
REV7 counteracts DNA double-strand break resection and affects PARP inhibition

Molecular Cell, Vol. 9, 835–846, April, 2002, Copyright 2002 by Cell Press
Wild-Type Levels of Spo11-Induced DSBsAre Required for Normal Single-StrandResection during Meiosis
homolog as repair template (e.g., Hunter and Kleckner,2001; Schwacha and Kleckner, 1997; Thompson andStahl, 1999; Zenvirth et al., 1997; and references within).
The coordinated nature of chromosome pairing, synap-sis, and recombination suggests that these processes are
Matthew J. Neale,1 Madhu Ramachandran,1
Edgar Trelles-Sticken,2,4 Harry Scherthan,2,4
and Alastair S.H. Goldman1,3
1Department of Molecular Biology andBiotechnology
mechanistically linked. This view is supported by a num-The University of Sheffieldber of observations. Mutations in various yeast andSheffield, S10 2TNmouse recombination genes affect chromosome pairingUnited Kingdomand/or synapsis (see Burgess, 2002; Hunter and Kleck-2 Department of Human Biologyner, 2001; Zickler and Kleckner, 1998). NonmutationalUniversity of Kaiserslauternloss of allelic recombination between divergent chromo-D-67663 Kaiserslauternsomes is also sufficient to reduce the effectiveness ofGermanymeiotic chromosome pairing (Goldman and Lichten,2000). On the other hand, genes whose products arestructural components of the SC are required for a nor-Summarymal recombination frequency and normal crossover dis-tribution (see Zickler and Kleckner, 1998). The coordina-We have studied the repair of a DNA-DSB created bytion of recombination with chromosome organizationthe VMA1-derived endonuclease in mutants that haveinvolves checkpoint-type monitoring (Lydall et al., 1996;different levels of Spo11-DSBs: WT (sae2), few (hop1),Roeder and Bailis, 2000). Interruption of DSB repair dur-and none (spo11-Y135F). In spo11-Y135F and hop1 cells,ing meiosis leads to checkpoint-dependent arrest. Es-intrachromosomal repair is more frequent than in WTtablishing arrest requires surveillance of the recombina-
and sae2 cells. In spo11-Y135F cells there was notion process and thus requires Spo11-DSB formation
chromosome pairing or synapsis and a faster turnover(Lydall et al., 1996; McKee and Kleckner, 1997a; Roeder
of resected DNA. Compared to WT and sae2 cells, and Bailis, 2000; and references within).spo11-Y135F and hop1 cells have a greater proportion It is possible that Spo11-DSBs also create a signal thatof long resection tracts. The data suggest that high stimulates interchromosomal repair. Genetic studies oflevels of Spo11-DSBs are required for normal regula- meiotic recombination at HO-induced DSBs show thattion of resection, even at a DSB created by another repair is substantially normal when Spo11-DSBs are ab-protein. WT control over resection could be important sent but that gene conversion rates are reduced andfor directing repair to be interchromosomal, increas- conversion tracts are lengthened (Malkova et al., 1996,ing the chance of creating interhomolog connections 2000). We have tested the idea that Spo11-DSBs stimu-essential to meiotic segregation. late wild-type (WT) interchromosomal DSB repair during
meiosis using genetic and molecular analyses of aIntroduction Spo11-independent DSB induced by the VMA1-derived
endonuclease (VDE, otherwise known as PI-SceI; BremerSpecialized functions are required to segregate homolo- et al., 1992; Gimble and Thorner, 1993). The strains usedgous chromosomes during the meiotic reductional divi- exhibited three different levels of Spo11-DSB formation:sion; these include chromosome pairing, synapsis, and WT, up to 12% of WT, or no breaks. We have found thatrecombination (Zickler and Kleckner, 1998). During pair- repair of the VDE-DSB during meiosis is substantiallying, homologous chromosomes align and move close modified when few or no Spo11-DSBs are formed. Theto each other, eventually permitting synapsis when a modifications include the increased presence of longertripartite proteinaceous structure (the synaptonemal resection tracts and shorter life span of resected inter-complex, SC) forms between them (Zickler and Kleck- mediates. These changes show a positive correlation
with more frequent repair using flanking repeated DNA.ner, 1998). To avoid nondisjunction, the homologs areOverall, the data suggest that Spo11-DSBs have a nega-held together as bivalents until first anaphase (Uhlmann,tive regulatory role during DSB processing, which influ-2001). In most organisms, this is achieved by interchro-ences the outcome of DSB repair.mosomal recombination creating crossovers that link
nonsister chromatids together. The progression of inter-chromosomal recombination from induction to com- Resultspletion is concomitant with chromosome pairing andsynapsis. Meiotic recombination is induced by double- We set out to determine whether Spo11-DSBs are re-strand DNA breaks (DSBs) catalyzed by the Spo11 pro- quired to stimulate normal meiotic DSB repair of atein (Keeney et al., 1997). Genetic control, ensuring re- Spo11-independent DSB induced by VDE. We initiallycombination in meiosis is interchromosomal, is indi- studied repair in a heterozygous strain expressing bothcated by mutants that show reduced bias for using the WT SPO11 and mutant spo11-Y135F-HA3His6 (referred
to as WT when no other mutation is present) and ina homozygous strain expressing only spo11-Y135F-3 Correspondence: [email protected] (referred to as spo11f ). The spo11f allele has4 Present address: MP1 Mol. Genetics, Dept. Ropers, D-14195, Ber-
lin, Germany. lost the active tyrosine that normally catalyzes formation

Molecular Cell836
either ARG4 or arg4-bgl (Figures 1C and 1D). Since theintegration at the ura3::Ty locus creates flanking repeatsof URA3 sequence, the DSB can also be repaired bydeletion of the intervening DNA to yield either ura3::Tyor URA3 (Figures 1E and 1F). We determined, from dis-section of 112 tetrads, that 96% of arg4-vde alleles arecut and repaired during meiosis (see Experimental Pro-cedures). A significant amount of repair leads to deletionusing flanking repeats (56%); this is consistent with pre-vious observations (Ray et al., 1988). Gene conversionto ARG4 took place in 18% of arg4-vde alleles, while21% were converted to arg4-bgl. The remaining 3.8%of arg4-vde alleles were unchanged. Of the tetrads dis-playing a gene conversion event, 71% of sister chroma-tids repaired to different products. Since damage in-curred and repaired before S phase leads to both sisterchromatids having the same modification after replica-tion, the observation that sister chromatids can be re-paired to different products indicates that, like Spo11-DSBs, VDE-DSBs are repaired after S phase.
spo11f Strains Are Less Able to Convertarg4-vde to ARG4The ability of arg4-vde/arg4-bgl diploid strains to pro-
Figure 1. Diagrams of the Parental Reporter Construct and Derived duce Arg� recombinants, characteristic of interchromo-Products somal recombination, was tested in return to growthThe thick black line represents plasmid DNA; the thin black line is (RTG) experiments (see Zenvirth et al., 1997). For thechromosomal DNA. RTG experiments, ndt80 strains that arrest in first pro-(A) Both donor (arg4-bgl) and recipient (arg4-vde) cassettes con-
phase (Xu et al., 1995) were used to avoid potentialtained an arg4 locus inserted into the natural ura3::Ty locus of SK1,problems associated with inviability of spo11f cells duecreating direct repeats of URA3 sequence.to chromosome nondisjunction during the first meiotic(B) The VDE-DSB can form only in the arg4-vde allele.
