
Somatic cell hybrid deletion map of human chromosome 18
6
GENOMICS 13, l-6 (1992) Somatic Cell Hybrid Deletion Map of Human Chromosome 18 ANTONIE D. KLINE,*‘t KATHERINE ROJAS, * REEMA MEWAR,* DEBORAH MOSHINSKY, * AND JOAN OVERHAUSER**’ *Department of Biochemistry and Molecular Biology and tDivision of Medical Genetics, Jefferson Medical College, Thomas Jefferson University, Philadelphia, Pennsylvania 19 107 Received November 5. 1991;revisedJanuary 7, 1992 The creation of a physical map of chromosome 18 will be useful for the eventual identification of specific chromosomal regions that are critical in the occurrence of Edwards syn- drome, the 18q- syndrome, and the 18p- syndrome. To begin the investigation of these syndromes, a physical map has been constructed to order random DNA fragments to specific por- tions of chromosome 18. A set of somatic cell hybrids that re- tain deletions or translocations involving chromosome 18 has been isolated and characterized. Over 200 lambda phage from a chromosome 18-specific library have been localized to 11 distinct regions of chromosome 18 using the chromosomal breakpoints present in the somatic cell hybrids. o 1992 Academic Press, Inc. INTRODUCTION Chromosome 18 is associated with specific aneuploi- dies including trisomy 18 or Edwards syndrome, 18p- syndrome, and 18q- syndrome. A range of partial dupli- cations and deletions have been reported in the litera- ture, suggesting that altered dosage of most of the chro- mosome produces potentially viable offspring (Fryns et al., 1978, Wilson et al., 1990, Turleau et al., 1980; Miller et al., 1990). It should be possible therefore to dissect the chromosome into regions containing genes involved in the distinctive phenotype associated with each syn- drome. Specific loci of medical significance have been mapped to chromosome 18. Chromosome 18 rearrangements ap- pear to play an important role in some types of cancer development. The bcl-2 gene located at 18q21.3 is in- volved in a 14;18 translocation found in human follicular B-cell lymphomas (Tsujimoto and Croce, 1986). Loss of expression of the tumor suppressor gene DCC, also mapped to 18q21.3, has been associated with colorectal cancers (Fearon et aZ.,1990). The differential expression of the genes involved in these cancers has not been in- vestigated in individuals with 18q deletions or duplica- tions. In addition, the myelin basic protein (MBP) gene has been localized to 18q22-q23 (Kamholz et al., 1987). Since both trisomy 18 and the 18q- syndrome can result ’ To whom correspondence should be addressed. in neurological dysfunction, an abnormality in the MBP gene may contribute to these phenotypes. Clarification of the potential role of the MBP gene in these aneu- ploidy syndromes may lead to increased understanding of the phenotypes associated with these disorders. To elucidate the roles of specific genes in the etiology of the chromosome 18 dosage syndromes, a physical map of chromosome 18 has been constructed. Individuals with deletions, duplications, and translocations involv- ing chromosome 18 have been identified. Transformed lymphoblasts derived from these individuals have been used to isolate somatic cell hybrids containing the rearranged human chromosome 18. DNA fragments from a chromosome 18-specific lambda phage library have been mapped with respect to the breakpoints in these individuals. MATERIALS AND METHODS Karyotypes andphenotypes of individuals with alterations of chromo- some 18. Chromosomal analysis on all of the individuals was per- formed using standard G-banding procedures (Verma and Babu, 1989). The phenotypes and karyotypes are shown in Table 1. Subject 1 is a normal individual with no alterations of chromosome 18. Subject 2 has a balanced translocation involving 5p and 18q and is clinically normal. She has a child previously described (Overhauser et ai, 1986), with the unbalanced karyotype 46,XY,-5,+der(5)t(5;18)(p15.3;qll.2), who has all the clinical characteristics of trisomy 18. Subjects 3 and 4 have balanced translocations, are clinically normal, and have no off- spring with unbalanced karyotypes. Subject 5 is clinically normal, with a mosaic karyotype; 78% of peripheral blood cells have a 1218 translocation and 22% have trisomy 12. Subjects 6, 7, and 12 all have the lap- syndrome. Subject 8 is a mildly mentally retarded female with a marker chromosome most likely of chromosome 18 origin. Sub- ject 9 has a balanced translocation involving chromosomes 14 and 18, is clinically normal, and has a child with the unbalanced karyotype 46,XY,-l8,+der(l8)t(l&l4)(q22;q23) displaying the 18q- syndrome. Subject 10 has a balanced translocation, is clinically normal, and has a child with the unbalanced karyotype 46,XY,-13,+t(13;18)(13qter- 13p13::18qlZ.l-laqter), who has clinical features consistent with tri- somy 18. Subject 11 has a terminal deletion of chromosome 18 result- ing in the 18q- syndrome phenotype. Cell lines and somatic cell hybrids. Peripheral leukocytes were ob- tained from the above individuals with chromosome 18 alterations. The lymphocytes were transformed with Epstein-Barr virus (EBV) and subsequently used for the isolation of somatic cell hybrids. The Chinese hamster ovary (CHO) cell line UCW206 contains a temperature-sensitive mutation in the aspargine-tRNA synthetase (AARS) gene, resulting in nonviability at 39°C (Cirullo et al., 1983). 1 osss-x43/92 $3.00 Copyright 0 1992 by Academic Press, Inc. All rights of reproduction in any form reserved.
