
Apigenin Extends Lifespan and Improves the Activity Pattern of Parkinson’s Disease Model Flies
Upload
independentCategory
view
1download
0
ROCK inhibition extends passage of human embryonic stem cell derived retinal pigmented
epithelium
Roxanne H. Crozea,b,c,d
, David E. Buchholza,b,c,d
, Monte Radekeb,c,d
, William Thia,b,c,d
, Peter J.
Coffeyb,c,d
and Dennis O. Clegga,b,c,d
aCenter for Stem Cell Biology and Engineering,
bCenter for the Study of Macular Degeneration,
cNeuroscience Research Institute,
dDepartment of Molecular, Cellular and Developmental
Biology, University of California, Santa Barbara, CA.
*Corresponding author: Dennis O. Clegg, Ph.D., Center for Stem Cell Biology and Engineering,
Neuroscience Research Institute, University of California, Santa Barbara, CA, 93106,
Telephone: 805-893-8490 Fax: 805-893-2005, Email: [email protected]
Abstract
Human embryonic stem cell derived retinal pigmented epithelium (hESC-RPE) are in
clinical trials for the treatment of macular diseases. Currently, these cells take over three months
to derive and subsequent months to mature and characterize. After only five to six passages the
cells begin to undergo an epithelial to mesenchymal transition and are unsuitable for cellular
therapies. We show that inhibition of Rho coiled coil kinase (ROCK) using a specific inhibitor,
Y-27632, allows extended passage of hESC-RPE in serum-free culture with maintenance of the
RPE phenotype. After 30 population doublings, hESC-RPE at passage 13 maintain normal
karyotype, display typical, polarized epithelial morphology, and continue to express RPE-
specific genes. Passage 13 hESC-RPE show protein localization and secretion patterns similar to
passage 2 cells, and display similar levels of phagocytosis of photoreceptor outer segments.
Microarray analysis from day two shows several key pathways are altered by ROCK inhibition,
including suppression of TGFβ and Wnt signaling. These findings describe a means to greatly
increase the yield of functional hESC-RPE for use in research and clinical trials.
Introduction
Human embryonic stem cells (hESC) offer a potentially unlimited supply of cells for
emerging cell-based therapies. Unfortunately, the process of deriving distinct cell types can be
time consuming and expensive. Furthermore, differentiated cells typically have a finite lifespan
and can only be passaged a limited number of times. Expanding the propagation potential of

stem cell-derived cells would be extremely useful for transplantation procedures and drug
screening efforts.
Clinical trials are currently under way using hESC derived retinal pigmented epithelium
(hESC-RPE) as a treatment for age related macular degeneration (AMD)1,2
. In the developed
world, AMD is the leading cause of blindness in the elderly with over 7.2 million people
afflicted in the US alone3,4
. It is generally agreed that AMD pathology is linked to dysfunction
and death of the RPE, which function to support the photoreceptor cells. Therefore, when the
RPE degenerate, the photoreceptors in turn cannot function and eventually die, resulting in loss
of vision5,6
. There are two forms of disease, exudative (wet) AMD and atrophic (dry) AMD. Wet
AMD can be successfully treated using anti-vascular endothelial growth factor drugs such as
Lucentis, Avastin or Eylea7. However, there is no effective treatment for the dry form, which
progresses to macular patches without RPE and photoreceptors (geographic atrophy), resulting in
extreme visual impairment.
Both hESC-RPE and iPS-RPE are being developed for AMD therapies by multiple
groups, using either cell suspensions or differentiated monolayers on scaffolds (Bharti et al., In
Press). The current preferred derivation process for RPE follows a spontaneous differentiation
protocol involving the removal of b-FGF in continuously adherent cultures of pluripotent stem
cells1,8
. This method can take up to 100 days before enrichment of the pigmented RPE
population followed by several months of additional culture to obtain relatively homogeneous,
mature RPE ready for transplantation. hESC-RPE (and fetal RPE) are passaged every 30 days
and can only be maintained in culture for five to six passages before the cells display abnormal
characteristics and are thought to undergo an epithelial to mesenchymal (EMT) transition9-11
.
These late passage cells lose their pigmented, cobblestone morphology and appear more
fibroblastic, and often senesce and die. Senescence in cultured primary somatic cells is
commonly observed as telomeres shorten and the Hayflick limit is reached12
.
To attempt to overcome this passage limitation, we examined the involvement of Rho-
associated coiled-coil serine/threonine kinase (ROCK) in RPE EMT/senescence. ROCK is
activated by RhoA GTPase and initiates signal transduction cascades to modulate several central
cell functions, including proliferation, apoptosis, cytoskeletal rearrangements, and migration13-15
.
There are two distinct ROCK isoforms, ROCK1 and ROCK2 that are activated by RhoA;
however, specific isoform functions remain poorly defined16
. The RhoA/ROCK pathway has

been studied extensively for its downstream effects on stress fiber formation17,18
and actin
filament stabilization13,19
. Rock inhibition using synthetic compounds is currently being tested
for pulmonary hypertension treatments with positive animal model results15,20,21
. Current
research suggests increasing roles for ROCK inhibitors, including ROCK1/2 inhibitor Y-27632,
in regulating various cell processes15,16,22,23
. Previous studies have showed ROCK inhibition
increased passage abilities of other epithelial cell types24-28
, as well as the clonability of
hESC29,30
; we have shown the beneficial effects of Y-27632 treatment following initial
enrichment of RPE during directed differentiation from hESC31
.
Here we report that inhibiting ROCK1/2 with Y-27632 allows extended passage of
hESC-RPE. Importantly, the resulting cells maintain proper gene expression, protein
localization, function, and karyotype. The simple culture methods described here allow a much
greater yield of hESC-RPE and provide abundant cells for disease modeling, drug screening and
further development of cellular therapies.
Materials and Methods
Cell Culture
Pluripotent Stem Cell Culture
H9 hESC were obtained from the WiCell Research Institute in Madison, WI. hESC were
maintained in mTESR media (STEMCELL Technologies, Vancouver, Canada) and grown on
Matrigel coated plates (BD Biosciences, San Jose, CA, USA) at 37°C 5% CO2.
Continuously Adherent Retinal Pigmented Epithelium Differentiation and Enrichment
H9 hESC were passaged onto Matrigel coated plates and allowed to overgrow for 8-14
days. mTESR media was then changed to XVIVO10 (Lonza, Basal, Switzerland) in the absence
of b-FGF. XVIVO10 was changed every other day for 90 days. Pigmented cells appeared after 4-
6 weeks. After 90 days in XVIVO10, hESC-RPE were enriched by first removing unpigmented
cells via mechanical dissection and then harvesting remaining (mostly pigmented) cells using
TrypLE Express (Life Technologies, Grand Island, NY, USA) for 5 minutes at 37°C 5% CO2.
Cells were passed through a 30µm strainer and replated on Matrigel coated plates creating an