(C–F) Different products of DSB repair at the recipient locus: (C) division. Use of ndt80 mutants equalized the viability ofgene conversion to ARG4, (D) gene conversion to arg4-bgl, (E) repair spo11f and WT strains during RTG (M.J.N. and A.S.H.G.,using flanking repeated sequences leading to ura3::Ty deletion unpublished data). Our expectation was that if WTproduct, and (F) repair using flanking repeated sequences leading
SPO11 were required to stimulate interhomolog geneto URA3 deletion product.conversion, then the frequency of Arg� colonies wouldbe lower in spo11f cells. There was no difference be-tween the Arg� frequencies derived from the SPO11of meiotic DSBs (Bergerat et al., 1997; Cha et al., 2000).ndt80 and spo11f ndt80 strains before VDE-DSBs wereFor WT, we used a SPO11/spo11f heterozygote strain,induced (Figure 2A). By 8 hr, the frequency of Arg� colo-so that any dominant affects of the spo11-Y135F-nies produced by the SPO11 ndt80 strain was �3-foldHA3His6 allele would be present in both strains. Re-greater than that produced by the spo11f ndt80 strain.cently it has been shown that expression of SPO11 andGene conversion due to Spo11-DSBs in this region isspo11-Y135F-HA3His6 in the same cell leads to WT lev-at least 40-fold lower than that induced by VDE (asels of DSB formation and spore viability (Diaz et al.,measured by random spore analyses using strains not2002). When the HA3His6 tag is associated with WTexpressing VDE; data not shown). Thus, loss of Spo11-SPO11, there is a reduction in DSB frequency, but thereinduced gene conversion in spo11f cells cannot directlyis no effect on either spore viability (Diaz et al., 2002)account for the lower yield of Arg� colonies.or chromosome pairing, as assessed by fluorescence
in situ hybridization (FISH; Peoples et al., 2002). Thus,the presence of the HA3His6 tag is not significant in our The Rate of VDE-DSB Turnover Is Different in WT
and spo11f Strainsexperiments.Southern blot analysis was used to assess the amountof cutting at the parental arg4-vde allele. This was doneDSBs Induced by VDE Lead to High Levels
of Recombination during Meiosis to check that the different frequencies of Arg� recombi-nants in RTG assays did not simply reflect different lev-VDE cuts DNA naturally in a site-specific manner at
approximately the time when Spo11-DSBs form (Bremer els of VDE activity. Quantification of the parental bandrevealed that it is cut by VDE with the same kinetics inet al., 1992; Gimble and Thorner, 1992). We used a re-
porter cassette containing an arg4-vde allele (with the WT and spo11f cells (Figures 2C–2E). In other words,the arg4-vde allele is cut by VDE in a Spo11-independentVDE cutsite) inserted into one natural ura3::Ty locus. An
arg4-bgl allele was inserted at the homologous ura3::Ty manner.Physical analysis was also used to examine the VDE-locus to create an allelic donor for interhomolog repair
of the VDE-induced DSB (VDE-DSB; Figure 1). Allelic DSB molecules. The quantity of DNA visible in a DSBband depends on both the amount of parental DNArepair of the VDE-DSB can result in gene conversion to
Spo11-DSBs Regulate Resection837
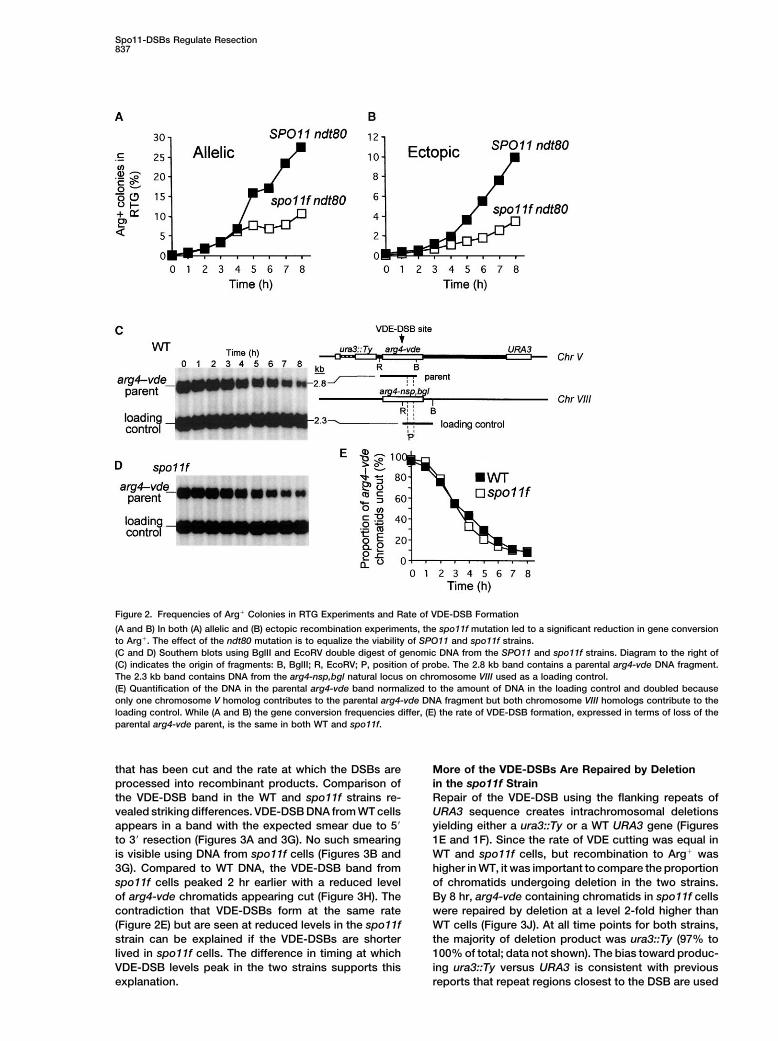
Figure 2. Frequencies of Arg� Colonies in RTG Experiments and Rate of VDE-DSB Formation
(A and B) In both (A) allelic and (B) ectopic recombination experiments, the spo11f mutation led to a significant reduction in gene conversionto Arg�. The effect of the ndt80 mutation is to equalize the viability of SPO11 and spo11f strains.(C and D) Southern blots using BglII and EcoRV double digest of genomic DNA from the SPO11 and spo11f strains. Diagram to the right of(C) indicates the origin of fragments: B, BglII; R, EcoRV; P, position of probe. The 2.8 kb band contains a parental arg4-vde DNA fragment.The 2.3 kb band contains DNA from the arg4-nsp,bgl natural locus on chromosome VIII used as a loading control.(E) Quantification of the DNA in the parental arg4-vde band normalized to the amount of DNA in the loading control and doubled becauseonly one chromosome V homolog contributes to the parental arg4-vde DNA fragment but both chromosome VIII homologs contribute to theloading control. While (A and B) the gene conversion frequencies differ, (E) the rate of VDE-DSB formation, expressed in terms of loss of theparental arg4-vde parent, is the same in both WT and spo11f.
that has been cut and the rate at which the DSBs are More of the VDE-DSBs Are Repaired by Deletionin the spo11f Strainprocessed into recombinant products. Comparison of
the VDE-DSB band in the WT and spo11f strains re- Repair of the VDE-DSB using the flanking repeats ofURA3 sequence creates intrachromosomal deletionsvealed striking differences. VDE-DSB DNA from WT cells
appears in a band with the expected smear due to 5� yielding either a ura3::Ty or a WT URA3 gene (Figures1E and 1F). Since the rate of VDE cutting was equal into 3� resection (Figures 3A and 3G). No such smearing
is visible using DNA from spo11f cells (Figures 3B and WT and spo11f cells, but recombination to Arg� washigher in WT, it was important to compare the proportion3G). Compared to WT DNA, the VDE-DSB band from
spo11f cells peaked 2 hr earlier with a reduced level of chromatids undergoing deletion in the two strains.By 8 hr, arg4-vde containing chromatids in spo11f cellsof arg4-vde chromatids appearing cut (Figure 3H). The
contradiction that VDE-DSBs form at the same rate were repaired by deletion at a level 2-fold higher thanWT cells (Figure 3J). At all time points for both strains,(Figure 2E) but are seen at reduced levels in the spo11f
strain can be explained if the VDE-DSBs are shorter the majority of deletion product was ura3::Ty (97% to100% of total; data not shown). The bias toward produc-lived in spo11f cells. The difference in timing at which
VDE-DSB levels peak in the two strains supports this ing ura3::Ty versus URA3 is consistent with previousreports that repeat regions closest to the DSB are usedexplanation.
