
Taxonomic characterization and plant colonizing abilities of some bacteria related to Bacillus...
11
Taxonomic characterization and plant colonizing abilities of some bacteria related to Bacillus amyloliquefaciens and Bacillus subtilis Oleg N. Reva a , Christina Dixelius b , Johan Meijer b , Fergus G. Priest c, * a Department of Antibiotics, Institute of Microbiology and Virology, 154 Zabolotnogo Str., 03143 Kyiv, Ukraine b Department of Plant Biology and Forest Genetics, Genetics Centre, Box 7080, Swedish University of Agricultural Sciences, SE-750 07 Uppsala, Sweden c School of Life Sciences, Heriot-Watt University, Riccarton, Edinburgh, Scotland EH14 4AS, UK Received 12 November 2003; received in revised form 20 January 2004; accepted 2 February 2004 First published online 1 March 2004 Abstract The phylogenetic relationships of 17 Bacillus strains isolated from plants and soil were determined from partial sequences of genes encoding 16S rRNA, gyraseA (gyrA) and the cheA histidine kinase. Five strains were closely related to Bacillus subtilis subsp. subtilis, three strains were more closely related to B. subtilis subsp. spizizeni and two strains were identified as B. mojavensis. The remaining seven strains formed a cluster closely related to, but distinct from, Bacillus amyloliquefaciens. Some of these strains formed red-pigmented colonies. The abilities of selected strains to survive in the rhizosphere and to colonize plants were studied using oilseed rape (Brassica napus), barley (Hordeum vulgare) and thale cress (Arabidopsis thaliana) as model plants. It was shown by following the titre of bacteria in seedlings and by scanning electron microscopy that survival of Bacillus cells on the roots of seedlings during the first week after treatment of seeds with spore suspensions was crucial for colonization of the rhizosphere and for biocontrol activity. The group of strains related to B. amyloliquefaciens were generally better adapted to colonization of the rhi- zosphere of plants than other members of the B. subtilis group and could be considered a distinct ecotype of B. amyloliquefaciens. Bacteria in this taxon could be recognized on the basis of amplification of a PCR product with primers directed to the tetB(L) locus but no product with primers directed to the a-amylase gene of B.amyloliquefaciens sensu stricto. Ó 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. Keywords: Bacillus amyloliquefaciens; Bacillus subtilis; 16S rRNA; gyrA; cheA; Identification; Plant colonization 1. Introduction It is generally recognized that the tissues of healthy plants can be colonized internally by microorganisms. The use of biopesticides based on these natural endo- phytic bacteria to control plant diseases is considered as a promising ecological alternative to chemical treat- ments. Several attempts to manipulate the development of plant-associated microbial communities of crop plants in order to protect the plants from disease or to promote plant growth have been made [1–3]. Usually these experiments involve the introduction of a large population of a specific microorganism to the seeds or to a furrow during planting [4–6]. Strains of Bacillus have several advantages over other biocontrol bacteria in that they are easy to cultivate and store. They can be applied as spores on plant seeds or in inoculants and they show significant protective effects towards several microbial pathogens as well as plant growth promoting activities. In particular, strains of Bacillus amyloliquefaciens and Bacillus subtilis have been shown to synthesize plant growth promoting substances such as gibberellins and * Corresponding author. Tel.: +44-131-451-3187/3464; fax: +44-131- 451-3009. E-mail address: [email protected] (F.G. Priest). 0168-6496/$22.00 Ó 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.femsec.2004.02.003 FEMS Microbiology Ecology 48 (2004) 249–259 www.fems-microbiology.org
Transcript of Taxonomic characterization and plant colonizing abilities of some bacteria related to Bacillus...

FEMS Microbiology Ecology 48 (2004) 249–259
www.fems-microbiology.org
Taxonomic characterization and plant colonizing abilities ofsome bacteria related to Bacillus amyloliquefaciens
and Bacillus subtilis
Oleg N. Reva a, Christina Dixelius b, Johan Meijer b, Fergus G. Priest c,*
a Department of Antibiotics, Institute of Microbiology and Virology, 154 Zabolotnogo Str., 03143 Kyiv, Ukraineb Department of Plant Biology and Forest Genetics, Genetics Centre, Box 7080, Swedish University of Agricultural Sciences,
SE-750 07 Uppsala, Swedenc School of Life Sciences, Heriot-Watt University, Riccarton, Edinburgh, Scotland EH14 4AS, UK
Received 12 November 2003; received in revised form 20 January 2004; accepted 2 February 2004
First published online 1 March 2004
Abstract
The phylogenetic relationships of 17 Bacillus strains isolated from plants and soil were determined from partial sequences of
genes encoding 16S rRNA, gyraseA (gyrA) and the cheA histidine kinase. Five strains were closely related to Bacillus subtilis subsp.
subtilis, three strains were more closely related to B. subtilis subsp. spizizeni and two strains were identified as B. mojavensis. The
remaining seven strains formed a cluster closely related to, but distinct from, Bacillus amyloliquefaciens. Some of these strains
formed red-pigmented colonies. The abilities of selected strains to survive in the rhizosphere and to colonize plants were studied
using oilseed rape (Brassica napus), barley (Hordeum vulgare) and thale cress (Arabidopsis thaliana) as model plants. It was shown by
following the titre of bacteria in seedlings and by scanning electron microscopy that survival of Bacillus cells on the roots of
seedlings during the first week after treatment of seeds with spore suspensions was crucial for colonization of the rhizosphere and for
biocontrol activity. The group of strains related to B. amyloliquefaciens were generally better adapted to colonization of the rhi-
zosphere of plants than other members of the B. subtilis group and could be considered a distinct ecotype of B. amyloliquefaciens.
Bacteria in this taxon could be recognized on the basis of amplification of a PCR product with primers directed to the tetB(L) locus
but no product with primers directed to the a-amylase gene of B.amyloliquefaciens sensu stricto.
� 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: Bacillus amyloliquefaciens; Bacillus subtilis; 16S rRNA; gyrA; cheA; Identification; Plant colonization
1. Introduction
It is generally recognized that the tissues of healthy
plants can be colonized internally by microorganisms.
The use of biopesticides based on these natural endo-
phytic bacteria to control plant diseases is considered as
a promising ecological alternative to chemical treat-ments. Several attempts to manipulate the development
of plant-associated microbial communities of crop
* Corresponding author. Tel.: +44-131-451-3187/3464; fax: +44-131-
451-3009.
E-mail address: [email protected] (F.G. Priest).
0168-6496/$22.00 � 2004 Federation of European Microbiological Societies
doi:10.1016/j.femsec.2004.02.003
plants in order to protect the plants from disease or to
promote plant growth have been made [1–3]. Usually
these experiments involve the introduction of a large
population of a specific microorganism to the seeds or to
a furrow during planting [4–6]. Strains of Bacillus have
several advantages over other biocontrol bacteria in that
they are easy to cultivate and store. They can be appliedas spores on plant seeds or in inoculants and they show
significant protective effects towards several microbial
pathogens as well as plant growth promoting activities.
In particular, strains of Bacillus amyloliquefaciens and
Bacillus subtilis have been shown to synthesize plant
growth promoting substances such as gibberellins and
. Published by Elsevier B.V. All rights reserved.

