
RsmY, a small regulatory RNA, is required in concert with RsmZ for GacA-dependent expression of...
19
Molecular Microbiology (2003) 50(4), 1361–1379 doi:10.1046/j.1365-2958.2003.03774.x © 2003 Blackwell Publishing Ltd Blackwell Science, LtdOxford, UKMMIMolecular Microbiology 1365-2958Blackwell Publishing Ltd, 200350 413611379Original ArticleSmall regulatory RNAs of P. fluorescensC. Valverde, S. Heeb, C. Keel and D. Haas Accepted 13 August, 2003. *For correspondence. E-mail [email protected]; Tel. (+41) 21 692 5639; Fax (+41) 21 692 5605. †Present address: School of Pharmaceutical Sciences, University Park, University of Nottingham, Nottingham NG7 2RD, UK. RsmY, a small regulatory RNA, is required in concert with RsmZ for GacA-dependent expression of biocontrol traits in Pseudomonas fluorescens CHA0 Claudio Valverde,* Stephan Heeb, † Christoph Keel and Dieter Haas Institut de Microbiologie Fondamentale, Bâtiment de Biologie, Université de Lausanne, CH-1015 Lausanne (Dorigny), Switzerland. Summary In the plant-beneficial soil bacterium and biocontrol model organism Pseudomonas fluorescens CHA0, the GacS/GacA two-component system upregulates the production of biocontrol factors, i.e. antifungal secondary metabolites and extracellular enzymes, under conditions of slow, non-exponential growth. When activated, the GacS/GacA system promotes the transcription of a small regulatory RNA (RsmZ), which sequesters the small RNA-binding protein RsmA, a translational regulator of genes involved in biocontrol. The gene for a second GacA-regulated small RNA (RsmY) was detected in silico in various pseudomonads, and was cloned from strain CHA0. RsmY, like RsmZ, contains several characteristic GGA motifs. The rsmY gene was expressed in strain CHA0 as a 118 nt transcript which was most abun- dant in stationary phase, as revealed by Northern blot and transcriptional fusion analysis. Transcription of rsmY was enhanced by the addition of the strain’s own supernatant extract containing a quorum- sensing signal and was abolished in gacS or gacA mutants. An rsmA mutation led to reduced rsmY expression, via a gacA -independent mechanism. Overexpression of rsmY restored the expression of target genes ( hcnA , aprA ) to gacS or gacA mutants. Whereas mutants deleted for either the rsmY or the rsmZ structural gene were not significantly altered in the synthesis of extracellular products (hydrogen cyanide, 2,4-diacetylphloroglucinol, exoprotease), an rsmY rsmZ double mutant was strongly impaired in this production and in its biocontrol properties in a cucumber- Pythium ultimum microcosm. Mobility shift assays demonstrated that multiple molecules of RsmA bound specifically to RsmY and RsmZ RNAs. In conclusion, two small, untranslated RNAs, RsmY and RsmZ, are key factors that relieve RsmA- mediated regulation of secondary metabolism and biocontrol traits in the GacS/GacA cascade of strain CHA0. Introduction The biocontrol strain Pseudomonas fluorescens CHA0 protects plant roots from pathogenic fungi (Keel and Déf- ago, 1997; Haas et al ., 2002). This ability depends to a considerable extent on the production of antifungal sec- ondary metabolites such as 2,4-diacetylphloroglucinol (DAPG), pyoluteorin and HCN (Laville et al ., 1992; 1998; Maurhofer et al ., 1994; Schnider-Keel et al ., 2000). The synthesis of these antifungal compounds is under the tight positive control of the GacS/GacA two-component system (Heeb and Haas, 2001; Haas et al ., 2002). As a conse- quence, a gacA mutant of P. fluorescens CHA0 no longer protects various dicotyledoneous plants from infection by root-pathogenic fungi (Laville et al ., 1992; Maurhofer et al ., 1995). The GacS/GacA regulatory system also acti- vates the synthesis of tryptophan side-chain oxidase and of extracellular enzymes such as phospholipase C and exoprotease in strain CHA0 (Oberhänsli and Défago, 1991; Sacherer et al ., 1994). A quorum-sensing signal, which is produced by strain CHA0 and is unrelated to N- acylhomoserine lactones (AHLs), activates the GacS/ GacA regulatory cascade in this bacterium (Heeb et al ., 2002), possibly by interaction with the sensor kinase GacS (Zuber et al ., 2003). Upon autophosphorylation, GacS transfers the phosphoryl group to the DNA-binding response regulator GacA (Pernestig et al ., 2001). Then, phosphorylated GacA is assumed to activate target gene expression, resulting in the production of antifungal com- pounds and extracellular enzymes (Heeb and Haas, 2001; Haas et al ., 2002). These target genes include the hcn , phl and apr operons, coding for HCN synthase, DAPG biosynthetic enzymes and the major exoprotease respec- tively (Laville et al ., 1998; Schnider-Keel et al ., 2000; Heeb et al ., 2002). The GacS/GacA signal transduction cascade positively controls transcription of a small, untranslated regulatory
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of RsmY, a small regulatory RNA, is required in concert with RsmZ for GacA-dependent expression of...

Molecular Microbiology (2003)
50
(4), 1361–1379 doi:10.1046/j.1365-2958.2003.03774.x
© 2003 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology 1365-2958Blackwell Publishing Ltd, 200350
413611379
Original Article
Small regulatory RNAs of P. fluorescensC. Valverde, S. Heeb, C. Keel and D. Haas
Accepted 13 August, 2003. *For correspondence. [email protected]; Tel. (
+
41) 21 692 5639; Fax (
+
41) 21692 5605. †Present address: School of Pharmaceutical Sciences,University Park, University of Nottingham, Nottingham NG7 2RD, UK.
RsmY, a small regulatory RNA, is required in concert with RsmZ for GacA-dependent expression of biocontrol traits in
Pseudomonas fluorescens
CHA0
Claudio Valverde,* Stephan Heeb,
†
Christoph Keel and Dieter Haas
Institut de Microbiologie Fondamentale, Bâtiment de Biologie, Université de Lausanne, CH-1015 Lausanne (Dorigny), Switzerland.
Summary
In the plant-beneficial soil bacterium and biocontrolmodel organism
Pseudomonas fluorescens
CHA0,the GacS/GacA two-component system upregulatesthe production of biocontrol factors, i.e. antifungalsecondary metabolites and extracellular enzymes,under conditions of slow, non-exponential growth.When activated, the GacS/GacA system promotesthe transcription of a small regulatory RNA (RsmZ),which sequesters the small RNA-binding proteinRsmA, a translational regulator of genes involved inbiocontrol. The gene for a second GacA-regulatedsmall RNA (RsmY) was detected
in silico
in variouspseudomonads, and was cloned from strain CHA0.RsmY, like RsmZ, contains several characteristicGGA motifs. The
rsmY
gene was expressed in strainCHA0 as a 118 nt transcript which was most abun-dant in stationary phase, as revealed by Northernblot and transcriptional fusion analysis. Transcriptionof
rsmY
was enhanced by the addition of the strain’sown supernatant extract containing a quorum-sensing signal and was abolished in
gacS
or
gacA
mutants. An
rsmA
mutation led to reduced
rsmY
expression, via a
gacA
-independent mechanism.Overexpression of
rsmY
restored the expression oftarget genes (
hcnA
,
aprA
) to
gacS
or
gacA
mutants.Whereas mutants deleted for either the
rsmY
or the
rsmZ
structural gene were not significantly altered inthe synthesis of extracellular products (hydrogencyanide, 2,4-diacetylphloroglucinol, exoprotease), an
rsmY rsmZ
double mutant was strongly impaired inthis production and in its biocontrol properties in acucumber-
Pythium ultimum
microcosm. Mobility
shift assays demonstrated that multiple molecules ofRsmA bound specifically to RsmY and RsmZ RNAs.In conclusion, two small, untranslated RNAs, RsmYand RsmZ, are key factors that relieve RsmA-mediated regulation of secondary metabolism andbiocontrol traits in the GacS/GacA cascade of strainCHA0.
Introduction
The biocontrol strain
Pseudomonas fluorescens
CHA0protects plant roots from pathogenic fungi (Keel and Déf-ago, 1997; Haas
et al
., 2002). This ability depends to aconsiderable extent on the production of antifungal sec-ondary metabolites such as 2,4-diacetylphloroglucinol(DAPG), pyoluteorin and HCN (Laville
et al
., 1992; 1998;Maurhofer
et al
., 1994; Schnider-Keel
et al
., 2000). Thesynthesis of these antifungal compounds is under the tightpositive control of the GacS/GacA two-component system(Heeb and Haas, 2001; Haas
et al
., 2002). As a conse-quence, a
gacA
mutant of
P. fluorescens
CHA0 no longerprotects various dicotyledoneous plants from infection byroot-pathogenic fungi (Laville
et al
., 1992; Maurhofer
et al
., 1995). The GacS/GacA regulatory system also acti-vates the synthesis of tryptophan side-chain oxidase andof extracellular enzymes such as phospholipase C andexoprotease in strain CHA0 (Oberhänsli and Défago,1991; Sacherer
et al
., 1994). A quorum-sensing signal,which is produced by strain CHA0 and is unrelated to N-acylhomoserine lactones (AHLs), activates the GacS/GacA regulatory cascade in this bacterium (Heeb
et al
.,2002), possibly by interaction with the sensor kinaseGacS (Zuber
et al
., 2003). Upon autophosphorylation,GacS transfers the phosphoryl group to the DNA-bindingresponse regulator GacA (Pernestig
et al
., 2001). Then,phosphorylated GacA is assumed to activate target geneexpression, resulting in the production of antifungal com-pounds and extracellular enzymes (Heeb and Haas, 2001;Haas
et al
., 2002). These target genes include the
hcn
,
phl
and
apr
operons, coding for HCN synthase, DAPGbiosynthetic enzymes and the major exoprotease respec-tively (Laville
et al
., 1998; Schnider-Keel
et al
., 2000;Heeb
et al
., 2002).The GacS/GacA signal transduction cascade positively
controls transcription of a small, untranslated regulatory

1362
C. Valverde, S. Heeb, C. Keel and D. Haas
© 2003 Blackwell Publishing Ltd,
Molecular Microbiology
,
50
, 1361–1379
RNA, designated RsmZ in strain CHA0 (Heeb
et al
.,2002) or PrrB (an RsmZ homologue) in strain F113,another
P. fluorescens
biocontrol strain (Aarons
et al
.,2000). Overexpression of RsmZ or PrrB from multicopyplasmids can suppress
gacS
and
gacA
mutations in bothstrains (Aarons
et al
., 2000; Heeb
et al
., 2002). Theexpression of a transcriptional
rsmZ-lacZ
fusion dependson the function of both GacS and GacA, strictly requiresan upstream activating sequence (UAS) at
-
180 nt fromthe transcription start site, and is stimulated by the addi-tion of the non-AHL quorum-sensing signal extractedfrom CHA0 cultures (Heeb
et al
., 2002). The currentmodel proposes that RsmZ RNA produced during lateexponential phase under GacS/GacA control binds andtitrates the small RNA-binding protein RsmA, therebypreventing the translational repression exerted by thisprotein on GacS/GacA target genes (Heeb
et al
., 2002;Haas
et al
., 2002). This model is supported by the obser-vations that overexpression of RsmA results in strongdownregulation of translational
hcnA
¢
-‘
lacZ
and
aprA
¢
-‘
lacZ
fusions, and that point mutations in the
hcnA
untranslated leader region near the ribosome binding siteabolish control by GacA and RsmA (Blumer
et al
., 1999).A similar signal transduction pathway also occurs in
Erwinia carotovora
, where the GacS/GacA system pro-motes the expression of the
rsmB
gene, which codes fora small RNA that counteracts RsmA-mediated regulationof several target mRNAs (Liu
et al
., 1998; Cui
et al
.,2001; Chatterjee
et al
., 2002). In
Escherichia coli
, theGacS (BarA)/GacA (UvrY) system positively controls thelevel of the carbon storage regulator RNAs CsrB andCsrC (Liu
et al
., 1997; Suzuki
et al
., 2002; Pernestig
et al
., 2003; Weilbacher
et al
., 2003). CsrB makes a ribo-nucleoprotein complex with approximately 18 moleculesof the CsrA protein, which is a homologue of RsmA. Theformation of such a complex can reduce the amount ofCsrA which binds to target mRNAs. For instance, CsrAbinding to
glgCAP
mRNA causes translational repressionand facilitates mRNA decay (Liu
et al
., 1995; 1997; Liuand Romeo, 1997; Baker
et al
., 2002; Dubey
et al
.,2003). Homologous elements of the GacA/RsmA post-transcriptional regulatory cascade have also been foundin
Salmonella typhimurium
where they control theexpression of genes related to invasiveness (Altier
et al
.,2000). Thus, many Gram-negative bacteria use this sig-nal transduction pathway to adjust the expression of met-abolic pathways involved in carbon flow, secondarymetabolism and secretion of exoenzymes or virulencefactors, especially during the transition from exponentialto stationary growth phase (Heeb and Haas, 2001;Pernestig
et al
., 2003).In
P. fluorescens
, mutational inactivation of
rsmZ
or
prrB
only marginally affects target gene expression comparedto the drastic reduction associated with
gacS
or
gacA
mutations (Aarons
et al
., 2000; Heeb
et al
., 2002), sug-gesting that
rsmZ (prrB
) may not be the only regulatoryRNA through which GacS and GacA antagonize RsmA.In different pseudomonads the
rpoS-rsmZ
intergenicregion is well conserved, especially in the UAS of the
rsmZ
promoter. We found that a similar UAS is alsopresent in the thermoregulatory region (TRR) of
Pseudomonas syringae
pv.
phaseolicola
(Rowley
et al
.,1993). When present in multiple copies, the TRR locusderepresses phaseolotoxin synthesis at 28
∞
C in this bac-terium; however, no product of this locus has been iden-tified (Rowley
et al
., 1993). In
P. fluorescens
CHA0, wefound that a TRR-like locus codes for a regulatory RNAwhich we designate RsmY and which cooperates withRsmZ as an RsmA antagonist in the GacS/GacA regula-tory system.
