
Screening of pigmented Bacillus aquimaris SH6 from ... - Anabio
16
ORIGINAL ARTICLE Screening of pigmented Bacillus aquimaris SH6 from the intestinal tracts of shrimp to develop a novel feed supplement for shrimp H.T. Ngo 1,* , T.T.N. Nguyen 1,* , Q.M. Nguyen 2,* , A.V. Tran 2,* , H.T.V. Do 3 , A.H. Nguyen 4 , T.-N. Phan 1 and A.T.V. Nguyen 1 1 Key Laboratory of Enzyme and Protein Technology, VNU University of Science, Hanoi, Vietnam 2 High School for Gifted Students, VNU University of Science, Hanoi, Vietnam 3 Faculty of Chemistry, VNU University of Science, Hanoi, Vietnam 4 ANABIO Research & Development JSC, Hanoi, Vietnam Keywords Bacillus, carotenoid, gastrointestinal tract, pigmented, shrimp, supplement. Correspondence Anh Thi Van Nguyen, Key Laboratory of Enzyme and Protein Technology, VNU University of Science, 334 Nguyen Trai, Hanoi, Vietnam. E-mail: [email protected] * These authors contribute to this work equally. 2016/0625: received 24 March 2016, revised 24 July 2016 and accepted 16 August 2016 doi:10.1111/jam.13274 Abstract Aims: To develop a novel feed supplement for shrimp using pigmented spore- forming bacterial strains isolated from their gastrointestinal tracts. Methods and Results: Eight pigmented Bacillus strains were selected from the isolates based on high production of heat-stable spores, typical UV-Vis spectra of produced carotenoids (400–550 nm), and free radical scavenging activity of their extracts. Of the eight strains, the red-orange pigmented Bacillus aquimaris SH6 was selected because it showed the highest abundance in shrimp guts (70% population). Whiteleg shrimp (n = 30 per group) fed with SH6 spores, at >3 9 10 6 CFU g 1 pellet for 4 weeks had redder colour (score of 21–23 vs 20–22), 27-fold higher astaxanthin level (069 vs 025 lgg 1 shrimp), 34% higher weight gain (718 vs 532 g shrimp 1 ), and 85% higher phenoloxidase activity (OD490 = 0265 vs 0143) than shrimp in the control group. Conclusions: The result supports the potential use of B. aquimaris SH6 as a feed supplement for promoting the colourization and weight gain, and for enhancing innate immunity of whiteleg shrimp. Significance and Impact of the Study: This study demonstrates that carotenoids produced by B. aquimaris SH6 can be successfully absorbed and converted to astaxanthin in whiteleg shrimp. Introduction The need for sustainable shrimp aquaculture drives stud- ies on the use of probiotics in growing shrimp, such as whiteleg shrimp (Litopenaeus vannamei) or black-tiger shrimp (Litopenaeus monodon). Probiotics can have a beneficial effect on the digestive processes in shrimp because of their ability to synthesize extracellular enzymes such as proteases, amylases and lipases and to provide growth factors such as vitamins, carotenoids, fatty acids and amino acids (Araya et al. 2002). Therefore, nutrients are absorbed more efficiently when the feed is supple- mented with probiotics. The commercial value of shrimps is predominately based on the visual appeal of their body colour. The red carotenoid, astaxanthin, has been identified as the pre- dominant pigment in Penaeus shrimp (Yamada et al. 1990). However, these shrimp are unable to produce astaxanthin de novo; only plants and protists (bacteria, algae and fungi) are capable of synthesizing carotenoids (Schmidt-Dannert 2000). Therefore, astaxanthin must be available in either their native habitat or manufactured diet to meet metabolic nutritional requirements. The red colour of cooked shrimp is produced by the release of the individual carotenoid prosthetic group (astaxanthin) from the carotenoproteins when denatured by the heat of cooking. Lack of dietary astaxanthin in cultured shrimp has been shown to cause ‘blue colour syndrome’ (Latscha 1989). Supplementing shrimp feed with synthe- sized red carotenoid astaxanthin is common to improve Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology 1357 Journal of Applied Microbiology ISSN 1364-5072
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Screening of pigmented Bacillus aquimaris SH6 from ... - Anabio

ORIGINAL ARTICLE
Screening of pigmented Bacillus aquimaris SH6 from theintestinal tracts of shrimp to develop a novel feedsupplement for shrimpH.T. Ngo1,*, T.T.N. Nguyen1,*, Q.M. Nguyen2,*, A.V. Tran2,*, H.T.V. Do3, A.H. Nguyen4, T.-N. Phan1
and A.T.V. Nguyen1
1 Key Laboratory of Enzyme and Protein Technology, VNU University of Science, Hanoi, Vietnam
2 High School for Gifted Students, VNU University of Science, Hanoi, Vietnam
3 Faculty of Chemistry, VNU University of Science, Hanoi, Vietnam
4 ANABIO Research & Development JSC, Hanoi, Vietnam
Keywords
Bacillus, carotenoid, gastrointestinal tract,
pigmented, shrimp, supplement.
Correspondence
Anh Thi Van Nguyen, Key Laboratory of
Enzyme and Protein Technology, VNU
University of Science, 334 Nguyen Trai,
Hanoi, Vietnam.
E-mail: [email protected]
*These authors contribute to this work
equally.
2016/0625: received 24 March 2016, revised
24 July 2016 and accepted 16 August 2016
doi:10.1111/jam.13274
Abstract
Aims: To develop a novel feed supplement for shrimp using pigmented spore-
forming bacterial strains isolated from their gastrointestinal tracts.
Methods and Results: Eight pigmented Bacillus strains were selected from the
isolates based on high production of heat-stable spores, typical UV-Vis spectra
of produced carotenoids (400–550 nm), and free radical scavenging activity of
their extracts. Of the eight strains, the red-orange pigmented Bacillus aquimaris
SH6 was selected because it showed the highest abundance in shrimp guts
(70% population). Whiteleg shrimp (n = 30 per group) fed with SH6 spores,
at >3 9 106 CFU g�1 pellet for 4 weeks had redder colour (score of 21–23 vs
20–22), 2�7-fold higher astaxanthin level (0�69 vs 0�25 lg g�1 shrimp), 34%
higher weight gain (7�18 vs 5�32 g shrimp�1), and 85% higher phenoloxidase
activity (OD490 = 0�265 vs 0�143) than shrimp in the control group.
Conclusions: The result supports the potential use of B. aquimaris SH6 as a
feed supplement for promoting the colourization and weight gain, and for
enhancing innate immunity of whiteleg shrimp.
Significance and Impact of the Study: This study demonstrates that
carotenoids produced by B. aquimaris SH6 can be successfully absorbed and
converted to astaxanthin in whiteleg shrimp.
Introduction
The need for sustainable shrimp aquaculture drives stud-
ies on the use of probiotics in growing shrimp, such as
whiteleg shrimp (Litopenaeus vannamei) or black-tiger
shrimp (Litopenaeus monodon). Probiotics can have a
beneficial effect on the digestive processes in shrimp
because of their ability to synthesize extracellular enzymes
such as proteases, amylases and lipases and to provide
growth factors such as vitamins, carotenoids, fatty acids
and amino acids (Araya et al. 2002). Therefore, nutrients
are absorbed more efficiently when the feed is supple-
mented with probiotics.
The commercial value of shrimps is predominately
based on the visual appeal of their body colour. The red
carotenoid, astaxanthin, has been identified as the pre-
dominant pigment in Penaeus shrimp (Yamada et al.
1990). However, these shrimp are unable to produce
astaxanthin de novo; only plants and protists (bacteria,
algae and fungi) are capable of synthesizing carotenoids
(Schmidt-Dannert 2000). Therefore, astaxanthin must be
available in either their native habitat or manufactured
diet to meet metabolic nutritional requirements. The red
colour of cooked shrimp is produced by the release of
the individual carotenoid prosthetic group (astaxanthin)
from the carotenoproteins when denatured by the heat of
cooking. Lack of dietary astaxanthin in cultured
shrimp has been shown to cause ‘blue colour syndrome’
(Latscha 1989). Supplementing shrimp feed with synthe-
sized red carotenoid astaxanthin is common to improve
Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology 1357
Journal of Applied Microbiology ISSN 1364-5072

