edulis in Yurimaguas, Peru, in an
arrangement of 2 x 2 m and used allometric equations based on the measurement of
diameter of the trunk 0.1 m above soil (hereafter D10) for 10 harvested individuals
with a D10 between 1 and 5.5 cm to estimate the total tree biomass. They estimated
that at 17 months I. edulis had an average of 10 Mg ha-1 of biomass and by 29
months this had increased to 14.24 Mg ha-1. Shimamoto et al. (2014) measured the
biomass production of 12-year-old naturally regenerating I. edulis in the Parana
State in Southern Brazil. They sampled 9 trees and found annual accumulation of
20.73 kg (± 12.09) to 24.48 kg (± 19.71) biomass tree-1. Prasad (2011) found that in
southern India, after 51 months, a spacing of 0.8 x 5 m for L. leucocephala resulted
in a total biomass production of 47 kg tree-1 (2,500 trees ha-1) whereas 1 x 1 spacing
resulted in 38 kg tree-1, but that the per hectare biomass was higher in the 1 x 1 m
spacing (10,000 trees ha-1). López et al. (2008) planted L. leucocephala in

221
southwestern Spain at a density of 9,500 trees ha-1 and found a total biomass
production for three provenances of 23.4 Mg ha-1 (±2.9), 30.9 Mg ha-1 (±6.9) and
25.6 Mg ha-1 (±13.1) at 24 months. E. berteroana in a live fence arrangement in
Costa Rica was reported to produce 19.4 Mg ha-1 after 24 months, but the planting
arrangement was not specified (Pezo et al., 1990). Pennington (1998) planted I.
edulis at 3 x 3 m spacing at trial sites in on the Pacific Coast and the Amazon of
Ecuador. I. edulis consistently demonstrated an increased height and diameter
growth rate when compared to four other species of Inga, and produced 2.5 Mg ha-
1year-1 of woody biomass.
In this study, I. edulis had the highest rates of overall biomass production. While the
trunk biomass was relatively similar for all three species, I. edulis produced over six
times (660%) more foliar biomass than E. berteroana or L. leucocephala. Szott et al.
(1994) in Yurimaguas, Peru, found that for small I. edulis trees (D10 = 2.5 cm), trunk
accounted for 33% of total biomass, branches for 21% and leaves for 45%. In larger
trees (D10= 10 cm) the biomass was 51% trunk, 32% branches and 17% leaves. In
the present study, the contribution of biomass components to total biomass was 60%
trunk, 25% branches and 15% leaf. The differences between the present study with
that of Szott et al. (1994) may be as a result of the pollard > 2 m which may have
resulted in less trunk biomass being removed than branch and leaf components.
As expected, the log-linear models for all biomass parameters in all species had the
lowest Furnival’s Index and therefore can be considered the best models for the

222
prediction of biomass. The inclusion of multiple predictors in the model (RCD +
height) did not improve the model fit for I. edulis, but did improve the model fit for the
total and trunk biomass of E. berteroana and L. leucocephala. Lojka et al. (2005)
found that power relationships on untransformed variables were the best fit for the
prediction of I. edulis biomass. They destructively sampled 35 trees and measured
leaf litter, diameter of trunk at D10, age of trees and height. They found the highest
prediction of wood biomass (kg) by the measurement of diameter (y = 0.0466x2.3713;
R2= 0.92). D10 was also the strongest predictor of leaf biomass (kg) (y =
0.1564x1.1817; R2=0.79). Szott et al. (1994) developed allometric equations for I.
edulis based on the destructive harvest of 27 trees with a D10 of 1 – 5.5 cm and
develop allometric equations for the stem biomass (kg) (y = 0.089*(D10)2; R2=0.77),
branch biomass (kg) (y = 0.057*(D10)2; R2=0.96) and leaf biomass (kg) (y =
0.3003*(D10); R2=0.85). In the present study, the R2 of best performing models
shows poorer performance than the models present in the literature. It is likely that
the lower R2 in this study is a consequence of the difference in management
approach in this study which included pollarding all trees over 2 m at 18 months.
Managed forest plots, where trees undergo management interventions such as
coppicing or pollarding, could be expected to demonstrate different allometric
relationships when compared with trees that are free from such interference.
However, it is the effect of the management on the trees that is precisely of interest,
as the management will effect the production of key ecosystem services from the
systems. The pipe model theory asserts that the diameter of a tree is proportional to

223
the water requirements of the tree canopy and that the canopy size is determined by
the transpirational rate (Shinozaki et al., 1964). The pipe model is one of the theories
from which allometric equations are believed to derive their power.
Nygren et al. (1993) state that repeated pruning of the tree alters the development of
the trunk diameter as it changes the leaf area to trunk ratio. They propose the
formation of equations based on measurements leaf area and leaf mass related to
the diameter of branches that form following pruning. Their approach was successful
for four clones of E. berteroana, the average of which produced an R2 of 0.89 with
the equation: leaf biomass = 0.2209*CS, where CS is branch cross sectional area
measured in mm. The R2 for the leaf biomass of E. berteroana in the present study
was lower (R2= 0.58) and future studies could consider the methodological approach
of Nygren et al. (1993), which may be superior for the prediction of foliar biomass.
Lott et al. (2000) found that the approach followed by Nygren (1993) produced
reliable estimates of canopy leaf area and biomass for Grevilla robusta agroforestry
systems, but proved unsuitable for routine measurements due to the time consuming
nature of this approach. Lott et al. (2000) suggest that the measurement of trunk
cross sectional area immediately below the first branch was suitable for the
prediction of leaf area and canopy biomass in Grevilla robusta managed for poles.
There is evidence that regrowth of trees following pruning and pollarding may be in
part dependent on carbohydrate reserves, which can be depleted by coppicing or
pollarding regimes and in some cases there may also be effects of seasonality (Kays
and Canham, 1991; Raitio et al., 1994).

224
In temperate regions where plants may have well defined growing seasons, the
timing of biomass management (via, e.g. directly via pruning/pollarding/coppicing or
browsing) has been shown to have an effect on the production of biomass (Douglas,
1996; Opping, 2002). Burner (2006) studied the effect of harvest date on Robinia
pseudoacacia and found temporal variation in allometric equations post-pollard, as
foliar mass accumulated more quickly in some months than in others. Therefore,
they developed month specific equations. Seasonality would also have had a strong
effect on the trees in this study, and month specific equations may have been a more
appropriate approach but was not followed in this study. The present study could be
improved by characterising the effects of both the frequency of pollarding and also
how the timing of pollarding relative to how tropical seasonality effects foliar
production in an attempt to increase understanding of how to improve management
and production of these species.
It is a convention in forestry that DBH is the most commonly used measurement for
the development of allometric equations because it can be easily measured with high
accuracy (Kershaw et al., 2016). For all species in this experiment, when the three
heights of stem diameter measurements were compared, the RCD had the highest fit
to total biomass, followed by SD. DBH demonstrated the poorest fit. This suggests
that the RCD is a valuable parameter that could be included in future studies that
attempt to develop allometric equations for young trees in agroforestry systems. The
stem measurement of RCD was the best single predictor of total biomass for I. edulis

225
and L. leucocephala. RCD has been used when a high proportion of sample trees
are below the 1.3 m at which DBH is traditionally measured (Dutcă et al., 2018).
However, equations developed from RCD may be less accurate for trees with
buttress roots or stilts. C. pentandra was included in this experiment, and is a
species that develops buttress roots. As this tree grows the reliability of the
measurement of RCD for total biomass is likely to change. Further work would be
required to understand more about this relationship, and how RCD changes in
reliability between size classes across species. Rubilar et al. (2010) measured both
RCD and DBH for Pinus radiata and found similar results for both variables. Guedes
(2018) found that measurements of RCD were interchangeable with DBH in the
lowland miombo woodlands of Mozambique. This may be useful to estimate the
biomass of trees that have been removed from their stumps. Jagodziński et al.
(2018) developed biomass equations for Pinus sylvestris, a species they observed to
be prone to root collar swelling. Therefore, they used root collar diameter only when
trees were shorter than 0.5 m. Dutcă (2018) used root collar diameter as a
parameter on young Norway spruce trees with a maximum height of 5.5 m. Kuyah et
al. (2013) found that D10 was better at predicting below ground biomass (R2 = 0.91)
than above ground biomass (R2 = 0.88), but overestimated the biomass of most
components by 20%. They discussed that these errors were, in part, due to
uncertainties in the location of the measurement of D10. MacCracken and
Ballenberghe (1993) tested multiple models for moose browse on the Copper River
Delta, Alaska and found that mass of leaf and twig production was best predicted by
RCD.

226
Authors have suggested that the inclusion of additional variables, such as height, will
increase the predictive powers of models (Chave et al., 2005). When the additional
variable height was included in the RCD model this became the strongest predictive
model for the total biomass of L. leucocephala and E. berteroana. The inclusion of
height did not improve the model fit for I. edulis. Height and stem diameter
measurements are frequently too highly correlated for both to be included in the
same model, due to the issue of multicollinearity (Balehegn et al., 2012; Sileshi,
2014). To avoid multicollinearity, some authors have tested both height and diameter
independently and included whichever variable results in improved fit or is easiest to
measure (Pajtek, 2008). In some cases they have combined the two variables into a
single variable to improve model fit, such as the D2H in Dutcă et al. (2018) and the
DBH2H in Rubilar et al. (2010). It is possible that the inclusion of height did not
improve the model fit for I. edulis as a consequence of the pollard at 2 m. I. edulis
may have responded to the increased availability of light following pollarding by
investing more resources into the growth of lighter foliar biomass, as opposed to the
production of heavier woody biomass. Of all species, I. edulis demonstrated the
highest foliar biomass production relative to other biomass components. The foliar
biomass, which weighs less than the woody biomass, would not be represented by
the height of the tree. Therefore, it may be the case that tree species that have a
high proportion of foliar to total biomass have a greater need for species specific
models, particularly when the trees are managed by pollarding or removal of the
foliar biomass.

227
Stem-based models (RCD, SD, DBH) predicted with the greatest accuracy the stem
or total above ground biomass, and displayed poorer performance when estimating
the branch or foliar biomass. This may occur when the stem biomass, which is likely
to be highly correlated with the stem diameter, is the largest constituent of the total
biomass. That it was not the case in E. berteroana may be due to the fact that E.
berteroana had been grown from 1 m stakes, and this may have resulted in a
significantly different growth form. Manipulation of a tree via pollarding will alter both
the total biomass and the relative contribution to the total biomass of components.
Yet, pruning and pollarding interventions may not impact on measurements of stem
diameter (Kuyah et al., 2013).
Crown diameter and crown volume were strongly correlated with one another and
both displayed the greatest fit as a single predictors of the total biomass of E.
berteroana. It was hypothesised that measurements of the crown of the tree would
be the best predictor of total foliar biomass. For E. berteroana, crown volume and
crown diameter both showed marginally improved fit over the stem volume
measurements for the prediction of total biomass. This could be due to the way this
tree produces foliage, it does not rely on heavy proliferation of bifurcate branching
systems, and instead the leaves are generated from principal branches and directly
from the trunk (Nygren et al., 1993). As the measurement of the crown was taken
where 5% of the foliage began, and the foliage was observed to occur along the
entire trunk of E. berteroana, it is possible that the cylindrical measurement of crown

228
volume was able to capture the majority of the woody biomass in unison with the
foliage. This result highlights the need to consider the morphology of the tree when
choosing which elements of the tree should be measured to develop allometric
models. Where the tree has a high proportion of woody to foliar biomass, a
measurement of the trunk combined with some reference to the branching
architecture may be appropriate. Further work is needed to develop models that
consider different branching architectures of individual species.
Future studies seeking to develop transferrable estimations of biomass can include a
wide range of trees of different sizes, positioning within the stand and age (Lott et al.,
2000). Allometric equations may vary due to site specific agro ecological, climatic
and edaphic conditions (Yuen et al., 2016), altitude (Cienciala et al., 2006),
provenance (Poorter et al., 2012), input of fertiliser and the use of clones (Heinsoo et
al., 2002), spacing and stand density (Telenius and Verwijst, 1995), the composition
of mixed tree plantations (Laclau et al., 2008), the frequency and timing of biomass
removal (Oppong et al., 2002), and the age of the trees in the sample (Peichl and
Arain, 2007). Practitioners must be aware that these variables may limit the
transferability of allometric equations between sites, and therefore a process of
verification is suggested ( (Eggleston et al., 2006; Yuen et al., 2016).
The total and stem biomass of these species can be reliably predicted by the
presented equations, even when pollarding has manipulated the form of the tree.
However, the present study does not account for repeat pollarding or the temporal

229
effects of season on the trees. The log-linear transformation improved the predictive
power of allometric equations for all biomass components of all species. The rapid
accumulation of biomass of these species implies that the integration of trees into
cattle pastures in the Amazon could represent a major contribution to above ground
C storage. Further work is needed to understand the effects of management on
these species, and the models presented in this study are specific to the species and
conditions in the study area, and care should be taken when extrapolating the
results, which were developed on a homogenous group of small trees.
References
Akaike, H., 1974. A New Look at the Statistical Model Identification.
Springer, New York, NY, pp. 215–222.
https://doi.org/10.1007/978-1-4612-1694-0_16
Arlot, S., Celisse, A., 2010. A survey of cross-validation procedures
for model selection. Statistics Surveys 4, 40–79.
https://doi.org/10.1214/09-SS054
Balehegn, M., Eniang, E.A., Hassen, A., 2012. Estimation of browse
biomass of Ficus thonningii, an indigenous multipurpose fodder tree in
northern Ethiopia. African Journal of Range & Forage Science.
https://doi.org/10.2989/10220119.2012.687071

230
Blum, M., Lövbrand, E., 2019. The return of carbon offsetting? The
discursive legitimation of new market arrangements in the Paris
climate regime. Earth System Governance 100028.
https://doi.org/10.1016/J.ESG.2019.100028
Chave, J., Andalo, C., Brown, S., Cairns, M.A., Chambers, J.Q., Eamus,
D., Fölster, H., Fromard, F., Higuchi, N., Kira, T., Lescure, J.-P.,
Nelson, B.W., Ogawa, H., Puig, H., Riéra, B., Yamakura, T., 2005. Tree
allometry and improved estimation of carbon stocks and balance in
tropical forests. Oecologia 145, 87–99.
https://doi.org/10.1007/s00442-005-0100-x
Chave, J., Réjou-Méchain, M., Búrquez, A., Chidumayo, E., Colgan,
M.S., Delitti, W.B.C., Duque, A., Eid, T., Fearnside, P.M., Goodman,
R.C., Henry, M., Martínez-Yrízar, A., Mugasha, W.A., Muller-Landau,
H.C., Mencuccini, M., Nelson, B.W., Ngomanda, A., Nogueira, E.M.,
Ortiz-Malavassi, E., Pélissier, R., Ploton, P., Ryan, C.M.,
Saldarriaga, J.G., Vieilledent, G., 2014. Improved allometric models
to estimate the aboveground biomass of tropical trees. Global Change
Biology 20, 3177–3190. https://doi.org/10.1111/gcb.12629
Cienciala, E., Černý, M., Tatarinov, F., Apltauer, J., Exnerová, Z.,

231
2006. Biomass functions applicable to Scots pine. Trees - Structure
and Function 20, 483–495. https://doi.org/10.1007/s00468-006-0064-4
Claassen, R., Cattaneo, A., Johansson, R., 2008. Cost-effective design
of agri-environmental payment programs: U.S. experience in theory and
practice. Ecological Economics 65, 737–752.
https://doi.org/10.1016/J.ECOLECON.2007.07.032
Corbera, E., Soberanis, C.G., Brown, K., 2009. Institutional
dimensions of Payments for Ecosystem Services: An analysis of Mexico’s
carbon forestry programme. Ecological Economics 68, 743–761.
https://doi.org/10.1016/J.ECOLECON.2008.06.008
Dutcă, I., 2018. Biomass data for young, planted Norway spruce (Picea
abies (L.) Karst.) trees in Eastern Carpathians of Romania. Data in
Brief 19, 2384–2392. https://doi.org/10.1016/J.DIB.2018.07.033
Dutcă, I., Mather, R., Blujdea, V.N.B., Ioraș, F., Olari, M., Abrudan,
I.V., 2018. Site-effects on biomass allometric models for early growth
plantations of Norway spruce (Picea abies (L.) Karst.). Biomass and
Bioenergy 116, 8–17. https://doi.org/10.1016/J.BIOMBIOE.2018.05.013

232
Eggleston, S., Buendia, L., Miwa, K., Ngara, T. and Tanabe, K. eds., 2006. 2006
IPCC guidelines for national greenhouse gas inventories (Vol. 5). Hayama, Japan:
Institute for Global Environmental Strategies.
Furnival, G.M., 1961. An Index for Comparing Equations Used In
Constructing Volume Tables. Forest Science 7, 337–341.
https://doi.org/10.1093/forestscience/7.4.337
Gevorkiantz, S.R., Bruce, D., Schumacher, F.X., 1997. Forest
Mensuration. Journal of the American Statistical Association 30, 780.
https://doi.org/10.2307/2277755
Heinsoo, K., Sild, E., Koppel, A., 2002. Estimation of shoot biomass
productivity in Estonian Salix plantations. Forest Ecology and
Management 170, 67–74. https://doi.org/10.1016/S0378-1127(01)00784-8
Hissa, L. de B.V., Aguiar, A.P.D., Camargo, R.R., Lima, L.S. de,
Gollnow, F., Lakes, T., 2019. Regrowing forests contribution to law
compliance and carbon storage in private properties of the Brazilian
Amazon. Land Use Policy 88, 104163.
https://doi.org/10.1016/J.LANDUSEPOL.2019.104163
Huang, S., Price, D., J. Titus, S., 2000. Development of

233
ecoregion-based height–diameter models for white spruce in boreal
forests. Forest Ecology and Management 129, 125–141.
https://doi.org/10.1016/S0378-1127(99)00151-6
Huxley, J.S., 1932. Problems of Relative Growth. Nature 129, 775–777.
https://doi.org/10.1038/129775a0
Jagodziński, A.M., Dyderski, M.K., Gęsikiewicz, K., Horodecki, P.,
Cysewska, A., Wierczyńska, S., Maciejczyk, K., 2018. How do tree stand
parameters affect young Scots pine biomass? – Allometric equations and
biomass conversion and expansion factors. Forest Ecology and
Management 409, 74–83. https://doi.org/10.1016/J.FORECO.2017.11.001
James, G., Witten, D., Hastie, T. and Tibshirani, R., 2013. An introduction to
statistical learning (Vol. 112, p. 18). New York: springer
Kays, J., Canham, C., 1991. Effects of Time and Frequency of Cutting
on Hardwood Root Reserves and Sprout Growth. Forest Science 37,
524–539. https://doi.org/10.1093/forestscience/37.2.524
Kuyah, S., Dietz, J., Muthuri, C., van Noordwijk, M., Neufeldt, H.,
2013. Allometry and partitioning of above- and below-ground biomass in
farmed eucalyptus species dominant in Western Kenyan agricultural

234
landscapes. Biomass and Bioenergy 55, 276–284.
https://doi.org/10.1016/J.BIOMBIOE.2013.02.011
Laclau, J.-P., Bouillet, J.-P., Gonçalves, J.L.M., Silva, E.V.,
Jourdan, C., Cunha, M.C.S., Moreira, M.R., Saint-André, L., Maquère,
V., Nouvellon, Y., Ranger, J., 2008. Mixed-species plantations of
Acacia mangium and Eucalyptus grandis in Brazil: 1. Growth dynamics
and aboveground net primary production. Forest Ecology and Management
255, 3905–3917. https://doi.org/10.1016/J.FORECO.2007.10.049
Lojka, B.., Preininger, D.., Lojkova, J., Banout, J., Polesny, Z.,
2005. Biomass growth and farmer knowledge of Inga edulis in the
Peruvian Amazon. AGRICULTURA TROPICA ET SUBTROPICA 38, 44–51.
López, F., García, M.M., Yánez, R., Tapias, R., Fernández, M., Díaz,
M.J., 2008. Leucaena species valoration for biomass and paper
production in 1 and 2 year harvest. Bioresource Technology 99,
4846–4853. https://doi.org/10.1016/J.BIORTECH.2007.09.048
Lott, J.E., Howard, S.B., Black, C.R., Ong, C.K., 2000. Allometric
estimation of above-ground biomass and leaf area in managed Grevillea
robusta agroforestry systems. Agroforestry Systems 49, 1–15.
https://doi.org/10.1023/A:1006330830109

235
MacCracken, J.G., Ballenberghe, V. Van, 1993. Mass-Diameter
Regressions for Moose Browse on the Copper River Delta, Alaska.
Journal of Range Management 46, 302. https://doi.org/10.2307/4002462
Martin, A.R., Thomas, S.C., 2011. A reassessment of carbon content in tropical
trees. PLoS ONE 6. https://doi.org/10.1371/journal.pone.0023533
Mascaro, J., Litton, C.M., Hughes, R.F., Uowolo, A., Schnitzer, S.A.,
2014. Is logarithmic transformation necessary in allometry? Ten,
one-hundred, one-thousand-times yes. Biological Journal of the Linnean
Society 111, 230–233. https://doi.org/10.1111/bij.12177
Mauricio, R.M., Ribeiro, R.S., Paciullo, D.S.C., Cangussú, M.A.,
Murgueitio, E., Chará, J., Estrada, M.X.F., 2019. Silvopastoral
Systems in Latin America for Biodiversity, Environmental, and
Socioeconomic Improvements, in: Agroecosystem Diversity. Elsevier, pp.
287–297. https://doi.org/10.1016/B978-0-12-811050-8.00018-2
MEA, 2005. Millenium Ecosystem Assessment: Ecosystems and Human Well-Being
(Volume 4). Washington, DC. Island Press.
Montagnini, F., Nair, P.K.R., 2004. Carbon sequestration: An

236
underexploited environmental benefit of agroforestry systems.
Springer, Dordrecht, pp. 281–295.
https://doi.org/10.1007/978-94-017-2424-1_20
Nahed-Toral, J., Valdivieso-Pérez, A., Aguilar-Jiménez, R.,
Cámara-Cordova, J., Grande-Cano, D., 2013. Silvopastoral systems with
traditional management in Southeastern Mexico: a prototype of
livestock agroforestry for cleaner production. Journal of Cleaner
Production 57, 266–279. https://doi.org/10.1016/j.jclepro.2013.06.020
Nygren, P., Rebottaro, S., Chavarra, R., 1993. Application of the
pipe model theory to non-destructive estimation of leaf biomass and
leaf area of pruned agroforestry trees. Agroforestry Systems 23,
63–77. https://doi.org/10.1007/BF00704851
Oppong, S.K., Kemp, P.D., Douglas, G.B., Foote, A.G., 2002. Effects of
season and frequency of harvest on browse yield and root carbohydrate
reserves of willows (Salix spp.) in New Zealand. New Zealand Journal
of Agricultural Research 45, 87–95.
https://doi.org/10.1080/00288233.2002.9513497
Packard, G.C., 2017a. Misconceptions about logarithmic transformation
and the traditional allometric method. Zoology 123, 115–120.