Results
The
rsmY
gene (TRR locus) of
Pseudomonas
spp. codes for a small RNA under GacS/GacA control
The overexpressed TRR locus of
P. syringae
pv.
phaseoli-cola
overrides thermoregulation of phaseolotoxin produc-tion, which is normally repressed at 28
∞
C (Rowley
et al
.,1993). TRR contains a conserved palindromic sequence(TGTAAGcN
4
tCtTACA; Fig. 1A) which is similar to asequence element present in the UAS of the GacA-regulated
rsmZ
promoter (Heeb
et al
., 2002). Moreover,TRR displays several GGA motifs which are characteristicof the riboregulators CsrB, RsmB, CsrC and RsmZ(Romeo, 1998; Cui
et al
., 2001; Heeb
et al
., 2002; Weil-bacher
et al
., 2003). The TRR locus also occurs in severalother
Pseudomonas
species whose genomes have beensequenced partially or entirely (Fig. 1A) and was pickedas a good candidate for encoding an RNA with affinity forRsmA. In the following, we will refer to the TRR locus asthe
rsmY gene. To demonstrate the existence of RsmYRNA, we carried out Northern blot analysis of total RNAfrom P. aeruginosa PAO1 grown in rich medium; thisrevealed a single transcript of about 120 nt, which accu-mulated in stationary phase (Fig. 1B). This value is ingood agreement with the size predicted (124 nt) from thealignment of the rsmY promoters and r-independent ter-minators (Fig. 1A).
An alignment of TRR-like loci in fluorescentpseudomonads (Fig. 1A) was used to design degeneratePCR oligonucleotides to amplify the corresponding rsmYregion from P. fluorescens strain CHA0 (see Experimentalprocedures), for which genomic information is currentlynot available. A 1.6 kb product was cloned in pBluescript(pBLS) producing pME6912, and the insert was confirmedby sequencing. In Northern blot experiments with totalRNA extracted from strain CHA0 at various cell densities,

Small regulatory RNAs of P. fluorescens 1363
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
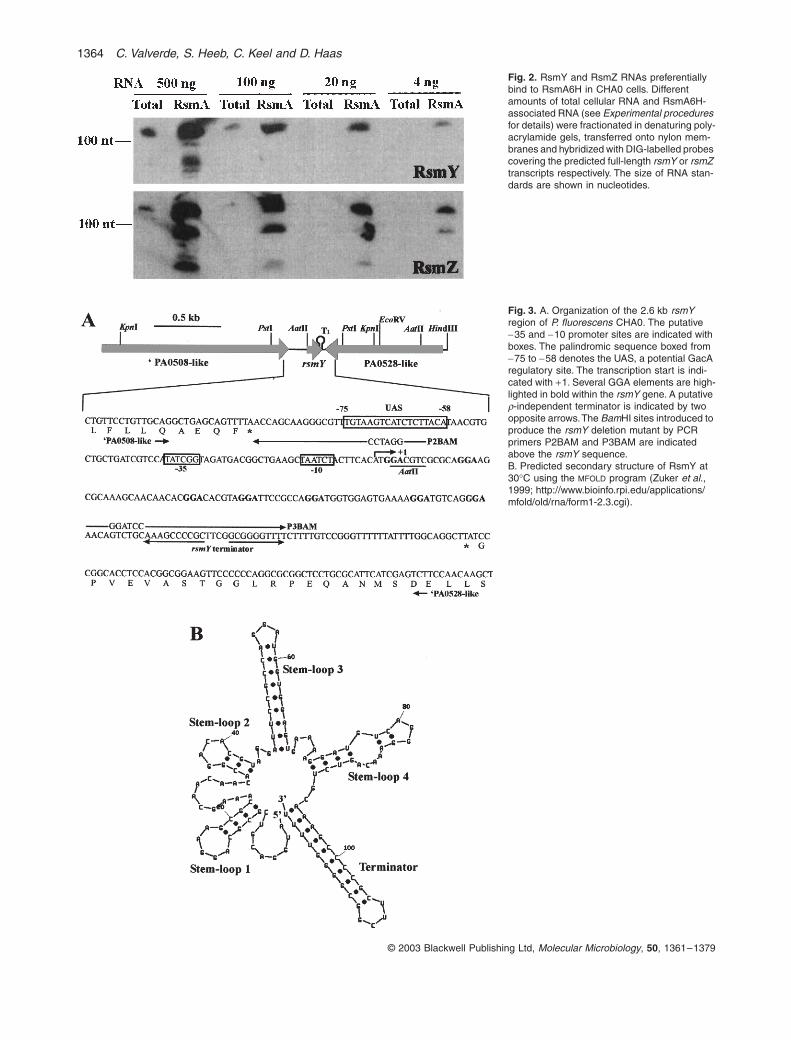
a single rsmY transcript of ~ 120 nt progressively accu-mulated in CHA0 grown at 20∞C (Fig. 1C, lanes 3–6), butwas not detected in gacS (Fig. 1C, lane 2) or gacAmutants (data not shown). Moreover, when the histidine-tagged RsmA protein (RsmA6H) was overexpressed inCHA0 carrying pME6078, the RsmY and RsmZ tran-scripts were found to be bound preferentially to this pro-tein in vivo, and consequently they could be detected bya Northern blot in highly diluted RsmA-associated RNA,whereas similar dilutions of total cellular RNA did notcontain detectable RsmY or RsmZ transcripts (Fig. 2).Some degradation or premature termination products ofRsmY and RsmZ could also be detected in the RsmAfraction (Fig. 2). In conclusion, these results strongly sug-gest that the rsmY gene codes for a GacS/GacA-con-trolled small RNA capable to interact with the translationalregulator RsmA.
Genomic context and promoter mapping of rsmY in P. fluorescens CHA0
In order to determine the genetic context of rsmY in strainCHA0, the 1.6 kb PCR fragment contained in pME6912was subcloned into the suicide vector pME3087 to per-form a chromosome walking experiment (see Experimen-tal procedures). The rsmY gene was located between aputative acyl-CoA dehydrogenase gene (homologous toP. aeruginosa PA0508) and a putative transcriptional reg-ulator gene of the LysR family (homologous to P. aerugi-nosa PA0528) (Fig. 3A). With the exception of P. syringaepv. tomato, the rsmY gene is always found next to thistranscriptional regulator in the Pseudomonas genomes(data not shown). In three P. fluorescens strains which arecurrently being sequenced (strains Pf0–1, Pf-5 andSBW25), the organization of the rsmY region is most
Fig. 1. A. Alignment of the thermoregulatory region (TRR) encompassing the rsmY gene of several pseudomonads. Psyr-phas, P. syringae pv phaseolicola (Rowley et al., 1993); Psyr-tom, P. syringae pv tomato (http://pseudomonas-syringae.org; Buell et al., 2003); Pput-KT2440, P. putida KT2440 (Nelson et al., 2002; http://www.ncbi.nlm.nih.gov/genomes/framik.cgi?db=Genome&gi=266); Pf-5, P. fluo-rescens Pf-5 (http://tigrblast.tigr.org/ufmg/index.cgi?database=p_fluorescens/seq), Pf0–1, P. fluorescens Pf0–1 (http://www.jgi.doe.gov/JGI_microbial/html/pseudomonas/pseudo_homepage.html); Pf-SBW25, P. fluorescens SBW25 (http://www.sanger.ac.uk/Projects/P_fluorescens); PAO1, P. aeruginosa PAO1 (Stover et al., 2000; http://www.pseudomonas.com). The nucleotide sequence of this DNA region for strain CHA0 is identical to that of strain Pf-5 (results of this study). The transcription start (indicated as +1) was deter-mined for strain CHA0 in this study. Asterisks denote conserved nucleotides. UAS, upstream activating sequence; -35 and -10, pro-moter elements.B. Northern blot analysis of the rsmY transcript of P. aeruginosa PAO1. Total RNA was extracted from cells grown at 37∞C in NYB and hybridized with a DIG-labelled probe covering the predicted full-length rsmY transcript. In each lane, 1 mg of RNA was loaded. OD600 values at the time of harvesting were, in lanes 1–5: 0.3, 0.9, 1.2, 1.5, and 2.0 respectively.C. Northern blot of P. fluorescens rsmY transcript. Total RNA was extracted from CHA0 (lane 1) and CHA19 (gacS; lane 2) cells grown at 30∞C or CHA0 cells grown at 20∞C (lanes 3–6), and hybridized with a DIG-labelled probe (121 bp) covering the predicted full-length rsmY transcript. In each lane, 2 mg of RNA was loaded. OD600 values at the time of harvesting were, in lanes 1–6, 3.0, 3.2, 0.3, 0.7, 1.0, and 1.5, respectively. Std, RNA standards; sizes are shown in nucleotides.
1364 C. Valverde, S. Heeb, C. Keel and D. Haas
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
Fig. 2. RsmY and RsmZ RNAs preferentially bind to RsmA6H in CHA0 cells. Different amounts of total cellular RNA and RsmA6H-associated RNA (see Experimental procedures for details) were fractionated in denaturing poly-acrylamide gels, transferred onto nylon mem-branes and hybridized with DIG-labelled probes covering the predicted full-length rsmY or rsmZ transcripts respectively. The size of RNA stan-dards are shown in nucleotides.
Fig. 3. A. Organization of the 2.6 kb rsmY region of P. fluorescens CHA0. The putative -35 and -10 promoter sites are indicated with boxes. The palindromic sequence boxed from -75 to -58 denotes the UAS, a potential GacA regulatory site. The transcription start is indi-cated with +1. Several GGA elements are high-lighted in bold within the rsmY gene. A putative r-independent terminator is indicated by two opposite arrows. The BamHI sites introduced to produce the rsmY deletion mutant by PCR primers P2BAM and P3BAM are indicated above the rsmY sequence.B. Predicted secondary structure of RsmY at 30∞C using the MFOLD program (Zuker et al., 1999; http://www.bioinfo.rpi.edu/applications/mfold/old/rna/form1-2.3.cgi).

Small regulatory RNAs of P. fluorescens 1365
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
strongly conserved. In particular, the 2.6 kb fragmentcloned from strain CHA0 has 98% nucleotide sequenceidentity with the corresponding region of strain Pf-5(http://www.tigrblast.tigr.org/ufmg/index.cgi?database=p_fluorescens/seq). As previously described for the TRRlocus of P. syringae pv. phaseolicola (Rowley et al., 1993),rsmY does not appear to contain any obvious open read-ing frames and has a potential r-independent terminator(Fig. 3A). The transcription start of rsmY was mapped byRACE (rapid amplification of its cDNA end; Tillett et al.,2000) (Fig. 3A; Fig. S1). A computer-simulated folding(Zuker et al., 1999) of the rsmY transcript at 30∞C predictsfour stem-loop structures and an additional terminatorhairpin, with characteristic GGA motifs in the loops andother unpaired regions (Fig. 3B). This modelled second-ary structure resembles that of RsmZ in strain CHA0(Heeb et al., 2002), PrrB in strain F113 (Aarons et al.,2000), and CsrB and CsrC in E. coli (Liu et al., 1997;Weilbacher et al., 2003).