enriched hESC-RPE population. hESC-RPE were maintained on Matrigel at 37°C 5% CO2 and
XVIVO10 was changed every other day. hESC-RPE were passaged every 30 days and seeded at
1.0x105cells/cm
2. Passage 2 (p2) hESC-RPE used for comparisons to passage 13 (p13) hESC-
RPE were cultured in this method.
Extended Passage Protocol of hESC-RPE
Directly following enrichment, hESC-RPE were plated into four Matrigel coated wells at
a lower density of 0.25x105cells/cm
2 in XVIVO10. Two wells were treated with Y-27632
(10µM, Tocris Bioscience, Bristol, UK) and the other two received water, as a control. One day
post confluence (~4-5 days), one Y-27632 treated well and one control well were passaged and
seeded again at 0.25x105cells/cm
2. Passaging continued in this fashion until cells failed to reach
confluence. The two wells not undergoing passaging continued to be supplemented with 10µM
Y-27632 or water for 14 days. On day 30 RNA was harvested. Images were taken on the
Olympus CKX41 (Olympus, Center Valley, PA, USA) on day 30. Passage 13 (p13) hESC-RPE
used in all experiments were derived using this protocol. The extended passage experiment was
performed with seven separate enrichment cultures of H9 hESC-RPE.
Population Doubling Calculation
Cell number was determined at each passage in the extended passage protocol using a
hemocytometer. Population doubling was calculated according to the following equation: 3.32
(log (number of cells counted at time of passage/number of cells plated))28
.
MTT Assay
To determine the effect of ROCK inhibition on hESC-RPE cell proliferation, cells were
plated into 96 well plates in XVIVO10 and allowed to attach for 2 hours. XVIVO10 was then
changed and Y-27632 treatment or water was administered. Media was changed every other day
with continuous 10µM Y-27632 or water treatment. MTT (3-(4,5-Dimethylthiazol-2-yl)-2,5-
Diphenyltetrazolium Bromide) assays were performed on day 2, 4, 10, 15, 20, 25, and 30
according to supplier instructions. A 5mg/ml MTT (Life Technologies) stock solution was added
to cells and incubated for four hours at 37°C 5% CO2. 10mM hydrochloric acid was then added
and incubated for four hours at 37°C 5% CO2. Absorbance was read at 570nm, on a Benchmark

Microplate Reader, (BioRad, Hercules, CA, USA) indicative of the amount of living cells. The
MTT assay was performed with hESC-RPE at passage 4, passaged every 30 days from
enrichment, seeded at a density of 1.0x105cells/cm
2.
Reverse Transcription PCR and Quantitative Real Time PCR (QRT-PCR)
Day 30 RNA was isolated using the Qiagen RNeasy Plus Extraction Kit (Qiagen,
Valencia, CA, USA). The iScript cDNA synthesis kit (BioRad) transformed 1µg RNA into
cDNA for QRT-PCR analysis. The following TaqMan gene expression assay IDs were loaded in
triplicate onto a BioRad QRT-PCR plate (BioRad): RPE specific protein 65kDa (RPE65)
Hs01071462_m1; bestrophin 1 (BEST1) Hs00188249_ml; retinaldehyde binding protein 1
(RLBP1) Hs00165632_ml; microphthalmia-associated transcription factor (MITF) isoform 2
AJD1S3G; premelanosome protein (PMEL) Hs00173854_m1; tyrosinase related protein 1
(TYRP1) Hs00167051_m1; tyrosinase (TYR) Hs00165976_ml; paired box 6 (PAX6)
Hs01088112_m1; marker of proliferation Ki-67 (MKI67) Hs01032443_m1; zinc finger protein
42 (REX1) Hs01124465_m1; spalt-like transcription factor 4 (SALL4) Hs00360675_m1;
microtubule-associated protein 2 (MAP2) Hs00258900_m1; integrin, alpha 2 (ITGA2)
Hs00158127_m1; platelet/endothelial cell adhesion molecule 1 (PECAM1) Hs00169777_m1;
S100 calcium binding protein A4 (S100A4) Hs00243202_m1; and housekeepers: eukaryotic
translation initiation factor 2B, subunit 2 beta (EIF2B2) Hs00204540_m1; ubiquitin-conjugating
enzyme E2R 2 (UBE2R2) Hs00215107_m1; and small EDRK-rich factor 2 (SERF2)
Hs00428481_m1 (Life Technologies). cDNA was combined with TaqMan Gene Expression
Master Mix (Life Technologies) and loaded onto the plate containing the gene expression assay
IDs. Amplification was carried out using a Bio-Rad CFX96 Real-Time System with the CFX
Manager Software and Excel used for analysis. Ct values were normalized to the geometric mean
of the housekeeping gene set.
Immunocytochemistry
Passage 2 and 13 hESC-RPE were seeded onto Matrigel coated 8-chambered slides at
1.0x105cells/cm
2. Passage 13 hESC-RPE were treated with 10µM Y-27632 for 14 days. 45 days
after plating, cells were washed with phosphate-buffered saline (PBS), fixed in 4%
paraformaldehyde in 0.1M sodium cacodylate buffer (pH 7.4) for seven minutes at 4°C. Cells

were washed with PBS and blocked with PBS containing 5% bovine serum albumin (BSA) and
0.2% Triton X-100 for 1 hour at 4°C. Cells were probed with primary antibodies against
MITFC5 (1:1000, Abcam, Cambridge, MA, USA), orthodenticle homeobox 2 (OTX2) (1:4000
Millipore, Billerica, MA, USA), RPE65 (1:100, Abcam), BEST1 (1:100, Abcam), PMEL (1:100,
Abcam), zona occludens (ZO-1) (1:100, Life Technologies), and MKI67 (1:1000, Abcam), in
PBS with 5% BSA overnight at 4°C. Cells were incubated with appropriate Alexa Fluor
conjugated secondary antibody (1:300, Life Technologies) for 1 hour at 4°C. Following
secondary antibody incubation, cells were stained with Hoescht (2µg/ml, Life Technologies) for
5 minutes at room temperature. Cells were then washed with PBS and imaged at room
temperature with the Olympus BX51 fluorescence microscope (Olympus). Five separate
enrichments of hESC-RPE were probed for each passage with this panel of primary antibodies.
Rod Outer Segment Phagocytosis Assay
21-week old human fetal RPE (fRPE), kindly provided by D. Bok (Donor: 042607-3914,
University of California, Los Angeles) were cultured according to Maminishkis et al.32
. fRPE
and hESC-RPE were grown in alpha modified minimum essential medium (MEM-α, Sigma-
Aldrich, St. Louis MO, USA) supplemented with fetal bovine serum (FBS), 15% for the first
three days and 5% for the remaining culture period, (Atlas Biologicals, Fort Collins, CO, USA),
N1 supplement (1X, Sigma-Aldrich), NEAA (1X, Life Technologies, Carlsbad, CA, USA),
GlutaMAX-I (2mM, Life Technologies), taurine (250µg/ml, Sigma-Aldrich), triiodothyronin
(0.013µg/ml, Sigma-Aldrich), and hydrocortisone (20ng/ml, Sigma-Aldrich). The ARPE19 cell
line was cultured in Dublecco’s modified eagle medium with nutrient mix F12 and sodium
pyruvate (DMEM/F12, Life Technologies), supplemented with GlutaMAXI (1X, Life
Technologies), 10% FBS (Atlas Biologicals) and 15mM HEPES (Life Technologies). Human
umbilical vein endothelial cells (HUVEC) were grown in endothelial cell growth medium with
supplement mix (EGM, PromoCell, Heidelberg, Germany). All cells were plated at
1.0x105cells/cm
2 onto 0.1% gelatin-coated, black walled 96 well plates in quadruplicate and
cultured for 30 days. hESC-RPE p13 were treated with 10µM Y-27632 for 14 days.
Rod outer segments (ROS) were isolated from bovine retinal extracts (Sierra Medical,
Whittier, CA, USA) and fluorescently labeled with FluoReporter FITC Protein Labeling Kit
(Life Technologies). hESC-RPE p2, hESC-RPE p13, fRPE, ARPE19 and HUVEC were cultured