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Somatic cell hybrid deletion map of human chromosome 18

GENOMICS 13, l-6 (1992)
Somatic Cell Hybrid Deletion Map of Human Chromosome 18
ANTONIE D. KLINE,*‘t KATHERINE ROJAS, * REEMA MEWAR,* DEBORAH MOSHINSKY, * AND JOAN OVERHAUSER**’
*Department of Biochemistry and Molecular Biology and tDivision of Medical Genetics, Jefferson Medical College, Thomas Jefferson University, Philadelphia, Pennsylvania 19 107
Received November 5. 1991;revisedJanuary 7, 1992
The creation of a physical map of chromosome 18 will be useful for the eventual identification of specific chromosomal regions that are critical in the occurrence of Edwards syn- drome, the 18q- syndrome, and the 18p- syndrome. To begin the investigation of these syndromes, a physical map has been constructed to order random DNA fragments to specific por- tions of chromosome 18. A set of somatic cell hybrids that re- tain deletions or translocations involving chromosome 18 has been isolated and characterized. Over 200 lambda phage from a chromosome 18-specific library have been localized to 11 distinct regions of chromosome 18 using the chromosomal breakpoints present in the somatic cell hybrids. o 1992 Academic
Press, Inc.
INTRODUCTION
Chromosome 18 is associated with specific aneuploi- dies including trisomy 18 or Edwards syndrome, 18p- syndrome, and 18q- syndrome. A range of partial dupli- cations and deletions have been reported in the litera- ture, suggesting that altered dosage of most of the chro- mosome produces potentially viable offspring (Fryns et al., 1978, Wilson et al., 1990, Turleau et al., 1980; Miller et al., 1990). It should be possible therefore to dissect the chromosome into regions containing genes involved in the distinctive phenotype associated with each syn- drome.
Specific loci of medical significance have been mapped to chromosome 18. Chromosome 18 rearrangements ap- pear to play an important role in some types of cancer development. The bcl-2 gene located at 18q21.3 is in- volved in a 14;18 translocation found in human follicular B-cell lymphomas (Tsujimoto and Croce, 1986). Loss of expression of the tumor suppressor gene DCC, also mapped to 18q21.3, has been associated with colorectal cancers (Fearon et aZ., 1990). The differential expression of the genes involved in these cancers has not been in- vestigated in individuals with 18q deletions or duplica- tions. In addition, the myelin basic protein (MBP) gene has been localized to 18q22-q23 (Kamholz et al., 1987). Since both trisomy 18 and the 18q- syndrome can result
’ To whom correspondence should be addressed.
in neurological dysfunction, an abnormality in the MBP gene may contribute to these phenotypes. Clarification of the potential role of the MBP gene in these aneu- ploidy syndromes may lead to increased understanding of the phenotypes associated with these disorders.
To elucidate the roles of specific genes in the etiology of the chromosome 18 dosage syndromes, a physical map of chromosome 18 has been constructed. Individuals with deletions, duplications, and translocations involv- ing chromosome 18 have been identified. Transformed lymphoblasts derived from these individuals have been used to isolate somatic cell hybrids containing the rearranged human chromosome 18. DNA fragments from a chromosome 18-specific lambda phage library have been mapped with respect to the breakpoints in these individuals.