colourization of shrimp. D’Abramo et al. (1983) have
demonstrated that pure carotenoids such as b-carotene,echinenone and canthaxanthin are converted to astaxan-
thin in cultured lobsters and that the level of pigmenta-
tion produced by these biosynthetic precursors is related
to the proximity to the astaxanthin end product. In
recent years, many research groups have attempted to
isolate pigmented bacteria that can produce carotenoids
to develop novel natural food and feed supplements
(Pane et al. 1996; Duc et al. 2006; Khaneja et al. 2009; Sy
et al. 2013). Yellow, orange, red and pink Bacillus species
have been isolated from seawater, sand and soil. For
example, Pane and his colleagues have identified a red-
pigmented Bacillus firmus strain producing astaxanthin,
which was isolated from a seawater rock pool. Their
result suggests the potential use of this bacterium in
aquaculture and in the pharmaceutical field (Pane et al.
1996). Yoon and his colleagues have isolated two yellow-
pigmented strains from seawater and identified them as
new species Bacillus aquimaris and Bacillus marisflavi
(Yoon et al. 2003). In another study by Suresh et al. an
arsenic-resistant yellowish-orange pigmented bacterium
was isolated from a sand sample obtained from an
arsenic-contaminated aquifer. The strain was identified as
a new species named Bacillus indicus (Suresh et al. 2004).
Khaneja et al. (2009) isolated several other carotenoid-
producing Bacillus species from seawater, soil and fer-
mented rice condiment. These included B. marisflavi,
Bacillus cibi and Bacillus altitudinis strains producing yel-
low and yellow-orange pigments; B. aquimaris and Bacil-
lus pumilus strains producing orange and orange-red
pigments; and some B. firmus strains producing pink and
deep pink pigments. Carotenoids produced by these
strains were determined to have absorption maxima at
455, 467 and 492 nm, corresponding to the visible col-
ours yellow, orange and pink respectively (Khaneja et al.
2009). In other studies, yellow-pigmented B. indicus
HU36 of human intestinal origin and red-pigmented
B. firmus GB1 isolated from soil have been investigated
for their ability to produce natural antioxidant carote-
noids (Duc et al. 2006; Hong et al. 2008; Khaneja et al.
2009; Cutting 2011); this was followed by in vivo studies
on the bioaccessibility and bioavailability of these carote-
noids, which were found to be even better than those of
commercial available synthesized carotenoids (Sy et al.
2013).
Many protocols on the isolation and characterization
of pigmented Bacillus species from seawater, shrimp
ponds, and human faeces have been implemented (Yoon
et al. 2003; Duc et al. 2006; Khaneja et al. 2009); how-
ever, the isolation of pigmented Bacillus species from gas-
trointestinal tracts (GITs) of shrimp, to develop
probiotics for use as feed supplements for shrimp
themselves, has not been reported. This is especially
important as it is generally recommended that probiotic
strains should be isolated from the GITs of their host;
such strains are thought to have the best chance of sur-
viving and colonizing the intestine, allowing beneficial
microbial flora to thrive (Dunne et al. 2001). Therefore,
we investigated whether reintroduction of pigmented
Bacillus sp. strains, isolated from the shrimp gut, back
into shrimp, would create beneficial microbiota. Thus, we
attempted to isolate pigmented strains of Bacillus species
directly from the GITs of shrimp to screen for the pro-
duction of high levels of antioxidant carotenoids and for
abundance in the shrimp gut. Spores of the selected
strain were produced for laboratory-20 scale blind trials
in whiteleg shrimp to evaluate shrimp colourization (col-
our score and astaxanthin level), weight gain and phe-
noloxidase activity, reflecting innate immune status.
Materials and methods
Reference strains
Yellow-pigmented B. indicus HU36 and Bacillus subtilis
HU58 isolated from human faeces (Duc et al. 2006; Tam
et al. 2006) and nonpigmented laboratory B. subtilis
PY79 was used as the reference strain in most experi-
ments described here; Bacillis cereus ATCC 10876 was
used as a control for the haemolysis test.
Preparation of intestinal samples and isolation of
pigmented Bacillus colonies
Twelve natural shrimp of different species including
L. vannamei and L. monodon were collected from rivers
and coastal regions (Binh Dai, Phu An, Thanh Thuy) in
Ben Tre Province, Vietnam. Each shrimp GIT was pre-
pared to collect the mucosa in 0�9% NaCl and then vig-
orously resuspended by vortexing until a homogenous
suspension was obtained. To recover heat-resistant
spores, 1 ml of the suspension was heated at 65°C for
20 min and serial dilutions were made with 0�9% NaCl
before plating on tryptone soy agar (TSA; Oxoid, Hamp-
shire, UK) and incubating for 1 day at 37°C to obtain
individual colonies. For each sample, different pigmented
colonies were picked randomly and transferred to new
Difco Sporulation Medium (DSM) agar plates, and then
checked by microscopy for morphological traits and pres-
ence of spores.
General methods
Each isolate was grown on DSM agar or in DSM broth
(Oxoid) for 48 h and assessed for sporulation percentage
Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology1358
Bacillus from shrimp intestine H.T. Ngo et al.

by determining the titre of heat-resistant cells (from 40°Cto 80°C, 20 min) vs the total viable cell count. Bacterial
growth was monitored in tryptone soy broth (TSB;
Oxoid) at 30°C, or at 37°C; absorbance of cell cultures
was measured after 24 h at 600 nm (OD600) using a
spectrophotometer Biomate 3 (Thermo Scientific, Wal-
tham, MA). Characterization of Bacillus strains using
microbiological and biochemical methods was performed
following Bergey’s Manual of Systematic Bacteriology
(Vos et al. 2009). The shapes of bacteria and positions of
spores of selected strains were observed under a conven-
tional microscope Primo Star (Carl Zeiss, Oberkochen,
Germany). To determine the ability of bacterial isolates
to grow under aerobic or anaerobic conditions, strains
were streaked into test tubes of thioglycolate semi-solid
media with 0�3% agar and then incubated at 37°C for
24 h (Evans and Kloos 1972). Metabolism of glucose and
citrate was assessed using a conventional Voges-Proskauer
(VP) reaction and citrate utilization test. To test for
haemolysis, isolates were inoculated on tryptose agar con-
taining sheep blood and incubated for 24 h at 37°C to
observe haemolysis zones. To evaluate antibiotic resis-
tance and sensitivity, antibiograms were obtained for each
strain using the radial diffusion method, according to the
recommendations of the National Committee for Clinical
Laboratory Standards (NCCLS 1997). Inhibition zones
were measured for 14 common antibiotics as listed in
Table 1. Amylase activity was assayed using the starch
hydrolysis test on agar plates containing 1% soluble
starch. Proteolytic activity was examined with casein and
gelatin hydrolysis tests on agar plates.
16S rRNA sequence analysis
Total genomic DNA was extracted from Bacillus species
overnight LB cultures. The genomic DNA was used as
template for PCR and partial 16S rRNA sequencing of a
PCR-amplified 1500 bp fragment using primers 27F: 50-AGAGTTTGATCMTGGCTCAG-30 and 1527R: 50-AAAGGAGGTGATCCAGCC-30 (Khaneja et al. 2009; Nguyen
et al. 2015). The resultant partial 16S rRNA sequences
were assembled and aligned with APE� software (The
University of Utah, Salt Lake, UT). Obtained sequences
were compared to sequences in the GenBank nonredun-
dant nucleotide database by BLAST analysis. The closest
species were identified and the per cent identity was
recorded. Phylogeny was inferred from aligned nucleotide
sequences of the 16S rRNA genes using MEGA ver. 6 soft-
ware (Tamura et al. 2013). Evolutionary relationships of
the Bacillus strains were estimated using the neighbour-
joining method (Saitou and Nei 1987) with 1000 boot-
strap replicates. Evolutionary distance was computed
using the p-distance model and are given in units of
number of base differences per site. Branch lengths are
proportional to the amount of evolutionary change.
Pigment extraction and analysis
Pigment extraction and analysis from Bacillus isolates
were performed following a previously reported method
(Khaneja et al. 2009). Extracts (200-ll) from 1�2 mg dry
weight of cells or 1-ml cultures were prepared to measure
absorbance values at optimal peaks, such as OD460 for
the SH8 and SH20 strains, OD470 for the SH6 and SH14
strains, or OD490 for the SH1, SH4, and SH5 strains.
Concentrations of carotenoids in the extracts were deter-
mined from the standard curve of astaxanthin at concen-
trations ranging from 1�05 to 16�75 lmol l�1. High
performance liquid chromatography (HPLC) analysis of
the selected SH6 strain was performed on 20 AD-UFLC
system (Shimadzu, Kyoto, Japan) consisting of a photodi-
ode array (PDA) detector using a reverse-phase (RP) C30,
5-lm column (250 mm 9 4�6 mm) coupled to a
C30 guard column (20 mm 9 4�6 mm; Thermo Scien-
tific). The mobile phase and elution conditions were
identical to those described by Khaneja et al. (2009). The
carotenoids were identified by comparison of spectral and
chromatographic characteristics with those published for
reference carotenoids (Britton et al. 2004).
Free radical scavenging activity of pigmented extractions
2,2-Diphenyl-1-picrylhydrazyl (DPPH) antioxidant activ-
ity was measured following a standard method (Sharma
and Bhat 2009). In brief, individual 1�5-ml extracts from
approx. 0�5 g (wet weight) of vegetative cells were incu-
bated with 500 ll of 250 lmol l�1 1,1-diphenyl-2-picryl-
hydrazyl (DPPH; Sigma, St. Louis, MO). The DPPH
absorbance at 517 nm was measured before and after the
reactions. Inhibition level was calculated using following
equation: Inhibition (%) = [(ODcontrol � ODsample)/
ODcontrol] 9 100. In parallel experiments, 1�5 ml of
ascorbic acid (6�25–25 lmol l�1) was added as a quanti-
tative standard. The experiment was repeated three times.
Production of pigmented spores
Growth of bacterial strains and spore formation were
optimized using various media (DSM, LB, TSB), pH (6–9), and temperatures (25–45°C) in a flask. To induce
spore production under the optimized conditions, Bacil-
lus strains were cultured in suitable medium for 48 h in
a fermenter (ANABIO R&D built in-house, Hanoi, Viet-
nam) at the optimal temperature (30–37°C) and pH
(7�0–8�0) for each strain. The purification of spores has been
described previously (Nicholson and Setlow 1990). The
Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology 1359
H.T. Ngo et al. Bacillus from shrimp intestine