Regulation of rsmY expression
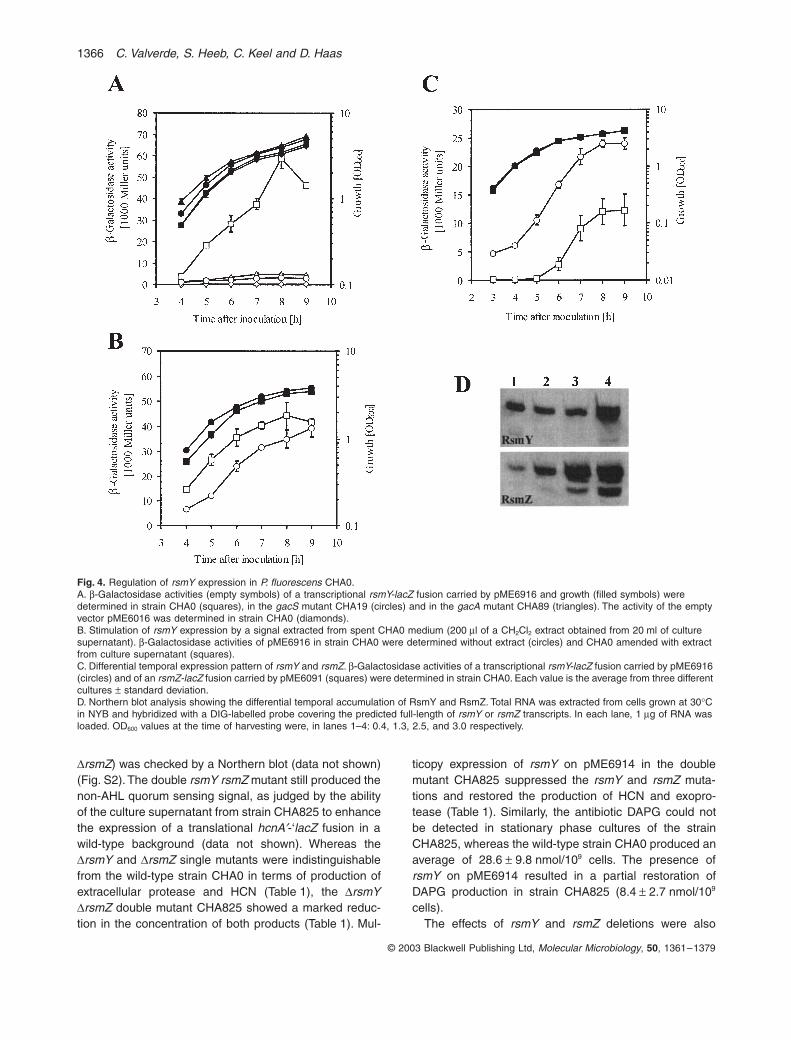
Northern blot experiments indicate that the expressionof RsmY RNA depends on GacS/GacA and increaseswith cell density (Fig. 1C). To quantify this regulation,we constructed a transcriptional fusion by placing thelacZ gene precisely at the +1 site under the control ofthe rsmY promoter in a vector that has five to sevencopies per cell (Heeb et al., 2000). The activity of thersmY promoter increased with cell density and, in latestationary phase, was 12–18 times higher in the wild-type strain CHA0 than in gacS and gacA mutants(Fig. 4A), confirming that the GacS/GacA system isstrongly involved in rsmY expression. In addition, whena culture extract containing the CHA0 quorum-sensingsignal was added to the growth medium, the activity ofthe rsmY promoter was enhanced, especially at lowcell densities (Fig. 4B). The rsmY-lacZ transcriptionalfusion was expressed earlier and at a higher basallevel than was an rsmZ-lacZ fusion (Fig. 4C). Suchtemporal difference in promoter activity was alsoreflected by the intracellular content of RsmY andRsmZ RNAs as detected by Northern hybridization atvarious cell densities (Fig. 4D). In conclusion, RsmY ispart of the GacS/GacA regulon of strain CHA0 and ispredicted to control the expression of target genes ear-lier than does RsmZ (Heeb et al., 2002).
In E. coli, CsrA activates transcription of csrB and csrCby an indirect mechanism involving the BarA (GacS)/UvrY(GacA) system (Suzuki et al., 2002; Weilbacher et al.,2003). In addition, disruption of either RNA gene resultsin a modest compensatory increase in the concentrationof the other (Weilbacher et al., 2003). We investigated ifsuch regulatory circuitry exists also in strain CHA0 by
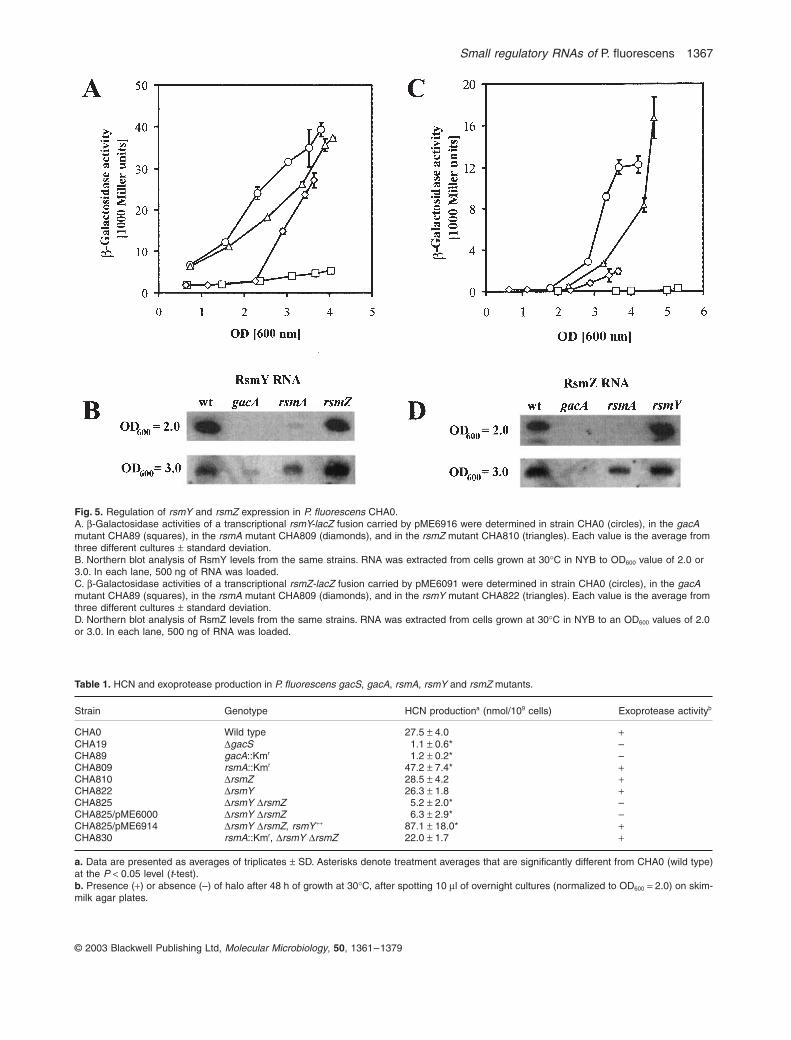
determining the effects of rsmA, rsmY and rsmZ mutationson the promoter activity and transcript levels of bothRNAs. RsmA was required for rsmY and rsmZ expressionat low cell densities (OD600 < 3.0; Fig. 5). However, thiseffect appears to be independent of the GacA level,because an rsmA mutation had no effect on the expres-sion of a chromosomal gacA¢-‘lacZ translational fusionalong the entire growth curve (data not shown) (Fig. S3).By contrast, absence of either rsmZ or rsmY led only tosome transient changes in rsmY or rsmZ transcriptionefficiencies respectively (Fig. 5A and C).
Overexpression of rsmY suppresses gacS and gacA mutations
Translational hcnA¢-‘lacZ and aprA¢-‘lacZ reporterfusions have been useful to monitor cell density-depen-dent regulation by the GacS/GacA cascade in strainCHA0. In gacS or gacA mutants, the expression ofthese fusions is extremely low (Blumer et al., 1999).When the rsmY gene was expressed from its own pro-moter in the multicopy vector pME6914 and introducedinto strain CHA806 (chromosomal aprA¢-‘lacZ fusion,DgacS), almost no b-galactosidase activity could bedetected and the rsmY transcript was detected at areduced level by Northern blotting (data not shown). It islikely that the amount of RsmY RNA produced from theplasmid in this strain lacking gacS was insufficient tocounteract the RsmA-mediated translational repressionof aprA mRNA, which reflects the requirement of theGacS/GacA system for a correct expression of rsmY(Figs 4 and 5). For this reason, the rsmY gene wasplaced under the strong tac promoter in pME6918, suchthat the transcription start site of this promoter coincidedwith the first nucleotide of rsmY. The presence ofpME6918 in strain CHA89.207 (chromosomal hcnA¢-‘lacZ fusion, gacA::Kmr) resulted in a higher b-galactosi-dase activity than in strain CHA207 (chromosomalhcnA¢-‘lacZ fusion, otherwise wild type), even withoutIPTG induction of the tac promoter (Fig. 6A). Overex-pression of rsmY in the gacS-defective strain CHA806also suppressed the gacS mutation, albeit only partially,as evidenced by an aprA¢-‘lacZ fusion (Fig. 6B). North-ern blot analysis suggests that this difference in the sup-pressor ability of the RsmY overexpressing plasmidpME6918 correlated with the level of RsmY RNA, whichfor unknown reasons was higher in CHA89.207 than instrain CHA806 (data not shown).
Phenotypes of rsmY and rsmY rsmZ mutants
A chromosomal rsmY deletion was created in the wild typeand in DrsmZ background. The absence of RsmY in theresulting strains CHA822 (DrsmY) and CHA825 (DrsmY
1366 C. Valverde, S. Heeb, C. Keel and D. Haas
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
DrsmZ) was checked by a Northern blot (data not shown)(Fig. S2). The double rsmY rsmZ mutant still produced thenon-AHL quorum sensing signal, as judged by the abilityof the culture supernatant from strain CHA825 to enhancethe expression of a translational hcnA¢-‘lacZ fusion in awild-type background (data not shown). Whereas theDrsmY and DrsmZ single mutants were indistinguishablefrom the wild-type strain CHA0 in terms of production ofextracellular protease and HCN (Table 1), the DrsmYDrsmZ double mutant CHA825 showed a marked reduc-tion in the concentration of both products (Table 1). Mul-
ticopy expression of rsmY on pME6914 in the doublemutant CHA825 suppressed the rsmY and rsmZ muta-tions and restored the production of HCN and exopro-tease (Table 1). Similarly, the antibiotic DAPG could notbe detected in stationary phase cultures of the strainCHA825, whereas the wild-type strain CHA0 produced anaverage of 28.6 ± 9.8 nmol/109 cells. The presence ofrsmY on pME6914 resulted in a partial restoration ofDAPG production in strain CHA825 (8.4 ± 2.7 nmol/109
cells).The effects of rsmY and rsmZ deletions were also
Fig. 4. Regulation of rsmY expression in P. fluorescens CHA0.A. b-Galactosidase activities (empty symbols) of a transcriptional rsmY-lacZ fusion carried by pME6916 and growth (filled symbols) were determined in strain CHA0 (squares), in the gacS mutant CHA19 (circles) and in the gacA mutant CHA89 (triangles). The activity of the empty vector pME6016 was determined in strain CHA0 (diamonds).B. Stimulation of rsmY expression by a signal extracted from spent CHA0 medium (200 ml of a CH2Cl2 extract obtained from 20 ml of culture supernatant). b-Galactosidase activities of pME6916 in strain CHA0 were determined without extract (circles) and CHA0 amended with extract from culture supernatant (squares).C. Differential temporal expression pattern of rsmY and rsmZ. b-Galactosidase activities of a transcriptional rsmY-lacZ fusion carried by pME6916 (circles) and of an rsmZ-lacZ fusion carried by pME6091 (squares) were determined in strain CHA0. Each value is the average from three different cultures ± standard deviation.D. Northern blot analysis showing the differential temporal accumulation of RsmY and RsmZ. Total RNA was extracted from cells grown at 30∞C in NYB and hybridized with a DIG-labelled probe covering the predicted full-length of rsmY or rsmZ transcripts. In each lane, 1 mg of RNA was loaded. OD600 values at the time of harvesting were, in lanes 1–4: 0.4, 1.3, 2.5, and 3.0 respectively.
Small regulatory RNAs of P. fluorescens 1367
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
Fig. 5. Regulation of rsmY and rsmZ expression in P. fluorescens CHA0.A. b-Galactosidase activities of a transcriptional rsmY-lacZ fusion carried by pME6916 were determined in strain CHA0 (circles), in the gacA mutant CHA89 (squares), in the rsmA mutant CHA809 (diamonds), and in the rsmZ mutant CHA810 (triangles). Each value is the average from three different cultures ± standard deviation.B. Northern blot analysis of RsmY levels from the same strains. RNA was extracted from cells grown at 30∞C in NYB to OD600 value of 2.0 or 3.0. In each lane, 500 ng of RNA was loaded.C. b-Galactosidase activities of a transcriptional rsmZ-lacZ fusion carried by pME6091 were determined in strain CHA0 (circles), in the gacA mutant CHA89 (squares), in the rsmA mutant CHA809 (diamonds), and in the rsmY mutant CHA822 (triangles). Each value is the average from three different cultures ± standard deviation.D. Northern blot analysis of RsmZ levels from the same strains. RNA was extracted from cells grown at 30∞C in NYB to an OD600 values of 2.0 or 3.0. In each lane, 500 ng of RNA was loaded.
Table 1. HCN and exoprotease production in P. fluorescens gacS, gacA, rsmA, rsmY and rsmZ mutants.
Strain Genotype HCN productiona (nmol/109 cells) Exoprotease activityb
CHA0 Wild type 27.5 ± 4.0 +CHA19 DgacS 1.1 ± 0.6* –CHA89 gacA::Kmr 1.2 ± 0.2* –CHA809 rsmA::Kmr 47.2 ± 7.4* +CHA810 DrsmZ 28.5 ± 4.2 +CHA822 DrsmY 26.3 ± 1.8 +CHA825 DrsmY DrsmZ 5.2 ± 2.0* –CHA825/pME6000 DrsmY DrsmZ 6.3 ± 2.9* –CHA825/pME6914 DrsmY DrsmZ, rsmY++ 87.1 ± 18.0* +CHA830 rsmA::Kmr, DrsmY DrsmZ 22.0 ± 1.7 +
a. Data are presented as averages of triplicates ± SD. Asterisks denote treatment averages that are significantly different from CHA0 (wild type)at the P < 0.05 level (t-test).b. Presence (+) or absence (–) of halo after 48 h of growth at 30∞C, after spotting 10 ml of overnight cultures (normalized to OD600 = 2.0) on skim-milk agar plates.
