
Signal Propagation in Oblique Dendrites of CA1
12
doi:10.1152/jn.00521.2005 94:4145-4155, 2005. J Neurophysiol Michele Migliore, Michele Ferrante and Giorgio A. Ascoli Pyramidal Cells Signal Propagation in Oblique Dendrites of CA1 You might find this additional info useful... 59 articles, 26 of which can be accessed free at: This article cites /content/94/6/4145.full.html#ref-list-1 9 other HighWire hosted articles, the first 5 are: This article has been cited by [PDF] [Full Text] [Abstract] , December , 2009; 102 (6): 3234-3250. J Neurophysiol Vladislav Volman, Herbert Levine, Eshel Ben-Jacob and Terrence J. Sejnowski Dendritic Conductances Locally Balanced Dendritic Integration by Short-Term Synaptic Plasticity and Active [PDF] [Full Text] [Abstract] , May 5, 2010; 30 (18): 6434-6442. J. Neurosci. Giorgio A. Ascoli, Sonia Gasparini, Virginia Medinilla and Michele Migliore Local Control of Postinhibitory Rebound Spiking in CA1 Pyramidal Neuron Dendrites [PDF] [Full Text] [Abstract] , July , 2011; 106 (1): 44-58. J Neurophysiol Marie L. Goeritz, Qing Ouyang and Ronald M. Harris-Warrick channels in a small neural network h I Localization and function of [PDF] [Full Text] [Abstract] , December 15, 2011; 589 (24): 6029-6038. J Physiol Mala M. Shah, Michele Migliore and David A. Brown compartments of rat hippocampal pyramidal neurons Differential effects of Kv7 (M-) channels on synaptic integration in distinct subcellular [PDF] [Full Text] [Abstract] , January 30, 2013; 33 (5): 2156-2165. J. Neurosci. Michele Ferrante, Michele Migliore and Giorgio A. Ascoli Functional Impact of Dendritic Branch-Point Morphology including high resolution figures, can be found at: Updated information and services /content/94/6/4145.full.html can be found at: Journal of Neurophysiology about Additional material and information http://www.the-aps.org/publications/jn This information is current as of March 7, 2014. American Physiological Society. ISSN: 0022-3077, ESSN: 1522-1598. Visit our website at http://www.the-aps.org/. (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by the publishes original articles on the function of the nervous system. It is published 12 times a year Journal of Neurophysiology on March 7, 2014 Downloaded from on March 7, 2014 Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Signal Propagation in Oblique Dendrites of CA1

doi:10.1152/jn.00521.2005 94:4145-4155, 2005.J NeurophysiolMichele Migliore, Michele Ferrante and Giorgio A. AscoliPyramidal CellsSignal Propagation in Oblique Dendrites of CA1
You might find this additional info useful...
59 articles, 26 of which can be accessed free at:This article cites /content/94/6/4145.full.html#ref-list-1
9 other HighWire hosted articles, the first 5 are:This article has been cited by
[PDF] [Full Text] [Abstract]
, December , 2009; 102 (6): 3234-3250.J NeurophysiolVladislav Volman, Herbert Levine, Eshel Ben-Jacob and Terrence J. SejnowskiDendritic ConductancesLocally Balanced Dendritic Integration by Short-Term Synaptic Plasticity and Active
[PDF] [Full Text] [Abstract], May 5, 2010; 30 (18): 6434-6442.J. Neurosci.
Giorgio A. Ascoli, Sonia Gasparini, Virginia Medinilla and Michele MiglioreLocal Control of Postinhibitory Rebound Spiking in CA1 Pyramidal Neuron Dendrites
[PDF] [Full Text] [Abstract], July , 2011; 106 (1): 44-58.J Neurophysiol
Marie L. Goeritz, Qing Ouyang and Ronald M. Harris-Warrick channels in a small neural networkhILocalization and function of
[PDF] [Full Text] [Abstract]
, December 15, 2011; 589 (24): 6029-6038.J PhysiolMala M. Shah, Michele Migliore and David A. Browncompartments of rat hippocampal pyramidal neuronsDifferential effects of Kv7 (M-) channels on synaptic integration in distinct subcellular
[PDF] [Full Text] [Abstract], January 30, 2013; 33 (5): 2156-2165.J. Neurosci.
Michele Ferrante, Michele Migliore and Giorgio A. AscoliFunctional Impact of Dendritic Branch-Point Morphology
including high resolution figures, can be found at:Updated information and services /content/94/6/4145.full.html
can be found at:Journal of Neurophysiologyabout Additional material and information http://www.the-aps.org/publications/jn
This information is current as of March 7, 2014.
American Physiological Society. ISSN: 0022-3077, ESSN: 1522-1598. Visit our website at http://www.the-aps.org/.(monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by the
publishes original articles on the function of the nervous system. It is published 12 times a yearJournal of Neurophysiology
on March 7, 2014
Dow
nloaded from on M
arch 7, 2014D
ownloaded from

Signal Propagation in Oblique Dendrites of CA1 Pyramidal Cells
Michele Migliore,1,2 Michele Ferrante,3 and Giorgio A. Ascoli3,4
1Department of Neurobiology, Yale University School of Medicine, New Haven, Connecticut; 2Institute of Biophysics, National ResearchCouncil, Palermo, Italy; 3Krasnow Institute for Advanced Study and 4Psychology Department, George Mason University,Fairfax, Virginia
Submitted 19 May 2005; accepted in final form 17 August 2005
Migliore, Michele, Michele Ferrante, and Giorgio A. Ascoli. Signalpropagation in oblique dendrites of CA1 pyramidal cells. J Neurophysiol 94:4145–4155, 2005; doi:10.1152/jn.00521.2005. The electrophysiologicalproperties of the oblique branches of CA1 pyramidal neurons arelargely unknown and very difficult to investigate experimentally.These relatively thin dendrites make up the majority of the apical treesurface area and constitute the main target of Schaffer collateral axonsfrom CA3. Their electrogenic properties might have an important rolein defining the computational functions of CA1 neurons. It is thusimportant to determine if and to what extent the back- and forwardpropagation of action potentials (AP) in these dendrites could bemodulated by local properties such as morphology or active conduc-tances. In the first detailed study of signal propagation in the fullextent of CA1 oblique dendrites, we used 27 reconstructed three-dimensional morphologies and different distributions of the A-typeK� conductance (KA), to investigate their electrophysiological prop-erties by computational modeling. We found that the local KA distri-bution had a major role in modulating action potential back propaga-tion, whereas the forward propagation of dendritic spikes originatingin the obliques was mainly affected by local morphological properties.In both cases, signal processing in any given oblique was effectivelyindependent of the rest of the neuron and, by and large, of the distancefrom the soma. Moreover, the density of KA in oblique dendritesaffected spike conduction in the main shaft. Thus the anatomicalvariability of CA1 pyramidal cells and their local distribution ofvoltage-gated channels may suit a powerful functional compartmen-talization of the apical tree.
I N T R O D U C T I O N
Pyramidal neurons in hippocampal area CA1 and neocorticallayer V have many small oblique dendrites stemming out of themain apical trunk at different distances from the soma. Themain apical trunk of CA1 pyramidal cells has been intensivelyinvestigated with respect to the distribution and kinetics ofvoltage-gated conductances (e.g., Bekkers 2000; Chen andJohnston 2004; Magee and Johnston 1995; Shao et al. 1999),electrogenic characteristics (e.g., Bernard and Johnston 2003;Gasparini et al. 2004; Golding et al. 2001; Kasuka et al. 2003),and synaptic properties (e.g., Frick et al. 2004; Otmakhova etal. 2002; Spruston et al. 1995; Zamanillo et al. 1999). Incontrast, the experimental investigation of the biophysicalproperties of oblique dendrites is exceedingly difficult becauseof the technical unfeasibility of patching such fine branches inuncertain positions. Their possible functional roles thus remainlargely unknown.
This is an important issue because in CA1 cells, obliquedendrites constitute the majority of the apical tree surface and
represent the main site of excitatory synaptic target from CA3Schaffer collaterals. A recent single-cell computational studyshowed that the CA1 pyramidal neuron’s subthreshold integra-tion of synaptic inputs can be characterized as a sum ofnonlinear responses, each from an individual oblique dendrite(Poirazi et al. 2003a,b). Experimental investigations in corticalpyramidal cells confirmed that synchronous inputs are inte-grated sigmoidally within the same dendritic branch, but thesummation becomes linear among inputs located on widelyseparate dendrites (Polsky et al. 2004). These data suggest thatoblique dendrites constitute fairly independent input/outputunits that can be viewed as a separate computational layer fromthe soma and main trunk. Such views point to the need for adeeper understanding of suprathreshold signal propagation toand from the oblique trees as an essential step to characterizethe computational power of CA1 pyramidal cells. This isespecially important in light of the direct experimental evi-dence (from the main trunk) that spike back propagation cansubserve Hebbian plasticity (Magee and Johnston 1997;Markram et al. 1997). If the invasion of oblique dendrites byantidromic spikes can be independently modulated, synapseson various trees could have different susceptibility to associa-tive plasticity. Similarly, relative isolation of oblique treesfrom the rest of the neuron would enable local Hebbianplasticity mediated by dendritic spikes (e.g., Alkon 1999).
It is well established, with both models and experiments, thataction potential (AP) propagation in the main trunk of pyra-midal neurons can be regulated by active conductances (e.g.,Hoffman et al. 1997; Johnston et al. 1999; Migliore et al. 1999)and local morphology (Schaefer et al. 2003; Vetter et al. 2001).However, a detailed picture of how APs propagate into/fromthe oblique dendrites is still missing. The major problem is thatexperimental data on CA1 oblique biophysics is limited to afew studies on the interaction of subthreshold inputs (Cash andYuste 1999; Liu 2004) and imaging studies of Ca2� concen-tration and dynamics (Frick et al. 2003; Nakamura et al. 2002).The local distribution of active channels is still largely un-known with the notable exception of the work by Frick et al.(2003), demonstrating the presence of a full set of activechannels with a possible major role for the A-type K� con-ductance (KA). Because this current is modulated by proteinkinases (Johnston et al. 1999), and thus ultimately by localneuronal activity, signal propagation into/from oblique den-drites cannot be expected to follow along the same lines as inthe trunk. It will depend on the interaction of factors that, inaddition to the local KA density, include distance from soma,
Address for reprint requests and other correspondence: M. Migliore, Dept.of Neurobiology, Yale University School of Medicine, New Haven, CT06520(E-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
J Neurophysiol 94: 4145–4155, 2005;doi:10.1152/jn.00521.2005.