250 O.N. Reva et al. / FEMS Microbiology Ecology 48 (2004) 249–259
indole-acetic acid [7], extracellular phytase [8], chitinase
[9,10] and antifungal peptides [11–14]. Seed inoculants
containing strains of B. subtilis and related species have
been shown to increase yields of various crops [1,5,
15–17].Recent taxonomic studies have revealed that B. sub-
tilis is heterogeneous and should be considered as a
complex of closely related species. Thus, isolates previ-
ously classified as B. subtilis are now recognized as Ba-
cillus atrophaeus, Bacillus mojavensis, and Bacillus
vallismortis [18–20] and B. subtilis in turn has been di-
vided into two subspecies B. subtilis subsp. subtilis and
B. subtilis subsp. spizizenii [21]. B. subtilis and B. amy-
loliquefaciens are phenotypically similar species and can
easily be confused. It is therefore not surprising that
most large-scale studies of the endophytic microflora
refer to isolates as B. subtilis without deeper analysis of
their taxonomy. However, definitive identification is
important because biocontrol and plant promotion ac-
tivities including antibacterial and antifungal polypep-
tide production, and synthesis of extracellular phytaseand chitinase, are strain-specific and may be associated
with certain species and subspecies of B. subtilis sensu
lato [8,12,14].
In this study we have used partial 16S rRNA, gyrA
and cheA sequences to assign several endophytic Bacillus
strains to a group closely associated with B. amyloliq-
uefaciens and with many of the features of a distinct
ecotype. We further show that selected strains, in par-ticular those of the B. amyloliquefaciens ecotype, colo-
nize plant roots effectively and can apparently withstand
the antibacterial effects of plant root exudates.
2. Materials and methods
2.1. Bacterial strains
The bacterial strains used in this study and the
GenBank accession numbers of corresponding se-
quences are listed in Table 1. The collection included
isolates from plants and soil (Ukranian Collection of
Microorganisms, UCM strains) which had previously
been identified to the B. subtilis/B. amyloliquefaciens
group on the basis of phenotype [22,23]. A further fivestrains of Bacillus were isolated from Arabidopsis thali-
ana seedlings which had been surface sterilized as seeds
and germinated under gnotobiotic conditions (see At
strains in Table 1). Bacteria were grown at 37 �C on
nutrient agar (Oxoid).
The antagonistic activity of Bacillus strains was tested
in vitro on potato dextrose agar (PDA). A spore sus-
pension of Leptosphaeria maculans (100 ll of 107
sporesml�1) was spread upon the plates and incubated
for 2 days at 28 �C. Bacillus cultures were inoculated on
to the centre of the same plates using a microbiological
loop and the plates incubated for 4 days at 28 �C. Zonesof inhibition of fungal growth were measured.
2.2. Plant growth conditions and treatments
Oilseed rape (Brassica napus L. cv. Westar, suscepti-
ble to Leptosphaeria maculans), barley (Hordeum vulgare
cv. Optic) and thale cress (Arabidopsis thaliana An-1),
sensitive to the phytopathogenic fungus L. maculans [24]
were used in the study.
Seeds were surface sterilized with 10% NaHClO4 for
20 min, rinsed briefly with ethanol, washed with sterile
distilled water and treated for 2 h with Bacillus sporesuspension (approximately 107 spores ml�1) produced
from cultures grown on nutrient agar at 37 �C for 4 days
generally achieving more than 99% sporulation as
judged by microscopy. Culture was removed from the
plate with sterile distilled water and adjusted to the de-
sired titre with distilled water. To determine the growth
colonization ability of Bacillus strains, plants were
grown for 4 days in a climate chamber with 70% relativehumidity and 16 h daylight at 21 �C and 8 h darkness at
17 �C in plastic vessels with Silvaperl (William Sinclair
Horticulture Ltd., Lincoln) (10 g of Silvaperl soaked
with 70 ml of liquid MS medium and sterilized at 115 �Cfor 30 min), or in pots with twice autoclaved standard
sphagnum based soil. In all plant colonization experi-
ments we used 12 plants in each test group and a neg-
ative control group (seeds treated with sterile distilledwater). Parts of roots, steams, leaves, flowers and pods
were weighed, crushed, re-suspended in 1 ml of water,
heated at 60 �C for 20 min to remove non-sporeforming
bacteria and plated on nutrient agar. Mean values and
standard deviations were calculated.
The PG2 isolate PHW 950:14 (Leroy) of L. maculans
[25] was used for plant inoculations. The fungus was
grown on potato dextrose agarose (PDA, Sigma) fortwo weeks before harvesting pycnidiospores. To assess
the protective effects of Bacillus strains, seeds of A.
thaliana (An-1) were surface sterilized as above and
treated with Bacillus spore suspension mixed with L.
maculans pycnidiospores in proportion Bacillus/L.
maculans 108/107 and 107/105 sporesml�1. In positive
and negative controls, seeds were treated with sterile
water and spore suspensions of L. maculans (in con-centrations 107 and 105 sporesml�1) respectively. The
seeds were germinated in 24-well multiwell plates (2
seeds per well) with 2 ml Murashige and Skoog (MS)
medium (ICN Biomedicals) supplemented with 1%
glucose and 1% microbiological agar.
2.3. Electron microscopy
Surface sterilized seeds of A. thaliana were treated
with Bacillus spore suspensions (approximately 107
sporesml�1) as described above and placed on sterile

Table 1
Bacterial strains used in this study, their characteristics and GenBank accession numbers of partial sequences of genes analysed in this study
Strainsa ;b Identificationc Source of Isolation/ Plant colonizatione Accession numbers in GenBankf
Commentsd 16S rRNA gyrA cheA tetB(L) promoter
region
DSM10T B. subtilis subsp. subtilis nd AJ276351 AF272021 nd nd
168 B. subtilis subsp. subtilis nd Z99124 (http://genolist.pasteur.fr/SubtiList/)
DSM 347T B. subtilis subsp. spizizenii nd AF074970 AF272020 AY212966 nd
DSM 7T B. amyloliquefaciens – AY055225 AF272015 AY212964 nd
H B. amyloliquefaciens nd nd nd nd nd
DSM 7264T B. atrophaeus Black pigmented nd AB021181 AF272016 nd nd
DSM 9205T B. mojavensis nd AB021191 AY212986 AY212965 nd
DSM 11031T B. vallismortis nd AB021198 AF272025 nd nd
UCMB-5008 Bs subsp. spizizenii Hay, black pigmented – AY211477 AY212969 AY212952 nd
UCMB-5014 Bs subsp. spizizenii Soil nd AY211479 AY212971 AY212955 nd
UCMB-5017 B. amyloliquefaciens Cotton plant + AY211472 AY212970 AY212954 AY212948
UCMB-5033 B. amyloliquefaciens Cotton plant + AY211473 AY212973 AY212957 nd
UCMB-5036 B. amyloliquefaciens Cotton plant – AY211476 AY212975 AY212953 nd
UCMB-5044 B. amyloliquefaciens Cotton plant – Not published AY212976 AY212956 AY212949
UCMB-5049 Bs subsp. subtilis Cotton plant ++ AY211469 AY212972 AY212958 nd
UCMB-5051 B. mojavensis Cotton plant nd AY211481 Not published AY212959 nd
UCMB-5075 B. mojavensis Soil nd AY211478 AY212977 AY212963 nd
UCMB-5113 B. amyloliquefaciens Soil; red-pigmented +++ AY211475 AY212974 AY212962 AY212950
UCMB-5137 Bs subsp. subtilis Soil ++ AY211471 AY212980 AY212960 nd
UCMB-5184 Bs subsp. subtilis Soil – AY211470 AY212979 AY212961 nd
At1 B. amyloliquefaciens Seedling of A. thaliana;
red-pigmented
+++ AY211483 AY212978 nd nd
At2 Bs subsp. subtilis Seedling of A. thaliana; nd AY211484 AY212983 nd nd
At3 Bs subsp. spizizenii Seedling of A. thaliana;
black-pigmented
– AY211485 AY212982 nd nd
At4 B. amyloliquefaciens Seedlings of A. thaliana;
red-pigmented
+++ AY211486 AY212985 nd nd
At5 Bs subsp. subtilis Seedlings of A. thaliana nd AY211487 AY212984 nd ndaT in superscript indicates type strains.b Strains labelled DSM were obtained from the German Collection of Microorganisms and Cell Cultures; strains labeled UCM were from the Ukranian Collection of Microorganisms, all other
strains were from our own collections.c Tentative identifications for isolates described in this study.dColour refers to pigmentation of colony.e Plant colonizing ability, +++, high, ++, medium, +, poor, –, undetectable; nd, not determined.fAccession numbers of sequences obtained in this study begin AY.
