Silencing of an Anther-specific Zinc-finger Gene, MEZ1 , Causes Aberrant Meiosis and Pollen Abortion...
16
Silencing of an anther-specific zinc-finger gene, MEZ1, causes aberrant meiosis and pollen abortion in petunia Sanjay Kapoor 1,2 and Hiroshi Takatsuji 1, * 1 Developmental Biology Laboratory, Plant Physiology Department, National Institute of Agrobiological Sciences, 2-1-2 Kannondai, Tsukuba 305-8602, Ibaraki, Japan (*author for correspondence; e-mail takatsuh @nias.affrc.go.jp); 2 Department of Plant Molecular Biology, University of Delhi South Campus, Benito Juarez Road, New Delhi, 110021, India Received 9 November 2005; accepted in revised form 31 January 2006 Key words: male sterile, meiosis, petunia, ploidy, transcription factor, zinc-finger Abstract MEZ1 (MEiosis-associated Zinc-finger protein 1) was first isolated as an anther-specific cDNA from Petunia hybrida. In the present study, we report its functional characterization, including its spatial and temporal expression profiles and phenotypes in MEZ1-silenced plants. MEZ1 transcripts were specifically localized in pollen mother cells during early stages of anther development, and were later distributed in vegetative tissues in anthers. Silencing of MEZ1 by cosuppression resulted in several anomalies during male meiosis that included inability of chromosomes to condense, loss of meiotic synchrony, and premature and apparently uncontrolled cytokinetic events. Consequently, by the end of meiosis 8–10 cells, instead of the normal 4, with varying DNA contents were formed in the MEZ1-silenced meiocytes. Most of these aborted prematurely, and those that matured had a distinctive morphology. MEZ1-silenced plants were female sterile when pollinated with wild-type pollen but they infrequently produced a few seeds upon self-polli- nation. Resulting T 1 plants had increased ploidy levels and exhibited severe anomalies during male meiosis, rendering them completely sterile. We discuss possible role of MEZ1 in the proper progression of plant meiosis. Abbreviations: am1, ameiotic1; Bl, bud length; CaMV, cauliflower mosaic virus; DAP, days after pollination; DAPI, 4¢,6-diamidino-2-phenylindole; MEZ1 , MEiosis-associated Zinc-finger protein 1; cMEZ1 , MEZ1 cDNA; NLS, nuclear localization signal; PMC, pollen mother cell; SAP, STERILE APETALA; SPL/NZZ, SPOROCYTELESS/NOZZLE; SWI1, Switch1 Introduction The exact halving of chromosome number while preserving the entire genomic information is pivotal to the sexual life cycle of eukaryotes. To achieve this, homologous chromosomes in the sporogenous cells of a diploid organism must pair accurately and then segregate into two identical groups. In animals, the germ cells that undergo meiosis are set aside during early embryonic development, whereas in plants, the meiotic process begins late in the development with a de novo differentiation of archesporial cells in developing male (anthers) and female (ovules) organs of the flower. The female arche- sporial cell gives rise to a single megaspore mother cell. In anthers, the archesporial cells undergo a defined number of mitotic divisions Plant Molecular Biology (2006) 61:415–430 ȑ Springer 2006 DOI 10.1007/s11103-006-0020-0
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Silencing of an Anther-specific Zinc-finger Gene, MEZ1 , Causes Aberrant Meiosis and Pollen Abortion...

Silencing of an anther-specific zinc-finger gene, MEZ1, causes aberrantmeiosis and pollen abortion in petunia
Sanjay Kapoor1,2 and Hiroshi Takatsuji1,*1Developmental Biology Laboratory, Plant Physiology Department, National Institute of AgrobiologicalSciences, 2-1-2 Kannondai, Tsukuba 305-8602, Ibaraki, Japan (*author for correspondence; e-mail [email protected]); 2Department of Plant Molecular Biology, University of Delhi South Campus, BenitoJuarez Road, New Delhi, 110021, India
Received 9 November 2005; accepted in revised form 31 January 2006
Key words: male sterile, meiosis, petunia, ploidy, transcription factor, zinc-finger
Abstract
MEZ1 (MEiosis-associated Zinc-finger protein 1) was first isolated as an anther-specific cDNA fromPetunia hybrida. In the present study, we report its functional characterization, including its spatial andtemporal expression profiles and phenotypes in MEZ1-silenced plants. MEZ1 transcripts were specificallylocalized in pollen mother cells during early stages of anther development, and were later distributed invegetative tissues in anthers. Silencing ofMEZ1 by cosuppression resulted in several anomalies during malemeiosis that included inability of chromosomes to condense, loss of meiotic synchrony, and premature andapparently uncontrolled cytokinetic events. Consequently, by the end of meiosis 8–10 cells, instead of thenormal 4, with varying DNA contents were formed in the MEZ1-silenced meiocytes. Most of these abortedprematurely, and those that matured had a distinctive morphology. MEZ1-silenced plants were femalesterile when pollinated with wild-type pollen but they infrequently produced a few seeds upon self-polli-nation. Resulting T1 plants had increased ploidy levels and exhibited severe anomalies during male meiosis,rendering them completely sterile. We discuss possible role of MEZ1 in the proper progression of plantmeiosis.
Abbreviations: am1, ameiotic1; Bl, bud length; CaMV, cauliflower mosaic virus; DAP, days afterpollination; DAPI, 4¢,6-diamidino-2-phenylindole; MEZ1 , MEiosis-associated Zinc-finger protein 1;cMEZ1 , MEZ1 cDNA; NLS, nuclear localization signal; PMC, pollen mother cell; SAP, STERILEAPETALA; SPL/NZZ, SPOROCYTELESS/NOZZLE; SWI1, Switch1
Introduction
The exact halving of chromosome number whilepreserving the entire genomic information ispivotal to the sexual life cycle of eukaryotes.To achieve this, homologous chromosomes inthe sporogenous cells of a diploid organism mustpair accurately and then segregate into twoidentical groups. In animals, the germ cells that
undergo meiosis are set aside during earlyembryonic development, whereas in plants, themeiotic process begins late in the developmentwith a de novo differentiation of archesporialcells in developing male (anthers) and female(ovules) organs of the flower. The female arche-sporial cell gives rise to a single megasporemother cell. In anthers, the archesporial cellsundergo a defined number of mitotic divisions
Plant Molecular Biology (2006) 61:415–430 � Springer 2006DOI 10.1007/s11103-006-0020-0

and differentiate into anther wall layers, tapetalnurse tissue, and numerous pollen mother cells(Yang et al., 1999; Yang and Sundaresan, 2000).Finally, megaspore and pollen mother cellsundergo meiotic divisions to generate femaleand male gametophytes, respectively.
Meiosis and mitosis share various cytologicalprocesses in common. Both these processesinvolve condensation of chromosomes followedby their characteristic alignment at metaphase,separation of sister chromatids/homologouschromosomes at anaphase, grouping of twonucleoids at telophase, and finally physical par-titioning of the nucleo-cytoplasmic domains bycytokinesis. Molecular dissection of the meioticcell division in a number of organisms hasrevealed that a large number of the componentsinvolved are also shared between mitosis andmeiosis. There are, however, several importantdissimilarities that differentiate these two pro-cesses. The most important of them is the pairingand recombination of homologous chromosomesin meiosis that sets the stage for reductionaldivision. It is followed by attachment of sisterkinetochores to spindles with the same polarity,ensuring segregation of homologs and not sisterchromatids during the first meiotic division. Inthe last two decades, a number of cell divisioncomponents involved in chromosome condensa-tion, sister chromatid/homologous chromosomecohesion, kinetochore–spindle attachment/align-ment, and cytokinesis have been identified. How-ever, only limited information is available aboutthe regulatory factors that control functioning ofthese components in a mitosis- or meiosis-specificmanner.