41450022-3077/05 $8.00 Copyright © 2005 The American Physiological Societywww.jn.org
on March 7, 2014
Dow
nloaded from
local geometry at branch points, and dendritic AP initiationsite. These issues have not, and cannot be, easily exploredexperimentally. In this paper, we use realistic computationalmodels of CA1 pyramidal cells to determine how and to whatextent the local KA and morphological properties might regu-late the backward and forward propagation of action potentials(APs) in the full length of the oblique branches (i.e., �600 �mfrom the soma).
M E T H O D S
All simulations were implemented and run with the NEURONprogram (v5.7) (Hines and Carnevale 1997) using the variable timestep feature. In most cases, simulations were carried out using theParallelContext () feature of NEURON under MPI on a Beowulfcluster (34 processors) or a IBM Linux cluster (1024 processors). Themodel and simulation files are available for public download under theModelDB section (Davison et al. 2004; Migliore et al. 2003) of theSenselab database (http://senselab.yale.med.edu).
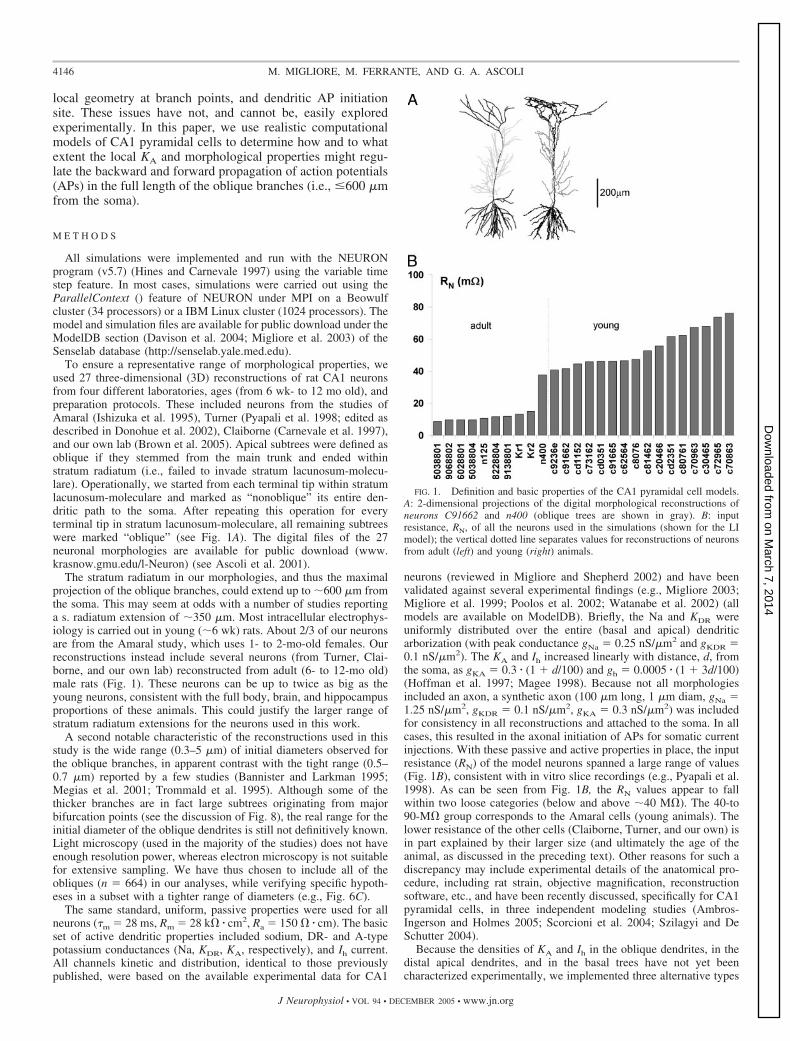
To ensure a representative range of morphological properties, weused 27 three-dimensional (3D) reconstructions of rat CA1 neuronsfrom four different laboratories, ages (from 6 wk- to 12 mo old), andpreparation protocols. These included neurons from the studies ofAmaral (Ishizuka et al. 1995), Turner (Pyapali et al. 1998; edited asdescribed in Donohue et al. 2002), Claiborne (Carnevale et al. 1997),and our own lab (Brown et al. 2005). Apical subtrees were defined asoblique if they stemmed from the main trunk and ended withinstratum radiatum (i.e., failed to invade stratum lacunosum-molecu-lare). Operationally, we started from each terminal tip within stratumlacunosum-moleculare and marked as “nonoblique” its entire den-dritic path to the soma. After repeating this operation for everyterminal tip in stratum lacunosum-moleculare, all remaining subtreeswere marked “oblique” (see Fig. 1A). The digital files of the 27neuronal morphologies are available for public download (www.krasnow.gmu.edu/l-Neuron) (see Ascoli et al. 2001).
The stratum radiatum in our morphologies, and thus the maximalprojection of the oblique branches, could extend up to �600 �m fromthe soma. This may seem at odds with a number of studies reportinga s. radiatum extension of �350 �m. Most intracellular electrophys-iology is carried out in young (�6 wk) rats. About 2/3 of our neuronsare from the Amaral study, which uses 1- to 2-mo-old females. Ourreconstructions instead include several neurons (from Turner, Clai-borne, and our own lab) reconstructed from adult (6- to 12-mo old)male rats (Fig. 1). These neurons can be up to twice as big as theyoung neurons, consistent with the full body, brain, and hippocampusproportions of these animals. This could justify the larger range ofstratum radiatum extensions for the neurons used in this work.
A second notable characteristic of the reconstructions used in thisstudy is the wide range (0.3–5 �m) of initial diameters observed forthe oblique branches, in apparent contrast with the tight range (0.5–0.7 �m) reported by a few studies (Bannister and Larkman 1995;Megias et al. 2001; Trommald et al. 1995). Although some of thethicker branches are in fact large subtrees originating from majorbifurcation points (see the discussion of Fig. 8), the real range for theinitial diameter of the oblique dendrites is still not definitively known.Light microscopy (used in the majority of the studies) does not haveenough resolution power, whereas electron microscopy is not suitablefor extensive sampling. We have thus chosen to include all of theobliques (n � 664) in our analyses, while verifying specific hypoth-eses in a subset with a tighter range of diameters (e.g., Fig. 6C).
The same standard, uniform, passive properties were used for allneurons (�m � 28 ms, Rm � 28 k� � cm2, Ra � 150 � � cm). The basicset of active dendritic properties included sodium, DR- and A-typepotassium conductances (Na, KDR, KA, respectively), and Ih current.All channels kinetic and distribution, identical to those previouslypublished, were based on the available experimental data for CA1
neurons (reviewed in Migliore and Shepherd 2002) and have beenvalidated against several experimental findings (e.g., Migliore 2003;Migliore et al. 1999; Poolos et al. 2002; Watanabe et al. 2002) (allmodels are available on ModelDB). Briefly, the Na and KDR wereuniformly distributed over the entire (basal and apical) dendriticarborization (with peak conductance gNa � 0.25 nS/�m2 and gKDR �0.1 nS/�m2). The KA and Ih increased linearly with distance, d, fromthe soma, as gKA � 0.3 � (1 � d/100) and gh � 0.0005 � (1 � 3d/100)(Hoffman et al. 1997; Magee 1998). Because not all morphologiesincluded an axon, a synthetic axon (100 �m long, 1 �m diam, gNa �1.25 nS/�m2, gKDR � 0.1 nS/�m2, gKA � 0.3 nS/�m2) was includedfor consistency in all reconstructions and attached to the soma. In allcases, this resulted in the axonal initiation of APs for somatic currentinjections. With these passive and active properties in place, the inputresistance (RN) of the model neurons spanned a large range of values(Fig. 1B), consistent with in vitro slice recordings (e.g., Pyapali et al.1998). As can be seen from Fig. 1B, the RN values appear to fallwithin two loose categories (below and above �40 M�). The 40-to90-M� group corresponds to the Amaral cells (young animals). Thelower resistance of the other cells (Claiborne, Turner, and our own) isin part explained by their larger size (and ultimately the age of theanimal, as discussed in the preceding text). Other reasons for such adiscrepancy may include experimental details of the anatomical pro-cedure, including rat strain, objective magnification, reconstructionsoftware, etc., and have been recently discussed, specifically for CA1pyramidal cells, in three independent modeling studies (Ambros-Ingerson and Holmes 2005; Scorcioni et al. 2004; Szilagyi and DeSchutter 2004).
Because the densities of KA and Ih in the oblique dendrites, in thedistal apical dendrites, and in the basal trees have not yet beencharacterized experimentally, we implemented three alternative types
FIG. 1. Definition and basic properties of the CA1 pyramidal cell models.A: 2-dimensional projections of the digital morphological reconstructions ofneurons C91662 and n400 (oblique trees are shown in gray). B: inputresistance, RN, of all the neurons used in the simulations (shown for the LImodel); the vertical dotted line separates values for reconstructions of neuronsfrom adult (left) and young (right) animals.
4146 M. MIGLIORE, M. FERRANTE, AND G. A. ASCOLI
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on March 7, 2014
Dow
nloaded from

of distributions, schematically reported in Table 1. In the “linearincrease” type (LI), we assumed that the experimental findings re-garding the distribution of active conductances in the main apicaltrunk were representative of the whole dendritic arborization. Accord-ingly, in this case, KA and Ih were linearly increased everywhere,including oblique, basal, and distal apical dendrites. In the “constant”distribution type (C1), we assumed no information beyond that ob-tained directly from dendritic patch-clamp recordings (Hoffmann etal. 1997; Magee 1998). The KA and Ih linearly increased in the maintrunk �350 �m from soma and were assumed to remain constant inthe rest of the dendrites at the same value of the closest location in thisrange. In particular, each oblique subtree had constant KA and Ih at thevalue of the point of attachment on the main trunk; distal apicaldendrites (all the subtrees of the main trunk beyond 350 �m) hadconstant active current distributions at the value of the main trunk at350 �m; and basal dendrites had constant values equal to the somaticdensities. Finally, the type C3 had the same distributions as C1 withthe exception of the KA values in the oblique. Following the indirectexperimental suggestion that the density of these channels could behigher in the obliques than in the trunk (Frick et al. 2003), in this case,KA was increased threefold in each oblique with respect to its value atthe point of attachment in the main trunk. Little differences wereobserved in the input resistance of any given neuron using the threekinds of distribution (data not shown).