Table
1Antibioticsuscep
tibility
ofpigmen
tedBacillusstrainsfrom
shrimpgastrointestinal
tracts
Antibioticdiscs*
SH1†
SH4†
SH5†
SH6†
SH8†
SH12†
SH14†
SH20†
Ampicillin
(10)
43�23
�0�42
S
31�73
�0�00
S
46�74
�0�33
S
22�44
�1�25
S
38�51
�0�49
S
47�55
�0�16
S
36�98
�0�19
S
18�60
�0�55
S
Chloramphen
icol(30)
29�18
�0�21
S
20�77
�0�35
S
31�02
�0�30
S
26�12
�0�92
S
28�83
�0�15
S
31�61
�0�14
S
27�01
�0�78
S
18�83
�0�50
S
Ciprofloxacin(5)
38�80
�0�45
S
17�10
�0�10
S
35�62
�0�58
S
30�87
�0�53
S
27�87
�0�21
S
31�48
�0�31
S
47�08
�0� 00
S
32�67
�0�47
S
Clindam
ycin
(2)
14�89
�0�01
R
12�70
�0�42
R
14�80
�0�01
R
9�57
�0�09
R
8�11
�0�02
I
18�22
�0�01
I
10�35
�0�20
R
17�83
�0�05
R
Co-trimoxazole
(25)
33�72
�0�07
S
23�97
�0�22
S
30�60
�0�30
S
24�63
�0�41
S
30�70
�0�22
S
29�02
�0�01
S
43�52
�0�67
S
26�74
�0�25
S
Erythromycin
(15)
21�18
�0�12
I
20�56
�0�30
I
30�55
�0�10
S
22�55
�0�61
S
25�30
�0�04
S
29�28
�0�17
S
40�80
�1�54
S
28�01
�0�41
S
Gen
tamicin
(10)
29�02
�0�29
S
19�15
�1�19
S
24�57
�0�00
S
19�24
�0�46
S
20�80
�0�07
S
26�49
�0�00
S
31�89
�0�45
S
25�01
�0�14
S
Kan
amycin
(30)
28�63
�0�16
S
17�49
�0�57
S
30�79
�0�06
S
25�33
�1�19
S
24�90
�0�63
S
28�54
�0�01
S
36�75
�0�87
S
23�94
�0�15
S
Neo
mycin
(30)
27�27
�0�67
S
17�14
�0�42
S
24�38
�0�01
S
17�15
�0�05
S
27�79
�0�24
S
21�31
�0�20
S
25�03
�0�30
S
22�55
�0�18
S
Rifam
picin
(30)
32�59
�0�26
S
16� 58
�0�04
I
36�73
�0�32
S
15�51
�0�35
S
30�70
�0�37
S
33�48
�0�30
R
29�14
�0�86
S
15�81
�0�34
S
Streptomycin
(10)
27�87
�0�33
S
14�28
�0�00
S
28�19
�0�01
S
29�40
�1�47
S
16�60
�0�26
S
25�85
�0�16
S
29�65
�0�58
S
21�00
�0�14
S
Tetracycline(30)
30�79
�0�18
S
26�74
�0�48
S
33�78
�0�21
S
28�28
�0�36
S
31�01
�0�05
S
33�41
�0�40
S
40�16
�0�00
S
23�27
�0�21
S
Trim
ethoprim
(5)
37�48
�0�00
S
6�45
�0�00
S
34�38
�0�16
S
24�01
�0�73
S
6�76
�0�00
S
31�52
�0�27
S
50�06
�0�54
S
30�28
�0�07
S
Van
comycin
(30)
19�57
�0�01
S
14�77
�0�26
S
19�51
�0�08
S
16�65
�0�44
S
16�70
�0�11
S
20�44
�0�06
S
24�29
�0�57
S
13�74
�0�09
S
S,sensitive;I,interm
ediate;R,resistan
t.
*Antibiotic-im
pregnated
discs
(6mm)witham
ountin
lgshownin
brackets.
†Averagediameter
ofinhibitionfrom
threeindividual
experim
ents.
Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology1360
Bacillus from shrimp intestine H.T. Ngo et al.

purified spore suspensions were then spray-dried at 160°Cat an atomizer speed of 28 000 g (ANABIO R&D built
in-house, Hanoi, Vietnam) for collection of spore powder.
Preparation of feed supplemented with spores or
synthesized astaxanthin
Commercial shrimp feed (Uni President, Tainan, Taiwan)
was used as the basal diet, supplemented with either
spores or astaxanthin. For selection of the best surviving
strain in shrimp guts, a mixture of spores from five
strains including SH1, SH5, SH6, SH8, and SH14 were
mixed with feed pellets at equal concentrations for each
spore (>3 9 106 CFU g�1 feed); the feed pellets were
then coated with cod liver oil Seven Seas� (Merck Con-
sumer Health, Darmstadt, Germany). For evaluating the
probiotic effects of SH6 spores, feed pellets were supple-
mented with either B. aquimaris SH6 spores or B. indicus
HU36 spores at concentrations of >3 9 106 CFU g�1
feed, followed by coating the pellets with cod liver oil. As
a negative control, the feed pellets were coated with cod
liver oil. As a positive control, the commercialized Caro-
phyll Pink� (DSM, Heerlen, the Netherlands) powder
containing 10% synthesized astaxanthin was used as sup-
plements. Three grams of Carophyll Pink� powder was
dissolved in 30 ml water (70°C) then sprayed on 600 g
commercial shrimp feed (ratio: 5 mg Carophyll powder
to 1 g pellets) to have a final astaxanthin concentration
of 0�5 mg g�1 pellets; feed pellets were then coated with
the liver oil. The feed pellets with or without spores were
coded and stored at room temperature for the duration
of the experimental trials in whiteleg shrimps.
Trials of probiotic treatment in whiteleg shrimp on a
laboratory scale
Forty-five-day-old whiteleg shrimp (L. vannamei), weigh-
ing approx. 3 g, were used for probiotic treatment on a
laboratory scale. Approval of animal care and use proto-
col form for conducting trials in shrimps is not required
as shrimp species are invertebrates. Trials in whiteleg
shrimp followed biosecurity guidelines of the Department
of Aquaculture, Vietnam Ministry of Agriculture and
Rural Development.
For the in vivo survival assays of Bacillus spores in
shrimp guts, shrimp were divided into two groups
(n = 10 shrimp per tank). Shrimp in each group were fed
as follows: commercial feed only (control group), feed
supplemented with a mixture of SH1, SH5, SH6, SH8,
and SH14 spores at >3 9 106 CFU g�1 pellet (Spore
group). After 7 days of feeding, five shrimp guts from
each group were taken at each time point: (i) 3 h after
the last feeding and (ii) 8 h after the last feeding. Shrimp
guts were prepared to collect the mucosa in 0�9% NaCl
and then vigorously resuspended by vortexing until a
homogenous suspension was obtained. Serial dilutions
with 0�9% NaCl were made before plating on TSA and
incubating for 1 day at 37°C to obtain individual colo-
nies. Colonies were observed based on their typical mor-
phologies and colours, and then counted to calculate
their initial population in shrimp guts.
For evaluating the probiotic activity of SH6 spores, a
blind trial was performed. Shrimps (n = 30 per tank) in
each group were fed with coded feed pellets as follows:
commercial feed only (control group), feed supplemented
with SH6 spores at >3 9 106 CFU g�1 pellet (SH6
group), feed supplemented with HU36 spores at
>3 9 106 CFU g�1 pellet (HU36 group), and feed sup-
plemented with the commercialized Carophyll� at 0�5 mg
synthesized astaxanthin g�1 pellet (Carophyll group). The
shrimp were maintained in a water bath thermostatically
controlled at 28°C and fed 5 g feed per day. For measur-
ing shrimp weight and phenoloxidase (PO) activity, ten
shrimp (n = 10) from each group were taken and data
were recorded after 14 and 28 days. The SD was calcu-
lated based on data collected from ten shrimp. For com-
paring colours, five shrimp (n = 5) in each group were
taken after 28 days, and then boiled comparing colour
using the Roche index, SalmoFanTM standard colour
(Brun and Fr�ed�eric 2006). For measuring astaxanthin
concentration, five shrimp (n = 5) in each group were
taken and data were recorded after 28 days. SD was cal-
culated based on data collected from five shrimp.
For evaluating the safety of the SH6 probiotics, a simi-
lar experimental design was set up for the four groups
(n = 30 per tank; 2 tanks per group) feed was supple-
mented with spores at an extremely high concentration
(>1 9 109 CFU g�1 pellet). Survival of shrimp in each
group was recorded after 7, 14, 21 and 28 days for calcu-
lation of survival rate at each time point. SD was calcu-
lated based on data collected from two tanks.
Extraction and measurement of astaxanthin in shrimp
Shrimp (n = 5) from each experimental group were treated
as follows. The chitin shell was removed, and the shrimp
were pulped well in liquid nitrogen. Pigments, including
astaxanthin, were extracted from 3-g shrimp samples by
adding 2 ml methanol, followed by 4 ml chloroform. The
suspension was incubated on ice for 20 min to minimize
degradation of astaxanthin. To the suspension, 1 ml water
was added and the sample was vortexed for 15 s. To form a
partition, the suspension was centrifuged for 3 min at
10 000 g. The lower phase (organic hypophase) was col-
lected and the upper phase (aqueous hyperphase)
re-extracted twice with chloroform until no colour was
Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology 1361
H.T. Ngo et al. Bacillus from shrimp intestine

