
Spectral properties of pseudo-resolvents under structured perturbations
Upload
independentCategory
view
0download
0
Mathematical Biosciences 220 (2009) 34–44
Contents lists available at ScienceDirect
Mathematical Biosciences
journal homepage: www.elsevier .com/locate /mbs
Periodic oscillation in delayed gene networks with SUM regulatory logicand small perturbations q
Yi Wang a,c, Zhongjun Ma b, Jianwei Shen a, Zengrong Liu d,*, Luonan Chen d,*
a Shanghai University College of Science, Shanghai University , Shanghai, 200444, Chinab School of Mathematical Science and Computing Technology, Central South University, Changsha, 410083, Chinac ZheJiang University of Finance and Economics, Hangzhou, 310012, Chinad Institute of Systems Biology, Shanghai University, Shanghai, 200444, China
a r t i c l e i n f o
Article history:Received 19 October 2008Received in revised form 22 March 2009Accepted 31 March 2009Available online 17 April 2009
Keywords:Gene networkPeriodic oscillationDistributed delay
0025-5564/$ - see front matter � 2009 Elsevier Inc. Adoi:10.1016/j.mbs.2009.03.010
q Supported by NSF of China with No.10701052 108Shanghai Municipality (S30104).
* Corresponding authors.E-mail addresses: [email protected] (Y. Wa
Liu), [email protected] (L. Chen).
a b s t r a c t
In this paper, we derive new criteria for evaluating the global stability of periodic oscillation in delayedgene networks with SUM regulatory logic and small perturbation, which appear in many biological sys-tems at biomolecular or cellular levels due to the weak coupling and signal diffusion (or transport) pro-cess. Our results rely on the Lipschtiz conditions of Hill function, topology of gene networks and delaykernels. In particular, Our method based on the proposed model transforms the original network intomatrix analysis problem, thereby not only significantly reducing the computational complexity but alsomaking analysis of periodic oscillation tractable for even large-scale nonlinear networks.
� 2009 Elsevier Inc. All rights reserved.
1. Introduction
It is well known that the gene networks (GNs) plays a key rolein regulating dynamics of processes transducing genetic signalsinto phenotypic variation and integrating genomic information,environmental cues, and physiological or developmental stimuli.Thus, understanding the architectures and their design principlesof GNs will fundamentally advance the study of core biologicalproblems [1,2]. In particular, since genetic regulatory networksare high-dimensional and nonlinear, it is indispensable to considerthe network dynamics from the viewpoint of nonlinear system the-ory. However, how to appropriately represent real gene regulatorysystems mathematically in terms of gene function, expressionmechanisms, and signal-transduction pathways remains unclear.Mathematical models are useful for discovering higher order struc-ture of an organism and for gaining deep insights into both staticand dynamic behaviors of gene networks by extracting functionalinformation from observation data [9].
With the rapid advances in theoretical study and biologicalexperiments concerning the underlying regulatory mechanisms,more sophisticated theoretical models and general techniquesare increasingly demanded to elucidate periodic behaviors, with
ll rights reserved.
02043 and Key Disciplines of
ng), [email protected] (Z.
the consideration of time delays that are particularly importantfor GNs due to long time durations of transcription, translation, dif-fusion, and active transport process. Until now, most theoreticalworks on the study of coupling of gene oscillators [5–8]. Therehave been some studies devoted to the stability and oscillationsof GNs with fixed time-delay [9–11]. It is shown that oscillationscan be induced by delay in both nonstochastic and stochasticGNs. If we considered the process of macromolecular transport,the distributed delay is biologically plausible owing to delay timeis drastic. Generally a distributed time-delay can be representedby an integral of a function of one or more variables over a speci-fied range of previous time. In this paper, one of main targets is toanalyze the global stability of GNs with the considerations of thedistributed delays. The proposed approach is general and can mod-el any macromolecular transport process. For example, if move-ment of mRNA from a transcription site to translation sites is anactive process with a significant range of transportation times forindividual molecules, a distributed delay is a proper modelingframework [12]. In addition, all cellular components exhibit intra-cellular noises owing to random births and deaths of individualmolecules, and extracellular noises owing to environment fluctua-tions, which also contribute to distributed delay.