It should be noted that a number of additional mechanisms reportedfor CA1 pyramidal cells were not included in the model. Many ofthem, such as additional K� conductances, persistent Na� current,Ca2�-dependent currents, but also activity-dependent changes inchannel density or kinetic, nonuniform distribution of various Ca2�
currents, and intracellular Ca2� dynamics, may modulate the signalpropagation process (Golding et al. 1999; Lazarewicz et al. 2002;Magee and Carruth 1999). This is precisely why we did not includethem in the model at this stage. In fact, we were interested in studyingsignal propagation to and from the oblique branches in CA1 neurons,a process that is exceedingly difficult to study experimentally and forthe underlying mechanisms of which very little empirical evidence isavailable. For these reasons, we included only the main conductancesinvolved in action potential generation and regulation (Na, KDR, andKA), and in subthreshold signal modulation (Ih). Such choice impliesthat these simulations can provide little definitive evidence of howspike initiation in oblique dendrites might occur and restricts the mainobject of this study to propagation, rather than initiation, of actionpotentials. It would be interesting to include additional cell propertiesin future studies to investigate how and to what extent they affect thebasic findings reported in this paper.
Oblique stimulation was implemented with a double exponentialtime course (using the Exp2Syn built-in function of NEURON) with 1and 5 ms for the rise and decay time, respectively, and a reversalpotential of 0 mV. These values are consistent with experimentalfindings (Andrasfalvy and Magee 2001), and no qualitative differ-ences were observed using different values in preliminary test simu-lations. The peak conductance was adjusted for each oblique com-partment as the minimum value sufficient to elicit a local spike withinthe same oblique tree. The resulting means � SD for the LI, C1, and
C3 distribution types were (all in nS) 4.35 � 2.76, 5.01 � 4.69, and10.15 � 8.89, respectively. In a set of simulations, a dendritic spikewas elicited in each of the oblique compartments while injecting asubthreshold steady depolarizing current in the main trunk at the pointof attachment of the oblique tree. Trunk current injection valuesstarted from 0.01 nA and were increased by 0.01 nA at each iterationof the simulation. Membrane voltage was monitored both in the maintrunk and in the soma, and the local spike was elicited in the obliquetree after stabilization of the somatic voltage. In particular, the localspike was elicited after 200 ms from the beginning of the depolarizingcurrent step in the trunk (this delay was found to be long enough toallow for both decay of transients and steady-state conditions for allthe currents). The iteration stopped either when a spike was detectedin the soma in response to oblique stimulation or when a spike waselicited in the trunk by the current injection (i.e., the injected currentwas no longer subthreshold).
R E S U L T S
The CA1 pyramidal cell computational model was con-structed and validated based on 3D anatomical reconstructionsand available experimental data on the dendritic distribution ofactive and passive properties, as detailed in METHODS. Becausethe majority of experimental recordings from CA1 pyramidalcell dendrites are collected from the main trunk (Fig. 1A: blackportion of the apical tree within �350 �m from the soma),three alternative distributions of conductance densities wereemployed in all simulations for the rest of the dendrites, LI, C1,and C3 (see Table 1 and METHODS for details). In all cases, theneurons exhibited a plausible range of input resistances (Fig.1B shows the values for the LI model).
We first investigated the factors controlling AP invasion ofobliques. The simulation protocol to study AP back-propaga-tion was to elicit a single somatic spike with a short (2 ms)current pulse in the soma of each neuron and record the peakamplitude of the AP during somatofugal propagation along themain trunk and each of the oblique subtrees (Fig. 2A). Thevalues of the current pulses for the LI, C1, and C3 distributionswere 0.93 � 0.76, 0.85 � 0.74, and 0.86 � 0.77 nA, respec-tively. The distribution of the average peak depolarization ofthe obliques (Fig. 2B) suggests that back propagation dependson the local KA distribution and tends to be all-or-none. Backpropagation does not depend, in contrast, on the local morpho-logical properties, such as the ratio between the initial obliquediameter and the diameter of the trunk at the point of attach-ment (Fig. 2C) or the oblique surface area (Fig. 2D). Note that,in most cases (e.g., Fig. 2A), the spike amplitude increases oninvasion of a small oblique dendrite attached to a large mainbranch as would be expected for reasons of alterations in thesurface-to-volume ratio. Figure 2C, in contrast, shows that theaverage over the peak values of the membrane voltage reached
TABLE 1. Schematic description of the distribution for the peak KA and Ih conductances in the configurations explored inthis work (LI, C1, and C3)
KA Ih
Trunk Oblique Trunk Oblique
LI Linear increase (L.i.) L.i. L.i. L.i.C1 �350 �m: L.i. �350 �m: same
value as 350 �mConstant: same value as branchpoint �350 �m: L.i. �350 �m: same
value as 350 �mConstant: same value as branchpoint
C3 Same as C1 Constant: 3 � the value atbranchpoint
Same as C1 Same as C1
4147SIGNAL PROPAGATION IN CA1 OBLIQUES
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on March 7, 2014
Dow
nloaded from
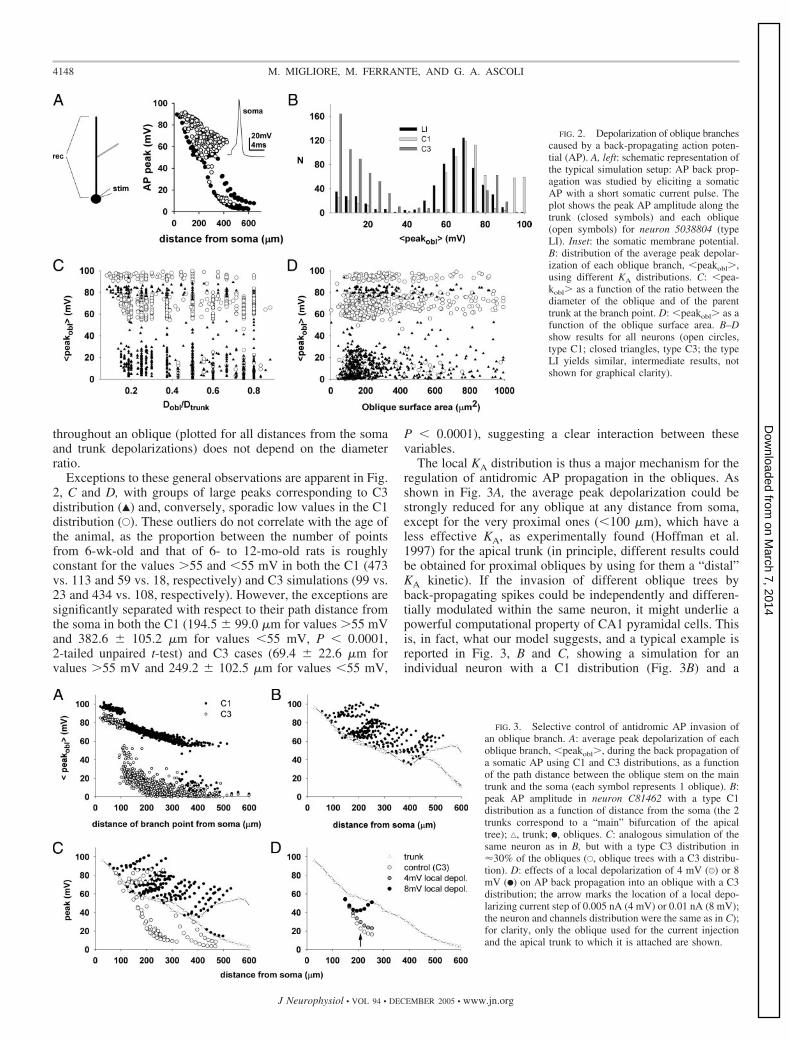
throughout an oblique (plotted for all distances from the somaand trunk depolarizations) does not depend on the diameterratio.
Exceptions to these general observations are apparent in Fig.2, C and D, with groups of large peaks corresponding to C3distribution (Œ) and, conversely, sporadic low values in the C1distribution (E). These outliers do not correlate with the age ofthe animal, as the proportion between the number of pointsfrom 6-wk-old and that of 6- to 12-mo-old rats is roughlyconstant for the values �55 and �55 mV in both the C1 (473vs. 113 and 59 vs. 18, respectively) and C3 simulations (99 vs.23 and 434 vs. 108, respectively). However, the exceptions aresignificantly separated with respect to their path distance fromthe soma in both the C1 (194.5 � 99.0 �m for values �55 mVand 382.6 � 105.2 �m for values �55 mV, P � 0.0001,2-tailed unpaired t-test) and C3 cases (69.4 � 22.6 �m forvalues �55 mV and 249.2 � 102.5 �m for values �55 mV,
P � 0.0001), suggesting a clear interaction between thesevariables.
The local KA distribution is thus a major mechanism for theregulation of antidromic AP propagation in the obliques. Asshown in Fig. 3A, the average peak depolarization could bestrongly reduced for any oblique at any distance from soma,except for the very proximal ones (�100 �m), which have aless effective KA, as experimentally found (Hoffman et al.1997) for the apical trunk (in principle, different results couldbe obtained for proximal obliques by using for them a “distal”KA kinetic). If the invasion of different oblique trees byback-propagating spikes could be independently and differen-tially modulated within the same neuron, it might underlie apowerful computational property of CA1 pyramidal cells. Thisis, in fact, what our model suggests, and a typical example isreported in Fig. 3, B and C, showing a simulation for anindividual neuron with a C1 distribution (Fig. 3B) and a
FIG. 2. Depolarization of oblique branchescaused by a back-propagating action poten-tial (AP). A, left: schematic representation ofthe typical simulation setup: AP back prop-agation was studied by eliciting a somaticAP with a short somatic current pulse. Theplot shows the peak AP amplitude along thetrunk (closed symbols) and each oblique(open symbols) for neuron 5038804 (typeLI). Inset: the somatic membrane potential.B: distribution of the average peak depolar-ization of each oblique branch, �peakobl�,using different KA distributions. C: �pea-kobl� as a function of the ratio between thediameter of the oblique and of the parenttrunk at the branch point. D: �peakobl� as afunction of the oblique surface area. B–Dshow results for all neurons (open circles,type C1; closed triangles, type C3; the typeLI yields similar, intermediate results, notshown for graphical clarity).
