
Soil microbial community structure and function relationships: A heat stress experiment
13
Soil microbial community structure and function are significantly affected by long-term organic and mineral fertilization regimes in the North China Plain Juan Li a , Julia Mary Cooper b , Zhi'an Lin a , Yanting Li a , Xiangdong Yang a , Bingqiang Zhao a, * a Key Laboratory of Plant Nutrition and Fertilizer, Ministry of Agriculture / Institute of Agricultural Resources and Regional Planning, Chinese Academy of Agricultural Sciences, Beijing 100081, PR China b Nafferton Ecological Farming Group, Newcastle University, Nafferton Farm, Stocksfield, Northumberland NE43 7XD, UK A R T I C L E I N F O Article history: Received 21 February 2015 Received in revised form 2 June 2015 Accepted 1 July 2015 Available online xxx Keywords: Long term fertilization Organic manure Soil extracellular enzyme activity Soil bacterial community Soil quality indicators A B S T R A C T An improved understanding of the complex interactions and relationships in the soil ecosystem is essential to predict the impact of farming practices on soil quality and its capacity for agricultural production. This study aims to improve our understanding of the impacts of fertilization strategy on key indicators of soil biological and chemical quality. We studied soils from a winter wheat-summer maize rotational experiment in the North China Plain with six different fertility treatments: no amendments (CK); standard mineral fertilizer treatment (SMF) or standard organic manure treatment (SMA) reflecting local farmer practice; mixed treatment with fertilizer and manure at half the rates for the SMF and SMA treatments (1/2 SMF + 1/2 SMA); double mineral fertilizer treatment (DMF); and double organic manure treatment (DMA). Soil organic C (SOC), total N (TN), total P (TP), pH, and dissolved organic C (DOC) and N (DON) and microbial biomass C (Cmic) and N (Nmic) were determined using standard methods. Soil bacterial community structure was assessed by denaturing gradient gel electrophoresis (DGGE), and activities for 10 extracellular enzymes (EEAs) were measured as indicators of soil function. Repeated application of either organic manure or mineral fertilizer increased SOC, TN, TP, DOC, DON, Cmic and Nmic, and decreased soil pH. Higher rates of organic manure fertilization significantly affected soil chemical properties compared to the lower rate. Soil bacterial community structure was significantly altered by the long-term fertilization regimes and diversity was significantly higher in the double manure rate treatment relative to mineral fertilizer. The higher urease, a-glycosidase, N-acetyl-b-glucosami- nidase, L-leucine aminopeptidase (involved in N cycling), b-glucosidase, b-xylosidase and b-cellobio- sidase (involved in C cycling), and alkaline phosphatase (involved in P cycling) activities for organic manure fertilized soils reflected a higher nutrient cycling capacity compared to mineral fertilized and control plots. Soil bacterial community diversities increased with Cmic and variations in EEAs were strongly correlated with soil DOC availability. Our study has demonstrated that a long-term fertilization strategy can be used to improve soil quality. Clearly, the use of organic fertilizers where available, is a win–win strategy for maintaining soil quality and crop productivity, while ensuring the delivery of soil ecosystem services into the future. ã 2015 Elsevier B.V. All rights reserved. 1. Introduction Chinese agriculture has intensified greatly since the early 1980s on a limited land area with large inputs of mineral fertilizers and other resources to meet the food demand of its increasing population (Guo et al., 2010). Cereal grain yields have increased by 65% between 1980 and 2010 (Zhang et al., 2012), but this success has come at a cost: overuse of mineral fertilizer not only induces low fertilizer use efficiency and the rapid depletion of known * Corresponding author at: Key Laboratory of Plant Nutrition and Fertilizer, Ministry of Agriculture / Institute of Agricultural Resources and Regional Planning, Chinese Academy of Agricultural Sciences, Beijing, 100081, China. Fax: +86 10 82108664. E-mail address: [email protected] (B. Zhao). http://dx.doi.org/10.1016/j.apsoil.2015.07.001 0929-1393/ ã 2015 Elsevier B.V. All rights reserved. Applied Soil Ecology 96 (2015) 75–87 Contents lists available at ScienceDirect Applied Soil Ecology journal homepage: www.elsevier.com/locate/apsoil
Transcript of Soil microbial community structure and function relationships: A heat stress experiment

Applied Soil Ecology 96 (2015) 75–87
Soil microbial community structure and function are significantlyaffected by long-term organic and mineral fertilization regimes in theNorth China Plain
Juan Lia, Julia Mary Cooperb, Zhi'an Lina, Yanting Lia, Xiangdong Yanga,Bingqiang Zhaoa,*aKey Laboratory of Plant Nutrition and Fertilizer, Ministry of Agriculture / Institute of Agricultural Resources and Regional Planning, Chinese Academy ofAgricultural Sciences, Beijing 100081, PR ChinabNafferton Ecological Farming Group, Newcastle University, Nafferton Farm, Stocksfield, Northumberland NE43 7XD, UK
A R T I C L E I N F O
Article history:Received 21 February 2015Received in revised form 2 June 2015Accepted 1 July 2015Available online xxx
Keywords:Long term fertilizationOrganic manureSoil extracellular enzyme activitySoil bacterial communitySoil quality indicators
A B S T R A C T
An improved understanding of the complex interactions and relationships in the soil ecosystem isessential to predict the impact of farming practices on soil quality and its capacity for agriculturalproduction. This study aims to improve our understanding of the impacts of fertilization strategy on keyindicators of soil biological and chemical quality. We studied soils from a winter wheat-summer maizerotational experiment in the North China Plain with six different fertility treatments: no amendments(CK); standard mineral fertilizer treatment (SMF) or standard organic manure treatment (SMA) reflectinglocal farmer practice; mixed treatment with fertilizer and manure at half the rates for the SMF and SMAtreatments (1/2 SMF + 1/2 SMA); double mineral fertilizer treatment (DMF); and double organic manuretreatment (DMA). Soil organic C (SOC), total N (TN), total P (TP), pH, and dissolved organic C (DOC) and N(DON) and microbial biomass C (Cmic) and N (Nmic) were determined using standard methods. Soilbacterial community structure was assessed by denaturing gradient gel electrophoresis (DGGE), andactivities for 10 extracellular enzymes (EEAs) were measured as indicators of soil function. Repeatedapplication of either organic manure or mineral fertilizer increased SOC, TN, TP, DOC, DON, Cmic andNmic, and decreased soil pH. Higher rates of organic manure fertilization significantly affected soilchemical properties compared to the lower rate. Soil bacterial community structure was significantlyaltered by the long-term fertilization regimes and diversity was significantly higher in the double manurerate treatment relative to mineral fertilizer. The higher urease, a-glycosidase, N-acetyl-b-glucosami-nidase, L-leucine aminopeptidase (involved in N cycling), b-glucosidase, b-xylosidase and b-cellobio-sidase (involved in C cycling), and alkaline phosphatase (involved in P cycling) activities for organicmanure fertilized soils reflected a higher nutrient cycling capacity compared to mineral fertilized andcontrol plots. Soil bacterial community diversities increased with Cmic and variations in EEAs werestrongly correlated with soil DOC availability. Our study has demonstrated that a long-term fertilizationstrategy can be used to improve soil quality. Clearly, the use of organic fertilizers where available, is awin–win strategy for maintaining soil quality and crop productivity, while ensuring the delivery of soilecosystem services into the future.
ã 2015 Elsevier B.V. All rights reserved.
Contents lists available at ScienceDirect
Applied Soil Ecology
journal homepage: www.elsevier .com/ locate /apsoi l
* Corresponding author at: Key Laboratory of Plant Nutrition and Fertilizer,Ministry of Agriculture / Institute of Agricultural Resources and Regional Planning,Chinese Academy of Agricultural Sciences, Beijing, 100081, China.Fax: +86 10 82108664.
E-mail address: [email protected] (B. Zhao).
http://dx.doi.org/10.1016/j.apsoil.2015.07.0010929-1393/ã 2015 Elsevier B.V. All rights reserved.
1. Introduction
Chinese agriculture has intensified greatly since the early 1980son a limited land area with large inputs of mineral fertilizers andother resources to meet the food demand of its increasingpopulation (Guo et al., 2010). Cereal grain yields have increasedby 65% between 1980 and 2010 (Zhang et al., 2012), but this successhas come at a cost: overuse of mineral fertilizer not only induceslow fertilizer use efficiency and the rapid depletion of known

76 J. Li et al. / Applied Soil Ecology 96 (2015) 75–87
P-deposits, but also has led to the degradation of the environment,through increased greenhouse gas emissions, nutrient run-off andbiodiversity loss (Zhu and Chen, 2002; Kahrl et al., 2010; Miao et al.,2011; Zhang et al., 2013). All these phenomena have led Chinesepeople to become increasingly concerned about the sustainabilityof current intensive agricultural management practices.
In order to ensure the future sustainability of agriculturalproduction, maintenance of soil quality is essential. Soil quality is“the capacity of a specific kind of soil to function, within natural ormanaged ecosystem boundaries, to sustain plant and animalproductivity, maintain or enhance water and air quality, andsupport human health and habitation” (Doran and Parkin, 1994).Soil quality can be assessed by measuring a range of biological,physical and chemical indicators which are sensitive to changes inmanagement (Fließbach et al., 2007), and linked to key soilfunctions. Measurement of soil quality indicators is therefore auseful approach for assessing and comparing the sustainability ofdifferent crop production strategies.
The replacement of mineral fertilizer with organic manurecould be one way to solve the problems of excessive mineralfertilizer use while at the same time improving soil quality such asphysical and chemical properties, carbon stocks, and biologicalproperties including soil biodiversity (Mäeder et al., 2002;Edmeades, 2003; Hole et al., 2005; Fließbach et al., 2007;Gattinger et al., 2012; Sradnick et al., 2013). Use of organicmanures as fertilizers can also reduce nutrient losses (Zhao et al.,2011) and climate change impacts (Kustermann et al., 2008), andsupport similar or higher crop yields than mineral fertilization incertain contexts (Melero et al., 2006; Lin et al., 2009; Seufert et al.,2012). Mäeder et al. (2002) reported that while yields were 20%lower in organic systems relying on manure fertilization, they havehigher nutrient use efficiencies than conventional systems. Linet al. (2009) in our research group reported similar yields inmanure fertilized treatments compared to mineral fertilizedtreatments after fifteen years in the same experiment.