as described above. Cells were treated with or without αvβ5 function blocking antibody
(62.5µg/ml, Abcam) or IgG (62.5µg/ml Abcam) isotype control for 30 minutes at 37°C 5% CO2.
Following the initial antibody incubation, the cells were challenged with 1x106
FITC-ROS/well
for 5 hours at 37°C 5% CO231,33
in media supplemented with a fresh aliquot of antibody. After
ROS incubation, cells were washed 6X with PBS and then 0.4% trypan blue was added for 20
minutes to quench external ROS. Each well was imaged using an Olympus inverted fluorescent
IX71 microscope (Olympus) to visualize internalized ROS fluorescence. Integrated pixel density
of photomicrographs was determined using Image J Software (National Institute of Health,
Bethesda, MD) with a rolling pixel radius of 50. Experiment was performed with three different
enrichments of hESC-RPE. fRPE and ARPE19 were positive control lines and the HUVEC line
was used as a negative control. All experiments were normalized to a single ARPE19 ROS
experimental data set.
Pigment Epithelium Derived Factor and Vascular Endothelial Growth Factor Enzyme-Linked
Immunosorbent Assay
hESC-RPE p2 and hESC-RPE p13 were grown on Matrigel coated 0.45µm HA inserts
(0.6cm2, Millipore) seeded at 1.0x10
5cells/cm
2. 10µM Y-27632 was added to hESC-RPE p13
cells for 14 days. On day 30, media was collected from both the apical and basal chambers
following a 72 hour exposure to the cells. Samples were snap frozen in liquid nitrogen and stored
at -80°C until ELISAs were run. Human pigment epithelium derived factor (PEDF) ELISA
(BioProducts MD, LLC., Middletown, MD, USA) and human vascular endothelial growth factor
(VEGF) ELISA (Life Technologies) were run according to manufacturer’s instructions. Both
apical and basal collections were subjected to each ELISA. Absorbance for both was read at
450nm on a VERSAmax tunable microplate reader (Molecular Devices, Sunnyvale, CA, USA).
A standard curve was produced and experimental concentrations were extrapolated.
Karyotype analysis
Karyotyping of passage two and passage 13 hESC-RPE, from the same enrichment, was
performed by Cell Line Genetics. Cells were analyzed prior to reaching confluence.
Agilent Whole Human Genome Microarray

hESC-RPE were cultured following the extended passage protocol. At passage five,
control and Y-27632 lysates were collected two days after plating, prior to reaching confluence.
RNA from four separate biological replicates was isolated using the Qiagen miRNeasy Kit
(Qiagen) for each treatment group. Global transcriptome analysis was performed with the
Agilent Whole Human Genome 4 × 44 K in situ oligonucleotide array platform (G4112F,
Agilent Technologies, Inc., Santa Clara, CA, USA) using the reagents and methods of the
manufacturer.
Results
ROCK inhibition allows extended passage of hESC-RPE
hESC-RPE plated at high density (1.0x105cells/cm
2) can typically only be passaged five
to six times, before senescing and undergoing an epithelial to mesenchymal transition9-11
. We
sought to identify culture conditions that allow extended passage of hESC-RPE and found that
inhibition of ROCK allowed at least 13 passages. When hESC-RPE were seeded at a low density
of 0.25x105cells/cm
2, they can only survive 5 passages before senescing (Fig.1). To test the role
of ROCK in this process, the synthetic compound, Y-27632, which inactivates the ATP binding
domain of both ROCK isoforms15
, was added to the culture medium, and cells were continuously
passaged. For each passage, a portion of cells were grown for 30 days to allow them to mature
and reach confluence, with removal of the ROCK inhibitor at day14. The extended passage cells
were able to assume their typical RPE cobblestone morphology up to passage 14. However, after
15 passages, patches of larger cells displaying a mesenchymal morphology (arrows) were
intermixed with areas of cells with epithelial morphology. By passage 18, cells failed to reach
confluence following 30 days in culture and never regain the typical RPE morphology (Fig. 1).
To determine the effects of Y-27632 on the rate of proliferation, cell numbers were
monitored and population doublings were calculated according to Chapman et al.28
hESC-RPE
were passaged in the presence of Y-27632 at low density, and cell counts were taken at each
passage. Y-27632-treated cells showed a substantial increase in population doublings, allowing
an average of 30 doublings compared to only 9 doublings in control cultures; each point
represents a passage (Fig. 2A). In the presence of the ROCK inhibitor, cell numbers increase
exponentially, with an average doubling time of 2.4 days, whereas controls increase in number

more significantly slower, with an average doubling time of 3.8 days, and cell numbers begin to
plateau as cells begin to senesce after 5 passages.
Y-27632 effects within a single passage, over 30 days, were also monitored using an
MTT assay to quantify living cells. Passage four hESC-RPE, without prior exposure to Y-27632,
were grown in the presence or absence of Y-27632 over a period of 30 days. At various time
points, MTT assays were performed to assess the number of living cells through the amount of
reduced dye, which was quantified by absorbance at 570nm (*p>0.05, **p>0.01) (Fig. 2B).
Treatment with Y-27632 increases the number of viable hESC-RPE significantly beginning at
ten days. This experiment confirmed that ROCK inhibition speeds up the rate of proliferation in
hESC-RPE.
Gene expression during extended passage of hESC-RPE
To gain a better understanding of any alterations in transcript levels in extended passaged
cells, we analyzed each passage after growing cells to confluence. Day 30 control and Y-27632
RNA was collected and subjected to q-PCR. Control hESC-RPE showed a drop in critical RPE
genes (RPE65, BEST1 and CRALBP) beginning at passage three, with significant differences
observed at passage five compared to passage one (**p>0.01) (Fig. 3). In addition, MITF
isoform 2, specific to RPE, decreased with passage in the control. Interestingly, levels of pigment
related mRNAs, PMEL, TYRP1, and TYR, remained constant in control treated hESC-RPE.
PAX6, a neural retina and immature RPE marker, increased over passage, but not significantly.
In Y-27632 treated hESC-RPE, mature RPE genes, RPE65 and BEST1 did not change
significantly between passage one and passage 13. Levels of mRNAs encoding CRALBP and
MITF2 (RPE specific), and PMEL, TYRP1, and TYR (pigment related) showed steady
expression over extended passage. In addition, PAX6 remained low throughout the time course
(Fig. 3).
Importantly MKI67, a marker of mitosis did not increase in Y-27632 treated cells at day
30. MKI67 mRNA levels in Y-27632 treated cells were barely detectable over passage. A slight
increase was detected at passage 13, but it is not significant. Thus, although cells proliferated
more rapidly in the presence of Y-27632 (Fig. 2), once the ROCK inhibitor was removed and