Molecular Cell838
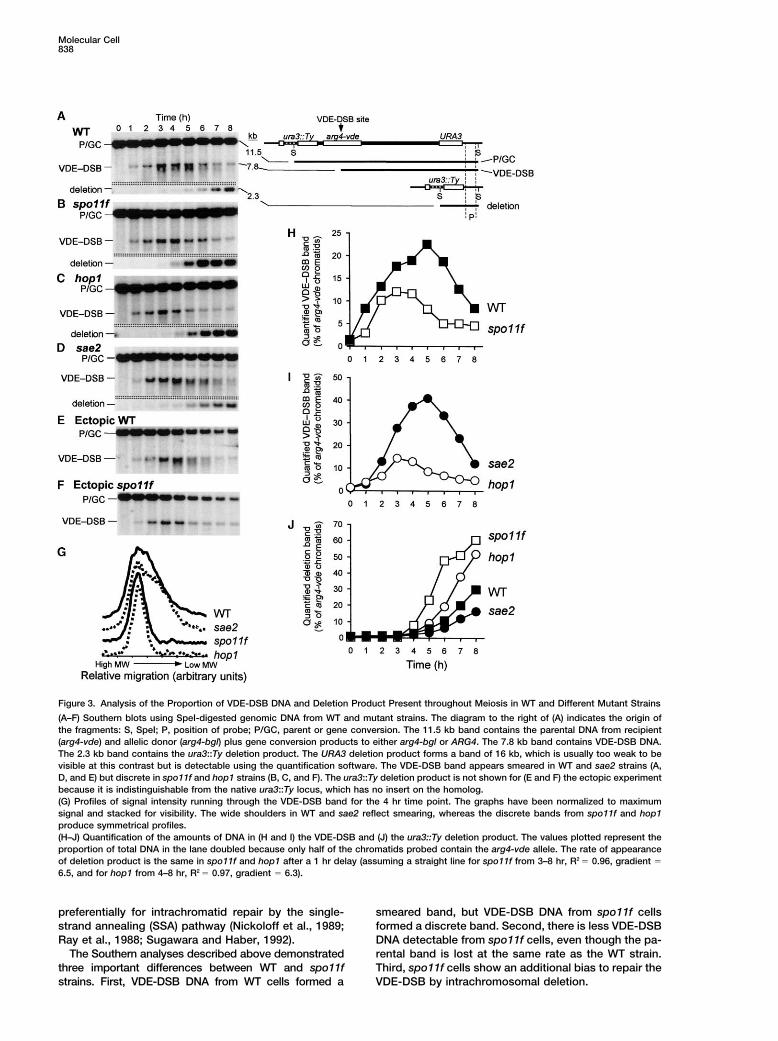
Figure 3. Analysis of the Proportion of VDE-DSB DNA and Deletion Product Present throughout Meiosis in WT and Different Mutant Strains
(A–F) Southern blots using SpeI-digested genomic DNA from WT and mutant strains. The diagram to the right of (A) indicates the origin ofthe fragments: S, SpeI; P, position of probe; P/GC, parent or gene conversion. The 11.5 kb band contains the parental DNA from recipient(arg4-vde) and allelic donor (arg4-bgl) plus gene conversion products to either arg4-bgl or ARG4. The 7.8 kb band contains VDE-DSB DNA.The 2.3 kb band contains the ura3::Ty deletion product. The URA3 deletion product forms a band of 16 kb, which is usually too weak to bevisible at this contrast but is detectable using the quantification software. The VDE-DSB band appears smeared in WT and sae2 strains (A,D, and E) but discrete in spo11f and hop1 strains (B, C, and F). The ura3::Ty deletion product is not shown for (E and F) the ectopic experimentbecause it is indistinguishable from the native ura3::Ty locus, which has no insert on the homolog.(G) Profiles of signal intensity running through the VDE-DSB band for the 4 hr time point. The graphs have been normalized to maximumsignal and stacked for visibility. The wide shoulders in WT and sae2 reflect smearing, whereas the discrete bands from spo11f and hop1produce symmetrical profiles.(H–J) Quantification of the amounts of DNA in (H and I) the VDE-DSB and (J) the ura3::Ty deletion product. The values plotted represent theproportion of total DNA in the lane doubled because only half of the chromatids probed contain the arg4-vde allele. The rate of appearanceof deletion product is the same in spo11f and hop1 after a 1 hr delay (assuming a straight line for spo11f from 3–8 hr, R2 � 0.96, gradient �
6.5, and for hop1 from 4–8 hr, R2 � 0.97, gradient � 6.3).
preferentially for intrachromatid repair by the single- smeared band, but VDE-DSB DNA from spo11f cellsformed a discrete band. Second, there is less VDE-DSBstrand annealing (SSA) pathway (Nickoloff et al., 1989;
Ray et al., 1988; Sugawara and Haber, 1992). DNA detectable from spo11f cells, even though the pa-rental band is lost at the same rate as the WT strain.The Southern analyses described above demonstrated
three important differences between WT and spo11f Third, spo11f cells show an additional bias to repair theVDE-DSB by intrachromosomal deletion.strains. First, VDE-DSB DNA from WT cells formed a
Spo11-DSBs Regulate Resection839
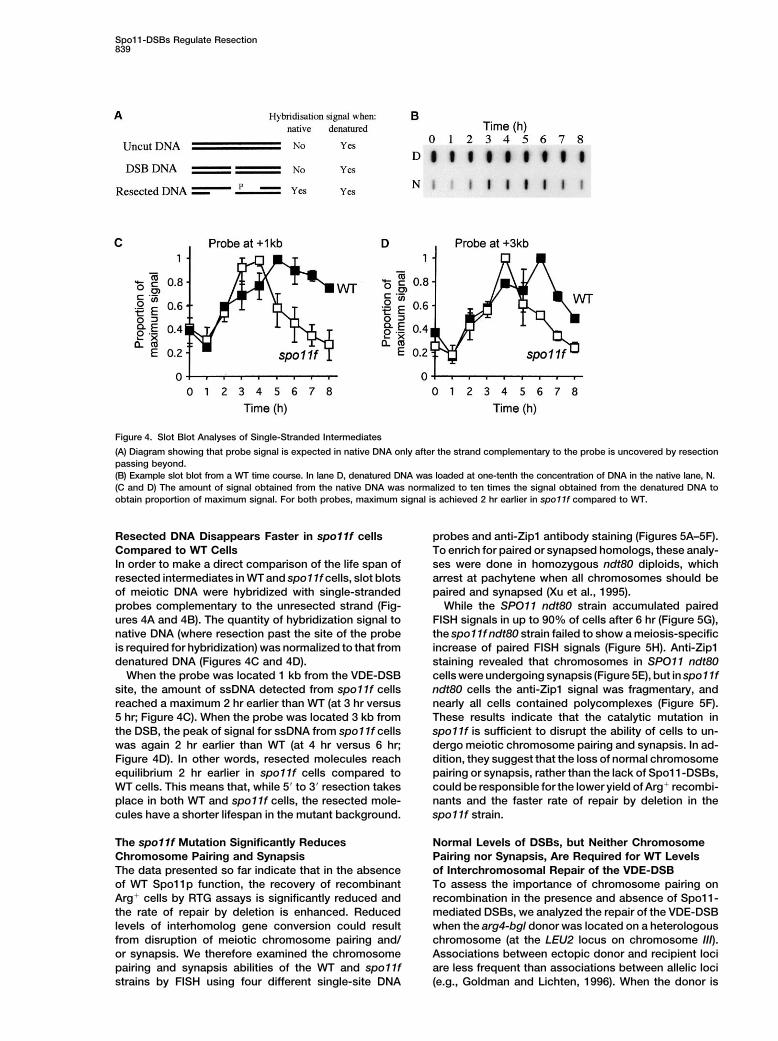
Figure 4. Slot Blot Analyses of Single-Stranded Intermediates
(A) Diagram showing that probe signal is expected in native DNA only after the strand complementary to the probe is uncovered by resectionpassing beyond.(B) Example slot blot from a WT time course. In lane D, denatured DNA was loaded at one-tenth the concentration of DNA in the native lane, N.(C and D) The amount of signal obtained from the native DNA was normalized to ten times the signal obtained from the denatured DNA toobtain proportion of maximum signal. For both probes, maximum signal is achieved 2 hr earlier in spo11f compared to WT.
Resected DNA Disappears Faster in spo11f cells probes and anti-Zip1 antibody staining (Figures 5A–5F).To enrich for paired or synapsed homologs, these analy-Compared to WT Cells
In order to make a direct comparison of the life span of ses were done in homozygous ndt80 diploids, whicharrest at pachytene when all chromosomes should beresected intermediates in WT and spo11f cells, slot blots
of meiotic DNA were hybridized with single-stranded paired and synapsed (Xu et al., 1995).While the SPO11 ndt80 strain accumulated pairedprobes complementary to the unresected strand (Fig-
ures 4A and 4B). The quantity of hybridization signal to FISH signals in up to 90% of cells after 6 hr (Figure 5G),the spo11f ndt80 strain failed to show a meiosis-specificnative DNA (where resection past the site of the probe
is required for hybridization) was normalized to that from increase of paired FISH signals (Figure 5H). Anti-Zip1staining revealed that chromosomes in SPO11 ndt80denatured DNA (Figures 4C and 4D).
When the probe was located 1 kb from the VDE-DSB cells were undergoing synapsis (Figure 5E), but in spo11fndt80 cells the anti-Zip1 signal was fragmentary, andsite, the amount of ssDNA detected from spo11f cells
reached a maximum 2 hr earlier than WT (at 3 hr versus nearly all cells contained polycomplexes (Figure 5F).These results indicate that the catalytic mutation in5 hr; Figure 4C). When the probe was located 3 kb from
the DSB, the peak of signal for ssDNA from spo11f cells spo11f is sufficient to disrupt the ability of cells to un-dergo meiotic chromosome pairing and synapsis. In ad-was again 2 hr earlier than WT (at 4 hr versus 6 hr;
Figure 4D). In other words, resected molecules reach dition, they suggest that the loss of normal chromosomepairing or synapsis, rather than the lack of Spo11-DSBs,equilibrium 2 hr earlier in spo11f cells compared to
WT cells. This means that, while 5� to 3� resection takes could be responsible for the lower yield of Arg� recombi-nants and the faster rate of repair by deletion in theplace in both WT and spo11f cells, the resected mole-
cules have a shorter lifespan in the mutant background. spo11f strain.