In yeast, the initiation of meiosis is controlledby a master regulator, Ime1. IME2, one ofdownstream regulatory factors of Ime1, encodesa serine–threonine protein kinase (Mitchell et al.,1990), and is required for meiotic DNA replication(Foiani et al., 1996), G1-S transition, and phos-phorylation-dependent degradation of Ime1 there-by shutting down the meiotic transcriptionalcascade. In Arabidopsis, the SPOROCYTELESS/NOZZLE (SPL/NZZ) gene, which encodes anuclear protein with limited sequence similarity tothe MADS-box domain, has been implicated inearly germ-line specification (Yang et al., 1999).Two other Arabidopsis gene, TAPETUM DETER-MINANT1 (Yang et al., 2003), which codes for a
small protein of 176 amino acids, and EXCESSMICROSPOROCYTES1/EXTRA SPOROGE-NOUS CELLS (Canales et al., 2002; Zhao et al.,2002), which codes for a putative leucine-richrepeat receptor protein kinase (LRRRPK), havebeen implicated in the specification of the correctnumber of male archesporial initials and subse-quent specification of tapetal and middle cell layeridentities. Ameiotic1 (am1) has been proposed tobe involved in the mitosis-to-meiosis switch inmaize. The am1 locus is crucial for the initiation ofboth male and female meiosis: am1 meiocytes,instead of entering meiotic prophase, undergo amitosis-like division and then degenerate (Gol-ubovskaya et al., 1997). Like Ime1 of yeast, plantAM1 could be involved in the regulation of earlymeiosis genes, such as Switch1 (SWI1), which inturn are involved in the establishment of sisterchromatid cohesion (Mercier et al., 2001, 2003).STERILE APETALA (SAP) is another transcrip-tion factor that is implicated in female meiosis(Byzova et al., 1999). Sap ovules progress tomeiosis I, but fail to proceed to the second meioticdivision.
We have previously reported identification andexpression profiling of several zinc-finger tran-scription-factor genes that express in a floral-organ specific manner in Petunia hybrida(Takatsuji, 1999). Seven such genes express in ananther-specific/preferential manner, while two ex-press specifically in the pistil (Kobayashi et al.,1998; Kubo et al., 2000). Of the anther-specific/preferential zinc-finger genes, the expression ofonly two – TAZ1 and MEZ1 (Meiosis-associated Zinc-finger protein 1; renamed fromPEThy:ZPT2-5) – was detected during pre-meioticstages of anther development (Kobayashi et al.,1998; Kapoor et al., 2002). Silencing of TAZ1caused defective development of tapetum (Kapooret al., 2002).
Here we show that silencing of MEZ1 severelyaffects the processes of male meiosis. MEZ1-silenced transgenic plants show severe defects inchromosome condensation. The aberrant meiosisusually resulted in unequal distribution of DNAinto daughter cells and excessive cell division,yielding up to 10 microspores with varyingamounts of DNA. Aberrance was also found infemale gametophytes. The data suggest thatMEZ1 is essential for proper progression of malemeiosis and possibly also in female meiosis.
416

Materials and methods
Plasmid construction
35S::MEZ1A 790-bp SalI–NotI fragment containing thecomplete MEZ1 cDNA (cMEZ1) sequence (Ko-bayashi et al., 1998) was excised from the pSPORTvector by using KpnI and SacI. This fragment wasinserted between the CaMV35S promoter and theNos terminator, which were already cloned in theSalI and EcoRI sites, respectively, of pUCAP (vanEngelen et al., 1995). This synthetic gene contain-ing 35S:cMEZ1:Tnos was excised from pUCAPby using AscI and PacI, and was inserted intopBINPLUS (van Engelen et al., 1995) with thesame set of enzymes.
35S:GFP:MEZ1The complete MEZ1 coding sequence was PCR-amplified by using two oligonucleotide primers,5¢-ACTAGGGCCCATGGTGGCTCTATCAACGAAGAGA-3¢ and 5¢-ACTAGGGCCCTTCATAAAAAGCAATCGACCATAGGA-3¢. The under-lined regions denote XmaI sites that were added tofacilitate the cloning of amplified DNA. Afterdigestion with XmaI the coding region of MEZ1was ligated to the XmaI-linearized psGFPcs vector(Jiang et al., 2001), between the green fluorescentprotein (GFP) coding sequence and the Nosterminator.
Plant transformation
The DNA constructs were introduced into petuniaplants by Agrobacterium tumefaciens (GV3101)-mediated transformation (Jorgensen et al., 1996).Transient expression of the 35S:GFP:MEZ1 genein onion peel cells was performed as described byJiang et al. (2001). The fluorescence of syntheticGFP fusion proteins was observed under a confo-cal microscope (Bio-Rad Laboratories, Hercules,CA, USA), and the resultant digital micrographswere assembled in Photoshop software (AdobeSystems Inc., San Jose, CA, USA).
Expression analysis
RNA gel blot analysisTotal RNA from petunia anthers was isolated byusing the procedure developed by Logemann et al.
(1987) and stored at )70 �C in 95% ethanol.Electrophoretic separation of RNA, blotting ontomembranes, and hybridization with a labeledprobe were performed as described previously(Kapoor et al., 2002). An antisense RNA probewas prepared from the 190-bp KpnI–SpeI subcl-oned fragment (Figure 4) by using a digoxigeninRNA labeling kit according to the manufacturer’sinstructions (Boehringer Mannheim, Mannheim,Germany).
RT-PCRRT-PCR was carried out by using SuperscriptTM
one-step RT-PCR system (Invitrogen) followingmanufacturer’s instructions. The cDNA synthesiswere done using 5 lg (anther) or 20 lg (ovary andseed) of total RNA as templates for 30 min at50 �C followed by 35 cycles of PCR (94 �C, 15 s;63 �C, 30 s; 72 �C, 1 min) using sense (5¢-ATGGTGGCTCTATCAACGAAGAGA-3¢) and anti-sense (5¢-TTCATAAAAAGCAATCGACCATAGGA-3¢) primers.
In situ hybridizationIn situ hybridization was carried out as describedpreviously (Kapoor et al., 2002). Antisense andsense RNA probes was prepared from threeregions – 190-bp KpnI–SpeI, 190-bp SpeI–HindIII,and 410-bp HindIII–SacI (Figure 4) – by using adigoxigenin RNA labeling kit (Boehringer Mann-heim), and then pooled before hybridization.
Light microscopy
Nucleus size measurementsThe anthers from the Bl-9-mm stage were fixed inethanol:acetic acid (9:1) for 3 h. These antherswere opened with the help of two needles directlyon a glass slide, and the released microspores werestained with 10 ll of 4¢,6-diamidino-2-phenylin-dole (DAPI) solution (1 lg/ml in 10% glyc-erol). Fluorescent signals were viewed under aLeica DMR microscope (Leica Microsystems,Inc., Wetzlar, Germany), and the fluorescingregion in each microspore was measured with thehelp of the QWIN Image Processing and AnalysisSystem (Leica).
Microscopic characterization of meiocytesChromosome spreads of male meiocytes from theBl-3- to 5-mm stages were prepared essentially
417

according to Ross et al. (1996) and stained with10 ll DAPI (1 lg/ml). Fluorescent signals wereviewed and photographed through a Leica DMRmicroscope.