cells attained confluence, increased expression of this proliferation marker was not observed
(Fig. 3).
We also examined markers for pluripotency, and potential contaminating cell types.
mRNA encoding pluripotency markers REX1 and SALL4 remained negligible with extended
passage, as did the neuronal marker MAP2, the smooth muscle marker ITGA2, the endothelial
marker PECAM, and the fibroblastic marker S100A4.
Protein expression and localization during extended passage of hESC-RPE
To examine protein localization in hESC-RPE passage 2 and passage 13, day 30 cells
were fixed and permeabilized for immunocytochemistry. Cells were probed with RPE-specific
antibodies against MITF, OTX2, RPE65, BEST1, PMEL, and ZO-1. Mature RPE markers
BEST1 and RPE65 were both expressed in the cytoplasm of passage 2 and passage 13 cells. In
addition, anti-PMEL was associated with distinct melanosomes and the tight junction marker
ZO-1 was confined to cell borders in passage 2 (Fig. 4, left) and passage 13 cells (Fig. 4, right).
MKI67 expression was not detected in either passage 2 or passage 13 cells.
We also examined the karyotype of passage 2 and passage 13, and both were found to be
normal (Supp. Fig. 1).
Functional analysis of extended passage hESC-RPE
Two major RPE functions are the diurnal phagocytosis of shed photoreceptor outer
segments and the proper apical, basal secretion of PEDF and VEGF, respectively34
. To examine
these functions in extended passage hESC-RPE treated with the ROCK inhibitor, phagocytosis
assays and ELISAs were performed. For phagocytosis assays, hESC-RPE p2, hESC-RPE p13,
negative and positive control cells were challenged with FITC labeled bovine rod outer
segments. Internalization of segments was quantified after trypan blue quenching of extracellular
segments. Passage 13 hESC-RPE showed levels of phagocytosis similar to that seen in passage 2
cells (Fig. 5A). To test whether phagocytosis occurred through the receptor mechanism as RPE
in vivo, cells were incubated with an integrin αvβ5 function blocking antibody prior to
challenging with FITC labeled ROS. This integrin has been shown to be important for RPE
phagocytosis35
. There was a significant decrease in the amount of ROS internalization compared

to IgG control treated cells (*p>0.05) for fRPE, hESC-RPE p2 and hESC-RPE p13 cells after
αvβ5 activity was blocked. This shows that phagocytosis requires the same integrin receptor after
extended passage, and that passage 13 hESC-RPE retain normal phagocytosis activity.
Apical and basal media was collected from hESC-RPE grown in transwells and subjected
to human PEDF and VEGF to analyze factor secretion levels. There was no significant
difference between p2 and p13 hESC-RPE for either apical PEDF release (Fig. 5B) or basal
VEGF secretion (Fig. 5C). Specific polarized release of these factors is showed by the dramatic
decrease of basally detected PEDF and apical VEGF levels; reaching significance between apical
and basal values for passage 2 PEDF release and passage 13 VEGF release (*p>0.05, **p>0.01).
ROCK inhibition affects transcript levels within many cell pathways
ROCK is known to be involved in numerous pathways within the cell, but its effect on
gene transcription has not been fully examined. To gain more insight into how ROCK inhibition
allows for extended passage of hESC-RPE, we assayed global transcription patterns using cDNA
microarray analysis. hESC-RPE RNA was collected two days after passage five from both
control and ROCK inhibited cells. Microarray analysis revealed that around 12,200 mRNAs
changed between control and Y-27632 samples. From those, just over 700 transcripts showed at
least a two-fold change, calculated from the mean ratio of Y-27632 over control; 369 genes
decreased after treatment and 356 increased. (See NCBI GEO for complete data set). Data was
further scrutinized by eliminating all genes where the individual experimental value did not
reach a 1.5 fold change. This removes the impact of potential outliers from affecting the mean
value.
Our microarray data reiterates the published literature that ROCK plays a role in several
different pathways that all could contribute to the ability of hESC-RPE to undergo extended
passage (Table 1)14,19,36,37
. Table 1 summarizes the microarray data that corresponds to the
known Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways containing ROCK and
other pathways that Y-27632 treatment is affecting, according to our data. These pathways
include: Actin Cytoskeleton Rearrangements, Adherens Junctions, Tight Junctions, Cell
Adhesion Molecules, ECM Receptor Interactions/Focal Adhesions, Vascular Smooth Muscle