1368 C. Valverde, S. Heeb, C. Keel and D. Haas
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
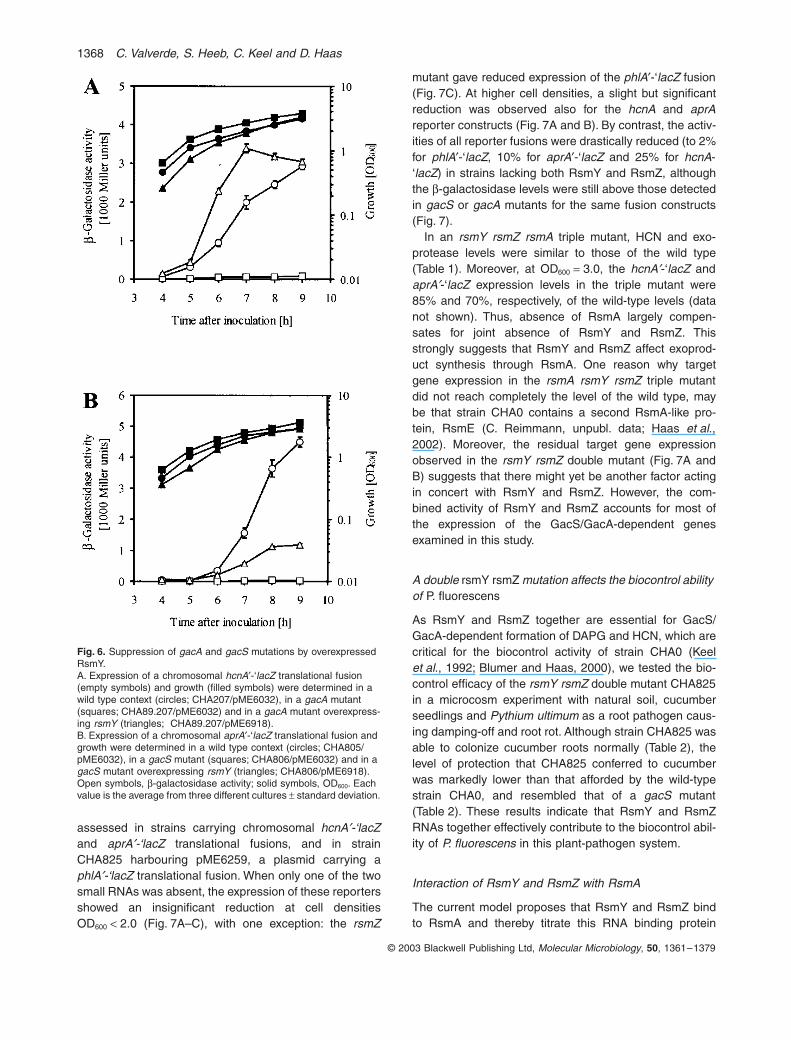
assessed in strains carrying chromosomal hcnA¢-‘lacZand aprA¢-‘lacZ translational fusions, and in strainCHA825 harbouring pME6259, a plasmid carrying aphlA¢-‘lacZ translational fusion. When only one of the twosmall RNAs was absent, the expression of these reportersshowed an insignificant reduction at cell densitiesOD600 < 2.0 (Fig. 7A–C), with one exception: the rsmZ
mutant gave reduced expression of the phlA¢-‘lacZ fusion(Fig. 7C). At higher cell densities, a slight but significantreduction was observed also for the hcnA and aprAreporter constructs (Fig. 7A and B). By contrast, the activ-ities of all reporter fusions were drastically reduced (to 2%for phlA¢-‘lacZ, 10% for aprA¢-‘lacZ and 25% for hcnA-‘lacZ) in strains lacking both RsmY and RsmZ, althoughthe b-galactosidase levels were still above those detectedin gacS or gacA mutants for the same fusion constructs(Fig. 7).
In an rsmY rsmZ rsmA triple mutant, HCN and exo-protease levels were similar to those of the wild type(Table 1). Moreover, at OD600 = 3.0, the hcnA¢-‘lacZ andaprA¢-‘lacZ expression levels in the triple mutant were85% and 70%, respectively, of the wild-type levels (datanot shown). Thus, absence of RsmA largely compen-sates for joint absence of RsmY and RsmZ. Thisstrongly suggests that RsmY and RsmZ affect exoprod-uct synthesis through RsmA. One reason why targetgene expression in the rsmA rsmY rsmZ triple mutantdid not reach completely the level of the wild type, maybe that strain CHA0 contains a second RsmA-like pro-tein, RsmE (C. Reimmann, unpubl. data; Haas et al.,2002). Moreover, the residual target gene expressionobserved in the rsmY rsmZ double mutant (Fig. 7A andB) suggests that there might yet be another factor actingin concert with RsmY and RsmZ. However, the com-bined activity of RsmY and RsmZ accounts for most ofthe expression of the GacS/GacA-dependent genesexamined in this study.
A double rsmY rsmZ mutation affects the biocontrol ability of P. fluorescens
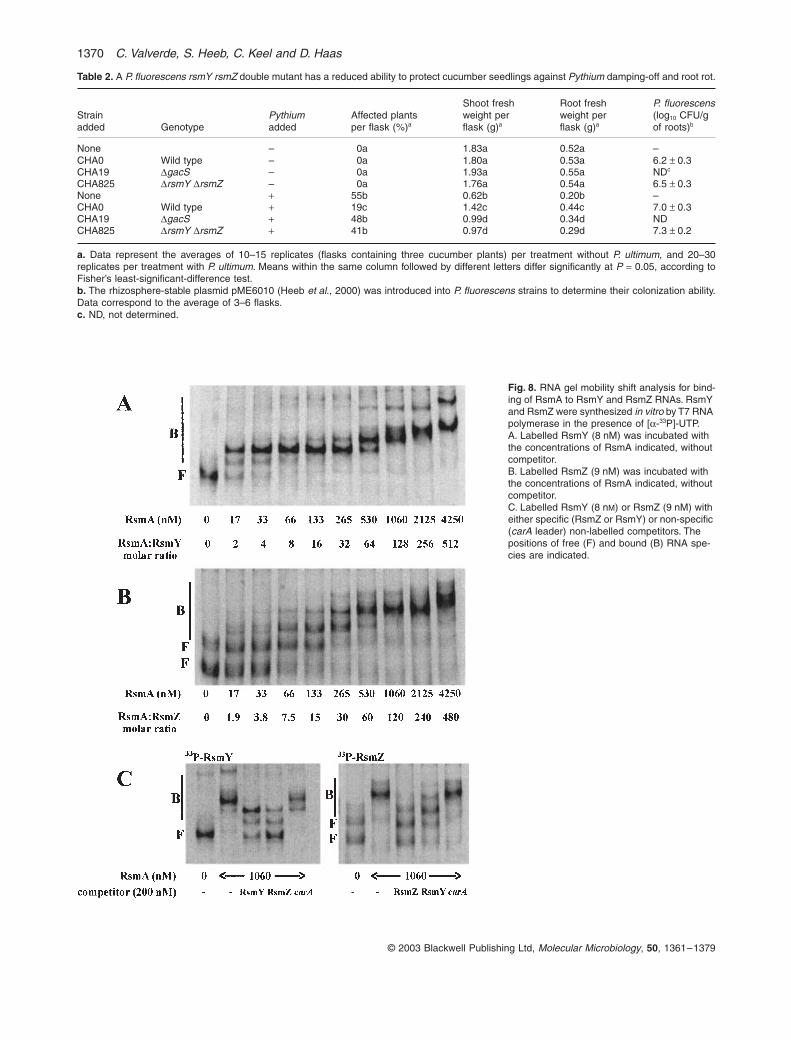
As RsmY and RsmZ together are essential for GacS/GacA-dependent formation of DAPG and HCN, which arecritical for the biocontrol activity of strain CHA0 (Keelet al., 1992; Blumer and Haas, 2000), we tested the bio-control efficacy of the rsmY rsmZ double mutant CHA825in a microcosm experiment with natural soil, cucumberseedlings and Pythium ultimum as a root pathogen caus-ing damping-off and root rot. Although strain CHA825 wasable to colonize cucumber roots normally (Table 2), thelevel of protection that CHA825 conferred to cucumberwas markedly lower than that afforded by the wild-typestrain CHA0, and resembled that of a gacS mutant(Table 2). These results indicate that RsmY and RsmZRNAs together effectively contribute to the biocontrol abil-ity of P. fluorescens in this plant-pathogen system.
Interaction of RsmY and RsmZ with RsmA
The current model proposes that RsmY and RsmZ bindto RsmA and thereby titrate this RNA binding protein
Fig. 6. Suppression of gacA and gacS mutations by overexpressed RsmY.A. Expression of a chromosomal hcnA¢-‘lacZ translational fusion (empty symbols) and growth (filled symbols) were determined in a wild type context (circles; CHA207/pME6032), in a gacA mutant (squares; CHA89.207/pME6032) and in a gacA mutant overexpress-ing rsmY (triangles; CHA89.207/pME6918).B. Expression of a chromosomal aprA¢-‘lacZ translational fusion and growth were determined in a wild type context (circles; CHA805/pME6032), in a gacS mutant (squares; CHA806/pME6032) and in a gacS mutant overexpressing rsmY (triangles; CHA806/pME6918). Open symbols, b-galactosidase activity; solid symbols, OD600. Each value is the average from three different cultures ± standard deviation.

Small regulatory RNAs of P. fluorescens 1369
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
from the recognition sequences present in targetmRNAs, e.g. of aprA, hcnA or phlA (Blumer et al., 1999;Haas et al., 2002). Support for this model comes fromthe observation that when RsmA6H was overexpressed
in CHA0 cells, it co-purified with important amounts ofRNA (about 60% [w/w]; Heeb et al., 2002) and it wasenriched for RsmY and RsmZ transcripts (Fig. 2). Tocharacterize better the interaction between RsmA andRsmY or RsmZ, we prepared in vitro transcripts of RsmYand RsmZ and performed mobility shift assays usingRsmA6H purified from E. coli. RsmA binding to RsmY(8 nM) was detected as two distinct complexes already at17 nM RsmA. The band corresponding to free RsmY wascompletely shifted between 33 and 66 nM RsmA. AtRsmA concentrations ≥66 nM, additional retarded bandswere detected, which probably correspond to highermolecular weight species with several molecules ofRsmA bound to RsmY (Fig. 8A). The RsmA-RsmY shiftpattern suggests that the affinity of the first binding siteon RsmY is higher than that of the remaining sites, as nosignificant change in the shifted species was observedbetween 17 and 265 nM RsmA (Fig. 8A). In vitro tran-scription of rsmZ yielded two differently sized transcripts,similar to those observed in Northern blots of cellularRNA preparations (Figs 2 and 4D). RsmA binding toRsmZ (9 nM) followed a similar pattern (Fig. 8B). In thiscase, however, a complete shift of the free RNA speciesoccurred between 133 and 265 nM RsmA. Higher RsmAconcentrations also led to the appearance of shiftedRsmA–RsmZ complexes having higher molecular weight(Fig. 8B).
Competition experiments were performed to assessthe specificity of RsmA binding to RsmY and RsmZ. Asshown in Fig. 8C, unlabelled RsmY and RsmZ tran-scripts were both able to compete with RsmA–RsmY orRsmA–RsmZ complexes, whereas the same amount ofa similarly sized RNA lacking the putative RsmA bindingelements (i.e. the untranslated leader of the P. fluore-scens carA gene) (Fig. S4) did not modify the interac-tion of RsmA with RsmY or RsmZ (Fig. 8C). Thisdemonstrates that RsmY and RsmZ bind to RsmAspecifically.
Fig. 7. Effect of rsmY and rsmZ deletions on the expression of the hcnA, aprA, and phlA genes.A. Expression of a chromosomal hcnA¢-‘lacZ translational fusion and growth were determined in a wild-type context (circles; CHA207), in a gacA mutant (squares; CHA89.207), in an rsmY mutant (triangles; CHA823), in an rsmZ mutant (inverted triangles; CHA811) and in an rsmY rsmZ double mutant (diamonds; CHA826).B. Expression of a chromosomal aprA¢-‘lacZ translational fusion and growth were determined in a wild type context (circles; CHA805), in a gacA mutant (squares; CHA806), in an rsmY mutant (triangles; CHA824), in an rsmZ mutant (inverted triangles; CHA812) and in an rsmY rsmZ double mutant (diamonds; CHA827).C. Expression of a plasmid-encoded phlA¢-‘lacZ translational fusion (pME6259) and growth were determined in a wild type context (cir-cles; CHA0), in a gacS mutant (squares; CHA19), in an rsmY mutant (triangles; CHA822), in an rsmZ mutant (inverted triangles; CHA810) and in an rsmY rsmZ double mutant (diamonds; CHA825). Open symbols, b-galactosidase activitiy; solid symbols, OD600. Each value is the average from three different cultures ± standard deviation.