Soil microbiota regulate biogeochemical nutrient processes andtheir activities play a critical role in terrestrial ecosystems.Meanwhile, microbial properties such as microbial biomass C, Nand P, respiration, and metabolic quotient are often used asindicators of soil quality because of their sensitivity to environ-mental changes, land use (Ndiaye et al., 2000; Acosta-Martinezet al., 2008; Xu et al., 2009) and agricultural management (Li et al.,2008). Numerous reports indicated that fertilization managementsignificantly affected soil microbial properties and communitycomposition (Mäder et al., 2002; Enwall et al., 2005; Gu et al.,2009; Feng et al., 2015). The addition of different amounts ofnitrogen fertilizer induced a shift in microbial communitystructure resulting in an increase in fungal biomarkers but asignificant decrease in the soil microbial community’s growth-response in treatments with higher rates of N addition (e.g.2,000 mg N g�1 soil (Yevdokimov et al., 2008)). Following additionof organic manure, soil microbial communities usually experienceincreases in biomass and activity that release nutrients in plant-available forms promoting plant vegetative growth and contribut-ing to crop productivity (Kallenbach and Grandy, 2011; Jacksonet al., 2012). Feng et al. (2015) revealed that the size of soil bacterialcommunity increased in a long-term organic manure fertilizedalkaline soil in the North China Plain (NCP).
Soil microorganisms depolymerize and mineralize organicmatter by producing extracellular enzymes (Allison et al., 2007).These enzymes have been recognized by soil scientists as one of themore sensitive components of the soil ecosystem and theiractivities (EEA) provide an early indication of the soil's functionalstatus and microbial nutrient demand (Allison et al., 2007;Giacometti et al., 2014). Previous research has shown that soilenzyme activities were influenced by tillage, land use and farming
practices (Li et al., 2008; Fan et al., 2012). For example, soilhydrolytic enzyme activities became higher after regular organicmanure application (García-Ruiz et al., 2008; Li et al., 2008). Fanet al. (2012) demonstrated that long-term mineral fertilizer inputdecreased cellobiohydrolases (CBH) activity, while long-termmanure input increased CBH activity.
This evidence demonstrates that different fertilization strate-gies can alter soil physical, chemical and biological quality, andhave an impact on crop yields. It is also clear that there is a linkbetween soil chemical/biological properties and soil biodiversityand function. Assessing the impact that organic manure andmineral fertilizers and their combined application could have onsoil quality within a Chinese production system will therefore helpto contribute to the development of more sustainable foodproduction systems that meet the demands of a growingpopulation while preventing soil degradation.
The objective of this study is to determine how long-termcontrasting organic and mineral fertilization regimes haveimpacted on indicators of soil quality including: soil chemical/biological properties, bacterial community structure and extracel-lular enzyme activities (EEAs). We hypothesize that long-termorganic manure fertilization will enhance soil chemical andbiological quality indicators, and alter soil microbial communitystructure by increasing diversity. We also hypothesize that thechanges in soil chemical and biological properties will drivechanges in the soil microbial community function (EEAs). Thisresearch should result in an improved understanding of therelationship between soil fertilization strategy and soil qualitywhich will contribute to the development of guidelines for moreeffective use of organic and mineral fertilizers in Chineseagricultural production systems.
2. Material and methods
2.1. Site descriptions and experimental design
This experiment was started in 1986 at Dezhou ExperimentalStation (116�340 E, 36�500 N, altitude: 20 m), Chinese Academy ofAgricultural Sciences (CAAS), which is located in Yucheng City,Shandong Province, in North China Plain (NCP), China. The site’smean annual precipitation and temperature are 569 mm and13.4 �C, respectively. The experimental soil is a Fluvo-aquic typeformed from the sediments of the Yellow River with light loamtexture (clay 21.4%; silt 65.6%; sand 13.0%). Baseline soil chemicalproperties at the start of the experiment in 1985 were 3.93 g totalorganic carbon kg�1, 0.51 g total nitrogen kg�1,7.50 mg Olsen Pkg�1, 1,73.00 mg ammonium acetate-extractable K kg�1, 0.96 gsoluble salt kg�1. The experiment mimicked the standard winterwheat-summer maize double cropping system which is widelyused in the NCP and produces on average a total grain yield (wheatyield plus maize yield) of 15 t ha�1 annum�1 (Lin et al., 2009).Standard commercial tillage and irrigation regimes are used.
2.1. Fertilization treatments
The experiment consists of 24 plots of six fertilization treat-ments with four replicates arranged in a randomized completeblock design. Each plot is 28 m2 (4 m � 7 m), and is separated by a0.8 m concrete slab to prevent the flow of water and fertilizerbetween the plots. The fertilization treatments include a control(CK) with no amendments added, a standard mineral fertilizer(SMF) treatment and a standard organic manure treatment (SMA)that both reflect local farmer practice, a mixed treatment(1/2SMF + 1/2SMA) with fertilizer and manure both applied athalf the rates for the SMF and SMA treatments, a double mineralfertilizer treatment (DMF) and a double organic manure treatment

J. Li et al. / Applied Soil Ecology 96 (2015) 75–87 77
(DMA). The organic manure was cattle manure from the dairyindustry nearby and was composted by regular turning (3–4 times)over a 4 month period before application. All manure applicationsin the experiment are made based on manure total N content.Mineral fertilizers and organic manures were uniformly broadcastonto the soil surface by hand and immediately incorporated intothe plowed soil (0–20 cm depth) by tillage before sowing. Details ofthe rates and timings of manure and fertilizer treatments areincluded in Table 1.
2.2. Soil sample collection and analysis of chemical properties andmicrobial biomass
Soils were sampled on October 7, 2011, prior to the annualapplication of organic manure to minimize effects of recentfertilizer additions on the soil chemical and biological propertiesmeasured. At least five cores (3 cm diameter, 0–20 cm depth) weretaken per plot and mixed to create one combined soil sample andthen separated into 3 parts. The first part was air-dried and sieved(0.25 mm); the second part was sieved (2 mm) and stored at 4 �C,while the third part was sieved (2 mm) and stored at �80 �C. Theair-dried portion was analyzed for organic C by vitriol acid–potassium dichromate oxidation method (SOC), total N by theKjeldahl method (TN) and total P by the HClO4-H2SO4method (TP),and soil pH was measured in a 1:2.5 (soil:water) mixture using thepotentiometric method (Lu, 2000). The second part (fresh soilstored at 4 �C) was used for measuring soil dissolved organiccarbon (DOC) and dissolved organic nitrogen (DON) by shakingwith 0.5 mol/L K2SO4 (1:4 soil:K2SO4 solution) for 30 min and
Table 1Summary of composition, rates and timing of applications for treatments that received orare not shown in this table). Rates represent the total amount applied for the wheat and murea (47% N); fertilizer P is mono-calcium phosphate (17% P2O5); fertilizer K is potaconcentrations 1.00–1.84% N; 0.58–1.67% P2O5; 0.98–1.34% K2O.
Treatment Standard mineralfertilizer (SMF)
Standard organicmanure (SMA)
Organic manure – 375 kg N ha�1 (19861993); 450 kg N ha�
(1994–present)
Timing of manure application – 50% of total beforewheat sowing and
before maize sowin(1986–1996); 100%before wheat sowi(1997–present)
Mineral fertilizer 375 kg N ha�1, 225 kgP2O5ha�1(1986–1993);450 kg N ha�1, (1994–present); 300 kgP2O5ha�1 (1994–1999);150 kg K2O ha�1 (1993–present); 240 kgP2O5ha�1 (2000–present)
–
Timing of mineral fertilizer application Total amount offertilizer N was appliedtwice annually. 50% oftotal N was appliedbefore winter wheatsowing and 50% beforemaize sowing. Winterwheat—40% N, 100%P2O5 and K2O beforesowing ; 60% N GS31;Maize—40% N beforesowing; 60% GS31
–
filtering. DOC and DON were measured in the filtrates bydichromate oxidation and Kjeldahl methods respectively. Micro-bial biomass C (Cmic) and biomass N (Nmic) contents wereassessed on the 4 �C component using the chloroform-fumigation–extraction method (Vance et al., 1987) as we described previously(Li et al., 2008).
2.3. Soil bacterial community structure and diversity analysis
The structure and diversity of bacterial groups of different long-term fertilized soil samples were investigated by a PCR-DGGE(polymerase chain reaction denaturing gradient gel electrophore-sis) approach. The soil samples maintained at �80 �C were used forthe extraction of soil microbial DNA. Soil DNA was extracted usingthe Ultraclean Soil DNA Isolation Kit (Mo Bio Laboratories, SolanaBeach, USA) according to the manufacturer’s instructions. The PCRamplification of the 16S rRNA gene was performed with theuniversal bacterial primers (V3 region of 16SrRNA gene) GC-clamp-338F and 518R (Muyzer et al., 1993) and the extracted DNA astemplate. The reaction mixture (50 ml) containing 25 ml EasyTaqMix (TransGen Biotech, Beijing), 0.2 mM of each primer, 2 ml bovineserum albumin (100 mg/ml), and 10 ng/ml DNA template was runon a Thermo Scientific Arktik Thermal Cycler. The cycling programconsisted of a 95 �C initial denaturating step for 5 min, followed by20 cycles of 94 �C denaturation for 1 min, 65 �C annealing for 30 s(reduced by 0.5 �C each cycle), and 72 �C extension for 3 min,followed by 10 cycles of 94 �C for 1 min, 55 �C annealing for 1 min,72 �C for 3 min; and a final 72 �C extension for 10 min. DGGE wasperformed using about 200 ng PCR product on a 6% (w/v)
ganic manure or mineral fertilizer (control treatment received no amendments andaize crops in a year. All N application rates based on total N contents; fertilizer N isssium sulphate (50% K2O). Organic manure is composted with typical nutrient
Mixed mineral fertilizerand organic manure (1/2SMF + 1/2SMA)
Double standardmineral fertilizer (DMF)
Double standard organicmanure (DMA)
–1
187.5 kg N ha�1 (1986–1993); 225 kg N ha�1
(1994–present)
– 750 kg N ha�1 (1986–1993); 900 kg N ha�1
(1994–present)
50%g
ng
50% of total beforewheat sowing and 50%before maize sowing(1986–1996); 100%before wheat sowing(1997–present)
– 50% of total beforewheat sowing and 50%before maize sowing(1986–1996); 100%before wheat sowing,(1997–present)
187.5 kg N ha�1, 112.5 kgP2O5ha�1 (1986–1993);225 kg N ha�1 (1994–present); 150 kgP2O5ha�1 (1994–1999);75 kg K2O ha�1 (1993–present);120 kg P2O5 ha�1
(2000–present)
750 kg N ha�1, 450 kgP2O5ha�1 (1986–1993);900 kg N ha�1, (1994–present); 600 kgP2O5ha�1 (1994–1999);300 kg K2O ha�1 (1993–present); 480 kgP2O5ha�1 (2000–present)
–
Total amount offertilizer N was appliedtwice annually. 50% oftotal N was appliedbefore winter wheatsowing and 50% beforemaize sowing. Winterwheat—40% N, 100%P2O5 and K2O beforesowing; 60% N GS31;Maize—40% N beforesowing; 60% GS31
Total amount offertilizer N was appliedtwice annually. 50% oftotal N was appliedbefore winter wheatsowing and 50% beforemaize sowing. Winterwheat—40% N, 100%P2O5 and K2O beforesowing; 60% N GS31;Maize—40% N beforesowing; 60% GS31
–

78 J. Li et al. / Applied Soil Ecology 96 (2015) 75–87
acrylamide:bisacrylamide (37.5:1, m:m) gel with 35–60% denatur-ing gradient at 120 V and 60 �C for 8 h using a DCode universalmutation detection system (Bio-Rad Laboratories, Hercules, USA).The gels were stained with 1:10,000 SYBR Green I DNA stainsolution (Sigma, USA) for 30 min and photographed using the GelDocumentation System and analyzed using the software QuantityOne (Bio-Rad Laboratories, Hercules, USA). The gel included12 lanes which allowed 2 replicates of each of the six treatmentsto be run (a total of two gels were used, see the other one in Fig. S1).