On the other hand, periodic perturbations are widespread in theexternal environment(e.g. daily light-dark cycle and Moon’s gravi-tational) and internal circumstance (e.g. cell division cycle or cellu-lar motility). Up to now, many theoretical models have beensuccessfully developed to understand rhythmic generators orinherent oscillations, but few studies consider the biological

Y. Wang et al. / Mathematical Biosciences 220 (2009) 34–44 35
oscillation driving by small perturbations (or synchronization),which are ubiquitous in the living cells, e.g. many cells or organsin mammals exhibit periodic oscillations with the same phase asrhythmic generators. Besides regular perturbations, there alwaysexist many irregular oscillations in the GNs, such as the stochasticregulation, production and decay processes which are due to tem-porary chemical or physical changes in the environment. Gene reg-ulation is an intrinsically noisy process, which is subjectintracellular and extracellular noise perturbations and environ-ment fluctuations [13–18]. Such perturbations and environmentfluctuations will undoubtedly affect the dynamics of networksboth quantitatively and qualitatively. Unfortunately, previousworks mainly focused on the perturbations of the linear systems,and there is little result on the perturbations of nonlinear biologi-cal systems due to the mathematical difficulties.
Motivated by the above discussions, rather than rhythmic gen-erators, the purpose of this paper are to study the periodic oscilla-tion of GNs with small periodic perturbation and time-delays. Inparticular, in this paper we derive new criteria for checking theglobal stability of periodic oscillation of GNs with SUM regulatorylogic by using the continuation theorem of Mawhin’s coincidencedegree theory and Lyapunov functional. The proposed approachis general and can be applied to analyze many biological oscilla-tions in an accurate manner. In other words, the theoretical resultsare able to cover a large range of nonlinear GNs even under uncer-tain environments. The paper is organized as follows. In Section 2,a framework of the general gene networks is given. In Section 3, wederive some results and new criteria about stability of periodicoscillations with fixed time-delay. Section 4 provides an exampleto illustrate the application of these criteria. Several summary re-marks are given in Section 5. In appendix, we derive the theoreticalresults for a general GN with perturbation and distributed delay.
2. Model of gene network
The activity of a gene is regulated by other genes through theconcentrations of their gene products, i.e. the transcription factors.Regulation can be quantified by the response characteristics, i.e.the level of gene expression as a function of the concentrationsof transcription factors. In this paper, based on the structure ofthe gene network (GN) or the genetic regulatory network pre-sented in [9], we consider a differential equation model describedas follows [3,9]:
_miðtÞ ¼ �aimiðtÞ þ biðp1ðtÞ; . . . ;pnðtÞÞ;_piðtÞ ¼ �cipiðtÞ þ dimiðtÞ; i ¼ 1; . . . ;n:
ð2:1Þ
where miðtÞ;piðtÞ 2 R are the concentrations of mRNA and protein ofthe ith node, respectively. In (2.1), ai and ci are the degradation ratesof the mRNA and protein, di is synthesize rate of the protein, andbiðtÞ is the regulatory function of the ith gene, which is a nonlinearfunction of the variables ðp1ðtÞ; . . . ; pnðtÞÞ but generally has a form ofmonotonicity with each variable [1,4]. In this network, there is oneoutput but multiple inputs for a single node or gene. A directed edgeis linked from node j to i if the transcriptional factor or protein j reg-ulates gene i.