Seed clearingWhole seeds at various stages of development werefixed in ethanol:acetic acid (9:1) overnight, washedtwice in 90% ethanol, and then mounted inclearing solution (2.5 g chloral hydrate in 1 ml30% glycerol) (Ruzin, 1999). The samples wereobserved and photographed through Nomarskioptics on a Leica DMR microscope. Color mask-ing of the relevant cells or tissues in digital pictureswas done in Adobe Photoshop.
Endosperm cellularizationSelf-fertilized seeds (11 DAP) from wild-type andMEZ1-silenced plants were fixed in HistochoiceTissue Fixative (Sigma, St. Louis, MO, USA),dehydrated through a series of ethanol-aqueoussolutions (10%, 30%, 50%, 70%, 90%, and 100%,30 min each), and embedded in Technovit 7100resin (Heraeus Kulzer GmbH, Wertheim, Ger-many). The embedded tissues were sectioned on arotary microtome (Mikron Instruments Inc., SanMarcos, CA, USA). For visualization of cellularstructure, the sections were stained with toluidineblue (Ruzin, 1999) and observed with Nomarskioptics. For assessing the extent of cellulosic wallformation in endosperm tissue, some sections werestained with calcofluor and observed under the UVoption of the Leica DMR microscope (Ruzin,1999).
Acetocarmine staining of chromosomesShoot apices containing three or four young leaveswere fixed in 90% ethanol:10% acetic acid over-night and softened in 45% acetic acid at roomtemperature for 5 min. The softened tissue wasplaced on a glass slide, and the meristematicportion was dissected out in a drop of 45% aceticacid. After the liquid was blotted off, a drop of 1%acetocarmine/45% acetic acid solution was over-laid to stain the chromosomes. After 2–5 min, acover slip was firmly pressed onto the tissue, excessliquid was blotted off, and the tissue was sealedwith clear nail varnish. The slides were observedand photographed by using bright field optics on aLeica DMR microscope.
Results
MEZ1 expression in anther
MEZ1 (PEThy:ZPT2-5) expresses specifically inanthers during early stages of flower development(Kobayashi et al., 1998). Figure 1A presents adetailed RNA gel blot analysis of MEZ1 fromanthers at 17 stages of flower development asdescribed in Kapoor et al. (2002). The first stage(buds lengths=3 mm; Bl-3 mm) corresponds tothe pollen mother cell (PMC) stage of microsporedevelopment, which is followed by meiosis in theBl-4- to 5-mm stages. The microspore tetradsresulting from male meiosis are visible in 5- to 5.5-mm buds. In 6- to 30-mm buds, the microsporesgrow but remain uninucleate. Later, an unequalmitotic division divides the pollen into a vegetativecell and a generative cell (binucleate). Meanwhile,the tapetum, which is evident even during thePMC stage, reaches the peak of its developmentby the Bl-6- to 8-mm stages, and then graduallystarts to degenerate after buds are 10–11 mm. Bythe Bl-35-mm stage, the tapetum completelydisappears from anthers. A small amount ofMEZ1 transcript was detectable in the 3-mmbuds. However, coinciding with the onset ofmeiosis (Bl-4-mm) the MEZ1 transcript levelincreased by approximately 10 times. The high-level MEZ1 expression continued until Bl-11-mm,and then declined precipitously to a level equiv-alent to that in Bl-3-mm stage. This decline inMEZ1 expression appears to coincide with theonset of tapetum degeneration. We verified thatthe hybridization probes used in this and follow-ing experiments did not cross-hybridize with otherzinc-finger genes in separate RNA and DNA gelblot experiments.
The cell-type-specific expression pattern ofMEZ1 was analyzed by in situ hybridization ontransverse sections of developing flower buds(Figure 2). Before Bl-3-mm stage, we did notdetect any expression (data not shown). At theBl-3-mm stage, MEZ1 was found to be expressedpredominantly in the PMCs, and partly in theouter tapetal cells. The other cell types, includingwall layers, stomium, vascular tissue, and the innertapetum, did not show any MEZ1 transcripts atthis stage (Figure 2B). In later stages, MEZ1transcripts were also detected in the vegetativetissues, such as tapetum, connective and anther
418

wall cells and appeared to decline in the microsp-ores as they developed (Figure 2D–H).
GFP–MEZ1 chimeric protein accumulatesin the nucleus
TFIIIA-type zinc finger motifs have been found ina number of eukaryotic transcription factors andimplicated in both protein–DNA and protein–protein interactions (Mackay and Crossley, 1998;Takatsuji, 1999). The PSORT program predictedMEZ1 to be a nuclear-localized polypeptide with a
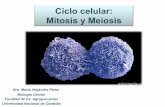
certainty of 94.1%, because it contains the classi-cal type of nuclear localization signals (NLSs),with 3 pat4 and 1 pat7 sequence patterns startingat amino acids 72, 90, 115, and 90, respectively(http://psort.nibb.ac.jp/). The sub-cellular locali-zation of MEZ1 was experimentally examined byvisualizing the accumulation of transientlyexpressed GFP–MEZ1 fusion proteins in onionpeel cells. As shown in Figure 3 (right panels),almost all the fluorescence of GFP–MEZ1 fusionproteins was concentrated in the nucleus, whileGFP alone was distributed throughout the
Figure 1. Expression patterns of MEZ1. (A) Temporal expression pattern of MEZ1 in anther. RNA gel blot analysis using total
RNA from anthers of wild-type plants and antisense digoxigenin-labeled MEZ1 RNA as a probe. The numbers at the top of each
lane represent the length of flower buds in millimeters. A graphic representation of the development of tapetum and pollen corre-
sponding to the size of flower buds is displayed at bottom. PMC, pollen mother cells. (B) RT-PCR analysis of MEZ1 expression in
anther, ovary and seeds. Anthers (Bl-2- to 3-, 4- to 5-, and 10- to 15-mm) and ovaries (Bl-10- to 15-mm) of young flower buds and
seeds (0–10 DAP) were analyzed.
419

cytoplasm (Figure 3; left panels). These resultssuggest that MEZ1 protein functions in thenucleus presumably as a transcription factor.
Production of MEZ1-silenced plants
To characterize the function of MEZ1, we placedthe 790-bp MEZ1 cDNA downstream of thecauliflowermosaic virus 35S (CaMV 35S) promoter
(Figure 4A) and introduced into petunia byAgrobacterium tumefaciens-mediated transforma-tion. The presence of the transgene in each line oftransformants was confirmed by PCR amplificationof the neomycin-phosphotransferase gene (data notshown). Sixteen transgenic lines were raised tomaturity; none of them displayed any morpholog-ical aberrance under normal greenhouse condi-tions. All these plants were analyzed for the
Figure 2. In situ hybridization for MEZ1. Transverse sections of anthers from Bl-3- (A and B), Bl-5- (C and D), Bl-8-mm (E and
F) and Bl-13-mm (G and H) stages were probed with sense (A, C, E and G) and antisense (B, D, F and H) MEZ1 probes. PMC,
pollen mother cells; T, tapetum; Mi, microspore; Co, connective tissue; W, wall layer. Bars=200 lm.