Normal Levels of DSBs, but Neither ChromosomeThe spo11f Mutation Significantly ReducesChromosome Pairing and Synapsis Pairing nor Synapsis, Are Required for WT Levels
of Interchromosomal Repair of the VDE-DSBThe data presented so far indicate that in the absenceof WT Spo11p function, the recovery of recombinant To assess the importance of chromosome pairing on
recombination in the presence and absence of Spo11-Arg� cells by RTG assays is significantly reduced andthe rate of repair by deletion is enhanced. Reduced mediated DSBs, we analyzed the repair of the VDE-DSB
when the arg4-bgl donor was located on a heterologouslevels of interhomolog gene conversion could resultfrom disruption of meiotic chromosome pairing and/ chromosome (at the LEU2 locus on chromosome III).
Associations between ectopic donor and recipient locior synapsis. We therefore examined the chromosomepairing and synapsis abilities of the WT and spo11f are less frequent than associations between allelic loci
(e.g., Goldman and Lichten, 1996). When the donor isstrains by FISH using four different single-site DNA
Molecular Cell840
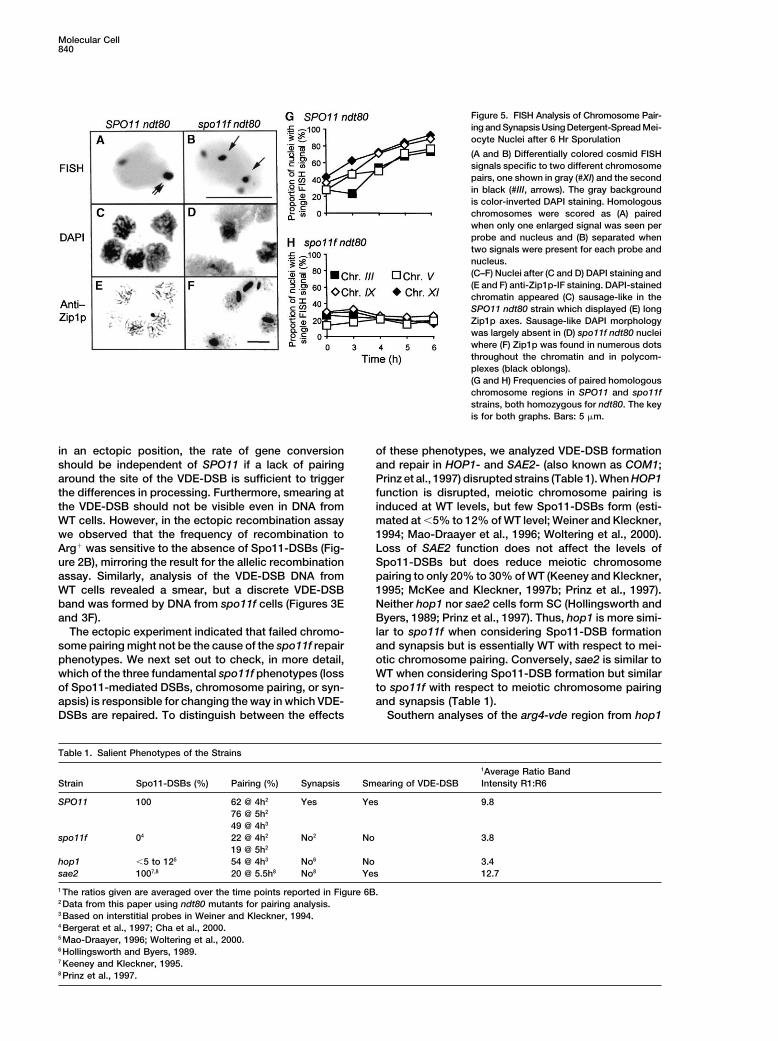
Figure 5. FISH Analysis of Chromosome Pair-ing and Synapsis Using Detergent-Spread Mei-ocyte Nuclei after 6 Hr Sporulation
(A and B) Differentially colored cosmid FISHsignals specific to two different chromosomepairs, one shown in gray (#XI) and the secondin black (#III, arrows). The gray backgroundis color-inverted DAPI staining. Homologouschromosomes were scored as (A) pairedwhen only one enlarged signal was seen perprobe and nucleus and (B) separated whentwo signals were present for each probe andnucleus.(C–F) Nuclei after (C and D) DAPI staining and(E and F) anti-Zip1p-IF staining. DAPI-stainedchromatin appeared (C) sausage-like in theSPO11 ndt80 strain which displayed (E) longZip1p axes. Sausage-like DAPI morphologywas largely absent in (D) spo11f ndt80 nucleiwhere (F) Zip1p was found in numerous dotsthroughout the chromatin and in polycom-plexes (black oblongs).(G and H) Frequencies of paired homologouschromosome regions in SPO11 and spo11fstrains, both homozygous for ndt80. The keyis for both graphs. Bars: 5 �m.
in an ectopic position, the rate of gene conversion of these phenotypes, we analyzed VDE-DSB formationand repair in HOP1- and SAE2- (also known as COM1;should be independent of SPO11 if a lack of pairing
around the site of the VDE-DSB is sufficient to trigger Prinz et al., 1997) disrupted strains (Table 1). When HOP1function is disrupted, meiotic chromosome pairing isthe differences in processing. Furthermore, smearing at
the VDE-DSB should not be visible even in DNA from induced at WT levels, but few Spo11-DSBs form (esti-mated at �5% to 12% of WT level; Weiner and Kleckner,WT cells. However, in the ectopic recombination assay
we observed that the frequency of recombination to 1994; Mao-Draayer et al., 1996; Woltering et al., 2000).Loss of SAE2 function does not affect the levels ofArg� was sensitive to the absence of Spo11-DSBs (Fig-
ure 2B), mirroring the result for the allelic recombination Spo11-DSBs but does reduce meiotic chromosomepairing to only 20% to 30% of WT (Keeney and Kleckner,assay. Similarly, analysis of the VDE-DSB DNA from
WT cells revealed a smear, but a discrete VDE-DSB 1995; McKee and Kleckner, 1997b; Prinz et al., 1997).Neither hop1 nor sae2 cells form SC (Hollingsworth andband was formed by DNA from spo11f cells (Figures 3E
and 3F). Byers, 1989; Prinz et al., 1997). Thus, hop1 is more simi-lar to spo11f when considering Spo11-DSB formationThe ectopic experiment indicated that failed chromo-
some pairing might not be the cause of the spo11f repair and synapsis but is essentially WT with respect to mei-otic chromosome pairing. Conversely, sae2 is similar tophenotypes. We next set out to check, in more detail,
which of the three fundamental spo11f phenotypes (loss WT when considering Spo11-DSB formation but similarto spo11f with respect to meiotic chromosome pairingof Spo11-mediated DSBs, chromosome pairing, or syn-
apsis) is responsible for changing the way in which VDE- and synapsis (Table 1).Southern analyses of the arg4-vde region from hop1DSBs are repaired. To distinguish between the effects
Table 1. Salient Phenotypes of the Strains
1Average Ratio BandStrain Spo11-DSBs (%) Pairing (%) Synapsis Smearing of VDE-DSB Intensity R1:R6
SPO11 100 62 @ 4h2 Yes Yes 9.876 @ 5h2
49 @ 4h3
spo11f 04 22 @ 4h2 No2 No 3.819 @ 5h2
hop1 �5 to 125 54 @ 4h3 No6 No 3.4sae2 1007,8 20 @ 5.5h8 No8 Yes 12.7
1 The ratios given are averaged over the time points reported in Figure 6B.2 Data from this paper using ndt80 mutants for pairing analysis.3 Based on interstitial probes in Weiner and Kleckner, 1994.4 Bergerat et al., 1997; Cha et al., 2000.5 Mao-Draayer, 1996; Woltering et al., 2000.6 Hollingsworth and Byers, 1989.7 Keeney and Kleckner, 1995.8 Prinz et al., 1997.