2.4. Soil extracellular enzyme activities (EEAs) analysis
Fluorometric substrates linked to 4-methylumbelliferone (MUB)and 7-amino-4-methylcoumarin (AMC) from Sigma (St Louis, MO,USA) were used for assays of seven hydrolytic enzymes (Table 2).Enzyme assays were performed as described by DeForest (2009)with slight modifications of the buffer concentrations due to thealkalinity of the soil in this experiment. Briefly, assays wereconducted by homogenizing each fresh soil sample (equivalentweight to 1.0 g dry mass soil) in 125 ml of 50 mM Tris buffer (pH 8.2)in a 200 ml screw-cap Nalgene bottle, and then stirring the mixturevigorously to maintain a uniform suspension. For the hydrolyticenzymes, the soil sample, Tris buffer, 10 mM references and 200 mMfluorometric substrates were distributed into a black 96-well plate inthe order as described by DeForest (2009). Plates were incubated inthe dark at 25 �C for 4 h until 10 ml 0.5 M NaOH was added to stop thereaction by bringing the pH in thewell to 10. Fluorescence(excitation360 nm; emission 450 nm) was read using a Thermo fluorometer.
The non-fluorometric enzymes (phenol oxidase and peroxi-dase) were measured spectrophotometrically using 3, 4-dihydrox-yphenylalanine (L-DOPA) from Sigma (St Louis, MO, USA) as asubstrate (Table 2). For phenol oxidase and peroxidase, the soilsample, Tris buffer and 25 mM L-DOPA were pipetted into a clear96-well microplate as described by DeForest (2009). Peroxidaseactivity required the addition of 10 ml 0.3% H2O2 solution into everywell. Plates were incubated in the dark at 25 �C for 20 h, and theabsorbance at 450 nm was measured using a Thermo fluorometer.
Urease activity was assayed using urea as the substrate asdescribed by Lu (2000). Briefly, fresh soil equivalent to 5 g oven-drysoil was incubated with 10 ml buffered urea solution (pH 9.0) and0.2 ml toluene in a 50-ml volumetric flask at 37 �C for 2 h, anddiluted with 38 �C water to 50 ml then filtered. The releasedammonium in the soil suspension was determined by theindophenol blue reaction and measured spectrophotometricallyat 578 nm (Crowther and Large, 1956).
Table 2Extracellular enzymes assayed in soils under long-term fertilization regimes, their edihydroxyphenylalanine, 4-MUB = 4-methylumbelliferyl).
Nutrient Cycle Enzyme Abbrev.
N cycle Urease Urease
a-glucosidase AG
L-leucine aminopeptidase LAP
N-acetyl-b-glucosaminidase NAG
C cycle b-1,4-glucosidase BG
b-D-cellobiosidase CBH
b-xylosidase BXYL
P cycle Alkaline Phosphatase AP
phenolic compounds oxidase Phenol oxidase POX
Peroxidase PER
a EC, Enzyme Commission number describing enzymatic function at increasing levhydrolases, 4-lyases, 5-isomerases, and 6-ligases)
2.5. Calculation and Statistical analyses
The degree of similarity in soil bacterial community structure(DGGE profiles) among the fertilizer treatments was determinedby cluster analysis using the Diversity Database Fingerprintingsoftware (Bio-Rad Laboratories) to generate similarity dendro-grams among banding patterns. The Dice similarity index based onUPGAMA (Unweighted Pair Group Method using ArithmeticAverages) clustering method was used. The microbial communityrichness (S, number of bands in each lane of DGGE gels) andmicrobial diversity index (Shannon–Wiener’s diversity index, H’)were calculated as Zak et al. (1994) described.
The hydrolytic enzymes activities and non-fluorometric enzymesactivities were calculated as described by DeForest (2009) andurease activity was calculated as described by Lu (2000). All enzymeactivities were expressed as nmol h�1 g�1 dry soil.
SPSS 11.5 software and MS Excel 2007 were used to carry outdata processing and statistical analysis (ANOVA). The full set oftreatments was used to determine the effect of fertility treatmenton response variables using one-way analysis of variance (ANOVA)with means compared using the least-significant differences (LSD)test. A subset of four treatments (SMF, SMA, DMF, DMA) was usedto test the effects of the interaction between fertilizer input typeand fertilizer input level using two-way ANOVA (p < 0.05).
A constrained ordination technique (CANOCO for Windows4.5 and CANODRAW for Windows) was used to evaluate therelationships between soil enzyme data (the absolute activities ofsoil extracellular enzymes), soil bacterial diversity indices and soilchemical/biological properties using redundancy discriminantanalysis (RDA). Forward selection of soil biological and chemicalproperties (9 variables: SOC, TN, TP, C/N ratio, pH, DOC, DON, Cmic,Nmic) was performed independently for each set of responsevariables (either enzyme activities or diversity indices) to generatea minimum set of significant explanatory variables.
3. Results
3.1. Soil chemical characteristics
Soil organic C and N were affected by fertilization treatments(Table 3). SOC and TN content increased in all treatments incomparison with the original soil after 25 years, and the CKtreatment had 87.3% more C and 149.0% more N than the originalsoil. On average, soils receiving manure had greater amounts ofSOC and TN than mineral fertilized soils, and the SOC contentincreased markedly with increasing rates of manure addition. SoilC/N ratio ranged from 5.82 to 8.84 with significantly higher values
nzyme commission number (ECa) and corresponding substrate (L-DOPA = L-3,4-
Substrate ECa
Urea 3.5.1.54-MUB-a-D-glucoside 3.2.1.20L-Leucine-7-amino-4-methylcoumarin hydrochloride 3.4.11.14-MUB-N-acetyl-b-D-glucosaminide 3.2.1.30
4-MUB-b-D-glucoside 3.2.1.214-MUB-b-D-cellobioside 3.2.1.914-MUB-b-D-xyloside 3.2.1.37
4-MUB-phosphate 3.1.3.1
L-DOPA 1.10.3.2L-DOPA 1.11.1.7
el of detail (the first number distinguishes 1-oxidoreductases, 2-transferases, 3-
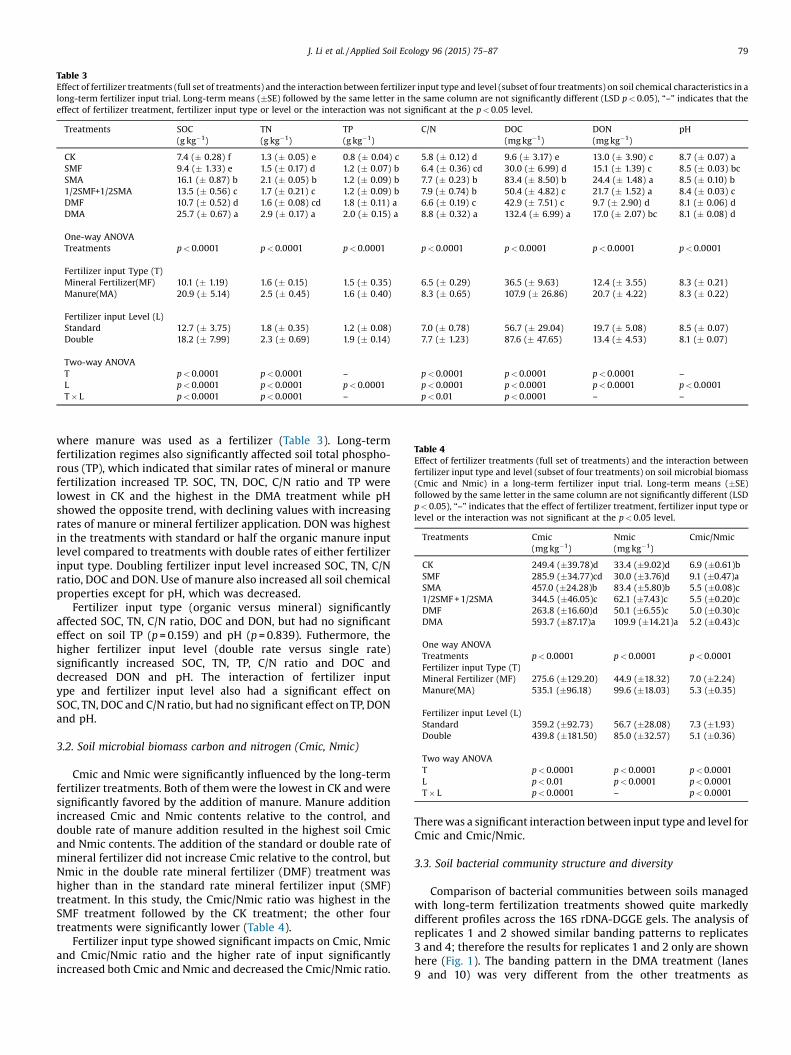
Table 3Effect of fertilizer treatments (full set of treatments) and the interaction between fertilizer input type and level (subset of four treatments) on soil chemical characteristics in along-term fertilizer input trial. Long-term means (�SE) followed by the same letter in the same column are not significantly different (LSD p < 0.05), “–” indicates that theeffect of fertilizer treatment, fertilizer input type or level or the interaction was not significant at the p < 0.05 level.