420
accumulation of MEZ1 transcripts by gel-blotanalysis of the total RNA extracted from pooledanthers of Bl-3- to 8-mm stages (two buds at every1-mm interval), representing both pre- and post-meiotic phases of anther development (Figure 4B).Thirteen of these lines (1, 3, 8, 10–16, 18–20)showed either increased, comparable, or somewhatreduced levels of MEZ1 transcript accumulationrelative to that in the wild-type plants (Figure 4B).These MEZ1-expressing transgenic lines did notshow any alterations in pollen development orfertility. Meanwhile, three independent lines (6, 7,and 9) did not show any detectable MEZ1 tran-scripts, indicating silencing of endogenous MEZ1by cosuppression in these plants. The expression ofTAZ1 (Kobayashi et al., 1998; Kapoor et al., 2002),which has the highest sequence similarity toMEZ1among anther-specific zinc-finger genes of petuniaand is expressed in a pattern overlapping spatiallyand temporally with that ofMEZ1, was unaffectedin these MEZ1-silenced lines (data not shown),indicating that the silencing was specific to MEZ1.These transformant lines showed anomalies inpollen development. They had extremely low pollen
counts at the time of dehiscence and the seed setresulting from self-pollination was reduced to lessthan 1% of that of the wild type (data not shown).These results indicate that MEZ1 expression isessential for normal development of pollen.
Phenotypic characterization of MEZ1-silencedplants
Pollen developmentIn the MEZ1-silenced lines, the number of maturepollen grains was reduced by approximately 80%,possibly by premature abortion of large number ofpollen grains: average pollen count per antherwere 30,309±3030 (n=6, from three flowers) inwild type plants and only 6342±444 (n=6, fromthree flowers) in MEZ1-silenced plants. Whenimmature microspores (Bl-9-mm stage) wereobserved after their nuclei were stained withDAPI, there was extensive variation in the sizesof nuclei. In wild-type petunia, most microsporenuclei (�90%) ranged in size from 1.6 to 3.4 lm2.By contrast, only 40% of microspore nuclei in theMEZ1-silenced plants fell into this size category(Figure 5A). Many of those in the MEZ1-silencedplants were very small (about 39% were<1.6 lm2), and most of the rest were somewhatlarger than those in wild-type plants on average(about 21% were >3.6 lm2). Almost all thesurviving microspores in MEZ1-silenced anthersappeared to be ‘‘square’’, in contrast to the usualtriangular shape of wild-type microspores(Figures 5B, C). Under the scanning electronmicroscope, the wild-type microspores were invari-ably tricolpate (Figure 5D), whereas those inMEZ1-silenced anthers had an extra germina-tion pore (colpus), making them tetracolpate(Figure 5E). At the end of meiotic division, insteadof four microspores of equal size forming a tetradin the wild-type anther, there were usually 7–9 cellswith variable DNA content enclosed in the tetrad-like structure in MEZ1-silenced anthers (Fig-ures 5F, G).
Microscopic observation of the sections ofMEZ1-silenced anthers revealed no morphologicalaberrations in the sporophytic tissues, in contrastto TAZ1-silenced anthers, in which tapetumdevelopment was defective (Kapoor et al., 2002).However, it needs further investigation to con-clude whether or not MEZ1 plays any role in thesetissues.
Figure 3. MEZ1 is a nuclear-localized protein. DNA con-
structs encoding synthetic GFP (left) and chimeric synthetic
GFP–MEZ1 (right) proteins under the control of the CaMV
35S promoter were bombarded into onion peel cells, and the
subcellular localization of transiently expressed proteins was
observed by visualization of GFP fluorescence through a
confocal microscope. Top, GFP fluorescence; middle, light
images; bottom, digital overlays.
421

Anomalies during male meiosis
The abnormal structure of meiocytes at tetradstage in MEZ1-silenced anthers led us to investi-gate the processes of meiosis. In wild type, thecondensation of chromatin starts when PMCs areattached to each other by common callose walls(Figures 6A, B). Condensation begins synchro-nously in all the PMCs within a locule. Thissynchrony is maintained throughout the process ofmeiosis, even after the separation of the meiocytesafter metaphase I (Figure 6C). Thereafter, thechromosomes are pulled to opposite poles (Fig-ure 6D), partially decondense at telophase I (Fig-ure 6E), then recondense and realign in the centerof the cells at metaphase II (Figure 6F). The sisterchromatids separate from each other at anaphaseII (Figure 6G) and form four daughter cells bycytokinesis (Figures 6H, I). By characterizing alarge number of anthers from the three MEZ1-silenced plants, the whole process of meiosis inthese plants was found to be distinctly differentfrom that in the wild type. Typical phenotypes atvarious stages of meiosis observed in the anthers(Bl-3- to 6-mm) of MEZ1-silenced lines are shownin Figure 6M–U. In accordance with MEZ1
expression in wild-type PMCs, the first distin-guishing feature recognized in these cells was anaberration in chromatin condensation in the veryearly meiocytes. There were no chromatin threadsvisible in any meiocytes at this stage (Figure 6M).We did not observe any structures that apparentlycorresponded to sub-stages from uncondensed tofully condensed chromosomes in the MEZ1-silencedlines, in spite of extensive microscopic observation.Subsequently, cells were seen with partial distribu-tion of the nuclear materials into unequal nucle-oid-like structures (Figure 6N–P). In panel N, thenuclear material is in the process of dividing into twounequal bodies without any visible separation ofindividual chromosomes. In the anthers of thisstage, at least 50% cells showed division of nuclearmaterials into two or three unequal parts in all theanthers from the three transformants. Each chro-mosomal separation (and nuclear division)appeared to be immediately followed by cellulardivision in these cells (Figure 6Q–S), whereascellular division occurs only after separation intofour sets of chromosomes in wild-type meiocytes.Moreover, the nuclear and cellular divisions con-tinued until there were eight or nine cells in morethan 90% of meiocytes (Figure 6T, U).
Figure 4. Analysis of MEZ1 transcripts in 35S::MEZ1 transgenic petunia plants. 35S::MEZ1 transgenic petunia plants were ana-
lyzed RNA gel blot hybridization using total anther RNA and antisense digoxigenin-labeled MEZ1 RNA as a probe. The numbers
at the top of each lane represent individual transgenic lines and a wild-type (WT) plant.
422

The microspore development program beyondmeiosis seems to have been largely unaffected bythe abolition of MEZ1 expression. Soon after thedissolution of the callose wall the individual wild-type microspores appear as uniform uninucleatestructures (Figure 6J). These undergo a mitoticdivision to form a vegetative cell and a generative
cell. By this stage, pollen apertures also becomedistinctly visible (Figure 6K). Further develop-ment is marked by an increase in the volume ofindividual microspores (Figure 6L). In MEZ1-silenced plants, the separation of individual mi-crospores suggests that decomposition of callosewalls occurred as in the wild type (Figure 6V). Theresultant microspores, however, were of varioussizes and contained variable amounts of DAPI-stainable nuclear material (Figure 6V). Mitoticdivision in these microspores gave rise to two-celled mature pollen grains similar to those in thewild type (Figure 6X). Of note, smaller microsp-ores with less DNA content than wild type haddisappeared from the Bl-35-mm stage onwards,and those that survived had more than the usualDNA content and a distinct morphology(Figure 6W).
Female fertility and seed development
After self-pollination, the fruits in MEZ1-silencedlines initiated seed development and grew, but thesize of the fruit and the number of developing seedswere approximately 1/3 of those in the wild type(Figure 7H). Then, 9 DAP, most of the seeds beganto shrink and eventually aborted by 13 DAP,leaving few mature seeds per fruit (Figure 7I). Asimilar seed development pattern followed evenwhen wild-type pollen was used to pollinateMEZ1-silenced flowers, except that the size of the fruit wasmuch smaller and no viable seed was obtained(data not shown). To investigate whether theaborted seeds bore any embryos, we comparedthe seeds from self-pollinated MEZ1-silencedplants with those of the wild type (Figure 7A–G).