Spo11-DSBs Regulate Resection841
and sae2 strains undergoing meiosis yielded signifi- hop1), there was a 1.7- to 3.6-fold increase in the propor-tion of resected molecules in both bands R3 and R6,cantly different results (Figures 3C and 3D). The VDE-
DSB DNA from hop1 cells appeared as a discrete band representing significantly longer resection tracts. Theratio of signal in bands R1:R6 is a useful indicator ofand at a level similar to that observed with DNA from
spo11f cells (Figures 3G and 3I). The amount of deletion the relative amounts of shortest:longest resection tractsdetected (Table 1). Overall, there is a clear trend reveal-product generated by hop1 cells was intermediate rela-
tive to WT and spo11f cells (Figure 3J). However, it is ing that in strains deficient for Spo11-DSB formation, agreater proportion of resected molecules have longerimportant to note that from 4 to 8 hr, the kinetics of
deletion product formation in the hop1 strain are the regions of ssDNA.same as that for spo11f. In other words, in hop1 cells theprocessing and repair of the VDE-DSB appears similar to Discussionspo11f but different from WT. In contrast, the VDE-DSBband for sae2 cells appeared smeared, as in WT, and We have examined repair of a DNA DSB induced duringaccumulated to higher levels (Figures 3G and 3I). The meiosis by the naturally occurring protein, VDE. Theamount of deletion product was similar to, but lower reporter cassette consisted of an arg4 allele containingthan, WT (Figure 3J). Thus, in sae2 cells the processing the VDE cutsite flanked by direct repeats of the URA3and repair of the VDE-DSB appears similar to WT but gene. The VDE-DSB can repair either by interchromo-different from spo11f. somal gene conversion or by deletion. The mode of
These data indicate that during repair of VDE-DSBs, repair was compared in WT and spo11f strains. Thecells with few Spo11-DSBs that are able to undergo latter cannot produce Spo11-DSBs, which are normalmeiotic chromosome pairing behave like the spo11f mu- to meiosis. It was found in RTG experiments that com-tant. Conversely, cells with normal levels of Spo11-DSBs mitment to gene conversion was significantly reducedbut without the ability to pair chromosomes behave like in the spo11f strain. This result is similar to earlier geneticWT. From this, we conclude that normal levels of Spo11- studies of HO-induced DSBs formed during meiosis inDSBs, but not normal levels of chromosome pairing or spo11� and rad50� strains, neither of which producesynapsis (since none of the mutants form SC), are re- Spo11-DSBs (Malkova et al., 1996, 2000).quired for WT repair of the VDE-DSB. In our study, the flanking repeated URA3 sequences
provided an alternative method for DSB repair that leadsto deletion of the intervening sequence. Compared toResection at the VDE-DSB Is More ExtensiveWT, there was more repair by deletion in the spo11fin Strains Lacking Spo11-DSBsstrain. During meiosis, repair using repeated regions isAn important difference between strains with Spo11-preferentially intrachromosomal (Ray et al., 1988). OneDSBs (WT and sae2) and the strains with no or fewway to check the likelihood of intrachromosomal recom-Spo11-DSBs (spo11f and hop1) is in the rate of repairbination is to determine the need for the meiosis-specificusing flanking repeats. Almost all repair by deletionRecA homolog, DMC1. Various studies have implicatedmade use of the repeats closest to each other, implyingDMC1 in stimulating interchromosomal repair (Arbel etthat deletion results from SSA (Sugawara and Haber,al., 1999; Schwacha and Kleckner, 1997; Thompson and1992). Since repair by SSA would require longer resec-Stahl, 1999). More recently, Hunter and Kleckner (2001)tion tracts than those needed for gene conversion, weproposed that interchromosomal DSB repair begins withdetermined whether the increased rate of repair by dele-a single end invasion by ssDNA associated with Dmc1p.tion in spo11f and hop1 strains is associated with anFormation of deletion products is independent of DMC1increased proportion of long resection tracts.in spo11f cells (M.J.N. and A.S.H.G., unpublished data)To measure resection, we modified previously usedand therefore unlikely to involve strand invasion. Thismethods (Sun et al., 1991; White and Haber, 1990). Ourobservation supports the idea that the deletion productassay detected the destruction of HaeII restriction endo-arises from an intrachromosomal event.nuclease sites as they became single stranded due to
5� to 3� resection. HaeII cleaves dsDNA at sites flankingthe VDE cutsite and at seven locations within 10 kb Processing of the VDE-DSB Ends
Our assay for detecting ssDNA revealed that a greaterdownstream of the arg4 alleles (Figure 6A). After diges-tion with HaeII, unresected VDE-DSB molecules are 389 proportion of resected molecules in spo11f and hop1
cells have longer single-stranded regions than in WT orbp in length (the distance from the VDE-DSB to theclosest HaeII site). Resection beyond the closest HaeII sae2 cells. Our finding of increased resection at a VDE-
DSB in strains with no or few Spo11-DSBs could explainsite destroys it, leading to a 1.8 kb fragment generatedby cutting at the next HaeII site downstream. Similarly, why Malkova et al. (1996, 2000) saw longer than normal
gene conversion tracts during repair of an HO-DSB infragments of 2.2, 2.7 (a 2.65 and 2.7 kb doublet), 3.3, 3.7,and 8.5 kb were predicted to appear as the population of Spo11-DSB-deficient cells. We found that increased re-
section was associated with elevated repair by deletionVDE-DSBs gained progressively longer ssDNA tails.The data are presented for the three most abundant using flanking direct repeats. This is consistent with the
deletion products resulting from the SSA pathway. Thebands in Figure 6A and are expressed as a proportionof total signal in the bands representing resected mole- longer resection tracts were also associated with faster
turnover of repair intermediates. Thus, when no or fewcules (Figure 6B). In strains with WT levels of Spo11-DSBs (WT and sae2), the majority of molecules were in Spo11-DSBs are present, resection cannot be a rate-
limiting step during repair of the VDE-DSB.band R1, representing a relatively short resection tract.For strains with no or few Spo11-DSBs (spo11f and Regulating the rate and degree of resection could

Molecular Cell842
Figure 6. Assay to Determine the Distance Distribution of Resection Downstream of the VDE-DSB
(A) Sample Southern blot from denaturing gel of WT DNA digested with HaeII. Bands labeled are: V, VDE-DSB; N/S, Nonspecific signal; P,Parent; R1–R6, progressively larger resection bands derived from successive destruction of HaeII sites as the length of ssDNA increases. Inthe diagram, the position of the VDE-DSB site, V, and HaeII sites, H, are indicated.(B) Quantification of the signal in bands R1, R3, and R6 expressed as a proportion of total signal in bands R1–R6. The blocks indicate theaverage data from two independent time courses; the range of data is shown by error bars.
be an important factor in determining the outcome of sites in the region, but the DNA digested is devoid ofendogenous proteins, and therefore this explanationmeiotic DSB repair. Slower resection in WT cells could
bias the chances of interchromosomal repair by reduc- seems unlikely. Digestion of the region in a plasmid,with HaeII, also suggests the enzyme has equal accessing the risk of either uncovering flanking homology or
initiating break-induced replication (Haber, 2000). If ei- to all sites (data not shown). Another possibility is thatthe resection does not proceed at a uniform rate atther of these occur before there has been time for homol-
ogous chromosomes to come into contact, preferred different distances from the VDE-DSB. Previous studieshave not found unequal rates of resection (Sun et al.,interchromosomal connections would not be generated.
Interestingly, exo1 mutants suffer less resection, and 1991; White and Haber, 1990), but our analysis is ofhigher resolution, measuring more bands and a longerthe outcome of recombination is changed to bias gene
conversion over reciprocal exchange (Khazanehdari and distance.Borts, 2000; Kirkpatrick et al., 2001; Tsubouchi andOgawa, 2000). This change may not reflect the sug- WT SPO11 Is Required for Normal Levels of Meiotic
Chromosome Pairing and Synapsisgested role of Exo1p in forming crossovers but, instead,a more general phenomenon whereby the length of re- Chromosome pairing errors could make the homolog
less available than normal to act as a donor during DSBsection tract influences the outcome of DSB repair. Thetiming of movement from repair intermediates to prod- repair and lead to an increase in intrachromosomal re-
pair. In S. cerevisiae, premeiotic homologous chromo-ucts may also be an important issue. Disruption of DNAdamage checkpoint genes during meiosis increases the some associations are lost during S phase to be reestab-
lished during first prophase, which culminates inchances of ectopic repair when there is a choice of twodonor sequences (one allelic, one ectopic) on the same synapsis (Weiner and Kleckner, 1994). Many mutations
inhibit the ability of meiotic cells to bring homologoushomolog (Grushcow et al., 1999). This form of changein repair partner of choice could be due to a more rapid chromosomes together during prophase I (see Zickler
and Kleckner, 1998). Among these are mutations intransition to product before there has been time to en-sure optimal choice of repair partner. genes that abolish or reduce the frequency of native
Spo11-DSBs. It is well established that meiotic chromo-In the assay to reveal loss of HaeII restriction endonu-clease sites due to resection, the relative abundance of some pairing and synapsis fail in spo11� S. cerevisiae
cells (Loidl et al., 1994; Weiner and Kleckner, 1994).bands varied with molecular weight in a nonuniformmanner. One predicted band (R5; Figure 6A) was not However, there has been some debate over whether a
function of Spo11, other than creating DSBs, is requireddetectable, although resection clearly passed beyondthe relevant restriction site causing formation of a larger for normal chromosome pairing.