237
https://doi.org/10.1016/J.ZOOL.2017.07.005
Packard, G.C., 2017b. Why allometric variation in mammalian metabolism
is curvilinear on the logarithmic scale. Journal of Experimental
Zoology Part A: Ecological and Integrative Physiology 327, 537–541.
https://doi.org/10.1002/jez.2129
Packard, G.C., 2014. Multiplicative by nature: Logarithmic
transformation in allometry. Journal of Experimental Zoology Part B:
Molecular and Developmental Evolution 322, 202–207.
https://doi.org/10.1002/jez.b.22570
Pagiola, S., Ramírez, E., Gobbi, J., de Haan, C., Ibrahim, M.,
Murgueitio, E., Ruíz, J.P., 2007. Paying for the environmental
services of silvopastoral practices in Nicaragua. Ecological Economics
64, 374–385. https://doi.org/10.1016/j.ecolecon.2007.04.014
Parang, K., Wiebe, L., Knaus, E., 2012. Springer Texts in Statistics
An Introduction To Statistical Learning. Current Medicinal Chemistry
7, 995–1039. https://doi.org/10.2174/0929867003374372
Peichl, M., Arain, M.A., 2007. Allometry and partitioning of above-
and belowground tree biomass in an age-sequence of white pine forests.

238
Forest Ecology and Management 253, 68–80.
https://doi.org/10.1016/J.FORECO.2007.07.003
Penman, J., Gytarsky, M., Hiraishi, T., Krug, T., Kruger, D., Pipatti,
R., Buendia, L., Miwa, K., Ngara, T., Tanabe, K., Wagner, F., 2003.
Good practice guidance for land use, land-use change and forestry.
Good practice guidance for land use, land-use change and forestry.
Pezo, D., Kass, M., Benavides, J., Romero, F. and Chaves, C., 1990. Potential of
legume tree fodders as animal feed in Central America. In Shrubs and tree fodders
for farm animals: proceedings of a workshop in Denpasar, Indonesia, 24-29 July
1989. IDRC, Ottawa, ON, CA.
Picard, N., Saint-André, L. and Henry, M., 2012. Manual for building tree volume and
biomass allometric equations: from field measurement to prediction. Manual for
building tree volume and biomass allometric equations: from field measurement to
prediction, FAO; Food and Agricultural Organization of the United Nations (2012).
Poorter, H., Niklas, K.J., Reich, P.B., Oleksyn, J., Poot, P., Mommer,
L., 2012. Biomass allocation to leaves, stems and roots: meta-analyses
of interspecific variation and environmental control. New Phytologist
193, 30–50. https://doi.org/10.1111/j.1469-8137.2011.03952.x

239
Prasad, J.V.N.S., Korwar, G.R., Rao, K.V., Mandal, U.K., G.R.Rao,
Srinivas, I., Venkateswarlu, B., Rao, S.N., Kulkarni, H.D., 2011.
Optimum stand density of Leucaena leucocephala for wood production in
Andhra Pradesh, Southern India. Biomass and Bioenergy 35, 227–235.
https://doi.org/10.1016/J.BIOMBIOE.2010.08.012
Raitio, H., Paukkonen, K., Kauppi, A., 1994. Effects of defoliation,
nitrogen nutrition, and temperature on leafing and root carbohydrates
of birch seedlings. Canadian Journal of Forest Research 24, 1914–1920.
https://doi.org/10.1139/x94-246
Rubilar, R.A., Allen, H.L., Alvarez, J.S., Albaugh, T.J., Fox, T.R.,
Stape, J.L., 2010. Silvicultural manipulation and site effect on above
and belowground biomass equations for young Pinus radiata. Biomass and
Bioenergy 34, 1825–1837.
https://doi.org/10.1016/J.BIOMBIOE.2010.07.015
Serrão, E.A.S., Nepstad, D., Walker, R., 1996. Upland agricultural and
forestry development in the Amazon: sustainability, criticality and
resilience. Ecological Economics 18, 3–13.
https://doi.org/10.1016/0921-8009(95)00092-5
Sharma, E., Ambasht, R.S., 1991. Biomass, productivity and energetics

240
in himalayan alder plantations. Annals of Botany 67, 285–293.
https://doi.org/10.1093/oxfordjournals.aob.a088138
Sharrow, S.H., Ismail, S., 2004. Carbon and nitrogen storage in
agroforests, tree plantations, and pastures in western Oregon, USA.
Agroforestry Systems 60, 123–130.
https://doi.org/10.1023/B:AGFO.0000013267.87896.41
Shimamoto, C.Y., Botosso, P.C., Marques, M.C.M., 2014. How much carbon
is sequestered during the restoration of tropical forests? Estimates
from tree species in the Brazilian Atlantic forest. Forest Ecology and
Management 329, 1–9. https://doi.org/10.1016/J.FORECO.2014.06.002
Shinozaki, K., Yoda, K., Hozumi, K., Kira, T., 1964. A Quantative
Analysis of plant form - The Pipe Model Theory: Basic Analyses.
Japanese Journal Of Ecology 14, 97–105.
https://doi.org/10.18960/seitai.14.3_97
Sileshi, G.W., 2014. A critical review of forest biomass estimation
models, common mistakes and corrective measures. Forest Ecology and
Management 329, 237–254. https://doi.org/10.1016/J.FORECO.2014.06.026
Smit, C., Ruifrok, J.L., van Klink, R., Olff, H., 2015. Rewilding with

241
large herbivores: The importance of grazing refuges for sapling
establishment and wood-pasture formation. Biological Conservation 182,
134–142. https://doi.org/10.1016/J.BIOCON.2014.11.047
Steinfeld, H., Food and Agriculture Organization of the United
Nations., Livestock, E. and D. (Firm), 2006. Livestock’s long shadow :
environmental issues and options. Food and Agriculture Organization of
the United Nations.
Szott, L.T., Palm, C.A., Davey, C.B., 1994. Biomass and litter
accumulation under managed and natural tropical fallows. Forest
Ecology and Management 67, 177–190.
https://doi.org/10.1016/0378-1127(94)90015-9
Takimoto, A., Nair, P.K.R., Nair, V.D., 2008. Carbon stock and
sequestration potential of traditional and improved agroforestry
systems in the West African Sahel. Agriculture, Ecosystems &
Environment 125, 159–166. https://doi.org/10.1016/j.agee.2007.12.010
Telenius, B., Verwijst, T., 1995. The influence of allometric
variation, vertical biomass distribution and sampling procedure on
biomass estimates in commercial short-rotation forests. Bioresource
Technology 51, 247–253. https://doi.org/10.1016/0960-8524(94)00133-L

242
Tzilivakis, J., Warner, D.J., Holland, J.M., 2019. Developing
practical techniques for quantitative assessment of ecosystem services
on farmland. Ecological Indicators 106, 105514.
https://doi.org/10.1016/J.ECOLIND.2019.105514
Yuen, J.Q., Fung, T., Ziegler, A.D., 2016. Review of allometric
equations for major land covers in SE Asia: Uncertainty and
implications for above- and below-ground carbon estimates. Forest
Ecology and Management 360, 323–340.
https://doi.org/10.1016/J.FORECO.2015.09.016

243
Appendices
Appendix 1. Table. The wet to dry relationship for biomass components of subsamples.taken from Inga edulis, Erythrina berteroana and Leucaena leucocephala, where R2 is the coefficient of determination, RSE is the residual square error and n is the sample size.
Species Biomass component Intercept Slope R2 RSE n
I. edulis Trunk 0.629 2.073 0.992 1.6 118
Branches -21.642 2.470 0.985 180.9 118
Leaf 7.478 2.679 0.997 49.5 118 E. berteroana Trunk 2.054 2.674 0.949 4.9 117
Branches 62.000 3.112 0.989 126.0 117
Leaf -0.480 4.918 0.966 62.1 117 L. leucocephala Trunk -0.624 1.795 0.996 1.1 115
Branches -24.161 2.246 0.998 51.2 115
Leaf 1.675 2.870 0.999 19.4 115

244
Appendix 2. Figure. Total biomass (g) and root collar diameter (mm) of Inga edulis plotted using the linear regression line (y = aX + b) showing clockwise from left (a) scatter plot and linear regression line, (b) departure from normal Q-Q plot and (c) residuals vs. leverage
20 40 60 80 100 120
01
00
02
00
03
00
04
00
05
00
06
00
0
RCD
TO
TA
LB
IOM
AS
S
−2 −1 0 1 2
−2
−1
01
23
Theoretical Quantiles
Sta
nd
ard
ized
re
sid
ua
ls
lm(TOTALBIOMASS ~ RCD)
Normal Q−Q
2
34
11
0.00 0.02 0.04 0.06 0.08 0.10
−3
−2
−1
01
23
4
Leverage
Sta
nd
ard
ized
re
sid
ua
ls
lm(TOTALBIOMASS ~ RCD)
Cook's distance 0.5
0.5
1Residuals vs Leverage
11
2
37

245
Appendix 3. Figure. Relationship between total biomass (g) and root collar diameter (mm) for Inga edulis plotted using the quadratic regression model (equation y = ax2 + bx + c) showing clockwise from left (a) scatter plot and polynomial regression line, (b) departure from normal Q-Q plot and (c) residuals vs. leverage
20 40 60 80 100 120
01
00
02
00
03
00
04
00
05
00
06
00
0
RCD
TO
TA
LB
IOM
AS
S
−2 −1 0 1 2
−2
−1
01
23
4
Theoretical Quantiles
Sta
nd
ard
ized
re
sid
ua
ls
lm(TOTALBIOMASS ~ poly(RCD, 2))
Normal Q−Q
2
34
11
0.0 0.1 0.2 0.3 0.4
−3
−2
−1
01
23
4
Leverage
Sta
nd
ard
ized
re
sid
ua
ls
lm(TOTALBIOMASS ~ poly(RCD, 2))
Cook's distance
1
0.5
0.5
1
Residuals vs Leverage
11
2
76

246
Appendix 4. Figure. Relationship between total biomass (g) and root collar diameter (mm) for I. edulis plotted using the log-linear transformation ln(y) = a + b * lnx showing clockwise from left (a) scatter plot and regression line, (b) departure from normal Q-Q plot and (c) residuals vs. leverage
2.5 3.0 3.5 4.0 4.5
67
8
LNRCD
LN
TO
TA
LB
IOM
AS
S
−2 −1 0 1 2
−2
−1
01
23
Theoretical Quantiles
Sta
nd
ard
ized
re
sid
ua
ls
lm(log(TOTALBIOMASS) ~ log(RCD))
Normal Q−Q
77
79
37
0.00 0.05 0.10 0.15 0.20
−2
−1
01
23
Leverage
Sta
nd
ard
ized
re
sid
ua
ls
lm(log(TOTALBIOMASS) ~ log(RCD))
Cook's distance
1
0.5
0.5
1
Residuals vs Leverage
77
76
79

247
Appendix 5. Table. Allometric equations following the linear model for biomass components of 0.5 x 7.5 m alley rows of Erythrina berteroana 6 months post pollarding to a height of 2 m. Where (x) is the measurement used as a predictor of biomass: Root collar diameter (RCD mm, 0.05 m), diameter at breast height (DBH (mm), 1.3 m), stump diameter (SD mm, 0.3 m), crown volume (CV cm3) and crown diameter (CD cm), (y) is the log transformation of total biomass(g), trunk biomass (g), branch biomass (g) or leaf biomass (g). b0 as the intercept coefficient, b1 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), root mean square error of model (Model RMSE, which is the same as Furnival’s Index for untransformed data) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
(x) (y) Biomass b0 (S.E.) P b1 (S.E.) P R2 MODEL
RMSE (F.I.) CV RMSE
RCD Total -698.4(265.4)0.01 32.2(4.0)<0.01 0.48 748.75 669.41
Trunk -335.3(234.2)0.2 21.6(3.5)<0.01 0.34 659.16 575.19
Branch -322.8(91.8)<0.01 9.3(1.4)<0.01 0.39 249.39 224.39
Leaf -45.6(15.8)<0.01 1.4(0.2)<0.01 0.33 43.09 41.24
DBH Total -136.6(303.3)0.7 71.9(13.4)<0.01 0.31 814.15 777.36
Trunk -118.3(241.2)0.6 56.1(10.6)<0.01 0.3 647.29 592.11
Branch 9.0(113.0)0.9 12.5(5.0)0.01 0.09 303.41 276.70
Leaf -27.3(16.8)0.1 3.4(0.7)<0.01 0.24 45.20 43.41
SD Total -221.7(305.0)0.5 32.4(5.7)<0.01 0.33 802.47 737.63
Trunk -236.7(237.9)0.3 26.3(4.5)<0.01 0.34 625.86 575.16
Branch 46.7(116.9)0.7 4.6(2.2)<0.05 0.06 307.62 268.01
Leaf -31.7(17.0)0.07 1.5(0.3)<0.01 0.26 44.67 42.56
CV Total 707.7(115.1)<0.01 0.0(0.0)<0.01 0.53 668.19 626.49
Trunk 637.8(107.3)<0.01 0.0(0.0)<0.01 0.35 623.07 569.37
Branch 68.6(40.7)0.1 0.0(0.0)<0.01 0.45 236.39 228.02
Leaf 1.3(4.6)0.8 0.0(0.0)<0.01 0.74 26.45 26.02
CD Total -144.2(217.0)0.5 16.7(2.1)<0.01 0.48 704.99 639.73
Trunk 127.4(199.9)0.5 10.3(2.0)<0.01 0.29 649.50 586.90
Branch -227.8(70.5)<0.01 5.4(0.7)<0.01 0.48 229.04 212.20
Leaf -43.8(10.6)<0.01 1.0(0.1)<0.01 0.56 34.39 32.69

248
Appendix 6. Table. Allometric equations following the linear model for biomass components of 0.5 x 7.5 m alley rows of Erythrina berteroana 6 months post pollarding to a height of 2 m. Where (x) is the root collar diameter (RCD mm, 0.05 m), or RCD and Height (cm) as predictors of biomass. (y) is the log transformation of total biomass (g), trunk biomass (g), branch biomass (g) or leaf biomass (g), b0 as the intercept coefficient, b1 and b2 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), root mean square error of model (Model RMSE, which is the same as Furnival’s Index F.I. for untransformed data) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
(x) (y)
Biomass b0 (S.E.) P b1 (S.E.) P b2 (S.E.) P R2 MODEL
RMSE (F.I.) CV RMSE
RCD + HEIGHT Total -2,138.7(464.9)<0.01 25.2(4.5)<0.01 8.4(2.1)<0.01 0.51 670.39 625.55
Trunk -1,485.3(413.3)<0.01 14.8(4.0)<0.01 7.1(1.9)<0.01 0.39 595.98 554.43
Branch -513.8(171.6)<0.01 9.3(1.7)<0.01 0.8(0.8)0.3 0.37 247.40 225.83
Leaf -139.6(26.3)<0.01 1.1(0.3)<0.01 0.5(0.1)<0.01 0.45 37.99 38.66

249
Appendix 7. Table. Allometric equations following the linear model for biomass components of 0.5 x 7.5 m alley rows of Inga edulis 6 months post pollarding to a height of 2 m. Where (x) is the measurement used a predictor of biomass: Root collar diameter (RCD mm, 0.05 m), diameter at breast height (DBH mm, 1.3 m), stump diameter (SD mm, 0.3 m), crown volume (CV cm3) and crown diameter (CD cm), (y) is the log transformation of total biomass (g), trunk biomass (g), branch biomass (g) or leaf biomass (g), b0 as the intercept coefficient, b1 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), root mean square error of model (Model RMSE, which is the same as Furnival’s Index for untransformed data) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
(x) (y)
Biomass b0 (S.E.) P b1 (S.E.) P R2 MODEL
RMSE (F.I.) CV
RMSE
RCD Total -931.6(332.5)<0.01 47.7(4.8)<0.01 0.56 956.09 954.3
Trunk -415.5(185.5)0.03 26.6(2.7)<0.01 0.56 533.50 510.3
Branch -351.0(168.8)0.04 13.8(2.4)<0.01 0.29 485.25 474.5
Leaf -165.1(98.8)0.1 7.3(1.4)<0.01 0.25 283.99 279.8
DBH Total 1,100.2(398.9)<0.01 31.9(10.7)<0.01 0.1 1360.22 1348.4
Trunk 662.7(220.1)<0.01 19.3(5.9)<0.01 0.12 750.59 756.8
Branch 216.0(163.7)0.2 9.8(4.4)0.03 0.06 558.17 552.4
Leaf 221.4(95.5)0.02 2.7(2.6)0.3 0.01 325.59 317.5
SD Total -1,054.7(345.0)<0.01 64.3(6.5)<0.01 0.55 957.32 957
Trunk -571.1(180.4)<0.01 37.6(3.4)<0.01 0.61 500.51 484.4
Branch -380.8(175.4)0.03 18.5(3.3)<0.01 0.29 486.68 473.4
Leaf -102.8(107.3)0.34 8.3(2.0)<0.01 0.17 297.86 291.3
CV Total 1.529e+03 (1.940e+02)<0.01 2.162e-04 (4.332e-05)<0.01 0.24 1250.69 1257.4
Trunk 1.001e+03 (1.120e+02)<0.01 1.057e-04 (2.502e-05)<0.01 0.18 722.35 733.2
Branch 3.357e+02 (8.180e+01)<0.01 7.102e-05( 1.827e-05)<0.01 0.16 527.37 525.7
Leaf 1.923e+02 (4.679e+01)<0.01 3.951e-05(1.045e-05)<0.01 0.15 301.68 293.2
CD Total 600.5(350.5)0.09 10.7(2.2)<0.01 0.24 1251.90 1291.8
Trunk 553.2(202.7)<0.01 5.2(1.3)<0.01 0.18 724.05 750.8
Branch 26.4(147.5)0.9 3.6(0.9)<0.01 0.16 526.87 534.5
Leaf 20.9(84.4)0.9 2.0(0.5)<0.01 0.15 301.56 303.5