Treatments SOC(g kg�1)
TN(g kg�1)
TP(g kg�1)
C/N DOC(mg kg�1)
DON(mg kg�1)
pH
CK 7.4 (� 0.28) f 1.3 (� 0.05) e 0.8 (� 0.04) c 5.8 (� 0.12) d 9.6 (� 3.17) e 13.0 (� 3.90) c 8.7 (� 0.07) aSMF 9.4 (� 1.33) e 1.5 (� 0.17) d 1.2 (� 0.07) b 6.4 (� 0.36) cd 30.0 (� 6.99) d 15.1 (� 1.39) c 8.5 (� 0.03) bcSMA 16.1 (� 0.87) b 2.1 (� 0.05) b 1.2 (� 0.09) b 7.7 (� 0.23) b 83.4 (� 8.50) b 24.4 (� 1.48) a 8.5 (� 0.10) b1/2SMF+1/2SMA 13.5 (� 0.56) c 1.7 (� 0.21) c 1.2 (� 0.09) b 7.9 (� 0.74) b 50.4 (� 4.82) c 21.7 (� 1.52) a 8.4 (� 0.03) cDMF 10.7 (� 0.52) d 1.6 (� 0.08) cd 1.8 (� 0.11) a 6.6 (� 0.19) c 42.9 (� 7.51) c 9.7 (� 2.90) d 8.1 (� 0.06) dDMA 25.7 (� 0.67) a 2.9 (� 0.17) a 2.0 (� 0.15) a 8.8 (� 0.32) a 132.4 (� 6.99) a 17.0 (� 2.07) bc 8.1 (� 0.08) d
One-way ANOVATreatments p < 0.0001 p < 0.0001 p < 0.0001 p < 0.0001 p < 0.0001 p < 0.0001 p < 0.0001
Fertilizer input Type (T)Mineral Fertilizer(MF) 10.1 (� 1.19) 1.6 (� 0.15) 1.5 (� 0.35) 6.5 (� 0.29) 36.5 (� 9.63) 12.4 (� 3.55) 8.3 (� 0.21)Manure(MA) 20.9 (� 5.14) 2.5 (� 0.45) 1.6 (� 0.40) 8.3 (� 0.65) 107.9 (� 26.86) 20.7 (� 4.22) 8.3 (� 0.22)
Fertilizer input Level (L)Standard 12.7 (� 3.75) 1.8 (� 0.35) 1.2 (� 0.08) 7.0 (� 0.78) 56.7 (� 29.04) 19.7 (� 5.08) 8.5 (� 0.07)Double 18.2 (� 7.99) 2.3 (� 0.69) 1.9 (� 0.14) 7.7 (� 1.23) 87.6 (� 47.65) 13.4 (� 4.53) 8.1 (� 0.07)
Two-way ANOVAT p < 0.0001 p < 0.0001 – p < 0.0001 p < 0.0001 p < 0.0001 –
L p < 0.0001 p < 0.0001 p < 0.0001 p < 0.0001 p < 0.0001 p < 0.0001 p < 0.0001T � L p < 0.0001 p < 0.0001 – p < 0.01 p < 0.0001 – –
Table 4Effect of fertilizer treatments (full set of treatments) and the interaction betweenfertilizer input type and level (subset of four treatments) on soil microbial biomass(Cmic and Nmic) in a long-term fertilizer input trial. Long-term means (�SE)followed by the same letter in the same column are not significantly different (LSDp < 0.05), “–” indicates that the effect of fertilizer treatment, fertilizer input type orlevel or the interaction was not significant at the p < 0.05 level.
Treatments Cmic(mg kg�1)
Nmic(mg kg�1)
Cmic/Nmic
CK 249.4 (�39.78)d 33.4 (�9.02)d 6.9 (�0.61)bSMF 285.9 (�34.77)cd 30.0 (�3.76)d 9.1 (�0.47)aSMA 457.0 (�24.28)b 83.4 (�5.80)b 5.5 (�0.08)c1/2SMF + 1/2SMA 344.5 (�46.05)c 62.1 (�7.43)c 5.5 (�0.20)cDMF 263.8 (�16.60)d 50.1 (�6.55)c 5.0 (�0.30)cDMA 593.7 (�87.17)a 109.9 (�14.21)a 5.2 (�0.43)c
One way ANOVATreatments p < 0.0001 p < 0.0001 p < 0.0001Fertilizer input Type (T)Mineral Fertilizer (MF) 275.6 (�129.20) 44.9 (�18.32) 7.0 (�2.24)Manure(MA) 535.1 (�96.18) 99.6 (�18.03) 5.3 (�0.35)
Fertilizer input Level (L)Standard 359.2 (�92.73) 56.7 (�28.08) 7.3 (�1.93)Double 439.8 (�181.50) 85.0 (�32.57) 5.1 (�0.36)
Two way ANOVAT p < 0.0001 p < 0.0001 p < 0.0001L p < 0.01 p < 0.0001 p < 0.0001T � L p < 0.0001 – p < 0.0001
J. Li et al. / Applied Soil Ecology 96 (2015) 75–87 79
where manure was used as a fertilizer (Table 3). Long-termfertilization regimes also significantly affected soil total phospho-rous (TP), which indicated that similar rates of mineral or manurefertilization increased TP. SOC, TN, DOC, C/N ratio and TP werelowest in CK and the highest in the DMA treatment while pHshowed the opposite trend, with declining values with increasingrates of manure or mineral fertilizer application. DON was highestin the treatments with standard or half the organic manure inputlevel compared to treatments with double rates of either fertilizerinput type. Doubling fertilizer input level increased SOC, TN, C/Nratio, DOC and DON. Use of manure also increased all soil chemicalproperties except for pH, which was decreased.
Fertilizer input type (organic versus mineral) significantlyaffected SOC, TN, C/N ratio, DOC and DON, but had no significanteffect on soil TP (p = 0.159) and pH (p = 0.839). Futhermore, thehigher fertilizer input level (double rate versus single rate)significantly increased SOC, TN, TP, C/N ratio and DOC anddecreased DON and pH. The interaction of fertilizer inputype and fertilizer input level also had a significant effect onSOC, TN, DOC and C/N ratio, but had no significant effect on TP, DONand pH.
3.2. Soil microbial biomass carbon and nitrogen (Cmic, Nmic)
Cmic and Nmic were significantly influenced by the long-termfertilizer treatments. Both of them were the lowest in CK and weresignificantly favored by the addition of manure. Manure additionincreased Cmic and Nmic contents relative to the control, anddouble rate of manure addition resulted in the highest soil Cmicand Nmic contents. The addition of the standard or double rate ofmineral fertilizer did not increase Cmic relative to the control, butNmic in the double rate mineral fertilizer (DMF) treatment washigher than in the standard rate mineral fertilizer input (SMF)treatment. In this study, the Cmic/Nmic ratio was highest in theSMF treatment followed by the CK treatment; the other fourtreatments were significantly lower (Table 4).
Fertilizer input type showed significant impacts on Cmic, Nmicand Cmic/Nmic ratio and the higher rate of input significantlyincreased both Cmic and Nmic and decreased the Cmic/Nmic ratio.
There was a significant interaction between input type and level forCmic and Cmic/Nmic.
3.3. Soil bacterial community structure and diversity
Comparison of bacterial communities between soils managedwith long-term fertilization treatments showed quite markedlydifferent profiles across the 16S rDNA-DGGE gels. The analysis ofreplicates 1 and 2 showed similar banding patterns to replicates3 and 4; therefore the results for replicates 1 and 2 only are shownhere (Fig. 1). The banding pattern in the DMA treatment (lanes9 and 10) was very different from the other treatments as

80 J. Li et al. / Applied Soil Ecology 96 (2015) 75–87
illustrated by the band labeled A on the figure. This band waspresent in all treatments except for DMA, indicating a cleardifference in community structure for this treatment. This wasreflected in the results of the cluster analysis. This analysis showedeach pair of replicates grouped together for each fertilizertreatment and CK (Fig. 2), indicating a good replication of bacterialcommunity structure within the same treatment.
Patterns of the treatments receiving the same type of fertilizers(only manure or only mineral fertilizer) clustered together.Treatments which had received a mixture of manure and mineralfertilizer (1/2SMF + 1/2SMA) were separated from the treatmentsreceiving only manure (SMA and DMA). Control treatments werean even more separated from the 1/2SMF + 1/2SMA treatments andthe treatments receiving only manure (SMA and DMA) (Fig. 2).Finally, patterns of the treatments receiving only mineral fertilizers(SMF and DMF) were separated from the other treatments.
For soil bacteria community richness (S) and diversity (H’) theDMA treatment had the highest levels with lowest values for thecontrol (Table 5). Fertilizer input type significantly affected S and H’but fertilizer input level had no impact on them, although therewas a significant interaction between type and level.
Fig. 1. DGGE profile of amplified 16S rRNA genes V3 fragments from soils underdifferent long term fertilizer treatments. (block 1 and 2). CK: control with noamendments added; SMF: standard mineral fertilizer treatment reflect local farmerpractice; SMA: standard organic manure treatment reflect local farmer practice; 1/2 SMF + 1/2 SMA: mixed treatment with fertilizer and manure both applied at halfthe rates for the SMF and SMA treatments; DMF: double mineral fertilizertreatment; DMA: double organic manure treatment. Subscripts refer to blocknumber.