Figure 5. Pollen phenotypes in MEZ1-silenced plants. (A)
Two-dimensional size measurement of DAPI-stained mature
pollen nuclei in wild-type (WT) and MEZ1-silenced (mez1)
plants. The graph represents scores for �1000 pollen nuclei
obtained from three individual undehisced anthers of the line
9 T0 plant at the Bl-65-mm stage. (B, C) Safranine-stained
mature pollen in wild-type (B) and MEZ1-silenced (C) plants
as visualized under Nomarski optics. Germination pores are
indicated by black arrowheads. (D, E) Scanning electron
microscopic images of mature pollen in wild-type (D) and
MEZ1-silenced (E) plants. (F, G) DAPI-stained microspores
of tetrad stage from 4-mm flower buds from wild-type (F)
and MEZ1-silenced (G) plants. Small cells with low DNA
content are marked with white arrowheads. Bars=20 lm.
The line 9 T0 plant was used for the data in the panels B–G.
b
423

424

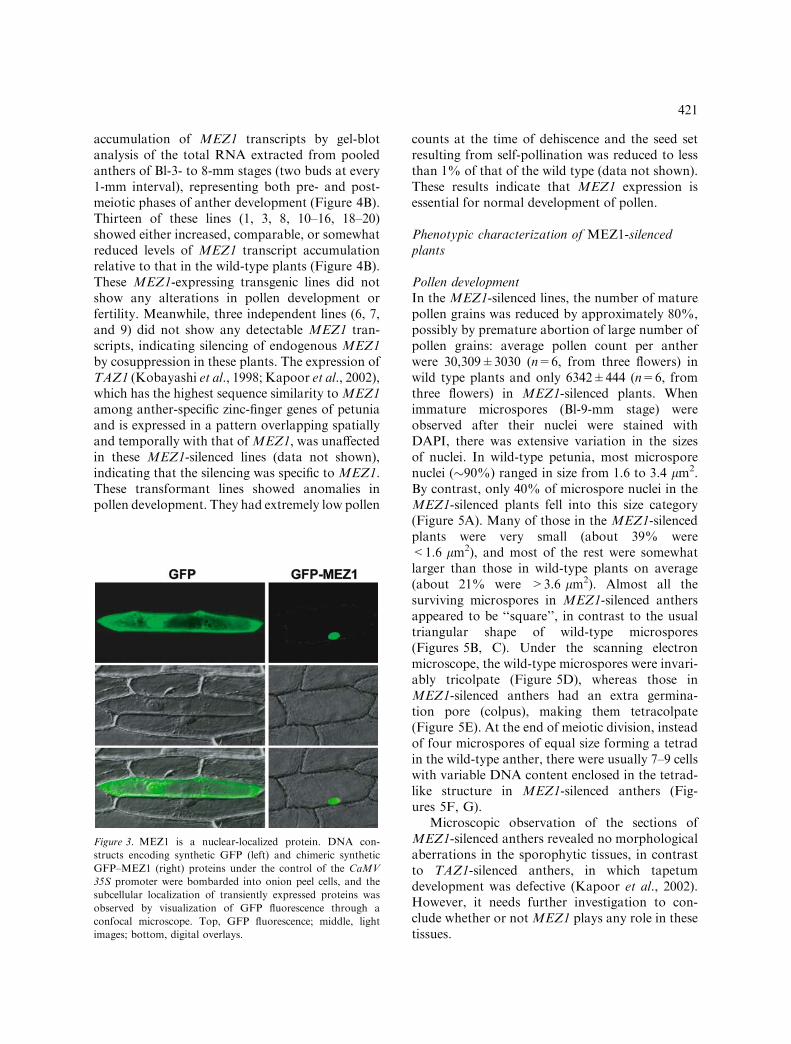
In wild-type seeds the first zygotic division occurredat 4 DAP and the quadrant stage was seen at6 DAP. By 10 DAP the embryo was at the lateglobular stage. In the heart-shaped embryo at11 DAP, the cells destined to become cotyledons,hypocotyl, and shoot apical meristem could bedistinguished. The embryos were fully developedby 16 DAP.
The initial development of embryos in theMEZ1-silenced seeds was indistinguishable fromthat in the wild type (data not shown). Theembryos developed to the late globular or earlyheart-shape stage by 10 or 11 DAP (Figure 7G).However, no further development was observed.When the seeds at 11 DAP were pressed open torelease the endosperm, the embryo and the endo-sperm appeared much more rigid than those in thewild-type seeds at the same developmental stage.Transverse sections of the seeds revealed that theendosperm was highly cellularized in the 11 DAPMEZ1-silenced seeds in comparison to the wild-type ones (Figure 7J, L). Staining of the tissuesections with calcofluor indicated that there wasextensive deposition of cellulose in endosperm cellwalls ofMEZ1-silenced seeds (Figure 7K, M). Thecellulose deposition was exceptionally high on the
outer surface of the peripheral cell layers of theendosperm. We attempted to detect MEZ1 expres-sion in developing pistils and female gametophytesby RT-PCR (Figure 1B) and in situ hybridization(data not shown), however, we did not detect anyexpression in these organs.
Phenotype of T1 plants
Asmentioned above, the self-pollination ofMEZ1-silenced plants resulted in very few seeds. Twentyof these seeds were germinated and grown tomaturity under greenhouse conditions. Morpho-logically, these T1 plants were indistinguishablefrom both the wild-type and theMEZ1-silenced T0
plants. However, analysis of male meiosis revealeda more severe phenotype than in the T0 plants.There were usually more than 10 cells in the tetrad-like structure after meiosis, and the mature pollencount was markedly reduced (data not shown).These T1 plants did not produce any viable seeds.
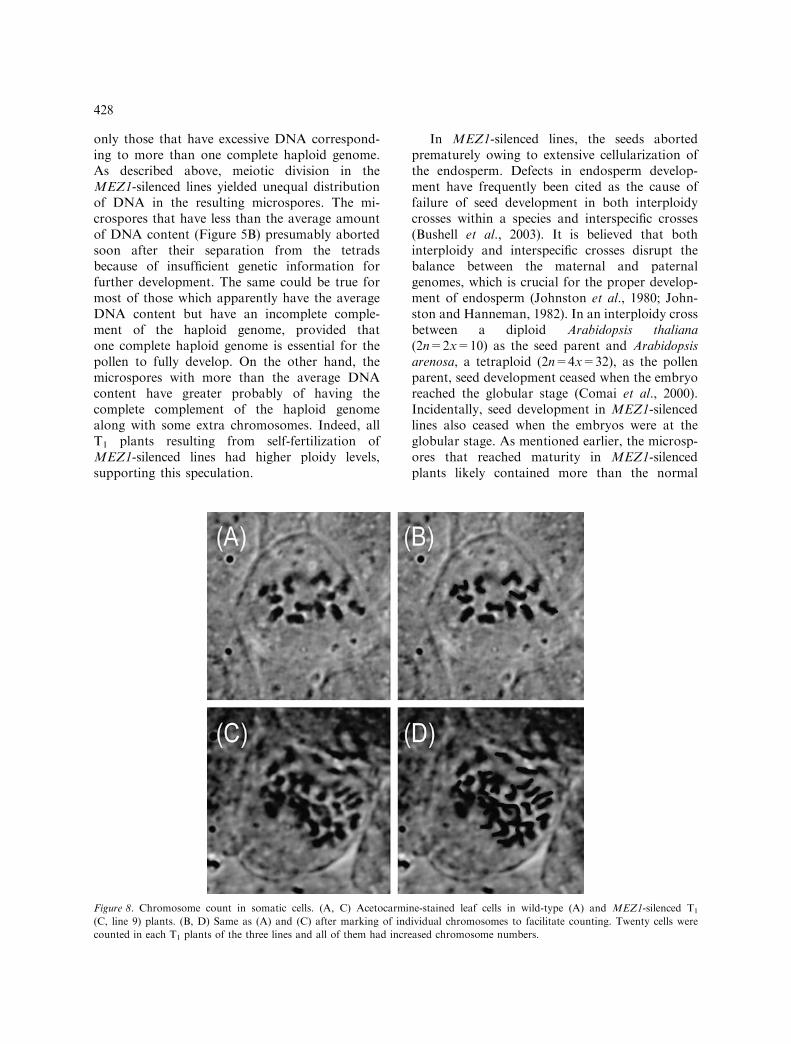
The T1 plants were suspected to have resultedfrom pollen with altered DNA content, so weinvestigated chromosome number in the cells ofshoot apical meristems and young leaves. Aceto-carmine staining of the chromosomes in thesevegetative tissues reveal increases in the chromo-some number in all the T1 plants analyzed.Representative shoot apical meristem cells in aT1 plant show 33 chromosomes (Figure 8C, D)instead of the 14 in the wild type (Figure 8A, B).