250
Appendix 8. Table. Allometric equations following the linear model for biomass components of 0.5 x 7.5 m alley rows of Inga edulis 6 months post pollarding to a height of 2 m. Where (x) is the root collar diameter (RCD mm, 0.05 m), or RCD and Height (cm) as predictors of biomass, (y) is the log transformation of total biomass (g), trunk biomass (g), branch biomass (g) or leaf biomass (g). b0 as the intercept coefficient, b1 and b2
as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), root mean square error of model (Model RMSE, which is the same as Furnival’s Index F.I. for untransformed data) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
(x) (y) Biomass b0 (S.E.) P b1 (S.E.) P b2 (S.E.) P R2 MODEL
RMSE (F.I.) CV
RMSE
RCD + HEIGHT Total -2,139.6(531.7)<0.01 41.9(5.0)<0.01 6.5(2.3)<0.01 0.6 910.26 922.6
Trunk -996.0(300.6)<0.01 23.8(2.8)<0.01 3.1(1.3)0.02 0.59 514.65 506.3
Branch -718.5(278.7)0.01 12.1(2.6)<0.01 2.0(1.2)0.1 0.31 477.03 473.3
Leaf -425.1(161.8)0.01 6.1(1.5)<0.01 1.4(0.7)<0.05 0.29 276.93 276.9

251
Appendix 9. Table. Allometric equations following the linear model for biomass components of 0.5 x 7.5 m alley rows of Leucaena leucocephala 6 months post pollarding to a height of 2 m. Where (x) is the measurement used as predictor of biomass: Root collar diameter (RCD mm, 0.05 m), diameter at breast height (DBH mm, 1.3 m), stump diameter (SD mm, 0.3 m), crown volume (CV cm3) and crown diameter (CD cm), (y) is the log transformation of total biomass (g), trunk biomass (g), branch biomass (g) or leaf biomass (g), b0 as the intercept coefficient, b1 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), root mean square error of model (Model RMSE, which is the same as Furnival’s Index for untransformed data) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
(x) (y) Biomass b0 (S.E.) P b1 (S.E.) P R2 MODEL RMSE (F.I.) CV RMSE
RCD Total 774.8(146.0)<0.01 46.3(3.2)<0.01 0.78 320.3 311.1
Trunk -685.9(95.9)<0.01 39.6(2.1)<0.01 0.85 210.4 198.9
Branch -55.7(94.4)0.6 5.2(2.1)0.02 0.09 207.1 178.2
Leaf .-33.250(14.4)0.02 1.4(0.3)<0.01 0.26 31.6 29.4
DBH Total 285.7(196.3)0.2 43.8(8.2)<0.01 0.32 558.6 545.8
Trunk 178.3(151.7)0.3 39.5(6.4)<0.01 0.39 431.7 415.2
Branch 109.8(76.0)0.2 2.8(3.2)0.4 0.01 216.2 189.9
Leaf -2.4(12.1)0.9 1.5(0.5)<0.01 0.12 34.4 32.3
SD Total -390.7(190.7)<0.05 47.2(5.2)<0.01 0.57 441.5 411.9
Trunk -447.3(127.1)<0.01 43.0(3.5)<0.01 0.71 294.4 279.8
Branch 76.5(93.1)0.4 2.7(2.5)0.3 0.02 215.6 189.1
Leaf -19.9(14.4)0.2 1.4(0.4)<0.01 0.18 33.2 30.5
CV Total 953.2(95.3)<0.01 0.0(0.0)<0.01 0.28 571.9 562.4
Trunk 837.7(81.2)<0.01 0.0(0.0)<0.01 0.22 487.6 471.2
Branch 104.9(33.8)<0.01 0.0(0.0)<0.01 0.13 202.6 177.2
Leaf 10.6(4.7)0.03 0.0(0.0)<0.01 0.40 28.4 25.7
CD Total 282.8(149.0)0.06 10.0(1.4)<0.01 0.46 496.0 474.3
Trunk 424.9(139.5)<0.01 6.5(1.3)<0.01 0.29 464.6 439.1
Branch -115.9(51.2)0.03 3.0(0.5)<0.01 0.39 170.3 161.7
Leaf -26.2(7.6)<0.01 0.6(0.1)<0.01 0.52 25.3 23.1

252
Appendix 10. Table. Allometric equations following the linear model for biomass components of 0.5 x 7.5 m alley rows of Leucaena leucocephala 6 months post pollarding to a height of 2 m. Where (x) is the root collar diameter (RCD mm, 0.05 m), or RCD and Height (cm) as predictors of biomass, (y) is the log transformation of total biomass (g), trunk biomass (g), branch biomass (g) or leaf biomass (g). b0 as the intercept coefficient, b1 and b2 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), root mean square error of model (Model RMSE, which is the same as Furnival’s Index F.I. for untransformed data) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
(x) (y) Biomass b0 (S.E.) P b1 (S.E.) P b2 (S.E.) P R2 MODEL
RMSE (F.I.) CV
RMSE
RCD + HEIGHT Total -1,209.0(188.1)<0.01 41.7(3.3)<0.01 2.2(0.7)<0.01 0.81 296.9 283.9
Trunk -908.3(127.9)<0.01 37.3(2.2)<0.01 1.1(0.4)0.02 0.87 201.9 192.7
Branch -215.4(129.0)0.1 3.5(2.2)0.1 0.8(0.4)0.08 0.14 203.5 170.9
Leaf -85.2(17.8)<0.01 0.9(0.3)<0.01 0.3(0.1)<0.01 0.43 28.0 25.5

253
Appendix 11. Table. Allometric equations following a quadratic model for biomass components of 0.5 x 7.5 m alley rows of Erythrina berteroana 6 months post pollarding to a height of 2 m. Where (x) is the measurement used as a predictor of biomass: Root collar diameter (RCD mm, 0.05 m), diameter at breast height (DBH mm, 1.3 m), stump diameter (SD mm, 0.3 m), crown volume (CV, cm3) and crown diameter (CD cm), (y) is the log transformation of total biomass (g), trunk biomass (g), branch biomass (g) or leaf biomass (g), b0 as the intercept coefficient, b1 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), root mean square error of model (Model RMSE, which is the same as Furnival’s Index for untransformed data) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
(x) (y) Biomass b0 (S.E.) P b1 (S.E.) P b2 (S.E.) P R2 MODEL RMSE
(F.I.) CV RMSE
RCD Total 1,405.2(15.1)<0.01 5,173.0(6.8)<0.01 158.8(0.2)0.8 0.39 748.5 726.3
Trunk 1,084.3(13.3)<0.01 3,315.9(4.9)<0.01 -251.4(-0.4)0.7 0.25 658.5 638.0
Branch 276.1(9.1)<0.01 1,620.2(6.5)<0.01 414.5(1.7)0.1 0.39 244.3 215.1
Leaf 44.9(8.4)<0.01 236.9(5.4)<0.01 -4.3(-0.1)0.9 0.29 43.1 44.5
DBH Total 1,405.2(14.2)<0.01 4,450.7(5.5)<0.01 -1,409.8(-1.7)0.09 0.32 796.0 765.4
Trunk 1,084.3(13.6)<0.01 3,471.3(5.3)<0.01 -449.7(-0.7)0.5 0.28 645.0 600.0
Branch 276.1(7.8)<0.01 771.1(2.6)0.01 -859.1(-2.9)<0.01 0.17 285.0 260.4
Leaf 44.9(8.3)<0.01 208.4(4.7)<0.01 -101.0(-2.3)0.03 0.27 43.5 42.1
SD Total 1,405.2(14.1)<0.01 4,592.7(5.6)<0.01 -245.9(-0.3)0.8 0.31 801.9 771.5
Trunk 1,084.3(14.0)<0.01 3,729.0(5.8)<0.01 124.7(0.2)0.8 0.32 625.7 604.8
Branch 276.1(7.3)<0.01 647.6(2.1)0.04 -403.7(-1.3)0.2 0.06 303.7 272.0
Leaf 44.9(8.1)<0.01 216.1(4.7)<0.01 33.1(0.7)0.5 0.26 44.5 45.5
CV Total 1,405.2(17.0)<0.01 5,875.5(8.6)<0.01 -124.6(-0.2)0.9 0.52 668.0 645.6
Trunk 1,084.2(14.1)<0.01 3,760.6(5.9)<0.01 -294.5(-0.5)0.6 0.33 622.0 574.8
Branch 276.1(9.4)<0.01 1,747.8(7.2)<0.01 97.5(0.4)0.7 0.43 236.1 246.7
Leaf 44.9(14.5)<0.01 367.1(14.4)<0.01 72.5(2.8)<0.01 0.76 24.9 24.8

254
Appendix 12. Table. Allometric equations following the quadratic model for biomass components of 0.5 x 7.5 m alley rows of Erythrina berteroana 6 months post pollarding to a height of 2 m. Where (x) is the root collar diameter (RCD mm, 0.05 m), or RCD and Height (cm) as predictors of biomass, (y) is the log transformation of total biomass (g), trunk biomass (g), branch biomass (g) or leaf biomass (g). b0 as the intercept coefficient, b1 and b2 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), root mean square error of model (Model RMSE, which is the same as Furnival’s Index F.I. for untransformed data) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
(x) (y)
Biomass b0 (S.E.) P b1 (S.E.) P b2 (S.E.) P b3 (S.E.) P b4 (S.E.) P R2
MODEL RMSE (F.I.)
CV RMSE
RCD +
HEIGHT Total 522.7(0.8)0.4 276.1(9.3)<0.01 1,468.6(5.5)<0.01 496.5(2.0)0.05 373.8(1.4)0.2 0.52 653.8 638.5
Trunk 366.8(1.5)0.1 44.9(11.0)<0.01 160.9(4.5)<0.01 35.8(1.1)0.3 189.4(5.2)<0.01 0.37 592.4 572.9
Branch 164.3(4.9)<0.01 0.20(14.52)<0.01 0.35(3.12)<0.01 0.02(0.18)0.9 0.16(11.75)<0.01 0.41 236.8 220.5
Leaf 0.19(1.71).0.9 -0.01(-0.11)0.9 0.04(10.44)<0.01 0.14(4.95)<0.01 0.03(1.20)0.2 0.59 32.3 34.7

255
Appendix 13. Table. Allometric equations following a quadratic model for biomass components of 0.5 x 7.5 m alley rows of Inga edulis 6 months post pollarding to a height of 2 m. Where (x) is the measurement used as a predictor of biomass: Root collar diameter (RCD mm, 0.05 m), diameter at breast height (DBH mm, 1.3 m), stump diameter (SD mm, 0.3 m), crown volume (CV, cm3) and crown diameter (CD cm), (y) is the log transformation of total biomass (g), trunk biomass (g), branch biomass (g) or leaf biomass (g), b0 as the intercept coefficient, b1 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), root mean square error of model (Model RMSE, which is the same as Furnival’s Index for untransformed data) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
(x) (y) Biomass b0 (S.E.) P b1 (S.E.) P b2 (S.E.) P R2 MODEL RMSE
(F.I.) CV RMSE
RCD Total 2,195.7(107.9)<0.01 9,623.1(971.1)<0.01 -699.9(971.1)0.5 0.55 952.9 981.9
Trunk 1,327.0(60.3)<0.01 5,362.0(542.9)<0.01 -253.7(542.9)0.6 0.54 493.4 529.1
Branch 554.6(54.8)<0.01 2,786.7(493.0)<0.01 -339.3(493.0)0.5 0.28 462.7 482.1
Leaf 314.1(32.1)<0.01 1,474.4(289.2)<0.01 -106.8(289.2)0.7 0.23 275.4 288.2
DBH Total 2,195.7(137.9)<0.01 4,096.6(1,241.4)<0.01 5,446.4(1,241.4)<0.01 0.26 1218.2 1,208.0
Trunk 1,327.0(73.1)<0.01 2,484.0(657.8)<0.01 -3,447.2(657.8)<0.01 0.33 645.5 634.7
Branch 554.6(60.5)<0.01 1,266.1(544.1)0.02 -1,464.4(544.1)<0.01 0.12 533.9 543.5
Leaf 314.1(36.3)<0.01 346.5(326.2)0.3 -534.8(326.2)0.1 0.02 320.1 318.2
SD Total 2,195.7(105.6)<0.01 9,613.2(950.1)<0.01 1,954.8(950.1)0.04 0.57 932.4 938.2
Trunk 1,327.0(55.6)<0.01 5,613.7(500.0)<0.01 889.5(500.0)0.08 0.61 490.7 487.0
Branch 554.6(54.3)<0.01 2,766.4(489.0)<0.01 731.3(489.0)0.1 0.29 479.9 467.0
Leaf 314.1(33.5)<0.01 1,233.1(301.2)<0.01 334.0(301.2)0.3 0.17 295.5 296.2
CV Total 2,195.7(114.8)<0.01 6,320.2(1,033.4)<0.01 6,588.3(1,033.4)<0.01 0.49 1014.1 1,050.5
Trunk 1,327.0(70.6)<0.01 3,088.7(635.5)<0.01 -3,280.8(635.5)<0.01 0.38 623.6 634.7
Branch 554.6(54.0)<0.01 2,076.3(485.6)<0.01 -2,033.5(485.6)<0.01 0.30 476.5 484.8
Leaf 314.1(30.2)<0.01 1,155.2(271.5)<0.01 -1,274.1(271.5)<0.01 0.32 266.4 270.9
CD Total 2,195.7(119.5)<0.01 6,300.7(1,075.8)<0.01 6,056.0(1,075.8)<0.01 0.44 1055.7 1,223.3
Trunk 1,327.0(72.6)<0.01 3,056.3(653.6)<0.01 -3,023.2(653.6)<0.01 0.34 641.4 774.8
Branch 554.6(54.1)<0.01 2,086.5(487.0)<0.01 -1,997.3(487.0)<0.01 0.29 477.9 540.2
Leaf 314.1(31.6)<0.01 1,157.9(284.1)<0.01 -1,035.4(284.1)<0.01 0.26 278.8 320.3

256
Appendix 14. Table. Allometric equations following the quadratic model for biomass components of 0.5 x 7.5 m alley rows of Inga edulis 6 months post pollarding to a height of 2 m. Where (x) is the root collar diameter (RCD mm, 0.05 m), or RCD and Height (cm) as predictors of biomass, (y) is the log transformation of total biomass (g), trunk biomass (g), branch biomass (g) or leaf biomass (g). b0 as the intercept coefficient, b1 and b2 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), root mean square error of model (Model RMSE, which is the same as Furnival’s Index F.I. for untransformed data) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
(x) (y)
Biomass b0 (S.E.) P b1 (S.E.) P b2 (S.E.) P b3 (S.E.) P b4 (S.E.) P R2
MODEL RMSE (F.I.)
CV RMSE
RCD +
HEIGHT Total 2,195.7(100.6)<0.01 8,336.5(992.2)<0.01 -853.7(906.6)0.3 2,956.2(991.8)<0.01 2,040.0(906.9)0.03 0.61 877.2 904.6
Trunk 1,327.0(56.6)<0.01 4,729.4(558.0)<0.02 -336.0(509.9)0.5 1,423.9(557.8)0.01 1,286.0(510.1)0.02 0.60 493.4 493.8
Branch 554.6(53.1)<0.01 2,375.0(523.3)<0.03 -396.2(478.2)0.4 912.1(523.2)0.09 978.3(478.4)0.04 0.32 462.7 468.0
Leaf 314.1(31.6)<0.01 1,232.1(311.5)<0.04 -121.5(284.7)0.7 620.2(311.4)0.05 620.2(311.4)0.05 0.26 275.4 299.3

257
Appendix 15. Table. Allometric equations following a quadratic model for biomass components of 0.5 x 7.5 m alley rows of Leucaena leucocephala 6 months post pollarding to a height of 2 m. Where (x) is the measurement used as a predictor of biomass: Root collar diameter (RCD mm, 0.05 m), diameter at breast height (DBH mm, 1.3 m), stump diameter (SD mm, 0.3 m), crown volume (CV cm3) and crown diameter (CD cm), (y) is the log transformation of total biomass (g), trunk biomass (g), branch biomass (g) or leaf biomass (g), b0 as the intercept coefficient, b1 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), root mean square error of model (Model RMSE, which is the same as Furnival’s Index for untransformed data) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
(x) (y) Biomass b0 (S.E.) P b1 (S.E.) P b2 (S.E.) P R2 MODEL RMSE
(F.I.) CV RMSE
RCD Total 1,259.5(40.6)<0.01 4,645.9(322.4)<0.01 148.8(322.4)0.6 0.77 314.6 313.6
Trunk 1,056.5(26.3)<0.01 3,979.3(208.8)<0.01 289.2(208.8)0.2 0.85 203.8 199.0
Branch 172.6(26.2)<0.01 521.3(208.1)0.02 -137.1(208.1)0.5 0.07 203.1 178.8
Leaf 30.4(4.0)<0.01 145.3(31.9)<0.01 -3.3(31.9)0.9 0.23 31.1 29.7
DBH Total 1,259.5(70.7)<0.01 2,968.3(561.3)<0.01 -362.0(561.3)0.5 0.30 547.7 552.9
Trunk 1,056.5(54.2)<0.01 2,677.0(429.9)<0.01 -530.3(429.9)0.2 0.38 419.5 413.3
Branch 172.6(27.4)<0.01 191.3(217.3)0.4 136.2(217.3)0.5 -0.01 212.1 192.5
Leaf 30.4(4.3)<0.01 100.1(34.4)<0.01 32.1(34.4)0.4 0.11 33.6 33.5
SD Total 1,259.5(50.5)<0.01 3,994.2(400.9)<0.01 1,498.8(400.9)<0.01 0.64 391.2 409.7
Trunk 1,056.5(35.1)<0.01 3,639.9(278.3)<0.01 798.3(278.3)<0.01 0.74 271.6 269.2
Branch 172.6(25.1)<0.01 232.6(199.3)0.2 672.2(199.3)<0.01 0.15 194.5 204.5
Leaf 30.4(4.2)<0.01 121.7(33.3)<0.01 28.3(33.3)0.4 0.16 32.5 31.4
CV Total 1,259.4(63.9)<0.01 2,809.7(507.5)<0.01 2,119.9(507.5)<0.01 0.43 495.3 498.9
Trunk 1,056.5(57.9)<0.01 2,007.5(459.5)<0.01 1,355.8(459.5)<0.01 0.29 448.4 444.3
Branch 172.5(23.2)<0.01 620.7(183.7)<0.01 -691.8(183.7)<0.01 0.28 179.3 170.4
Leaf 30.4(3.4)<0.01 181.4(27.1)<0.01 -72.3(27.1)<0.01 0.45 26.4 24.5
CD Total 1,259.5(61.3)<0.01 3,582.4(486.4)<0.01 -902.8(486.4)0.07 0.47 474.7 470.4
Trunk 1,056.5(55.3)<0.01 2,316.9(438.8)<0.01 1,270.4(438.8)<0.01 0.36 428.2 427.6
Branch 172.6(20.9)<0.01 1,057.9(165.6)<0.01 351.8(165.6)0.04 0.41 161.7 169.9
Leaf 30.4(3.2)<0.01 207.6(25.4)<0.01 15.9(25.4)0.5 0.51 24.8 24.5