3.4. Patterns of soil microbial community structure
RDA was carried out using soil chemical and biologicalproperties (TN, SOC, TP, C/N ratio, pH, DOC, DON, Cmic and Nmic)as explanatory variables and the relative abundance of 2 indicators(S and H’) of soil bacterial community structure as responsevariables (Fig. 3a). The explanatory variables accounted for 67.1% ofthe total variation (adjusted explained variation is 46.0%). Axis1 and axis 2 accounted for 66.6% and 0.6% of the variationrespectively. The most significant factor selected by forwardselection was Cmic (p < 0.01) and the variance of soil bacterialcommunity structure data explained by Cmic was 52.8%. Increasesin S and H’ along axis 1 strongly correlated with Cmic (Fig. 3a).Variations along axis 1 were also driven by DOC, TN, SOC, Nmic,and C/N ratio, while TP was positively associated with both axis1 and axis 2. Soil pH had a negative effect on axis 1 and axis 2 andDON was positively associated with axis 1 while negativelyaffecting axis 2. The circles on Fig. 3b indicate the separationbetween treatments which applied with manure (DMA treatmentand SMA treatment) and the other treatments along axis1 indicating different soil bacterial community structures whenorganic manure was applied for a prolonged period. CK treatmentwith no fertilizers input showed the lowest values along axis 1 andhad the highest correlation with pH. Within the same standardfertilizer input level, the SMA treatment had the highest valuesalong axis 1 and did not overlap with 1/2SMF + 1/2SMA and SMF.
3.5. Soil extracellular enzyme activities (EEAs)
To clarify the patterns of extracellular enzyme activities underthe different treatments we expressed each enzyme as apercentage of the control (Ai et al., 2012). Extracellular enzymeactivities (EEAs) were significantly affected by long-term fertilizertreatments (p < 0.05). Fertilizer input type significantly affectedeight EEAs with the exception of POX and PER. All ten EEAs weresignificantly affected by fertilizer input level.
Urease activity, which is involved in N acquisition, was highestin the treatments which received only manure (Fig. 4a). At thesame standard N input level, the greatest urease activity was foundin the SMA treatment followed by the 1/2SMF + 1/2SMA treatment,and the lowest urease activity was measured in the SMF treatment.Compared to CK, long-term fertilized treatments had higher AGactivity except for DMF treatment in which AG activity was lowerthan the control. AG activity was significantly higher in treatmentswhich received manure compared to the other treatments.Similarly, treatments with manure addition had a positive effecton activities for the carbohydrate-degrading enzymes BG, BXYLand CBH. Compared to the control, the chitin degradation enzyme(NAG) activity declined and the polypeptide degradation enzymeLAP increased in the SMF treatment. In both cases application ofdouble the manure rate resulted in the highest increase in EEAsrelative to the control. Alkaline phosphatase (AP) activity showed asimilar pattern of responses to the urease activity. POX and PERdecreased with increasing N input level, showing an opposite trendcompared with the other enzymes.
3.6. Patterns of soil enzyme activities
RDA was performed using soil chemical and biologicalproperties (TN, SOC, TP, C/N ratio, pH, DOC, DON, Cmic and Nmic)as factors and all ten enzyme activities as response variables(Fig. 5a). Results showed that the explanatory variables accountedfor 75% of the total variation (adjusted explained variation is58.9%). Based on permutation tests, DOC was the most significantvariable selected by forward selection explaining 53.4% of thevariance of enzyme data (p < 0.01, padj< 0.05). The first two RDA

Fig. 2. DGGE Cluster analysis (UPGMA) of 16SrDNA profiles of bacterial communities in soils under different long-term fertilizer treatments. (Block 1 and 2). CK: control withno amendments added; SMF: standard mineral fertilizer treatment reflect local farmer practice; SMA: standard organic manure treatment reflect local farmer practice; 1/2 SMF + 1/2 SMA: mixed treatment with fertilizer and manure both applied at half the rates for the SMF and SMA treatments; DMF: double mineral fertilizer treatment; DMA:double organic manure treatment. Subscripts refer to block number.
Table 5Effect of fertilizer treatments (full set of treatments) and the interaction between fertilizer input type and level (subset of fourtreatments) on diversity properties of soil bacteria using DGGE bands pattern data associated with different long-term fertilizertreatments. Long-term means (�SE) followed by the same letter in the same column are not significantly different (LSD p < 0.05), “–”
indicates that the effect of fertilizer treatment, fertilizer input type or level or the interaction was not significant at the p < 0.05 level.
Fertilizer Treatments Richness (S detected band) Shannon Weiner index (H’)
CK 38 (�3)c 3.60 (�0.09)cSMF 42 (�1)b 3.70 (�0.03)abSMA 43 (�2)ab 3.73 (�0.03)ab1/2SMF + 1/2SMA 41 (�2)b 3.68 (�0.03)bDMF 41 (�1)b 3.66 (�0.00)bDMA 45 (�1)a 3.76 (�0.03)a
One-way ANOVATreatments p < 0.001 p < 0.01
Fertilizer input Type (T)Mineral Fertilizer (MF) 41 (�1) 3.68 (�0.03)Manure(MA) 44 (�2) 3.74 (�0.03)
Fertilizer input Level (L)Standard 42 (�1) 3.71 (�0.03)Double 43 (�2) 3.71 (�0.06)
Two way ANOVAT p < 0.0001 p � 0.001L – –
T � L p < 0.05 p < 0.05
J. Li et al. / Applied Soil Ecology 96 (2015) 75–87 81
axes accounted for 55.34% and 8.08% of the total variation. The soilproperties TP, TN, SOC, DOC, C/N ratio, Cmic and Nmic werepositively correlated with the first axis (1), while pH loadednegatively along axis 1 and axis 2.
Axis 1 largely differentiated activities of C-cycling enzymesCBH, BG and BXYL, N-cycling enzymes AG, Urease, NAG and LAP,and the P-cycling enzyme AP. Axis 2 primarily explained variabilityin POX and PER which oxidize phenolic compounds, and werelargely driven by pH.
The clear separation of fertility treatments based on theirextracellular enzyme activities is shown in Fig. 5b with the CK,SMA, and DMA treatments differentiated from the other treat-ments. Soil properties driving these differences in enzyme activitywere primarily soil organic carbon and nitrogen which positively
affected axis 1, and pH which negatively affected axis 1. Within thesame standard fertilizer input level, the SMA treatment showedhigher values along axis 1 compared to 1/2SMF + 1/2SMA and SMF.The SMF treatment had slightly lower values than the 1/2SMF + 1/2SMA treatment along axis 1 due to its lower soil C and N contentsand higher pH.
4. Discussion
This research approach provides insight into how soil chemicaland biological properties, total bacterial community structure andsoil extracellular enzyme activities respond to the contrasting 25-year organic and mineral fertilization regimes. Long-term fertili-zation significantly impacted soil nutrient availability, and in turn
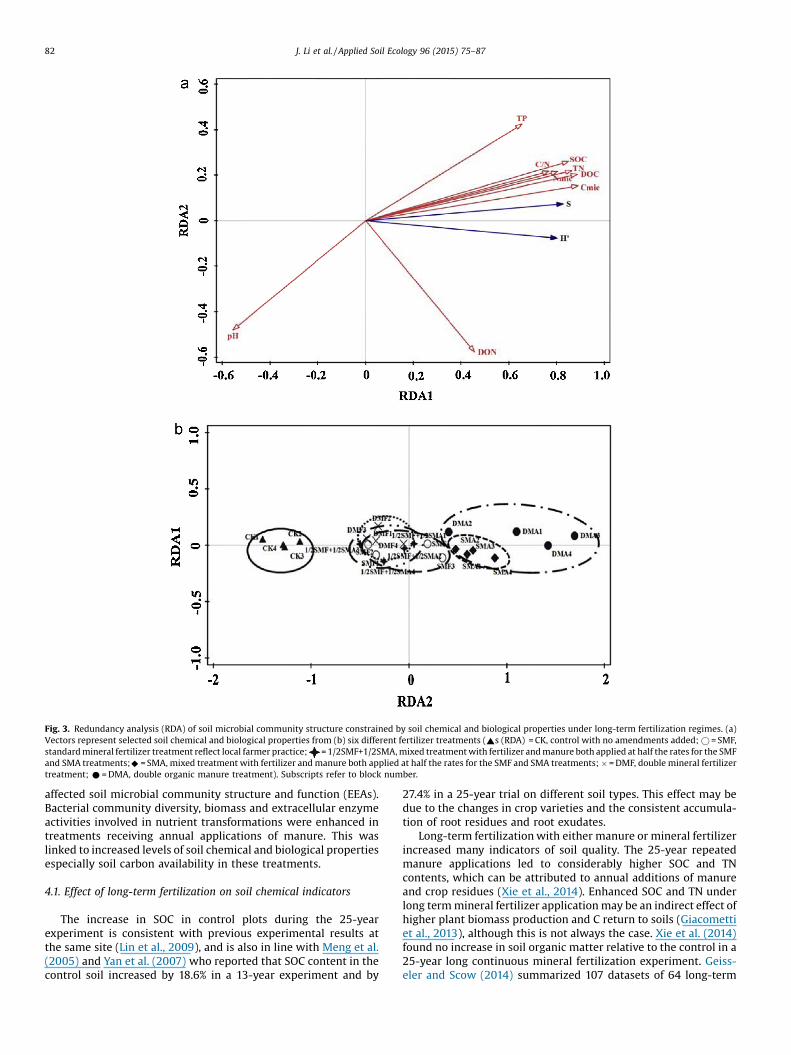
Fig. 3. Redundancy analysis (RDA) of soil microbial community structure constrained by soil chemical and biological properties under long-term fertilization regimes. (a)Vectors represent selected soil chemical and biological properties from (b) six different fertilizer treatments (~s (RDA) = CK, control with no amendments added; � = SMF,standard mineral fertilizer treatment reflect local farmer practice; = 1/2SMF+1/2SMA, mixed treatment with fertilizer and manure both applied at half the rates for the SMFand SMA treatments;^ = SMA, mixed treatment with fertilizer and manure both applied at half the rates for the SMF and SMA treatments; � = DMF, double mineral fertilizertreatment; * = DMA, double organic manure treatment). Subscripts refer to block number.
82 J. Li et al. / Applied Soil Ecology 96 (2015) 75–87
affected soil microbial community structure and function (EEAs).Bacterial community diversity, biomass and extracellular enzymeactivities involved in nutrient transformations were enhanced intreatments receiving annual applications of manure. This waslinked to increased levels of soil chemical and biological propertiesespecially soil carbon availability in these treatments.
4.1. Effect of long-term fertilization on soil chemical indicators
The increase in SOC in control plots during the 25-yearexperiment is consistent with previous experimental results atthe same site (Lin et al., 2009), and is also in line with Meng et al.(2005) and Yan et al. (2007) who reported that SOC content in thecontrol soil increased by 18.6% in a 13-year experiment and by
27.4% in a 25-year trial on different soil types. This effect may bedue to the changes in crop varieties and the consistent accumula-tion of root residues and root exudates.