Contraction, Axon Guidance, Chemokine Guidance, HIPPO Signaling, TGFβ Signaling, Wnt
Signaling and the Cell Cycle.
In Table 1, the left column displays the quantile normalized mean of the four biological
replicates of control and Y-27632 for each gene. The right column indicates a Y-27632:Control
ratio of fold change, and the color corresponds to the amount of quantitative difference. Red
indicates an increase of expression in Y-27632 treated cells, while green portrays a decrease in
transcript levels compared to control.
The data depicted a decrease in tight junctions, cell adhesion molecules, and most
mRNAs encoding proteins involved in ECM receptor interactions and focal adhesion, at day two,
prior to cell confluence, following ROCK inhibition. HIPPO, TGFβ and Wnt signaling analysis
showed transcript suppression of key ligands that are normally involved in pathway activation
after ROCK inhibition. No changes in SMADs were detected, however they are activated
through phosphorylation, therefore more work is needed to uncover whether ROCK inhibition
affects SMAD activity. In addition, transcripts tied to progression through the cell cycle
increased substantially after Y-27632 treatment.
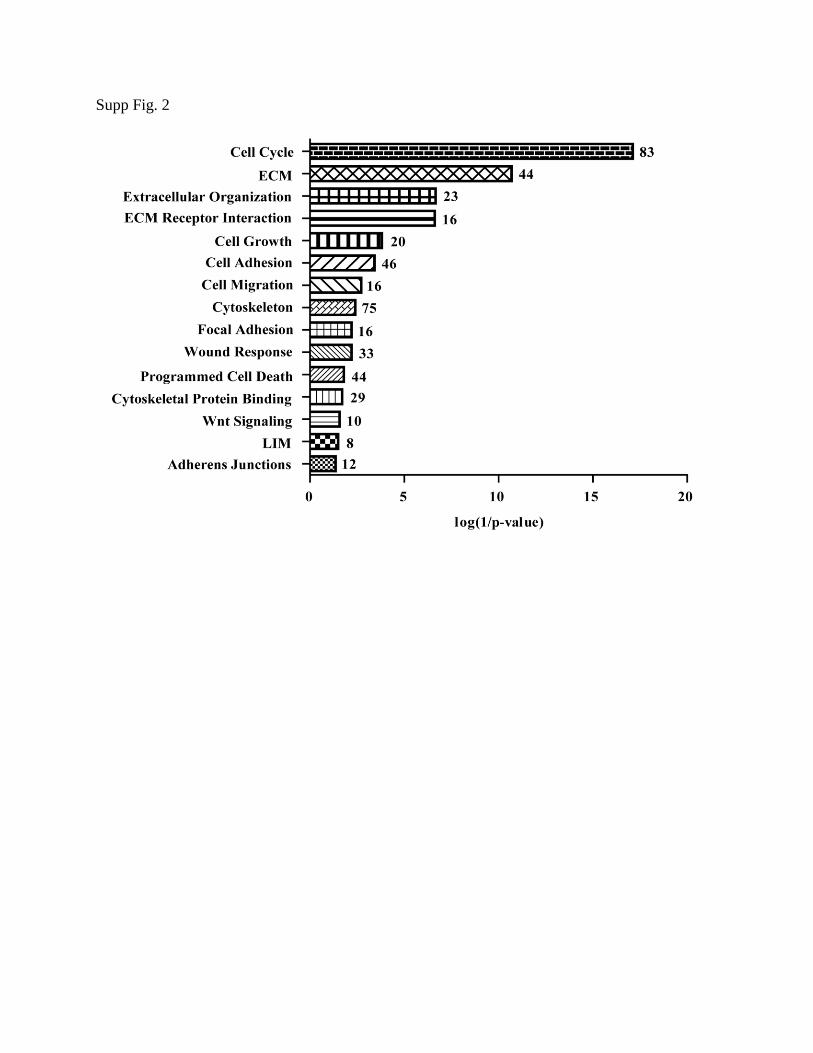
These data were also analyzed by clustering software DAVID38,39
. It revealed several
significant gene groups affected by ROCK inhibition, many which overlap the KEGG pathway
analysis (Supp. Fig. 2).
Discussion
RPE were first derived from hESC in 2004 and since then groups have been attempting to
optimize the process8,40
. The main obstacles of culturing hESC-RPE have always been long
derivation time and limited potential for expansion once differentiated. Recently, progress has
been made in developing more rapid methods for differentiation of hESC into RPE 31,41-43
. For
example, Buchholz et al.31
described a method to direct differentiation using a series of growth
factors known to play a role in RPE differentiation in vivo. However, once RPE are obtained,
they still lack the ability for extended passage.
We report here that inhibition of ROCK using the compound, Y-27632, has the capability
to increase the proliferation and extend the passage of hESC-RPE for up to 13 passages, resulting

in 30 doublings. This phenomenon occurs even when the cells are seeded at a fourth of the
seeding density that is typically used. Thus, one can employ less starting material, and passage
cells longer, leading to a drastic increase in number of cells produced from a single culture of
hESC.
Extended passage hESC-RPE maintained appropriate morphology and gene expression of
key RPE markers without an increase in pluripotent, fibroblastic or endothelial transcripts.
Immunocytochemistry of passage 13 hESC-RPE exhibited proper localization of RPE
transcription factors, proteins involved in pigmentation, tight junctions, and proteins important
for RPE-specific functions. Importantly, no aberrant proliferation was detected at day 30. hESC-
RPE p13 were compared to p2 to ensure functionality through a variety of tests. hESC-RPE p13
internalized bovine outer segments at the same rate as early passage hESC-RPE. hESC-RPE p13
also secreted PEDF apically, and VEGF basally, at values consistent with p2 hESC-RPE. In
addition, the karyotyping of late passage ROCK inhibited hESC-RPE did not reveal any genomic
abnormalities when compared to early passage hESC-RPE.
How does ROCK inhibition lead to extended passage? Microarray analysis suggests that
ROCK inhibition could be suppressing an epithelial to mesenchymal transition (EMT) through
various pathways. These include inhibition of key ligands of the TGFβ pathway, TGFβ1 and
GDF6 44
and Wnt signaling, WNT5A and WNT5B45,46
, along with decreasing levels of collagens
1A1 and 4A2,47,48
known biomarkers of EMT induction.49
RPE are thought to undergo EMT after repeated passages in culture, a main reason why
hESC-RPE have a finite ability to expand, limiting their production for use in transplantation11,50
.
TGFβ signaling has been shown to be the implicated in activating EMT in RPE11,44,51,52
.
Furthermore, several investigations have predicted the Rho/ROCK pathway is involved in
regulating EMT in multiple cell types53-56 , possibly through TGFβ signaling
57. There has also
been evidence linking Wnt activation and β-catenin accumulation to the increased expression of
EMT related genes45,58
. Inhibition of ROCK, through addition of Y-27632 or other synthetic
compounds has already been shown in some cases to reverse this transition59,60
. Our results are
consistent with the idea that decreased expression of TGFβ and Wnt signaling transcripts lead to
the suppression of EMT.

Normally, it appears hESC-RPE undergo EMT after five to six passages; however if
ROCK is inhibited, two important processes are affected allowing for an increase in hESC-RPE
expansion. First, ROCK inhibition promotes proliferation by inducing multiple components
involved in cell cycle progression. This allows RPE to quickly reform tight junctions, which is
critical to RPE health61
. Second, ROCK inhibition affects many pathways that could be
converging to suppress RPE EMT. This allows hESC-RPE to remain functional for an extended,
but finite, period in culture. Future experiments will elucidate the components involved in these
pathways in RPE and other cell types.
The prevalence of AMD for all individuals over the age of 40 in the US alone was
calculated to be 7.2 million people. That number is projected to increase 50% by 2020 totaling to
11.1 million4. Assuming that a 3 by 5 mm patch of a hESC-RPE monolayer on a scaffold might
be developed for dry AMD patients, it is estimated that 300,000 hESC-RPE cells will be required
for each patient. These numbers comes from determining the actual size of the macula, and then
striving to transplant a patch that covers the same area, seeded at confluence. The macula is
around 2.35mm2, and seeding at 1.0x10
5cells/cm
2, yields 235,000 cells needed for a single
transplantation62
. Slightly overestimating to 300,000 and multiplying by the number of patients
expected in 2020, results in 3.3x1012
cells required to treat all those affected. Following the
extended passage protocol, a single starting source of 237,500 hESC-RPE, the number of cells
plated from enrichment into a single well of a 6 well plate, will reach 4.0x1012
cells following 24
population doublings. According to the population doublings graph, it will only take ~55 days to
reach this amount and generate enough cells to treat all those diagnosed with the disease in 2020.
Compared to the current method of derivation, the extended passage protocol using ROCK
inhibition has the potential to increase the yield of mature hESC-RPE by over 5000 times within
the first 60 days, in addition to using ¼ of the starting cell number.
From a cell engineering perspective, these discoveries will facilitate the production of
functional hESC-RPE and possibly other epithelial cells derived from pluripotent stem cells, for
transplantation purposes. Currently, passage 2 or 3 hESC-RPE are being employed in clinical
trials. Longer passaged cells deserve further experimentation to characterize their efficacy and
potential tumorigenicity in animal studies. In addition, defining the mechanisms of EMT and
devising methods to inhibit this process could be extremely beneficial in creating therapeutics for