observed in the debris. Protein contamination was
removed from extractions, which were concentrated in
PBS. The extracts were stored at �20°C at this stage. Before
measuring, 1-ml samples were concentrated fivefold using
0�2 ml acetonitrile/methanol/dichloromethane (ratio
75 : 20 : 5) and injected onto the HPLC column. Astaxan-
thin in extract (A) was first separated and then detected
online, using the Waters Alliance (Milford, MA) HPLC sys-
tem with an online photo diode array (PDA 2996 at
450 nm) detector. Injections were made, and separations
performed on a reverse-phase (RP) C18 (2) 5-lm column
(150 mm 9 4�6 mm; Agilent, San Francisco, CA). The
mobile phase, acetonitrile/methanol/dichloromethane
(75 : 20 : 5), eluted at a rate of 1�5 ml min�1, was main-
tained at a constant temperature of 30°C. A defined sample
of astaxanthin (2�68 lmol l�1), used as control, was mea-
sured at the same time to compare retention time (Rt).
Concentrations of astaxanthin in the extract were mea-
sured based on height and area of peaks. Total astaxanthin
concentration per 1 g of shrimp Cs (lg g�1 shrimp�1) was
calculated using the equation: Cs = k 9 1�67 9 Ce/W,
where k was the dilution ratio of the samples (k = 6), Ce
was astaxanthin concentration in the extract (lmol l�1),
and W was the weight of shrimp (g).
Phenoloxidase activity measurement
PO activities in shrimp were measured following the pro-
tocol of a previous report by Luciane and Margherita
(1997) and Nguyen et al. (2014).
Statistical analysis
Data of astaxanthin concentrations, weight gains and PO
activities among the four treatment groups were compared
using a Student’s t-test at a significance level of 0�05, 0�01,0�001 and 0�0001. Statistical analyses were performed using
the Analysis ToolPak in MICROSOFT EXCEL Software (Micro-
soft, Redmond, WA). An F-test for two-sample variance
was used before performing a t-test for two unpaired sam-
ples. ANOVA single factor analysis was used to compare more
than two samples. Survival rate data among the four
treatment groups were compared using a Chi-Squared X2
test and P values were considered significant at a level of
<0�05.
Results
Characterization of pigmented Bacillus strains isolated
from shrimp gastrointestinal tracts
We isolated 23 pigmented, spore-forming aerobic strains
of bacteria from the guts of 12 shrimp collected from
rivers and coastal regions of Ben Tre (Vietnam). To
develop heat-stable probiotics as feed supplements to
colourize shrimp and improve shrimp health, we screened
strains of Bacillus. We first screened strains based on the
following criteria: (i) diverse pigment found in bacterial
strains ranging from yellow, orange, red and pink, (ii)
high absorbance value of methanol-chloroform extracted
carotenoids at a typical UV-Vis wavelength of 400–550 nm (equivalent to more than 250 lmol l�1), and
(iii) a high sporulation efficiency of more than 85%. Our
primary investigation of the 23 pigmented Bacillus strains
indicated that they had much different characteristics. As
shown in Table 2, we screened eight representative pig-
mented, spore-forming strains named SH1, SH4, SH5,
SH6, SH8, SH12, SH14, SH20 having different colours
and peaks of absorbance wavelengths. For example, the
pink-red extract of SH1 had the highest peak at 495 nm,
the red-orange extract of SH6 had the highest peak at
468 nm, and the yellow extract of SH8 had three peaks at
435, 465 and 487 nm. The data were confirmed by a neg-
ative absorbance value obtained from the reference non-
pigmented B. subtilis PY79 strain and positive VIS
absorbance peak of extract from strain HU36 at 454 nm
(data not shown). Among the eight strains, SH1, SH5,
SH6, SH8, SH12 and SH20 produced high levels of caro-
tenoids equivalent to 250 lg ml�1 or higher. All strains
were able to sporulate quickly in DSM medium with
sporulation efficiency ranging from 85 to 100%. Most of
the pigmented spores were not very heat resistant, except
that of SH8, which retained its haft-count (50% survival)
after treatment at 80°C for 20 min. With lower tempera-
ture treatment (55°C for 20 min), 50% survival of SH5,
SH6 and SH12 spores were retained, whereas live counts
of SH1, SH14, and SH20 spores remained at only 1–10%.
The eight pigmented strains were further characterized
based on physiological and biochemical properties
according to Bergey’s Manual of Systematic Bacteriology
and 16S rRNA sequence analysis (Table 2). In all strains,
width of vegetative cells was less than 1 lm. The spore
position and characteristics of isolates were different from
each other; some were subterminal, and others were ter-
minal and central. All strains were able to grow aerobi-
cally at 30°C, the temperature for culturing shrimp. Only
the SH8 strain was able to growth in both aerobic and
anaerobic conditions. In the presence of 6�5% NaCl, all
strains also grew well. Most strains were able to hydrolyse
starch at different levels except SH8. Different strains
exhibited different levels of caseinase, except SH5. In the
lipase test, only three strains, SH1, SH6 and SH8, could
hydrolyse Tween 80, as indicated by observable opaque
halos around these colonies. Among the eight screened
strains, SH6 exhibited strong amylase, caseinase and
lipase activities. For assessing the safety and toxicity of
Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology1362
Bacillus from shrimp intestine H.T. Ngo et al.

these eight pigmented strains, we used sheep blood agar
and observed haemolysis zones; all isolates were found to
be negative in haemolysis. All strains were evaluated for
their antibiotic sensitivity to a panel of antibiotics,
including those highlighted by the European Food Safety
Authority (EFSA 2005) and recommended by the NCCLS
(1997). As shown in Table 1, all strains were sensitive to
most of the 14 tested antibiotics. Among these, 11 antibi-
otic groups were included with different modes of action,
including a b-lactam (ampicillin), a quinolone (ciproflox-
acin), a lincosamide (clindamycin), a macrolide (ery-
thromycin), four aminoglycosides (gentamicin,
streptomycin, kanamycin, neomycin), a glycopeptide
(vancomycin) and rifampicin, tetracycline, chlorampheni-
col and cotrimoxazol (trimethoprim and sulfamethoxa-
zole). Unexpectedly, all strains were either resistant or
moderately sensitive to clindamycin; SH4 was moderately
sensitive to erythromycin and rifampicin, and SH12 was
resistant to rifampicin. To determine species identity,
strains were assessed by 16S rRNA sequencing and BLAST
analysis. Identification at the species level using 16S
rRNA sequence analysis indicated that strains were closely
related to B. aquimaris (SH1, SH4, SH6, SH14, SH20),
B. firmus (SH5, SH12) and B. marisflavi (SH8) with iden-
tity scores equal to or greater than 0�98.Assessing all characterization data listed in Tables 1
and 2, we determined that SH6 was the most promising
strain due to the following: (i) high production level of
carotenoids, (ii) sporulation efficiency of 100%, and (iii)
good production of three kinds of enzymes including
amylase, protease and lipase. By analysing the phyloge-
netic relationship (based on rRNA sequences) between
SH6 (GenBank accession No: KF443807) and other refer-
ence strains (Fig. 1), SH6 was confirmed to belong to the
B. aquimaris species.
Antioxidant activities of pigmented extracts
An important criterion for identifying carotenoids is
antioxidant activity. Thus, to confirm that the pigments
found in selected strains were carotenoids, we used a
standard scavenging assay of free DPPH radicals. As
shown in Fig. 2, pigmented extracts of four isolated
strains including SH1, SH5, SH6 and SH8 exhibited
Table 2 Characterization of pigmented Bacillus strains isolated from shrimp gastrointestinal tracts
Strains SH1 SH4 SH5 SH6 SH8 SH12 SH14 SH20
Colour* PR PO PO RO Y PO YO RP
UV-Vis spectral
characteristics (nm)
495 494 490 468 435, 465, 487 504 470 460
Carotenoid
production
++++ +++ ++++ ++++ ++++ ++++ ++ ++++
Sporulation (%) 85 80 95 100 100 95 80 90
Survival during
heat treatment†
++ + +++ +++ ++++ +++ + ++
Size of bacteria
(lm)‡
<1 <1 <1 <1 <1 <1 <1 <1
Spore position S S T C S T S T
Aerobic + + + + + + + +
Anaerobic � � � � + � � �Growth at 30°C + + +++ +++ ++ +++ + +++
6�5% NaCl + + + + + + + +
Amylase ++ ++ +++ ++ � ++ ++ ++
Caseinase + ++ � ++ + + + +
Lipase ++ + � ++++ ++ + � �Haemolysis c c c c c c c c
VP Test � + + � � � � �Closest match§ Bacillus aquimaris
(0�99)B. aquimaris
(0�99)Bacillus
firmus
(0�98)
B. aquimaris
(0�99)Bacillus
marisflavi
(0�99)
B. firmus
(0�98)B. aquimaris
(0�99)B. aquimaris
(0�99)
T, Terminal; S, subterminal; C, central.
�, negative; +, weak or positive; ++, average; +++, good/high; ++++, very good/very high.
*Colour of vegetative cell, P, pink; R, red; Y, yellow.
†Heat treatment at 55°C, 20 min.
‡Width size of vegetative cell.
§Using 16S rDNA sequence analysis in this work. The similarity score is shown in brackets.
Peaks having higher absorbance values were underlined
Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology 1363
H.T. Ngo et al. Bacillus from shrimp intestine