O.N
.Reva
etal./FEMSMicro
biologyEcology48(2004)249–259
251
252 O.N. Reva et al. / FEMS Microbiology Ecology 48 (2004) 249–259
tissue saturated with water and kept under light for 3
days in the climate chamber as described above. Bacillus
treated seeds and 4-day-old embryos of A. thaliana were
fixed in 1% glutaraldehyde in PBS buffer (20 g NaCl, 2 g
KCl, 2 g KH2PO4, 14.1 g NaH2PO4 per l�1 of distilledwater, pH 7.2) for 2 h. Samples were treated with 1%
OsO4 for 1 h, rinsed with distilled water, dehydrated in
acetone gradually by 20% (15 min per each stage) and
finally dried by the critical-point method. They were
coated with platinum/palladium before viewing in a
JEOL SEM 6320 microscope.
2.4. DNA fragment amplification, sequencing and phylo-
genetic analyses
Chromosomal DNA was isolated from late expo-
nential phase nutrient broth cultures by phenol extrac-
tion as described previously [26]. PCRs were performed
on a Hybaid PCR Express thermal cycler using Bi-
oTaqe DNA Polymerase (BioLine). The primers used
and annealing temperatures are shown in Table 2. PCRproducts were electrophoresed in 1.5% agarose (Seakem
LE agarose, Flowgen). PCR products were sequenced in
both directions using an ABI 310 capilliary sequencing
system as described previously [27]. Sequences were
aligned using BioEdit 5.0.9 (http://www.mbio.ncsu.edu/
BioEdit/bioedit.html) and edited manually.
The chromosomal loci upstream to tetB(L) were
prepared by PCR amplification with two inverted PCRprimers TetL_Up and TetB_Dn (Table 2). Chromo-
somal DNA preparations from B. amyloliquefaciens
strains UCMB-5017, B-5113 and B-5044 were cleaved
with HindIII for which no sites were present in the part
of the gene to be amplified, ligated using T4 DNA ligase
and used as template for PCR reactions. The PCR
Table 2
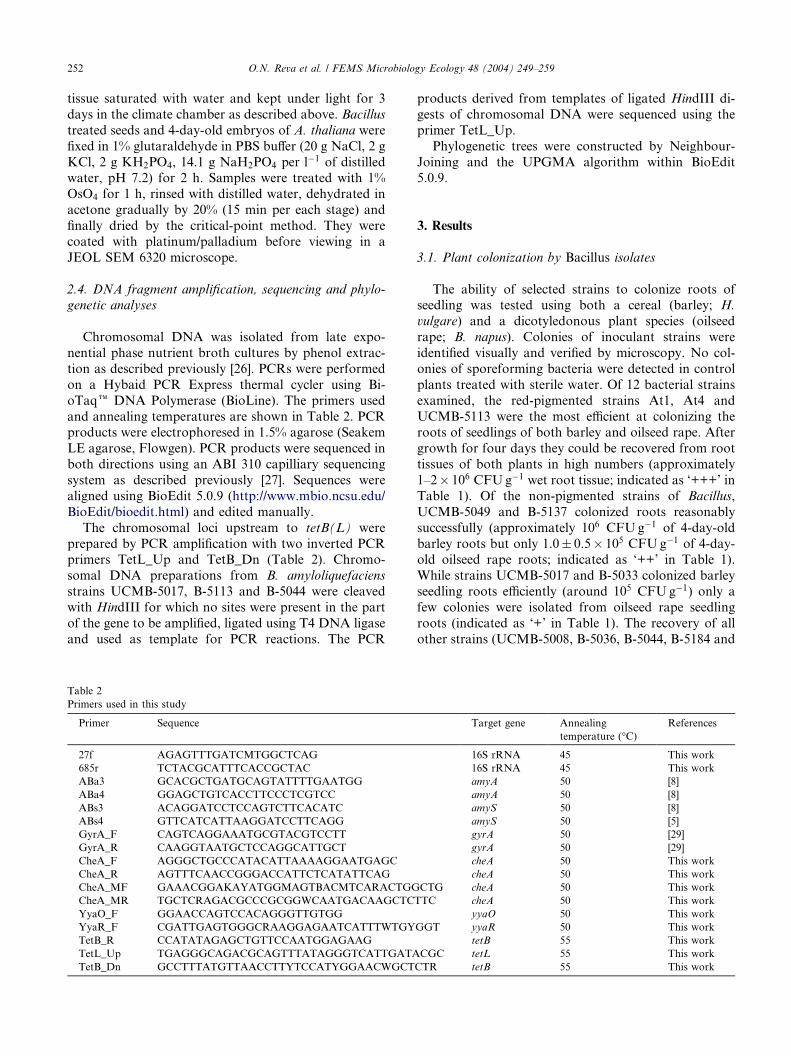
Primers used in this study
Primer Sequence
27f AGAGTTTGATCMTGGCTCAG
685r TCTACGCATTTCACCGCTAC
ABa3 GCACGCTGATGCAGTATTTTGAATGG
ABa4 GGAGCTGTCACCTTCCCTCGTCC
ABs3 ACAGGATCCTCCAGTCTTCACATC
ABs4 GTTCATCATTAAGGATCCTTCAGG
GyrA_F CAGTCAGGAAATGCGTACGTCCTT
GyrA_R CAAGGTAATGCTCCAGGCATTGCT
CheA_F AGGGCTGCCCATACATTAAAAGGAATGAGC
CheA_R AGTTTCAACCGGGACCATTCTCATATTCAG
CheA_MF GAAACGGAKAYATGGMAGTBACMTCARACTGG
CheA_MR TGCTCRAGACGCCCGCGGWCAATGACAAGCTC
YyaO_F GGAACCAGTCCACAGGGTTGTGG
YyaR_F CGATTGAGTGGGCRAAGGAGAATCATTTWTGY
TetB_R CCATATAGAGCTGTTCCAATGGAGAAG
TetL_Up TGAGGGCAGACGCAGTTTATAGGGTCATTGATA
TetB_Dn GCCTTTATGTTAACCTTYTCCATYGGAACWGCT
products derived from templates of ligated HindIII di-
gests of chromosomal DNA were sequenced using the
primer TetL_Up.