other RPE dysfunction diseases such as proliferative vitreal retinopathy, which is characterized
by RPE EMT followed by retinal detachment and loss of vision63
.
Conclusions
Inhibition of ROCK through addition of the synthetic compound Y-27632 allows
extended passage of hESC-RPE. This effect seems, in part, to be achieved through the inhibition
of an epithelial to mesenchymal transition normally seen after 5 passages. We have shown that
inhibiting ROCK allows hESC-RPE to be seeded at a fourth the normal density and grow for up
to 13 passages. The extended passaged hESC-RPE are comparable to untreated early passage
hESC-RPE by all parameters tested. This simple technique, in combination with the published
rapid differentiation protocols, could lead to a faster and more efficient way of producing hESC-
RPE for clinical trials, basic disease research and drug screening.
Acknowledgments
This work was supported by the Garland Initiative for Vision, grant DR1-0144 from the
California Institute for Regenerative Medicine, Fight for Sight, The Foundation Fighting
Blindness Wynn-Gund Translational Research Acceleration Program, and by the UCSB Institute
for Collaborative Biotechnologies through grant W911NF-09-0001 from the U.S. Army
Research Office. The content of the information does not necessarily reflect the position or the
policy of the Government, and no official endorsement should be inferred. We would like to
thank Carolyn Radeke for technical support while performing the microarray and Dean Bok for
the generous gift of fetal RPE cells.
Disclosure Statement
Dennis O. Clegg is a co-founder of Regenerative Patch Technologies, LLC.

References
1 Croze, R. H. and Clegg, D. O. Cell-Based Therapy for Retinal Degenerative Disease in Developmental Opthalmology Vol. 53 (ed Zarbin MA Casaroli-Marano RP) 81-96 (Karger, 2014).
2 Schwartz, S. D. et al. Embryonic stem cell trials for macular degeneration: a preliminary report. Lancet 379, 713-720, doi:10.1016/S0140-6736(12)60028-2 (2012).
3 Klein, R. et al. Prevalence of age-related macular degeneration in the US population. Archives of ophthalmology 129, 75-80, doi:10.1001/archophthalmol.2010.318 (2011).
4 Friedman, D. S. et al. Prevalence of age-related macular degeneration in the United States. Archives of ophthalmology 122, 564-572, doi:10.1001/archopht.122.4.564 (2004).
5 Curcio, C. A., Medeiros, N. E. & Millican, C. L. Photoreceptor loss in age-related macular degeneration. Investigative ophthalmology & visual science 37, 1236-1249 (1996).
6 Young, R. W. Pathophysiology of age-related macular degeneration. Survey of ophthalmology 31, 291-306 (1987).
7 Gehrs, K. M., Anderson, D. H., Johnson, L. V. & Hageman, G. S. Age-related macular degeneration--emerging pathogenetic and therapeutic concepts. Annals of medicine 38, 450-471, doi:10.1080/07853890600946724 (2006).
8 Rowland, T. J., Buchholz, D. E. & Clegg, D. O. Pluripotent human stem cells for the treatment of retinal disease. Journal of cellular physiology 227, 457-466, doi:10.1002/jcp.22814 (2012).
9 Burke, J. M. Epithelial phenotype and the RPE: is the answer blowing in the Wnt? Progress in retinal and eye research 27, 579-595, doi:10.1016/j.preteyeres.2008.08.002 (2008).
10 Grisanti, S. & Guidry, C. Transdifferentiation of retinal pigment epithelial cells from epithelial to mesenchymal phenotype. Investigative ophthalmology & visual science 36, 391-405 (1995).
11 Lee, S. C. et al. Epitheliomesenchymal transdifferentiation of cultured RPE cells. Ophthalmic research 33, 80-86, doi:55648 (2001).
12 Shay, J. W. & Wright, W. E. Hayflick, his limit, and cellular ageing. Nature reviews. Molecular cell biology 1, 72-76, doi:10.1038/35036093 (2000).
13 Riento, K. & Ridley, A. J. Rocks: multifunctional kinases in cell behaviour. Nature reviews. Molecular cell biology 4, 446-456, doi:10.1038/nrm1128 (2003).
14 Amano, M., Fukata, Y. & Kaibuchi, K. Regulation and functions of Rho-associated kinase. Experimental cell research 261, 44-51, doi:10.1006/excr.2000.5046 (2000).
15 Liao, J. K., Seto, M. & Noma, K. Rho kinase (ROCK) inhibitors. Journal of cardiovascular pharmacology 50, 17-24, doi:10.1097/FJC.0b013e318070d1bd (2007).
16 Zhou, Q., Gensch, C. & Liao, J. K. Rho-associated coiled-coil-forming kinases (ROCKs): potential targets for the treatment of atherosclerosis and vascular disease. Trends in pharmacological sciences 32, 167-173, doi:10.1016/j.tips.2010.12.006 (2011).
17 Leung, T., Chen, X. Q., Manser, E. & Lim, L. The p160 RhoA-binding kinase ROK alpha is a member of a kinase family and is involved in the reorganization of the cytoskeleton. Molecular and cellular biology 16, 5313-5327 (1996).
18 Somlyo, A. P. & Somlyo, A. V. Signal transduction by G-proteins, rho-kinase and protein phosphatase to smooth muscle and non-muscle myosin II. The Journal of physiology 522 Pt 2, 177-185 (2000).
19 Street, C. A. & Bryan, B. A. Rho kinase proteins--pleiotropic modulators of cell survival and apoptosis. Anticancer research 31, 3645-3657 (2011).
20 Doggrell, S. A. Rho-kinase inhibitors show promise in pulmonary hypertension. Expert opinion on investigational drugs 14, 1157-1159, doi:10.1517/13543784.14.9.1157 (2005).