Generally, the form of (2.1) may be very complicated, depend-ing on all biochemical reactions involved in this regulation. Typicalregulatory logics include AND-like gates and OR-like gates [19,20]for bi. In this paper, we focus on a model of gene networks whereeach transcription factor acts additively to regulate a gene. That is,the regulatory function is of the form biðp1ðtÞ; :::; pnðtÞÞ ¼Rn
j¼1bijðpjðtÞÞ, which is also called SUM logic [9,21], i.e. the regula-tory function sums over all the inputs. Such a SUM logic does existin many natural genetic networks [19]. In synthetic gene networks,one of the simplest ways to implement such an additive input
function is to provide a gene with multiple promoters, eachresponding to one of the inputs [10,21]. Such a regulation by multi-ple promoters is indeed found in many gene systems.
The function bijðpjðtÞÞ is generally expressed by a monotonicfunction of the Hill form
bijðpjðtÞÞ ¼
eijpjðtÞ=kð ÞH
1þ pjðtÞ=kð ÞH;
if transcription factor j is an activator of gene i;
eij1
1þ pjðtÞ=kð ÞH;
if transcription factor j is a repressor of gene i:
8>>>>>>>>><>>>>>>>>>:
ð2:2Þ
where H is the Hill coefficient, k is a positive constant, and eij P 0 isthe dimensionless transcriptional rate of transcription factor j togene i, which is a bounded constant.
In the following paper, the active transports which can be mod-eled with a time delay are introduced into system (2.1) to repre-sent biological processes. The time delays can be assumed to bedistributed delays which means that each macromolecule takes adifferent time to translocate from its place of synthesis to the loca-tion. Here, the derivative of a variable, which can be the concentra-tion of a macromolecule, depends on an integral of a function ofone or more variables over a specified range of previous time.For example, a general distributed delay for one variable takesthe form
dxðtÞdt¼ F x;
Z x
0f ðsÞxðt � sÞds
� �; with
Z x
0f ðsÞds ¼ 1:
The equation expresses a normalization condition imposed for bio-logical realism [12,22].
Hence, (2.1) can be rewritten into the following equations withthe distributed time-delay:
_miðtÞ ¼ �aimiðtÞ þPnj¼1
eijhijR tij
0 fijðsÞpjðt � sÞds� �
þ aiðtÞ;
_piðtÞ ¼ �cipiðtÞ þ diR sji
0 giðsÞmiðt � sÞdsþ biðtÞ;
ð2:3Þ
where aiðtÞ ¼ aiðt þ TÞ and biðtÞ ¼ biðt þ TÞði ¼ 1; . . . ; nÞ are smallperiodic perturbation which is considered as coupled periodic fluc-tuations from external environment or other rhythmic generators.Such a process include mechanisms may be relate to the cell divi-sion cycle or cellular motility [23].
For the sake of simplicity, we first consider the following equa-tions with the fixed time-delay which assumes that each macro-molecule takes the same length of time to translocate from itsplace of synthesis to the location and the general model with dis-tributed delay will be analyzed in the Appendix. That is
_miðtÞ ¼ �aimiðtÞ þPnj¼1
eijhij pjðt � sjÞ� �
þ aiðtÞ;
_piðtÞ ¼ �cipiðtÞ þ dimiðt � riÞ þ biðtÞ;ð2:4Þ
where aiðtÞ ¼ aiðt þ TÞ and biðtÞ ¼ biðt þ TÞði ¼ 1; . . . ;nÞ.Throughout this paper, we always make the following
assumption:A1. There exists Mij > 0 such that j hijðuÞ � hijðvÞ j6 Mijju� v j
for each u;v 2 Rði; j ¼ 1; . . . ;nÞ.Assumption A1 is generally satisfied in GNs due to the satura-
tion effects of transcription and translation processes.The following assumption is reasonable for the system (2.3)
which is sure sfijðsÞ and sgiðsÞ are integrable:A2. The delay kernels fijðsÞ; giðsÞ : ½0;þ1Þ ! ½0;þ1Þ
ði:j ¼ 1;2; . . . ; nÞ are continuous and integrable, and satisfy

36 Y. Wang et al. / Mathematical Biosciences 220 (2009) 34–44
Rþ10 fijðsÞds ¼ 1;
Rþ10 sfijðsÞds < þ1;Rþ1
0 giðsÞds ¼ 1;Rþ1
0 sgiðsÞds < þ1:
In order to simplify the proofs and compare our results, we give fol-lowing lemma and definition:
Assume that Tn�n ¼ A ¼ ðaijÞn�n : aij 6 0; i–j�
.