MATERIALS AND METHODS
Karyotypes andphenotypes of individuals with alterations of chromo- some 18. Chromosomal analysis on all of the individuals was per- formed using standard G-banding procedures (Verma and Babu, 1989). The phenotypes and karyotypes are shown in Table 1. Subject 1 is a normal individual with no alterations of chromosome 18. Subject 2 has a balanced translocation involving 5p and 18q and is clinically normal. She has a child previously described (Overhauser et ai, 1986), with the unbalanced karyotype 46,XY,-5,+der(5)t(5;18)(p15.3;qll.2), who has all the clinical characteristics of trisomy 18. Subjects 3 and 4 have balanced translocations, are clinically normal, and have no off- spring with unbalanced karyotypes. Subject 5 is clinically normal, with a mosaic karyotype; 78% of peripheral blood cells have a 1218 translocation and 22% have trisomy 12. Subjects 6, 7, and 12 all have
the lap- syndrome. Subject 8 is a mildly mentally retarded female with a marker chromosome most likely of chromosome 18 origin. Sub- ject 9 has a balanced translocation involving chromosomes 14 and 18, is clinically normal, and has a child with the unbalanced karyotype 46,XY,-l8,+der(l8)t(l&l4)(q22;q23) displaying the 18q- syndrome. Subject 10 has a balanced translocation, is clinically normal, and has a child with the unbalanced karyotype 46,XY,-13,+t(13;18)(13qter- 13p13::18qlZ.l-laqter), who has clinical features consistent with tri- somy 18. Subject 11 has a terminal deletion of chromosome 18 result- ing in the 18q- syndrome phenotype.
Cell lines and somatic cell hybrids. Peripheral leukocytes were ob- tained from the above individuals with chromosome 18 alterations. The lymphocytes were transformed with Epstein-Barr virus (EBV) and subsequently used for the isolation of somatic cell hybrids.
The Chinese hamster ovary (CHO) cell line UCW206 contains a temperature-sensitive mutation in the aspargine-tRNA synthetase (AARS) gene, resulting in nonviability at 39°C (Cirullo et al., 1983).
1 osss-x43/92 $3.00 Copyright 0 1992 by Academic Press, Inc.
All rights of reproduction in any form reserved.

2 KLINE ET AL.
TABLE 1
Phenotypes and Karyotypes of Individuals with Chromosome 18 Rearrangements
Subject Karyotype
1 46,XY 2 46,XX,t@;l@(p15.2;q12.2)
3 46,XY,t(l8;2l)(pll;qll.2) 4 46,XY,t(7;18)(q22;q22.1) 5 45,XX,-18,+der(l2)t(12;18)(pll;qll)/47,XX,+12 6 46,XX,del(l8)(pll)/47,XX,+iso(18q) 1 45,Xx;-13;-18;+t(13;18)(13qter-cent-18qter)
8 47,XX,+de1(18)(ql1.2) 9 46,XY,t(14;18)(q32;q21)
10 46,XY,t(13;18)(p13;q12.1) 11 46,XX,de1(18)(21.3) 12 46,XX,de1(18)(pll)
u Offsnrina with unbalanced translocation has Edwards svndrome uhenotype. . L
h Unbalanced translocation phenotype unknown. ”
Phenotype
Normal Normal” Normal* Normal”
Normal 18p- syndrome 18p- syndrome Trisomy 18p syndrome Normal* Normal” 18q- syndrome 18p- syndrome
Hybrid cell line
HHW324 HHW659 HHW778
HHW813 JH176 JH158 dH184
JH211 JH226 JH232 JH237 JH252
The corresponding human gene is located on human chromosome 18 (Cirullo et al., 1983). Each transformed cell line (subjects 1-7, 9-12)
was fused with UCW206 in the presence of polyethylene glycol and placed at the restrictive temperature, allowing only the somatic cell
hybrids that retain a human chromosome 18 to survive.
The transformed cell line derived from subject 8 contained a marker chromosome thought to be of 18~ origin. To obtain this marker chro- mosome in a cell hybrid, without including either normal human chro-
mosome 18, somatic cell fusions were performed using the CHO cell line, UCW56, which contains a temperature-sensitive mutation in the leucyl-tRNA synthetase gene and results in nonviability at 39°C (Dana and Wasmuth, 1982). The equivalent human gene is located on human chromosome 5 (Dana and Wasmuth, 1982). The resulting so- matic cell hybrids derived from the fusion with this hamster cell line
were grown at the restrictive temperature and screened for the pres- ence of the marker chromosome.