21 Pankey, E. A. et al. The Rho kinase inhibitor azaindole-1 has long-acting vasodilator activity in the pulmonary vascular bed of the intact chest rat. Canadian journal of physiology and pharmacology 90, 825-835, doi:10.1139/y2012-061 (2012).
22 Okumura, N. et al. Enhancement on primate corneal endothelial cell survival in vitro by a ROCK inhibitor. Investigative ophthalmology & visual science 50, 3680-3687, doi:10.1167/iovs.08-2634 (2009).
23 Koizumi, N., Okumura, N. & Kinoshita, S. Development of new therapeutic modalities for corneal endothelial disease focused on the proliferation of corneal endothelial cells using animal models. Experimental eye research 95, 60-67, doi:10.1016/j.exer.2011.10.014 (2012).
24 Liu, X. et al. ROCK inhibitor and feeder cells induce the conditional reprogramming of epithelial cells. The American journal of pathology 180, 599-607, doi:10.1016/j.ajpath.2011.10.036 (2012).
25 Horani, A., Nath, A., Wasserman, M. G., Huang, T. & Brody, S. L. Rho-associated protein kinase inhibition enhances airway epithelial Basal-cell proliferation and lentivirus transduction. American journal of respiratory cell and molecular biology 49, 341-347, doi:10.1165/rcmb.2013-0046TE (2013).
26 Miyashita, H. et al. Long-term maintenance of limbal epithelial progenitor cells using rho kinase inhibitor and keratinocyte growth factor. Stem cells translational medicine 2, 758-765, doi:10.5966/sctm.2012-0156 (2013).
27 van den Bogaard, E. H. et al. Rho kinase inhibitor Y-27632 prolongs the life span of adult human keratinocytes, enhances skin equivalent development, and facilitates lentiviral transduction. Tissue engineering. Part A 18, 1827-1836, doi:10.1089/ten.TEA.2011.0616 (2012).
28 Chapman, S., Liu, X., Meyers, C., Schlegel, R. & McBride, A. A. Human keratinocytes are efficiently immortalized by a Rho kinase inhibitor. The Journal of clinical investigation 120, 2619-2626, doi:10.1172/JCI42297 (2010).
29 Ohgushi, M. et al. Molecular pathway and cell state responsible for dissociation-induced apoptosis in human pluripotent stem cells. Cell stem cell 7, 225-239, doi:10.1016/j.stem.2010.06.018 (2010).
30 Ohgushi, M. & Sasai, Y. Lonely death dance of human pluripotent stem cells: ROCKing between metastable cell states. Trends in cell biology 21, 274-282, doi:10.1016/j.tcb.2011.02.004 (2011).
31 Buchholz, D. E. et al. Rapid and efficient directed differentiation of human pluripotent stem cells into retinal pigmented epithelium. Stem cells translational medicine 2, 384-393, doi:10.5966/sctm.2012-0163 (2013).
32 Maminishkis, A. et al. Confluent monolayers of cultured human fetal retinal pigment epithelium exhibit morphology and physiology of native tissue. Investigative ophthalmology & visual science 47, 3612-3624, doi:10.1167/iovs.05-1622 (2006).
33 Rowland, T. J. et al. Differentiation of human pluripotent stem cells to retinal pigmented epithelium in defined conditions using purified extracellular matrix proteins. Journal of tissue engineering and regenerative medicine 7, 642-653, doi:10.1002/term.1458 (2013).
34 Strauss, O. The retinal pigment epithelium in visual function. Physiological reviews 85, 845-881, doi:10.1152/physrev.00021.2004 (2005).
35 Lin, H. & Clegg, D. O. Integrin alphavbeta5 participates in the binding of photoreceptor rod outer segments during phagocytosis by cultured human retinal pigment epithelium. Investigative ophthalmology & visual science 39, 1703-1712 (1998).
36 Duong-Quy, S., Bei, Y., Liu, Z. & Dinh-Xuan, A. T. Role of Rho-kinase and its inhibitors in pulmonary hypertension. Pharmacology & therapeutics 137, 352-364, doi:10.1016/j.pharmthera.2012.12.003 (2013).
37 Amano, M., Nakayama, M. & Kaibuchi, K. Rho-kinase/ROCK: A key regulator of the cytoskeleton and cell polarity. Cytoskeleton 67, 545-554, doi:10.1002/cm.20472 (2010).

38 Huang da, W., Sherman, B. T. & Lempicki, R. A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nature protocols 4, 44-57, doi:10.1038/nprot.2008.211 (2009).
39 Huang da, W., Sherman, B. T. & Lempicki, R. A. Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucleic acids research 37, 1-13, doi:10.1093/nar/gkn923 (2009).
40 Klimanskaya, I. et al. Derivation and comparative assessment of retinal pigment epithelium from human embryonic stem cells using transcriptomics. Cloning and stem cells 6, 217-245, doi:10.1089/clo.2004.6.217 (2004).
41 Idelson, M. et al. Directed differentiation of human embryonic stem cells into functional retinal pigment epithelium cells. Cell stem cell 5, 396-408, doi:10.1016/j.stem.2009.07.002 (2009).
42 Zahabi, A. et al. A new efficient protocol for directed differentiation of retinal pigmented epithelial cells from normal and retinal disease induced pluripotent stem cells. Stem cells and development 21, 2262-2272, doi:10.1089/scd.2011.0599 (2012).
43 Zhu, Y. et al. Three-dimensional neuroepithelial culture from human embryonic stem cells and its use for quantitative conversion to retinal pigment epithelium. PloS one 8, e54552, doi:10.1371/journal.pone.0054552 (2013).
44 Lee, S. C., Kim, S. H., Koh, H. J. & Kwon, O. W. TGF-betas synthesized by RPE cells have autocrine activity on mesenchymal transformation and cell proliferation. Yonsei medical journal 42, 271-277 (2001).
45 Son, H. & Moon, A. Epithelial-mesenchymal Transition and Cell Invasion. Toxicological research 26, 245-252, doi:10.5487/TR.2010.26.4.245 (2010).
46 Xiao, L., Wang, M., Yang, S., Liu, F. & Sun, L. A Glimpse of the Pathogenetic Mechanisms of Wnt/ -Catenin Signaling in Diabetic Nephropathy. BioMed research international 2013, 987064, doi:10.1155/2013/987064 (2013).
47 Billings, P. C. et al. The transforming growth factor-beta-inducible matrix protein (beta)ig-h3 interacts with fibronectin. The Journal of biological chemistry 277, 28003-28009, doi:10.1074/jbc.M106837200 (2002).
48 Kim, J. E. et al. Molecular properties of wild-type and mutant betaIG-H3 proteins. Investigative ophthalmology & visual science 43, 656-661 (2002).
49 Scanlon, C. S., Van Tubergen, E. A., Inglehart, R. C. & D'Silva, N. J. Biomarkers of epithelial-mesenchymal transition in squamous cell carcinoma. Journal of dental research 92, 114-121, doi:10.1177/0022034512467352 (2013).
50 Tamiya, S., Liu, L. & Kaplan, H. J. Epithelial-mesenchymal transition and proliferation of retinal pigment epithelial cells initiated upon loss of cell-cell contact. Investigative ophthalmology & visual science 51, 2755-2763, doi:10.1167/iovs.09-4725 (2010).
51 Stocks, S. Z., Taylor, S. M. & Shiels, I. A. Transforming growth factor-beta1 induces alpha-smooth muscle actin expression and fibronectin synthesis in cultured human retinal pigment epithelial cells. Clinical & experimental ophthalmology 29, 33-37 (2001).
52 Gamulescu, M. A. et al. Transforming growth factor beta2-induced myofibroblastic differentiation of human retinal pigment epithelial cells: regulation by extracellular matrix proteins and hepatocyte growth factor. Experimental eye research 83, 212-222, doi:10.1016/j.exer.2005.12.007 (2006).
53 Hu, Y. B. et al. Roles of Rho/Rock signaling pathway in silica-induced epithelial-mesenchymal transition in human bronchial epithelial cells. Biomedical and environmental sciences : BES 26, 571-576, doi:10.3967/0895-3988.2013.07.008 (2013).