Lemma 1. ([24]): Let A 2Tn�n. Then, each of the following condi-tions is equivalent to the statement ‘ A is a non-singular M-matrix’:
(1) All of the principal minors of A are positive.(2) A has all positive diagonal elements and there exists a posi-
tive diagonal matrix D such that AD is strictly diagonallydominant, that is:
aiidi >Xi–j
jaijjdi; i ¼ 1;2; . . . ;n:
(3) A is inverse-positive, that is, A�1 exists and A�1 P 0.
The above Lemma 1 is widely used in the theoretical analysis of opti-mization design, finance mathematics, and neural networks. In thispaper, we adopt Lemma 1 to estimate the norm of miðtÞ and piðtÞ inthe main results, i.e. Theorem 1 of next section. By similar argumentin the [25], we have following lemma:
Lemma 2. Assume that aiðtÞ ¼ biðtÞ ¼ 0; ðA1Þ and ðA2Þ hold. Then,the system (2.3) stability, if
C ¼In C12
C21 In
� �
is a non-singular M-matrix, where In is a unit matrix andC12 ¼ðuijÞn�n;uij¼�eijMij=ai;C21 =diag f�d1=c1;�d2=c2; . . . ;�dn=cnÞ.For details, see in the [25].
Definition. ([26,27]): The T-periodic solution of GN (2.3) is said tobe globally asymptotically stable if it is stable in the sense ofLyapunov and global attractive, where global attractivity meansthat every trajectory tends to x�ðtÞ ¼ ðm�1ðtÞ; . . . ;m�nðtÞ; p�1ðtÞ; . . . ;
p�nðtÞÞ> of GN (2.3) as t ! þ1.
3. Some results of gene networks with fixed delay
In this section, we derive the main results which ensure theexistence and stability of periodic oscillators in GNs with perturba-tions and delay, where perturbation aiðtÞ and biðtÞ are assumed tobe small and periodic. Letting aþi ¼ suptPojaiðtÞj and bþi ¼suptPojbiðtÞj, we have the following sufficient conditions on theexistence of periodic solutions:
Theorem 1. Assume that aþi and bþi are bounded number, ðA1Þ andðA2Þ hold. Then, the system (2.4) has at least one T-periodic solutioncorresponding to the same periodic of perturbation, if
C ¼In C12
C21 In
� �is a non-singular M-matrix, where In is a unit matrix andC12 ¼ðuijÞn�n;uij¼�eijMij=ai; C21=diag f�d1=c1;�d2=c2; . . . ;�dn=cnÞ.
The detail proof of this result is given in Appendix A. To ensurenon-singular M-matrix, all of the elements C12 and C21 should berelatively small, comparing with the diagonal elements. In otherwords, ai and ci are required to be relatively large, comparing withthe synthetic rates. With those conditions, (2.4) is ensured to haveone T-periodic solution.
In following theorem, we show that under certain conditionssystem (2.4) has at least one T-periodic solution which is globallyattractive. It means that all the trajectories of (2.4) eventuallytends to the unique periodic solution.
Theorem 2. Assume that all conditions of Theorem 1 hold. Thesystem (2.4) has a unique T- periodic solution, which is globallyattractive, if
O ¼O11 O12
O21 O22
� �
is a non-singular M-matrix, where O11 ¼ diag ð2a1 �Pn
j¼1e1jM1j;
2a2 �Pn
j¼1e2jM2j; . . . ;2an �Pn
j¼1enjMnjÞ;O12 ¼ ðv ijÞn�n; v ij ¼ �eijMij;
O21 ¼ diag ð�d1;�d2; . . . ;�dnÞ;O22 ¼ diag ð2c1 � d1;2c2 � d2; . . . ;
2cn � dnÞ.