Screening of the somatic cell hybrids. Somatic cell hybrids were
screened using the polymerase chain reaction (PCR). Cell clones were transferred to 24-well plates and grown until confluent. The cells were trypsinized and half were transferred to a new well. The remaining cells were placed in a microfuge tube and sedimented. The cells were resuspended in 50 ~11X PCR buffer (10 mM Tris-HCl, pH 8.3,50 mM KCl, 1.5 mM MgCl,, 0.01% w/v gelatin) containing 100 @g/ml pro- teinase K. The cells were incubated at 55°C for 1 h and then at 94°C
for 10 min. The PCR was carried out in a total volume of 50 ~1 with 5 ~1 of the DNA sample, 1 WM each primer, in 10 mMTris-HCl, pH 8.3,50 mM KCl, 3 n&f MgCl,, 0.01% w/v gelatin, 2% formamide, 250 fiLM each dNTP, and 0.6 unit AmpliTaq DNA polymerase (Perkin-Elmer Cetus, Norwalk, CT). The sets of primers, the size of each PCR prod- uct, and their location are shown in Table 2. Amplification was per- formed by 35 cycles of denaturation at 96°C (1.5 min), annealing at 55°C (45 s), extension at 72°C (30 s), and a final extension time of 7 min at 72°C. Fifteen microliters of each sample was loaded onto a 1.5% agarose gel in 1X TAE buffer.
Somatic cell hybrids which gave the appropriate PCR amplification products, suggesting retention of the rearranged chromosome 18, were then examined by alkaline-Giemsa staining and trypsin-Giemsa banding to confirm the PCR results. The cell hybrids were maintained at 39°C to ensure retention of the human chromosome 18. Because the process of cell fusion can lead to random chromosomal breakages and rearrangements, at least two independent cell hybrids with the deleted chromosome 18 were isolated from each cell fusion. This was to ensure that the chromosome 18 in the cell hybrids was an accurate representa- tion of the deleted chromosome 18 present in the individual. Only when the subsequent molecular analyses of the deleted chromosomes 18 in the cell hybrids were identical, were the somatic cell hybrids considered an accurate representation of the patients’ deleted chro-
mosomes 18. For simplicity, we have included only the data from one somatic cell hybrid from each cell fusion.
Identification of lambda phage containing chromosome 18-specific inserts. A somatic cell hybrid, HHW324, contains a normal chromo- some 18 as its only human genetic material. Using high-molecular- weight DNA from this somatic cell hybrid, a lambda phage library was constructed in the vector EMBL4 as previously described (Carlock and Wasmuth, 1985). The lambda phage were plated and screened for the presence of human repetitive sequences as described previously (Carlock and Wasmuth, 1985). Over 200 positive plaques were puri- fied and lambda phage DNA was isolated using the procedure de- scribed by Helms et al. (1985).
Blotting and hybridization. High-molecular-weight DNA was ex- tracted from hamster cells, human cells, and the somatic cell hybrids, digested to completion with EcoRI, and electrophoresed through a 0.8% agarose gel. The DNA was transferred to nylon membranes by capillary action in the presence of 0.4 M NaOH. Lambda phage DNA was labeled with “P using the random primer method (Feinberg and Vogelstein, 1989). Prehybridization was performed in 7% SDS, 1% BSA, 10% PEG 8000, 0.125 M Na,PO, heptahydrate, 2 mM EDTA, 250 mM NaCI, 0.2% sodium pyrophosphate for at least 1 h. Hybridiza- tion was performed in the same solution containing the denatured labeled DNA, with 100 pg sheared denatured human placenta DNA and 400 fig sheared denatured salmon sperm DNA at 65°C for 16-24 h. The membranes were washed in 2X SSC, 0.2% SDS at 65°C for 30 min, followed by a wash in 0.5X SSC, 0.2% SDS at 65°C for 30 min. The membranes were subjected to autoradiography for 1 to 3 days.
RESULTS
Analysis of Somatic Cell Hybrids Using PCR
To create a physical map of chromosome 18, trans- formed cells were produced from individuals with break- points in various regions of the chromosome, including individuals with the lap- syndrome, Edwards syndrome (trisomy 18), and the 18q- syndrome. The karyotypes and phenotypes of these individuals have been described under Materials and Methods. Since the exact location of the selectable marker AARS was not known, initial fusions were performed using transformed lymphoblasts containing balanced translocations, ensuring retention of one of the derivative chromosomes containing the se- lectable marker. Somatic cell hybrids were isolated from

MAPPING OF CHROMOSOME 18 3
DNA fragment
TABLE 2
PCR Primers for Somatic Cell Hybrid Screening
Location Primer PCR size (kb)
324-44E-G
324-22E-H
324-128E-G
92H-A
18~11.3
18q12.2-q22.3
18q23
5pll-p13
CAGACAACCTACAAAGGAGC 0.40 GGACTGTTGATAATTTCTCA GGATCTGAGCTAGAATAACT 0.48 TGTTTGGCTGAGTACGTGTGTA ATGACAGCACTCTTCACACA 0.30 CCACTAAACAGTAACTTCCT TTGTCAACAGCAGCGACTCA 0.21 GAGCTGTTGCGTGTTAATTC
all of these lymphoblast lines and used for analysis of the chromosome 18-specific lambda phage inserts.