1370 C. Valverde, S. Heeb, C. Keel and D. Haas
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
Table 2. A P. fluorescens rsmY rsmZ double mutant has a reduced ability to protect cucumber seedlings against Pythium damping-off and root rot.
Strainadded Genotype
Pythiumadded
Affected plantsper flask (%)a
Shoot freshweight perflask (g)a
Root freshweight perflask (g)a
P. fluorescens(log10 CFU/gof roots)b
None – 0a 1.83a 0.52a –CHA0 Wild type – 0a 1.80a 0.53a 6.2 ± 0.3CHA19 DgacS – 0a 1.93a 0.55a NDc
CHA825 DrsmY DrsmZ – 0a 1.76a 0.54a 6.5 ± 0.3None + 55b 0.62b 0.20b –CHA0 Wild type + 19c 1.42c 0.44c 7.0 ± 0.3CHA19 DgacS + 48b 0.99d 0.34d NDCHA825 DrsmY DrsmZ + 41b 0.97d 0.29d 7.3 ± 0.2
a. Data represent the averages of 10–15 replicates (flasks containing three cucumber plants) per treatment without P. ultimum, and 20–30replicates per treatment with P. ultimum. Means within the same column followed by different letters differ significantly at P = 0.05, according toFisher’s least-significant-difference test.b. The rhizosphere-stable plasmid pME6010 (Heeb et al., 2000) was introduced into P. fluorescens strains to determine their colonization ability.Data correspond to the average of 3–6 flasks.c. ND, not determined.
Fig. 8. RNA gel mobility shift analysis for bind-ing of RsmA to RsmY and RsmZ RNAs. RsmY and RsmZ were synthesized in vitro by T7 RNA polymerase in the presence of [a-33P]-UTP.A. Labelled RsmY (8 nM) was incubated with the concentrations of RsmA indicated, without competitor.B. Labelled RsmZ (9 nM) was incubated with the concentrations of RsmA indicated, without competitor.C. Labelled RsmY (8 nM) or RsmZ (9 nM) with either specific (RsmZ or RsmY) or non-specific (carA leader) non-labelled competitors. The positions of free (F) and bound (B) RNA spe-cies are indicated.

Small regulatory RNAs of P. fluorescens 1371
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
Discussion
Several small untranslated RNAs act as regulators of var-ious cellular processes in bacteria (Argaman et al., 2001;Rivas et al., 2001; Wassarman et al., 2001; Wassarman,2002; Storz, 2002; Wagner et al., 2002). These regulatoryelements, also termed riboregulators, fall into severalclasses (Gottesman, 2002). One major class comprisesriboregulators that are able to establish base pairing withtarget mRNAs, e.g. DsrA, RprA, Spot42, MicF, UptR andRhyB of E. coli (Majdalani et al., 1998; Wassarman et al.,1999; Guigueno et al., 2001; Majdalani et al., 2002;Massé and Gottesman, 2002). In general, such RNAsmodify the behaviour of the target mRNA upon binding.Many of them require the RNA-binding protein Hfq as acofactor in the antisense RNA–RNA interaction (Gottes-man, 2002; Møller et al., 2002). A second important groupof riboregulators, e.g. CsrB and CsrC in E. coli (Liu et al.,1997; Weilbacher et al., 2003), CsrB in S. typhimurium(Altier et al., 2000), RsmB in Erwinia spp. (Ma et al., 2001)and RsmZ (PrrB) in P. fluorescens (Aarons et al., 2000;Heeb et al., 2002), are antagonists of small translationalregulators (CsrA, RsmA). It is thought that an excess ofthese small RNAs can titrate the CsrA/RsmA protein.Thus, RsmA-mediated regulation of target mRNAs can berelieved (Romeo, 1998; Haas et al., 2002). Many Gram-negative bacteria use the GacS/GacA two-componentsystem to regulate the expression of secondary metabo-lites and extracellular enzymes under the influence ofenvironmental and biological signals (Heeb and Haas,2001). Interestingly, riboregulators of the second classmentioned above are important elements within the GacS/GacA cascade. In the plant pathogen Erwinia carotovorassp. carotovora, the GacS/GacA system positively con-trols the expression of the rsmB riboregulator gene andthereby modulates bacterial virulence (Cui et al., 2001).By antagonizing the translational regulator RsmA, RsmBnot only allows the translation of target mRNAs such ashrpL, but also prevents RsmA-mediated mRNA decay(Chatterjee et al., 2002). RsmA-like proteins are alsopresent in some Gram-positive bacteria (White et al.,1995) but their role remains to be investigated in thesespecies, although there is evidence for a small RNA of theRsmB/CsrB type in Bacillus subtilis (Ando et al., 2002).
In this study, we describe a novel small non-coding RNA(designated RsmY) whose transcription is activated by theGacS/GacA two-component system and which actstogether with RsmZ to regulate positively the expressionof the aprA, hcnA and phlA genes in P. fluorescens CHA0,apparently at a post-transcriptional level. Although thersmY locus has been previously described as a ther-moregulatory region (TRR) in P. syringae pv. phaseolicola,where multiple copies of TRR derepress phaseolotoxinsynthesis at 28∞C (Rowley et al., 1993), its mechanism of
action has remained obscure. In the present study, thepredicted rsmY RNA product was detected by Northernblot in both P. aeruginosa PAO1 and in P. fluorescensCHA0 as a single transcript of about 120 nt (Fig. 1). Sev-eral properties of RsmY qualify it as a component of theGacS/GacA post-transcriptional regulatory cascade in P.fluorescens. (i) The rsmY promoter was positively regu-lated by the GacS/GacA system and the non-AHL quo-rum-sensing signal (Fig. 4). (ii) Overexpression of rsmYactivated the expression of reporter fusions which areknown to be negatively regulated by RsmA (Blumer et al.,1999) in gacS or gacA mutants (Fig. 6). (iii) Mutationalinactivation of both rsmY and rsmZ led to low expressionof exoproduct genes and reduced biocontrol ability, closeto the properties of gacS and gacA mutants (Fig. 7;Tables 1 and 2). (iv) The effects of rsmY and rsmZ ontarget genes are mediated (at least partially) throughRsmA (Table 1). (v) RsmY, like RsmZ, specifically boundmultiple molecules of RsmA in vitro (Fig. 8), which sup-ports the idea that these riboregulators act throughsequestration of the translational regulator RsmA, therebypermitting the translation of the target genes examined.Thus, the GacS/GacA-dependent signal transductionpathway model now incorporates RsmY as a second ribo-regulator, in addition to RsmZ.
The fact that an rsmY rsmZ double mutant was notcompletely devoid of exoproducts as in the case of gacSor gacA mutants (Table 1, Fig. 7), suggests that therecould still be a third riboregulator in the system. Redun-dancy of riboregulators has also been observed recentlyin E. coli with the discovery of CsrC, a second riboregu-lator analogous to CsrB (Weilbacher et al., 2003). Thesignificance of such a redundancy of regulatory RNAswhich apparently fulfill similar functions in strain CHA0 ispresently not understood. Nevertheless, in this study, wehave observed that RsmY and RsmZ can have differen-tial effects on GacS/GacA-dependent genes that maypoint to a distinctive role for each riboregulator: (i) RsmZappears at higher cell densities than does RsmY (Fig. 4Cand D); (ii) an rsmZ mutation led to a 50% reduction ofphlA¢-‘lacZ expression whereas the effect of an rsmYmutation was minimal (Fig. 7C). The hcnA and aprAgenes are expressed earlier than the phlA operon and itis therefore not surprising to find that mutation of rsmZaffects phlA expression but had only marginal effects onhcnA and aprA expression at the end of growth (Fig. 7Aand B). The sequential appearance of RsmY and RsmZalso suggests that the promoters of the rsmY and rsmZgenes respond differentially to negative and/or positivecontrol elements in addition to GacA. In addition, RsmYand RsmZ appear to differ in their affinity for RsmA. Atlow RsmA concentration (~ 20 nM), RsmA preferentiallyinteracted with RsmY (Fig. 8A and B), whereas itappears that RsmZ was a more effective competitor than
1372 C. Valverde, S. Heeb, C. Keel and D. Haas
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
RsmY for RsmA binding at high RsmA concentrations(~ 1 mM) (Fig. 8C).
Contrary to CsrA/RsmA protein sequences (which arewell conserved amongst Gram-negative and Gram-posi-tive bacteria), the riboregulators CsrB and CsrC, RsmB,RsmY and RsmZ have little sequence identity betweenthemselves and differ in their size, but all have similarsecondary structures (Liu et al., 1997; Heeb et al., 2002;Weilbacher et al., 2003; Fig. 3B. In silico analysis of sev-eral rsmY genes from fluorescent pseudomonads showsthat a GGA motif occurs seven times (Fig. 1A) and aprediction of RsmY folding for CHA0 places four of thesemotifs in the single-stranded regions of several hairpinsand a fifth one between hairpins (Fig. 3B). Similarly, RsmZappears to present five of these motifs in loops or othersingle-stranded regions (Heeb et al., 2002). CsrB andCsrC RNAs of E. coli being larger than RsmY and RsmZ,have 18 and 9 stem-loops, respectively, exposing CAG
GAUG imperfect repeats that also contain the minimalGGA trinucleotide sequence (Liu et al., 1997; Weilbacheret al., 2003). From these structural features, it can behypothesized that repeated single-stranded, GGA ribonu-cleic elements exposed in unpaired regions couldrepresent a minimal requirement for multiple binding ofRsmA/CsrA. However, this still needs to be demonstratedexperimentally.
Experimental procedures
Bacterial strains and growth conditions
The bacterial strains, vectors and oligonucleotides used inthis study are listed in Table 3. Strains were routinelygrown in nutrient yeast broth [NYB; 2.5% (wt/vol) nutrientbroth, 0.5% (wt/vol) yeast extract] with shaking, or onnutrient agar [NA; 4% (wt/vol) blood agar base, 0.5% (wt/vol) yeast extract] amended with the following antibiotics,
Table 3. Bacterial strains, plasmids and oligonucleotides used in this study.
Strain, plasmidor oligonucleotide Genotype, relevant properties Reference or source
E. coliDH5a, HB101 Laboratory strains Sambrook and Russell (2001)P. aeruginosaPAO1 Wild type Holloway (1955)P. fluorescensCHA0 Wild type Voisard et al. (1994)CHA19 DgacS Zuber et al. (2003)CHA89 gacA::Kmr Laville et al. (1992)CHA96 Chromosomal translational gacA¢-‘lacZ fusion Laville et al. (1992)CHA207 Chromosomal translational hcnA¢-‘lacZ fusion Blumer et al. (1999)CHA89.207 Chromosomal translational hcnA¢-‘lacZ fusion, gacA::Kmr Blumer et al. (1999)CHA805 Chromosomal translational aprA¢-‘lacZ fusion Blumer et al. (1999)CHA806 Chromosomal translational aprA¢-‘lacZ fusion, DgacS Heeb et al. (2002)CHA809 rsmA::Kmr Blumer et al. (1999)CHA810 DrsmZ Heeb et al. (2002)CHA811 Chromosomal translational hcnA¢-‘lacZ fusion, DrsmZ Heeb et al. (2002)CHA812 Chromosomal translational aprA¢-‘lacZ fusion, DrsmZ Heeb et al. (2002)CHA822 DrsmY This studyCHA823 Chromosomal translational hcnA¢-‘lacZ fusion, DrsmY This studyCHA824 Chromosomal translational aprA¢-‘lacZ fusion, DrsmY This studyCHA825 DrsmY, DrsmZ This studyCHA826 Chromosomal translational hcnA¢-‘lacZ fusion, DrsmY, DrsmZ This studyCHA827 Chromosomal translational aprA¢-‘lacZ fusion, DrsmY, DrsmZ This studyCHA829 Chromosomal translational gacA¢-‘lacZ fusion, rsmA::Kmr This studyCHA830 DrsmY, DrsmZ, rsmA::Kmr This studyCHA831 Chromosomal translational hcnA¢-‘lacZ fusion, DrsmY, DrsmZ, rsmA::Kmr This studyCHA832 Chromosomal translational aprA¢-‘lacZ fusion, DrsmY, DrsmZ, rsmA::Kmr This studyPlasmidspBLS pBluescript II KS + cloning vector, ColE1 replicon, Apr StratagenepME497 Mobilizing plasmid; IncP-1, Tra; RepA– (Ts); Apr Voisard et al. (1988)pME3087 Suicide vector; ColE1 replicon, IncP-1–, Mob; Tcr Voisard et al. (1994)pME6000 Cloning vector, pBBR1MCS derivative; Tcr Maurhofer et al. (1998)pME6001 Cloning vector, pBBR1MCS derivative; Gmr Blumer et al. (1999)pME6010 pVS1-p15A shuttle cloning vector; Tcr Heeb et al. (2000)pME6016 pVS1-p15A shuttle vector for transcriptional lacZ fusions; Tcr Schnider-Keel et al. (2000)pME6032 lacIQ-Ptac expression vector; pVS1-p15A shuttle vector; Tcr Heeb et al. (2002)pME6032rsmY rsmY under Ptac promoter at position +42 in pME6032 This studypME6078 P. fluorescens CHA0 rsmA (encoding RsmA6H), in pME6032 Heeb et al. (2002)pME6081 Suicide plasmid for gene replacement containing rsmA::Kmr; Tcr Blumer et al. (1999)pME6091 Transcriptional rsmZ-lacZ fusion; Tcr Heeb et al. (2002)pME6259 Translational phlA¢-‘lacZ fusion; Tcr Schnider-Keel et al. (2000)pME6530 Translational hcnA¢-‘lacZ fusion; Tcr Blumer et al. (1999)
a. Specified restriction sites are underlined.