Arabidopsis homologue of MEZ1
In an attempt to see whether Arabidopsis has acounterpart of MEZ1, we characterized Ds inser-tion lines for ZAT12 (AT5G59820), an Arabidop-sis gene that is the most homologous to MEZ1.However, the two insertion lines from RIKENthat have Ds insertions in 5¢ UTR (53-3682-1) andin the coding region (13-1968-1) did not show anyphenotype like those of MEZ1-silenced petunia.Arabidopsis has some other MEZ1 homologuegenes that are nearly as homologous to MEZ1 asZAT12, therefore, it is possible that one or more ofthem is responsible for the functions correspond-ing to those of MEZ1 in petunia. It is also possiblethat MEZ1-like function of ZAT12 might beobscured due to redundancy of gene functions.
Figure 6. Processes of meiosis in wild-type and MEZ1-
silenced plants. Fluorescence micrographs of DAPI-stained mi-
crosporocytes at meiotic and post-meiotic stages in wild-type
(A–L) and MEZ1-silenced (M–X) plants. Sixty anthers each
from three MEZ1-silenced transformant lines were observed
and all the three lines showed consistent results. Shown in the
panels M–X are those of the line 9 T0 plant. (A–L) Meiosis in
wild-type plants. (A) Early prophase I. (B) Late prophase I.
(C) Metaphase I. (D) Anaphase I. (E) Telophase I. (F) Meta-
phase II. (G) Early anaphase II. (H) Tetrads. (I) Single
wild-type pollen tetrad. (J) Uninucleate microspores soon after
their separation from tetrads at Bl-6- to 7-mm. (K) Binucleate
microspores at Bl-30-mm. (L) Mature pollen at Bl-65-mm.
(M–X) Meiosis in MEZ1-silenced plants. (M) Chromosome
condensation has not begun. (N) Initiation of chromosome
condensation; distribution of DNA into two unequal nucleo-
plasmic domains in the central cell. (O, P) Formation of two or
three nucleoplasmic domains. (Q) Unequal nucleoplasmic divi-
sion caused by invagination of cell wall. (R) Further nuclear
division in one of the daughter cells. (S) Non-synchronous cell
divisions. (T) Formation of tetrad-like structures with numer-
ous unequal cells and nucleoplasmic domains. (U) Single
tetrad-like structure with nine unequal cells. (V) Uninucleate
microspores soon after their separation from tetrads showing
no uniformity in size. (W) Three of the larger microspores with
excessive DNA content. (X) Mature binucleate pollen with
four germination pores.
b
425
426

Discussion
The data presented here indicate that expression ofMEZ1 is essential for the proper progression ofmale meiosis. In accordance with its expression inPMCs, the effects ofMEZ1 silencing are evident atas early as the PMC stage. The male meiocytes inMEZ1-silenced plants displayed very limited chro-mosome condensation during early stages ofprophase I. A pentameric protein complex, con-densin, has been implicated in the premitoticcondensation of chromosomes in yeast, Xenopus,Drosophila, human, and Arabidopsis (Hirano et al.,1997; Kimura and Hirano, 2000; Hirano, 2002;Page and Hawley, 2003). A recent report bySiddiqui et al. (2003) showed that transgenic andmutant Arabidopsis plants with reduced levels ofAtCAPs, components of condensin complex,showed extensive defects in chromosome conden-sation and segregation during meiosis and wide-spread abnormalities during vegetative tissues. Themeiotic phenotype of these plants shows consider-able similarity to that of the MEZ1-silencedplants. This could provide further clues to thefunction of MEZ1.
Another notable feature of meiosis in MEZ1-silenced plants was an aberrant cytokinesis thatimmediately followed every nuclear division in-stead of taking place once after telophase II.Microspore cytokinesis is a unique event that isnot observed elsewhere in plant development. It
starts with the distinctive deposition of callosebetween the plasma membrane and cell wall duringdifferentiation of microspore mother cells. Thecallose remains a major constituent of the inters-poral walls in pollen tetrads formed after meiosis,and is later degraded, resulting in the release ofindividual haploid microspores (Echlin and God-win, 1968; Stieglitz, 1977). Two kinds of mecha-nism of intersporal cell-wall formation are known– successive and simultaneous. The successive typeis more common in monocots, where cell walls areassembled after both telophases I and II. Thesimultaneous type is a regular feature in dicots,including Arabidopsis and petunia, where cell wallsbetween all four resulting nuclei form at the sametime, that is, after telophase II. Successive cytoki-nesis is generally characterized by the formation ofphragmoplast-associated cell plates after eachnuclear division that extend from the center tothe periphery of the cell. By contrast, simultaneouscytokinesis initiates as ingrowths from the callosewall, which expand centripetally until the microsp-ores are separated (Brown and Lemmon, 1988,2001; Otegui and Staehelin, 2000). Notably, theinitiation of cell-wall formation in MEZ1-silencedmeiocytes was typical of the simultaneous-typecytokinesis observed in the wild-type petuniaplants, i.e., initiating as ingrowths from the cellwall. On the other hand, the timing of wallformation was similar to that in the successivetype, i.e., every nuclear division was followed bywall formation. However, unlike in the successive-type cytokinesis, wall formation did not stop aftertwo cycles of division, but continued until therewere eight or nine cells in a tetrad-like structure.Taken together, these observations suggest thatthe timing of cytokinesis and the way cells wallsare formed are two independent processes and thatMEZ1 silencing affected only one of the twoprocesses. It remains to be clarified whether thisphenotype reflects direct involvement of MEZ1 inthe cytokinesis or it is a secondary effect resultingfrom earlier meiotic defects.
The mature pollen grains in MEZ1-silencedlines exhibited distinctive morphology and largervolume: they had four apertures (colpi) instead ofthe three in the wild-type pollen and were roughlytwice as voluminous on average. It is generallyknown that the volume of pollen increases pro-portionally to the amount of DNA materialswithin them. These microspores are presumably
Figure 7. Development of embryo and endosperm in wild-
type and MEZ1-silenced plants. (A–F) Stages of embryo
development from 4 to 16 DAP during wild-type seed devel-
opment. (A) Zygote. (B) two-celled; dividing. (C) eight-celled.