The initial screening of somatic cell hybrids from pa- tients 2-4 was performed by karyotype analysis without the aid of PCR. Several lambda phage were subsequently mapped to the ends of chromosome 18 and a more proxi- mal region of 18q based on initial mapping data. Single- copy sequences from three of these lambda phage were subcloned and sequenced to identify appropriate PCR primer sequences. These 18~ and 18q primers were used to screen the remaining somatic cell hybrids by PCR analysis (Table 2).
DNA isolated from a somatic cell hybrid was initially subjected to PCR, using the 18p and two 18q primers. PCR analysis from somatic cell hybrids derived from a fusion with lymphoblasts from subject 5 and UCW206 is shown in Fig. 1A. The somatic cell hybrids obtained after selection should contain either the normal chro- mosome 18 (amplification between all three primer sets), the normal chromosome 18 and the derivative chromosome 12 (amplification between all three primer
1 2 1 2
r--l rlr-l
FIG. 1. PCR analysis of somatic cell hybrids. Cells were harvested and treated as stated under Materials and Methods. (A) Two hybrids from a fusion using lymphoblasts from subject 5, with deleted material of 18~. DNA from each cell hybrid was used to amplify DNA between three chromosome la-specific PCR primers described in Table 2. (B) Two cell hybrids from a fusion using the cells from subject 8. (See text for explanation.)
sets), or the derivative chromosome 12 alone (amplifica- tion between the 18q centromeric and distal 18q primer sets only). Sample 1 shows amplification between all three primer sets, indicating the presence of a normal chromosome 18 in this cell hybrid, and sample 2 shows amplification with only the two 18q primer sets, indicat- ing the presence of the derivative chromosome 12.
PCR screening was also used when the altered chro- mosome 18 could not be used to retain the selectable marker. In subject 8, interest was in the marker chromo- some. Initial karyotype analysis of the leukocytes sug- gested that the marker chromosome was an iso(l8p) or 18p with a portion of proximal 18q. Several attempts at fusing this cell line with UCW206 resulted in somatic cell hybrids which always retained a normal human chromosome 18. Results from cell fusions with other lymphoblast cell lines determined that the AARS select- able marker was located on the distal half of 18q, ex- plaining the inability to obtain somatic cell hybrids con- taining only the marker chromosome. To obtain the marker chromosome without retention of either normal chromosome 18, fusions were performed with UCW56, as described under Materials and Methods. In addition, sequencing and PCR primers were derived from an anon- ymous clone, 92H-A, which had been previously mapped to 5pll-p13 (Overhauser et al., 1986). The resulting so- matic cell hybrids were then screened by PCR for the nonselective retention of the marker chromosome dem- onstrated by the amplification of the 18p-specific primers but not the 18q primers. These results are shown in Fig. 1B. Sample 1 shows amplification from a somatic cell hybrid that has retained chromosome 18p material, but not 18q material, suggesting nonselective retention of the marker chromosome, whereas sample 2 shows DNA from a somatic cell hybrid that has retained a chromosome 5, with no retention of chromosome 18 material.