Small regulatory RNAs of P. fluorescens 1373
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
when required: ampicillin, 100 mg ml-1; gentamicin,10 mg ml-1; kanamycin, 25 mg ml-1; tetracycline, 25 mg ml-1
(100 mg ml-1 for P. fluorescens). Chloramphenicol wasused at a concentration of 15 mg ml-1 to select P. fluore-scens and to counterselect E. coli in mating experiments.When relevant, 5-bromo-4-chloro-3-indolyl-b-D-galactopyra-noside (X-gal) was added to plates at a final concentrationof 0.02%. Routine incubation temperatures were 37∞C forE. coli and P. aeruginosa, and 30∞C for P. fluorescens.Pseudomonas fluorescens was grown at 35∞C to improveits capacity to accept heterologous DNA (e.g. in transfor-
mation or triparental matings with E. coli). For the expres-sion of a phlA¢-‘lacZ translational fusion, P. fluorescensstrains were grown in a minimal glucose-ammoniummedium (OSG), containing 0.5% (wt/vol) D-glucose, 0.1%(wt/vol) ammonium sulphate, and 0.1% (wt/vol) Triton X-100, in addition to the salt solutions described by Ornstonand Stanier (1966).
DNA and RNA manipulations
Small- and large-scale plasmids preparations were done
pME6912 1.6 kb PCR product obtained with primers TRR5 and TRR6, containing the CHA0 rsmYgene, blunt-end cloned into EcoRV-cut pBLS
This study
pME6912hx pME3087 with the 1.6-kb HindIII/XbaI fragment from pME6912 This studypME6912b Plasmid rescued from CHA0::pME6912 This studypME6914 pME6000 with the XhoI/EcoRI 1.6-kb fragment from pME6912 This studypME6916 Transcriptional rsmY-lacZ fusion: pME6091 with the BamHI/PstI fragment containing
the rsmZ promoter replaced by the rsmY promoter as a PCR product obtained withprimers PTRRBAM and PTRRPST
This study
pME6917 pBLS with a 2.6-kb HindIII fragment from pME6912b, containing the rsmY gene, a1.4-kb DNA fragment upstream and a 0.9-kb genomic fragment downstream
This study
pME6917D pME3087 with a EcoRI/HindIII 2-kb PCR product derived from pME6917, containing adeletion of 92 bp in the rsmY gene flanked by 1-kb DNA fragments
This study
pME6918 Ptac-rsmY fusion at the +1 site This studypME6919 Template for in vitro transcription with rsmY under T7 promoter This studypME6920 Template for in vitro transcription with rsmZ under T7 promoter This studypME6926 Template for in vitro transcription with carA leader under T7 promoter This studypTZ19R Cloning vector for in vitro transcription assays FermentasOligonucleotidesa (5¢ 3¢)DT88 GAAGAGAAGGTGGAAATGGCGTTTTGG, 3¢-end anchor oligonucleotide. Tillett et al. (2000)DT89 CCAAAACGCCATTTCCACCTTCTCTTC, anchor-specific oligonucleotide. Tillett et al. (2000)FRSMYH TCTAAAGCTTATGGACGTCGCGCAGGAAGC, forward rsmY, HindIII site at -5. This studyFRSMZH TCAAAGCTTTGTCGACGGATAGACACAG, forward rsmZ, HindIII site at -5. This studyP1ECO GGCCGAATTCGAGCGTTCCTACCAGAACGC, forward upstream rsmY,
EcoRI site at -957.This study
P2BAM GACGGGATCCTGAAGTAGATTAGCTTCAGC, reverse rsmY, BamHI site at +4. This studyP3BAM AACAGGATCCAAAGCCCCGCTTCGGCGGGGTTTT, forward rsmY, BamHI site at +89. This studyP4HIN GATTAAGCTTAACTGCGTTATTTCGTCGCG, reverse downstream rsmY, HindIII site
at + 1070.This study
PAOTRR1 GTCAGGACATTGCGCAGGAAGC, forward rsmY of P. aeruginosa PAO1, annealsbetween +1 and +22.
This study
PAOTRR2 AAAACCCCGCCTTTTGGGCG, reverse rsmY of P. aeruginosa, anneals between +124and +105.
This study
PTRRBAM TACCTGGATCCATTTGGCTACACCGCC, forward rsmY promoter, BamHI site at -299. This studyPTRRPST TCCATCTGCAGTAGATTAGCTTCAGCCG, reverse rsmY promoter, PstI site at -2. This studyRRSMYP GATTCTGCAGAGAAAACCCCGCCGAAGCGG, reverse rsmY, PstI site at +122. This studyRRSMZP AACTGCAGCGGGACTTTTCGACAGACG, reverse rsmZ, PstI site at +146. This studyTACAAT TCCGGACGTCCATCCACACATTATACGAGCCGA, reverse Ptac, natural AatII site in
rsmY at +8.This study
TETA1 AACCCAAAGGAAAGGCGCTGTC, anneals between the EcoRV and AatII sites oftetA in pME6032
Heeb et al. (2002)
TRR4 AAGAAAACCCCGCCGAAGCG, reverse rsmY; anneals between +121 and +102. This studyTRR4XHO GATTCTCGAGAGAAAACCCCGCCGAAGCGG, reverse rsmY, XhoI site at +126 This studyTRR5 GGYCTGTACSCGATGCTGAC, forward upstream rsmY of P. fluorescens Pf-1, P. putida
KT2440 and P. syringae pv tomato.This study
TRR6 AAGAAAACCCCGCCKAAGCG, reverse rsmY of P. fluorescens Pf-1, P. putida KT2440and P. syringae pv tomato; anneals between +121 and +102.
This study
TRR7 ATGGACGTCGCGCAGGAAGC, forward rsmY; anneals between +1 and +20. This studyTRR7ECO TCTAGAATTCATGGACGTCGCGCAGGAAGC, forward rsmY, EcoRI site at -6. This studyTRR10 TCCCTGACATCCTTTTCACTCCACC, reverse rsmY, anneals between +84 and +60. This studyTRR11 TGGCGGAATCCTACGTGTCCGTGTTG, reverse rsmY, anneals between +55 and +30. This studyT3, T7 pBLS primers Stratagene
Strain, plasmidor oligonucleotide Genotype, relevant properties Reference or source
a. Specified restriction sites are underlined.
Æ
Table 3. cont.

1374 C. Valverde, S. Heeb, C. Keel and D. Haas
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
with the cetyltrimethylammonium bromide method (Del Salet al., 1988) and the Jetstar kit (Genomed GmbH) respec-tively. Chromosomal DNA from P. fluorescens was preparedas previously described (Gamper et al., 1992). Total RNAwas extracted with the High Pure RNA isolation kit (Roche)according to manufacturer’s recommendations. DNA manip-ulations were carried out as described (Sambrook and Rus-sell, 2001). DNA fragments were purified from agarose gelswith the Qiagen Gel extraction kit (Qiagen). DNA sequenc-ing was performed with the Big Dye Terminator Cyclesequencing kit and an ABI-prism 373 automatic sequencer(Applied Biosystems).
Polymerase chain reactions were typically carried out with2 U of thermostable DNA polymerase (Extra-Pol II, Eurobio)in a reaction mixture containing target DNA, 200 mM of eachof the four dNTPs (Roche), 20 pmol of each of two primers,1.5 mM MgCl2 and 1¥ Extra-Pol buffer in a final volume of20 ml. For the amplification reaction, an initial denaturationstep of 2 min at 95°C was followed by 25–30 cycles [45 s at95°C, 45 s at 50–65°C (depending on the G + C content andlength of the primers), and 0.5–2 min at 72°C (depending onthe length of the amplicon)] and a final elongation step of5 min at 95°C.
Southern and Northern blots
DNA was blotted from agarose gels to Hybond-N nylon mem-branes (Amersham Biosciences). RNA for Northern blots waselectrophoretically separated on denaturing urea-polyacryla-mide gels (8.3 M urea, 8% acrylamide, 0.2% bisacrylamide)in 1¥ TBE buffer [50 mM Tris-borate (pH 8.3), 1 mM EDTA].The lane corresponding to the molecular weight markers (lowrange RNA ladder; Fermentas) was cut out, stained withethidium bromide (5 mg ml-1 in 1¥ TBE) and photographedunder UV light beside a reference ruler. The rest of the gelwas electroblotted at 150 mA (15–25 V) onto a Hybond-Nmembrane in 1¥ TBE buffer for 20 min. All membranes werewashed with 2¥ SSC (1¥ SSC is 0.15 M NaCl and 15 mMsodium citrate) before nucleic acids were cross-linked byexposing the membranes to UV light for 5 min (Sambrookand Russell, 2001). Southern and Northern hybridizationswere done with digoxigenin (DIG)-labelled DNA probesaccording to recommended protocols (Roche’s DIG SystemUser’s Guide for Filter Hybridization) or with DNA probesobtained by replacing dNTPs in a standard PCR with DIGDNA labelling mix (Roche).
Cloning of the rsmY gene (TRR locus) from P. fluorescens CHA0
The rsmY gene and its surroundings (1 kb on both sides)were obtained from available genome sequences for fluores-cent pseudomonads (see legend of Fig. 1). Two degenerateprimers (TRR5 and TRR6) were designed based on the
sequence alignment and used to PCR-amplify the rsmYlocus from P. fluorescens CHA0 DNA. A 1.6 kb band wasobtained with an annealing temperature of 65∞C. The PCRproduct (10 ml) was blunt-end cloned into pBluescript (pBLS)in a reaction mixture containing 500 ng of the cloning vector,5 U of EcoRV, 5 U of T4 DNA polymerase, 6.25 U of T4 DNAligase, 200 mM of each of the four dNTPs and 1¥ ligationbuffer in a final volume of 17.5 ml. This resulted in pME6912.The identity of the cloned 1.6 kb fragment was confirmed bysequencing.
Chromosome walking downstream of rsmY
A 1.6 kb HindIII/XbaI fragment from pME6912 containing theCHA0 rsmY gene locus was subcloned into the HindIII/XbaI-digested vector pME3087. After integration of the resultingsuicide plasmid into the chromosome of strain CHA0 by tri-parental mating using E. coli HB101/pME497 as the mobiliz-ing strain (Voisard et al., 1988), the integrated plasmid wasrescued by digesting the chromosomal DNA with BamHIand ligation. The resulting plasmid, pME6912b, had anadditional 6.5 kb fragment downstream of the initial 1.6 kbHindIII/XbaI fragment. A HindIII 2.6 kb fragment frompME6912b, containing the rsmY gene bordered by 1.4 kb ofupstream and 0.9 kb of downstream chromosomal DNA,was subcloned into HindIII-digested pBLS to producepME6917. The 2.6 kb HindIII insert in pME6917 wassequenced by Microsynth (Balgach, Switzerland; GenBankaccession AY266632, Fig. 3A).
Determination of rsmY 5¢-end
The 5¢-end of the rsmY transcript was mapped by RACE(Tillett et al., 2000). Total RNA from strain CHA0 was isolatedfrom cells collected at late stationary phase. Reverse tran-scription (RT) reaction was performed using 30 pmol ofprimer TRR10 and 6 U of reverse transcriptase (OmniscriptRT kit, Qiagen Inc), in a final volume of 50 ml containing 1¥Qiagen RT buffer, 20 mM DTT, 45 U of RNAse inhibitor(Amersham Biosciences), 1 mM dNTPs and 10 mg of totalRNA from CHA0 cells collected in stationary phase. Theprimer was annealed stepwise (100∞C for 3 min, 60∞C for 2 h,37∞C for 1 h and 25∞C for 30 min) in 1¥ annealing buffer[20 mM Tris.HCl, 0.2 M NaCl, 0.1 M EDTA (pH 8.0)] in 30 mlbefore addition of the other RT components. The RT reactionwas performed at 42∞C for 60 min. After RNA removal byalkaline hydrolysis and neutralization, cDNA was precipitatedand resuspended in 20 ml of water. T4 RNA ligase (NewEngland Biolabs) was used to anchor a 5¢-phosphorylated,3¢-end ddATP-blocked oligonucleotide DT88 (P-DT88-ddATP) to the single-stranded cDNA. The ligation was doneovernight at room temperature with 1.3 ml of 10¥ T4 RNAligase buffer, 6.5 pmol of P-DT88-ddATP, 10 ml of the cDNAand 8 U of T4 RNA ligase. The resulting ligation mixture wasused directly without purification in the subsequent semin-ested PCRs. The anchor-ligated cDNA (1 ml) was amplifiedfirst with primers DT89 (anchor-specific primer) and TRR10,followed by amplification with primer DT89 and the internalprimer TRR11. An 82 bp band was purified from an agarosegel and blunt-end cloned into pBLS as described above.

Small regulatory RNAs of P. fluorescens 1375
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
Three independent clones were sequenced using pBLSprimer T7 (data not shown; Fig. S1).