(D) Late globular. (E) Heart stage. (F) Fully developed
embryo with cotyledons (green), meristem (yellow), and hypo-
cotyl (blue). Suspensor cells are marked by red in (B), (C),
(D), (E) and (G). (G) An embryo in a MEZ1-silenced plant
arrested at the globular stage. (H, I) Opened fruits of wild-
type (WT) and MEZ1-silenced (mez1) self-pollinated plants at
5 DAP (H) and 13 DAP (I); most of the seeds aborted by
13 DAP. (J, L) Toluidine blue-stained transverse sections of
seeds from self-pollinated wild-type (J) and MEZ1-silenced
(L) plants; excessive cellularization of endosperm in MEZ1-
silenced seeds. (K, M) Calcofluor-stained transverse sections
of wild-type (K) and MEZ1-silenced (M) seeds visualized un-
der UV; brightly fluorescing calcofluor shows areas with high
cellulose deposition. Three MEZ1-silenced transformant lines
were characterized and all of them showed consistent results.
The images from the line 9 T0 plant are shown. En, endo-
sperm; Em, embryo; Sc, seed coat; Wp, wall parenchyma.
b
427
only those that have excessive DNA correspond-ing to more than one complete haploid genome.As described above, meiotic division in theMEZ1-silenced lines yielded unequal distributionof DNA in the resulting microspores. The mi-crospores that have less than the average amountof DNA content (Figure 5B) presumably abortedsoon after their separation from the tetradsbecause of insufficient genetic information forfurther development. The same could be true formost of those which apparently have the averageDNA content but have an incomplete comple-ment of the haploid genome, provided thatone complete haploid genome is essential for thepollen to fully develop. On the other hand, themicrospores with more than the average DNAcontent have greater probably of having thecomplete complement of the haploid genomealong with some extra chromosomes. Indeed, allT1 plants resulting from self-fertilization ofMEZ1-silenced lines had higher ploidy levels,supporting this speculation.
In MEZ1-silenced lines, the seeds abortedprematurely owing to extensive cellularization ofthe endosperm. Defects in endosperm develop-ment have frequently been cited as the cause offailure of seed development in both interploidycrosses within a species and interspecific crosses(Bushell et al., 2003). It is believed that bothinterploidy and interspecific crosses disrupt thebalance between the maternal and paternalgenomes, which is crucial for the proper develop-ment of endosperm (Johnston et al., 1980; John-ston and Hanneman, 1982). In an interploidy crossbetween a diploid Arabidopsis thaliana(2n=2x=10) as the seed parent and Arabidopsisarenosa, a tetraploid (2n=4x=32), as the pollenparent, seed development ceased when the embryoreached the globular stage (Comai et al., 2000).Incidentally, seed development in MEZ1-silencedlines also ceased when the embryos were at theglobular stage. As mentioned earlier, the microsp-ores that reached maturity in MEZ1-silencedplants likely contained more than the normal
Figure 8. Chromosome count in somatic cells. (A, C) Acetocarmine-stained leaf cells in wild-type (A) and MEZ1-silenced T1
(C, line 9) plants. (B, D) Same as (A) and (C) after marking of individual chromosomes to facilitate counting. Twenty cells were
counted in each T1 plants of the three lines and all of them had increased chromosome numbers.
428

number of chromosomes. Upon self-fertilization,these pollen grains would have caused a geneticimbalance in the endosperm in favor of thepaternal genome. Paternal genome excess haspreviously been reported to correlate with over-growth of the endosperm, resulting in embryonicarrest in Arabidopsis (Scott et al., 1998). Thus, theobserved arrest of embryo development in MEZ1-silenced plants could be a consequence of theaberrant meiosis. All the T1 plants (resulting fromself-pollination) analyzed showed varying degreesof increased ploidy levels in the sporophytic cells.This finding indicates the presence of femalegametophytes having more than the normal num-ber of chromosomes in the MEZ1-silenced lines,and strongly suggests that female meiosis is alsodefective in these plants. Seed development wasalso arrested when wild-type pollen was used topollinate the MEZ1-silenced plants. We did notobtain any seeds from this cross-pollination,presumably because of discordance of chromo-some numbers upon any pairing of male andfemale gametophytes. We did not detect anyexpression of MEZ1 in developing pistils andfemale gametophytes that could account for thefunction in female gametogenesis. This couldsuggest that some MEZ1-derived signals fromthe anther or other part(s) of plants are involved inthe MEZ1 function in female gametogenesis.However, we cannot exclude the possibility oftransient MEZ1 expression in a very narrow timewindow and/or at very low levels that are belowthe detection limits.
MEZ1 is presumably a transcription factor,considering its nuclear localization and the pre-viously reported sequence-specific DNA-bindingactivities of its closely related family members(Takatsuji et al., 1994). Loss of MEZ1 expres-sion affected multiple aspects of meiosis; viz.,chromosome condensation, meiotic synchrony,distribution of chromosomes, and cytokinesis.This might mean that MEZ1 could play a role ata high position in the hierarchy of regulatorygenes that control the processes just after themitosis-to-meiosis switch. Alternatively, it couldbe involved only in the regulation of earlymeiotic events, of which the disturbance affectslater events. Provided that MEZ1 is a transcrip-tion factor, identification of its target genes willbe essential for further understanding of itsfunction.
Acknowledgments
Hiroshi Kouchi and James Doughty are gratefullyacknowledged for their valuable suggestionsregarding in situ hybridization experiments. Weare also grateful to Akiko Baba-Kasai for provid-ing us with the psGFPcs vector, and OsamuUeno, Shoji Sugano, and Kenichi Kubo for valu-able discussions. This work was supported by theCenter of Excellence Special Coordination Fundsfor the Promotion of Science and Technologyfrom the Science and Technology Agency ofJapan, a PROBRAIN grant from the Bio-OrientedTechnology Research Advancement Institutionof Japan and Grant-in-Aid for Scientific Researchfrom the Japan Society for the Promotion ofScience.
References
Brown, R.C. and Lemmon, B.E. 1988. Microtubules associatedwith simultaneous cytokinesis of coenocytic microsporo-cytes. Am. J. Bot. 75: 1848–1856.
Brown, R.C. and Lemmon, B.E. 2001. The cytoskeleton andspatial control of cytokinesis in the plant life cycle. Protopl-asma 215: 35–49.
Bushell, C., Spielman, M. and Scott, R.J. 2003. The basis ofnatural and artificial postzygotic hybridization barriers inArabidopsis species. Plant Cell 15: 1430–1442.
Byzova, M.V., Franken, J., Aarts, M.G., de Almeida-Engler, J.,Engler, G., Mariani, C., Van Lookeren Campagne, M.M.and Angenent, G.C. 1999. Arabidopsis STERILE APET-ALA, a multifunctional gene regulating inflorescence, flower,and ovule development. Genes Dev. 13: 1002–1014.
Canales, C., Bhatt, A.M., Scott, R. and Dickinson, H. 2002.EXS, a putative LRR receptor kinase, regulates malegermline cell number and tapetal identity and promotesseed development in Arabidopsis. Curr. Biol. 12: 1718–1727.
Comai, L., Tyagi, A.P., Winter, K., Holmes-Davis, R.,Reynolds, S.H., Stevens, Y. and Byers, B. 2000. Phenotypicinstability and rapid gene silencing in newly formed arabid-opsis allotetraploids. Plant Cell 12: 1551–1568.