Long-term fertilization with either manure or mineral fertilizerincreased many indicators of soil quality. The 25-year repeatedmanure applications led to considerably higher SOC and TNcontents, which can be attributed to annual additions of manureand crop residues (Xie et al., 2014). Enhanced SOC and TN underlong term mineral fertilizer application may be an indirect effect ofhigher plant biomass production and C return to soils (Giacomettiet al., 2013), although this is not always the case. Xie et al. (2014)found no increase in soil organic matter relative to the control in a25-year long continuous mineral fertilization experiment. Geiss-eler and Scow (2014) summarized 107 datasets of 64 long-term

.b.a
c. d.
.f.e
d
ab
c
b
a
0
50
100
150
200
250
300
350
SMF
SMA
1/2SMF+1/2SM
A
DMF
DMA
Urease
Perc
ent
diff
eren
ce (%
) fro
m C
K
c
b
c
bc
a
0
50
100
150
200
250
SMF
SMA
1/2SMF+1/2SMA
DMF
DMA
Alkaline Phosphatase (AP)
Perc
ent
diff
eren
ce (%
) fro
m C
K
bc
a
ab
c
a
-60
-40
-20
0
20
40
60
80
100
SMF
SMA
1/2SMF+1/2SM
A
DMF
DMA
α-Glucosidase(AG)
Perc
ent
diffe
renc
e (%
) fro
m C
K
c
b
c
b
a
0
50
100
150
200
250
SMF
SMA
1/2SMF+1/2SM
A
DMF
DMA
β-Xylosidase (BXYL)
Perc
ent
diff
eren
ce (%
) fro
m C
K
c
b
cc
a
0
50
100
150
200
250
300
350
SMF
SMA
1/2SMF+1/2SM
A
DMF
DMA
β-Cellobiosidase (CBH)
Perc
ent
diff
eren
ce (%
) fro
m C
K
c
b
bc
bc
a
0
50
100
150
200
250
SMF
SMA
1/2SMF+1/2SM
A
DMF
DMA
β-Glucosidase (BG)
Perc
ent
diff
eren
ce (%
) fro
m C
K
Fig. 4. Percent difference of soil Extracellular Enzyme Activities (EEAs) in the fertilizer treatments compared to the corresponding control treatments (CK) calculated as[(fertilizer treatment�CK)/CK � 100]. CK: control with no amendments added; SMF: standard mineral fertilizer treatment reflect local farmer practice; SMA: standard organicmanure treatment reflect local farmer practice; 1/2 SMF + 1/2 SMA: mixed treatment with fertilizer and manure both applied at half the rates for the SMF and SMA treatments;DMF: double mineral fertilizer treatment; DMA: double organic manure treatment. Means labeled with the same letter are not significantly different at p < 0.05 (LSD test onactual values of the EEAs).
J. Li et al. / Applied Soil Ecology 96 (2015) 75–87 83
experiments and found that the mineral fertilization increased SOCcontent relative to the control by a mean of 12.8%, which agreeswell with our results.
The long-term application of fertilizers can decrease pH inalkaline soils, as found in another 15-year trial in the NCP where pHdeclined after repeated fertilizer application, especially when

.h.g
.j.iFertil izer Trea tments
d
b
cc
a
-20
0
20
40
60
80
100
120
140
160
180
SMF
SMA
1/2SMF+1/2SM
A
DMF
DMA
N-acetyl-β-Glucosaminidase (NAG)
Perc
ent
diff
eren
ce (%
) fro
m C
K
d
c c
b
a
-20
-15
-10
-5
0
5
10
15
20
25
30
SMF
SMA
1/2SMF+1/2SMA
DMF
DMA
L-leucine aminopeptidase (LAP)
Perc
ent
diff
eren
ce (%
) fro
m C
Kb
a
d
c
d-20
-15
-10
-5
0
5
10
15
20
SMF
SMA
1/2SMF+1/2SM
A
DMF
DMA
Peroxidase (PER)
Perc
ent
diff
eren
ce (%
) fro
m C
K b
a
b
c
d-35
-30
-25
-20
-15
-10
-5
0
5
SMF
SMA
1/2SMF+1/2
SMA
DMF
DMA
Phenol oxidase (POX)
Perc
ent
diff
eren
ce (%
) fro
m C
K
Fig. 4. (Continued)
84 J. Li et al. / Applied Soil Ecology 96 (2015) 75–87
manure was used (Liang et al., 2012). This was also observed in ourtrial where pH declined with increasing application rate, regard-less of the input type. The mechanism for this pH decline may notbe the same for each input type. Although hydrolysis of urea resultsin a slight increase in pH, this is followed by a decrease asammonium is converted to nitrate through nitrification processes,which could result in a pH decline in our experiment. Manureapplications can increase soil pH due to the liming effect of addedcarbonates and organic matter (Cooper and Warman, 1997);however, in this trial the long-term additions of manure may haveresulted in an accumulation of organic acids resulting in a declinein pH (Liang et al., 2012).
In this study, standard rate fertilizer input treatments hadsimilar effects on TP, which is consistent with the previous report(Lin et al., 2009). Fertilizer input level was the predominant factorinfluencing TP (Table 2). Fertilizer input type and level also had animpact on soil DOC and DON contents, which are key indicators ofagricultural soil quality (Haynes, 2000). At very high rates oforganic manure (DMA) DOC was highest, but DON was reducedrelative to the SMA treatment. This suggests that the high levels ofDOC were stimulating microbial activity leading to immobilizationof N from the DON pool. This is supported by the evidence from the
EEAs which were significantly higher for enzymes involved in Ccycling (e.g. BG, CBH and BXYL).
4.2. Effect of long-term fertilization on soil microbial biomass
The higher soil microbial biomass in the manure treatmentsmay have been due to the addition of microbial populations in themanure and the additional C, which activates the soil indigenousmicrobiota. Enhancement of soil microbial biomass after organicmanure addition has been reported during long-term experimentswith the additional carbon sources which are benefit for thegrowth of soil microorganisms and the increases of soil nutrientscontents (Li et al., 2008).
Several studies have shown that mineral fertilizer applicationcan increase the crop residues returned back to the soil andstimulate soil microbial growth and activity (Giacometti et al.,2013). This has been demonstrated even when the abovegroundcrop was removed at harvest, possibly because of not onlyincreased root biomass but also the stimulation of bulk orrhizospheric microbial activity (Gu et al., 2009). In our experiment,this effect was not observed, and even the double mineral fertilizerrate input treatment had a Cmic similar with that of the control.This is in line with previous studies which found lower Cmic at
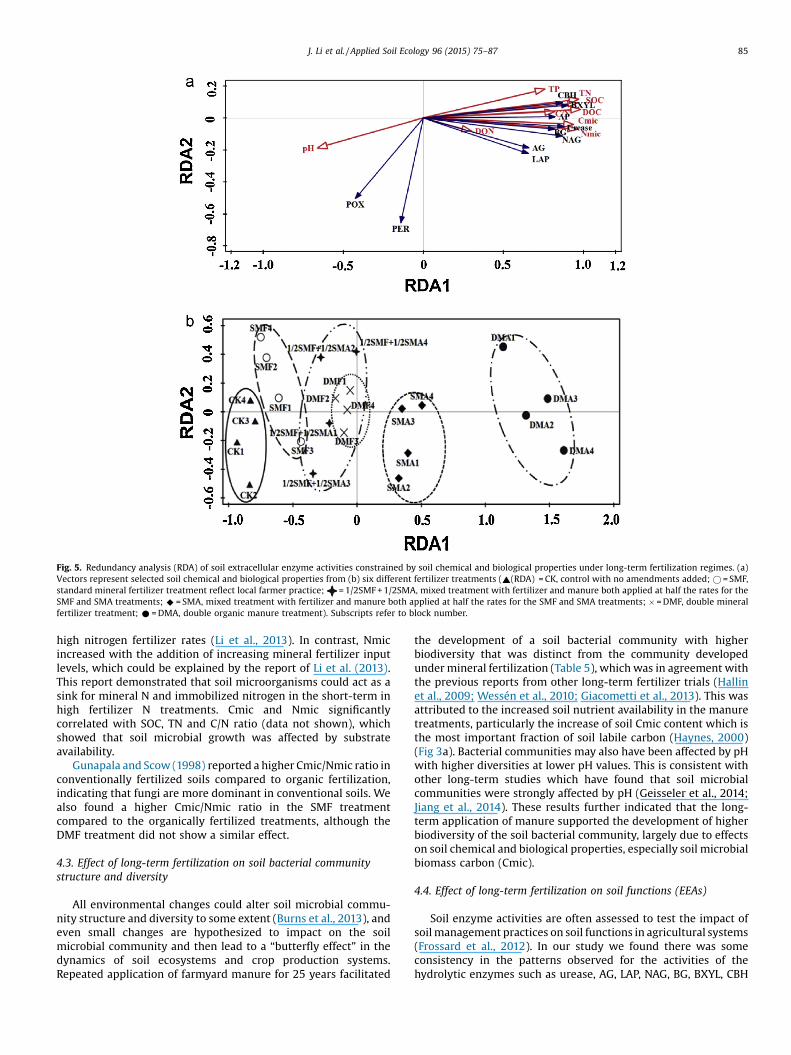
Fig. 5. Redundancy analysis (RDA) of soil extracellular enzyme activities constrained by soil chemical and biological properties under long-term fertilization regimes. (a)Vectors represent selected soil chemical and biological properties from (b) six different fertilizer treatments (~(RDA) = CK, control with no amendments added; � = SMF,standard mineral fertilizer treatment reflect local farmer practice; = 1/2SMF + 1/2SMA, mixed treatment with fertilizer and manure both applied at half the rates for theSMF and SMA treatments; ^ = SMA, mixed treatment with fertilizer and manure both applied at half the rates for the SMF and SMA treatments; � = DMF, double mineralfertilizer treatment; * = DMA, double organic manure treatment). Subscripts refer to block number.
J. Li et al. / Applied Soil Ecology 96 (2015) 75–87 85
high nitrogen fertilizer rates (Li et al., 2013). In contrast, Nmicincreased with the addition of increasing mineral fertilizer inputlevels, which could be explained by the report of Li et al. (2013).This report demonstrated that soil microorganisms could act as asink for mineral N and immobilized nitrogen in the short-term inhigh fertilizer N treatments. Cmic and Nmic significantlycorrelated with SOC, TN and C/N ratio (data not shown), whichshowed that soil microbial growth was affected by substrateavailability.