258
Appendix 16. Table. Allometric equations following the quadratic model for biomass components of 0.5 x 7.5 m alley rows of Leucaena leucocephala 6 months post pollarding to a height of 2 m. Where (x) is the root collar diameter (RCD mm, 0.05 m), or RCD and Height (cm) as predictors of biomass, (y) is the log transformation of total biomass (g), trunk biomass (g), branch biomass (g) or leaf biomass (g). b0 as the intercept coefficient, b1 and b2 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), root mean square error of model (Model RMSE, which is the same as Furnival’s Index F.I. for untransformed data) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
(x) (y) Biomass b0 (S.E.) P b1 (S.E.) P b2 (S.E.) P b3 (S.E.) P b4 (S.E.) P R2
MODEL RMSE (F.I.)
CV RMSE
RCD +
HEIGHT Total 1,259.5(34.6)<0.01 4,300.9(12.9)<0.01 358.4(1.2)0.2 1,111.7(3.4)<0.01 561.6(1.9)0.07 0.81 314.6 284.2
Trunk 1,056.5(42.8)<0.01 3,757.6(16.7)<0.01 412.2(2.0)0.05 635.3(2.8)<0.01 214.1(1.1)0.3 0.87 188.2 194.0
Branch 172.6(6.7)<0.01 444.9(1.9)0.06 -76.1(-0.4)0.7 345.1(1.5)0.1 308.1(1.5)0.1 0.11 194.9 170.3
Leaf 30.4(8.6)<0.01 98.4(3.1)<0.01 22.3(0.8)0.4 131.3(4.1)<0.01 39.4(1.4)0.2 0.41 26.8 25.3

259
Appendix 17. Table. Allometric equations following the log-linear model for biomass components of 0.5 x 7.5 m alley rows of Erythrina berteroana 6 months post pollarding to a height of 2 m. Where ln(x) is the log transformation of the following measurements used as predictors of biomass: Root collar diameter (RCD mm, 0.05 m), diameter at breast height (DBH mm, 1.3 m), stump diameter (SD mm, 0.3 m), crown volume (CV cm3) and crown diameter (CD cm) ln(y) is the log transformation of total biomass (g), trunk biomass (g), branch biomass (g) or leaf biomass (g). b0 as the intercept coefficient, b1 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), lambda (λ) and standard deviation (S.D.) as back transformation according to Marklund (1987), Furnival’s Index (F.I.) according to Kershaw et al. (2016), root mean square error of model (Model RMSE) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
ln(x) ln(y)
Biomass b0 (S.E.) P b1 (S.E.) P R2 λ S.D F.I. MODEL RMSE
CV RMSE
RCD Total -2.12 (0.72) 0.0045 2.19 (0.18) <0.001 0.68 1.24 1.33 585.4 0.54 0.46
Trunk -2.09 (0.76) 0.007 2.12 (0.19) <0.001 0.65 1.28 1.66 465.3 0.55 0.39
Branch -5.87 (1.30) <0.001 2.64 (0.32) <0.001 0.50 1.42 1.23 140.0 0.92 1.51
Leaf -8.80 (1.56) <0.001 2.86 (0.38) <0.001 0.44 1.70 1.78 25.6 1.19 1.20
DBH Total 2.40 (0.58) <0.001 1.52 (0.19) <0.001 0.49 1.36 0.75 608.0 0.56 0.57
Trunk 2.27 (0.56) <0.001 1.48 (0.18) <0.001 0.49 1.21 1.03 455.9 0.54 0.55
Branch -0.18 (1.08) 0.87 1.73 (0.36) <0.001 0.26 1.57 1.47 158.2 1.03 1.07
Leaf -3.52 (1.26) 0.007 2.19 (0.42) <0.001 0.29 1.67 1.52 25.9 1.21 1.25
SD Total -0.58 (0.67) 0.39 1.95 (0.17) <0.001 0.64 1.43 0.98 596.4 0.55 0.54
Trunk -0.96 (0.63) 0.13 1.98 (0.16) <0.001 0.67 1.21 1.15 433.6 0.51 0.39
Branch -2.21 (1.28) 0.09 1.87 (0.33) <0.001 0.31 1.69 2.05 163.5 1.07 1.62
Leaf -6.84 (1.37) <0.001 2.55 (0.36) <0.001 0.42 1.89 2.48 26.2 1.22 1.25
CV Total -0.32 (0.56) 0.57 0.55 (0.04) <0.001 0.70 1.30 0.86 579.1 0.53 0.55
Trunk -0.25 (0.60) 0.68 0.52 (0.05) <0.001 0.65 1.28 1.11 471.3 0.55 0.59
Branch -4.22 (0.91) <0.001 0.70 (0.07) <0.001 0.59 1.42 1.29 130.8 0.85 0.90
Leaf -7.60 (1.08) <0.001 0.81 (0.08) <0.001 0.57 1.53 1.99 22.6 1.05 1.07
CD Total 0.57 (0.50) 0.25 1.44 (0.11) <0.001 0.70 1.18 0.23 565.1 0.52 0.55
Trunk 0.76 (0.56) 0.18 1.33 (0.13) <0.001 0.62 1.29 1.44 478.9 0.56 0.63
Branch -3.99 (0.65) <0.001 2.04 (0.15) <0.001 0.73 0.25 0.19 104.8 0.68 0.74
Leaf -7.06 (1.07) <0.001 2.28 (0.24) <0.001 0.58 1.49 1.36 20.1 0.94 0.93

260
Appendix 18. Table. Allometric equations following the log-linear model for biomass components of 0.5 x 7.5 m alley rows of Erythrina berteroana 6 months post pollarding to a height of 2 m. Where ln(x) is the log transformation of root collar diameter (RCD mm, 0.05 m), or RCD and Height (cm) as predictors of biomass. ln(y) is the log transformation of total biomass (g), trunk biomass (g), branch biomass (g) or leaf biomass (g). b0 as the intercept coefficient, b1 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), lambda (λ) and standard deviation (S.D.) as back transformation according to Marklund (1987), Furnival’s Index (F.I.) according to Kershaw et al. (2016), root mean square error of model (Model RMSE) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
ln(x) ln(y)
(Biomass) b0 (S.E.) P b1 (S.E.) P b2 (S.E.) P R2 λ S.D F.I. Model RMSE
CV RMSE
RCD + Height Total -6.41 (1.02) <0.001 1.53 (0.2) <0.001 1.31 (0.25) <0.001 0.77 1.12 0.83 523.2 0.47 0.49
Trunk -6.91 (1.03) <0.001 1.38 (1.38) <0.001 1.47 (0.25) <0.001 0.76 1.13 0.94 409.5 0.48 0.50
Branch -7.40 (2.99) <0.001 2.4 (0.4) <0.001 0.44 (0.59) 0.46 0.41 1.66 1.47 140.5 0.91 0.96
Leaf -14.50 (2.46) <0.001 1.98 ( 0.47) <0.001 1.74 (0.6) 0.005 0.49 1.58 1.49 24.1 1.12 1.17

261
Appendix 19. Table. Allometric equations following the log-linear model for biomass components of 0.5 x 7.5 m alley rows of Inga edulis 6 months post pollarding to a height of 2 m. Where ln(x) is the log transformation of the following measurements used as predictors of biomass: Root collar diameter (RCD mm, 0.05 m), diameter at breast height (DBH mm, 1.3 m), stump diameter (SD mm, 0.3 m), crown volume (CV cm3) and crown diameter (CD cm) ln(y) is the log transformation of total biomass(g), trunk biomass (g), branch biomass (g) or leaf biomass (g). b0 as the intercept coefficient, b1 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), lambda (λ) and standard deviation (S.D.) as back transformation according to Marklund (1987), Furnival’s Index (F.I.) according to Kershaw et al. (2016), root mean square error of model (Model RMSE) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
ln(x) ln(y)
(Biomass) b0 (S.E.) P b1 (S.E.) P R2 λ S.D. F.I. Model RMSE
CV RMSE
RCD Total 0.56 (0.51) 0.28 1.68 (0.12) <0.001 0.70 1.07 0.54 759.2 0.44 0.46
Trunk 0.95 (0.43) 0.03 1.47 (0.10) <0.001 0.71 1.07 0.56 409.6 0.38 0.39
Branch -8.89 (1.67) <0.001 3.45 (0.40) <0.001 0.48 2.37 4.64 287.9 1.45 1.51
Leaf -6.47 (1.33) <0.001 2.78 (0.32) <0.001 0.48 2.19 6.52 169.1 1.16 1.20
DBH Total 3.78 (0.71) <0.001 1.06 (0.20) <0.001 0.25 1.21 0.72 1192.5 0.70 0.72
Trunk 3.39 (0.59) <0.001 1.03 (0.17) <0.001 0.32 1.20 0.66 629.3 0.58 0.60
Branch -2.31 (1.86) 0.22 2.19 (0.53) <0.001 0.18 2.65 2.83 362.3 1.82 1.87
Leaf -0.31 (1.53) 0.84 1.53 (0.44) <0.001 0.13 2.27 3.47 219.2 1.50 1.55
SD Total 0.95 (0.62) 0.13 1.68 (0.16) <0.001 0.59 2.56 1.34 888.9 0.52 0.54
Trunk 0.78 (0.45) 0.08 1.60 (0.12) <0.001 0.71 1.12 0.65 411.0 0.38 0.39
Branch -7.83 (1.86) <0.001 3.40 (0.48) <0.001 0.39 2.56 4.61 312.3 1.57 1.62
Leaf -4.88 (1.56) <0.01 2.55 (0.40) <0.001 0.34 2.61 8.16 191.4 1.31 1.37
CV Total 0.84 (0.96) 0.38 0.46 (0.06) <0.001 0.37 1.13 0.60 1092.0 0.64 0.66
Trunk 1.62 (0.86) 0.06 0.37 (0.06) <0.001 0.33 1.18 0.64 626.3 0.57 0.59
Branch -9.32 (2.53) <0.001 1.01 (0.18) <0.001 0.30 2.81 5.14 334.6 1.68 1.73
Leaf -9.30 (1.81) <0.001 0.99 (0.13) <0.001 0.44 1.93 3.83 176.1 1.21 1.26
CD Total 1.83 (0.78) 0.02 1.14 (0.16) <0.001 0.40 1.18 0.62 1070.2 0.63 0.66
Trunk 2.47 (0.70) <0.001 0.92 (0.14) <0.001 0.34 1.16 0.65 619.3 0.57 0.59
Branch -8.63 (1.93) <0.001 2.83 (0.39) <0.001 0.40 2.33 3.57 309.7 1.56 1.62
Leaf -7.14 (1.46) <0.001 2.47 (0.27) <0.001 0.47 2.04 5.29 171.8 1.18 1.24

262
Appendix 20. Table. Allometric equations following the log-linear model for biomass components of 0.5 x 7.5 m alley rows of Inga edulis 6 months post pollarding to a height of 2 m. Where ln(x) is the log transformation of root collar diameter (RCD mm, 0.05 m), or RCD and Height (cm) as predictors of biomass. ln(y) is the log transformation of total biomass(g), trunk biomass (g), branch biomass (g) or leaf biomass (g). b0 as the intercept coefficient, b1 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), lambda (λ) and standard deviation (S.D.) as back transformation according to Marklund (1987), Furnival’s Index (F.I.) according to Kershaw et al. (2016), root mean square error of model (Model RMSE) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
ln(x) ln(y)
(Biomass) b0 (S.E.) P b1 (S.E.) P b2 R2 λ S.D. F.I. Model RMSE
CV RMSE
RCD + Height Total -0.27 (1.26) 0.83 1.64 (0.13) <0.001 0.17 (0.24) 0.71 0.69 1.14 0.57 761.1 0.44 0.46
Trunk 0.49 (1.07) 0.65 1.45 (0.11) <0.001 0.1 (0.20) 0.64 0.71 1.06 0.56 409.6 0.37 0.40
Branch -8.96 (4.14) 0.03 3.45 (0.43) <0.001 0.01 (0.8) 0.99 0.47 2.41 4.72 287.9 1.45 1.52
Leaf -10.24 (3.28) <0.01 2.64 (0.34) <0.001 0.8 (0.64) 0.21 0.48 2.03 5.63 169.1 1.15 1.20

263
Appendix 21. Table. Allometric equations following the log-linear model for biomass components of 0.5 x 7.5 m alley rows of Leucaena leucocephala 6 months post pollarding to a height of 2 m. Where ln(x) is the log transformation of the following measurements used as predictors of biomass: Root collar diameter (RCD mm, 0.05 m), diameter at breast height (DBH mm, 1.3 m), stump diameter (SD mm, 0.3 m), crown volume (CV cm3) and crown diameter (CD cm) ln(y) is the log transformation of total biomass(g), trunk biomass (g), branch biomass (g) or leaf biomass (g). b0 as the intercept coefficient, b1 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), lambda (λ) and standard deviation (S.D.) as back transformation according to Marklund (1987), Furnival’s Index (F.I.) according to Kershaw et al. (2016), root mean square error of model (Model RMSE) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
ln(x) ln(y)
(Biomass) b0 (S.E.) P b1 (S.E.) P R2 λ S.D. F.I. Model RMSE
CV RMSE
RCD Total -0.19 (0.42) 0.66 1.91 (0.11) <0.001 0.82 1.05 0.30 292.6 0.27 0.28
Trunk -0.20 (0.35) 0.57 1.87 (0.09) <0.001 0.87 1.03 0.23 203.6 0.22 0.23
Branch -7.63 (2.17) <0.001 3.17 (0.58) <0.001 0.33 2.35 3.51 98.2 1.39 1.51
Leaf -8.42 (1.59) <0.001 2.96 (0.43) <0.001 0.44 1.61 1.54 14.9 1.03 1.20
DBH Total 3.89 (0.47) <0.001 1.02 (0.15) <0.001 0.42 1.12 0.56 531.8 0.50 0.51
Trunk 3.59 (0.41) <0.001 1.06 (0.14) <0.001 0.50 1.11 0.50 398.5 0.44 0.45
Branch 0.69 (1.54) 0.65 1.17 (0.51) 0.02 0.08 2.61 4.10 115.1 1.63 1.87
Leaf -1.10 (1.20) 0.36 1.25 (0.39) 0.002 0.14 1.92 2.18 18.5 1.28 1.55
SD Total 2.39 (0.59) <0.001 1.31 (0.17) <0.001 0.50 1.15 1.17 492.9 0.46 0.52
Trunk 1.89 (0.48) <0.001 1.40 (0.14) <0.001 0.63 1.10 0.57 464.0 0.38 0.39
Branch -2.26 (2.01) 0.27 1.86 (0.57) 0.002 0.15 4.80 23.82 110.8 1.57 1.62
Leaf -4.01 (1.53) 0.01 1.91 (0.42) <0.001 0.24 1.97 2.95 17.4 1.20 1.37
CV Total 3.00 (0.54) <0.001 0.30 (0.04) <0.001 0.47 1.14 0.54 507.1 0.47 0.49
Trunk 3.39 (0.55) <0.001 0.26 (0.042) <0.001 0.39 1.12 0.55 443.3 0.49 0.50
Branch -6.24 (1.40) <0.001 0.89 (0.11) <0.001 0.48 0.62 0.93 86.4 1.23 1.27
Leaf -6.43 (1.04) <0.001 0.79 (0.08) <0.001 0.56 0.42 0.51 13.3 0.92 0.95
CD Total 2.52 (0.52) <0.001 0.99 (0.12) <0.001 0.53 1.11 0.49 475.5 0.44 0.46
Trunk 3.06 (0.57) <0.001 0.84 (0.13) <0.001 0.42 1.09 0.51 431.9 0.47 0.49
Branch -7.16 (1.43) <0.001 2.55 (0.32) <0.001 0.51 1.86 2.33 83.7 1.19 1.23
Leaf -7.67 (0.98) <0.001 2.31 (0.22) <0.001 0.65 1.34 0.97 11.9 0.82 0.85

264
Appendix 22. Table. Allometric equations following the log-linear model for biomass components of 0.5 x 7.5 m alley rows of Leucaena leucocephala 6 months post pollarding to a height of 2 m. Where ln(x) is the log transformation of root collar diameter (RCD mm, 0.05 m), or RCD and Height (cm) as predictors of biomass. ln(y) is the log transformation of total biomass(g), trunk biomass (g), branch biomass (g) or leaf biomass (g). b0 as the intercept coefficient, b1 as the slope coefficient, both reported with standard error (S.E.) and significance (P). Coefficient of determination (R2), lambda (λ) and standard deviation (S.D.) as back transformation according to Marklund (1987), Furnival’s Index (F.I.) according to Kershaw et al. (2016), root mean square error of model (Model RMSE) and results of K-fold model cross validation (k=10) reported as root mean squared error (CV RMSE).
ln(x) ln(y)
(Biomass) b0 (S.E.) P b1 (S.E.) P b2 (S.E.) P R2 λ S.D F.I. Model RMSE
CV RMSE
RCD + Height Total -3.04 (0.79) <0.001 1.69 (0.12) <0.001 0.66 (0.16) <0.001 0.86 0.98 0.26 64.33 0.24 0.25
Trunk -2.26 (0.67) 0.001 1.71 ( 0.1) <0.001 0.47 (0.13) <0.001 0.89 1.03 0.21 39.02 0.20 0.21
Branch -22.3 (4.03) <0.001 2.01 (0.59) <0.001 3.36 (0.81) <0.001 0.46 2.53 6.44 109.94 1.23 1.29
Leaf -21.53 (2.75) <0.001 1.92 (0.40) <0.001 3.01 (0.55) <0.001 0.61 1.34 1.05 10.62 0.84 0.89

265
CHAPTER V
CATTLE BROWSING IMPACTS ON THREE
SILVOPASTORAL FORAGE LEGUMES IN AMAZONIA
Abstract
A lack of knowledge of locally adapted browse species limits the development of
more sustainable livestock production systems in the Amazon. Three replicates of a
control and three stands of forage legumes arranged in three alleys of 0.5 x 7.5 m
were each directly browsed by cattle to determine the forage potential of these
species. Destructive sampling of grass and tree foliage, canopy measurements and
leaf counts prior to and post browsing were used to estimate browse removal. Post-
trial, the damage of cattle on leaves and branches of trees of each species were
measured. Soil compaction was measured between treatments and prior to and
following the cattle trial. All three species were palatable to cattle and readily eaten
when fresh. Treatments containing trees and grass produced more available forage
(>2.2 Mg ha-1) for cattle than the grass only treatment (1.5 Mg ha-1). Destructive
sampling of foliar biomass below 1.6 m demonstrated that cattle consumed 99% of
Erythrina berteroana, 75% of Inga edulis and 80% of Leucaena leucocephala. When
leaves were counted from 0.5-1.5 m height the change from pre to post-trial was a
decrease of 95% for E. berteroana, 80% for I. edulis and 87% for L. leucocephala.
Cattle browsed at an average height of 1.5 m but reached a maximum height of 2.6

266
m for E. berteroana as they pulled down branches browse. This research highlights
the value of locally adapted browse species for cattle forage and the productive
potential of silvopastoral systems in the Amazon.
1. Introduction
Cattle are traditionally viewed as grazers that consume grass and other herbaceous
plants from the ground. It is often overlooked that they are also browsers and that
shrub and tree foliage can provide an important contribution to cattle diets
(Zampaligré et al., 2013; Dechnik-Vázquez et al., 2019). A number of environmental
factors are contributing to the global decline of grassland, including increased
frequency of drought, fire and changes in atmospheric CO2 levels (Morgan et al.,
2007). There is increased impetus to find browse species that can contribute to a
diverse and resilient system for livestock production in the face of climatic change
(Murgueitio et al., 2014). Under rising temperatures the forage quality of grass is
predicted to decline, whilst simultaneously methane emissions from cattle are
predicted to increase (Lee et al., 2017). In the Bragantina region of Northern Brazil,
Hohnwald et al. (2010) demonstrated that in the humid tropics, the integration of
woody components could enhance forage production, ecological stability and
sustainability when compared to grass monocultures. Historically, shrubs and trees
have been perceived as weeds that limit grass productivity and efforts have
concentrated on their elimination (Scoones, 1995). To date, little attention has been
given to developing livestock production systems in the Amazon biome that seek to
maximise the availability of tree and shrub browse to cattle, also known as

267
silvopastoral systems (Hohnwald et al., 2016). It remains to be proven in the region
that such a system could increase the production of edible biomass over traditional
models of monoculture grass pastures.
The inclusion of trees and shrubs for browse can present advantages for livestock
production and increase the provision of ecosystem services when compared with
pasture production systems. Extensive cattle production systems in the tropics lack
shade that could serve to reduce the incidence of heat stress in cattle. Heat stress in
cattle is associated with decreased feed intake (Mader and Davis, 2004),
reproductive health (Klinedinst et al., 1993), milk yield (Parsons et al., 2001) and
fitness and longevity (King et al., 2006). Certain trees and shrubs can serve to
reduce methane emissions when ingested by cattle (Pal et al., 2015). The benefits of
trees are numerous and include enhanced nutrient cycling and soil formation, the
recovery of compacted soils, the reduction of runoff, additional habitat to support
biodiversity, increased ecological connectivity and the mitigation of climate change
via the sequestration of carbon (Fisher, 1995; Montagnini and Nair, 2004; Sharrow
and Ismail, 2004; MEA, 2005; Udawatta and Kallenbach, 2011; Mauricio et al.,
2019). Farmers may experience direct benefits of planting trees, such as the
increased longevity of livestock farms. Such benefits may reduce the need for
expansion into primary forest for new pasture (Murgueitio et al., 2011).
In the Amazon, perpetual pasture degradation and abandonment has resulted in an
expanding agricultural frontier of deforestation for the continued production of cattle