Echlin, P. and Godwin, H. 1968. The ultrastructure andontogeny of pollen in Helleborus foetidus L. II. Pollen graindevelopment through the callose special wall stage. J Cell Sci3: 175–186.
Foiani, M., Capone, R., Nadjar-Boger, E., Sagee, S., Hash-imshoni, T. and Kassir, Y. 1996. A meiosis-specific proteinkinase, Ime2, is required for the correct timing of DNAreplication and for spore formation in yeast meiosis. Mol.Gen. Genet. 293: 278–288.
Golubovskaya, I., Avalkina, N. and Sheridan, W.F. 1997. Newinsights into the role of the maize ameiotic1 locus. Genetics147: 1339–1350.
Hirano, T. 2002. The ABCs of SMC proteins: two-armedATPases for chromosome condensation, cohesion, andrepair. Genes Dev. 16: 399–414.
429

Hirano, T., Kobayashi, R. and Hirano, M. 1997. Condensins,chromosome condensation protein complexes containingXCAP-C, XCAP-E and a Xenopus homolog of the Drosoph-ila Barren protein. Cell 89: 511–521.
Jiang, C.J., Shoji, K., Matsuki, R., Baba, A., Inagaki, N.,Ban, H., Iwasaki, T., Imamoto, N., Yoneda, Y., Deng,X.W. and Yamamoto, N. 2001. Molecular cloning ofa novel importin alpha homologue from rice, by whichconstitutive photomorphogenic 1 (COP1) nuclear localiza-tion signal (NLS)-protein is preferentially nuclearimported. J. Biol. Chem. 276: 9322–9329.
Johnston, S.A., den Nijs, T.P.M., Peloquin, S.J. and Hann-eman, J.R.E. 1980. The significance of genic balance toendosperm development in interspecific crosses. Theor.Appl. Genet. 57: 5–9.
Johnston, S.A. and Hanneman, R.E. 1982. Manipulations ofendosperm balance number overcome crossing barriersbetween diploid Solanum species. Science 217: 446–448.
Jorgensen, R.A., Cluster, P.D., English, J., Que, Q. and Napoli,C.A. 1996. Chalcone synthase cosuppression phenotypes inpetunia flowers: comparison of sense vs. antisense constructsand single-copy vs. complex T-DNA sequences. Plant Mol.Biol. 31: 957–973.
Kapoor, S., Kobayashi, A. and Takatsuji, H. 2002. Silencing ofthe tapetum-specific zinc finger gene TAZ1 causes prematuredegeneration of tapetum and pollen abortion in petunia.Plant Cell 14: 2353–2367.
Kimura, K. and Hirano, T. 2000. Dual roles of the 11Sregulatory subcomplex in condensin functions. Proc. Natl.Acad. Sci. USA 97: 11972–11977.
Kobayashi, A., Sakamoto, A., Kubo, K., Rybka, Z., Kanno, Y.and Takatsuji, H. 1998. Seven zinc-finger transcriptionfactors are expressed sequentially during the developmentof anthers in petunia. Plant J. 13: 571–576.
Kubo, K., Kanno, Y., Nishino, T. and Takatsuji, H. 2000.Zinc-finger genes that specifically express in pistil secretorytissues of petunia. Plant Cell Physiol. 41: 377–382.
Logemann, J., Schell, J. and Willmitzer, L. 1987. Improvedmethod for the isolation of RNA from plant tissues. Anal.Biochem. 163: 16–20.
Mackay, J.P. and Crossley, M. 1998. Zinc fingers are stickingtogether. Trends Biochem. Sci. 23: 1–4.
Mercier, R., Armstrong, S.J., Horlow, C., Jackson, N.P.,Makaroff, C.A., Vezon, D., Pelletier, G., Jones, G.H. andFranklin, F.C. 2003. The meiotic protein SWI1 is requiredfor axial element formation and recombination initiation inArabidopsis. Development 130: 3309–3318.
Mercier, R., Vezon, D., Bullier, E., Motamayor, J.C., Sellier, A.,Lefevre, F., Pelletier, G. and Horlow, C. 2001. SWITCH1(SWI1): a novel protein required for the establishment ofsister chromatid cohesion and for bivalent formation atmeiosis. Genes Dev. 15: 1859–1871.
Mitchell, A.P., Driscoll, S.E. and Smith, H.E. 1990. Positivecontrol of sporulation-specific genes by the IME1 and IME2products in Saccharomyces cerevisiae. Mol. Cell. Biol. 10:2104–2110.
Otegui, M. and Staehelin, L.A. 2000. Cytokinesis in floweringplants: more than one way to divide a cell. Curr. Opin. PlantBiol. 3: 493–502.
Page, S.L. and Hawley, R.S. 2003. Chromosome choreography:the meiotic ballet. Science 301: 785–789.
Ross, K.J., Fransz, P. and Jones, G.H. 1996. A lightmicroscopic atlas of meiosis in Arabidopsis thaliana. Chro-mosome Res. 4: 507–516.
Ruzin, S.E. 1999. Plant Microtechnique and Microscopy.Oxford University Press, New York.
Scott, R.J., Spielman, M., Bailey, J. and Dickinson, H.G. 1998.Parent-of-origin effects on seed development in Arabidopsisthaliana. Development 125: 3329–3341.
Siddiqui, N.U., Stronghill, P.E., Dengler, R.E., Hasenkampf,C.A. and Riggs, C.D. 2003. Mutations in Arabidopsiscondensin genes disrupt embryogenesis, meristem organiza-tion and segregation of homologous chromosomes duringmeiosis. Development 130: 3283–3295.
Stieglitz, H. 1977. Role of beta-1,3-glucanase in postmeioticmicrospore release. Dev. Biol. 57: 87–97.
Takatsuji, H. 1999. Zinc-finger proteins: the classical zinc fingeremerges in contemporary plant science. Plant Mol. Biol. 39:1073–1078.
Takatsuji, H., Nakamura, N. and Katsumoto, Y. 1994. A newfamily of zinc finger proteins in Petunia: structure, DNAsequence recognition, and floral organ-specific expression.Plant Cell 6: 947–958.
van Engelen, F.A., Molthoff, J.W., Conner, A.J., Nap, J.P.,Pereira, A. and Stiekema, W.J. 1995. pBINPLUS: animproved plant transformation vector based on pBIN19.Transgenic Res. 4: 288–290.
Yang, W.C. and Sundaresan, V. 2000. Genetics of gameto-phyte biogenesis in Arabidopsis. Curr. Opin. Plant Biol. 3:53–57.
Yang, S.-L., Xie, L.-F., Mao, H.-Z., Puah, C.S., Yang, W.-C.,Jiang, L., Sundaresan, V. and Ye, D. 2003. TAPETUMDETERMINANT1 is required for cell specialization in theArabidopsis anther. Plant Cell 15: 2792–2804.
Yang, W.C., Ye, D., Xu, J. and Sundaresan, V. 1999. TheSPOROCYTELESS gene of Arabidopsis is required forinitiation of sporogenesis and encodes a novel nuclearprotein. Genes Dev. 13: 2108–2117.
Zhao, D.-Z., Wang, G.-F., Speal, B. and Ma, H. 2002. TheEXCESS MICROSPOROCYTES1 gene encodes a putativeleucine-rich repeat receptor protein kinase that controlssomatic and reproductive cell fates in the Arabidopsis anther.Genes Dev. 16: 2021–2031.
430