The proof is given in Appendix B. The conditions for Theorem 2are similar to those of Theorem 1, i.e. the degradation rates are re-quired to be larger than those of the synthesis rates so that O is anon-singular M-matrix. Next, we further give the conditions forthe asymptotical stability of the T-periodic solution.
Theorem 3. Assume that all conditions of Theorem 1 hold. Thesystem (2.4) has a unique T- periodic solution, which is globally andasymptotically stable if O 2T and is a weakly column diagonallydominant matrices.
The detail proof is given in Appendix C. The conditions of The-orem 3 also require large degradation rates, comparing with thesynthesis rates. As indicated by the theoretical results, a networkwith perturbation (even with sufficiently small perturbation) hasglobally stable oscillation under certain conditions. It means thatall the trajectories of (2.4) will eventually converge to the uniqueperiodic solution. It’s worth noting that, even if the parametersand regulatory functions are uncertain, we may design a robustGN, which can be driven by any periodic signals and thereby is ableto be synchronized with the driven forces even under uncertainenvironment, as long as the parameter intervals are finite.
4. Numerical example
In this section, we present an example of a gene network toshow the effectiveness and correctness of our theoretical results.
We consider the dynamics of the repressilator, which has beentheoretically predicted and experimentally investigated in Esche-richia coli [3]. The repressilator is a cyclic negative-feedback loopcomprising three repressor genes (lacl; tetR and cl) and their pro-moters. The kinetics of the system are determined by six coupledfirst-order differential equations
_miðtÞ ¼ �aimiðtÞ þ e1þpjðt�siÞÞ2
þ aiðtÞ;
_pjðtÞ ¼ �cjpjðtÞ þ djmjðt � rjÞ þ bjðtÞ;i ¼ lacl; tetR; cl; j ¼ cl; lacl; tetR;
ð4:1Þ
where mi and pi are the concentrations of the three mRNAs andrepressor-proteins, and e denotes the ratio of the protein decay rateto the mRNA decay rate. We select a set of biologically plausibleparameters as e ¼ 1:5; ai ¼ ci ¼ 1; di ¼ 0:9; si ¼ 0:2;rj ¼ 0:8. Conve-niently, we give two simple periodical perturbation asaiðtÞ ¼ 0:02 cos t;biðtÞ ¼ 0:02 sin t with T ¼ 2p, which are assumedto be weakly coupled from external environment or other rhythmicgenerators.
By appropriate computation, we have max h0 ¼ 3ffiffi3p
8 < 21=32,
C ¼
1 0 0 0 0 � 6364
0 1 0 � 6364 0 0
0 0 1 0 � 6364 0
� 910 0 0 1 0 0
0 � 910 0 0 1 0
0 0 � 910 0 0 1
0BBBBBBBBB@
1CCCCCCCCCA
and

0 2 4 6 8 10 12 14 16 18 200.5
0.6
0.7
0.8
0.9
1
1.1
Time( π) →
___ mRNA concentration...... protein concentration
Fig. 1. Time evolution of three mRNA and protein concentrations of GN (4.1)
Y. Wang et al. / Mathematical Biosciences 220 (2009) 34–44 37
O ¼
6564 0 0 0 0 � 63
64
0 6564 0 � 63
64 0 00 0 65
64 0 � 6364 0
� 910 0 0 11
10 0 00 � 9
10 0 0 1110 0
0 0 � 910 0 0 11
10
0BBBBBBBBB@
1CCCCCCCCCA:
It is easy to check that C and O are non-singular M-matrices. FromTheorem 3, the repressilator with this set of parameters has a T-Periodic oscillation, which is globally and asymptotically stable. InFig. 1 we plot the trajectories of mRNAs and proteins concentra-tions, which confirm the theoretical prediction. By Lemma 2, wefind that GN have only stable stationary solution absent small per-turbation(i.e., aiðtÞ ¼ biðtÞ ¼ 0), see Fig. 2. It is sufficiently demon-strate that periodic environment perturbation can induce geneoscillation under certain conditions.