Phylogenetic trees were constructed by Neighbour-
Joining and the UPGMA algorithm within BioEdit5.0.9.
3. Results
3.1. Plant colonization by Bacillus isolates
The ability of selected strains to colonize roots ofseedling was tested using both a cereal (barley; H.
vulgare) and a dicotyledonous plant species (oilseed
rape; B. napus). Colonies of inoculant strains were
identified visually and verified by microscopy. No col-
onies of sporeforming bacteria were detected in control
plants treated with sterile water. Of 12 bacterial strains
examined, the red-pigmented strains At1, At4 and
UCMB-5113 were the most efficient at colonizing theroots of seedlings of both barley and oilseed rape. After
growth for four days they could be recovered from root
tissues of both plants in high numbers (approximately
1–2� 106 CFUg�1 wet root tissue; indicated as ‘+++’ in
Table 1). Of the non-pigmented strains of Bacillus,
UCMB-5049 and B-5137 colonized roots reasonably
successfully (approximately 106 CFUg�1 of 4-day-old
barley roots but only 1.0� 0.5� 105 CFUg�1 of 4-day-old oilseed rape roots; indicated as ‘++’ in Table 1).
While strains UCMB-5017 and B-5033 colonized barley
seedling roots efficiently (around 105 CFUg�1) only a
few colonies were isolated from oilseed rape seedling
roots (indicated as ‘+’ in Table 1). The recovery of all
other strains (UCMB-5008, B-5036, B-5044, B-5184 and
Target gene Annealing
temperature (�C)References
16S rRNA 45 This work
16S rRNA 45 This work
amyA 50 [8]
amyA 50 [8]
amyS 50 [8]
amyS 50 [5]
gyrA 50 [29]
gyrA 50 [29]
cheA 50 This work
cheA 50 This work
CTG cheA 50 This work
TTC cheA 50 This work
yyaO 50 This work
GGT yyaR 50 This work
tetB 55 This work
CGC tetL 55 This work
CTR tetB 55 This work

Table 3
Protection of Arabidopsis thaliana (An-1) against Leptosphaeria maculans (Leroy) and in vitro inhibition of L. maculans by Bacillus strains
Bacillus strains Zone of inhibitiona
(mm)
Protective effect of Bacillus spores applied with a spore suspension of L. maculans in the following
proportions (bacterium/fungus)
107/107 107/105
Dead plants Healthy plants % Dead plants Dead plants Healthy plants % Dead plants
UCMB-5017 15� 3 10 2 83 3 8 27
UCMB-5137 18� 4 8 3 73 0 12 0
UCMB-5033 16� 3 10 2 83 0 12 0
UCMB-5036 14� 3 11 1 92 6 6 50
UCMB-5113 9� 2 0 11 0 0 12 0
Control (infected) 11 1 92 11 1 92
Control (non-infected) 0 12 0 0 12 0a Zone of inhibition of growth of the fungus around colony of the Bacillus (see Section 2 for details).
O.N. Reva et al. / FEMS Microbiology Ecology 48 (2004) 249–259 253
At3) was too low to allow an accurate estimation of
bacterial numbers (indicated as ‘–’ in Table 1).
Oilseed rape plants grown from seeds treated with
spores of Bacillus did not differ from control plants, with
no signs of disease or growth inhibition. However, the
red-pigmented colonies of strains At1, At4 and UCMB-
5113 could be isolated from stems, leaves, flowers and
pods of the treated plants in titres of around1.0� 103CFUg�1 tissue after one month of growth.
Fig. 1. Micrograph of A. thaliana (An-1) seed surface with cells of B.
amyloliquefaciens UCMB-5017 one day after seed treatment with
spores.
Fig. 2. Micrograph of two cells of B. amyloliquefaciensUCMB-5113 on
the surface of A. thaliana (An-1) seeds showing apparent attachment
structures between bacterium and root.
3.2. Plant protection by Bacillus isolates
The abilities of Bacillus strains to protect plants
against phytopathogens relates to antibacterial and
antifungal polypeptide biosynthesis, and to the abilities
of these strains to survive on plant roots. In our exper-iments, seeds of A. thaliana were treated with Bacillus
spore suspensions mixed with spores of the phytopath-
ogenic fungus L. maculans. The fungal growth was
registered after incubation for one week. Concomi-
tantly, the antagonistic activity of the Bacillus strains
was tested in vitro as zones of inhibition of fungal
growth. Strains UCMB-5017 and UCMB-5036 showed
high antifungal activity on plates but protected only halfof the phytopathogen-treated A. thaliana seeds (Table
3), while the red-pigmented strain (UCMB-5113) was
characterized by a relatively weak antagonistic activity
on plates but complete protection of the plant seedlings.
Plant–microbe interactions often involve release of
bacteriocidal substances by the plant to suppress bac-
terial growth in the rhizosphere. We investigated the
effects of root secretions by A. thaliana on the develop-ment of two strains, the successful root colonizer,
UCMB-5113 (red-pigmented) and strain UCMB-5017
which is non-pigmented and a poor root colonizer. After
treatment of the seeds with Bacillus spores, the seed
surface was periodically examined during seed germi-
nation for the presence of spores and vegetative cells by
SEM. Spores of both strains successfully germinated on
the seed surface after one day (Fig. 1) and, in the case of
the more successful plant-colonizing strain, structures
apparently attaching the bacterium to the root surfacewere apparent (Fig. 2). Seed germination started 2–3
days after treatment (Fig. 3(a) and (b)). At this time
changes in cell morphology were apparent in strain
UCMB-5017. The cells near the root of the seedling
were depressed (Fig. 3(c)) perhaps due to the release of
bacteriocidal substances by plant roots. One day later,
no cells of strain UCMB-5017 were visible, only empty
cell walls were observed on the seed surface (Fig. 4(a)).Conversely, cells of strain UCMB-5113 formed a cell

Fig. 3. A general view of a 3-day-old seedling of A. thaliana (An-1); (b)
magnified view of a region marked by white rectangle on picture (a)
with multiple cells of B. amyloliquefaciens UCMB-5017 apparent (in-
dicated by arrows); (c) bacterial cells with decomposed structures.
254 O.N. Reva et al. / FEMS Microbiology Ecology 48 (2004) 249–259
layer on the seed surface (Fig. 4(b)) and microcolonies
were apparent in the root hair zone (not shown).