In a study by Cha et al. (2000) using the same spo11fband. This could be explained if the HaeII restrictionenzyme does not have equal access to all available point mutation, there was significant recovery of paired

Spo11-DSBs Regulate Resection843
signals following the normal loss of premeiotic pairing. Spo11-DSBs in sae2 cells is associated with greaterthan WT accumulation of VDE-DSBs and lower levelsHowever, we could not detect any induction of pairing
or synapsis during meiosis in our spo11f ndt80 strain. of repair by deletion. These findings could result fromthe sae2 mutation reducing the relative abundance ofA genetic assay for chromosome juxtaposition using the
same mutation agrees with this result (Peoples et al., resection proteins per VDE-DSB by causing resectionmachinery to be tied to the unprocessed Spo11-DSBs.2002). It has recently been shown that disruption of
the SPO11 homolog, rec12�, also causes a defect in Evidence for this possibility comes from the observationthat rad50S cells (which have also blocked Spo11-DSBchromosome pairing (Nabeshima et al., 2001). The pair-
ing defect reported for rec12� S. pombe is not as severe ends) contain foci of proteins required for resection(Mre11, Rad50, and Xrs2), implying that the resectionas we report for spo11f S. cerevisiae, possibly because
the characteristic nuclear movements which aid chro- machinery may not be freely diffusible when Spo11-DSBs are present (Usui et al., 1998).mosome pairing (horse-tailing) are still present in rec12�
cells (Nabeshima et al., 2001). We conclude that fully The data are also consistent with a model in which therate of resection is regulated by protein modifications. Itfunctional SPO11 is required to induce normal mei-
otic chromosome pairing in yeast. In other organisms, has recently been demonstrated that blocking pro-cessing of DNA damaged by MMS in mitosis activatesSpo11-DSBs are not required to induce meiotic chromo-
some pairing or synapsis (Dernburg et al., 1998; McKim Tel1, leading to phosphorylation of Mre11 and Xrs2 (Usuiet al., 2001). Since Spo11 covalently binds to DNA as itand Hayashi-Hagihara, 1998). However, other organ-
isms may facilitate chromosome pairing and synapsis makes DSBs, it is possible that Spo11-DSBs representsimilarly blocked damage to the cell, activating the Tel1through more robust premeiotic homolog associations
or specialized pairing centers (discussed extensively in pathway and meiotic regulation of the Mre11 complex.By this model, strains that are unable to remove Spo11Dernburg et al., 1998).from the DNA ends, such as sae2 mutants, might be-come hyperregulated. This could lead to the slower thanPossible Causes for Modified End ProcessingWT turnover of VDE-DSBs and the reduced intrachromo-The ectopic recombination experiment indicated thatsomal repair observed in sae2 cells.the lack of chromosome pairing and synapsis around
The evidence that DNA damage checkpoint proteinsthe VDE-DSB is not sufficient to bring about the reducedalter repair partner of choice indicates another possiblefrequency of Arg� or the discrete VDE-DSB band. Weroute to regulation of resection (Grushcow et al., 1999).made use of sae2 and hop1 mutant strains to separateNormally during meiosis the presence of Spo11-DSBsthe phenotype of failure to form Spo11-DSBs from theleads to checkpoint dependent activation of Mek1,inability to induce meiotic chromosome pairing (see Ta-which is required to regulate exit from pachyteneble 1 for salient phenotypes). The hop1 strain behaved(Roeder and Bailis, 2000). Mek1 and other proteins invery much like spo11f, whereas sae2 cells behaved morethe pachytene checkpoint pathway could be involvedlike WT. These correlations strongly support the ideain regulating components of the resection complex.that the modified VDE-DSB repair, seen in spo11f cells,
is due to the failure of Spo11-DSB formation (as seenExperimental Proceduresin hop1 cells) rather than the lack of chromosome pairing
(as seen in the ectopic recombination experiment and Strains and Plasmidssae2 cells). Diploid yeast strains of the SK1 background used in this study
are listed in the supplemental data at http://www.molecule.org/cgi/The presence of multiple (around 200) DSBs, rathercontent/full/9/4/835/DC1. A list of haploid strains used to constructthan Spo11-DSBs per se, could reduce the rate andthe appropriate diploids is available on request. Yeast strains weredegree of resection and encourage interchromosomalmaintained using standard laboratory techniques. All incubationsrepair. The viability of spores and recombination levelswere at 30�C, and liquid cultures were grown with vigorous agitation.
are improved in spo11� cells by lesions resulting from Strains were made to express the VMA1-derived endonucleaseX-irradiation (Thorne and Byers, 1993). This does not (VDE, also known as PI-SceI) by sporulation of strains transformed
with plasmid pAG70 that expresses TFP1/VMA1::VDE (YEpVMA1;conclusively show that exogenous DSBs induce WT endprovided by R. Borts). Meiosis-specific VDE cleavage causes geneprocessing because the modified end processing seenconversion of its reading frame into an empty TFP1/VMA1 genein spo11f was also associated with some interchromo-(Bremer et al., 1992). Spores were dissected, cured of the plasmid,somal repair.and checked for insertion of the VDE coding sequence at TFP1/
One factor that might influence the rate and degree of VMA1 by PCR across the insertion site.resection at a DSB is the relative abundance of resection The arg4-vde recipient cassette was created in plasmid pAG404
based on pMJ113 (Wu and Lichten, 1995), but the ARG4 regionmachinery compared to double-strand ends. Sincelacks the NspI mutation and includes a 73 bp insertion of VDEthere are approximately 200 less DSBs per meiosis incleavage sequence at EcoRV. The arg4-vde construct was insertedspo11f compared to WT (Bishop, 1994), the increasedat the ura3::Ty locus by transformation of haploid strains with linear-
abundance of resection proteins per VDE-DSB might ized pAG404.cause hyperresection. On the other hand, any mutation The ndt80::ADE2 mutation contains a 2.2 kb insertion of pMJ412that decreases the abundance of resection proteins per (provided by M. Lichten) at the BamHI site located 1224 bp inside the
NDT80 open reading frame. Genomic disruptions were confirmedVDE-DSB could reduce the amount of resection. In thisby PCR and Southern blot analysis. An SK1 strain containing therespect, the sae2 experiments are worth considering.nuc1::LEU2 disruption and spo11-Y135F-HA3His6 allele was obtainedIn sae2 cells the Spo11-DSBs are not resected becausefrom S. Keeney. An SK1 strain containing hop1::LEU2 was obtained
the Spo11p remains covalently bound, protecting the from N. Hollingsworth. An SK1 strain containing sae2::KanMX wasbroken ends (Keeney et al., 1997; Keeney and Kleckner, obtained from M. Lichten. All mutations were subsequently crossed
into relevant strains using standard techniques.1995; Liu et al., 1995). We found that persistence of

Molecular Cell844
Sporulation Procedures to Allers and Lichten (2001); hexamine cobalt (III) chloride was ex-cluded from solutions. Restriction endonuclease digested DNA wasFor DNA and genetic analyses, single diploid colonies were inocu-
lated into 5 ml liquid YEPD and grown overnight. Liquid cultures separated under: native conditions (0.5% 1TAE agarose gels for12–14 hr at 70V) or denaturing conditions (0.7% agarose gels in 50were diluted into multiple flasks of PSP2 (0.67% yeast nitrogen
base/0.1% yeast extract/1% KAc/50mM K-biphthalate [pH 5.0]) pre- mM NaOH and 1 mM EDTA for 20 hr at 46V at 5�C). Separated DNAwas blotted to Zetaprobe membrane (Bio-Rad) under denaturingsporulation medium and grown for 24 hr. 2PSP2 cultures with an
OD600 of 1.5 were rapidly harvested for synchronous sporulation. conditions with a Vacugene-XL system (PharmaciaBiotech). Theprobe used to display EcoRV and BglII double-digested DNA afterCells were resuspended in two-thirds of the presporulation volume
of SPM (1% KAc) which was prewarmed to 30�C and supplemented native separation is specific to ARG4 (chromosome VIII coordinates140767 to 141130). The probe used to display SpeI-digested DNAwith adenine (10 mg/l), arginine (10 mg/l), and PPG1000 (0.001%).