FIG. 3. Selective control of antidromic AP invasion ofan oblique branch. A: average peak depolarization of eachoblique branch, �peakobl�, during the back propagation ofa somatic AP using C1 and C3 distributions, as a functionof the path distance between the oblique stem on the maintrunk and the soma (each symbol represents 1 oblique). B:peak AP amplitude in neuron C81462 with a type C1distribution as a function of distance from the soma (the 2trunks correspond to a “main” bifurcation of the apicaltree); ‚, trunk; F, obliques. C: analogous simulation of thesame neuron as in B, but with a type C3 distribution in30% of the obliques (E, oblique trees with a C3 distribu-tion). D: effects of a local depolarization of 4 mV (U) or 8mV (F) on AP back propagation into an oblique with a C3distribution; the arrow marks the location of a local depo-larizing current step of 0.005 nA (4 mV) or 0.01 nA (8 mV);the neuron and channels distribution were the same as in C);for clarity, only the oblique used for the current injectionand the apical trunk to which it is attached are shown.
4148 M. MIGLIORE, M. FERRANTE, AND G. A. ASCOLI
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on March 7, 2014
Dow
nloaded from
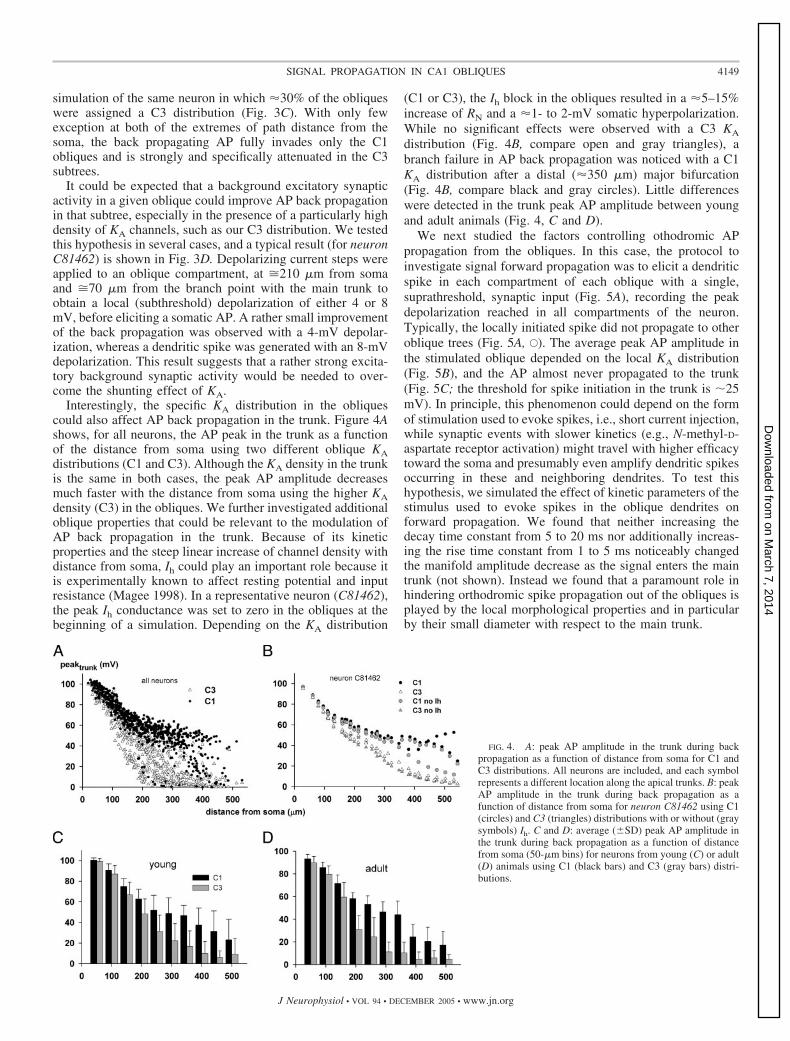
simulation of the same neuron in which 30% of the obliqueswere assigned a C3 distribution (Fig. 3C). With only fewexception at both of the extremes of path distance from thesoma, the back propagating AP fully invades only the C1obliques and is strongly and specifically attenuated in the C3subtrees.
It could be expected that a background excitatory synapticactivity in a given oblique could improve AP back propagationin that subtree, especially in the presence of a particularly highdensity of KA channels, such as our C3 distribution. We testedthis hypothesis in several cases, and a typical result (for neuronC81462) is shown in Fig. 3D. Depolarizing current steps wereapplied to an oblique compartment, at �210 �m from somaand �70 �m from the branch point with the main trunk toobtain a local (subthreshold) depolarization of either 4 or 8mV, before eliciting a somatic AP. A rather small improvementof the back propagation was observed with a 4-mV depolar-ization, whereas a dendritic spike was generated with an 8-mVdepolarization. This result suggests that a rather strong excita-tory background synaptic activity would be needed to over-come the shunting effect of KA.
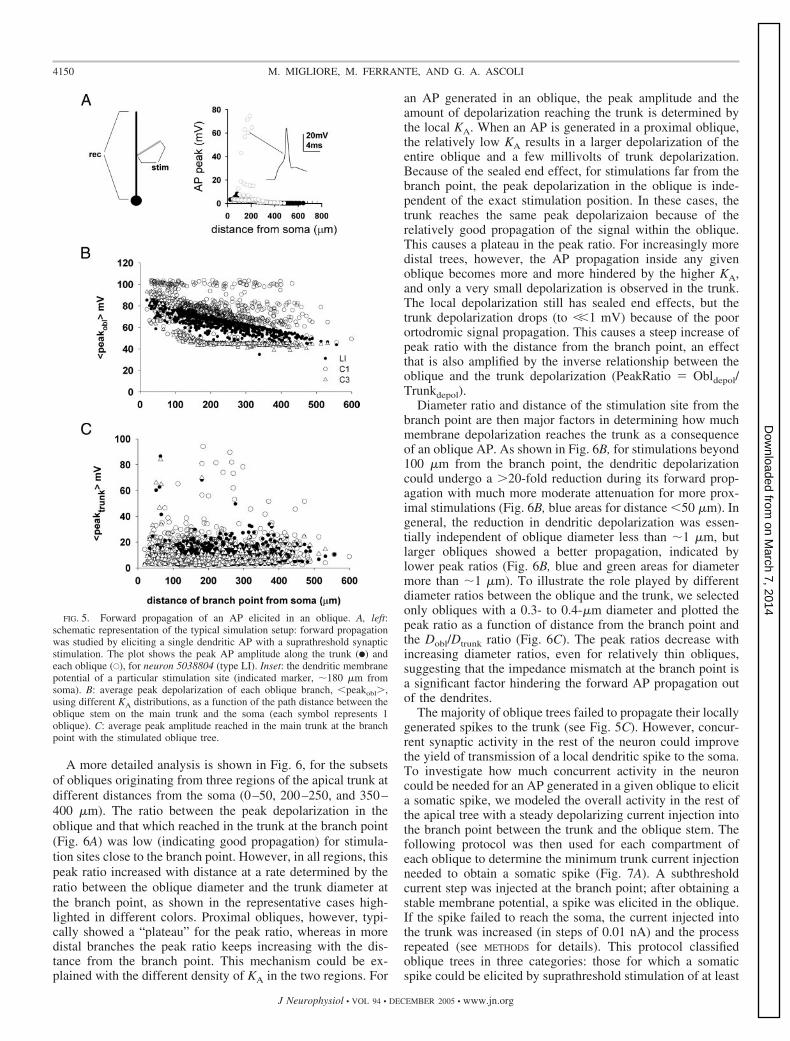
Interestingly, the specific KA distribution in the obliquescould also affect AP back propagation in the trunk. Figure 4Ashows, for all neurons, the AP peak in the trunk as a functionof the distance from soma using two different oblique KAdistributions (C1 and C3). Although the KA density in the trunkis the same in both cases, the peak AP amplitude decreasesmuch faster with the distance from soma using the higher KAdensity (C3) in the obliques. We further investigated additionaloblique properties that could be relevant to the modulation ofAP back propagation in the trunk. Because of its kineticproperties and the steep linear increase of channel density withdistance from soma, Ih could play an important role because itis experimentally known to affect resting potential and inputresistance (Magee 1998). In a representative neuron (C81462),the peak Ih conductance was set to zero in the obliques at thebeginning of a simulation. Depending on the KA distribution
(C1 or C3), the Ih block in the obliques resulted in a 5–15%increase of RN and a 1- to 2-mV somatic hyperpolarization.While no significant effects were observed with a C3 KA
distribution (Fig. 4B, compare open and gray triangles), abranch failure in AP back propagation was noticed with a C1KA distribution after a distal (350 �m) major bifurcation(Fig. 4B, compare black and gray circles). Little differenceswere detected in the trunk peak AP amplitude between youngand adult animals (Fig. 4, C and D).
We next studied the factors controlling othodromic APpropagation from the obliques. In this case, the protocol toinvestigate signal forward propagation was to elicit a dendriticspike in each compartment of each oblique with a single,suprathreshold, synaptic input (Fig. 5A), recording the peakdepolarization reached in all compartments of the neuron.Typically, the locally initiated spike did not propagate to otheroblique trees (Fig. 5A, E). The average peak AP amplitude inthe stimulated oblique depended on the local KA distribution(Fig. 5B), and the AP almost never propagated to the trunk(Fig. 5C; the threshold for spike initiation in the trunk is �25mV). In principle, this phenomenon could depend on the formof stimulation used to evoke spikes, i.e., short current injection,while synaptic events with slower kinetics (e.g., N-methyl-D-aspartate receptor activation) might travel with higher efficacytoward the soma and presumably even amplify dendritic spikesoccurring in these and neighboring dendrites. To test thishypothesis, we simulated the effect of kinetic parameters of thestimulus used to evoke spikes in the oblique dendrites onforward propagation. We found that neither increasing thedecay time constant from 5 to 20 ms nor additionally increas-ing the rise time constant from 1 to 5 ms noticeably changedthe manifold amplitude decrease as the signal enters the maintrunk (not shown). Instead we found that a paramount role inhindering orthodromic spike propagation out of the obliques isplayed by the local morphological properties and in particularby their small diameter with respect to the main trunk.