significantly high scavenging activity 76–98%, whereas the
four remaining extracts of SH4, SH12, SH14, and SH20
had less than 70% scavenging activities (data not shown).
The pigmented extracts of SH5 and SH6 had the highest
DPPH radical scavenging activities (SH5: 98�5 � 0�2%;
SH6: 87�0 � 0�9%) among all strains, which was equal to
that of ascorbic acid (96�0 � 0�2% at a concentration of
9�38 lmol l�1) and higher than that of the positive con-
trol HU36 (78�4 � 0�9%). The data were confirmed
using nonpigmented B. subtilis PY79 as a negative con-
trol, wherein no detectable DPPH scavenging activity was
observed. Based on this data, we concluded that the pig-
ments found in the eight selected strains showed the
potential to be antioxidant carotenoids, and that carote-
noids of the SH5 and SH6 strains exhibited stronger
antioxidant activity than those of the other strains (SH1:
77�1 � 1�1%; SH8: 84�8 � 0�5%).
Survival and colonization of strain SH6 in shrimp
intestines
After the in vitro characterization of Bacillus strains, we
wanted to screen strains for the in vivo characteristics of
survival and colonization in shrimp guts, which are
important probiotic properties. For convenience, we
selected five strains with distinguished colony morphol-
ogy, including SH1, SH5, SH6, SH8, and SH14, for
assessing viability in shrimp guts. We set up two time
points for counting bacterial populations in the shrimp
gut, 3 and 8 h after the last feeding. This was because
transit time of feed in the shrimp intestine is 4–5 h. At
3 h, our results showed that colonies of the SH6 strain
were the most abundant, accounting for 70% of the total
microbiota (Fig. 3a). Meanwhile, SH1 accounted for only
for 30% of the population, and colonies of SH5, SH8
Bacillus marisflavi strain TF-11 (NR025240)
Bacillus marisflavi strain SNS4 (LC009453)
Bacillus aquimaris strain CH9 (KM516787.1)
Bacillus aquimaris strain SH6 (KF443807)
Bacillus aquimaris strain TF-12 (NR025241.1)
Bacillus lichenifomis strain AIX64 (GU967449)
Bacillus pumilis strain GC43 (KF158227)
Bacillus megaterium isolate OS-223 (AM237398)
Bacillus cereus strain LKT 1/2 (AJ577284)
Lactobacillus acidiphillus strain ATCC 4356 (AB008203)
0·01
93
97
87
98
Bacillus lichenifomis strain LZBL-13 (JX847119)
76
Bacillus pumilis strain ESR21 (KC915229)
Bacillus megaterium strain WN606 (DQ275183)
Bacillus cereus strain G56 (KM019745)
100
100
100
100
Figure 1 Phylogenetic relationship between the selected pigmented Bacillus strains. Dendrograms of strains based on 16S rRNA sequence align-
ment using MEGA ver. 6 software. A selected Bacillus strain is highlighted in bold and GenBank accession numbers are shown in brackets. Statisti-
cal (bootstrap) values and a scale bar representing evolutionary distance are shown. The 16S rRNA gene sequence of the lactic acid bacterium
Lactobacillus acidophilus, an out-group of the Bacillus genus, was used as the root of the phylogenetic tree.
Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology1364
Bacillus from shrimp intestine H.T. Ngo et al.

and SH14 were not detectable. Thus, it was concluded
that SH6 spores survive better than spores of other pig-
mented strains and were the most abundant population
in the shrimp gut during the passage of feed. To further
access adhesion and colonization capability of pigmented
Bacillus strains in the shrimp intestinal mucosa, we col-
lected spores 8 h after feeding, when feed was completely
extruded from the shrimp intestine. The results (Fig. 3b)
indicated that there was an increase in nonpigmented
Bacillus colonies (~79%) and a significant reduction in
pigmented Bacillus colonies (21%). However, colonies of
the SH6 strain were still the most abundant among pig-
mented strains (20% of the population), followed by SH1
(1% of the population). Thus, even though it was not
comparable to that of nonpigmented Bacillus strains, col-
onization of the SH6 strain in the shrimp gut was still
the best among pigmented Bacillus strains. As SH6
showed promising probiotic properties in vitro (as men-
tioned above) and in vivo (based on colonization assays),
we decided to characterize the SH6 strain for the devel-
opment of probiotics for use in whiteleg shrimp.
Carotenoid profiling of SH6 and characterization of SH6
spores
To clarify the type of carotenoids produced by the SH6
strain, the carotenoid extract was analysed by HPLC-
PDA. Typical HPLC chromatographic profiles for the
SH6 extract were recorded at 450 nm and are presented
in Fig. 4. The physical characteristics of the carotenoids
detected were concordant with the visible colour of the
orange-red SH6 colonies, as fraction no. 3 (Rt at
29�437 min), showing absorbance peaks at 492 and
522 nm (orange, pink, red), was the most abundant.
Bacillus strains are conventionally thought to form
heat-resistant spores. However, our primary data, shown
in Table 2, indicate that pigmented Bacillus strains do
not possess this ability, as most were not stable during
heat treatment at 80°C for 20 min. However, to utilize
SH6 as a probiotic strain in feed production, we needed
to assess sporulation and its exact level of heat stability.
For this, we attempted to culture SH6 spores in DSM
broth for 48 h using a fermenter; the resulting spore sus-
pension was spray-dried to obtain a heat-stable spore
powder. As controls, we also produced spore powders of
the medium-sporulating B. indicus HU36 yellow-pigmen-
ted strain (about 50% sporulation efficiency) and the
high-sporulating nonpigmented B. subtilis HU58 strain
(almost 100% sporulation efficiency), both reference
strains. As expected, from 50 l of we obtained 500 g of
red-orange SH6 spore powder at the very high concentra-
tion of 4�5 9 1011 CFU g�1 (Fig. 5a) with almost 100%
spore purity (Fig. 5b). This concentration was almost
equal to that obtained for HU58 spores
(5�8 9 1011 CFU g�1), and was 4-fold higher than that
obtained for HU36 spores (data not shown). To explain
this difference in spore concentration, the heat stability of
the SH6 spores was compared to that of HU36 and
HU58. As shown in Fig. 5c, HU36 spores were surpris-
ingly heat-sensitive as the live count was reduced by 8%
at only 50°C. In contrast, SH6 spores were stable at 50°C(60% survival), and 15% survival was maintained at
70°C, equivalent to 50% survival at 55°C. However, the
heat stability of SH6 spores was much less than that of
the B. subtilis HU58 spores (55% survival at 80°C, 23%survival at 90°C); the heat stability of HU58 has been
characterized for use as a food ingredient (Permpoonpat-
tana et al. 2012).
0
10
20
30
40
50
60
70
80
90
100
SH1SH5
SH6SH8
HU36PY79
Sca
veng
ing
perc
enta
ge o
f DP
PH
(%
)
Group
37·5
µm
ol l–1
18·7
5 µm
ol l–1
9·38
µm
ol l–1
4·69
µm
ol l–1
Figure 2 Inhibition of DPPH free radical
formation by pigmented extracts. (■) Samples
include extracts of SH1, SH5, SH6, SH8 and
reference strains (HU36, PY79). ( ) Control
was ascorbic acid at concentrations of 18�75,9�38 and 4�69 lmol l�1. Inhibition (%) =
[(ODcontrol � ODsample)/ODcontrol] 9 100.
Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology 1365
H.T. Ngo et al. Bacillus from shrimp intestine

Improved pigmentation of whiteleg shrimp by
supplementing with SH6 spores
We first evaluated the effects of SH6 probiotics on the
pigmentation of whiteleg shrimp. Four groups were
designed for pigmentation assays, including SH6 (fed
with SH6 spores at >3 9 106 CFU g�1 pellet), HU36
(fed with the reference HU36 spores at
>3 9 106 CFU g�1 pellet), control (without supple-
ments), and Carophyll (fed with commercial Carophyll�,
at 0�5 mg synthesized astaxanthin g�1 pellet). After
4 weeks of feeding, shrimp colour after boiling was com-
pared for the four groups (Fig. 6a). The Carophyll group
had the reddest colour (score: 24–25), followed by the
SH6 group (score: 21–23), the HU36 group (score: 21–23), and the control group (score: 20–22).
To correlate the relationship between the shrimp colour
and the presence of astaxanthin in shrimp tissue, we deter-
mined the astaxanthin levels in shrimp of each group. As
shown in Fig. 6b, we found that the level of astaxanthin in
the SH6 group (0�69 � 0�03 lg g�1 shrimp�1) was 2�7-fold higher than that of the control (0�25 � 0�09 lg g�1
shrimp�1; P < 0�05) but 2�3-fold lower than that of
the Carophyll group (1�59 � 0�08 lg g�1 shrimp�1;
P < 0�01). Unexpectedly, the difference in astaxanthin
levels between the HU36 group (0�47 � 0�13lg g�1 shrimp�1) and control group was not statistically
significant (P > 0�05).
Weight gain in whiteleg shrimp by supplementing with
SH6 spores
At time points of 2 and 4 weeks, there was an apparent
disparity between spore and nonspore groups. Shrimp in
both the SH6 and HU36 groups gained weight much fas-
ter than the control and Carophyll groups (P < 0�0001)at both time points. For example, the SH6 group gained
5�36 � 0�3 g and 7�18 � 0�24 g at 2 and 4 weeks
SH1: 30%
SH6: 70%
SH1: 1%
SH6: 20%
Nonpigmented Bacillus: 79%
(b)(a)
Figure 3 Survival and colonization of
pigmented Bacillus strains in the shrimp gut.
Panel (a) Pie chart of the major population of
SH6 & SH1 strains isolated from shrimp guts
after 3-h-feeding of the experimental group
(fed with a mixture of SH1, SH5, SH6, SH8
and HU36 spores at >3 9 106 CFU g�1).
Panel (b) Pie chart of population (B2) of major
nonpigmented Bacillus strains and SH6 and
SH1 strains isolated from shrimp guts after 8-
h-feeding in the experimental group.
5·0 10·0 15·0 20·0 25·0 30·0 min
0·0
2·5
5·0
7·5
10·0
mAU450 nm,4 nm (1·00)
20·1
09
22·2
39
29·4
37
Figure 4 High-performance liquid
chromatography profiles of SH6 pigment
extract. UV/VIS recorded at 450 nm indicating
two carotenoid types at low absorbance
levels of peak no. 1, Rt at 20�109 min, k max
(484; 496), peak no. 2, Rt at 22�239 min, k
max (466; 489) and one carotenoid type at
high absorbance level of peak no. 3, Rt at
29�437 min, k max (492; 522). Peaks having
higher absorbance values were underlined.
Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology1366
Bacillus from shrimp intestine H.T. Ngo et al.