268
(Martínez and Zinck, 2004). This process is viewed as the antithesis of forest
conservation. The Amazon hosts many native trees that have shown potential for
domestication in agroforestry systems, some of which are reported to be palatable to
cattle (Serrão et al., 1996; Hohnwald, 2016). A well managed browse component
could help ameliorate the chronic degradation of pastures and even mitigate some
effects of climatic change of the Amazon (Serrão et al.,1996). Furthermore, the
degraded pastureland in the Amazon represents an opportunity for afforestation
projects that could restore landscapes to productivity and simultaneously increase
the provision of ecosystem services. A key limitation to the adoption of this approach
is the lack of studies that document the impact of cattle browsing on native plants.
The development of silvopastoral systems could be an important incentive for
landowners in the Amazon to reduce reliance on extensive grass monocultures and
the use of fire in the management of pastures for cattle production (Serrão et al.,
1996; Hohnwald, 2016). Few studies have documented the forage potential of locally
adapted species. An increased understanding of which plants are palatable to cattle
and readily eaten when fresh would contribute to the development of appropriate
silvopastoral systems. The business of cattle farming seeks to increase per hectare
livestock productivity. If the inclusion of forage trees in pastures can be shown to
increase productivity over current management then there is an economic incentive
for the adoption of silvopastoral systems. A key limitation to developing such
approaches is a lack of methodologies that quantify the productive potential of
forage for cattle from trees and shrubs. In contrast to studies that consider the total

269
biomass production of trees, this study aims to identify which proportion of the total
biomass is available to cattle via a series of direct browsing experiments, and
therefore this study attempts to quantify the productive increase, as opposed to the
total increase in biomass.
This study seeks to compare novel and existing approaches to assess the forage
potential of trees and shrubs for cattle production by quantifying the browsing impact
of cattle on trees. The study compares trees destructively harvested prior to and post
cattle browsing, with alternative, non-destructive approaches. The non-destructive
approaches include measurements of the crown and leaf counts. A range of
destructive and non-destructive methodologies have been attempted here in order to
compare the consistency of the results of each method and provide a series of
experimental approaches that can be attempted in future studies with reference to
the present study. The browsing impact on the trees is considered as the diameter of
woody material consumed or broken via cattle interactions, for three species in a
random block field trial in a pasture established in the Peruvian Amazonian region of
Madre de Dios. These approaches can be used to determine the total edible
biomass available to cattle and the amount of biomass consumed, and crucially,
evaluates the accuracy of non-destructive approaches that may be used as
methodologies in on-going field trials to measure biomass removal by cattle. Such
studies can contribute to our understanding of management and productivity of
alternative silvopastoral livestock systems.

270
It was hypothesised that:
(a) Erythrina berteroana, Inga edulis and Leucaena leucocephala are palatable to
cattle and will be consumed when fresh.
(b) Tree and grass arrangements can surpass the amount of edible biomass
than that found in monoculture Brachiaria brizantha cv. Marandú pastures.
(c) The availability of browse to cattle is limited by the height of the foliage.
2. Methods
2.1 Experimental Design and Cattle Herd
A detailed description of the experimental design and establishment is available in
Chapter II. The experiment sought to evaluate the browse potential of 24-month old
Inga edulis, Erythrina berteroana and Leucaena leucocephala. These species had
previously been shown to survive and grow at the study site (Chapter II). These
three species are considered experimental treatments. The fourth experimental
treatment was a control of pasture without trees. There were five replicate plots of
each treatment available at the beginning of this study. Three replicate plots were
selected at random for the present study due to limited resources. In total, there
were 12 experimental plots, composed of three replicates of four treatments.
Eight non-lactating Bos taurus indicus heifers of approximately equal size and body
condition were selected for the browsing experiment (Edmonson et al., 1989). The
cattle were drawn from a herd that were managed via grazing on pastures and

271
browsing a 4 km roadside strip adjacent to the property. The cattle had previous
experience of browsing several different species of trees and shrubs but were
unfamiliar with the study site. The cattle were bought onto the experimental property
and into a training corral where they had a 15-day period of acclimatization on the
Brachiaria brizantha cv. Marandú grass. An electric fence was introduced to the
corral so that the cows would become accustomed to the fence.
The experiment was carried out in three eight-day phases (Table 15). Phase I began
on the 13th February 2019. The cattle were selected in pairs and each pair was
introduced into a replicate plot. After 8 days, the pairs of cattle were moved to the
next replicate plot that had been chosen at random where they remained for eight
further days. Then the cattle were moved into the last plot where they remained for a
further eight days. Phase III of the experiment concluded on the 8th March 2019.
Table 15. Pairing combinations and random rotation of cattle (8 cattle numbered 1-8) between four treatments in three, eight day phases of a cattle trial.
I. edulis L. leucocephala E. berteroana Control Grass
Phase I 1,6 7,8 2,5 3,4 Phase II 3,4 1,6 7,8 2,5 Phase III 7,8 2,5 3,4 1,6
Two cattle were allocated per treatment, for a total of eight cattle in the four
replicates at any one time (Figure 52). A one-meter wide strip was cut around all
replicates for the installation of an electric fence and to prevent cattle from browsing
adjacent plots. A single red polyester wire electric fence was installed around each
replicate plot at a height of 0.8 m. Permanent water (provided ad libitum) and 5 x 5 m

272
shade stations were installed at the edge of each plot. The area that contained the
water and shade station was excluded from all sampling.
Figure 52. Map of the cattle trial from east to west Phase I (red), Phase II (blue) and Phase III (yellow).
2.2. Browsing Detection Using Destructive Measurements
At the beginning of each eight-day sampling period the replicate number, tree
species, date, observer and subsample codes were recorded. The root collar
diameter (RCD, measured at the widest part 0.05 m above the ground) of trees
within each plot was measured and divided into ten size classes. Six trees from each
size class were randomly selected for destructive sampling; three to be harvested
prior to the introduction of cattle, and three to be harvested after the cattle had left
the replicate. Trees were harvested up to two days before the cattle were introduced,
and were harvested up to two days after they had left. The biomass was harvested in

273
two stages to consider the effect of resource height on resource intake. It was
assumed that cattle could reach a maximum height of 1.6 m. All biomass greater
than 1.6 m from ground level was harvested separately from biomass less than 1.6
m. Biomass was classified as stem, branch or foliage, and weighed using an
appropriately sized spring-loaded weight scale.
Statistical Analysis
To determine whether an effect of browsing on foliar biomass could be detected, the
total foliar biomass was compared prior to and post the cattle trial. Data were first
checked for normality using the Shapiro-Wilk test for normality and the Levene’s test
for variance. In the case of I. edulis, the data were not normally distributed but had
comparable variances. Therefore, a Mann-Whitney was used to test for differences.
In the case of E. berteroana the Levene’s test demonstrated that the variance
between the results prior to and post-trial was too great to consider using the Mann-
Whitney test, therefore a Mood’s Median test was used.
To test the hypothesis that a greater proportion of foliar biomass was consumed at a
height less than 1.6 m the ratio of foliar biomass under and over 1.6 m was
compared for each tree when sampled prior to and post the cattle trial. The
proportional change was compared for each species using a proportional test. To
assess the relative amount of tree forage that had been consumed by animals
between species, the proportion of foliar biomass below 1.6 m prior to and post the
introduction of cattle was compared using a one-way Analysis of Variance (ANOVA).

274
This established the relative amount of foliage that had been consumed by the
animals between treatments and was considered as a measure of palatability.
2.3. Browsing Detection Using Non-Destructive Measurements
Within each replicate, 30 trees were selected to represent ten size classes. Tree
height was measured from the ground to the top of the highest meristem. Leaf
emergence height (LEH) was measured from the ground to where foliage volume
assumed 5% of the total. The same measurement was called canopy height when
used in the equations presented below. A 5 m measuring tape was used to measure
crown diameter (CD), taken as the mean of three horizontal measurements of the
crown perpendicular to the ground.
The crown radius (r) was calculated as the crown diameter divided by two. Crown
depth (CD) was derived using equation 1. Crown area (CA) approximates a circle
and was derived using equation 2. Crown volume (CV) was assumed to equate to a
cylinder, the best fit for the pollarded trees, and was calculated using equation 3:
(1) Crown Depth = Height – Canopy Height
(2) CA = πr2
(3) CV =πr2CD
Statistical Analysis

275
To test for an impact of the cattle browsing on crown volume, the crown volumes for
each tree measured prior to and post the cattle trial were compared within a species.
The crown volume data were checked for normality using the Shapiro-Wilk test.
Crown volume was not normally distributed and therefore the homogeneity of
variances was tested using Levene’s test. The crown volumes of both I. edulis and L.
leucocephala demonstrated a homogeneous variance and therefore the Mann
Whitney U test was used with the crown volume as a continuous response variable
and prior to or post cattle trial as a categorical predictor. For E. berteroana the
Levene’s test showed that the variance within the data were not homogeneously
distributed. Therefore, the decision was taken to use the Kruskal-Wallis test to
determine if there was an effect of cattle browsing on crown volume.
To determine whether cattle had an effect on the height at which leaves were
present, the continuous response variable of LEH was tested for normality using the
Shapiro-Wilk test for each species. The LEH of both I. edulis and E. berteroana were
normally distributed and therefore paired t-tests were used to compare for each
species the LEH prior to and post the cattle trial. The LEH data for L. leucocephala
were not normally distributed and therefore a Levene’s test was employed to confirm
homogeneity of the variance. The canopy height data for L. leucocephala were of
equal variance and therefore a Mann Whitney U test was used test for an effect of
cattle browsing on LEH.

276
2.4 Browsing Detection Using Leaf Counts
Leaves were counted prior to and post the cattle trial to detect an effect of browsing
on the number of leaves. Within each species 60 branches were chosen at random
from 30 trees per replicate. Two sections of the branch were chosen. The first
section was 1 m long, and was measured from 0.5 m from the ground. Assuming the
branches were vertical, this can be described as a section 0.5 m to approximately
1.5 m above the ground. The second section was 0.5 m long and measured from 1.5
m to approximately 2 m above the ground. The replicates were exposed to cattle
within two days of sampling, and the sampling procedure was repeated the day after
the cattle left the replicates. The difference in the number of leaves prior to and post-
trial was assumed to be due to utilization by cattle.
Statistical Analysis
The total number of leaves on a tree was a continuous variable. A Shapiro-Wilk test
was used to test for normality and the distribution of the variable was found to be
non-normal. A non-parametric test, the Wilcoxon signed rank test was used to
compare the total number of leaves prior to and post the cattle trial for each species.
2.5 Measurement Of Browsing Impact
Immediately following each eight-day cattle trial, 30 trees in every replicate were
visually assessed for signs of browsing or cattle impact. The height of the impact
was recorded and categorised as branch or leaf. Two damage types were reported,
either browsed or broken. The diameter of the impact was recorded with callipers.

277
Statistical Analysis
The Shapiro-Wilk test was used to check the normality of the distribution of the
continuous response variables: the diameter of point of leaf browsing, diameter at
point of branch browsing, diameter of branch broken, height of leaf browsed and the
height of branch browsed. The height of leaf browsing was found to have a normal
distribution therefore a one-way ANOVA was used to determine if there was a
difference between the heights of the browsing impact between the different tree
species. Tukey’s Honest Significant Differences test was applied post-hoc to
determine which species were different. Shapiro-Wilk tests indicated the following
response variables were not normally distributed: diameter at point of branch
browsing, diameter of branch broken and the height of branch browsed. Non-
parametric tests were used for these response variables. The Kruskal-Wallis test
was applied, using species as a categorical predictor. When the result was found to
be significant, Dunn’s test was used to determine which species were different.
Dunn’s test is considered an appropriate test for groups with unequal numbers of
observations (Zar, 2010). Mortality as a result of the cattle trial was calculated as the
percentage of total trees that had died as a result of the trial.
2.6 Estimation Of Grass Forage
Prior and post-feeding trial, all ground biomass was harvested within five randomly
located 1 m2 quadrats. Sampling prior to the feeding trial subjectively considered
biomass as either an ‘edible’ or ‘nonedible’ fraction. ‘Edible fraction’ was visually

278
assessed as the leaf or stem (< 25 mm) component of the grass no less than 10 cm
from the ground. This fraction was considered to be the ‘leaf’ part of the grass and
the part most commonly favoured by cattle. The inedible fraction was the remaining
grass biomass, which was considered to be the ‘woody stem’ part of the grass and of
lower grazing value to the cattle. Each fraction was weighed separately as fresh
weight. Three subsamples were taken of each fraction and dried to a constant
weight. Post feeding trial there was no ‘edible’ fraction available to sample, therefore
sampling consisted solely of the non-edible fraction.
2.7 Effect Of Trees and Cattle on Soil Bulk Density
To determine the effect of trees on soil bulk density, samples were taken from either
within one meter of the tree line or between the rows of trees to a depth of either 0 –
10 cm or 10 – 20 cm. Five samples were taken from each of these categories, for a
total of 20 samples per replicate. A cylindrical steel corer of 5 x 9 cm was used to
give a volume of 318 cm3. The core was driven vertically into the soil and excavated
with a trowel. Once removed, the soil core was transferred to a paper bag. The core
was oven dried at 55 °C until a constant dry weight was reached and this value was
reported. Bulk density was calculated as the weight of the sample divided by the
volume of the sample and is reported in g cm3 (Swift and Bignell, 2001). To
determine the effect of cattle on soil bulk density the aforementioned sampling
procedure was repeated following the introduction of cattle. In control replicates with
no trees, sample sites were chosen randomly and no closer than 10 m to an edge of
each plot.

279
Statistical Analysis
The effect of trees on soil bulk density was determined within treatments by
comparing the bulk density within the tree line with the bulk density between the
rows of trees at the 0 – 10 cm or 10 – 20 cm depth profile. The bulk density was the
continuous response variable to the categorical treatment of species. Within each
species and depth, the bulk density was tested for normality by using Shapiro-Wilk
tests. The data were found to be normally distributed. Paired t-tests were used for
each species and depth to compare the compaction of the soils within the tree line
with the compaction of the soils between the rows of trees.
Subsequent to the cattle trial the sampling methodology was repeated to test for an
effect of cattle on soil compaction. Using each treatment as a predictor variable, the
bulk density post-cattle trial at two depths was tested for normality by using Shapiro-
Wilk tests. Where the data were normally distributed, paired t-tests were used to
compare the bulk density prior to or post the introduction of cattle. The data were not
normally distributed for L. leucocephala at 5-10 cm depth, or for E. berteroana at
either the 0-10 cm or 10-20 cm depths. In these cases Kruskal-Wallis tests were
used. If an effect of cattle on soil bulk density was found, it was explored via t-tests
to determine whether there was an effect of trees on soil bulk density by comparing
the soil bulk density from within the tree line with that taken from between the rows of
trees.

280
3. Results
Cattle browsing reduced the amount of available foliage for all species (Figures 53-
56). This effect was detected by all methodologies. When foliar biomass was
destructively sampled below 1.6 m, the results show foliar biomass reductions for all
three tree species following the introduction of the cattle. A greater proportion of
biomass was lost below 1.6 m than over 1.6 m for E. berteroana (P< 0.05), I. edulis
(P <0.01) and L. leucocephala (P< 0.01). Destructively sampled L. leucocephala
showed an increase in total foliar biomass of 20% (P<0.01) post-browsing; this result
is discussed further in the discussion section. When foliar biomass below 1.6 m was
compared prior to and post the cattle trial, E. berteroana showed a 99% decrease, I.
edulis a 75% decrease and L. leucocephala an 80% decrease (Table 16). When the
change in foliar biomass below 1.6 m was compared prior to and post cattle trial
there was no effect of species detected. When total biomass (i.e. below and above
1.6 m) was considered there was a reduction in total foliar biomass of E. berteroana
of 92% (P<0.01) and of I. edulis by 30% (P<0.05).
Table 16. Dry foliar biomass for individual trees (dry matter g tree-1), standard deviation (S.D.) and number of individuals sampled (n) prior to and post cattle browsing either mean total tree foliar biomass or foliar biomass harvested <1.6 m on potential browse species Erythrina berteroana, Inga edulis and Leucaena leucocephala. Significant differences (P<0.05) from prior value to post value following cattle browsing denoted by an asterisk (*).
Prior or Post Foliar biomass Foliar biomass <1.6 m
Species Cattle Trial n g tree-1 S.D. g tree-1 S.D.
E. berteroana Prior 87 45.3 66.2 15.5 66.2
Post 90 3.8* 11.8 0.2* 11.8
I. edulis Prior 88 297.1 347.7 141.7 347.7
Post 90 204.5* 348.2 34.9* 348.2
L. leucocephala Prior 87 60.4 193.0 6.6 193.0

281
Post 90 72.3* 277.9 1.3* 277.9
Figure 53. Cattle browsing Inga edulis

282
Figure 54. Browsing line where cattle consumed all the foliage they could reach.
Figure 55. Post-browsing, an Erythrina berteroana row is missing several trees

283
Figure 56. Leucaena leucocephala defoliated on low branches
E. berteroana demonstrated the greatest loss of crown volume due to browsing, a
67% decrease (P <0.01), followed by I. edulis (51% decrease, P <0. 01) (Table 17).
For the species L. leucocephala there was a 31% reduction but this was a
statistically weaker effect of browsing on crown volume (P = 0.06). Post browsing,
the leaf emergence height increased in E. berteroana by 49% (P<0.01), in I. edulis
by 50% (P <0.001) and in L. leucocephala by 20% (P <0.01) during the browsing
period.

284
Table 17. Mean average (Av) and standard deviation (S.D.) of change per individual tree in crown volume (M3) and leaf emergence height (LEH, cm) prior to and post a cattle browsing experiment on individuals (n) of potential browse species, Erythrina berteroana, Inga edulis and Leucaena leucocephala. Significant differences between sampling period prior to and post (P<0.05) denoted by an asterix (*).
Species
Prior to cattle trial Post cattle trial
Parameter Av S.D. n Av SD n
Crown Volume (M3) E. berteroana 0.94 1.15 88 0.31* 0.51 50
I. edulis 2.01 2.05 48 1.02* 1.65 46
L. leucocephala 1.54 2.6 83 1.06 2.05 75
LEH (cm) E. berteroana 112.6 40.1 88 167.8* 74.2 50
I. edulis 101.6 33.3 48 152.5* 35.1 46
L. leucocephala 191 45.9 83 230.7* 53.5 75
The mean number of leaves decreased among all species as a result of cattle
browsing (Table 18). E. berteroana lost the greatest proportion of leaves in both the
0.5 – 1.5 m and the 1.5 – 2.0 m category (95% and 91% respectively). I. edulis and
L. leucocephala both lost more leaves under 1.5 m (80%, 87%) than over 1.5 m
(48%, 80%). When the methodologies used to measure browsing impact prior to and
post cattle trial are compared between species they show comparable results
between species, as shown for E. berteroana in Figure 57, I. edulis in Figure 58 and
L. leucocephala in Figure 59.