FIG. 4. A: peak AP amplitude in the trunk during backpropagation as a function of distance from soma for C1 andC3 distributions. All neurons are included, and each symbolrepresents a different location along the apical trunks. B: peakAP amplitude in the trunk during back propagation as afunction of distance from soma for neuron C81462 using C1(circles) and C3 (triangles) distributions with or without (graysymbols) Ih. C and D: average (�SD) peak AP amplitude inthe trunk during back propagation as a function of distancefrom soma (50-�m bins) for neurons from young (C) or adult(D) animals using C1 (black bars) and C3 (gray bars) distri-butions.
4149SIGNAL PROPAGATION IN CA1 OBLIQUES
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on March 7, 2014
Dow
nloaded from
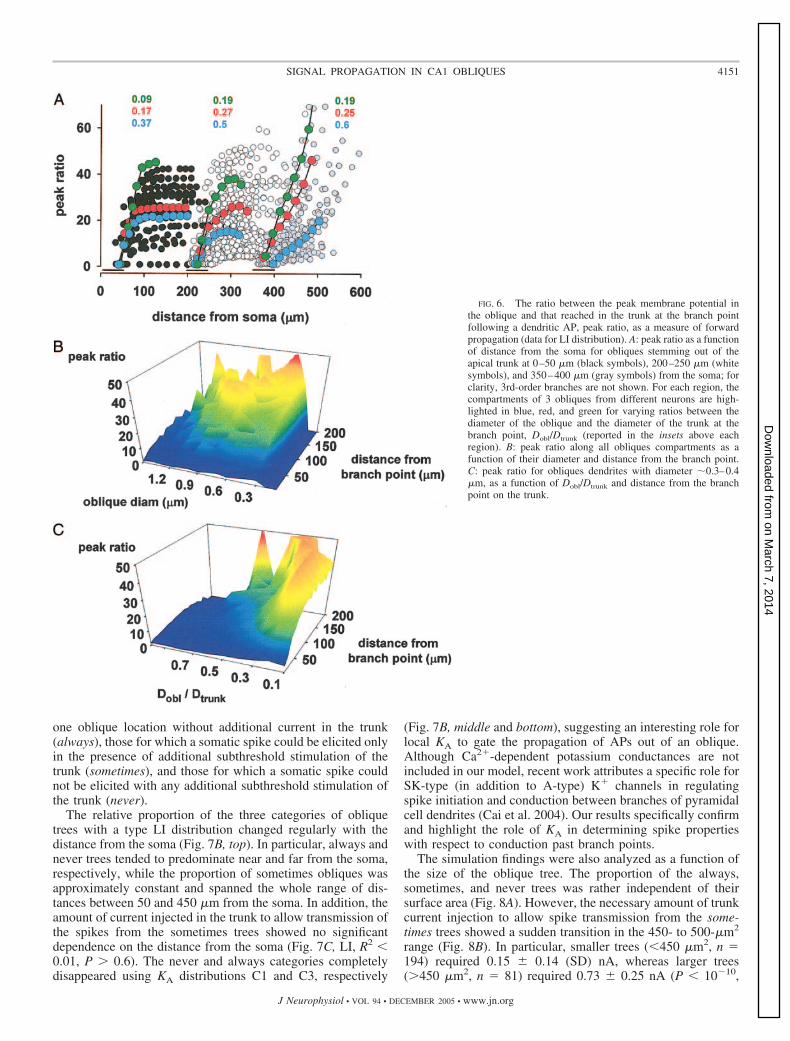
A more detailed analysis is shown in Fig. 6, for the subsetsof obliques originating from three regions of the apical trunk atdifferent distances from the soma (0–50, 200–250, and 350–400 �m). The ratio between the peak depolarization in theoblique and that which reached in the trunk at the branch point(Fig. 6A) was low (indicating good propagation) for stimula-tion sites close to the branch point. However, in all regions, thispeak ratio increased with distance at a rate determined by theratio between the oblique diameter and the trunk diameter atthe branch point, as shown in the representative cases high-lighted in different colors. Proximal obliques, however, typi-cally showed a “plateau” for the peak ratio, whereas in moredistal branches the peak ratio keeps increasing with the dis-tance from the branch point. This mechanism could be ex-plained with the different density of KA in the two regions. For
an AP generated in an oblique, the peak amplitude and theamount of depolarization reaching the trunk is determined bythe local KA. When an AP is generated in a proximal oblique,the relatively low KA results in a larger depolarization of theentire oblique and a few millivolts of trunk depolarization.Because of the sealed end effect, for stimulations far from thebranch point, the peak depolarization in the oblique is inde-pendent of the exact stimulation position. In these cases, thetrunk reaches the same peak depolarizaion because of therelatively good propagation of the signal within the oblique.This causes a plateau in the peak ratio. For increasingly moredistal trees, however, the AP propagation inside any givenoblique becomes more and more hindered by the higher KA,and only a very small depolarization is observed in the trunk.The local depolarization still has sealed end effects, but thetrunk depolarization drops (to ��1 mV) because of the poorortodromic signal propagation. This causes a steep increase ofpeak ratio with the distance from the branch point, an effectthat is also amplified by the inverse relationship between theoblique and the trunk depolarization (PeakRatio � Obldepol/Trunkdepol).
Diameter ratio and distance of the stimulation site from thebranch point are then major factors in determining how muchmembrane depolarization reaches the trunk as a consequenceof an oblique AP. As shown in Fig. 6B, for stimulations beyond100 �m from the branch point, the dendritic depolarizationcould undergo a �20-fold reduction during its forward prop-agation with much more moderate attenuation for more prox-imal stimulations (Fig. 6B, blue areas for distance �50 �m). Ingeneral, the reduction in dendritic depolarization was essen-tially independent of oblique diameter less than �1 �m, butlarger obliques showed a better propagation, indicated bylower peak ratios (Fig. 6B, blue and green areas for diametermore than �1 �m). To illustrate the role played by differentdiameter ratios between the oblique and the trunk, we selectedonly obliques with a 0.3- to 0.4-�m diameter and plotted thepeak ratio as a function of distance from the branch point andthe Dobl/Dtrunk ratio (Fig. 6C). The peak ratios decrease withincreasing diameter ratios, even for relatively thin obliques,suggesting that the impedance mismatch at the branch point isa significant factor hindering the forward AP propagation outof the dendrites.
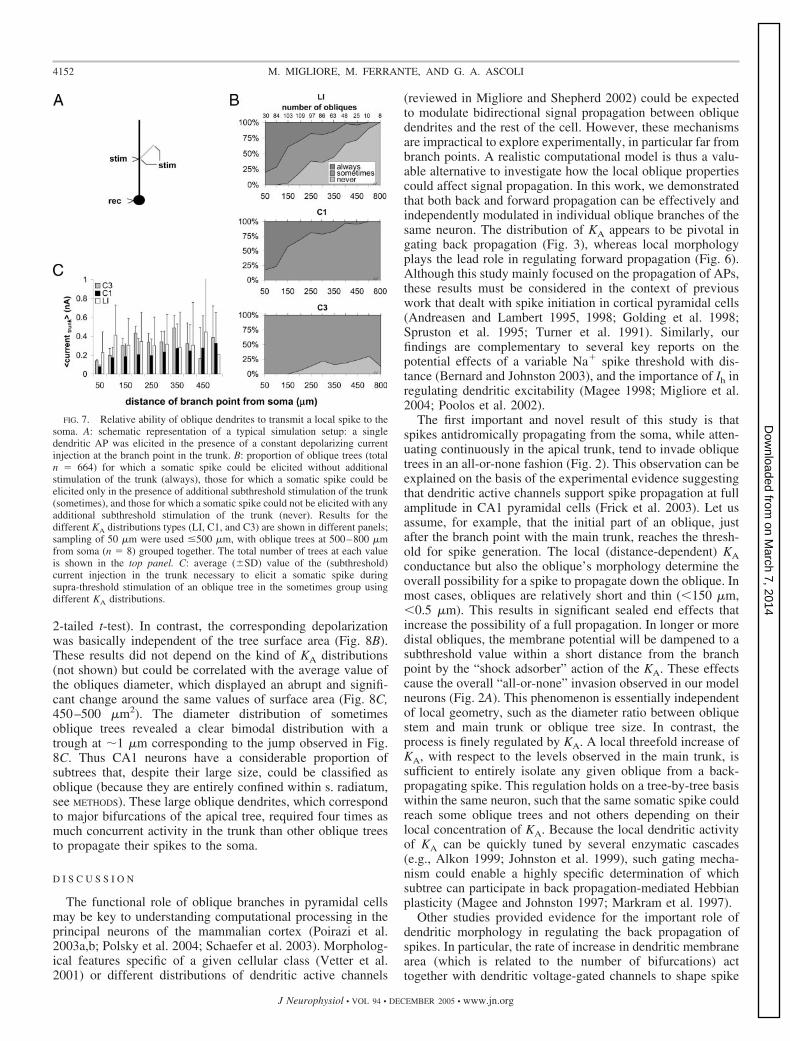
The majority of oblique trees failed to propagate their locallygenerated spikes to the trunk (see Fig. 5C). However, concur-rent synaptic activity in the rest of the neuron could improvethe yield of transmission of a local dendritic spike to the soma.To investigate how much concurrent activity in the neuroncould be needed for an AP generated in a given oblique to elicita somatic spike, we modeled the overall activity in the rest ofthe apical tree with a steady depolarizing current injection intothe branch point between the trunk and the oblique stem. Thefollowing protocol was then used for each compartment ofeach oblique to determine the minimum trunk current injectionneeded to obtain a somatic spike (Fig. 7A). A subthresholdcurrent step was injected at the branch point; after obtaining astable membrane potential, a spike was elicited in the oblique.If the spike failed to reach the soma, the current injected intothe trunk was increased (in steps of 0.01 nA) and the processrepeated (see METHODS for details). This protocol classifiedoblique trees in three categories: those for which a somaticspike could be elicited by suprathreshold stimulation of at least
FIG. 5. Forward propagation of an AP elicited in an oblique. A, left:schematic representation of the typical simulation setup: forward propagationwas studied by eliciting a single dendritic AP with a suprathreshold synapticstimulation. The plot shows the peak AP amplitude along the trunk (F) andeach oblique (E), for neuron 5038804 (type LI). Inset: the dendritic membranepotential of a particular stimulation site (indicated marker, �180 �m fromsoma). B: average peak depolarization of each oblique branch, �peakobl�,using different KA distributions, as a function of the path distance between theoblique stem on the main trunk and the soma (each symbol represents 1oblique). C: average peak amplitude reached in the main trunk at the branchpoint with the stimulated oblique tree.