Gunapala and Scow (1998) reported a higher Cmic/Nmic ratio inconventionally fertilized soils compared to organic fertilization,indicating that fungi are more dominant in conventional soils. Wealso found a higher Cmic/Nmic ratio in the SMF treatmentcompared to the organically fertilized treatments, although theDMF treatment did not show a similar effect.
4.3. Effect of long-term fertilization on soil bacterial communitystructure and diversity
All environmental changes could alter soil microbial commu-nity structure and diversity to some extent (Burns et al., 2013), andeven small changes are hypothesized to impact on the soilmicrobial community and then lead to a “butterfly effect” in thedynamics of soil ecosystems and crop production systems.Repeated application of farmyard manure for 25 years facilitated
the development of a soil bacterial community with higherbiodiversity that was distinct from the community developedunder mineral fertilization (Table 5), which was in agreement withthe previous reports from other long-term fertilizer trials (Hallinet al., 2009; Wessén et al., 2010; Giacometti et al., 2013). This wasattributed to the increased soil nutrient availability in the manuretreatments, particularly the increase of soil Cmic content which isthe most important fraction of soil labile carbon (Haynes, 2000)(Fig 3a). Bacterial communities may also have been affected by pHwith higher diversities at lower pH values. This is consistent withother long-term studies which have found that soil microbialcommunities were strongly affected by pH (Geisseler et al., 2014;Jiang et al., 2014). These results further indicated that the long-term application of manure supported the development of higherbiodiversity of the soil bacterial community, largely due to effectson soil chemical and biological properties, especially soil microbialbiomass carbon (Cmic).
4.4. Effect of long-term fertilization on soil functions (EEAs)
Soil enzyme activities are often assessed to test the impact ofsoil management practices on soil functions in agricultural systems(Frossard et al., 2012). In our study we found there was someconsistency in the patterns observed for the activities of thehydrolytic enzymes such as urease, AG, LAP, NAG, BG, BXYL, CBH

86 J. Li et al. / Applied Soil Ecology 96 (2015) 75–87
and AP in the organically fertilized and the combined organic andmineral fertilized soils. Soil enzyme activities could be explainedby soil chemical and biological properties such as nutrientavailability and microbial activity, which were strongly influencedby long-term fertilization regime. Hojjati and Nourbakhsh (2007)reported a positive correlation between the activity of hydrolyticenzymes and total organic carbon content, DOC and TN whichincreased significantly under organic fertilization. The addition oforganic fertilizers activates microorganisms to produce hydrolasesinvolved in the nutrient cycles, which respond to the increasedsupply of easily degradable organic matter (Hojjati and Nour-bakhsh, 2007). In addition, long-term mineral fertilizer applicationpromoted the production of hydrolytic enzyme activities related toC, N and P cycles, which is consistent with several previous studies(Melero et al., 2006; Li et al., 2008). The potential activity of theP-cycling enzyme AP (alkaline phosphatase) was previouslydemonstrated to have a strong positive relationship with soilmicrobial biomass carbon (Bowles et al., 2014) and an inverserelationship with high P availability (Olander and Vitousek, 2000;Allison et al., 2007). Our analysis indicated that 8 soil chemical andbiological quality indicators (TN, SOC, TP, C/N ratio, pH, DOC, Cmicand Nmic) (data not shown) and soil bacterial community diversityindicators S (p < 0.01) and H’ (p < 0.01) had strongly positiverelationships with AP activity. Oxidases such as phenol oxidase(POX) and peroxidase (PER) which mainly catalyze the process oflignin degradation are generally produced by fungi (Burns et al.,2013), and their activities were significantly different from those ofall other enzymes in this experiment. Sinsabaugh (2010) reportedthat POX and PER activities increased with soil pH, and candecrease with N amendment. We also found significant reductionsin POX activity with increasing rates of either organic or mineralfertilizers, as well as increasing PER and POX activities withincreasing pH (Fig. 4, Fig. 5).
Our results have demonstrated that key soil functions relatingto nutrient cycling are enhanced by long-term manure addition.These changes in soil function may indicate the evolution of a soilmicrobial community after long-term manure application that isbetter adapted to the metabolism of organic fertilizers. Thisadapted community could contribute to more efficient cycling ofnutrients from organic sources in the long-term.
5. Conclusions
In a Fluvo-aquic soil of the North China Plain, long-term organicmanure application enhanced various chemical indicators of soilquality (SOC, TN, TP, DOC, DON content) as well as microbiologicalindicators (Cmic and Nmic content). The bacterial structures anddiversities were significantly affected by long-term fertilizationstrategy, and were constrained by alterations of soil resourceavailability (SOC). Soil function related to nutrient cycling was alsoenhanced in the soil with a history of manure amendment.
These results indicate that use of organic fertilizers can result inthe development of a soil adapted to metabolism of a variety oforganic substrates. This has relevance for future crop productionwhere it is expected that more organic waste materials will berecycled to the land. An additional benefit of cropping systems thatrely on organic fertilizers for nutrients is the enhancement of soilecosystem service provision. Soil organic C is expected to increaseunder systems reliant on organic fertilizers, with the concomitantecosystem services of greenhouse gas (GHG) mitigation (due to Csequestration) and water regulation (drought and flood mitigation).Enhanced SOC will also lead to improvements in soil structure withreduced risk of off-site damage due to soil erosion and sedimenta-tion. In addition, soils with high SOC levels and microbial activitieshave been shown to have a higher degree of resilience to both
environmental (floods, drought) and manmade (intensification ofagricultural production) stresses (Kumar et al., 2014).
Our study has demonstrated that a long-term fertilizationstrategy can be used to improve soil quality in a variety of ways.Clearly, the use of organic fertilizers where available, is a win–winstrategy for maintaining soil quality and crop productivity, whileensuring the delivery of soil ecosystem services into the future.
Acknowledgements
The authors gratefully acknowledge funding from NationalNatural Science Foundation of China (NSFC, Grant No. 31301843),National Nonprofit Institute Research Grant of CAAS (Grant No.IARRP-202-5) and the European Community financial participa-tion under the Seventh Framework Programme for Research,Technological Development and Demonstration Activities, for theIntegrated Project NUE-CROPS FP7-CP-IP 222645. The viewsexpressed in this publication are the sole responsibility of theauthor(s) and do not necessarily reflect the views of the EuropeanCommission. Neither the European Commission nor any personacting on behalf of the Commission is responsible for the use whichmight be made of the information contained herein.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.apsoil.2015.07.001.
References
Acosta-Martinez, V., Acosta-Mercado, D., Sotomayor-Ramırez, D., Cruz-Rodrıguez,L., 2008. Microbial communities and enzymatic activities under differentmanagement in semiarid soils. Appl. Soil Ecol. 38 (3), 249–260.
Ai, C., Liang, G., Sun, J., Wang, X., Zhou, W., 2012. Responses of extracellular enzymeactivities and microbial community in both the rhizosphere and bulk soil tolong-term fertilization practices in a fluvo-aquic soil. Geoderma 173, 330–338.
Allison, S.D., Gartner, T., Holland, K., Weintraub, M., Sinsabaugh, R.L., 2007. Soilenzymes: linking proteomics and ecological process. Man. Environ. Microbiol.704–711.
Bowles, T.M., Acosta-Martínez, V., Calderón, F., Jackson, L.E., 2014. Soil enzymeactivities, microbial communities, and carbon and nitrogen availability inorganic agroecosystems across an intensively-managed agricultural landscape.Soil Biol. Biochem. 68, 252–262.
Burns, R.G., DeForest, J.L., Marxsen, J., Sinsabaugh, R.L., Stromberger, M.E.,Wallenstein, M.D., Weintraub, M.N., Zoppini, A., 2013. Soil enzymes in achanging environment: Current knowledge and future directions. Soil Biol.Biochem. 58, 216–234.
Cooper, J.M., Warman, P.R., 1997. Effects of three fertility amendments on soildehydrogenase activity, organic C and pH. Can. J. Soil Sci. 77 (2), 281–283.
Crowther, A.B., Large, R.S., 1956. Improved conditions for the sodium phenoxide-sodium hypochlorite method for the determination of ammonia. Analyst 81,64–65.
DeForest, J., 2009. The influence of time, storage temperature, and substrate age onpotential soil enzyme activity in acidic forest soils using MUB-linked substratesand L-DOPA. Soil Biol. Biochem. 41, 1180–1186.
Doran, J.W., Parkin, T.B., 1994. Defining and assessing soil quality. In: Doran, J.W.,Coleman, D.C., Bezdicek, D.F., Stewart, B.A. (Eds.), Defining Soil Quality for aSustainable Environment. Soil Science Society Americca Journal, Madison, pp.3–21.
Edmeades, D.C., 2003. The long-term effects of manures and fertilizers on soilproductivity and quality: a review. Nutr. Cycling Agroecosyst. 66 (2), 165–180.
Enwall, K., Philippot, L., Hallin, S., 2005. Activity and composition of the denitrifyingbacterial community respond differently to long-term fertilization. Appl.Environ. Microbiol. 71, 8335–8343.
Fan, F., Li, Z., Wakelin, S.A., Yu, W., Liang, Y., 2012. Mineral fertilizer alters cellulolyticcommunity structure and suppresses soil cellobiohydrolase activity in a long-term fertilization experiment. Soil Biol. Biochem. 55, 70–77.
Feng, Y., Chen, R., Hu, J., Zhao, F., Wang, J., Chu, H., Zhang, J., Dolfing, J., Lin, X., 2015.Bacillus asahii comes to the fore in organic manure fertilized alkaline soils. SoilBiol. Biochem. 81, 186–194.
Fließbach, A., Oberholzer, H.R., Gunst, L., Mäder, P., 2007. Soil organic matter andbiological soil quality indicators after 21 years of organic and conventionalfarming. Agric. Ecosyst. Environ. 118, 273–284.

J. Li et al. / Applied Soil Ecology 96 (2015) 75–87 87
Frossard, A., Gerull, L., Mutz, M., Gessner, M.O., 2012. Disconnect of microbialstructure and function: enzyme activities and bacterial communities in nascentstream corridors. ISME J. 6 (3), 680–691.
García-Ruiz, R., Ochoa, V., Hinojosa, M.B., Carreira, J.A., 2008. Suitability of enzymeactivities for the monitoring of soil quality improvement in organic agriculturalsystems. Soil Biol. Biochem. 40 (9), 2137–2145.