5. Insight
Generally speaking, a cellular system is not only affected by var-ious external fluctuations but also characterized with significanttime delays in gene regulation, in particular, for the transcription,
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 300.4
0.5
0.6
0.7
0.8
0.9
1
1.1
Time(π) →
___ mRNA concertration...... protein concertration
Fig. 2. Time evolution of three mRNA and protein concentrations of GN (4.1)without perturbation.
translation, diffusion, and active transport. Such time delays mayaffect the dynamics of the entire biological system both qualita-tively and quantitatively. In this paper, we derived new criteriafor checking the global stability of periodic oscillation of GNs withdelays and periodic perturbations by using the continuation theo-rem of Mawhin’s coincidence degree theory, the non-singular M-matrix and Lyapunov function. Theoretical results ensure that theGN has a stable periodic oscillation provided that there is a smallperiodic driving force, which can be used to analyze cellular oscil-lations, to understand synchronization phenomena, and even todesign a robust synthetic oscillator.
5.1. Designing a large scale GN
As indicated in this paper, the networks with perturbation havestable oscillation under certain conditions which relay on theLyapunov constants of the regulatory functions, dimensionlesstranscriptional rate, and the degradation rates of mRNAs or pro-teins. When analyzing or designing a large scale GN, we need thefollowing steps based on Theorems 1–3.
(1) Firstly, we select a set of biologically plausible parametersand compute the Lipschitz constants Mij of the function bi.
(2) Check whether C and O are non-singular M-matrixes.
If all of the conditions are satisfied, there exists a stable periodicoscillation, which coincides with the weak periodic signal in phase.Owing to the fact that our results are robust for genetic oscillatornetworks with perturbation and distributed delay, we can designa robust oscillation in the GN within the defined parameter inter-vals or ranges.
5.2. Small perturbation can also induce oscillation
Since genetic regulatory networks are high-dimensional andnonlinear, it is indispensable to consider the network dynamicsfrom the viewpoint of nonlinear systems theory. Up to now, severaltheoretical models have been successfully developed to understandcircadian phenomena of GNs [28,36], but few studies consider thesmall perturbations. On the other hand, these perturbations alwaysexist in the GNs, such as the stochastic regulation, production anddecay processes which are due to temporary chemical or physicalchanges in the environment, or periodic perturbations from othercells. Unfortunately, previous results are mainly for the perturba-tions on linear systems, and there are few results on the perturba-tions of nonlinear systems due to the mathematical difficulties.
In [23], a model for a synthetic gene oscillator is present, andthe coupling of the small oscillator to a periodic process that isintrinsic to the cell is considered. They studied the synchronizationproperties of the coupled system, and showed how the oscillatorcan be constructed to yield a significant amplification of cellularoscillations both numerically and experimentally.
In our paper, we consider a general nonlinear GN model andshow that under certain conditions, the nonlinear system coupledwith even small perturbation can always oscillate with the signals.
5.3. GNs with distributed delay
Transportation of biomolecules can be modeled as a diffusive oractive process in nature. In the framework of differential equations,such processes can be modeled as a time delay. The time delay isusually assumed to be discrete for the sake of simplification, i.e.each macromolecule is assumed to take the same length of timeto translocate from one place to another where it exerts an effect,such as (2.4). Clearly, such assumption is not biologically plausible,in particular, when there are a small number of the molecules in

38 Y. Wang et al. / Mathematical Biosciences 220 (2009) 34–44
the system. In this paper, we adopt a distributed delay to repre-sent such processes, i.e. (2.3) so as to analyze the GN in a moreaccurate manner. In principle, this approach can model any mech-anism of macromolecular transport. For example, if movement ofmRNA from a transcription site to translation sites is an activeprocess with a significant range of transport times for individualmolecules, a distributed delay would be the proper modelingframework.