The cells were incubated with maximum aeration at 30�C. For cytol- after native separation is specific to a region downstream of URA3(chromosome V coordinates 117126 to 117992). A single-strandedogy, cultures were grown in presporulation medium to a density of
2 107 cells/ml and then transferred to sporulation medium (2% probe specific to the above ARG4 region and complementary to theunresected strand was made by multiple round primer extensionKAc) at a density of 4 107 cells/ml. Synchrony was monitored by
DAPI staining. (Liu et al., 1995) and used to display resection products followingdenaturing electrophoresis of HaeII-digested DNA. The slot blotassay was based on a method described previously (Sugawara andCytology
Aliquots were immediately transferred to tubes on ice containing Haber, 1992). In brief, 1 �g of native genomic DNA dissolved in 10
SSC (1.5 M sodium chloride and 0.15 M trisodium citrate) was blottedone-tenth volume of acid-free 37% formaldehyde (Merck). After 0.5hr, cells were washed with 1 SSC and spheroplasted with Zymoly- on to Zetaprobe membrane (Bio-Rad) using slot blot manifold under
vacuum pressure. Heat-denatured DNA was used at one-tenth thease 100 T (100 �g/ml) in 0.8 M sorbitol, 2% KAc, and 10mM DTT.After adding 10 volumes of ice-cold 1 M sorbitol, spread prepara- native DNA concentration. DNA was UV crosslinked to the mem-
brane and hybridized using single-stranded probes (made as above)tions were obtained as described (Scherthan and Trelles-Sticken,2002). specific to the unresected strand, 1018 bp and 3044 bp downstream
of the VDE-DSB site. The amount of probe hybridizing to each bandCosmid DNA probes were used to FISH-tag regions on #III (ATCCnumber 70884), #IX (ATCC number 70895), and the middle of the was quantified using scanning densitometry after exposure to Ko-
dak phosphor screens and using a Personal-FX phosphorimagerlong arm of #XI (pUKG151; Thierry et al., 1995; Trelles-Sticken etal., 2000). The middle region of #V was probed with a composite with QuantityOne software (Bio-Rad).
All genetic and physical data presented from time courses areplasmid probe (Scherthan et al., 1992). Probes were labeled eitherwith digoxigenin-11-dUTP (Roche Biochem) or with biotin-14-dCTP averaged for two independent experiments; there was no significant
variation unless indicated.(Invitrogen) using a nick translation (Invitrogen). All preparationswere subjected to two-color FISH as described previously (Scher-than et al., 1992; Trelles-Sticken et al., 2000). Acknowledgments
Signal patterns in spread nuclei were investigated using a ZeissAxioskop epifluorescence microscope equipped with a double- We would like to thank S. Burgess, N. Hollingsworth, C. Price, andband-pass filter for simultaneous observation of red and green fluo- A. Villeneuve for discussions and comments on an early version ofrescence, or single-band-pass filters for excitation of red, green, and the manuscript. We are also grateful to T. Allers, V. Borde, R. Borts,blue (Chroma Technologies). Digital images were obtained using a L. Gilbertson, J. Haber, S. Keeney, M. Lichten, A. Malkova, and P.cooled gray-scale CCD camera (Hamamatsu) controlled by the ISIS Sudbery for discussions and/or technical advice. Yeast strains and/fluorescence image analysis system (MetaSystems). For all time or plasmids were generously provided by R. Borts, N. Hollingsworth,points and probe combinations, more than 100 nuclei of randomized S. Keeney, and M. Lichten. This work was supported by BBSRCpreparations were examined directly in the microscope for single grant 50/G12839 to A.S.H.G.; H.S. was supported by the DFG (SCHEor double FISH signals of the various probes. Only nuclei containing 350/8-3).signals of both probes were scored.
Received: August 13, 2001Genetic Assay of Recombination in SPO11 Strain by Dissection Revised: March 6, 2002Spores of dAG206 were dissected to determine the repair patternof the VDE-DSB. Spore colonies were replica plated to synthetic Referencesmedium lacking arginine (SC-Arg) or uracil (SC-Ura) to check to seeif they were: (1) arginine and uracil prototrophs (meaning there had Allers, T., and Lichten, M. (2001). Differential timing and control ofbeen a gene conversion event of arg4-vde to ARG4), (2) arginine noncrossover and crossover recombination during meiosis. Celland uracil auxotrophs (meaning the spore contained the prominent 106, 47–57.class of deletion product to ura3::Ty), and (3) arginine auxotrophs
Arbel, A., Zenvirth, D., and Simchen, G. (1999). Sister chromatid-but uracil prototrophs (meaning the spore contained either parentalbased DNA repair is mediated by RAD54, not by DMC1 or TID1.arg4-bgl, gene conversion of arg4-vde to arg4-bgl, unchangedEMBO J. 18, 2648–2658.arg4-vde, or rare deletion to URA3). The spore colonies from class 3Bergerat, A., de Massy, B., Gadelle, D., Varoutas, P.C., Nicolas, A.,were further tested by mating in patches with arg4-nsp and arg4-bgland Forterre, P. (1997). An atypical topoisomerase II from Archaeatester strains on YEPD. Matings were replica plated to SC-Arg me-with implications for meiotic recombination. Nature 386, 414–417.dium and exposed to a pulse of UV light to stimulate recombination.
Recombination between arg4-bgl and arg4-nsp alleles yielded Arg� Bishop, D.K. (1994). RecA homologs Dmc1 and Rad51 interact topapillae. No such papillae could arise from an arg4-bgl and arg4-bgl form multiple nuclear complexes prior to meiotic chromosome syn-mating. Spore colonies with uncut arg4-vde were able to produce apsis. Cell 79, 1081–1092.Arg� papillae when mated with both tester strains. Spore colonies Bremer, M.C., Gimble, F.S., Thorner, J., and Smith, C.L. (1992). VDEcontaining a URA3 deletion product were unable to form papillae endonuclease cleaves Saccharomyces cerevisiae genomic DNA atwith either tester strain. a single site: physical mapping of the VMA1 gene. Nucleic Acids
Res. 20, 5484.Return to Growth
Burgess, S.M. (2002). Homologous chromosome associations andAliquots of synchronously sporulating cells were serial diluted innuclear order in meiotic and mitotically dividing cells of buddingwater and plated onto YEPD and SC-Arg medium to assess theyeast. Adv. Genet. 46, 49–90.proportion of Arg� prototrophs.Cha, R.S., Weiner, B.M., Keeney, S., Dekker, J., and Kleckner, N.(2000). Progression of meiotic DNA replication is modulated by inter-DNA Isolation and Southern and Slot Blot Analyseschromosomal interaction proteins, negatively by Spo11p and posi-Twenty-five milliliter samples of sporulating culture were removed
at intervals and processed for storage and DNA isolation according tively by Rec8p. Genes Dev. 14, 493–503.

Spo11-DSBs Regulate Resection845
Dernburg, A.F., McDonald, K., Moulder, G., Barstead, R., Dresser, fying recessive diploid-specific mutations in Saccharomyces cere-visiae, its application to the isolation of mutants blocked at interme-M., and Villeneuve, A.M. (1998). Meiotic recombination in C. elegans
initiates by a conserved mechanism and is dispensable for homolo- diate stages of meiotic prophase and characterization of a new geneSAE2. Genetics 146, 797–816.gous chromosome synapsis. Cell 94, 387–398.
Diaz, R.L., Alcid, A.D., Berger, J.M., and Keeney, S. (2002). Identifica- McKim, K.S., and Hayashi-Hagihara, A. (1998). mei-W68 in Drosoph-tion of residues in yeast spo11p critical for meiotic DNA double- ila melanogaster encodes a spo11 homolog: evidence that thestrand break formation. Mol. Cell. Biol. 22, 1106–1115. mechanism for initiating meiotic recombination is conserved. Genes
Dev. 12, 2932–2942.Gimble, F.S., and Thorner, J. (1992). Homing of a DNA endonucleasegene by meiotic gene conversion in Saccharomyces cerevisiae. Na- Nabeshima, K., Kakihara, Y., Hiraoka, Y., and Nojima, H. (2001). Ature 357, 301–306. novel meiosis-specific protein of fission yeast, Meu13p, promotes
homologous pairing independently of homologous recombination.Gimble, F.S., and Thorner, J. (1993). Purification and characteriza-EMBO J. 20, 3871–3881.tion of VDE, a site-specific endonuclease from the yeast Saccharo-
myces cerevisiae. J. Biol. Chem. 268, 21844–21853. Nickoloff, J.A., Singer, J.D., Hoekstra, M.F., and Heffron, F. (1989).Double-strand breaks stimulate alternative mechanisms of recombi-Goldman, A.S.H., and Lichten, M. (1996). The efficiency of meioticnation repair. J. Mol. Biol. 207, 527–541.recombination between dispersed sequences in Saccharomyces
cerevisiae depends upon their chromosomal location. Genetics 144, Peoples, T.L., Dean, E.W., Gonzalez, O., Lambourne, L., and Bur-43–55. gess, S.M. (2002). Close, stable homolog juxtaposition during meio-
sis in budding yeast is dependent on meiotic recombination, occursGoldman, A.S.H., and Lichten, M. (2000). Restriction of ectopic re-combination by interhomolog interactions during Saccharomyces independent of synapsis and is distinct from DSB-independent pair-
ing contacts. Genes Dev., in press.cerevisiae meiosis. Proc. Natl. Acad. Sci. USA 97, 9537–9542.