285
Table 18. Mean leaf count per tree (number of leaves tree-1) and standard deviation (S.D.) prior to and post a cattle-browsing trial on individuals (n) of potential browse species Erythrina berteroana, Inga edulis and Leucaena leucocephala. Significant differences (P<0.05) between leaves counted prior to and post browsing are denoted by an asterix (*).
Sampling Range Species
Prior/ Post n
Number of leaves tree-1 S.D.
Mean reduction (%)
1.5 m - 2 m E. berteroana Prior 65 26.3 18.6
Post 65 1.4* 3.5 -95
I. edulis Prior 70 17.6 18.8
Post 70 9.2* 14.4 -48
L. leucocephala Prior 37 16.1 14.4
Post 37 3.9* 7.2 -76
0.5 m - 1.5 m E. berteroana Prior 132 29.3 23.6
Post 132 2.6* 4.6 -91
I. edulis Prior 163 24.6 23.2
Post 163 5.0* 6.6 -80
L. leucocephala Prior 99 16.2 13.6
Post 99 2.1* 5.0 -87
Figure 57. Percentage change of Erythrina berteroana biomass for four assessment methodologies used to detect biomass removal following cattle browsing.
-100
-80
-60
-40
-20
0
20
40
60
80
100
Destructive <160cm
Crown Volume Leaf Count 50-150 cm
Leaf EmergenceHeight
Pro
po
rtio
na
l ch
an
ge

286
Figure 58. Percentage change of Inga edulis biomass components between methodologies used to detect biomass removal following a cattle-browsing experiment.
Figure 59. Percentage change of Leucaena leucocephala biomass components between methodologies used to detect biomass removal following a cattle-browsing experiment.
Browsing damage was recorded on branches for all species (Table 19). The
browsed branches of E. berteroana showed an increased diameter compared with
-120
-100
-80
-60
-40
-20
0
20
40
60
Destructive <160cm
Crown Volume Leaf Count 50-150 cm
Leaf EmergenceHeight
Pro
po
rtio
na
l ch
an
ge
-120
-100
-80
-60
-40
-20
0
20
40
Destructive <160cm
Crown Volume Leaf Count 50-150 cm
Leaf EmergenceHeight
Pro
po
rtio
na
l ch
an
ge

287
other species. The branches of E. berteroana had a 35% greater diameter than I.
edulis and 65% greater than L. leucocephala. There was no effect of species on
maximum height of branches browsed. E. berteroana had the highest rate of
mortality when measured immediately post browsing (12.5%) that was six times
greater than that of the species with the lowest mortality, I. edulis (2.1%). L.
leucocephala had a mortality of 4.8%.
Table 19. Impacts measured following a cattle trial on three potential browse species Erythrina berteroana, Inga edulis and Leucaena leucocephala. Impact types were classified as browsing-type damage (Browsed) or breakage-type damage (Broken) of two categories, branches (all species) and leaves (I. edulis). Average diameter (mm), average height (cm) and maximum height (cm) and respective standard deviations (S.D.) reported on a number (n) of individual trees. Significant differences between species (P<0.05) denoted by asterisk (*).
Impact Category
Impact Type
Diameter (mm) Height (cm)
Species Average S.D. Average Max S.D. n
E. berteroana Branch Broken 12.8* 7.2 153 240 41.7 126
Branch Browsed 5.0* 1.8 156 256 42.7 222
I. edulis Branch Broken 9.0* 4.7 134 195 37.0 39
Branch Browsed 3.9* 1.4 150 246 36.3 181
Leaf Browsed 2.5 0.7 144 197 55.3 245
L. leucocephala Branch Broken 6.5* 3.8 135 260 44.1 41
Branch Browsed 2.8* 0.9 154 225 36.2 113
There was no effect of treatment on the amount of edible or non-edible biomass
available prior to the cattle trial (Table 20). When grass biomass was compared prior
to and post the cattle trial, 100% of the edible fraction was consumed in all
treatments. If the edible fraction of grass in each treatment was combined with the
results of the destructive foliage estimates <1.6 m as a measure of total available
edible foliage, the no-tree control treatment would produce 1.5 Mg ha-1 less available
edible forage than the tree treatments of E. berteroana (2.3 Mg ha-1), I. edulis (2.3
Mg ha-1) or L. leucocephala (2.2 Mg ha-1).

288

289
Table 20. Edible and non-edible fractions of mean average dry weight grass biomass (Mg ha-1) produced in four treatments, reported for sampling carried out prior to and post the introduction of cattle.
Prior (n = 15) Post (n = 15)
Treatment Fraction Average (Mg ha-1) S.D.
Average (Mg ha-1) SD
Control Edible 1.5 0.6 0
Non-edible 30.2 9.7 38.8 9.1
E. berteroana Edible 2.3 0.8 0
Non-edible 29.4 10.9 29.8 10.0
I. edulis Edible 1.9 0.5 0
Non-edible 22.7 8.5 30.7 11.8
L. leucocephala Edible 2.2 0.9 0
Non-edible 30.2 12.9 31.4 7.8
Prior to the cattle trial there were no differences in the soil bulk density within the line
of trees or between the rows of trees of any of the species tested. Following the
cattle trial, the soils of the I. edulis replicates had become compacted at both the 0-
10 cm (P <0.01) and the 10-20 cm horizon (P <0.01). This effect was found to occur
within the tree line at both the 0-10 cm (P <0.05) and 10-20 cm horizon (P <0.01),
and between the rows of trees at the 10-20 cm horizon (P <0.05). No effect of the
introduction of cattle on soil compaction was found in any of the other treatments.
4. Discussion
This study sought to identify the forage potential of three tree species. The results
show that the three tree species are palatable when fresh and readily eaten by
cattle. When the biomass of traditional monoculture was compared with the available
biomass of the silvopastoral system the results showed that the inclusion of these
tree species would increase the forage available to cattle. Therefore, the

290
silvopastoral system represents an increase in per hectare productivity and has the
potential to support more cattle per hectare than the pasture-only conventional
system. The reported increase in production per hectare is a conservative estimate
as this figure only considers the foliar biomass up to a height of 1.6 m. Over the
course of the experiment, it was found that cattle were able to access browse up to a
height of 2.6 m. The estimation only considered foliar biomass as the sole
component of edible biomass. In practice, it was found that cattle also browsed
branches. The trees therefore provided a greater amount of edible biomass to the
cattle than that reported in this study.
Destructive sampling was able to describe and characterise the impact of browsing
on the foliar biomass of all species in this experiment. Although samples were drawn
from the same population there was a degree of natural deviation between the
samples of trees destructively harvested prior to and post the introduction of cattle.
An example of this in the present study was that destructive sampling detected an
increase in total foliar biomass for L. leucocephala. This result is a methodological
artefact and implies that a sample size must be chosen that is proportional to the
variation within the population and the predicted browsing impact. Often this is
difficult to establish in advance and the present study detected an increase in
biomass despite a reasonable sample size. A drawback to this approach is that there
is a high input of labour for the results.

291
Destructive sampling of foliar biomass does not have to destroy the tree. It would be
possible to strip the trees of foliar biomass prior to an experiment and compare it
with the foliar biomass harvested subsequent to a browsing experiment. This is also
known as simulated browsing. The advantage of this is the tree can be measured
over time, a possibility that is removed when the tree is destructively harvested. A
disadvantage is that simulated browsing may not accurately represent the height of
browsing which may be greater or lower than expected, or the animal’s interaction
with the browse, for example, an animal’s use of woody branches or ability to
manipulate branches by pulling on them. Destructive sampling may be a necessary
approach to determine the total biomass of trees and this approach is often used to
calibrate or evaluate alternative, non-destructive techniques (Barnes, 1976).
Sanon et al. (2007) found that the mean height reached by browsing cattle was 1.47
m and the maximum was 1.9 m. Schoenbaum et al. (2018) found in Israel that cattle
(described as local breed x Hereford) could reach a maximum grazing height of 2.2
m. Ouédraogo-Koné et al. (2006) found that cattle could browse to a height of 1.1 m.
In the present study, cattle were found to browse on average between 1.3 and 1.6 m.
However, the maximum heights reported to have reached cattle in this study
exceeded 2.5 m. It was only possible to identify the maximum diameter of leaf
browsing impact in the species I. edulis due to partial consumption of the leaf by
cattle. When cattle browsed E. berteroana or L. leucocephala leaves the entire leaf
was removed. Therefore, there was no visible post-trial impact to record. In
anticipation of the effect of browsing height on resource availability, the destructive

292
sampling had considered all above ground biomass in two height categories, over
1.6 m and below 1.6 m. A greater proportion of biomass was consumed below 1.6 m.
If the total amount of foliar biomass were considered as the estimation of total
available forage, the results would have underestimated the proportion of available
forage consumed. If the resource is to be directly browsed a study must seek to
establish the effect of resource height on direct browsing by the animal as this will
influence estimations of the availability of forage (Renaud et al., 2003). This is
because when large trees are considered, only a fraction of the total foliar biomass is
actually within reach and therefore available to the cattle. Alternatively, forage
systems such as cut and carry or lopping systems may aim to determine the edible
fraction of biomass available to animals with little regard for the browsing height
(Balehegn et al., 2012).
The non-destructive approach to measuring a browsing impact using crown volume
prior to and post cattle trial is limited by the fact that crown volume does not consider
whether biomass is within reach of the cattle and an effect of light browsing may not
be detected. The impact of browsing was detectable in the present study,
presumably due to the trees having a low crown due to pollarding. Trees in cattle
pastures may exhibit browse lines; this term describes a visible line of defoliation
caused by cattle consuming all foliage within their reach. This study introduces the
concept of leaf emergence height (LEH), which was conceptualised as attempt to
capture the occurrence and height of browse lines on the tree species in the study.
The LEH increased for all species post cattle trial, which suggests the trees were

293
consumed by the cattle. However, the LEH does not indicate how much forage was
consumed. Furthermore, trees that may offer some supplemental forage value but
are not palatable enough to be completely defoliated may be overlooked by this
technique. The LEH could be of use when determining an appropriate pollard height
for species.
The number of leaves lost as a result of the cattle trial was strikingly similar to the
results of the destructive harvest of trees below 1.6 m. The methodology similar to
that described by Rees (1974). The present study initially planned to employ the leaf
counting technique only in the 0.5-1.5 m range. However, after the first round of the
cattle trial it became apparent that in certain instances the cattle were browsing
above this range. Therefore, an additional category of 1.5 – 2 m was specified. A
greater proportion of leaves were lost in the 0.5 – 1.5 m range for all species, as this
range was the easiest for cattle to reach. Rees (1974) did not establish a uniform
height for sampling within the experiment. This is possibly due to different
management approaches between their study and this study. In Rees’ (1974) study
the plants were pollarded to 1 m height in the season in which the study was carried
out and it can be assumed that all tree forage was accessible to cattle.
The specific management of this silvopastoral system is related to the quantities of
biomass produced by both the tree and grass elements. For instance, this study was
limited to one planting arrangement of 0.5 x 7.5 m, equating to 2667 trees ha-1. Six
months prior to the cattle trial, all trees had been pollarded to 2 m height to reduce

294
competition with grass (Chapter III). It was found that the pollarding reduced the
negative effects of trees on the grass (presumed to be due to light competition).
However, the pollarding may also have impacted the tree’s production and
distribution of foliar biomass and these effects were not studied. The browsing
resulted in a substantial reduction in foliage on the trees, and this may have
increased the light available to the grass. Further work is needed to understand how
the tree and grass elements interact in silvopastoral systems. For example, the
planting arrangement, the selection of tree and grass companion species, the use of
fertilisers and pruning or pollarding management can all effect the production per
hectare and were not evaluated in the present experiment.
A limitation of this study was that it did not consider how the availability of grass
affected the practice of tree browsing. While it was observed that the trees were
consumed at the same time as the grass there was no data collected to suggest that
consumption of browse was as a result of the depletion of the edible grass biomass.
The results of this study indicate a substantial portion of inedible biomass. The
Brachiaria brizantha cv. Marandú pasture had been sown by hand in March 2017
and it was first grazed 24 months later at the time of the present experiment. This
meant that a large fraction of the grass biomass had become inedible, as the mature
stems of grass are fibrous and unpalatable to cattle. This study employed a
subjective estimation of edible biomass fraction. The total edible fraction was
completely devoured by the completion of the trial, which suggests the methodology
underestimated the total edible fraction of grass. Future studies may consider

295
simulating the grazing of the grass by repeatedly cutting the grass, an approach that
was beyond the resources of this study.
The foliage of all tree species was consumed by cattle and there were no differences
detected between species in the amount of foliage consumed. This suggests that all
species were palatable and that relative palatability was not detected in the present
study. I. edulis produced the most overall biomass and the most accessible and
edible foliage for the cattle of all the tested species. The results of this study confirm
that I. edulis is palatable to cattle and readily eaten when available. It observed that
the cattle preferred to be in the shade of I. edulis than in the sun. I. edulis
experienced branch browsing and damage to branches as a result of cattle
interaction. However, I. edulis demonstrated the lowest immediate mortality following
cattle interaction. This may be because stems of I. edulis were of an increased
diameter when compared with those of E. berteroana or L. leucocephala (Chapter
III). It is possible that stem size and mortality following the cattle trial were related.
While beyond the scope of the present study, this could be of importance in
determining an appropriate stage at which silvopastoral systems could be exposed
to cattle.
Hohnwald et al., (2015) reported that I. edulis was a high producer of woody and
leafy biomass, a frequently occurring tree in Capoeira, a nitrogen fixing legume and
palatable for cattle. Hohnwald (2016) reported that I. edulis had ‘bad performance’
on a scale of palatability and biomass, which they described as a ‘positive value’ for

296
forage quality and ‘intermediate’ values for mortality. Hohnwald (2016) found that I.
edulis suffered from the overgrown grass sward, but when established it was robust.
He also mentions that the small trees may have not been obvious to browsing cattle
amongst the tall grass and may have been overlooked, ultimately resulting in a
negative palatability value. The present study differs from Hohnwald (2016) in the
experimental design, establishment and management of trees and finds contrary to
Hohnwald (2016) that I. edulis is a suitable candidate for further investigation for
forage potential. Dechnik-Vázquez et al. (2019) rated I. edulis 11th of 268 species on
the amount of times the species appeared browsed. They found tannin content of
0.31% but no presence of toxic cyanogenic glycosides.
E. berteroana consistently exhibited the greatest proportion of foliar biomass lost, the
greatest diameter of branches consumed and broken and the highest mortality
following the cattle trial when compared with the other species. Unforeseen in the
experimental design was that cattle would employ unique browsing strategies for
each species. It was observed that the cattle displayed a browsing habit for E.
berteroana of pulling down branches on which to feed. The branches would
frequently break during this process and often sections of the broken branches
would also be chewed and consumed. Post-trial, almost all the branches of E.
berteroana trees had been broken, in some cases entire plants were missing,
presumed destroyed by the cattle. This phenomenon was also reflected in the post-
impact monitoring, where E. berteroana reported the highest average and maximum

297
point of branch browsing, and the greatest increase in leaf emergence height
following the experiment.
This study was limited to the planting of 1 m stakes of E. berteroana. This may have
contributed to a substantially different growth form from than that observed when E.
berteroana is grown from seed, where it tends to form a more robust and resistant
tree (personal observation). The 1 m stakes produced adventitious shoots formed
from the callus tissue between the bark and the wood of the cut surface that may
have resulted in weaker branches than a natural growth form of E. berteroana. This
may have made the trees of this species susceptible to cattle damage. Future
studies could consider propagation from 15 cm cuttings or from seed.
Of the three tree species, L. leucocephala produced the least available foliar
biomass. It was observed that this was due to L. leucocephala growing out of reach
of the cattle as the pollard at 2 m, which favoured the production of adventitious buds
that grew at a height of 2 m. This resulted in a minimal contribution to available
forage for cattle. If the tree is produced with the intention of being directly browsed
by cattle, a pollarding height under 2 m may be considered to encourage leaf
production at accessible heights. Guevarra et al. (1978) compared three harvest
frequencies of 2.5, 3 and 4 months, three planting arrangements of 15 x 30 cm, 30 x
50 cm or 45 x 50 cm and two accessions of L. leucocephala in North Kohala, Hawaii.
They found that for optimal forage production, a planting arrangement of 15 x 50 cm
and pollarding when the plants were approximately 1 m in height were desirable

298
management practices to increase forage yield. The present study reports the ability
of this species to grow in the Amazon in association with B. brizantha cv. Marandú.
L. leucocephala has been widely documented as a tropical forage tree but it has not
yet been reported to grow successfully in the Amazon. Szott et al. (1991) speculated
that L. leucocephala failed to establish due to high levels of exchangeable aluminium
at their experimental site in Yurimaguas, Peru. L. leucocephala is considered a
highly invasive tree (Richardson, 1998). However, no incidences of natural
regeneration were observed in the invasive B. brizantha cv. Marandú pasture.
Prior to the cattle trial there were no within species differences in soil bulk density
between or within the rows of trees. This may be due to the relatively undisturbed
nature of the site at the beginning of the study, as cattle had not been present on the
land for 30 years and no machinery was used in the preparation of the experiment.
After the cattle trial the soils had a higher bulk density in the I. edulis parcels. This
corresponds with the observation that the cattle were spending their time in the
shade of I. edulis.
Research has shown that in traditional pasture models of the Amazon the highest
nutrient concentrations occur in the most utilized and therefore compacted areas, for
example, surrounding the water trough. This may result in a net loss of nutrients over
time as the land’s nutrients are essentially exported and subsequently leached
(Boddey et al., 2004). If the cattle are spending time in the shade of I. edulis, these
nutrients may be circulated back into the tree and become available to the animals

299
via consumption of the foliage. Furthermore, over 5 years the roots of I. edulis have
been shown to reverse the compaction of soils (Fisher, 1995). A relevant extension
of this work would be to attempt to capture the effects of on-going compaction by
cattle on growth and mortality of I. edulis. For example, planting trees in alley rows
may result in different patterns of cattle movement and behaviour, which may
influence the distribution of soil compaction and nutrients across pastures. A logical
extension of the present work is a critical assessment of the chemical composition of
these foliages and their impact on animal health.
In conclusion, the inclusion of trees into pastures can provide an increase in
available forage for cattle. There are multiple techniques, both destructive and non-
destructive that can be used to estimate the utilisation of browse plants by cattle.
Cattle are generalist browsers and may include leaves and twigs into their diet.
Where resource availability is limited by resource height, cattle may find ways to
manipulate browse trees to access forage out of their reach. Locally adapted species
may be able to provide palatable forage for cattle. Further work is needed to
understand the impacts of these forage species on animal health.
References
Balehegn, M., Eniang, E.A., Hassen, A., 2012. Estimation of browse
biomass of Ficus thonningii, an indigenous multipurpose fodder tree in
northern Ethiopia. African Journal of Range & Forage Science.
https://doi.org/10.2989/10220119.2012.687071

300
Barnes, D.L., 1976. A review of plant‐based methods of estimating food
consumption, percentage utilisation, species preferences and feeding
patterns of grazing and browsing animals. Proceedings of the Annual
Congresses of the Grassland Society of Southern Africa 11, 65–71.
https://doi.org/10.1080/00725560.1976.9648781
Boddey, R.M., MacEdo, R., Tarré, R.M., Ferreira, E., De Oliveira,
O.C., De, C., Cantarutti, R.B., Pereira, J.M., Alves, B.J.R.,
Urquiaga, S., 2004. Nitrogen cycling in Brachiaria pastures: The key
to understanding the process of pasture decline. Agriculture,
Ecosystems and Environment 103, 389–403.
https://doi.org/10.1016/j.agee.2003.12.010
Dechnik-Vázquez, Y.A., García-Barrios, L., Ramírez-Marcial, N., van
Noordwijk, M., Alayón-Gamboa, A., 2019. Assessment of browsed plants
in a sub-tropical forest frontier by means of fuzzy inference. Journal
of Environmental Management 236, 163–181.
https://doi.org/10.1016/J.JENVMAN.2019.01.071
Edmonson, A.J., Lean, I.J., Weaver, L.D., Farver, T., Webster, G.,
1989. A Body Condition Scoring Chart for Holstein Dairy Cows. Journal
of Dairy Science 72, 68–78.