Cell hybrids that produced the pattern of PCR ampli- fication consistent with retention of the abnormal deriv- ative of chromosome 18 were analyzed by trypsin- Giemsa banding to ensure that chromosomal rearrange- ments had not occurred. At least two independent somatic cell hybrids were obtained from each lympho- blast cell line and analyzed. This strategy was used to confirm that the retained abnormal human chromosome 18 was an accurate representation of the aneuploidy
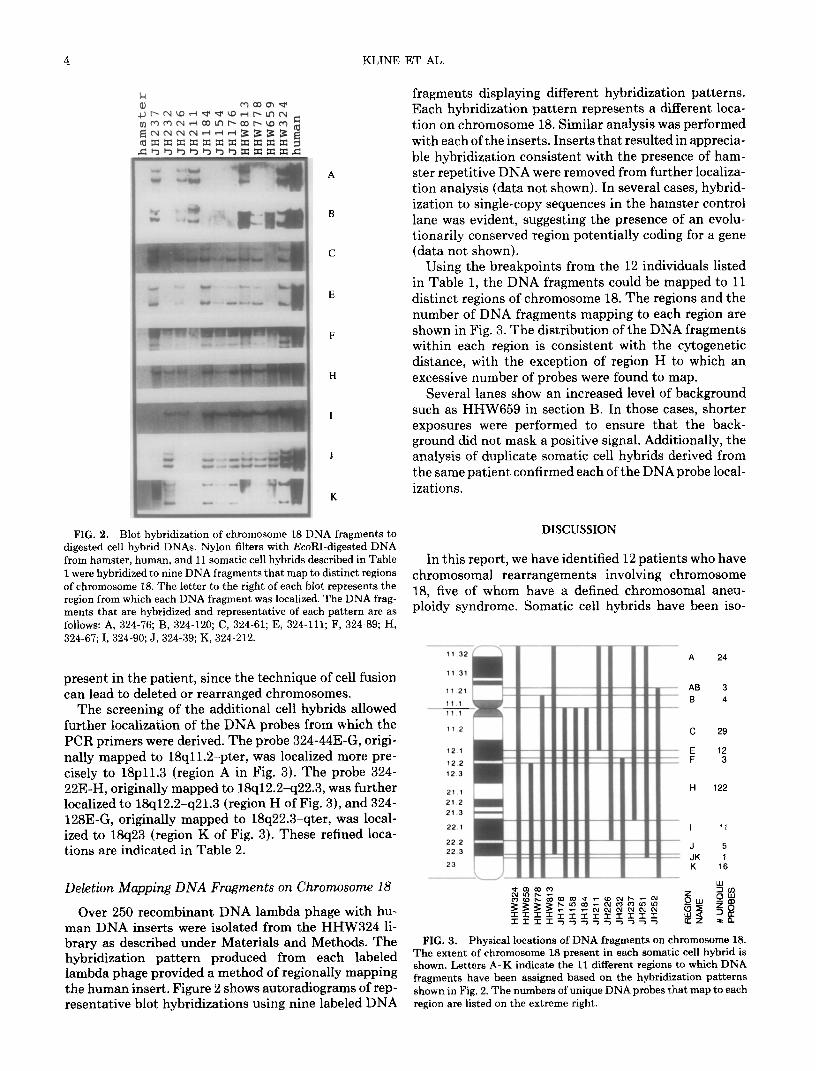
4 KLINE ET AL.
fragments displaying different hybridization patterns. Each hybridization pattern represents a different loca- tion on chromosome 18. Similar analysis was performed with each of the inserts. Inserts that resulted in apprecia- ble hybridization consistent with the presence of ham- ster repetitive DNA were removed from further localiza- tion analysis (data not shown). In several cases, hybrid- ization to single-copy sequences in the hamster control lane was evident, suggesting the presence of an evolu- tionarily conserved region potentially coding for a gene (data not shown).
Using the breakpoints from the 12 individuals listed in Table 1, the DNA fragments could be mapped to 11 distinct regions of chromosome 18. The regions and the number of DNA fragments mapping to each region are shown in Fig. 3. The distribution of the DNA fragments within each region is consistent with the cytogenetic distance, with the exception of region H to which an excessive number of probes were found to map.
Several lanes show an increased level of background such as HHW659 in section B. In those cases, shorter exposures were performed to ensure that the back- ground did not mask a positive signal. Additionally, the analysis of duplicate somatic cell hybrids derived from the same patient confirmed each of the DNAprobe local- izations.
FIG. 2. Blot hybridization of chromosome 18 DNA fragments to DISCUSSION digested cell hybrid DNAs. Nylon filters with EcoRI-digested DNA from hamster, human, and 11 somatic cell hybrids described in Table In this report, we have identified 12 patients who have 1 were hybridized to nine DNA fragments that map to distinct regions of chromosome 18. The letter to the right of each blot represents the
chromosomal rearrangements involving chromosome
region from which each DNA fragment was localized. The DNA frag- 18, five of whom have a defined chromosomal aneu-
ments that are hybridized and representative of each pattern are as ploidy syndrome. Somatic cell hybrids have been iso-
follows: A, 324-76; B, 324-120; C, 324-61; E, 324-111;.F, 324-89; H, 324-67; I, 324-90; J, 324-39; K, 324-212.
present in the patient, since the technique of cell fusion can lead to deleted or rearranged chromosomes.
The screening of the additional cell hybrids allowed further localization of the DNA probes from which the PCR primers were derived. The probe 324-44E-G, origi- nally mapped to 18qll.2-pter, was localized more pre- cisely to 18~11.3 (region A in Fig. 3). The probe 324- 22E-H, originally mapped to 18q12.2-q22.3, was further localized to 18q12.2-q21.3 (region H of Fig. 3), and 324- 128E-G, originally mapped to 18q22.3-qter, was local- ized to 18q23 (region K of Fig. 3). These refined loca- tions are indicated in Table 2.