Gattinger, A., Muller, A., Haeni, M., Skinner, C., Fliessbach, A., Buchmannb, N., Mäder,P., Stolze, M., Smith, P., Nadia El-Hage Scialabba, N.E.H., Nigglia, U., 2012.Enhanced top soil carbon stocks under organic farming. Proc. Natl. Acad. Sci. U.S. A. 109 (44), 18226–18231.
Geisseler, D., Scow, K.M., 2014. Long-term effects of mineral fertilizers on soilmicroorganisms–a review. Soil Biol. Biochem 75, 54–63.
Giacometti, C., Demyan, M.S., Cavani, L., Marzadori, C., Ciavatta, C., Kandeler, E.,2013. Chemical and microbiological soil quality indicators and their potential todifferentiate fertilization regimes in temperate agroecosystems. Appl. Soil. Ecol.64, 32–48.
Giacometti, C., Cavani, L., Baldoni, G., Ciavatta, C., Marzadori, C., Kandeler, E., 2014.Microplate-scale fluorometric soil enzyme assays as tools to assess soil qualityin a long-term agricultural field experiment. Appl. Soil. Ecol. 75, 80–85.
Gu, Y., Zhang, X., Tu, S., Lindström, K., 2009. Soil microbial biomass, crop yields, andbacterial community structure as affected by long-term fertilizer treatmentsunder wheat-rice cropping. Eur. J. Soil Biol. 45 (3), 239–246.
Gunapala, N., Scow, K.M., 1998. Dynamics of soil microbial biomass and activity inconventional and organic farming systems. Soil Biol. Biochem. 30 (6), 805–816.
Guo, J.H., Liu, X.J., Zhang, Y., Shen, J.L., Han, W.X., Zhang, W.F., Christie, P., Goulding, K.W.T., Vitousek, P.M., Zhang, F.S., 2010. Significant acidification in major Chinesecroplands. Science 327 (5968), 1008–1010.
Hallin, S., Jones, C.M., Schloter, M., Philippot, L., 2009. Relationship between N-cycling communities and ecosystem functioning in a 50-year-old fertilizationexperiment. ISME J. 3 (5), 597–605.
Haynes, R.J., 2000. Labile organic matter as an indicator of organic matter quality inarable and pastoral soils in New Zealand. Soil Biol. Biochem. 32 (2), 211–219.
Hojjati, S., Nourbakhsh, F., 2007. Effects of cow manure and sewage sludge onthe activity and kinetics of L-glutaminase in soil. Biol. Fertil. Soils 43 (4),491–494.
Hole, D.G., Perkins, A.J., Wilson, J.D., Alexander, I.H., Grice, P.V., Evans, A.D., 2005.Does organic farming benefit biodiversity? Biol. Conserv. 122 (1), 113–130.
Jackson, L.E., Bowles, T.M., Hodson, A.K., Lazcano, C., 2012. Soil microbial-root andmicrobial-rhizosphere processes to increase nitrogen availability and retentionin agroecosystems. Curr. Opin. Environ. Sustainability 4, 517–522.
Jiang, X., Hou, X., Zhou, X., Xin, X., Wright, A., Jia, Z., 2014. pH regulates key players ofnitrification in paddy soils. Soil Biol. Biochem. doi:http://dx.doi.org/10.1016/j.soilbio.2014.10.025.
Kahrl, F., Li, Y., Su, Y., Tennigkeit, T., Wilkes, A., Xu, J.C., 2010. Greenhouse gasemissions from nitrogen fertilizer use in China. Environ. Sci. Policy 13 (8),688–694.
Kallenbach, C., Grandy, A.S., 2011. Controls over soil microbial biomass responses tocarbon amendments in agricultural systems: A meta-analysis. Agric. Ecosyst.Environ. 144 (1), 241–252.
Kustermann, B., Kainz, M., Hulsbergen, K.J., 2008. Modeling carbon cycles andestimation of greenhouse gas emissions from organic and conventional farmingsystems. Renew. Agric. Food Syst. 23, 38–52.
Kumar, S., Patra, A.K., Singh, D., Purakayastha, T.J., 2014. Long-term chemicalfertilization along with farmyard manure enhances resistance and resilience ofsoil microbial activity against heat stress. J. Agron. Crop. Sci. 200 (2), 156–162.
Li, F., Liu, M., Li, Z., Jiang, C., Han, F., Che, Y., 2013. Changes in soil microbial biomassand functional diversity with a nitrogen gradient in soil columns. Appl. Soil.Ecol. 64, 1–6.
Li, J., Zhao, B.Q., Li, X.Y., Jiang, R.B., So, H.B., 2008. Effects of long-term combinedapplication of organic and mineral fertilizers on microbial biomass, soil enzymeactivities and soil fertility. Agric. Sci. China 7 (3), 336–343.
Liang, Q., Chen, H., Gong, Y., Fan, M., Yang, H., Lal, R., Kuzyakov, Y., 2012. Effects of15 years of manure and inorganic fertilizers on soil organic carbon fractions in awheat-maize system in the North China Plain. Nutr. Cycling Agroecosyst. 92 (1),21–33.
Lin, Z.A., Zhao, B.Q., Yuan, L., So, H.B., 2009. Effects of organic manure and fertilizerslong-term located application on soil fertility and crop yield. Sci. Agric. Sin. 42(8), 2809–2819 (in Chinese).
Lu, R.K., 2000. The Analysis Method of Soil Agricultural Chemistry. ChineseAgricultural Sciences and Technology Press (in Chinese).
Mäder, P., Fließbach, A., Dubois, D., Gunst, L., Fried, P., Niggli, U., 2002. Soil fertilityand biodiversity in organic farming. Science 296 (5573), 1694–1697.
Melero, S., Porras, J.C.R., Herencia, J.F., Madejon, E., 2006. Chemical and biochemicalproperties in a silty loam soil under conventional and organic management. SoilTillage Res. 90 (1), 162–170.
Meng, L., Ding, W.X., Cai, Z.C., 2005. Long-term application of organic manure andnitrogen fertilizer on N2O emissions, soil quality and crop production in a sandyloam soil. Soil Biol. Biochem. 37, 2037–2045.
Miao, Y., Stewart, B.A., Zhang, F.S., 2011. Long-term experiments for sustainablenutrient management in China. A review. Agron. Sustainable Dev. 31 (2),397–414.
Muyzer, G., De Waal, E.C., Uitterlinden, A.G., 1993. Profiling of complex microbialpopulations by denaturing gradient gel electrophoresis analysis of polymerasechain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol.59 (3), 695–700.
Ndiaye, E.L., Sandeno, J.M., McGrath, D., Dick, R.P., 2000. Integrative biologicalindicators for detecting change in soil quality. Am. J. Altern. Agric. 15 (1), 26–36.
Olander, L.P., Vitousek, P.M., 2000. Regulation of soil phosphatase and chitinaseactivity by N and P availability. Biogeochemistry 49, 175–190.
Seufert, V., Ramankutty, N., Foley, J.A., 2012. Comparing the yields of organic andconventional agriculture. Nature 485 (7397), 229–232.
Sinsabaugh, R.L., 2010. Phenol oxidase, peroxidase and organic matter dynamics ofsoil. Soil Biol. Biochem. 42 (3), 391–404.
Sradnick, A., Murugan, R., Oltmanns, M., Raupp, J., Joergensen, R.G., 2013. Changes infunctional diversity of the soil microbial community in a heterogeneous sandysoil after long-term fertilization with cattle manure and mineral fertilizer. Appl.Soil. Ecol. 63, 23–28.
Vance, E.D., Brookes, P.C., Jenkinson, D.S., 1987. An extraction method for measuringsoil microbial biomass C. Soil Biol. Biochem. 19, 703–707.
Wessén, E., Nyberg, K., Jansson, J.K., Hallin, S., 2010. Responses of bacterial andarchaeal ammonia oxidizers to soil organic and fertilizer amendments underlong-term management. Appl. Soil. Ecol. 45 (3), 193–200.
Xie, H., Li, J., Zhu, P., Peng, C., Wang, J., He, H., Zhang, X., 2014. Long-term manureamendments enhance neutral sugar accumulation in bulk soil and particulateorganic matter in a Mollisol. Soil Biol. Biochem. 78, 45–53.
Xu, Z., Chen, C., He, J., Liu, J., 2009. Trends and challenges in soil research linkingglobal climate change to local long-term forest productivity. J. Soils Sediments 9(2), 83–88.
Yan, D., Wang, D., Yang, L., 2007. Long-term effect of chemical fertilizer, straw, andmanure on labile organic matter fractions in a paddy soil. Biol. Fertil. Soils 44 (1),93–101.
Yevdokimov, I., Gattinger, A., Buegger, F., Munch, J.C., Schloter, M., 2008. Changes inmicrobial community structure in soil as a result of different amounts ofnitrogen fertilization. Biol. Fertil. Soils 44 (8), 1103–1106.
Zak, J.C., Willig, M.R., Moorhead, D.L., Wildman, H.G., 1994. Functional diversity ofmicrobial communities: a quantitative approach. Soil Biol. Biochem. 26 (9),1101–1108.
Zhang, F., Cui, Z., Chen, X., Ju, X., Shen, J., Chen, Q., Liu, X., Zhang, W., Mi, G., Fan, M.,Jiang, R., 2012. Integrated nutrient management for food security andenvironmental quality in China. Adv. Agron. 116, 1–40.
Zhang, W.F., Dou, Z.X., He, P., Ju, X.T., Powlson, D., Chadwick, D., Norse, D., Lu, Y.L.,Zhang, Y., Wu, L., Chen, X.P., Cassman, K.G., Zhang, F.S., 2013. New technologiesreduce greenhouse gas emissions from nitrogenous fertilizer in China. Proc.Natl. Acad. Sci. U. S. A. 110 (21), 8375–8380.
Zhao, B.Q., Li, X.Y., Liu, H., Wang, B.R., Zhu, P., Huang, S.M., Bao, D.J., Li, Y.T., So, H.B.,2011. Results from long-term fertilizer experiments in China: The risk ofgroundwater pollution by nitrate. NJAS-Wagen. J. Life Sci. 58 (3), 177–183.
Zhu, Z., Chen, D., 2002. Nitrogen fertilizer use in China–contributions to foodproduction, impacts on the environment and best management strategies. Nutr.Cycling Agroecosyst. 63 (2–3), 117–127.




