In a single cell, many species only have a small number of mol-ecules, which means that averaging effect on delay (e.g. fixed de-lay) may not be adequate. There are many evidences for activetransport of specific mRNAs and proteins in eukaryotic cytoplasm.Examples include microtubule-dependent movement of Vg1mRNA in Xenopus oocytes [29], microtubule-dependent localiza-tion of several maternal mRNAs for proteins that direct embryonicdevelopment (such as bicoid, bicuadal-D, and oskar) in Drosophilaoocytes [30], movement of tubulin monomers from the cell body tothe ends of growing axons, in a variety of vertebrate neuronaltypes, at rates well in excess of those that could be explained bydiffusion [31]; and, more generally, axonal and dendritic transportof various types. In reality, active macromolecular transport in pro-karyotic cells can also be important. For example, it has been ob-served that specific intracellular localization of proteinsdetermine the relative fates of daughter cells after cell division[32]. Above examples are sufficient to convince that the activetransport play important roles in dynamics of the biological sys-tems and may affect the dynamics of the entire biological system,both qualitatively and quantitatively.
In this paper, we considered the active macromolecule trans-port in nature, and show that the ability to couple naturally occur-ring genetic oscillations to a synthetically designed network maylead to possible strategies for entraining oscillations in cellularmRNA or protein levels.
Appendix A. Existence oscillation for the GN with thedistributed delay
In this Appendix, we will give some sufficient conditions ofexistence of periodic oscillation for the GN (2.3):
_miðtÞ ¼ �aimiðtÞ þXn
j¼1
eijhij
Z tij
0fijðsÞpjðt � sÞds
� �þ aiðtÞ;
_piðtÞ ¼ �cipiðtÞ þ di
Z sji
0giðsÞmiðt � sÞdsþ biðtÞ;
where aiðtÞ ¼ aiðt þ TÞ and biðtÞ ¼ biðt þ TÞði ¼ 1;2; . . . ;nÞ.Let the initial conditions associated with system (2.3) have the
form: miðsÞ ¼ /iðsÞ; piðsÞ ¼ wiðsÞ; s 2 ð�1; 0�; where /iðsÞ;wiðsÞare continuous functions (i ¼ 1;2; . . . n).
For any solution
xðtÞ ¼ ðm1ðtÞ;m2ðtÞ; . . . ;mnðtÞ;p1ðtÞ;p2ðtÞ; . . . ;pnðtÞÞ>
and T-periodic solution
xðtÞ� ¼ ðm�1ðtÞ;m�2ðtÞ; . . . ;m�nðtÞ; p�1ðtÞ; p�2ðtÞ; . . . ;p�nðtÞÞ>
of system (2.3), we define kxðtÞ � xðtÞ�k1 as
kðxðtÞ � x�ðtÞk1 ¼Xn
i¼1
jmiðtÞ �m�i ðtÞj þXn
j¼1
jpjðtÞ � p�j ðtÞj:
For convenience and completeness, we introduce some useful nota-tions, definitions and lemmas in this section. Let kuk is Lebesguenorm of u with kuk ¼ ð
R T0 juðsÞj
2dsÞ1=2 and �v is mean of v from 0 toT: �v ¼ 1
T
R T0 vðsÞds, where uðsÞ 2 CðR;RÞ and vðsÞ is a T-periodic
function.
Lemma 3. ([33]). Assume that A is a non-singular M-matrix andAw 6 d, then w 6 A�1d.
In the following, we introduce some concepts and results from[34] by Gaines and Mawhin.