4150 M. MIGLIORE, M. FERRANTE, AND G. A. ASCOLI
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on March 7, 2014
Dow
nloaded from
one oblique location without additional current in the trunk(always), those for which a somatic spike could be elicited onlyin the presence of additional subthreshold stimulation of thetrunk (sometimes), and those for which a somatic spike couldnot be elicited with any additional subthreshold stimulation ofthe trunk (never).
The relative proportion of the three categories of obliquetrees with a type LI distribution changed regularly with thedistance from the soma (Fig. 7B, top). In particular, always andnever trees tended to predominate near and far from the soma,respectively, while the proportion of sometimes obliques wasapproximately constant and spanned the whole range of dis-tances between 50 and 450 �m from the soma. In addition, theamount of current injected in the trunk to allow transmission ofthe spikes from the sometimes trees showed no significantdependence on the distance from the soma (Fig. 7C, LI, R2 �0.01, P � 0.6). The never and always categories completelydisappeared using KA distributions C1 and C3, respectively
(Fig. 7B, middle and bottom), suggesting an interesting role forlocal KA to gate the propagation of APs out of an oblique.Although Ca2�-dependent potassium conductances are notincluded in our model, recent work attributes a specific role forSK-type (in addition to A-type) K� channels in regulatingspike initiation and conduction between branches of pyramidalcell dendrites (Cai et al. 2004). Our results specifically confirmand highlight the role of KA in determining spike propertieswith respect to conduction past branch points.
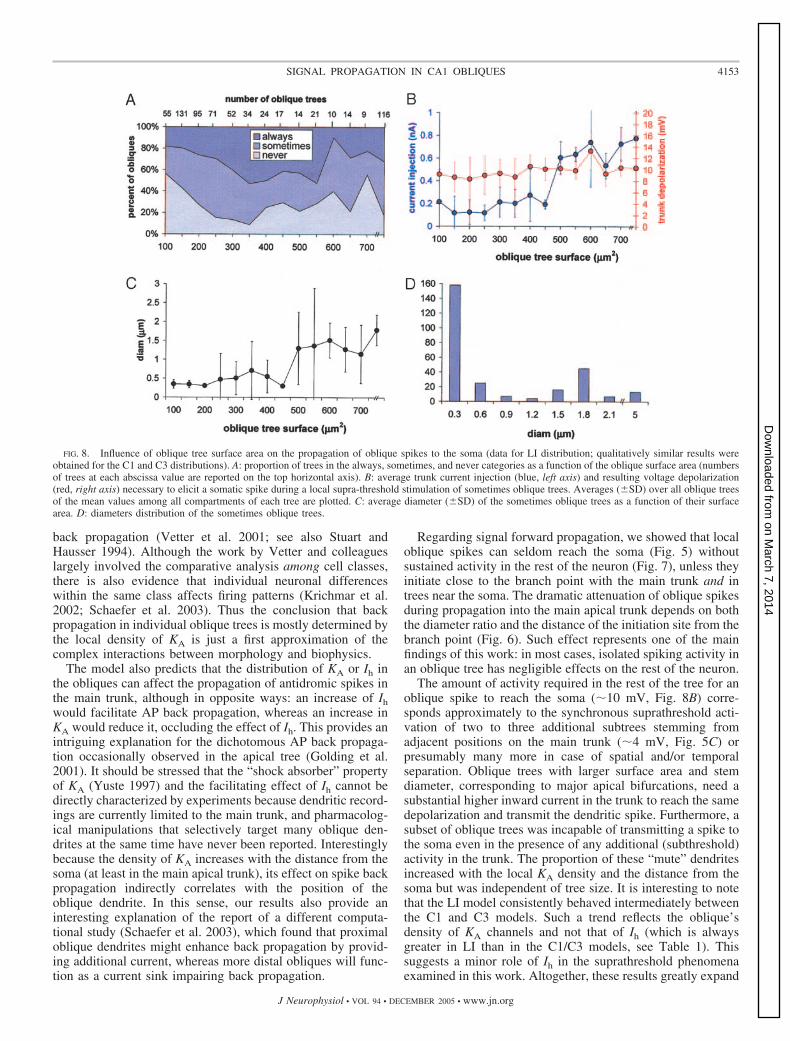
The simulation findings were also analyzed as a function ofthe size of the oblique tree. The proportion of the always,sometimes, and never trees was rather independent of theirsurface area (Fig. 8A). However, the necessary amount of trunkcurrent injection to allow spike transmission from the some-times trees showed a sudden transition in the 450- to 500-�m2
range (Fig. 8B). In particular, smaller trees (�450 �m2, n �194) required 0.15 � 0.14 (SD) nA, whereas larger trees(�450 �m2, n � 81) required 0.73 � 0.25 nA (P � 1010,
FIG. 6. The ratio between the peak membrane potential inthe oblique and that reached in the trunk at the branch pointfollowing a dendritic AP, peak ratio, as a measure of forwardpropagation (data for LI distribution). A: peak ratio as a functionof distance from the soma for obliques stemming out of theapical trunk at 0–50 �m (black symbols), 200–250 �m (whitesymbols), and 350–400 �m (gray symbols) from the soma; forclarity, 3rd-order branches are not shown. For each region, thecompartments of 3 obliques from different neurons are high-lighted in blue, red, and green for varying ratios between thediameter of the oblique and the diameter of the trunk at thebranch point, Dobl/Dtrunk (reported in the insets above eachregion). B: peak ratio along all obliques compartments as afunction of their diameter and distance from the branch point.C: peak ratio for obliques dendrites with diameter �0.3–0.4�m, as a function of Dobl/Dtrunk and distance from the branchpoint on the trunk.
4151SIGNAL PROPAGATION IN CA1 OBLIQUES
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on March 7, 2014
Dow
nloaded from
2-tailed t-test). In contrast, the corresponding depolarizationwas basically independent of the tree surface area (Fig. 8B).These results did not depend on the kind of KA distributions(not shown) but could be correlated with the average value ofthe obliques diameter, which displayed an abrupt and signifi-cant change around the same values of surface area (Fig. 8C,450–500 �m2). The diameter distribution of sometimesoblique trees revealed a clear bimodal distribution with atrough at �1 �m corresponding to the jump observed in Fig.8C. Thus CA1 neurons have a considerable proportion ofsubtrees that, despite their large size, could be classified asoblique (because they are entirely confined within s. radiatum,see METHODS). These large oblique dendrites, which correspondto major bifurcations of the apical tree, required four times asmuch concurrent activity in the trunk than other oblique treesto propagate their spikes to the soma.
D I S C U S S I O N
The functional role of oblique branches in pyramidal cellsmay be key to understanding computational processing in theprincipal neurons of the mammalian cortex (Poirazi et al.2003a,b; Polsky et al. 2004; Schaefer et al. 2003). Morpholog-ical features specific of a given cellular class (Vetter et al.2001) or different distributions of dendritic active channels
(reviewed in Migliore and Shepherd 2002) could be expectedto modulate bidirectional signal propagation between obliquedendrites and the rest of the cell. However, these mechanismsare impractical to explore experimentally, in particular far frombranch points. A realistic computational model is thus a valu-able alternative to investigate how the local oblique propertiescould affect signal propagation. In this work, we demonstratedthat both back and forward propagation can be effectively andindependently modulated in individual oblique branches of thesame neuron. The distribution of KA appears to be pivotal ingating back propagation (Fig. 3), whereas local morphologyplays the lead role in regulating forward propagation (Fig. 6).Although this study mainly focused on the propagation of APs,these results must be considered in the context of previouswork that dealt with spike initiation in cortical pyramidal cells(Andreasen and Lambert 1995, 1998; Golding et al. 1998;Spruston et al. 1995; Turner et al. 1991). Similarly, ourfindings are complementary to several key reports on thepotential effects of a variable Na� spike threshold with dis-tance (Bernard and Johnston 2003), and the importance of Ih inregulating dendritic excitability (Magee 1998; Migliore et al.2004; Poolos et al. 2002).
The first important and novel result of this study is thatspikes antidromically propagating from the soma, while atten-uating continuously in the apical trunk, tend to invade obliquetrees in an all-or-none fashion (Fig. 2). This observation can beexplained on the basis of the experimental evidence suggestingthat dendritic active channels support spike propagation at fullamplitude in CA1 pyramidal cells (Frick et al. 2003). Let usassume, for example, that the initial part of an oblique, justafter the branch point with the main trunk, reaches the thresh-old for spike generation. The local (distance-dependent) KAconductance but also the oblique’s morphology determine theoverall possibility for a spike to propagate down the oblique. Inmost cases, obliques are relatively short and thin (�150 �m,�0.5 �m). This results in significant sealed end effects thatincrease the possibility of a full propagation. In longer or moredistal obliques, the membrane potential will be dampened to asubthreshold value within a short distance from the branchpoint by the “shock adsorber” action of the KA. These effectscause the overall “all-or-none” invasion observed in our modelneurons (Fig. 2A). This phenomenon is essentially independentof local geometry, such as the diameter ratio between obliquestem and main trunk or oblique tree size. In contrast, theprocess is finely regulated by KA. A local threefold increase ofKA, with respect to the levels observed in the main trunk, issufficient to entirely isolate any given oblique from a back-propagating spike. This regulation holds on a tree-by-tree basiswithin the same neuron, such that the same somatic spike couldreach some oblique trees and not others depending on theirlocal concentration of KA. Because the local dendritic activityof KA can be quickly tuned by several enzymatic cascades(e.g., Alkon 1999; Johnston et al. 1999), such gating mecha-nism could enable a highly specific determination of whichsubtree can participate in back propagation-mediated Hebbianplasticity (Magee and Johnston 1997; Markram et al. 1997).