respectively; the HU36 group gained 5�22 � 0�19 g and
6�71 � 0�21 g at 2 and 4 weeks respectively. By contrast,
the control group gained only 4�26 � 0�35 at 2 weeks
and 5�32 � 0�26 g at 4 weeks (Fig. 7a). In the nonspore
Carophyll group, shrimp were heavier (4�79 � 0�22 g
and 6�00 � 0�26 g at 2 and 4 weeks respectively;
P < 0�0001) than those in the control group, but not as
heavy as those in the SH6 group.
Increased immune-related phenoloxidase activity by
supplementing with SH6 spores
Shrimp have no adaptive memory, and therefore depend
on innate defence systems to protect against pathogens.
PO activity is the parameter most used to represent the
innate immune response, which protests against patho-
genic attacks (Sarathi et al. 2007; Nguyen et al. 2014).
We found that PO activity in the SH6 group was clearly
higher than that of other groups after 2 weeks of contin-
uous feeding (OD490 = 0�134 for the SH6 group vs 0�09for the HU36 group, 0�076 for the control group, and
0�081 for the Carophyll group; P < 0�01). At week four,
an 85% increase in PO activity was observed in the SH6
group (OD490 = 0�265) compared to that of the control
group (OD490 = 0�143; P < 0�01), whereas the PO
activities in the HU36 (OD490 = 0�121) and Carophyll
groups (OD490 = 0�121) were not significant different
(P > 0�05) from those of the control group (Fig. 7b).
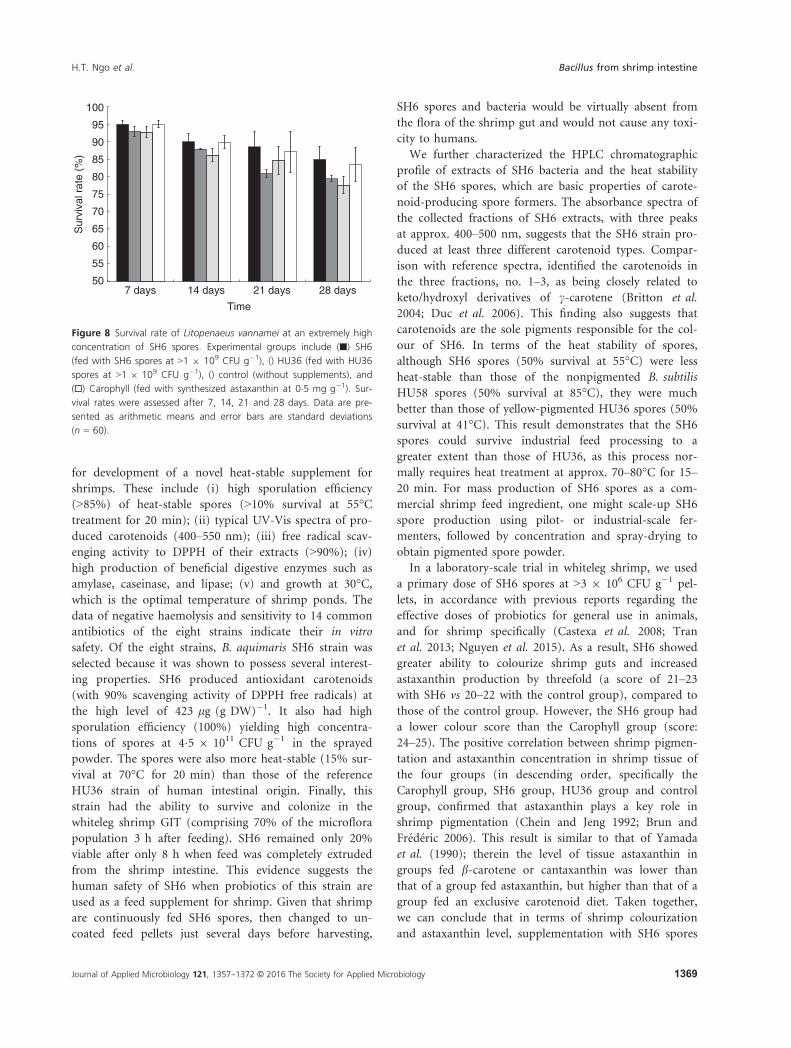
Safety of SH6 spores
SH6 is a new probiotic strain that has not been well doc-
umented in the Qualified Presume as Safe (QPS) list. We
therefore determined the toxicity dose of SH6 spores in
whiteleg shrimp through increasing the dose of probiotics
up to >1 9 109 CFU g�1, which was approx. 300-fold
higher than the probiotic concentration
(>3 9 106 CFU g�1). After 2 and 4 weeks of feeding, we
found that the SH6-fed or HU36-fed groups did not die,
but experienced an even better survival rate (SH6:
84�9 � 3�6% and HU36: 79�4 � 0�9%), compared to that
of the control group (77�4 � 2�4%; Fig. 8). The Caro-
phyll-fed group also had a good survival rate of
83�5 � 4�9%. Nevertheless, the increases in survival rates
for SH6-fed (9�5%), HU6-fed (2�3%), and Carophyll-fed
groups (7�4%) in comparison to those of the control
group were not statistically significant (P > 0�05). The
data confirm that the lethal doses of both SH6 and HU36
spores in whiteleg shrimp were above 1 9 109 CFU g�1
for up to a 4-week treatment.
(a) (b)
0
0.5CF
U (
× 1
09 m
l–1 )
1
1.5
2
2.5
3
Temperature (°C)
25 40 50 60 70 80 90
2 µm
(c)Figure 5 SH6 spores and their heat
resistance. Panel (a) Spray-dried powder of
orange SH6 spores (>3 9 1011 CFU g�1).
Panel (b) Microscopy observation of 100%
SH6 spores in the powder. Panel (c) Heat
stability of SH6 in comparison to that of
HU58 and HU36 spores. Heat-counts of (■)
SH6, ( ) HU58 and (□) HU36 after treatment
at various temperatures ranging from Rt to
90°C. Data are presented as arithmetic
means and error bars are standard deviations
(n = 3). Colour figure can be viewed at
wileyonlinelibrary.com.
Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology 1367
H.T. Ngo et al. Bacillus from shrimp intestine