3.3. Taxonomy of plant-associated Bacillus isolates
We were also interested to characterize the Bacillus
strains taxonomically to determine if plant colonizingand protective features correlated with phylogenetic
groups. We amplified and compared the hypervariable
50-regions (nucleotides 115–485) of the 16S rRNA genes
Fig. 4. A. thaliana (An-1) seed coats inoculated with spores of B. amyloliquef
the presence of Bacillus by SEM after 4 days of germination. Only empty cel
seed surface heavily.
of the strains since this region has been observed to be
useful for classification and identification of endospore-
forming bacteria [8,28]. Nine polymorphic taxon-spe-
cific sites within the partial 16S rRNA sequences of the
strains were determined (Table 4). Analysis of the sec-ondary structure of the rRNA molecules revealed that
the variable nucleotides were located outside of base
paired regions (loops), or that they comprised base pairs
within stem structures, for example at sites 185/202 and
465/483. The 465/483 nucleotide pair showed two al-
ternatives: A/T (B. mojavensis and B. subtilis) or G/C (B.
amyloliquefaciens, B. atrophaeus and B. vallismortis).
Similarly, the T/A pair at positions 185/202 was presentin most of the reference strains but it was altered to C/G
in the plant-associated strains of B. amyloliquefaciens.
However, the type strain of B. amyloliquefaciens had the
unmatched pair T/G at positions 185/202 (Table 4).
Analysis of the 16S rRNA gene sequences revealed
that strains UCMB-5017, B-5033, B-5036, B-5044, B-
5113, At1 and At4 were closely related to the type strain
of B. amyloliquefaciens but distinguished by the poly-morphisms noted above and the bright red colony pig-
mentation apparent with strains UCMB-5113, At1 and
At4. Strains UCMB-5051 and B-5075 were associated
with B. mojavensis species. Strains UCM B-5049, B-
5137, B-5184, At2 and At5 were assigned to B. subtilis
on the basis of the partial 16S rRNA sequence com-
parisons, but it was impossible to refer them to sub-
species subtilis or spizizenii.In order to clarify further the classification of these
bacteria we analysed the gyrA gene, which has previ-
ously been shown to be effective for resolving these
closely related taxa of the B. subtilis group [29]. We
amplified and aligned sequences from nucleotides 226 to
1015 of gyrA for our isolates and built a neighbour-
joining tree (Fig. 5). Most strains isolated from plants,
including the red-pigmented strains, comprised amonophyletic lineage encompassing the type strain of B.
amyloliquefaciens. A second cluster of strains (based on
UCMB-5137) was recovered close to B. subtilis subsp.
aciens strains UCMB-5017 (a) and UCMB-5113 (b) were analysed for
l walls of strain B-5017 were present while strain B-5113 colonized the

Table 4
Comparison of hypervariable regions of 16S rRNA sequences of selected plant-associated strains and reference strains
Strains and alleles Positions of variable nucleotides
180 185 202 234 271 285 465 472 483
B. amyloliquefaciens DSM 7T C T G G C G G A C
B. amyloliquefaciens UCMB-5017, B-5033,
B-5036, B-5044, B-5113, At1, At4
G C G G C G G A C
B. atrophaeus DSM 7264T C T A G C A G A C
B. mojavensis DSM 9205T, UCMB-5051 C T A G C A A G T
B. mojavensis UCMB-5075 C T A A C A A G T
B. subtilis subsp. subtilis DSM10T,
UCMB-5184, At2, At5
G T A G C A A G T
Bs subsp. subtilis UCMB-5049, B-5137 G T A G T A A G T
Bs subsp. subtilis 168 alleles
rrnB, D, G, H, W G T A G C A A G T
rrnE, I G T A G T A A G T
rrnA G T A G C G A G T
rrnO G T A G C G G G T
rrnJ G T A G T A G G T
B. subtilis subsp. spizizenii DSM 347T C T A G T A A G T
Bs subsp. spizizenii UCMB-5008 C T A A C A A G T
Bs subsp. spizizenii UCMB-5014 C T A A T A A G T
Bs subsp. spizizenii AT3 C T A G C A A G T
B. vallismortis DSM11031T C T A G T A G A C
O.N. Reva et al. / FEMS Microbiology Ecology 48 (2004) 249–259 255
subtilis (strains DSM 10T and 168). Strains UCMB-5051
(not shown, only a partial sequence was obtained) and
UCMB-5075 were recovered close to B. mojavensis. Fi-
nally, two black-pigmented strains, UCMB-5008 and
At3, had almost identical sequences and shared 95%
similarity with the type strain of B. subtilis subsp. spiz-
izenii. A non-pigmented strain UCMB-5014 was also
located near here.To evaluate the gyrA phylogeny, we analysed the
cheA gene for selected strains. This key enzyme of bac-
terial chemotaxis is broadly distributed among all
Gram-positive and Gram-negative eubacteria and is
crucial for effective plant colonization by endophytic
Pseudomonas [30]. This background suggested that cheA
may reveal particular variation in this collection of
Fig. 5. Phylogenetic tree based on partial gyrA nucleotide sequences.
The scale bar indicates 0.1 nucleotide substitutions per nucleotide
position. Letter T marks type strains.
plant-associated bacteria. Two primers (CheA_F/
CheA_R) were derived from the B. subtilis 168 genome
sequence targeted to positions 127–156 and 1048–1077,
respectively. Sequences of the PCR products obtained
for selected reference strains were then used to design
primers CheA_MF/CheA_MR (Table 2). PCR products
amplified with these new primers were sequenced,
aligned and a neighbour-joining tree was constructedfrom the region between positions 325 and 891 of the B.
subtilis 168 cheA gene (Fig. 6). The topology and allo-
cation of strains to clades in the cheA tree were similar
to the gyrA tree, and the group of plant-associated
strains related to B. amyloliquefaciens was again evident.
The well characterized a-amylase genes of B. subtilis
and B. amyloliquefaciens have been used for distin-
Fig. 6. Phylogenetic tree based on partial cheA nucleotide sequences.
The scale bar indicates 0.1 nucleotide substitutions per nucleotide
position. Letter T marks type strains.

Fig. 7. Gene organization of regions near tetB(L) in chromosomes of B. subtilis subsp. subtilis 168 and B. amyloliquefaciens strains UCMB-5113, B-
5017 and B-5044. Arrowheads show parts of DNA targeted by primers TetB_R,YyaO_F and YyaR_F.
256 O.N. Reva et al. / FEMS Microbiology Ecology 48 (2004) 249–259
guishing strains of these closely related species. We
consequently amplified this gene using two pairs of
standard primers ABa1/ABa2 and ABs1/ABs2 specific
for the a-amylase genes of B. amyloliquefaciens and B.
subtilis respectively [8]. Primers ABs1/ABs2 were diag-nostic for B. subtilis subsp. subtilis strains and failed to
produce a product when DNA from B. subtilis subsp.
spizizenii DSM 347T was used as template. Primers
ABa1/ABa2 produced a product of the correct size with
DNA from B. amyloliquefaciens strains DSM 7T and H
and B. mojavensis DSM 9205T and strains UCMB-5051
and B-5075, but failed with all the plant-associated
strains of B. amyloliquefaciens.The ability to survive in the plant rhizosphere envi-
ronment, which is replete with antibiotics, may rely on
an effective system of efflux proteins that limit the ac-
cumulation of antibacterial substances in the cytoplasm.