54 Wei, J. et al. Rho kinase pathway is likely responsible for the profibrotic actions of aldosterone in renal epithelial cells via inducing epithelial-mesenchymal transition and extracellular matrix excretion. Cell biology international 37, 725-730, doi:10.1002/cbin.10082 (2013).
55 Clay, M. R. & Halloran, M. C. Rho activation is apically restricted by Arhgap1 in neural crest cells and drives epithelial-to-mesenchymal transition. Development 140, 3198-3209, doi:10.1242/dev.095448 (2013).
56 Zhang, H., Liu, X., Liu, Y., Yi, B. & Yu, X. Epithelial-mesenchymal transition of rat peritoneal mesothelial cells via Rhoa/Rock pathway. In vitro cellular & developmental biology. Animal 47, 165-172, doi:10.1007/s11626-010-9369-0 (2011).
57 Cho, H. J. & Yoo, J. Rho activation is required for transforming growth factor-beta-induced epithelial-mesenchymal transition in lens epithelial cells. Cell biology international 31, 1225-1230, doi:10.1016/j.cellbi.2007.04.006 (2007).
58 Azar, K. M., Xiao, L. & Ma, J. Baseline obesity status modifies effectiveness of adapted diabetes prevention program lifestyle interventions for weight management in primary care. BioMed research international 2013, 191209, doi:10.1155/2013/191209 (2013).
59 Gu, L., Gao, Q., Ni, L., Wang, M. & Shen, F. Fasudil inhibits epithelial-myofibroblast transdifferentiation of human renal tubular epithelial HK-2 cells induced by high glucose. Chemical & pharmaceutical bulletin 61, 688-694 (2013).
60 Das, S., Becker, B. N., Hoffmann, F. M. & Mertz, J. E. Complete reversal of epithelial to mesenchymal transition requires inhibition of both ZEB expression and the Rho pathway. BMC cell biology 10, 94, doi:10.1186/1471-2121-10-94 (2009).
61 Rizzolo, L. J. Development and role of tight junctions in the retinal pigment epithelium. International review of cytology 258, 195-234, doi:10.1016/S0074-7696(07)58004-6 (2007).
62 Tribukait, A., Rosenhall, U. & Osterdahl, B. Morphological characteristics of the human macula sacculi. Audiology & neuro-otology 10, 90-96, doi:10.1159/000083364 (2005).
63 Chen, H. C., Zhu, Y. T., Chen, S. Y. & Tseng, S. C. Wnt signaling induces epithelial-mesenchymal transition with proliferation in ARPE-19 cells upon loss of contact inhibition. Laboratory investigation; a journal of technical methods and pathology 92, 676-687, doi:10.1038/labinvest.2011.201 (2012).

Figure Legends
Figure 1. Effect of ROCK inhibition on hESC-RPE passage. Phase contrast images are shown of
control and Y-27632-treated hESC-RPE, with passage number indicated in the upper right. All
images taken on day 30. Images are from a single experiment that is representative of nine
experiments. Arrows indicate cells with mesenchymal phenotype at passage 16 and 17. Scale
bar=100µm.
Figure 2. ROCK inhibition affects hESC-RPE proliferation. (A) Population doubling is plotted
versus time in cultures with and without addition of Y-27632. Each point represents a passage
and when a calculation was made. n=5. (B) P4 hESC-RPE grown in the presence or absence of
Y-27632 and cell number was quantified by measuring MTT reduction. Error bars represent ±
SEM. *p≤0.05 **p≤0.01 compared to control for the same time point. n=3 (same enrichment).
Figure 3. Gene expression in extended passage hESC-RPE. RPE specific, pigmentation, neural
retina/immature RPE, cell cycle, pluripotent and non-RPE genes were analyzed from day 30
RNA at various passages. All data was normalized to geometric mean of three housekeeper
mRNAs. Error bars represent ± SEM. *p≤0.05 **p≤0.01 compared to passage one within the
same treatment group. n=3.
Figure 4. Protein expression and localization in extended passage hESC-RPE. Passage 2 (left
panel) and passage13 (right panel) cells were stained for RPE markers after reaching confluence
at day 30. Scale bars=100µm. Images shown are representatives of four experiments for each
passage.
Figure 5. Function of extended passage hESC-RPE. (A) RPE phagocytosis of bovine
photoreceptor outer segments, as determined by pixel density analysis of photomicrographs, is
shown. fRPE and HUVECs serve as positive and negative controls, respectively. All
experiments are normalized to a control ARPE19 cell line included in each experiment. Isotype
matched IgG was added as a control for the αvβ5 function blocking antibody. (B,C) ELISA
analysis of PEDF and VEGF. Polarized hESC-RPE showed an increase in apical PEDF and basal
VEGF secretion, consistent between passages. Error bars represent ± SEM. *p≤0.05 (A)
compared to samples treated with anti-αvβ5 (B,C) compared to secretion from opposite cell side.
n=3 for all experiments.
Table 1. Changes in gene expression resulting from ROCK inhibition. Microarray data
comparing gene expression differences between control and Y-27632 treated hESC-RPE at day
two passage five. The KEGG pathways containing ROCK and those affected by Y-237632
addition are represented. Control and Y-27632 numerical values on the left represent the mean
quantile normalized data of four biological replicates. Color coded ratio on the right represents
log2(Y-27632/Control), 1=2 fold change. n=4.

Supplemental Figure 1. Cytogenetic analysis reveals normal human female 46, XX karyotype
for hESC-RPE passage 3 (left panel) and passage 13 (right panel) from the same enrichment. Y-
27632 treatment does not create chromosome abnormalities.
Supplemental Figure 2. The microarray data was transformed using the ratio: log2(Y-
27632/Control) and put into the DAVID clustering software. Cell processes were graphed
according to p-value. The number of genes within the cluster is indicated at the right of each bar.
Figure 1.

Figure 2.
Figure 3.

Figure 4.

Figure 5.

Table 1.

Supp Fig. 1
Supp Fig. 2
Copyright © 2022 FDOKUMEN




