Grushcow, J.M., Holzen, T.M., Park, K.J., Weinert, T., Lichten, M., Prinz, S., Amon, A., and Klein, F. (1997). Isolation of COM1, a newand Bishop, D.K. (1999). Saccharomyces cerevisiae checkpoint gene required to complete meiotic double-strand break-inducedgenes MEC1, RAD17, and RAD24 are required for normal meiotic recombination in Saccharomyces cerevisiae. Genetics 146, 781–795.recombination partner choice. Genetics 153, 607–620. Ray, A., Siddiqi, I., Kolodkin, A.L., and Stahl, F.W. (1988). Intra-Haber, J.E. (2000). Partners and pathways repairing a double-strand chromosomal gene conversion induced by a DNA double-strandbreak. Trends Genet. 16, 259–264. break in Saccharomyces cerevisiae. J. Mol. Biol. 201, 247–260.Hollingsworth, N.M., and Byers, B. (1989). HOP1: a yeast meiotic Roeder, G.S., and Bailis, J.M. (2000). The pachytene checkpoint.pairing gene. Genetics 121, 445–462. Trends Genet. 16, 395–403.Hunter, N., and Kleckner, N. (2001). The single-end invasion. An Scherthan, H., and Trelles-Sticken, E. (2002). Yeast FISH: delineationasymmetric intermediate at the double-strand break to double-Holli- of chromosomal targets in vegetative and meiotic yeast cells. Inday junction transition of meiotic recombination. Cell 106, 59–70. Springer Lab Manual on FISH Technology, B. Rautenstrauss and T.
Liehr, eds. (New York: Springer), pp. 329–345.Keeney, S., and Kleckner, N. (1995). Covalent protein-DNA com-plexes at the 5� strand termini of meiosis-specific double-strand Scherthan, H., Loidl, J., Schuster, T., and Schweizer, D. (1992). Mei-breaks in yeast. Proc. Natl. Acad. Sci. USA 92, 11274–11278. otic chromosome condensation and pairing in Saccharomyces cere-Keeney, S., Giroux, C.N., and Kleckner, N. (1997). Meiosis-specific visiae studied by chromosome painting. Chromosoma 101, 590–595.DNA double-strand breaks are catalyzed by Spo11, a member of a Schwacha, A., and Kleckner, N. (1997). Interhomolog bias duringwidely conserved protein family. Cell 88, 375–384. meiotic recombination: meiotic functions promote a highly differen-Khazanehdari, K.A., and Borts, R.H. (2000). EXO1 and MSH4 differ- tiated interhomolog-only pathway. Cell 90, 1123–1135.entially affect crossing-over and segregation. Chromosoma 109, Sugawara, N., and Haber, J.E. (1992). Characterization of double-94–102. strand break-induced recombination: homology requirements andKirkpatrick, D.T., Ferguson, J.R., Petes, T.D., and Symington, L.S. single-stranded DNA formation. Mol. Cell. Biol. 12, 563–575.(2001). Decreased meiotic intergenic recombination and increased
Sun, H., Treco, D., and Szostak, J.W. (1991). Extensive 3�-overhang-meiosis I nondisjunction in exo1 mutants of Saccharomyces cerevis-
ing, single-stranded DNA associated with the meiosis-specific dou-iae. Genetics 156, 1549–1557.
ble-strand breaks at the ARG4 recombination initiation site. Cell 64,Liu, J.H., Wu, T.C., and Lichten, M. (1995). The location and structure 1155–1161.of double-strand DNA breaks induced during yeast meiosis: evi-
Thierry, A., Gaillon, L., Galibert, F., and Dujon, B. (1995). Constructiondence for a covalently-linked DNA-protein intermediate. EMBO J.
of a complete genomic library of Saccharomyces cerevisiae and14, 4599–4608.
physical mapping of chromosome XI at 3.7 kb resolution. Yeast 11,Loidl, J., Klein, F., and Scherthan, H. (1994). Homologous pairing is 121–135.reduced but not abolished in asynaptic mutants of yeast. J. Cell
Thompson, D.A., and Stahl, F.W. (1999). Genetic control of recombi-Biol. 125, 1191–1200.nation partner preference in yeast meiosis. Isolation and character-
Lydall, D., Nikolsky, Y., Bishop, D.K., and Weinert, T. (1996). A mei- ization of mutants elevated for meiotic unequal sister-chromatidotic recombination checkpoint controlled by mitotic checkpoint recombination. Genetics 153, 621–641.genes. Nature 383, 840–843.
Thorne, L.W., and Byers, B. (1993). Stage-specific effects of X-irradi-Malkova, A., Ross, L., Dawson, D., Hoekstra, M.F., and Haber, J.E. ation on yeast meiosis. Genetics 134, 29–42.(1996). Meiotic recombination initiated by a double-strand break in
Trelles-Sticken, E., Dresser, M.E., and Scherthan, H. (2000). Meioticrad50� yeast cells otherwise unable to initiate meiotic recombina-telomere protein Ndj1p is required for meiosis-specific telomeretion. Genetics 143, 741–754.distribution, bouquet formation and efficient homologue pairing. J.
Malkova, A., Klein, F., Leung, W.Y., and Haber, J.E. (2000). HO endo-Cell Biol. 151, 95–106.
nuclease-induced recombination in yeast meiosis resemblesTsubouchi, H., and Ogawa, H. (2000). Exo1 roles for repair of DNASpo11-induced events. Proc. Natl. Acad. Sci. USA 97, 14500–14505.double-strand breaks and meiotic crossing over in SaccharomycesMao-Draayer, Y., Galbraith, A.M., Pittman, D.L., Cool, M., and Ma-cerevisiae. Mol. Biol. Cell 11, 2221–2233.lone, R.E. (1996). Analysis of meiotic recombination pathways in theUhlmann, F. (2001). Chromosome cohesion and segregation in mito-yeast Saccharomyces cerevisiae. Genetics 144, 71–86.sis and meiosis. Curr. Opin. Cell Biol. 13, 754–761.McKee, A.H., and Kleckner, N. (1997a). Mutations in SaccharomycesUsui, T., Ohta, T., Oshiumi, H., Tomizawa, J., Ogawa, H., and Ogawa,cerevisiae that block meiotic prophase chromosome metabolismT. (1998). Complex formation and functional versatility of Mre11 ofand confer cell cycle arrest at pachytene identify two new meiosis-budding yeast in recombination. Cell 95, 705–716.specific genes SAE1 and SAE3. Genetics 146, 817–834.
McKee, A.H., and Kleckner, N. (1997b). A general method for identi- Usui, T., Ogawa, H., and Petrini, J.H. (2001). A DNA damage response

Molecular Cell846
pathway controlled by Tel1 and the Mre11 complex. Mol. Cell 7,1255–1266.
Weiner, B.M., and Kleckner, N. (1994). Chromosome pairing via mul-tiple interstitial interactions before and during meiosis in yeast. Cell77, 977–991.
White, C.I., and Haber, J.E. (1990). Intermediates of recombinationduring mating type switching in Saccharomyces cerevisiae. EMBOJ. 9, 663–673.
Woltering, D., Baumgartner, B., Bagchi, S., Larkin, B., Loidl, J., delos Santos, T., and Hollingsworth, N.M. (2000). Meiotic segregation,synapsis, and recombination checkpoint functions require physicalinteraction between the chromosomal proteins Red1p and Hop1p.Mol. Cell. Biol. 20, 6646–6658.
Wu, T.C., and Lichten, M. (1995). Factors that affect the locationand frequency of meiosis-induced double-strand breaks in Sacchar-omyces cerevisiae. Genetics 140, 55–66.
Xu, L., Ajimura, M., Padmore, R., Klein, C., and Kleckner, N. (1995).NDT80, a meiosis-specific gene required for exit from pachytene inSaccharomyces cerevisiae. Mol. Cell. Biol. 15, 6572–6581.
Zenvirth, D., Loidl, J., Klein, S., Arbel, A., Shemesh, R., and Simchen,G. (1997). Switching yeast from meiosis to mitosis: double-strandbreak repair, recombination and synaptonemal complex. GenesCells 2, 487–498.
Zickler, D., and Kleckner, N. (1998). The leptotene-zygotene transi-tion of meiosis. Annu. Rev. Genet. 32, 619–697.
Copyright © 2022 FDOKUMEN




