Discussion
As mentioned in the introduction to this article, a num-
ber of publications have reported the identification and
characterization of pigmented bacilli isolated from seawa-
ter, water and mud in shrimp ponds, soil, and human
faeces. To our knowledge, this is the first study to isolate
and characterize carotenoid-producing Bacillus strains
from the shrimp GIT. To develop a novel supplement for
promoting the colourization and health of shrimp, we
hypothesized that probiotic strains for shrimp should be
isolated from the intestinal tracts of shrimp, because
these strains have the best chance of surviving and colo-
nizing in the intestine. We have tried isolating and
screening spore-forming bacteria, which belong to the
genus Bacillus, because their spores are normally stable in
acidic conditions of the stomach. Thus, they can pass
through the stomach unscathed and subsequently germi-
nate in the small intestine and grow (Tam et al. 2006).
Then, we can characterize and screen the best strain of
pigmented bacteria, which produce carotenoids, to
develop probiotics for improving the colour of whiteleg
shrimp and providing antioxidants and nutrition for
shrimp. Here, whiteleg shrimp was chosen for trials
because it is one of the most exported species from Viet-
nam (Lan 2013) and its colour change is easy to observe.
Based on this hypothesis, we set up a protocol to isolate
pigmented colonies from GIT of shrimp. All shrimp were
collected from wild rivers and coastal regions in the Ben
Tre province to avoid the use of any commercial probi-
otics and to obtain biodiversity of the intestinal micro-
organisms.
Among 23 coloured isolates, eight strains were selected
from in vitro conditions due to their potential properties
0
0·2
0·4
0·6
0·8
1
1·2
1·4
1·6
1·8
SH6 HU36 Control Carophyll
µg a
stax
anth
in g
–1 s
hrim
p–1
Group
*
**
Score: 21–23 21–23 20–22 24–25
Group: SH6 HU36 Control CarophyII(a)
(b)
Figure 6 Colour and astaxanthin concentration in Litopenaeus van-
namei after 28 days feeding with SH6 spores. Experiment groups
include SH6 (fed with SH6 spores at >3 9 106 CFU g�1), HU36 (fed
with HU36 spores at >3 9 106 CFU g�1), control (without supple-
ments), and Carophyll (fed with synthesized astaxanthin at
0�5 mg g�1). Panel (a). Image of boiled shrimps and their variable col-
our scores indicating the levels of red pigmentation. Panel (b) Astax-
anthin concentrations in shrimp. Data are presented as arithmetic
means and error bars are standard deviations (n = 5). P values were
generated by ANOVA using the Student’s t-test for multiple compar-
isons to the control (*P < 0�05; **P < 0�01). Colour figure can be
viewed at wileyonlinelibrary.com.
0·0
1·0
2·0
3·0
4·0
5·0
6·0
7·0
8·0
14 days 28 days
Wei
ght (
g sh
rimp–
1 )
Time
Time
**** ********
********
****
0
0·05
0·1
0·15
0·2
0·25
0·3
0·35
14 days 28 daysPhe
nolo
xida
se a
ctiv
ity (
O.D
. 490
nm
)
**
**
****
(a)
(b)
Figure 7 Weight gain (a) and phenoloxidase activity (b) of Litope-
naeus vannamei after 14 and 28 days feeding with SH6 spores.
Experimental groups include (■) SH6 (fed with SH6 spores at
>3 9 106 CFU g�1), ( ) HU36 (fed with HU36 spores at >3 9 106
CFU g�1), ( ) control (without supplements), and (□) Carophyll (fed
with synthesized astaxanthin at 0�5 mg g�1). Data are presented as
arithmetic means and error bars are standard deviations (n = 10). P
values were generated by ANOVA using the Student’s t-test for multiple
comparisons to the control (**P < 0�01; ****P < 0�0001).
Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology1368
Bacillus from shrimp intestine H.T. Ngo et al.
for development of a novel heat-stable supplement for
shrimps. These include (i) high sporulation efficiency
(>85%) of heat-stable spores (>10% survival at 55°Ctreatment for 20 min); (ii) typical UV-Vis spectra of pro-
duced carotenoids (400–550 nm); (iii) free radical scav-
enging activity to DPPH of their extracts (>90%); (iv)
high production of beneficial digestive enzymes such as
amylase, caseinase, and lipase; (v) and growth at 30°C,which is the optimal temperature of shrimp ponds. The
data of negative haemolysis and sensitivity to 14 common
antibiotics of the eight strains indicate their in vitro
safety. Of the eight strains, B. aquimaris SH6 strain was
selected because it was shown to possess several interest-
ing properties. SH6 produced antioxidant carotenoids
(with 90% scavenging activity of DPPH free radicals) at
the high level of 423 lg (g DW)�1. It also had high
sporulation efficiency (100%) yielding high concentra-
tions of spores at 4�5 9 1011 CFU g�1 in the sprayed
powder. The spores were also more heat-stable (15% sur-
vival at 70°C for 20 min) than those of the reference
HU36 strain of human intestinal origin. Finally, this
strain had the ability to survive and colonize in the
whiteleg shrimp GIT (comprising 70% of the microflora
population 3 h after feeding). SH6 remained only 20%
viable after only 8 h when feed was completely extruded
from the shrimp intestine. This evidence suggests the
human safety of SH6 when probiotics of this strain are
used as a feed supplement for shrimp. Given that shrimp
are continuously fed SH6 spores, then changed to un-
coated feed pellets just several days before harvesting,
SH6 spores and bacteria would be virtually absent from
the flora of the shrimp gut and would not cause any toxi-
city to humans.
We further characterized the HPLC chromatographic
profile of extracts of SH6 bacteria and the heat stability
of the SH6 spores, which are basic properties of carote-
noid-producing spore formers. The absorbance spectra of
the collected fractions of SH6 extracts, with three peaks
at approx. 400–500 nm, suggests that the SH6 strain pro-
duced at least three different carotenoid types. Compar-
ison with reference spectra, identified the carotenoids in
the three fractions, no. 1–3, as being closely related to
keto/hydroxyl derivatives of c-carotene (Britton et al.
2004; Duc et al. 2006). This finding also suggests that
carotenoids are the sole pigments responsible for the col-
our of SH6. In terms of the heat stability of spores,
although SH6 spores (50% survival at 55°C) were less
heat-stable than those of the nonpigmented B. subtilis
HU58 spores (50% survival at 85°C), they were much
better than those of yellow-pigmented HU36 spores (50%
survival at 41°C). This result demonstrates that the SH6
spores could survive industrial feed processing to a
greater extent than those of HU36, as this process nor-
mally requires heat treatment at approx. 70–80°C for 15–20 min. For mass production of SH6 spores as a com-
mercial shrimp feed ingredient, one might scale-up SH6
spore production using pilot- or industrial-scale fer-
menters, followed by concentration and spray-drying to
obtain pigmented spore powder.
In a laboratory-scale trial in whiteleg shrimp, we used
a primary dose of SH6 spores at >3 9 106 CFU g�1 pel-
lets, in accordance with previous reports regarding the
effective doses of probiotics for general use in animals,
and for shrimp specifically (Castexa et al. 2008; Tran
et al. 2013; Nguyen et al. 2015). As a result, SH6 showed
greater ability to colourize shrimp guts and increased
astaxanthin production by threefold (a score of 21–23with SH6 vs 20–22 with the control group), compared to
those of the control group. However, the SH6 group had
a lower colour score than the Carophyll group (score:
24–25). The positive correlation between shrimp pigmen-
tation and astaxanthin concentration in shrimp tissue of
the four groups (in descending order, specifically the
Carophyll group, SH6 group, HU36 group and control
group, confirmed that astaxanthin plays a key role in
shrimp pigmentation (Chein and Jeng 1992; Brun and
Fr�ed�eric 2006). This result is similar to that of Yamada
et al. (1990); therein the level of tissue astaxanthin in
groups fed b-carotene or cantaxanthin was lower than
that of a group fed astaxanthin, but higher than that of a
group fed an exclusive carotenoid diet. Taken together,
we can conclude that in terms of shrimp colourization
and astaxanthin level, supplementation with SH6 spores
50
55
60
65
70
75
80
85
90
95
100
7 days 14 days 21 days 28 days
Sur
viva
l rat
e (%
)
Time
Figure 8 Survival rate of Litopenaeus vannamei at an extremely high
concentration of SH6 spores. Experimental groups include (■) SH6
(fed with SH6 spores at >1 9 109 CFU g�1), () HU36 (fed with HU36
spores at >1 9 109 CFU g�1), () control (without supplements), and
(□) Carophyll (fed with synthesized astaxanthin at 0�5 mg g�1). Sur-
vival rates were assessed after 7, 14, 21 and 28 days. Data are pre-
sented as arithmetic means and error bars are standard deviations
(n = 60).
Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology 1369
H.T. Ngo et al. Bacillus from shrimp intestine

was less efficient than supplementation with Carophyll
(astaxanthin), but was slightly better than HU36 spores
and obviously better than uncoated feed.
In terms of weight gain, the SH6 group was heaviest
(7�18 � 0�24 g), followed by the HU36 group
(6�71 � 0�21 g), the Carophyll group (6�00 � 0�26 g),
and the control group (5�32 � 0�26 g). The improvement
on weight gain in groups treated with carotenoids or car-
otenoid-producing bacteria over the control group can be
explained by the fact that carotenoids may perform some
physiological function as an intracellular oxygen reserve
in low dissolved oxygen conditions (Chein and Jeng
1992). Thus, the dietary carotenoid requirement in aqua-
tic animals, which survive in low dissolved oxygen condi-
tions, may by higher than that for species living in
normal conditions. The SH6 and HU36 groups gained
more weight than the Carophyll group because Bacillus
strains not only produce carotenoids, but also produce
beneficial enzymes such as amylase and caseinase to maxi-
mize the digestion of shrimp feed (Table 2 of this work;
Duc et al. 2006). This could result in the observed 34 and
26% increased weight gain, by SH6 and HU36 strains,
respectively, compared to that of the control group, after
4 weeks of feeding. The better weight gain of the SH6
group compared to that of the HU36 group might be due
to better colonization and growth of SH6 in shrimp guts
(Fig. 3), which would result in better digestive health.
The most common parameter reflecting the innate
immune response of shrimp is active PO. PO catalyses
the oxidation of tyrosine to produce toxic quinones and
other short-lived reaction intermediates leading to the
formation of melanin. Melanin then binds to the surface
of bacteria and increases the adhesion of haemocytes to
bacteria, thus accelerating their removal through the for-
mation of small nodes (Cerenius and Soderhall 2004). In
this study, the slight increase in PO activity over time for
all groups (from week two to week four) suggests that
the immune system was improved during shrimp matu-
ration. Similar data were previously reported by Nguyen
et al. (2014), in which increasing PO activities in whiteleg
shrimp groups from day 0 to day 14 was observed during
feeding with pellets of either uncoated or coated spores
of B. subtilis PY79 reference strain. The effective 85%
increase in PO activity in the SH6 group can be attribu-
ted to the immunostimulatory effects of the cell wall
components of this strain, such as b-glucan and
lipopolysaccharide (Rengpipat et al. 2000; Gullian et al.
2004). Alternatively, mucosal immunity in the shrimp
might have been improved through the efficient coloniza-
tion of B. aquimaris SH6 in the shrimp gut. This could
nullify the deregulated and suppressed immune responses
induced by pathogenic bacteria, and optimize immune
parameters (Gatesoupe 1999).
In conclusion, the probiotic properties of the SH6
strain of shrimp origin conferred greater health benefits
to shrimp than those of the HU36 strain of human origin
in terms of shrimp colourization, weight gain and
immune enhancement. In comparison to conventional
supplementation with astaxanthin in shrimp feed, supple-
mentation with SH6 spores was more advantageous in
terms of weight gain and immune enhancement, although
less efficient in terms of colourization. Biosafety data
indicate that SH6 spores, at a 300-fold higher concentra-
tion than that of the conventional probiotic concentra-
tion, did not cause toxicity to shrimps. Thus, SH6 shows
potential as a carotenoid-producing probiotic strain for
further development of a novel feed supplement. Further
interesting questions to be addressed are how SH6 spores
germinate, proliferate and interact with the intestine of
whiteleg shrimp to produce carotenoids, to confer these
beneficial effects. In addition, dose- and time-dependent
trials on the effects of SH6 probiotics in whiteleg shrimps
are necessary to find an optimal feeding regime applicable
for shrimp aquaculture.
Acknowledgements
This research is funded by Vietnam Ministry of Science
and Technology, Viet Nam National University, Hanoi
(VNU) under project number KLEPT.12.03 to N.T.V.A.
We thank Simon M. Cutting for the kind gift of the
reference strains, Vu T. M. Duc and Tran T. My for
technical assistance, Le H. Dung for measuring carote-
noids and astaxanthin levels, Do M. Ha for statistical
analyses, Pham T.T. Huong and Bui T. V. Ha for fruitful
discussion.
Conflict of Interest
The authors report no declarations of interest.
References
Araya, M., Morelli, L., Reid, G., Sanders, M.E. and Stanton, C.
(2002) Guidelines for the evaluation of probiotics in food.
Joint FAO/WHO working group report. No. 1-11.
London, ON, Canada.
Britton, G., Liaaen-Jensen, S. and Pfander, H. (ed.) (2004)
Carotenoids, Handbook. Basel, Switzerland: Birkhauser
Verlag.
Brun, H.L. and Fr�ed�eric, V. (2006) Shrimp pigmentation with
natural carotenoids. Aquac Asia Pacific 2, 34–35.Castexa, M., Chima, L., Phama, D., Lemairea, P., Wabetea, N.,
Nicolasb, J.L., Schmidelyc, P. and Mariojoulsc, C. (2008)
Probiotic P. acidilactici application in shrimp Litopenaeus
Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology1370
Bacillus from shrimp intestine H.T. Ngo et al.