12.1
12.2
12.3
21.1
21 2
21.3
22.1
22.2
22.3
23
Deletion Mapping DNA Fragments on Chromosome 18
Over 250 recombinant DNA lambda phage with hu- man DNA inserts were isolated from the HHW324 li- brary as described under Materials and Methods. The hybridization pattern produced from each labeled lambda phage provided a method of regionally mapping the human insert. Figure 2 shows autoradiograms of rep- resentative blot hybridizations using nine labeled DNA
-
A 24
- AB 3 _
B 4 -
JK 1
K 16
FIG. 3. Physical locations of DNA fragments on chromosome 18. The extent of chromosome 18 present in each somatic cell hybrid is shown. Letters A-K indicate the 11 different regions to which DNA fragments have been assigned based on the hybridization patterns shown in Fig. 2. The numbers of unique DNA probes that map to each region are listed on the extreme right.

MAPPING OF CHROMOSOME 18 5
lated for each of the abnormal derivatives of chromo- some 18 to increase the efficiency of mapping a large number of DNA fragments to specific regions of the chromosome. Using over 200 random DNA fragments from a chromosome l&specific library, these DNA frag- ments have been divided into 11 distinct regions.
With the cells lines analyzed to date, the critical re- gions for several of the syndromes can be further de- fined. The critical segment for the 18p- syndrome pres- ently resides from 18pter + 18~11.21. Only the most proximal portion of short arm can be eliminated from involvement in the etiology of the syndrome using the patients analyzed in this paper. Additional patients with the 18p- syndrome are needed to characterize this criti- cal region further.
There has been controversy in the literature regarding the location of the critical region for trisomy 18 syn- drome. Matsuoka et al. (1981) analyzed 20 patients and concluded that the region that must be present in three copies to produce the trisomy 18 phenotype is located in 18q21. Analysis of additional partial trisomy 18 patients by Fryns et al. (1978,1979) and Miicke et al. (1982) con- cluded that the crucial bands for the appearance of the trisomy 18 phenotype were proximal to 18q12.2. There are several possible explanations for these findings: (1) There is no specific critical chromosomal segment that, when present in three copies, results in all of the clinical characteristics of trisomy 18. Wilson et al. (1990) have suggested that multiple chromosomal areas may be in- volved in the appearance of the complete trisomy 18 phe- notype. (2) The limitations of cytogenetics may make it difficult to accurately determine the exact chromosomal regions duplicated, rendering the correlation with the clinical phenotype inaccurate. The advantage with mo- lecular analyses of chromosomal duplications and trans- locations is that chromosomal regions in different pa- tients can be compared directly at the DNA level, elimi- nating possible misinterpretations of karyotypes.
Using the analyses of patients in this study, the criti- cal region for the Edwards syndrome appears to lie distal to 18q12.2. This is based on one patient with a duplica- tion from 18pter --f 18q12.1 without the typical pheno- type and two patients with duplications from 18q12.2 + 18qter and all of the characteristics of trisomy 18. Fur- ther patient samples are needed to localize the critical region to a more defined region.
In the present patient samples only one patient has the 18q- syndrome. The localization of the critical re- gion for this syndrome is limited to the region 18q21.3 + 18qter. A patient has been reported having a terminal deletion of chromosome 18 without the phenotype asso- ciated with this syndrome (Kamholz et al., 1988), sug- gesting that a critical chromosomal segment may in fact lie within 18q21.3-q22.3. It will be important to compare these samples and those from additional patients with the 18q- syndrome at the molecular level to confirm this hypothesis.
With the creation of a physical map and the sublocali- zation of multiple DNA fragments along chromosome
18, it will be of additional benefit to identify polymor- phic markers within these mapped DNA fragments. To- ward this end, each of the DNA fragments has been screened for the presence of large tracts of dinucleotide repeats which are known to be highly polymorphic (Weber and May, 1989) and have been previously mapped to chromosome 18 (Weber and May, 1990a,b). Preliminary data suggest that at least 50 of the lambda phage mapping to nine of the regions contain dinucleo- tide repeats (data not shown). Cloning, sequencing, and screening of these dinucleotide repeats will allow the identification of RFLPs that are known to span all re- gions of the chromosome.
ACKNOWLEDGMENTS
We thank Dr. Ladonna Immken, University of Texas Medical School at Houston; Dr. David Ledbetter, Baylor College of Medicine; Dr. Frank Greenberg, Baylor College of Medicine; Dr. Gary Silver- man, Washington University School of Medicine; and the Chromo- some 18 Registry and Research Society. This work was supported by NIH Grant HG-00167.
REFERENCES
Carlock, L. R., and Wasmuth, J. J. (1985). Molecular approach to analyzing the human 5p deletion syndrome, cri du chat. Somat. Cell Mol. Genet. 11: 267-276.