Other studies provided evidence for the important role ofdendritic morphology in regulating the back propagation ofspikes. In particular, the rate of increase in dendritic membranearea (which is related to the number of bifurcations) acttogether with dendritic voltage-gated channels to shape spike
FIG. 7. Relative ability of oblique dendrites to transmit a local spike to thesoma. A: schematic representation of a typical simulation setup: a singledendritic AP was elicited in the presence of a constant depolarizing currentinjection at the branch point in the trunk. B: proportion of oblique trees (totaln � 664) for which a somatic spike could be elicited without additionalstimulation of the trunk (always), those for which a somatic spike could beelicited only in the presence of additional subthreshold stimulation of the trunk(sometimes), and those for which a somatic spike could not be elicited with anyadditional subthreshold stimulation of the trunk (never). Results for thedifferent KA distributions types (LI, C1, and C3) are shown in different panels;sampling of 50 �m were used �500 �m, with oblique trees at 500–800 �mfrom soma (n � 8) grouped together. The total number of trees at each valueis shown in the top panel. C: average (�SD) value of the (subthreshold)current injection in the trunk necessary to elicit a somatic spike duringsupra-threshold stimulation of an oblique tree in the sometimes group usingdifferent KA distributions.
4152 M. MIGLIORE, M. FERRANTE, AND G. A. ASCOLI
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on March 7, 2014
Dow
nloaded from
back propagation (Vetter et al. 2001; see also Stuart andHausser 1994). Although the work by Vetter and colleagueslargely involved the comparative analysis among cell classes,there is also evidence that individual neuronal differenceswithin the same class affects firing patterns (Krichmar et al.2002; Schaefer et al. 2003). Thus the conclusion that backpropagation in individual oblique trees is mostly determined bythe local density of KA is just a first approximation of thecomplex interactions between morphology and biophysics.
The model also predicts that the distribution of KA or Ih inthe obliques can affect the propagation of antidromic spikes inthe main trunk, although in opposite ways: an increase of Ihwould facilitate AP back propagation, whereas an increase inKA would reduce it, occluding the effect of Ih. This provides anintriguing explanation for the dichotomous AP back propaga-tion occasionally observed in the apical tree (Golding et al.2001). It should be stressed that the “shock absorber” propertyof KA (Yuste 1997) and the facilitating effect of Ih cannot bedirectly characterized by experiments because dendritic record-ings are currently limited to the main trunk, and pharmacolog-ical manipulations that selectively target many oblique den-drites at the same time have never been reported. Interestinglybecause the density of KA increases with the distance from thesoma (at least in the main apical trunk), its effect on spike backpropagation indirectly correlates with the position of theoblique dendrite. In this sense, our results also provide aninteresting explanation of the report of a different computa-tional study (Schaefer et al. 2003), which found that proximaloblique dendrites might enhance back propagation by provid-ing additional current, whereas more distal obliques will func-tion as a current sink impairing back propagation.
Regarding signal forward propagation, we showed that localoblique spikes can seldom reach the soma (Fig. 5) withoutsustained activity in the rest of the neuron (Fig. 7), unless theyinitiate close to the branch point with the main trunk and intrees near the soma. The dramatic attenuation of oblique spikesduring propagation into the main apical trunk depends on boththe diameter ratio and the distance of the initiation site from thebranch point (Fig. 6). Such effect represents one of the mainfindings of this work: in most cases, isolated spiking activity inan oblique tree has negligible effects on the rest of the neuron.
The amount of activity required in the rest of the tree for anoblique spike to reach the soma (�10 mV, Fig. 8B) corre-sponds approximately to the synchronous suprathreshold acti-vation of two to three additional subtrees stemming fromadjacent positions on the main trunk (�4 mV, Fig. 5C) orpresumably many more in case of spatial and/or temporalseparation. Oblique trees with larger surface area and stemdiameter, corresponding to major apical bifurcations, need asubstantial higher inward current in the trunk to reach the samedepolarization and transmit the dendritic spike. Furthermore, asubset of oblique trees was incapable of transmitting a spike tothe soma even in the presence of any additional (subthreshold)activity in the trunk. The proportion of these “mute” dendritesincreased with the local KA density and the distance from thesoma but was independent of tree size. It is interesting to notethat the LI model consistently behaved intermediately betweenthe C1 and C3 models. Such a trend reflects the oblique’sdensity of KA channels and not that of Ih (which is alwaysgreater in LI than in the C1/C3 models, see Table 1). Thissuggests a minor role of Ih in the suprathreshold phenomenaexamined in this work. Altogether, these results greatly expand
FIG. 8. Influence of oblique tree surface area on the propagation of oblique spikes to the soma (data for LI distribution; qualitatively similar results wereobtained for the C1 and C3 distributions). A: proportion of trees in the always, sometimes, and never categories as a function of the oblique surface area (numbersof trees at each abscissa value are reported on the top horizontal axis). B: average trunk current injection (blue, left axis) and resulting voltage depolarization(red, right axis) necessary to elicit a somatic spike during a local supra-threshold stimulation of sometimes oblique trees. Averages (�SD) over all oblique treesof the mean values among all compartments of each tree are plotted. C: average diameter (�SD) of the sometimes oblique trees as a function of their surfacearea. D: diameters distribution of the sometimes oblique trees.
4153SIGNAL PROPAGATION IN CA1 OBLIQUES
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on March 7, 2014
Dow
nloaded from

on the recent experimental work on the propagation of locallyinitiated dendritic spikes in the main trunk of CA1 pyramidalcells (Gasparini et al. 2004).
The effective independence of oblique trees from each other,with respect to their spiking behavior, enables the independentimplementation of “localized” Hebbian plasticity (Alkon1999). A synapse may be strengthened when the presynapticactivity coincides not necessarily with the somatic spiking ofthe postsynaptic neuron but possibly just with a locally initi-ated oblique spike. Such form of synaptic plasticity could beongoing continuously and in parallel in many oblique branchesduring “silent” periods (lack of somatic spiking activity) of aCA1 pyramidal cell.
In summary, signal propagation in CA1 pyramidal cells isshaped by the interaction of unique morphological and bio-physical features. The dendritic distributions of all main activeconductances are known to be qualitatively and quantitativelydifferent in various neuronal classes (Migliore and Shepherd2002). Earlier work showed that the dendritic morphology ofdifferent classes can affect both back and forward propagationof action potentials and that different ratios of Na� and K�
conductances can partially compensate for the morphologicaldifferences (Vetter et al. 2001). Here we demonstrate thatsignal propagation varies within the homogenous group ofCA1 pyramidal cells and in fact within individual neurons.
The ability of specific morphological and biophysical pa-rameters to isolate and independently modulate subcellularcomponents of CA1 pyramidal neurons provides these cellswith peculiar computational properties. At the level of synapticintegration, the computational power of the apical tree isenhanced by a “double-step” nonlinear process, in whichindividual oblique trees undergo parallel electrogenic thresholddiscriminations before the resulting signals, brought subthresh-old into the main trunk, are integrated anew in the soma(Poirazi et al. 2003a,b). An even more important functionalconsequence of the oblique properties examined in this work,however, is the potential to independently modulate synapticefficacies in every tree in parallel. In an insightful theoreticalstudy, Poirazi and Mel (2001) analyzed the increased memorycapacity of a neuronal architecture with independent nonlinearunits. The present work suggests a physiologically plausibleimplementations of these mechanisms by means of the inde-pendent and parallel modulation of back and forward propa-gation in each oblique tree, based on developmental mecha-nisms (local morphological features) and dynamic rules (reg-ulation of active conductances).
A C K N O W L E D G M E N T S
We thank the Yale University Computer Science Department (New Haven,CT) and the CINECA consortium (Bologna, Italy) for granting access to theirparallel systems.
G R A N T S
This research was supported by National Institutes of Health R01 GrantNS-39600 jointly funded by National Institute of Neurological Disorders andStroke, National Institute of Mental Health, and National Science Foundationunder the Human Brain Project and AG-025633 as part of the NSF/NationalInstitutes of Health Collaborative Research in Computational NeuroscienceProgram.
R E F E R E N C E S
Alkon DL. Ionic conductance determinants of synaptic memory nets and theirimplications for Alzheimer’s disease. J Neurosci Res 58: 24–32, 1999.
Ambros-Ingerson J and Holmes WR. Analysis and comparison of morpho-logical reconstructions of hippocampal field CA1 pyramidal cells. Hip-pocampus 15: 302–315, 2005.
Andrasfalvy BK and Magee JC. Distance-dependent increase in AMPAreceptor number in the dendrites of adult hippocampal CA1 pyramidalneurons. J Neurosci 21: 9151–9159, 2001.
Andreasen M and Lambert JD. Regenerative properties of pyramidal celldendrites in area CA1 of the rat hippocampus. J Physiol 483: 421–441,1995.
Andreasen M and Lambert JD. Factors determining the efficacy of distalexcitatory synapses in rat hippocampal CA1 pyramidal neurons. J Physiol507: 441–462, 1998.
Ascoli GA, Krichmar JL, Nasuto SJ, and Senft SL. Generation, descriptionand storage of dendritic morphology data. Philos Trans R Soc Lond B BiolSci 356: 1131–1145, 2001.
Bannister NJ and Larkman AU. Dendritic morphology of CA1 pyramidalneurones from the rat hippocampus. I. Branching patterns. J Comp Neurol360: 150–160, 1995.
Bekkers JM. Distribution of slow AHP channels on hippocampal CA1pyramidal neurons. J Neurophysiol 83: 1756–1759, 2000.
Bernard C and Johnston D. Distance-dependent modifiable threshold foraction potential back-propagation in hippocampal dendrites. J Neurophysiol90: 1807–1816, 2003.
Brown KM, Donohue DE, D’Alessandro G, and Ascoli GA. A cross-platform freeware tool for digital reconstruction of neuronal arborizationsfrom image stacks. Neuroinformatics In Press.
Cai X, Liang CW, Muralidharan S, Kao JP, Tang CM, and ThompsonSM. Unique roles of SK and Kv4.2 potassium channels in dendriticintegration. Neuron 44: 351–364, 2004.