Overexpression of rsmY
A PCR product obtained from pME6912 with primersTRR7ECO and TRR4XHO was digested with EcoRI and XhoIand cloned into EcoRI/XhoI-cut pME6032, producingpME6032rsmY. To fuse the rsmY gene exactly to the +1transcription start site of Ptac (Amann et al., 1983), a 42 bpdeletion was created by PCR using oligonucleotides TETA1and TACAAT. Primer TACAAT joins the -1 nucleotide of Ptac
to the first nine nucleotides of rsmY, six of which naturallyform an AatII site (see Fig. 3A). Primer TETA1 is complemen-tary to a region in the tetA tetracycline resistance marker inpME6032rsmY, annealing upstream of an AatII site. Theresulting 3.2 kb product was digested with AatII and used toreplace the corresponding 3.2 kb AatII fragment inpME6032rsmY, producing pME6918. Because this constructwas deleterious to the growth of E. coli strains even whenthe tac promoter was not induced, the last subcloning stepwas done by transforming P. fluorescens CHA806, in whichthis construct did not have a detrimental effect. The Ptac-rsmYfusion on pME6918 was checked by sequencing.
Construction of an rsmY-lacZ fusion
A reporter plasmid was constructed carrying a transcriptionalrsmY-lacZ fusion in which the +1 nucleotide of lacZ corre-sponds to the +1 nucleotide of the rsmY promoter. First, thersmY promoter was PCR-amplified from pME6912 with prim-ers PTRRBAM and PTRRPST. This product was digestedwith BamHI (artificially introduced by PTRRBAM upstream ofthe rsmY promoter) and PstI (artificially introduced by PTR-RPST in the +1 region) before being exchanged by the cor-responding BamHI/PstI fragment of pME6091 containing thersmZ promoter fused to the lacZ ORF (Heeb et al., 2002), toproduce pME6916. This rsmY reporter construct waschecked by sequencing.
Chromosomal deletion of rsmY
Two PCR products flanking the rsmY structural gene wereobtained from pME6917 using primers P1ECO with P2BAMand P3BAM with P4HIN. Polymerase chain reaction productswere digested with EcoRI/BamHI (the 1.1 kb upstream frag-ment) and BamHI/HindIII (the 0.9 kb downstream fragment)and ligated to generate a 2 kb product that created a 92 bpdeletion of the rsmY gene (positions +1 to +92) but leavingintact its promoter and terminator (Fig. 3A). The strategyintroduced an artificial BamHI site downstream of the rsmYpromoter. To produce a suicide plasmid for gene replace-ment, pME6917D, the 2 kb EcoRI/HindIII fragment wascloned into EcoRI/HindIII-cut pME3087. rsmY-negativemutants were obtained by integration of the suicide plasmidpME6917D into the chromosome of strains CHA0, CHA207and CHA805 after triparental mating as above, with selectionfor tetracycline-resistant (Tcr), chloramphenicol-resistant(Cmr) recombinants. After a second crossing-over, Tcs
recombinants were obtained, producing strains CHA822,
CHA823 and CHA824, respectively. Similarly, the rsmY-negative/rsmZ-negative double mutants CHA825, CHA826and CHA827 were obtained by gene replacement in strainsCHA810, CHA811 and CHA812.
Chromosomal inactivation of rsmA
The rsmA-negative mutants CHA829, CHA830, CHA831 andCHA832 were obtained by gene replacement after integrationof the suicide plasmid pME6081 (rsmA::Kmr; Blumer et al.,1999) into the chromosome of strains CHA96, CHA825,CHA826 and CHA827, respectively, and selection for Tcs Kmr
recombinants.
Purification of histidine-tagged RsmA and associated RNAs
Purification of the RsmA6H protein was performed from P.fluorescens CHA0 cells bearing the expression vectorpME6078 as previously described (Heeb et al., 2002). RNAwas separated from the purified protein fractions by twoextractions with phenol: chloroform (1:1) and once with chlo-roform. RNA was ethanol-precipitated and resuspended indiethylpyrocarbonate-treated water. For the mobility shiftassays in polyacrylamide gels, RsmA6H was overexpressedin E. coli DH5a/pME6078 cells, and the protein eluate fromthe Ni-NTA column (Qiagen) was dialysed overnight against10 mM Tris-acetate (pH 8.0) at 4∞C. Protein content was esti-mated with the Bradford method with bovine serum albuminas the standard. The homogeneity of the preparation wasassessed by native polyacrylamide gel electrophoresis.RsmA6H accounted for ≥90% of the protein content (data notshown).
In vitro transcription of rsmY and rsmZ
Both genes were PCR-amplified and cloned under the controlof the T7 promoter in the vector pTZ19R, to produce plasmidspME6919 (rsmY; primers FRSMYH, RRSMYP) andpME6920 (rsmZ; primers FRSMZH, RRSMZP). Radioac-tively labelled RsmY and RsmZ transcripts were synthesizedin vitro with a T7 transcription kit (Fermentas) following themanufacturer’s instructions in the presence of [a-33P]-UTPand linearized pME6919 or pME6920. RNA was purified byphenol: chloroform extraction and desalted with Sephadex G-25 minicolumns (Amersham Biosciences). Unlabelled RsmYand RsmZ RNAs, and the 115-nt 5¢-leader RNA of the carAgene of strain CHA0, were synthesized following the sameprotocol, but with cold UTP, from linearized pME6919,pME6920, and pME6926. RNA concentration was estimatedby UV absorption at 260 nm.
Gel mobility shift assay
Binding reactions (10 ml) contained 10 mM Tris.acetate atpH 7.5, 10 mM MgCl2, 50 mM NaCl, 50 mM KCl, 10 mM DTT,5% (w/v) glycerol, 27.5 ng of yeast tRNA, 4 U of RNAseinhibitor (Roche), 8 nM RsmY (45000 c.p.m.) or 9 nM RsmZ(77000 c.p.m.) labelled with [a-33P]-UTP, and purified

1376 C. Valverde, S. Heeb, C. Keel and D. Haas
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
RsmA6H (at various concentrations). Assays were also car-ried out in the presence of various unlabelled RNA compet-itors (see text for details). Reaction mixtures were incubatedfor 30 min at 30∞C to allow complex formation. Samples werethen fractionated on native 10% polyacrylamide gels andradioactive bands were visualized by autoradiography afterdrying the gels.
Detection of HCN, exoprotease and DAPG
HCN production was measured as previously described(Voisard et al., 1989). Qualitative exoprotease activity wasdetected on skim-milk nutrient agar [0.4% (wt/vol) blood agarbase, 0.05% (wt/vol) yeast extract, 1.35% (wt/vol) agar, sup-plemented with 15% (v/v) of skim milk] (Sacherer et al.,1994). DAPG was extracted with ethyl acetate from cell cul-tures (3–5 replicates) grown in 100 ml of OSG medium tostationary phase (OD600 3.5–4.5). The extract was driedunder vacuum, resuspended in 200 ml of methanol and anal-ysed by HPLC as described (Keel et al., 1992).
Extraction of the non-AHL signal
Culture supernatants of strain CHA0, CHA89 and CHA825were prepared and extracted with dichloromethane asdescribed (Heeb et al., 2002).
b-Galactosidase assays
b-Galactosidase activities were quantified by the Millermethod (Miller, 1972), using cells permeabilized with 5%toluene. Pseudomonas fluorescens cells were grown in 20 mlof NYB (in 100-ml Erlenmeyers) with shaking at 30∞C, exceptfor expression of phlA¢-‘lacZ where growth was in OSG. TritonX-100 [0.1% (wt/vol)] was added to avoid cell aggregation.
Biocontrol assay in natural soil
For the assay aliquots of 60 g of natural sandy loam soil fromEschikon, Switzerland, were distributed into 200 ml Erlenm-eyer flasks with wide openings. When appropriate, the soilwas artificially infested with the pathogenic oomycete Pyth-ium ultimum strain 67–1 by adding 2.5 g of a five-day-oldmillet-seed inoculum of the pathogen per kg of soil. Threesterile-grown, 72 h-old cucumber seedlings (Cucumis sativuscv. Chinese Snake) were then placed in each flask as previ-ously described (Keel et al., 2002). Pseudomonas fluore-scens strains were added to soil as a suspension (5 ml perflask) of cells washed twice in sterile distilled water to give1 ¥ 107 CFU/g of soil. Control flasks received the sameamount of sterile water. Seedlings were covered with 10 g ofnon-treated soil and flasks were sealed with cotton stoppers.The microcosms were incubated in a randomized blockdesign in a growth chamber containing 80% relative humidityand 22°C with light (200 mmol/s m-2; ratio of 1.37 of light at
660 nm to 730 nm) for 16 h, followed by an 8 h dark periodat 15°C. No watering was necessary. At six days after inoc-ulation, the percentage of plants bearing symptoms of fungalattack was determined. Plants were removed from the flasks,washed, briefly dried with paper towels and weighed. Rootsfrom one flask were pooled and transferred into a 50 mlplastic tube containing 20 ml of sterile distilled water. Tubeswere vigorously shaken at 240 r.p.m. for 30 min and theresulting suspensions were used to determine CFU numbers.Data represent the means from 12 to 30 replicates per treat-ment (one replicate corresponds to one flask containing threeplants). Data were analysed for significance with varianceanalysis, followed by Fisher’s least-significant-difference test.
Acknowledgements
We thank Patrick Michaux and Maria Péchy for technicalhelp. Support was obtained through grants from the SwissNational Foundation for Scientific Research (projects 31–56608.99 and 3100A0-100180), the Roche Research Foun-dation, and the EU project ECOSAFE (QLK3-2000–31759).
Supplementary material
The following material is available from http://www.blackwellpublishing.com/products/journals/suppmat/mmi/mmi3774/mmi3774sm.htmFig. S1. Determination of the 5’-end of the rsmY transcript.Fig. S2. Northern blot of rsmY mutants.Fig. S3. rsmA does not regulate gacA.Fig. S4. 5’-leader region of the carA transcript of P. fluore-scens strain CHA0 as an unspecific competitor for electro-phoretic mobility shift assays.
References
Aarons, S., Abbas, A., Adams, C., Fenton, A., and O’Gara,F. (2000) A regulatory RNA (PrrB RNA) modulates expres-sion of secondary metabolites in Pseudomonas fluore-scens F113. J Bacteriol 182: 3913–3919.
Altier, C., Suyemoto, M., Ruiz, A.I., Burnham, K.D., and Mau-rer, R. (2000) Characterization of two novel regulatorygenes affecting Salmonella invasion gene expression. MolMicrobiol 35: 635–646.
Amann, E., Brosius, J., and Ptashne, M. (1983) Vectorsbearing a hybrid trp-lac promoter useful for regulatedexpression of cloned genes in Escherichia coli. Gene 25:167–178.
Ando, Y., Asari, S., Suzuma, S., Yamane, K., and Nakamura,K. (2002) Expression of a small RNA, BS203 RNA, fromthe yocI-yocJ intergenic region of Bacillus subtilis genome.FEMS Microbiol Lett 207: 29–33.
Argaman, L., Hershberg, R., Vogel, J., Bejerano, G., Wagner,E.G.H., Margalit, H., et al. (2001) Novel small RNA-encoding genes in the intergenic regions of Escherichiacoli. Curr Biol 11: 941–950.
Baker, C.S., Morozov, I., Suzuki, K., Romeo, T., andBabitzke, P. (2002) CsrA regulates glycogen biosynthesis

Small regulatory RNAs of P. fluorescens 1377
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
by preventing translation of glgC. Escherichia coli. MolMicrobiol 44: 1599–1610.
Blumer, C., and Haas, D. (2000) Mechanism, regulation, andecological role of bacterial cyanide biosynthesis. ArchMicrobiol 173: 170–177.
Blumer, C., Heeb, S., Pessi, G., and Haas, D. (1999) GlobalGacA-steered control of cyanide and exoprotease produc-tion in Pseudomonas fluorescens involves specific ribo-some binding sites. Proc Natl Acad Sci USA 96: 14073–14078.
Buell, C.R., Joardar, V., Lindeberg, M., Selengut, T.,Paulsen, I.T., Gwinn, M.L., et al. (2003) The completegenome sequence of the Arabidopsis and tomato patho-gen Pseudomonas syringae pv. tomato DC3000. Proc NatlAcad Sci USA 100: 10181–10186.
Chatterjee, A., Cui, Y., and Chatterjee, A.K. (2002) RsmAand the quorum-sensing signal, N-(3-oxohexanoyl)-L-homoserine lactone, control the levels of rsmB RNA inErwinia carotovora subsp. carotovora by affecting itsstability. J Bacteriol 184: 4089–4095.
Cui, Y., Chatterjee, A., and Chatterjee, A.K. (2001) Effectsof the two-component system comprising GacA andGacS of Erwinia carotovora subsp. carotovora on the pro-duction of global regulatory rsmB RNA, extracellularenzymes, and harpin (Ecc). Mol Plant-Microbe Interact14: 516–526.
Del Sal, G., Manfioletti, G., and Schneider, C. (1988) A one-tube plasmid DNA mini-preparation suitable for sequenc-ing. Nucleic Acids Res 16: 9878.