Cirullo, R. E., Arredondo-Vega, F. X., Smith, M., and Wasmuth, J. J. (1983). Isolation and characterization of interspecific heat-resistant hybrids between a temperature-sensitive Chinese hamster cell asparaginyl-tRNA synthetase mutant and normal human leuko- cytes: Assignment of human asnS gene to chromosome 18. Somat. Cell Genet. 9: 215-233.
Dana, S., and Wasmuth, J. J. (1982). Linkage of the Zeus, emtB and chr genes on chromosome 5 in humans and expression of human genes encoding protein synthetic components in human-Chinese
hamster hybrids. Somat. Cell Genet. 8: 245-264.
Fearon, E. R., Cho, K. R., Nigro, J. M., Kern, S. E., Simons, J. W., Ruppert, J. M., Hamilton, S. R., Preisinger, A. C., Thomas, G., Kinzler, K. W., and Vogelstein, B. (1990). Identification of a chro- mosome 18q gene that is altered in colorectal cancers. Science 247: 49-56.
Feinberg, A. P., and Vogelstein, B. (1989). Addendum: A technique for radiolabeling DNA restriction endonuclease fragments to high spe- cific activity. Anal. Biochem. 137: 266-267.
Fryns, J. P., Detavernier, F., Van Fleteren, A., and Van Den Berghe, H. (1978). Partial trisomy 18q in a newborn with typical 18 trisomy phenotype. Hum. Genet. 44: 201-205.
Fryns, J. P., Vinken, L., Marien, J., and Van Den Berghe, H. (1979). Partial trisomy 18q12, due to intrachromosomal duplication, is not associated with typical 18 trisomy phenotype. Hum. Genet. 46: 341- 344.
Helms, C., Graham, M. Y., Dutchik, J. E., and Olson, M. V. (1985). A new method for purifying lambda DNA from phage lysatas. DNA 4: 39-49.
Kamholz, J., Fischbeck, K., Ritter, A., Zackai, E., McDonald, D., and Emanuel, B. (1988). Segmental spinal muscular atrophy associated with a partial deletion of chromosome 18. Am. J. Hum. Genet. 43: AllO.
Kamholz, J., Spielman, R., Gogolin, K., Modi, W., O’Brien, S., and Lazzarini, R. (1987). The human myelin-basic-protein gene: Chro- mosomal localization and RFLP analysis. Am. J. Hum. Genet. 40: 365-373.

6 KLINE ET AL.
Matsuoka, R., Matsuyama, S., Yamamoto, Y., Kuroki, Y., and Matsui, I. (1981). Trisomy 18q. Hum. Genet. 57: 78-82.
Miller, G., Mowrey, P. N., Hopper, K. D., Frankel, C. A., and Ladda, R. L. (1990). Neurologic manifestations in 18q- syndrome. Am. J. Med. Genet. 37: 128-132.
Mticke, J., Trautmann, K., Sandig, K.-R., and Theile, H. (1982). The crucial band for phenotype of trisomy 18. Hum. Genet. 60: 205.
Overhauser, J., Beaudet, A. L., and Wasmuth, J. J. (1986). A fine structure physical map of the short arm of chromosome 5. Am. J. Hum. Genet. 39: 562-572.
Tsujimoto, Y., and Croce, C. M. (1986). Analysis of the structure, transcripts, and protein products of bcl-2, the gene involved in hu- man follicular lymphoma. Proc. N&l. Acud. Sci. USA 83: 5214- 5218.
Turleau, C., Chavin-Colin, F., Narbouton, R., Asensi, D., and de
Grouchy, J. (1980). Trisomy 18q-. Trisomy mapping of chromo- some 18 revisited. Clin. Genet. 18: 20-26.
Verma, R. S., and Babu, A. (1989). “Human Chromosomes: Manual of Basic Techniques,” pp. 47-49. Pergamon Press, Elmsford, NY.
Weber, J. L., and May, P. E. (1989). Abundant class of human DNA polymorphisms which can be typed using the polymerase chain reac- tion. Am. J. Hum. Genet. 44: 388-396.
Weber, J. L., and May, P. E. (1990). Dinucleotide repeat polymor- phism at the D18S34 locus. Nucl. Acids Res. 18: 2201.
Weber, J. L., and May, P. E. (1990). Dinucleotide repeat polymor- phism at the D18S35 locus. Nucl. Acids Res. 18: 6465.
Wilson, G. N., Heller, K. B., Elterman, R. D., and Schneider, N. R. (1990). Partial trisomy 18 with minimal anomalies: Lack of corre- spondence between phenotypic manifestations and triplicated loci along chromosome 18. Am. J. Med. Genet. 36: 506-510.




