One such transporter that confers maintenance of gen-
eral homeostasis and antibiotic resistance is the anti-
porter encoded by genes tetB-tetL [31,32]. These genes
are located near the origin of replication in B. subtilis
168 [33]. Chromosomal rearrangements were detected in
the strains of B. amyloliquefaciens compared to B. sub-
tilis 168 (Fig. 7). The gene yyaR (function unknown)
located 2003 bp downstream to tetB(L) in B. subtilis
168 appeared upstream next to tetB(L) in the B. amy-
loliquefaciens strains. In B. subtilis 168, tetB(L) are
adjacent to yyaO. To confirm this arrangement in the
other strains, two primers YyaO_F and YyaR_F de-rived from the 30-end of yyaO and yyaR were designed
and used for PCR amplification together with the re-
Table 5
Discrimination by PCR of plant-associated bacteria related to B. amylolique
Species and strains
B. amyloliquefaciens DSM 7T, H
B. amyloliquefaciens AT1, AT4, UCMB-5017, B-5033, B-5036, B-5044, B-5
B. atrophaeus DSM 7264T
B. mojavensis DSM 9205T UCMB-5051, B-5075
B. subtilis subsp. subtilis DSM 10T At2, At5, UCMB-5049, B-5137, B-518
B. subtilis subsp. spizizenii DSM 347T At3, UCMB-5008, B-5014
B. vallismortis DSM 11031T
‘+’, PCR products of expected size were detected by gel electrophoresis.
‘–’, PCR products of expected size were not detected by gel electrophore
versed primer TetB_R targeted to the 50-end of tetB.
PCR products were amplified using the primer pair
TetB_R/YyaR_F from all B. amyloliquefaciens strains
including the type strain and strain H, but no products
were amplified with strains of all other taxa used in thisstudy. Conversely, the primer pair TetB_R/YyaO_F
worked solely with B. subtilis subsp. subtilis strains and
did not amplify sequences from all other strains in-
cluding the type strain of B. subtilis subsp. spizizenii
(Table 5).
4. Discussion
The ability to colonize plants is a multifactorial
process requiring resistance to plant defence systems as
well as the ability to initiate growth on plant surfaces,
invade tissues and develop within the plant. In return for
a safe and nutrient rich environment, the bacterium may
provide the host with resistance to plant pathogens
through the synthesis of antibiotics and enzymes andpromote plant growth [1,5,15–17]. Indeed, it seems that
some bacteria become intimately associated with the
plant since strains of B. amyloliquefaciens (At1, At4 and
UCMB-5113) could be isolated in reasonably high titres
from stems, leaves and pods of oilseed rape plants one
month after initial treatment of the seed. Moreover,
vertical transmission of these strains from plant to plant
via the seeds was indicated by the isolation of severalstrains, including At1 and At4, from seedlings derived
faciens and B. subtilis
Target genes/regions and primer pairs
amyS (Abs3/
Abs4)
amyA (Aba3/
Aba4)
tetB YyaO_F/
TetB_R
tetB YyaR_F/
TetB_R
– + – +
113 – – – +
– – – –
– + – –
4 + – + –
– – – –
– – – –
sis.

O.N. Reva et al. / FEMS Microbiology Ecology 48 (2004) 249–259 257
from surface sterilized seeds and grown in a gnotobiotic
environment.
For effective initiation of colonization, the bacterium
must attach to the seed surface, primary roots and root
hairs, and survive the bacteriocidal substances releasedby roots. A comparison of a relatively poor colonizing
bacterium, B. amyloliquefaciens strain UCMB-5017, and
the more efficient colonizer, B. amyloliquefaciens
UCMB-5113, revealed that the latter formed a thick
layer on the seed surface while the former was appar-
ently inhibited and subsequently killed, maybe by bac-
teriocidal substances from the root. Moreover, strain
UCMB-5113 produced surface structures that appearedto anchor the bacterium to the root surface (Fig. 2). It
would seem that this bacterium is well adapted to seed
and root colonization. This effective plant colonization
ability for B. amyloliquefaciens UCMB-5113 was asso-
ciated with a high level of protection against the phy-
topathogen L. maculans, although it showed limited
antagonistic activity against the fungus in vitro. In
contrast other strains had distinct antifungal activity invitro but showed limited in vivo protection (Table 3).
The most probable explanations for this are that (i)
colonizing ability is a key to successful plant protection
and (ii) the environment within the plant may induce the
bacterium to synthesize antifungal compounds that are
not produced in vitro. Moreover, inoculation of plants
with endophytic bacteria can result in induced systemic
resistance in the host plant. The phenomenon of inducedsystemic resistance involves release of antibiotic sub-
stances in response to bacterial challenge and was first
demonstrated with an endophytic Pseudomonas strain,
but has also been noted when B. amyloliquefaciens was
used as a biocontrol agent [34–36]. These findings sug-
gest that in vitro antifungal bioassays can be misleading
in assessing the biocontrol properties of an organism.
The bacteria included in this study were largely iso-lated from plants or from soil intimately associated with
plant roots. However, the abilities to colonize plants in
the laboratory may not necessarily relate to their ori-
gins. For example, some strains isolated from the inner
tissues of cotton plants, such as B. amyloliquefaciens
UCMB-5036 and UCM B-5044, did not colonize the
roots of barley or oilseed rape plants to any appreciable
extent. This indicates specificity of bacterial strains to-wards certain host plants. Moreover, while various
strains, including some that produced red-pigmented
colonies, colonized our test plants effectively, others did
not. We therefore characterized the bacteria to deter-
mine any relationships between taxonomy and plant
colonizing ability.
Sequence analysis of partial 16S rRNA genes revealed
that all strains were closely related to B. amyloliquefac-
iens and B. subtilis. This is consistent with several pre-
vious studies in which strains of these species have been
commonly encountered as plant colonizing bacteria
[5,6,9,16,37]. While the partial 16S rRNA sequences
provided insufficient resolution to distinguish the vari-
ous subspecies and close relatives of B. subtilis, a group
of strains was evident characterized by a G residue at
position 202, typical of B. amyloliquefaciens, and yethaving a unique C at position 185. This suggested a
group of strains closely related to, but distinct from B.
amyloliquefaciens. All strains in this group were isolated
from plants and many had good plant colonizing
properties. Three red-pigmented strains were included in
this group. Such pigmentation is rare in B. amylolique-
faciens and in Bacillus in general.
In order to analyse more thoroughly this group ofplant-associated strains, we compared partial gyrA se-
quences which have been shown to be useful for dis-
tinguishing B. subtilis from its close relatives [29]. gyrA
sequences of the seven B. amyloliquefaciens-like strains
formed a monophyletic group with the sequences from
the three red-pigmented strains clustering together
(Fig. 5) and two strains (UCMB-5036 and B-5044)
which failed to colonize barley and oilseed rape rootsforming another subgroup. All seven bacteria were
closely related to the type strain of B. amyloliquefaciens
on the basis of gyrA sequences. Other isolates were
identified according to their nearest relative, although it
is apparent that there is some sequence variation in the
B. subtilis subsp. spizizenii, B. subtilis subsp. subtilis, and
B. mojavensis clades. It is not clear how much variation
in gyrA may be associated with a taxon since, with theexception of B. subtilis subsp. subtilis, branches in the
original study were represented by single strains. Al-
though the results from our strains support the use of
this gene for species recognition in this group since it
enabled approximate assignment of strains to a taxon,
we sought further evidence from a second gene, cheA.