Dubey, A.K., Baker, C.S., Suzuki, K., Jones, A.D., Pandit, P.,Romeo, T., and Babitzke, P. (2003) CsrA regulates trans-lation of the Escherichia coli carbon starvation gene, cstA,by blocking ribosome access to the cstA transcript. J Bac-teriol 185: 4450–4460.
Gamper, M., Ganter, B., Polito, M.R., and Haas, D. (1992)RNA processing modulates the expression of the arcDABCoperon in Pseudomonas aeruginosa. J Mol Biol 226: 943–957.
Gottesman, S. (2002) Stealth regulation: biological circuitswith small RNA switches. Genes Dev 16: 2829–2842.
Guigueno, A., Dassa, J., Belin, P., and Boquet, P.L. (2001)Oversynthesis of a new Escherichia coli small RNA sup-presses export toxicity of DsbA¢-PhoA unfoldable periplas-mic proteins. J Bacteriol 183: 1147–1158.
Haas, D., Keel, C., and Reimmann, C. (2002) Signal trans-duction in plant-beneficial rhizobacteria with biocontrolproperties. Antonie Van Leeuwenhoek 81: 385–395.
Heeb, S., and Haas, D. (2001) Regulatory roles of the GacS/GacA two-component system in plant-associated andother Gram-negative bacteria. Mol Plant-Microbe Interact14: 1351–1363.
Heeb, S., Itoh, Y., Nishijyo, T., Schnider, U., Keel, C., et al.(2000) Small, stable shuttle vectors based on the minimalpVS1 replicon for use in Gram-negative, plant-associatedbacteria. Mol Plant-Microbe Interact 13: 232–237.
Heeb, S., Blumer, C., and Haas, D. (2002) Regulatory RNAas mediator in GacA/RsmA-dependent global control ofexoproduct formation in Pseudomonas fluorescens CHA0.J Bacteriol 184: 1046–1056.
Holloway, B.W. (1955) Genetic recombination in Pseudomo-nas aeruginosa. J Gen Microbiol 13: 572–581.
Keel, C., and Défago, G. (1997) Interactions between bene-ficial soil bacteria and root pathogens: mechanisms andecological impact. In Multitrophic Interactions in TerrestrialSystems. Gange, A.C., and Brown, V.K. (eds). London:Blackwell Scientific Publishers, pp. 27–46.
Keel, C., Schnider, U., Maurhofer, M., Voisard, C., Laville, J.,Burger, U., et al. (1992) Suppression of root diseases byPseudomonas fluorescens CHA0: importance of the sec-ondary metabolite 2,4-diacetylphloroglucinol. Mol Plant-Microbe Interact 5: 4–13.
Keel, C., Ucurum, Z., Michaux, M., Adrian, M., and Haas, D.(2002) Deleterious impact of a virulent bacteriophage onsurvival and biocontrol activity of Pseudomonas fluore-scens strain CHA0 in natural soil. Mol Plant-Microbe Inter-act 15: 567–576.
Laville, J., Voisard, C., Keel, C., Maurhofer, M., Défago, G.,and Haas, D. (1992) Global control in Pseudomonas fluo-rescens mediating antibiotic synthesis and suppression ofblack root rot of tobacco. Proc Natl Acad Sci USA 89:1562–1566.
Laville, J., Blumer, C., Von Schroetter, C., Gaia, V., Défago,G., Keel, C., et al. (1998) Characterization of the hcnABCgene cluster encoding hydrogen cyanide synthase andanaerobic regulation by ANR in the strictly aerobic biocon-trol agent Pseudomonas fluorescens CHA0. J Bacteriol180: 3187–3196.
Liu, M.Y., and Romeo, T. (1997) The global regulator CsrAof Escherichia coli is a specific mRNA-binding protein. JBacteriol 179: 4639–4642.
Liu, M.Y., Yang, H., and Romeo, T. (1995) The product of thepleiotropic Escherichia coli gene csrA modulates glycogenbiosynthesis via effects on mRNA stability. J Bacteriol 177:2663–2672.
Liu, M.Y., Gui, G., Wei, B., Preston, J.F., III, Oakford, L.,Yüksel, Ü. et al. (1997) The RNA molecule CsrB binds tothe global regulatory protein CsrA and antagonizes itsactivity in Escherichia coli. J Biol Chem 272: 17502–17510.
Liu, Y., Cui, Y., Mukherjee, A., and Chatterjee, A.K. (1998)Characterization of a novel RNA regulator of Erwinia caro-tovora ssp. carotovora that controls production of extracel-lular enzymes and secondary metabolites. Mol Microbiol29: 219–234.
Ma, W., Cui, Y., Liu, Y., Dumenyo, K.C., Mukherjee, Y.C.A.,and Chatterjee, A.K. (2001) Molecular characterization ofglobal regulatory RNA species that control pathogenicityfactors in Erwinia amylovora and Erwinia herbicola pv. Gyp-sophilae. J Bacteriol 183: 1870–1880.
Majdalani, N., Cunning, C., Sledjeski, D., Elliott, T., and Got-tesman, S. (1998) DsrA RNA regulates translation of RpoSmessage by an anti-antisense mechanism, independent ofits action as an antisilencer of transcription. Proc Natl AcadSci USA 95: 12462–12467.
Majdalani, N., Hernandez, D., and Gottesman, S. (2002)Regulation and mode of action of the second small RNAactivator of RpoS translation, RprA. Mol Microbiol 46: 813–826.
Massé, E., and Gottesman, S. (2002) A small RNA regu-lates the expression of genes involved in iron metabolismin Escherichia coli. Proc Natl Acad Sci USA 99: 4620–4625.

1378 C. Valverde, S. Heeb, C. Keel and D. Haas
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
Maurhofer, M., Keel, C., Haas, D., and Défago, G. (1994)Pyoluteorin production by Pseudomonas fluorescens strainCHA0 is involved in the suppression of Pythium damping-off of cress but not of cucumber. Eur J Plant Pathol 100:221–232.
Maurhofer, M., Keel, C., Haas, D., and Défago, G. (1995)Influence of plant species on disease suppression byPseudomonas fluorescens strain CHA0 with enhancedantibiotic production. Plant Pathol 44: 40–50.
Maurhofer, M., Reimmann, C., Schmidli-Sacherer, P., Heeb,S., Haas, D., and Défago, G. (1998) Salicylic acid biosyn-thetic genes expressed in Pseudomonas fluorescens strainP3 improve the induction of systemic resistance in tobaccoagainst tobacco necrosis virus. Phytopathology 88: 678–684.
Miller, J.H. (1972) Experiments in Molecular Genetics. ColdSpring Harbor, NY: Cold Spring Harbor Laboratory Press.
Møller, T., Franch, T., Hojrup, P., Keene, D.R., Bächinger,H.P., Brennan, R.G., et al. (2002) Hfq: a bacterial Sm-likeprotein that mediates RNA–RNA interaction. Mol Cell 9:23–30.
Nelson, K.E., Weinel, C., Paulsen, I.T., Dodson, R.J., Hilbert,H., Martins Dos Santos, V.A., et al. (2002) Completegenome sequence and comparative analysis of the meta-bolically versatile Pseudomonas putida KT2440. EnvironMicrobiol 4: 799–808.
Oberhänsli, T., and Défago, G. (1991) Spontaneous loss oftryptophan side chain oxydase of Pseudomonas fluore-scens strain CHA0 – a marker for genetic instability. InPlant Growth-Promoting Rhizobacteria – Progress andProspects. Keel, C., Koller, B., and Défago, G. (eds).IOBC/WPRS Bulletin XIV/8, Zurich, Switzerland, pp. 392–398.
Ornston, L.N., and Stanier, R.Y. (1966) The conversion ofcathecol and protocatechuate to b-ketoadipate byPseudomonas putida. J Biol Chem 241: 3776–3786.
Pernestig, A.K., Melefors, O., and Georgellis, D. (2001) Iden-tification of UvrY as the cognate response regulator for theBarA sensor kinase in Escherichia coli. J Biol Chem 276:225–231.
Pernestig, A.K., Georgellis, D., Romeo, T., Suzuki, K., Tome-nius, H., Normark, S., et al. (2003) The Escherichia coliBarA-UvrY two-component system is needed for efficientswitching between glycolytic and gluconeogenic carbonsources. J Bacteriol 185: 843–853.
Rivas, E., Klein, R.J., Jones, T.A., and Eddy, S.R. (2001)Computational identification of noncoding RNAs in E. coliby comparative genomics. Curr Biol 11: 1369–1373.
Romeo, T. (1998) Global regulation by the small RNA-bindingprotein CsrA and the non-coding RNA molecule CsrB. MolMicrobiol 29: 1321–1330.
Rowley, K.B., Clements, D.E., Mandel, M., Humphreys, T.,and Patil, S.S. (1993) Multiple copies of a DNA sequencefrom Pseudomonas syringae pathovar phaseolicola abol-ish thermoregulation of phaseolotoxin production. MolMicrobiol 8: 625–635.
Sacherer, P., Défago, G., and Haas, D. (1994) Extracellularprotease and phospholipase C are controlled by the globalregulatory gene gacA in the biocontrol strain Pseudomo-nas fluorescens CHA0. FEMS Microbiol Lett 116: 155–160.
Sambrook, J., and Russell, D.W. (2001) Molecular Cloning:a Laboratory Manual, 3rd edn. Cold Spring Harbor, NY:Cold Spring Harbor Laboratory Press.
Schnider-Keel, U., Seematter, A., Maurhofer, M., Blumer, C.,Duffy, B., Gigot-Bonnefoy, C., et al. (2000) Autoinductionof 2,4-diacetylphloroglucinol biosynthesis in the biocontrolagent Pseudomonas fluorescens CHA0 and repression bythe bacterial metabolites salicylate and pyoluteorin. J Bac-teriol 182: 1215–1225.
Storz, G. (2002) An expanding universe of noncoding RNAs.Science 296: 1260–1263.
Stover, C.K., Pham, X.Q., Erwin, A.L., Mizoguchi, S.D., War-rener, P., Hickey, M.J., et al. (2000) Complete genomesequence of Pseudomonas aeruginosa PAO1, an opportu-nistic pathogen. Nature 406: 959–964.
Suzuki, K., Wang, X., Weilbacher, T., Pernestig, A.K., Mele-fors, O., Georgellis, D., et al. (2002) Regulatory circuitry ofthe CsrA/CsrB and BarA/UvrY systems of Escherichia coli.J Bacteriol 184: 5130–5140.
Tillett, D., Burns, B.P., and Neilan, B.A. (2000) Opti-mized rapid amplification of cDNA ends (RACE) formapping bacterial mRNA transcripts. Biotechniques 28:448–456.
Voisard, C., Rella, M., and Haas, D. (1988) Conjugativetransfer of plasmid RP1 to soil isolates of Pseudomonasfluorescens is facilitated by certain large RP1 deletions.FEMS Microbiol Lett 55: 9–14.
Voisard, C., Keel, C., Haas, D., and Défago, G. (1989) Cya-nide production by Pseudomonas fluorescens helpssupress black root rot of tobacco under gnotobiotic condi-tions. EMBO J 8: 351–358.
Voisard, C., Bull, C.T., Keel, C., Laville, J., Maurhofer, M.,Schnider, M., et al. (1994) Biocontrol of root diseases byPseudomonas fluorescens CHA0: current concepts andexperimental approaches. In Molecular Ecology of Rhizo-sphere Microorganisms. O’Gara, F., Dowling, D.N., andBoesten, B. (eds). Germany: VCH-VerlagsgesellschaftmbH, pp. 67–89.
Wagner, G.H., Altuvia, S., and Romby, P. (2002) AntisenseRNAs in bacteria and their genetic elements. Adv Genet46: 361–398.
Wassarman, K.M. (2002) Small RNAs in bacteria: diverseregulators of gene expression in response to environmen-tal changes. Cell 109: 141–144.
Wassarman, K.M., Zhang, A., and Storz, G. (1999)Small RNAs in Escherichia coli. Trends Microbiol 7:37–45.
Wassarman, K.M., Repoila, F., Rosenow, C., Storz, G., andGottesman, S. (2001) Identification of novel small RNAsusing comparative genomics and microarrays. Genes Dev15: 1637–1651.
Weilbacher, T., Suzuki, K., Dubey, A.K., Wang, X., Gudapaty,S., Morozov, I., et al. (2003) A novel sRNA component ofthe carbon storage regulatory system of Escherichia coli.Mol Microbiol 48: 657–670.
White, D., Hart, M.E., and Romeo, T. (1995) Phylogeneticdistribution of the global regulatory gene csrA amongeubacteria. Gene 182: 221–223.
Zuber, S., Carruthers, F., Keel, C., Mattart, A., Blumer, C., etal. (2003) GacS sensor domains pertinent to the regulationof exoproduct formation and to the biocontrol potential of

Small regulatory RNAs of P. fluorescens 1379
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 50, 1361–1379
Pseudomonas fluorescens CHA0. Mol Plant-Microbe Inter-act 16: 634–644.
Zuker, M., Mathews, D.H., and Turner, D.H. (1999) Algo-rithms and thermodynamics for RNA secondary structure
prediction: a practical guide. In RNA Biochemistry andBiotechnology. Barciszewski, J., and Clark, B.F.C. (eds).NATO ASI Series. Dordrecht: Kluwer Academic Publish-ers, pp. 11–43.




