301
https://doi.org/10.3168/JDS.S0022-0302(89)79081-0
Fisher, R.F., 1995. Amelioration of Degraded Rain Forest Soils by
Plantations of Native Trees. Soil Science Society of America Journal
59, 544. https://doi.org/10.2136/sssaj1995.03615995005900020039x
Guevarra, A.B., Whitney, A.S., Thompson, J.R., 1978. Influence of
Intra-Row Spacing and Cutting Regimes on the Growth and Yield of
Leucaena1. Agronomy Journal 70, 1033.
https://doi.org/10.2134/agronj1978.00021962007000060034x
Hohnwald, S., Rischkowsky, B., King, J.M., Camarão, A.P., Rodrigues
Filho, J.A., Zeppenfeld, T., 2015. Intensive cattle browsing did not
prevent fallow recuperation on smallholder grass-capoeira pastures in
the NE-Amazon. Agroforestry Systems 89, 813–828.
https://doi.org/10.1007/s10457-015-9815-9
Hohnwald, S., Trautwein, J., Camarão, A.P., Wollny, C.B., 2016.
Relative palatability and growth performance of capoeira species as
supplementary forages in the NE-Amazon. Agriculture, Ecosystems &
Environment 218, 107–115. https://doi.org/10.1016/J.AGEE.2015.11.015
King, J.M., Parsons, D.J., Turnpenny, J.R., Nyangaga, J., Bakari, P.,

302
Wathes, C.M., 2006. Modelling energy metabolism of Friesians in Kenya
smallholdings shows how heat stress and energy deficit constrain milk
yield and cow replacement rate. Animal Science 82, 705–716.
https://doi.org/10.1079/ASC200689
Klinedinst, P.L., Wilhite, D.A., Hahn, G.L., Hubbard, K.G., 1993. The
potential effects of climate change on summer season dairy cattle milk
production and reproduction. Climatic Change 23, 21–36.
https://doi.org/10.1007/BF01092679
Lee, M.A., Davis, A.P., Mizeck G.G.C., Manning, P., 2017. Forage
quality declines with rising temperatures, with implications for
livestock production and methane emissions. Biogeosciences 14,
1403–1417. https://doi.org/doi:10.5194/bg-14-1403-2017
Mader, T.L., Davis, M.S., 2004. Effect of management strategies on
reducing heat stress of feedlot cattle: Feed and water intake.
Journal of Animal Science 82, 3077–3087.
https://doi.org/10.2527/2004.82103077x
Martínez, L.J., Zinck, J.A., 2004. Temporal variation of soil
compaction and deterioration of soil quality in pasture areas of
Colombian Amazonia. Soil and Tillage Research 75, 3–17.

303
https://doi.org/10.1016/j.still.2002.12.001
Morgan, J.A., Milchunas, D.G., LeCain, D.R., West, M., Mosier, A.R.,
2007. Carbon dioxide enrichment alters plant community structure and
accelerates shrub growth in the shortgrass steppe. Proceedings of the
National Academy of Sciences 104, 14724–14729.
https://doi.org/10.1073/pnas.0703427104
Murgueitio, E., Calle, Z., Uribe, F., Calle, A., Solorio, B., 2011.
Native trees and shrubs for the productive rehabilitation of tropical
cattle ranching lands. Forest Ecology and Management 261, 1654–1663.
https://doi.org/10.1016/j.foreco.2010.09.027
Murgueitio, E.R., Chará, J.O., Cuartas, C.C., Naranjo, J.R., Barahona,
R.R., 2014. Intensive Silvopastoral Systems (ISPS), mitigation and
adaptation tool to climate change. Tropical and Subtropical
Agroecosystems 17, 501–507.
Ouédraogo-Koné, S., Kaboré-Zoungrana, C.Y., Ledin, I., 2006. Behaviour
of goats, sheep and cattle on natural pasture in the sub-humid zone of
West Africa. Livestock Science 105, 244–252.
https://doi.org/10.1016/J.LIVSCI.2006.06.010

304
Pal, K., Patra, A.K., Sahoo, A., Kumawat, P.K., 2015. Evaluation of
several tropical tree leaves for methane production potential,
degradability and rumen fermentation in vitro. Livestock Science 180,
98–105. https://doi.org/10.1016/J.LIVSCI.2015.07.011
Parsons, D.J., Armstrong, A.C., Turnpenny, J.R., Matthews, A.M.,
Cooper, K., Clark, J.A., 2001. Integrated models of livestock systems
for climate change studies. 1. Grazing systems. Global Change Biology
7, 93–112. https://doi.org/10.1046/j.1365-2486.2001.00392.x
Rees, W.A., 1974. Preliminary Studies into Bush Utilization by Cattle
in Zambia, Journal of Applied Ecology 11 (1), 207-214
Renaud, P.C., Verheyden-Tixier, H., Dumont, B., 2003. Damage to
saplings by red deer (Cervus elaphus): effect of foliage height and
structure. Forest Ecology and Management 181, 31–37.
https://doi.org/10.1016/S0378-1127(03)00126-9
Richardson, D.M., 1998. Forestry Trees as Invasive Aliens Article in
Conservation Biology. https://doi.org/10.1046/j.1523-1739.1998.96392.x
Sanon, H.O., Kaboré-Zoungrana, C., Ledin, I., 2007. Behaviour of
goats, sheep and cattle and their selection of browse species on

305
natural pasture in a Sahelian area. Small Ruminant Research 67, 64–74.
https://doi.org/10.1016/J.SMALLRUMRES.2005.09.025
Schoenbaum, I., Henkin, Z., Yehuda, Y., Voet, H., Kigel, J., 2018.
Cattle foraging in Mediterranean oak woodlands – Effects of management
practices on the woody vegetation. Forest Ecology and Management
419-420, 160–169. https://doi.org/10.1016/J.FORECO.2018.03.017
Scoones, I., 1995. Exploiting heterogeneity:habitat use by cattle in
dryland Zimbabwe. Journal of Arid Environments 29, 221–237.
https://doi.org/10.1016/S0140-1963(05)80092-8
Serrão, E.A.S., Nepstad, D., Walker, R., 1996. Upland agricultural and
forestry development in the Amazon: sustainability, criticality and
resilience. Ecological Economics 18, 3–13.
https://doi.org/10.1016/0921-8009(95)00092-5
Sharrow, S.H., Ismail, S., 2004. Carbon and nitrogen storage in
agroforests, tree plantations, and pastures in western Oregon, USA.
Agroforestry Systems 60, 123–130.
https://doi.org/10.1023/B:AGFO.0000013267.87896.41
Szott, L.T., Palm, C.A., Sanchez, P.A., 1991. Agroforestry In Acid

306
Soils Of The Humid Tropics. Advances in Agronomy 45, 275–301.
https://doi.org/10.1016/S0065-2113(08)60043-7
Udawatta, R.P., Kallenbach, R.L., 2011. Agroforestry Buffers for
Nonpoint Source Pollution Reductions from Agricultural Watersheds
Influence of Cover Crop Managements on X-Ray CT-Measured Pore
Parameters and Hydraulic Properties of Soil View project
Parameterization of the APEX model View project. Article in Journal of
Environmental Quality. https://doi.org/10.2134/jeq2010.0168
Zampaligré, N., Dossa, L.H., Schlecht, E., 2013. Contribution of
browse to ruminant nutrition across three agro-ecological zones of
Burkina Faso. Journal of Arid Environments 95, 55–64.
https://doi.org/10.1016/J.JARIDENV.2013.03.011

307
CHAPTER VI
CONCLUSIONS
1. Summary of Research Findings
Chapter I highlighted some of the advances and challenges in the current research
into silvopastoral systems. Chapter II outlined a framework of decision-making
processes for the design of agroforestry systems and used the contextual problem of
livestock production induced chronic pasture degradation to describe the
development of a novel silvopastoral system for the Amazon. The objective of the
silvopastoral system was to reduce the degradation of pastures via the inclusion of
locally adapted and native trees, and to offer sustainable improvements in
productivity when compared with the traditional model of extensive cattle farming in
the Amazon.
Chapter III followed the growth of five locally adapted species for 18 months from
when they were planted in a Brachiaria brizantha cv. Marandú pasture. It was found
that the tree species responded differently to the pasture environment. Erythrina
berteroana, Inga edulis and Leucaena leucocephala demonstrated positive increase
in height, diameter at breast height and root collar diameter when measured in all 6-

308
month intervals. After 12 months, Senegalia loretensis and Ceiba pentandra began
to decrease in height as mortality increased for these two species. At 18 months, I.
edulis was 4.4 m tall and was found to halve grass production. All species were
pollarded, and I. edulis produced more pollard biomass (0.42 Mg ha-1) than E.
berteroana (0.19 Mg ha-1) or L. leucocephala (0.27 Mg ha-1). Six months later, all
species but C. pentandra were found to negatively effect grass production. However,
the magnitude of this negative effect was less for I. edulis than in the previous
monitoring period.
It was concluded that some species are better adapted to the environmental
conditions of the study site than others. A shorter, 12-month experiment may have
incorrectly concluded that S. loretensis and C. pentandra were well adapted, but
these species had declined within 18 months. It should also be taken into
consideration that it is unknown how E. berteroana, I. edulis and L. leucocephala will
respond to the pasture environment after 18 months. In-situ field experiments can
assist in the evaluation of tree species in the context of the pests and pathogens
present at the site. It was reported that at the present site all species suffered bark
damage by rodent species, and that C. pentandra was susceptible to attacks from
ants of the Atta genus. These findings demonstrated some of the advantages of in-
situ long-term field experiments.
Rising levels of carbon in the atmosphere are driving global climatic changes and
this has resulted in a market for services and processes that can remove

309
atmospheric carbon, such as afforestation. The potential carbon sequestration that
could be provided by silvopastoral systems is an important market-benefit that could
enable payments to farmers for the adoption of more sustainable practices. A
commonly used non-destructive approach to determine the biomass of a tree, and by
extension the carbon sequestration of a tree, is the use of allometric equations to
determine the relationship between an easily measurable element of the tree and it’s
total biomass.
Chapter IV developed species and component specific allometric equations and
compared the predictive power of a variety of tree measurements against biomass.
The potential combinations were assessed for their fit to the data using linear,
quadratic and log-linear models and the models were compared using the Furnival’s
Index. Log-linear models were found to be superior to linear or quadratic models for
the prediction of biomass components in all species. The results showed that a
single parameter model of root collar diameter (RCD) was the best predictor of all
biomass components of I. edulis, whereas for E. berteroana and L. leucocephala a
multiple model that included RCD and height improved the predictive measurements
of total and trunk biomass. The models for the prediction of total biomass showed
the best fit for the species of L. leucocephala (R2= 0.86) followed by E. berteroana
(R2= 0.77) and I. edulis (R2= 0.70). After 24 months, I. edulis had accumulated the
most biomass (7.2 Mg ha-1) when compared to E. berteroana (4.4 Mg ha-1) or L.
leucocephala (4.3 Mg ha-1). The predicted carbon sequestration of these species
was greatest for I. edulis (3.4 Mg ha-1) when compared to E. berteroana (2.1 Mg ha-

310
1) or L. leucocephala (2.0 Mg ha-1). Chapter IV concludes that silvopasture could
offer increased sequestration of carbon in the form of trees that increase above
ground biomass. This finding is based on the assumption of active management of
trees to reduce negative impacts on the growth, and therefore the carbon
sequestration, of associated grasses. The rapid accumulation of above ground
biomass in I. edulis in the pasture environment and its resilience to pollarding makes
this species an attractive option for planting arrangements to maximize carbon
sequestration.
All three tree species were palatable to cattle and readily eaten when fresh.
Destructive sampling of foliar biomass below 1.6 m suggested that cattle consumed
99% of E. berteroana, 75% of I. edulis and 80% of L. leucocephala. When leaves
were counted from 0.5-1.5 m the change pre to post-trial was a decrease of 95% for
E. berteroana, 80% for I. edulis and 87% for L. leucocephala. Cattle browsed at an
average height of 1.5 m but reached a maximum height of 2.6 m for E. berteroana as
they pulled down branches browse. Cattle consumed the tips of branches and
leaves. E. berteroana trees had been developed from cuttings and they were found
to be the most damaged by exposure to cattle.
Treatments containing trees and grass were found to produce more available forage
(>2.2 Mg ha-1) than the grass only treatment (1.5 Mg ha-1). It is important to consider
the effect of pollarding on these results. After 18 months, there was a negative effect
of I. edulis on the production of pasture grass. This occurred when the I. edulis had

311
an average height of 4.4 m. Subsequently, pollarding of I. edulis at 18 months 2 m
reduced this negative effect. The increased production of available forage biomass
and was greater than the decreased production of the pasture grass biomass.
Pollarding is a useful strategy for managers of silvopastoral systems, as competition
for resources such as light can negatively affect plant productivity.
Practical recommendations for those seeking to plant a silvopastoral system for
cattle production in the Brachiaria pastures of the Amazon are as follows. Establish
80% I. edulis and 20% alternative species in a planting arrangement of 0.5 x 7.5-15
m. The first pollard should occur at the onset of a rainy season when 70% of all trees
are greater than 3 m height. The first pollard should apply to 60% of all I. edulis and
be to a height of 2.5 m. The rationale behind this decision is to make some I. edulis
foliage available as forage for cattle, also to leave some I. edulis to provide shade for
the months where the rain is less frequent and contribute to the continued provision
of ecosystem services. In the absence of cattle the pollard is still a useful way of
thinning the tree line and maximizing the production of litter to contribute to healthy
topsoil. Leaving a considerable proportion of trees will also ensure there are I. edulis
fruit available to replenish the I. edulis plant stocks and expand the planted area.
2. Comparison with Previous Research
This research contributes to a growing body of work that catalogues silvopastoral
systems as an approach to livestock production that can meet the challenge of
sustainably feeding a growing population. Murgueitio et al. (2011) assert that

312
silvopastoral systems are instrumental for the productive rehabilitation of cattle
production and biodiversity conservation in agricultural landscapes. They state that
native trees and shrubs can be integrated into intensive silvopastoral systems and
that this is an important approach for climate change adaptation and mitigation.
Dagang and Nair (2003) reviewed two decades of research on silvopastoral systems
in Central America and find that such systems were not widely implemented by
farmers in spite of the potential for silvopasture to improve pasture productivity and
buffer deforestation. They suggest future researchers must understand the mind-set
of farmers when managing their resources, which is to maximize perceived gains
within the constraints of the system dynamics. They assert that silvopastoral
research must seek to produce fast results, be of low risk for farmers to adopt and
have a favorable impact on farming systems (Dagang and Nair, 2003).
This study relied on an understanding of the forage potential of locally adapted
plants. Serrão et al. (1996) advocate the domestication of native Amazonian plants
for agroforestry systems and conclude that more research is needed to identify
optimal configuration of land use systems. The importance of locally adapted plants
for the development of silvopastoral systems has been highlighted by many authors
(including but not limited to Butterfield, 1995; Ibrahim et al., 2006; Guimarães-Beelen
et al., 2006; Murgueitio et al., 2011; Nunes et al., 2016; Dechnik-Vázquez et al.,
2019). However, research on the forage potential of trees that are adapted to
Amazon pastures is scarce and to date has concentrated on the North Eastern
Amazon in Brazil (Hohnwald et al., 2010; Hohnwald, 2016). The present study is in

313
the South Western Amazon, Peru, 1700 miles from Hohnwald’s (2016) study site. I.
edulis, a species described in Hohnwald’s (2016) experiment was also present in the
region of the present study site and therefore selected for the present experiment as
a parallel with Hohnwald’s (2016) study. However, a different planting design was
used to that of Hohnwald (2016). The planting design of the present silvopastoral
system was taken from the densest rows of alley planted I. edulis (0.5 m) found in
the literature (Alegre and Rao, 1996; Leblanc et al., 2006).
Devendra and Ibrahim (1999) consider that one of the greatest challenges facing the
development of silvopastoral systems is the need to improve integrated resource
movement to increase productivity on whole farm systems that are driven by market-
orientated access. The present study found an increase in two key benefits, that of
increased forage production, which is a benefit that accrues to the farmer, and that of
enhanced carbon sequestration which is experienced as a global good but has an emerging
market. Numerous studies have shown the potential of trees to produce high quality
fodder for animals (Haque et al., 1986; Pezo et al., 1990; Douglas et al., 1996; Ainalis and
Tsiouvaras, 1998; Rengsirikul et al., 2011). The majority of this work has concentrated on
the production of forage in fodder bank arrangements where the production of forage is
measured relative to a lopping, coppicing or pollarding approach (Muschler et al., 1993;
Larbi et al., 2005; Ortega-Vargas et al., 2013; Casanova-Lugo et al., 2014; Khedri et al.,
2018).

314
When trees are grown in rows they are known as alleys. Alleys are commonly used to
grow food crops where prunings from the rows of trees can be left between the rows for
nutrient inputs or weed control or the pruning can be cut and carried to livestock and
used as a fodder bank (Duguma et al., 1988; Leblanc et al., 2006; Ghezehei et al., 2015). In
the present study it was envisaged that the cattle could directly browse the alleys of trees.
The planting design of this study shares certain design characteristics (east to west
planting lines, objective of maximizing production) with the ‘intensive’ silvopastoral
systems described by Murgueitio et al. (2011). Murgueitio et al. (2011) suggest that in an
intensive silvopastoral system a planting density may be upwards of 10,000 plants ha-1,
whereas the present study planted forage trees at a density of 2667 plants ha-1.
The present study reports an increase in carbon sequestration due to enhanced
production of above ground biomass. The carbon storage of trees is directly related to the
production of total biomass, therefore this study may underestimate total carbon
sequestration (Nair et al., 2010). Sharrow and Ismail (2004) hypothesized that as grasses
store the majority of their carbon below ground and trees and shrubs store the majority of
their carbon above ground, combinations of trees and grasses would be able to provide
increased sequestration of carbon. The potential for and uncertainties of silvopastoral
systems to sequester carbon are well documented (see Nair et al., 2010; Feliciano et

315
al., 2018). In the present study it is not known what effect the pollarding may have on
the production of biomass of these species or how this may effect the carbon
sequestration in this system. Ultimately, managers must find a balance between the
benefits they wish to accrue with a silvopastoral system.
The present study only considers the above ground biomass of trees in relation to
potential for carbon sequestration and does not consider the below ground biomass or the
effect of trees in the present study on soil carbon. Feliciano et al., (2018) reviewed 88
studies on soil carbon. They found that the greatest soil carbon sequestration results
from the transition of a grassland system to a silvopastoral system (4.38 Mg
C ha−1 yr−1). Higher soil organic content is related to tree density (Saha et al., 2009).
Soils proximate to trees, compared to soils at a greater distance from trees, store
increased carbon (Howlett, 2009; Takimoto et al., 2009). Haile et al. (2010) found on
Spodosols and Ultisols in Florida that silvopastoral systems had greater potential to
store more stable carbon in the soil at all depths compared with the treeless system.
3. Theoretical Implications of Findings
Wilderness continues to be converted to agriculture to meet the food needs of a
growing global population. Livestock is a major driver of land cover change and while
awareness is of the impacts of meat consumption on the environment is growing in
western societies, the global demand for meat and dairy products is projected to
have increased by 15% in 2027 (OECD‐ FAO, 2018). As the agricultural frontier

316
expands into neotropical rainforest areas, it leaves in its a wake a mosaic of
pasturelands that have been degraded by poor management practices such as fire
regimes and overgrazing (Costa and Rehman, 1999). There is an urgent need to
develop new technology for the productive rehabilitation of these lands if wilderness
areas are to remain while simultaneously feeding a growing global population
(Murgueitio et al., 2011). It is theorised that intensification of existing cattle systems
will reduce the incentive to deforest additional land, in practice this is likely to depend
on political and economic circumstances, such as market saturation (Latawiec et al.,
2014; Müller-Hansen et al., 2019). However, silvopastoral systems would result in
net afforestation that would provide increased ecological connectivity in an
increasingly fragmented landscape (Cadavid-Florez et al., 2019; Vaca et al., 2019).
The practice of silvopasture has been described as a “triple win” land use (Chará,
2019). Silvopastoral systems have the potential to increase cattle stocking densities
by providing an increase in forage that can therefore intensify production in a way
that simultaneously increases carbon sequestration. The adoption of silvopastoral
systems may be a crucial strategy of farmer adaptation to projected climate impacts
which include a decline in the forage quality of grasses under increased
temperatures (Lee et al., 2017), and increased frequency and intensity of extreme
weather patterns, such as drought (Jat et al., 2016; Lawson et al., 2019). The
present study chose tree five species to assess their silvopastoral potential, of which
three were considered to be well adapted to the present study site. There are many
more species in the Amazon that represent good potential for forage (Hohnwald,

317
2016; Dechnik-Vázquez et al., 2019). Different species combinations may also be
more successful than others. The present study alludes to the potential of Inga edulis
as a nurse tree to assist in the establishment of silvopastoral systems in the
Brachiaria brizantha cv. Marandú pasture grasses, as it is able to rapidly accumulate
foliar biomass and open up a niche which due to a reduced presence of grass may
have a reduced influence of the allelopathic effects of the grass.
The findings of this study imply that there is an excellent potential for further
development of silvopastoral systems in the Amazon. Silvopastoral systems in the
Amazon could provide rapid and substantial increases in available forage biomass
and carbon sequestration over monoculture grass pastures. This combination of
benefits accruing at both the farm and global level supports the need to develop
payment-based incentives for landowners to adopt silvopastoral systems.
4. Limitations of the Study
The design of the present study was ambitious. The positive outcome was that it was
able to answer some general questions about the potential for silvopastoral systems
in the Amazon. However, a key limitation of this approach is that often the fine scale
interactions that are interesting design elements of such a system were overlooked.
For example, there was a single planting arrangement of 0.5 x 7.5 m. This meant
that the findings were less comparable with those of the literature, which commonly
focus on tree growth and biomass production in 2 x 2 m or 3 x 3 m arrangements.