Plant colonization is believed to be an active process
that requires bacteria to respond to the complex envi-ronment of the plant rhizosphere which is replete with
antibiotics and chemical signals. It may be expected,
therefore, that endophytic bacteria have evolved sensi-
tive and finely tuned chemotactic responses. Indeed,
cheA mutants of Pseudomonas fluorescens are defective
in both flagella-driven chemotaxis and tomato root
colonization [30]. We therefore used the cheA gene as a
phylogenetic marker for the B. amyloliquefaciens strainsisolated from plants. The resultant tree (Fig. 6) was to-
pologically consistent with the gyrA tree and confirmed
strain allocation to taxa. It also revealed that cheA was
changing approximately five times faster than gyrA in
these organisms. Moreover, while the gyrA alleles con-
tained many synonymous changes, the cheA alleles were
characterized by non-synonymous changes, insertions
and deletions, suggesting rapid adaptation to the rhi-zosphere environment by these bacteria.
A simple PCR-based diagnostic procedure for iden-
tification of B. amyloliquefaciens and the related plant-

258 O.N. Reva et al. / FEMS Microbiology Ecology 48 (2004) 249–259
associated taxa would be useful for identification of
strains isolated from plants or used for biocontrol pur-
poses. The existing PCR test for identification of B.
amyloliquefaciens strains based on the a-amylase gene of
the organism [5] was not useful since the primers failedto amplify a product from the plant-associated strains.
We therefore analysed the tetB(L) antiporter genes of
these strains. A different organization of these genes was
discovered in B. amyloliquefaciens strains compared
with strains of B. subtilis subsp. subtilis (Fig. 7) which
enabled the design of diagnostic primers YyaR_F/
TetB_R that amplified a product from B. amylolique-
faciens DNA (both the type strain and plant-associatedstrains) but not from DNA of strains of B. atrophaeus,
B. mojavensis, B. subtilis subsp. subtilis and subsp.
spizizenii. Thus the plant-associated strains can be
recognized by a positive reaction with the tetB(L)
primers and a negative reaction with those for a-amylase
(Table 5).
The precise taxonomic status of the plant-associated
strains of B. amyloliquefaciens remains unclear. Theseorganisms have much in common with B. amylolique-
faciens sensu stricto, the tetB(L) region of the chromo-
some has the same arrangement and cheA is very
similar. But in other aspects they are distinguishable,
there is a signature base in the 16S rRNA gene (C/G at
positions 185/202 in the plant-associated strains of B.
amyloliquefaciens compared with the unmatched pair T/
G in the type strain, see Table 4), the a-amylase geneappears to be dissimilar from that of the type strain
since it is not amplified by amyA primers, and the gyrA
sequences vary. In many respects these bacteria have the
properties of an ecotype which Cohan defines as ‘‘a set
of strains using the same or similar ecological resources’’
[38]. The ecotype concept includes consideration of
competition between adaptive mutants and is similar to
the clone in bacterial population genetics [39]. A bac-terial ecotype can be identified as a sequence cluster [38]
just as multilocus sequence typing indicates clones in
bacterial species [40]. On the basis of the cheA and the
gyrA sequence data and other information presented
here, it would seem that the plant-associated strains of
B. amyloliquefaciens could be considered a distinct
ecotype within this species.
Acknowledgements
Reva O. is grateful to the Society for General Mi-
crobiology of United Kingdom and the Swedish Insti-
tute (SI) with C.F. Lundstr€oms Stifftelse for financial
support of this work through the International Research
Fellowships Grant scheme and New VISBY ProgramFellowship Grant respectively. We are grateful to Hans
Ekwall, Dept. of Anatomy and Histology, the Swedish
University of Agricultural Sciences (SLU), for assistance
with the scanning electron microscopy. Inger Happsta-
dius, Sval€ov-Weibull AB is gratefully acknowledged for
providing B. napus seeds and the fungal isolate and we
thank Dr. Peter Morris, Heriot-Watt University, for
providing barley seeds and assistance with barley ex-periments.
References
[1] Chanway, C.P. (1998) Bacterial endophytes: ecological and
practical implication. Sydowia, 149–170.
[2] Cook, R.J. (1990) Twenty-five years of progress towards biolog-
ical control. In: Biological Control of Soil-borne Plant Pathogens
(Hornby, D., Ed.), pp. 1–14. CAB International, Wallingford,
UK.
[3] Gilbert, G.S. and Parke, J.L. (1993) Effects of an introduced
bacterium on bacterial communities on roots. Ecology 74, 840–
854.
[4] Chanway, C.P., Shishido, M., Nairn, J., Jungwirth, S., Markham,
J., Xiao, G. and Holl, F.B. (2000) Endophytic colonization and
field responses of hybrid spruce seedlings after inoculation with
plant growth-promoting rhizobacteria. Forest Ecol. Manag. 133,
81–88.
[5] Emmert, E.A.B. and Handelsman, J. (1999) Biocontrol of plant
disease: a (Gram-) positive perspective. FEMS Microbiol. Lett.
171, 1–9.
[6] Sharga, B.M. and Lyon, G.D. (1998) Bacillus subtilis BS 107 as an
antagonist of potato blackleg and soft rot bacteria. Can. J.
Microbiol. 44, 777–783.
[7] Turner, J.T. and Backman, P.A. (1991) Factors relating to peanut
yield increases after seed treatment with Bacillus subtilis. Plant
Dis. 75, 347–353.
[8] Idriss, E.E., Makarewicz, O., Farouk, A., Rosner, K., Greiner, R.,
Bochow, H., Richter, T. and Borriss, R. (2002) Extracellular
phytase activity of Bacillus amyloliquefaciens FZB45 contributes
to its plant-growth-promoting effect. Microbiology 148, 2097–
2109.
[9] Manjula, K. and Podile, A.R. (2001) Chitin-supplemented
formulations improve biocontrol and plant growth promoting
efficiency of Bacillus subtilis AF1. Can. J. Microbiol. 47, 618–
625.
[10] Wang, S.L., Shih, I.L., Liang, T.W. and Wang, C.H. (2002)
Purification and characterization of two antifungal chitinases
extracellularly produced by Bacillus amyloliquefaciens V656 in a
shrimp and crab shell powder medium. J. Agric. Food Chem. 50,
2241–2248.
[11] Leifert, C., Li, H., Chidburee, S., Hampson, S., Workman, S.,
Sigee, D., Epton, H.A. and Harbour, A. (1995) Antibiotic
production and biocontrol activity by Bacillus subtilis CL27 and
Bacillus pumilus CL45. J. Appl. Bacteriol. 78, 97–108.
[12] Pinchuk, I.V., Bressollier, P., Sorokulova, I.B., Verneuil, B. and
Urdaci, M.C. (2002) Amicoumacin antibiotic production and
genetic diversity of Bacillus subtilis strains isolated from different
habitats. Res. Microbiol. 153, 269–276.
[13] Raaijmakers, J.M., Vlami, M. and de Souza, J.T. (2002) Antibi-
otic production by bacterial biocontrol agents. Antonie Van
Leeuwenhoek 81, 537–547.