Carnevale NT, Tsai KY, Claiborne BJ, and Brown TH. Comparativeelectrotonic analysis of three classes of rat hippocampal neurons. J Neuro-physiol 78: 703–720, 1997.
Cash S and Yuste R. Linear summation of excitatory inputs by CA1pyramidal neurons. Neuron 22: 383–394, 1999.
Chen X and Johnston D. Properties of single voltage-dependent K� channelsin dendrites of CA1 pyramidal neurones of rat hippocampus. J Physiol 559:187–203, 2004.
Davison AP, Morse TM, Migliore M, Shepherd GM, and Hines ML.Semi-automated population of an online database of neuronal models(ModelDB) with citation information, using PubMed for validation. Neu-roinformatics 2: 327–332, 2004.
Donohue DE, Scorcioni R, and Ascoli GA. Generation and description ofneuronal morphology using L-Neuron: a case study. In: ComputationalNeuroanatomy: Principles and Methods, edited by Ascoli GA. Totowa, NJ:Humana, 2002, p. 49–70.
Frick A, Magee J, and Johnston D. LTP is accompanied by an enhancedlocal excitability of pyramidal neuron dendrites. Nat Neurosci 7: 126–135,2004.
Frick A, Magee J, Koester HJ, Migliore M, and Johnston D. Normalizationof Ca2� signals by small oblique dendrites of CA1 pyramidal neurons.J Neurosci 23: 3243–3250, 2003.
Gasparini S, Migliore M, and Magee JC. On the initiation and propagationof dendritic spikes in CA1 pyramidal neurons. J Neurosci 24: 11046–11056,2004.
Golding NL, Jung HY, Mickus T, and Spruston N. Dendritic calcium spikeinitiation and repolarization are controlled by distinct potassium channelsubtypes in CA1 pyramidal neurons. J Neurosci 19: 8789–8798, 1999.
Golding NL, Kath WL, and Spruston N. Dichotomy of action-potentialbackpropagation in CA1 pyramidal neuron dendrites. J Neurophysiol 86:2998–3010, 2001.
Golding NL and Spruston N. Dendritic sodium spikes are variable triggers ofaxonal action potentials in hippocampal CA1 pyramidal neurons. Neuron21: 1189–1200, 1998.
Hines M and Carnevale T. The NEURON simulation environment. NeuralComp 9: 1179–1209, 1997.
Hoffman DA, Magee JC, Colbert CM, and Johnston D. K� channelregulation of signal propagation in dendrites of hippocampal pyramidalneurons. Nature 387: 869–875, 1997.
Ishizuka N, Cowan WM, and Amaral DG. A quantitative analysis of thedendritic organization of pyramidal cells in the rat hippocampus. J CompNeurol 362: 17–45, 1995.
Johnston D, Hoffman DA, Colbert CM, and Magee JC. Regulation ofback-propagating action potentials in hippocampal neurons. Curr OpinNeurobiol 9: 288–292, 1999.
4154 M. MIGLIORE, M. FERRANTE, AND G. A. ASCOLI
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on March 7, 2014
Dow
nloaded from

Kasuga A, Enoki R, Hashimoto Y, Akiyama H, Kawamura Y, Inoue M,Kudo Y, and Miyakawa H. Optical detection of dendritic spike initiationin hippocampal CA1 pyramidal neurons. Neuroscience 118: 899–907, 2003.
Krichmar JL, Nasuto SJ, Scorcioni R, Washington SD, and Ascoli GA.Effects of dendritic morphology on CA3 pyramidal cell electrophysiology:a simulation study. Brain Res 941: 11–28, 2002.
Lazarewicz MT, Migliore M, and Ascoli GA. A new bursting model of CA3pyramidal cell physiology suggests multiple locations for spike initiation.Biosystems 67: 129–137, 2002.
Liu G. Local structural balance and functional interaction of excitatory andinhibitory synapses in hippocampal dendrites. Nat Neurosci 7: 373–379,2004.
Magee JC. Dendritic hyperpolarization-activated currents modify the integra-tive properties of hippocampal CA1 pyramidal neurons. J Neurosci 18:7613–7624, 1998.
Magee JC and Carruth M. Dendritic voltage-gated ion channels regulate theaction potential firing mode of hippocampal CA1 pyramidal neurons. J Neu-rophysiol 82: 1895–1901, 1999.
Magee JC and Johnston D. Characterization of single voltage-gated Na� andCa2� channels in apical dendrites of rat CA1 pyramidal neurons. J Physiol487: 67–90, 1995.
Magee JC and Johnston D. A synaptically controlled, associative signal forHebbian plasticity in hippocampal neurons. Science 275: 209–213, 1997.
Markram H, Lubke J, Frotscher M, and Sakmann B. Regulation ofsynaptic efficacy by coincidence of postsynaptic APs and EPSPs. Science275: 213–215, 1997.
Megias M, Emri Z, Freund TF, and Gulyas AI. Total number and distri-bution of inhibitory and excitatory synapses on hippocampal CA1 pyramidalcells. Neuroscience 102: 527–540, 2001.
Migliore M. On the integration of subthreshold inputs from perforant path andSchaffer collaterals in hippocampal CA1 neurons. J Comput Neurosci 14:185–192, 2003.
Migliore M, Hoffman DA, Magee JC, and Johnston D. Role of an A-typeK� conductance in the back-propagation of action potentials in the dendritesof hippocampal pyramidal neurons. J Comput Neurosci 7: 5–16, 1999.
Migliore M, Morse TM, Davison AP, Marenco L, Shepherd GM, andHines ML. ModelDB: making models publicly accessible to support com-putational neuroscience. Neuroinformatics 1: 135–139, 2003.
Migliore M, Messineo L, and Ferrante M. Dendritic Ih selectively blockstemporal summation of unsynchronized sistal inputs in CA1 pyramidalneurons. J Comput Neurosci 16: 5–13, 2004.
Migliore M and Shepherd GM. Emerging rules for the distributions of activedendritic conductances. Nature Rev Neurosci 3: 362–370, 2002.
Nakamura T, Lasser-Ross N, Nakamura K, and Ross WN. Spatial segre-gation and interaction of calcium signalling mechanisms in rat hippocampalCA1 pyramidal neurons. J Physiol 543: 465–480, 2002.
Otmakhova NA, Otmakhov N, and Lisman JE. Pathway-specific propertiesof AMPA and NMDA-mediated transmission in CA1 hippocampal pyrami-dal cells. J Neurosci 22:1199–1207, 2002.
Poirazi P, Brannon T, and Mel BW. Arithmetic of subthreshold synapticsummation in a model CA1 pyramidal cell. Neuron 37: 977–987, 2003a.
Poirazi P, Brannon T, and Mel BW. Pyramidal neuron as two-layer neuralnetwork. Neuron 37: 989–999, 2003b.
Poirazi P and Mel BW. Impact of active dendrites and structural plasticity onthe memory capacity of neural tissue. Neuron 29: 779–796, 2001.
Polsky A, Mel BW, and Schiller J. Computational subunits in thin dendritesof pyramidal cells. Nat Neurosci 7: 621–627, 2004.
Poolos NP, Migliore M, and Johnston D. Pharmacological upregulation ofh-channels selectively reduces the excitability of pyramidal neuron den-drites. Nat Neurosci 5: 767–774, 2002.
Pyapali GK, Sik A, Penttonen M, Buzsaki G, and Turner DA. Dendriticproperties of hippocampal CA1 pyramidal neurons in the rat: intracellularstaining in vivo and in vitro. J Comp Neurol 391: 335–352, 1998.
Schaefer AT, Larkum ME, Sakmann B, and Roth A. Coincidence detectionin pyramidal neurons is tuned by their dendritic branching pattern. J Neu-rophysiol 89: 3143–3154, 2003.
Scorcioni R, Lazarewicz MT, and Ascoli GA. Quantitative morphometry ofhippocampal pyramidal cells: differences between anatomical classes andreconstructing laboratories. J Comp Neurol 473: 177–193, 2004.
Shao LR, Halvorsrud R, Borg-Graham L, and Storm JF. The role ofBK-type Ca2�-dependent K� channels in spike broadening during repetitivefiring in rat hippocampal pyramidal cells. J Physiol 521:135–146, 1999.
Spruston N, Jonas P, and Sakmann B. Dendritic glutamate receptor channelsin rat hippocampal CA3 and CA1 pyramidal neurons. J Physiol 482:325–352, 1995.
Stuart G and Hausser M. Initiation and spread of sodium action potentials incerebellar Purkinje cells. Neuron 13: 703–712, 1994.
Szilagyi T and De Schutter E. Effects of variability in anatomical recon-struction techniques on models of synaptic integration by dendrites: acomparison of three Internet archives. Eur J Neurosci 19: 1257–1266, 2004.
Trommald M, Jensen V, and Andersen P. Analysis of dendritic spines in ratCA1 pyramidal cells intracellularly filled with a fluorescent dye. J CompNeurol 353: 260–274, 1995.
Turner RW, Meyers DE, Richardson TL, and Barker JL. The site forinitiation of action potential discharge over the somatodendritic axis of rathippocampal CA1 pyramidal neurons. J Neurosci 11: 2270–2280, 1991.
Vetter P, Roth A, and Hausser M. Propagation of action potentials indendrites depends on dendritic morphology. J Neurophysiol 85: 926–937,2001.
Watanabe S, Hoffman DA, Migliore M, and Johnston D. Dendritic K�
channels contribute to spike-timing dependent long-term potentiation inhippocampal pyramidal neurons. Proc Natl Acad Sci USA 99: 8366–8371,2002.
Yuste R. Potassium channels. Dendritic shock absorbers. Nature 387: 851–853, 1997.
Zamanillo D, Sprengel R, Hvalby O, Jensen V, Burnashev N, Rozov A,Kaiser KM, Koster HJ, Borchardt T, Worley P, Lubke J, Frotscher M,Kelly PH, Sommer B, Andersen P, Seeburg PH, and Sakmann B.Importance of AMPA receptors for hippocampal synaptic plasticity but notfor spatial learning. Science 284: 1805–1811, 1999.
4155SIGNAL PROPAGATION IN CA1 OBLIQUES
J Neurophysiol • VOL 94 • DECEMBER 2005 • www.jn.org
on March 7, 2014
Dow
nloaded from




