318
The results were drawn from a single site, which means the results cannot be
transferred to other areas. Species were arranged in monospecific stands, whereas
in practice mixed stands could increase biodiversity, reduce disease and exploit
more resources by inherent variation such as branching pattern, leaf area, niche
occupancy or rooting depth. All species were drawn from single accessions when in
reality there are multiple accessions of species some of which may have been more
successful than others. The experiment was only carried out once, and therefore is
temporally limited to the specific prevailing conditions of the time at which it was
carried out.
There were six dominant species in this experiment, consisting of five tree species
and one pasture grass. Ideally, there would have been a much more thorough
revision of potential species to use in the experiment but this was not possible due to
the time sensitive nature of the current project. However, there are many more tree
species with excellent potential not considered in the study. No consideration was
taken for the chemical constituents of the foliage, or potential toxicity. The decision to
use Brachiaria brizantha cv. Marandú was taken because it is one of the most
productive grasses available in the region and also presents some of the most
challenging conditions for tree establishment (Sobanski and Marques, 2014). The
decision was taken to arrange the trees in monospecific stands. All management
decisions that excluded an alternative approach can be considered as impact
limitations of the present study as they further narrow the scope of the present study.

319
The trees were pollarded to 2 m at 18 months. This was considered an essential
management strategy in keeping with the objective of optimal forage production, but
also had the effect of limiting the comparisons that could be drawn between the
present study and other studies in the literature. The pollarded biomass was weighed
as a total biomass. In retrospect, it would have been valuable to separate the stems,
branches and foliage. This would have been informative to consider the amount of
forage or fuel wood that could be gathered by the pollard. The original research
design allowed for repeat pollarding and manual defoliation at varying frequencies
and intensities. Due to unforeseen changes to the project timeline this experiment
was not possible.
In Chapter III there were two sampling approaches to measuring the effect of trees
on the regrowth of grass. The first approach considered a 30-day regrowth of grass
that had been cut to 10 cm. It was originally intended to measure consecutive 30-day
regrowth intervals. This approach would have also captured a seasonal effect on
grass growth. The second consideration of the grass biomass occurs after the
pollard event and considers the regrowth following a 6-month interval. Care was
taken to consider the two results separately and this occurrence was an
unintentional artefact of field sampling.
In Chapter V there was no treatment effect on the total fractions of grass biomass
prior to and post the cattle trial. Five 1 m2 grass quadrats were taken in each
replicate prior to the cattle trial and five post the cattle trial. This resulted in n=15 for

320
each treatment prior to the cattle trial and n=15 post the cattle trial. It is possible that
the sample sizes were too small to detect an effect of treatment. When a
retrospective power analysis was carried out it was found hat the number of samples
that would have been required to detect a treatment effect would have exceeded
0.09 ha, the size of each replicate. Therefore, this was not a feasible approach for
the present study. This occurred because the grass had been left to grow for 24
months and this may have resulted in a greater inter-sample variation than that
which may have been found in an active pasture. In anticipation of this, the study
design included a subjective visual assessment of ‘edible biomass’.
The estimation of the contribution of tree biomass to carbon stocks only considered
the above ground biomass and did not consider the below ground components of the
trees. The carbon fraction was derived from the literature. However, as detailed
biomass estimates were made for the three species it would have improved the
study to have component specific estimates of carbon for each tree species. This
was beyond the scope of the present study. No consideration was given to the soil
carbon potential of the silvopastoral system, which could have yielded interesting
results.

321
5. Future Research
5.1 Direct Extensions of the Study
In the present study, the species Erythrina berteroana, Inga edulis and Leucaena
leucocephala were able to establish in a pasture environment when planted as
saplings. This was a costly approach as each tree was planted as a seed in an
individual bag of soil and transported to the planting location. In the present study,
this was feasible due to an on-site nursery. If this technology were to be widely
adopted then a useful study would compare the cost and labour of the present
approach with direct seeding and on-going management of the surrounding pasture
grass.
5.2 Broader Issues for Future Work
A lack of studies document the forage potential of trees native to the Amazon.
Despite the relatively recent history of cattle farming in the Amazon, there is local
ecological knowledge of which species are preferentially browsed by cattle. Once
species have been identified, cafeteria type feeding trials could be used to evaluate
intake and effect on animal health. There is also an effect of the chemicals found in
milk following the consumption of certain forages that should be assessed.
The present study is primarily concerned with the production of forage and
sequestration of carbon. Future research could seek to increase on-farm revenue.
Incorporating a timber element into the silvopastoral system could generate added

322
economic incentives. Fast-growing leguminous timber trees with a low leaf area
index could be incorporated into the rows of trees and simultaneously enhance the
sequestration of carbon. Fruit trees could be an additional form of on-farm income.
Fertilisers could be used to account for the nutrients exported from the site in the
form of timber or fruits. The increased biodiversity of such stands could increase the
farm productivity and resilience by incorporating a more diverse selection of tree
products.
More work is required to understand the costs of implementing silvopastoral systems
and farmers perceptions of such systems. Outreach and education may be required
to explain the system to landowners (Latawiec et al., 2014). It is likely that some form
of credit or payment would be required to assist in the transition to silvopastoral
systems. Some examples of these approaches exist in the literature from other
regions and could be usefully applied in the study region of Madre de Dios, Peru
(Calle et al., 2009; Garbach et al., 2012; Cranford and Mourato, 2014).
5.3 Practical Next Steps
The growing demand for rehabilitation and reforestation projects could be met by the
extension of this research into real world scenarios. The following discussion
summarises some ideas and concepts that occurred as a result of this study and
may be of use when considering how to translate these studies into action. In order
to extend the interpretation of the results of this research it is necessary to replicate
silvopastoral systems to many more sites, which would allow a sample of

323
silvopastoral systems to be directly compared, both between silvopastoral sites with
common and contrasting site conditions and with alternative land uses. This could be
done with the participation and local ecological knowledge of cattle farmers. A survey
of farms could evaluate a farmer’s local ecological knowledge and their willingness
and conditions to engage with the development of such trial silvopastoral systems.
The present study has described numerous benefits of silvopastoral systems
accruing at the farm level. It is likely that landowners will require some assistance to
initially adopt the system and over time will see the benefits and be incentivised by
their own experience to continue to expand the system. Crucially, they would be
involved in every stage, including the production of the plants from seed. Such an
approach would ensure that in the future, once they are able to enjoy the benefits of
the system they have the experience of all stages of the process to continue
expanding the silvopastoral system on their farm and adapting the system to meet
their own needs. The next step is transferring this technology to the farmers and
supporting them to create the silvopastoral system that best fits their needs, and
simultaneously create a baseline from which the system can be measured over time,
in different contexts and thereby attract greater interest and investment.
7. Conclusion
Silvopastoral farming systems can make livestock farming more economically and
environmentally sustainable when compared with traditional models of cattle farming
in the Amazon. The use of robust and well-replicated on-farm experiments such as
the one recommended by the framework in this thesis can be used to evaluate the
considerable potential of locally adapted species for use in silvopastoral systems.

324
When compared with conventional models of farming in the region, the silvopastoral
system presented in this study provided gains in productivity, accruing as increased
total biomass, an increase in available forage for cattle and the sequestration of
carbon. Widespread adoption of silvopastoral systems on existing cleared land in the
Amazon could reduce the need for further deforestation, reverse the trend of pasture
degradation and soil erosion, increase biodiversity in agricultural landscapes and
contribute to the mitigation of climatic change.
References
Ainalis, A.B., Tsiouvaras, C.N., 1998. Forage production of woody
fodder species and herbaceous vegetation in a silvopastoral system in
northern Greece. Agroforestry Systems 42, 1–11.
https://doi.org/10.1023/A:1006112330453
Alegre, J.C., Rao, M.R., 1996. Soil and water conservation by contour
hedging in the humid tropics of Peru. Agriculture, Ecosystems &
Environment 57, 17–25. https://doi.org/10.1016/0167-8809(95)01012-2
Butterfield, R.P., 1995. Promoting biodiversity: advances in
evaluating native species for reforestation. Forest Ecology and
Management 75, 111–121.
Cadavid-Florez, L., Laborde, J., Zahawi, R.A., 2019. Using landscape

325
composition and configuration metrics as indicators of woody
vegetation attributes in tropical pastures. Ecological Indicators 101,
679–691. https://doi.org/10.1016/J.ECOLIND.2019.01.072
Calle, A., Montagnini, F., Andrés, F., 2009. Farmer’s perceptions of
silvopastoral system promotion in Quindío, Colombia. Bois et Forets
des Tropiques 300, 79 – 93.
Chará J., Reyes E., Peri P., Otte J., Arce E., S.F., 2019.
Silvopastoral Systems and their Contribution to Improved Resource Use
and Sustainable Development Goals (SDG): Evidence from Latin America.
Rome, Italy.
Costa, F.P., Rehman, T., 1999. Exploring the link between farmers’
objectives and the phenomenon of pasture degradation in the beef
production systems of Central Brazil. Agricultural Systems 61,
135–146. https://doi.org/10.1016/S0308-521X(99)00043-8
Cranford, M., Mourato, S., 2014. Credit-Based Payments for Ecosystem
Services: Evidence from a Choice Experiment in Ecuador. World
Development 64, 503–520.
https://doi.org/10.1016/j.worlddev.2014.06.019

326
Dagang, A.B.K., Nair, P.K.R., 2003. Silvopastoral research and
adoption in Central America: Recent findings and recommendations for
future directions, in: Agroforestry Systems. Kluwer Academic
Publishers, pp. 149–155. https://doi.org/10.1023/A:1026394019808
Dechnik-Vázquez, Y.A., García-Barrios, L., Ramírez-Marcial, N., van
Noordwijk, M., Alayón-Gamboa, A., 2019. Assessment of browsed plants
in a sub-tropical forest frontier by means of fuzzy inference. Journal
of Environmental Management 236, 163–181.
https://doi.org/10.1016/J.JENVMAN.2019.01.071
Devendra, C., Ibrahim, M., 1999. Silvopastoral systems as a strategy
for diversification and productivity enhancement from livestock in the
tropics. Centro Agronomico Tropical de Investigación y Enseñanza
(CATIE ) 2020, 8–22.
Douglas, G.B., Bulloch, B.T., Foote, A.G., 1996. Cutting management of
willows ( Salix spp.) and leguminous shrubs for forage during summer.
New Zealand Journal of Agricultural Research 39, 175–184.
https://doi.org/10.1080/00288233.1996.9513176
Duguma, B., Kang, B.T., Okali, D.U.U., 1988. Effect of pruning
intensities of three woody leguminous species grown in alley cropping

327
with maize and cowpea on an alfisol. Agroforestry Systems 6, 19–35.
https://doi.org/10.1007/BF02220106
Garbach, K., Lubell, M., DeClerck, F. a J., 2012. Payment for
Ecosystem Services: The roles of positive incentives and information
sharing in stimulating adoption of silvopastoral conservation
practices. Agriculture, Ecosystems and Environment 156, 27–36.
https://doi.org/10.1016/j.agee.2012.04.017
Ghezehei, S.B., Everson, C.S., Annandale, J.G., 2015. Can productivity
and post-pruning growth of Jatropha curcas in silvopastoral systems be
regulated by manipulating tree spacing/arrangement without changing
tree density? Biomass and Bioenergy 74, 233–243.
https://doi.org/10.1016/j.biombioe.2015.01.007
Grieser, D., 2013. Mathematisches Problemlösen und Beweisen : Eine
Entdeckungsreise in die Mathematik.
https://doi.org/10.1016/S0065-2113(10)08005-3
Guimarães-Beelen, P.M., Berchielli, T.T., Beelen, R., Araújo Filho,
J., Oliveira, S.G. de, 2006. Characterization of condensed tannins
from native legumes of the Brazilian northeastern semi-arid. Scientia
Agricola 63, 522–528. https://doi.org/10.1590/S0103-90162006000600002

328
Haile, S.G., Nair, V.D., Nair, P.K.R., 2010. Contribution of trees to
carbon storage in soils of silvopastoral systems in Florida, USA.
Global Change Biology 16, 427–438.
https://doi.org/10.1111/j.1365-2486.2009.01981.x
Haque, I., Jutzi, S., Neate, P.J.H., 1986. Potentials of forage
legumes in farming systems of sub-Saharan Africa, in: Proceedings of a
Workshopheld at ILCA, Addis Ababa, Ethiopia, 16-19 September 1985.
International Livestock Centre for Africa, p. 583.
Hohnwald, S., 2016. Relative palatability and growth performance of
capoeira species as supplementary forages in the NE-Amazonia.
Agriculture, Ecosystems & Environment 218, 107–115.
https://doi.org/10.1016/j.agee.2015.11.015
Hohnwald, S., Trautwein, J., Camarão, A.P., Gerold, G., Wollny,
C.B.A., 2010. Resource Management and Rural Development Biomass
Production and Relative Palatability of Possible Supplementary Forage
Plants of the Northeastern Amazon.
Ibrahim, M., Villanueva, C., Casasola, F., Rojas, J., 2006. Sistemas
silvopastoriles como una herramienta para el mejoramiento de la

329
productividad y restauración de la integridad ecológica de paisajes
ganaderos. Pastos y Forrajes 29, 383–419.
Jat, M.L., Dagar, J.C., Sapkota, T.B., Yadvinder-Singh, Govaerts, B.,
Ridaura, S.L., Saharawat, Y.S., Sharma, R.K., Tetarwal, J.P., Jat,
R.K., Hobbs, H., Stirling, C., 2016. Climate Change and Agriculture:
Adaptation Strategies and Mitigation Opportunities for Food Security
in South Asia and Latin America. pp. 127–235.
https://doi.org/10.1016/bs.agron.2015.12.005
Khedri, L., Ghahramany, L., Ghazanfari, H., Pulido, F., 2018. A
quantitative study of pollarding process in silvopastoral systems of
Northern Zagros, Iran. Forest Systems 26, e018.
https://doi.org/10.5424/fs/2017263-11433
Larbi, A., Anyanwu, N.J., Oji, U.I., Etela, I., Gbaraneh, L.D.,
Ladipo, D.O., 2005. Fodder Yield and Nutritive Value of Browse Species
in West African Humid Tropics: Response to Age of Coppice Regrowth.
Agroforestry Systems 65, 197–205.
https://doi.org/10.1007/s10457-005-0922-x
Latawiec, A.E., Strassburg, B.B.N., Valentim, J.F., Ramos, F.,
Alves-Pinto, H.N., 2014. Intensification of cattle ranching production

330
systems: socioeconomic and environmental synergies and risks in
Brazil. Animal 8, 1255–1263. https://doi.org/10.1017/S1751731114001566
Lawson, G., Dupraz, C., Watté, J., 2019. Can Silvoarable Systems
Maintain Yield, Resilience, and Diversity in the Face of Changing
Environments? Agroecosystem Diversity 145–168.
https://doi.org/10.1016/B978-0-12-811050-8.00009-1
Leblanc, H.A., Nygren, P., McGraw, R.L., 2006. Green mulch
decomposition and nitrogen release from leaves of two Inga spp. in an
organic alley-cropping practice in the humid tropics. Soil Biology and
Biochemistry 38, 349–358.
https://doi.org/10.1016/J.SOILBIO.2005.05.012
Lee, M.A., Davis, A.P., Mizeck G.G.C., Manning, P., 2017. Forage
quality declines with rising temperatures, with implications for
livestock production and methane emissions. Biogeosciences 14,
1403–1417. https://doi.org/doi:10.5194/bg-14-1403-2017
Müller-Hansen, F., Heitzig, J., Donges, J.F., Cardoso, M.F.,
Dalla-Nora, E.L., Andrade, P., Kurths, J., Thonicke, K., 2019. Can
Intensification of Cattle Ranching Reduce Deforestation in the Amazon?
Insights From an Agent-based Social-Ecological Model. Ecological

331
Economics 159, 198–211. https://doi.org/10.1016/J.ECOLECON.2018.12.025
Murgueitio, E., Calle, Z., Uribe, F., Calle, A., Solorio, B., 2011a.
Native trees and shrubs for the productive rehabilitation of tropical
cattle ranching lands. Forest Ecology and Management 261, 1654–1663.
https://doi.org/10.1016/J.FORECO.2010.09.027
Muschler, R.G., Nair, P.K.R., Meléndez, L., 1993. Crown development
and biomass production of pollarded Erythrina berteroana, E. fusca and
Gliricidia sepium in the humid tropical lowlands of Costa Rica.
Agroforestry Systems 24, 123–143. https://doi.org/10.1007/BF00706887
Nunes, A.T., Cabral, D.L.V., Amorim, E.L.C., Santos, M.V.F. dos,
Albuquerque, U.P., 2016. Plants used to feed ruminants in semi-arid
Brazil: A study of nutritional composition guided by local ecological
knowledge. Journal of Arid Environments 135, 96–103.
https://doi.org/10.1016/J.JARIDENV.2016.08.015
OECD-FAO (Ed.), 2018. Chapter 6. Meat Market situation, in:
Agricultural Outlook 2018-2027. p. 150.
https://doi.org/10.1787/agr-outl-data-en
Ortega-Vargas, E., López-Ortiz, S., Burgueño-Ferreira, J.A., Campbell,

332
W.B., Jarillo-Rodríguez, J., 2013. Date of pruning of Guazuma
ulmifolia during the rainy season affects the availability,
productivity and nutritional quality of forage during the dry season.
Agroforestry Systems 87, 917–927.
https://doi.org/10.1007/s10457-013-9608-y
Pezo, D., Kass, M., Benavides, J., Romero, F. and Chaves, C., 1990. Potential of
legume tree fodders as animal feed in Central America. In Shrubs and tree fodders
for farm animals: proceedings of a workshop in Denpasar, Indonesia, 24-29 July
1989. IDRC, Ottawa, ON, CA.
Rengsirikul, K., Kanjanakuha, A., Ishii, Y., Kangvansaichol, K.,
Sripichitt, P., Punsuvon, V., Vaithanomsat, P., Nakamanee, G., Tudsri,
S., 2011. Potential forage and biomass production of newly introduced
varieties of leucaena (Leucaena leucocephala (Lam.) de Wit.) in
Thailand. Grassland Science 57, 94–100.
https://doi.org/10.1111/j.1744-697X.2011.00213.x
Serrão, E.A.S., Nepstad, D., Walker, R., 1996. Upland agricultural and
forestry development in the Amazon: sustainability, criticality and
resilience. Ecological Economics 18, 3–13.
https://doi.org/10.1016/0921-8009(95)00092-5

333
Sharrow, S.H., Ismail, S., 2004. Carbon and nitrogen storage in
agroforests, tree plantations, and pastures in western Oregon, USA.
Agroforestry Systems 60, 123–130.
https://doi.org/10.1023/B:AGFO.0000013267.87896.41
Sobanski, N., Marques, M.C.M., 2014. Effects of soil characteristics
and exotic grass cover on the forest restoration of the Atlantic
Forest region. Journal for Nature Conservation 22, 217–222.
https://doi.org/10.1016/J.JNC.2014.01.001
Vaca, R.A., Rodiles-Hernández, R., Ochoa-Gaona, S., Taylor-Aquino,
N.E., Obregón-Viloria, R., Díaz-García, D.A., Navarrete-Gutiérrez,
D.A., 2019. Evaluating and supporting conservation action in
agricultural landscapes of the Usumacinta River Basin. Journal of
Environmental Management 230, 392–404.
https://doi.org/10.1016/J.JENVMAN.2018.09.055




