[14] Walker, R., Powell, A.A. and Seddon, B. (1998) Bacillus isolates
from the spermosphere of peas and dwarf French beans with
antifungal activity against Botrytis cinerea and Phytium species. J.
Appl. Microbiol. 84, 791–801.
[15] Hallmann, J., Quadt-Hallmann, A., Mahafee, W.F. and Kloep-
per, J.W. (1997) Bacterial endophytes in agricultural crops. Can. J.
Microbiol. 38, 303–308.

O.N. Reva et al. / FEMS Microbiology Ecology 48 (2004) 249–259 259
[16] Krebs, B., Hoding, B., Kubart, S., Workie, M.A., Junge, H.,
Schmiedeknecht, G., Grosch, R., Bochow, H. and Hevesi, M.
(1998) Use of Bacillus subtilis as biocontrol agents. I. Activity and
characterization of Bacillus subtilis strains. J. Plant Dis. Protect.
105, 181–197.
[17] Wipat, A. and Harwood, C.R. (1999) The Bacillus subtilis genome
sequence: the molecular blueprint of a soil bacterium. FEMS
Microbiol. Ecol. 28, 1–9.
[18] Nakamura, L.K. (1989) Taxonomic relationship of black-pig-
mented Bacillus subtilis strains and proposal for Bacillus atropha-
eus sp. nov. Int. J. Syst. Bacteriol. 39, 295–300.
[19] Roberts, M.S., Nakamura, L.K. and Cohan, F.M. (1994) Bacillus
mojavensis sp. nov., distinguishable from Bacillus subtilis by
sexual isolation, divergence in DNA sequence, and differences
in fatty acid composition. Int. J. Syst. Bacteriol. 44,
256–264.
[20] Roberts, M.S., Nakamura, L.K. and Cohan, F.M. (1996) Bacillus
vallismortis sp. nov., a close relative of Bacillus subtilis, isolated
from soil in Death Valley, California. Int. J. Syst. Bacteriol. 46,
470–475.
[21] Nakamura, L.K., Roberts, M.S. and Cohan, F.M. (1999) Rela-
tionship of Bacillus subtilis clades associated with strains 168 and
W23: a proposal for Bacillus subtilis subsp. subtilis subsp. nov. and
Bacillus subtilis subsp. spizizeni subsp. nov. Int. J. Syst. Evolut.
Microbiol. 49, 1211–1215.
[22] Reva, O.N., Sorokulova, I.B. and Smirnov, V.V. (2001) Simplified
technique for identification of the aerobic spore-forming
bacteria by phenotype. Int. J. Syst. Evolut. Microbiol. 51, 1361–
1371.
[23] Sorokulova, I.B., Reva, O.N., Smirnov, V.V., Pinchuk, I.V.,
Lapa, S.V. and Urdaci, M.C. (2003) Genetic diversity and
involvement in bread spoilage of Bacillus strains from flour and
ropy bread. Lett. Appl. Microbiol. 37, 169–173.
[24] Bohman, S., Staal, J., Thomma, B.P.H.J., Wang, M. and Dixelius,
C. (2004) Characterisation of an Arabidopsis–Leptosphaeria mac-
ulans pathosystem: resistance partially requires camalexin biosyn-
thesis and is independent of salicylic acid, ethylene and jasmonic
acid signalling. Plant J. 37, 9–20.
[25] Koch, E., Song, K., Osborn, T.C. and Williams, P.W. (1991)
Relationship between pathogenicity and phylogeny based on
restriction fragment length polymorphism in Leptosphaeria mac-
ulans. Mol. Plant–Microbe Interact. 4, 341–349.
[26] De Silva, S., Pettersson, B., Aquino de Muro, M. and Priest, F.G.
(1998) A DNA probe for the detection and identification of
Bacillus sporothermodurans using the 23S-16S rDNA spacer region
and phylogenetic analysis of some field isolates of Bacillus which
form highly heat resistant spores. Syst. Appl. Microbiol. 21, 398–
407.
[27] van Beek, S. and Priest, F.G. (2002) Evolution of the lactic acid
bacterial community during malt whisky fermentation: a poly-
phasic study. Appl. Environ. Microbiol. 68, 297–305.
[28] Goto, K., Omura, T., Hara, Y. and Sadaie, Y. (2000) Application
of the partial 16S rDNA sequence as an index for rapid
identification of species in the genus Bacillus. J. Gen. Appl.
Microbiol. 46, 1–8.
[29] Chun, J. and Bae, K.S. (2000) Phylogenetic analysis of Bacillus
subtilis and related taxa based on partial gyrA gene sequences.
Antonie van Leeuwenhoek 78, 123–127.
[30] de Weert, S., Vermeiren, H., Mulders, I.H., Kuiper, I., Hendrickx,
N., Bloemberg, G.V., Vanderleyden, J., De Mot, R. and Lugten-
berg, B.J. (2002) Flagella-driven chemotaxis towards exudate
components is an important trait for tomato root colonization by
Pseudomonas fluorescens. Mol. Plant–Microbe Interact. 15, 1173–
1180.
[31] Bechhofer, D.H. and Stasinopoulos, S.J. (1998) tet A(L) mutants
of a tetracycline-sensitive strain of Bacillus subtilis with the
polynucleotide phosphorylase gene deleted. J. Bacteriol. 180,
3470–3473.
[32] Cheng, J., Guffanti, A.A. and Krulwich, T.A. (1994) The
chromosomal tetracycline resistance locus of Bacillus subtilis
encodes a Naþ/Hþ antiporter that is physiologically important
at elevated pH. J. Biol. Chem. 44, 27365–27371.
[33] Kunst, F., Ogasawara, N. and Moszer, I., et al. (1997) The
complete genome sequence of the Gram-positive bacterium
Bacillus subtilis. Nature 390, 249–256.
[34] Ahn, I.P., Park, K. and Kim, C.H. (2002) Rhizobacteria-induced
resistance perturbs viral disease progress and triggers defense-
related gene expression. Mol. Cells 13, 302–308.
[35] Benhamou, N. and Nicole, M. (1999) Cell biology of plant
immunization against microbial infection: the potential of induced
resistance in controlling plant diseases. Plant Physiol. Biochem.
37, 703–719.
[36] Hammond-Kosack, K.E. and Jones, J.D.G. (1997) Plant disease
resistance genes. Annu. Rev. Plant Physiol. Plant Mol. Biol. 48,
575–607.
[37] Misaghi, I.J. and Dondelinger, C.R. (1990) Endophytic bacteria in
symptom-free cotton plants. Phytopathology 9, 808–811.
[38] Cohan, F.M. (2002) What are bacterial species? Annu. Rev.
Microbiol. 56, 547–587.
[39] Smith, M.J., Feil, E.J. and Smith, N.H. (2000) Population
structure and evolutionary dynamics of pathogenic bacteria.
BioEssays 22, 1115–1122.
[40] Feil, E.J., Maiden, M.C., Achtman, M. and Sprat, B.G. (1999)
The relative contributions of recombination and mutation to the
divergence of clones of Neisseria meningitides. Mol. Boil. Evol. 16,
1496–1502.




