stylirostris culture subject to vibriosis in New Caledonia.
Aquaculture 275, 182–193.Cerenius, L. and Soderhall, K. (2004) The prophenoloxidase-
activating system in invertebrates. Immunol Rev 198, 116–126.
Chein, Y.H. and Jeng, S.C. (1992) Pigmentation of kuruma
prawn, Penaeus japonicas Bate, by various pigment sources
and levels and feeding regimes. Aquaculture 102, 333–346.Cutting, S.M. (2011) Bacillus probiotics. Food Microbiol 28,
214–220.D’Abramo, L.R., Baum, N.A., Bordner, C.E. and Conlline, D.E.
(1983) Carotenoids as a source of pigmentation in
juvenile lobster. Can J Fish Aquat Sci 40,
699–704.Duc, L.H., Fraser, P.D., Tam, N.M.K. and Cutting, S.M.
(2006) Carotenoids present in halotolerant Bacillus spore
formers. FEMS Microbiol Lett 255, 215–224.Dunne, C., O’Mahony, L., Murphy, L., Thornton, G.,
Morrissey, D., O’Halloran, S., Feeney, M. and Flynn, S.
(2001) In vitro selection criteria for probiotic bacteria of
human origin: correlation with in vivo findings. Am J Clin
Nutr 73, 386–392.EFSA (2005) Opinion of the Scientific Committee on a request
from EFSA related to a generic approach to the safety
assessment by EFSA of microorganisms used in feed/food
and the production of feed/food additives. EFSA J 226, 1–12.Evans, J.B. and Kloos, W.E. (1972) Use of shake cultures in a
semisolid thioglycolate medium for differentiating
staphylococci from micrococci. Appl Microbiol 23, 326–331.
Gatesoupe, F.J. (1999) The use of probiotics in aquaculture.
Aquaculture 180, 147–165.Gullian, M., Thompson, F. and Rodriguez, J. (2004) Selection
of probiotic bacteria and study of their immuno-
stimulatory effect in Penaeus vannamei. Aquaculture 233,
1–14.Hong, H.A., Huang, J.M., Khaneja, R., Hiep, L.V., Urdaci,
M.C. and Cutting, S.M. (2008) The safety of Bacillus
subtilis and Bacillus indicus as food probiotics. J Appl
Microbiol 105, 510–520.Khaneja, R., Perez-Fons, L., Fakhry, S., Baccigalupi, L., Steiger, S.,
To, E., Sandmann, G., Dong, T.C. et al. (2009) Carotenoids
found in Bacillus. J Appl Microbiol 108, 1889–1902.Lan, N.T.P. (2013) Social and ecological challenges of market-
oriented shrimp farming in Vietnam. Springer Plus 2, 675–685.
Latscha, T. (1989) The role of astaxanthin in shrimp
pigmentation. Aquacop Ifremer 9, 319–325.Luciane, M.P. and Margherita, A.B. (1997) The
prophenoloxidase activating system of the shrimp Penaeus
paulensis and associated factors. Dev Comp Immunol 21,
385–395.NCCLS (1997) Performance Standards for Antimicrobial Disk
Susceptibility Tests. Wayne, PA: National Committee for
Clinical Laboratory Standards.
Nguyen, T.V.A., Pham, C.K., Pham, H.T.T., Pham, H.L.,
Nguyen, H.A., Dang, T.L., Hong, A.H., Cutting, S.M.
et al. (2014) Bacillus subtillic spores expressing the VP28
antigene: a potential oral treatment to protect Litopenaeus
vannamei against white spot syndrome. FEMS Microbiol
Lett 358, 202–208.Nguyen, T.V.A., Nguyen, D.V., Tran, T.M., Nguyen, T.L.,
Nguyen, H.A. and Phan, T.-N. (2015) Isolation and
characterization of Bacillus subtilis CH16 strain from
chicken gastrointestinal tracts for use as a feed supplement
to promote weight gain in broilers. Lett Appl Microbiol 60,
580–588.Nicholson, W.L. and Setlow, P. (1990) Sporulation,
germination, and outgrowth. In Molecular Biological
Methods for Bacillus ed. Harwood, C.R. and Cutting, S.M.
pp. 391–450. Sussex, UK: John Wiley and Sons.
Pane, L., Radin, L., Franconi, G. and Carli, A. (1996) The
carotenoid pigments of a marine Bacillus firmus strain.
Boll Soc Ital Biol Sper 72, 303–308.Permpoonpattana, P., Hong, H.A., Khaneja, R. and Cutting,
S.M. (2012) Evaluation of Bacillus subtilis strains as
probiotics and their potential as a food ingredient. Benef
Microbes 3, 127–35.Rengpipat, S., Rukpratanporn, S., Piyatiratitivorakul, S. and
Menasaveta, P. (2000) Immunity enhancement in black
tiger shrimp (Penaeus monodon) by a probiotic bacterium
(Bacillus S11). Aquaculture 191, 271–288.Saitou, N. and Nei, M. (1987) The neighbor-joining method: a
new method for reconstructing phylogenetic trees. Mol
Biol Evol 4, 406–425.Sarathi, M., Ahmed, V., Venkatesan, C., Balasubramanian, G.,
Prabavathy, J. and Hameed, A. (2007) Comparative study
on immune response of Fenneropenaeus indicus to Vibrio
alginolyticus and white spot syndrome virus. Aquaculture
271, 8–20.Schmidt-Dannert, C. (2000) Engineering novel carotenoids in
microorganisms. Curr Opin Biotechnol 11, 255–260.Sharma, O.P. and Bhat, T.K. (2009) DPPH antioxidant assay
revisited. Food Chem 113, 1202–1205.Suresh, K., Prabagaran, S.R., Sengupta, S. and Shivaji, S.
(2004) Bacillus indicus sp. nov., an arsenic-resistant
bacterium isolated from an aquifer in West Bengal, India.
Int J Syst Evol Microbiol 54, 1369–1375.Sy, C., Gleize, B., Chamot, S., Dangles, O., Carlin, F., Veyrat,
S.C. and Borel, P. (2013) Glycosyl carotenoids from
marine spore-forming Bacillus sp. strains are readily
bioaccessible and bioavailable. Food Res Int 51,
914–923.Tam, N.K., Uyen, N.Q., Hong, H.A., Duc, L.H., Hoa, T.T.,
Serra, C.R., Henriques, A.O. and Cutting, S.M. (2006) The
intestinal life cycle of Bacillus subtilis and close relatives.
J Bacteriol 188, 2692–2700.Tamura, K., Stecher, G., Peterson, D., Filipski, A. and Kumar,
S. (2013) MEGA6: molecular evolutionary genetics analysis
version 6.0. Mol Biol Evol 30, 2725–2729.
Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology 1371
H.T. Ngo et al. Bacillus from shrimp intestine

Tran, N.T., Pham, M.D. and Hatai, K. (2013) Overview of the use
of probiotics in aquaculture. Inter J Res Fish Aquac 3, 89–97.Vos P., Garrity G., Jones D., Krieg N.R., Ludwig W., Rainey
F.A., Schleifer K.H. and Whitman W.B. (2009) Bergy’s
Manual of Systematic Bacteriology, Volume 3. The
Firmicutes 1984, 2nd edn. New York, NY: Springer-Verlag.
Yamada, S., Tanaka, Y., Sameshima, M. and Ito, Y. (1990)
Pigmentation of prawn (Penaeus japonicus) with
carotenoids. I. Effect of dietary astaxanthin, b-caroteneand cantaxanthin on pigmentation. Aquaculture
87, 323–330.Yoon, J.H., Kim, I.G., Kang, K.H., Oh, T.K. and Park, Y.H.
(2003) Bacillus marisflavi sp. nov. and Bacillus aquimaris
sp. nov., isolated from sea water of a tidal flat of the
Yellow Sea in Korea. Int J Syst Evol Microbiol 53, 1297–1303.
Journal of Applied Microbiology 121, 1357--1372 © 2016 The Society for Applied Microbiology1372
Bacillus from shrimp intestine H.T. Ngo et al.




