Let X and Z be two Banach spaces, L : DomL � X ! Z be a linearmapping, and N : X ! Z be a continuous mapping. The mapping Lwill be called a Fredholm mapping of index zero if dim Ker L=codimIm L < þ1 and Im L is closed in Z. If L is a Fredholm mapping ofindex zero and there exist continuous projectors P : X ! X andQ : Z ! Z such that Im P=Ker L, Ker Q=Im L=Im ðI � Q), it followsthat mapping LjDomL
TKerP : ðI � PÞX ! ImL is invertible. We denote
the inverse of that mapping by KP . If X is an open bounded subsetof X, the mapping N will be called L-compact on if QNðXÞ isbounded and KPðI � QÞN : X! X is compact. Since Im Q is isomor-phic to Ker L, there exists an isomorphism J : Im Q ! Ker L.
Lemma 4. (Mawhin’s continuation theorem). Let X and Z be twoBanach spaces and L be a Fredholm mapping of index zero. Assumethat X � X is an open bounded set and N : X ! Z is a continuousoperator which is L-compact on X. Then Lx ¼ Nx has at least onesolution in Dom L
TX, if the following conditions are satisfied:
(1) For each k 2 ð0;1Þ; x 2 @XT
DomL; Lx – kNx;(2) For each x 2 @X
TKerL;QNx ¼ 0;
(3) degðJQNx;XT
KerL;0Þ– 0,
where J : ImQ ! KerL is an isomorphism.
Lemma 5. ([35]) Let f ð�Þ : ½0;þ1Þ ! Rþ be a continuous function. Iff ð�Þ is integrable and uniformly continuous on ½0;þ1Þ, thenlimt!þ1f ðtÞ ¼ 0.
Theorem 4. Assume that aþ; bþ are bounded number, ðA1Þ and ðA2Þhold. Then, the system (2.3) has at least one T-periodic solution corre-sponding to the same periodic of enteral perturbation, if
C ¼In C12
C21 In
� �
is a non-singular M-matrix, where In is a unit matrix andC12¼ðuijÞn�n;uij ¼�eijMij=ai;C21 =diag f�d1=c1;�d2=c2; . . . ;�dn=cnÞ.
Proof. Let X and Z is set of T-periodic solution:X ¼ Z ¼ fxðtÞ ¼ ðm1ðtÞ; . . . ;mnðtÞ;p1ðtÞ; . . . ;pnðtÞÞ
>jmiðtÞ ¼ miðt þ TÞ;piðtÞ ¼ piðt þ TÞ; i ¼ 1;2; . . . ;ng.Then X is a Banach space with the norm k � k1.
Let L : DomL � X ! Z; P : XT
DomL! KerL;Q : X ! X=Im L andN : X ! Z be given by the following:
Lx ¼ ð _m1ðtÞ; _m2ðtÞ; . . . ; _mnðtÞ; _p1ðtÞ; _p2ðtÞ; . . . ; _pnðtÞÞ>;
Px ¼ Qx ¼ �m1ðtÞ; �m2ðtÞ; . . . ; �mnðtÞ; �p1ðtÞ; �p2ðtÞ; . . . ; �pnðtÞð Þ>;
Nx ¼
�a1m1ðtÞ þPnj¼1
e1jh1jR t1j
0 f1jðsÞpjðt � sÞds� �
þ a1ðtÞ
� � �
�anmnðtÞ þPnj¼1
enjhnjR tnj
0 fnjðsÞpjðt � sÞds� �
þ anðtÞ
�c1p1ðtÞ þ d1R s1i
0 g1ðsÞm1ðt � sÞdsþ b1ðtÞ
� � ��cnpnðtÞ þ dn
R sni0 gnðsÞmnðt � sÞdsþ bnðtÞ
0BBBBBBBBBBBBBBB@
1CCCCCCCCCCCCCCCA
:
It is easy to see that L is a linear operator with KerL ¼ xðtÞjxðtÞ ¼ xð0Þ 2 R2n
n owhich mean Ker L is set of constant
mapping. Im L ¼ xðtÞjxðtÞ 2 Z;R T
0 xðtÞdt ¼ 0n o
is closed in Z and

Y. Wang et al. / Mathematical Biosciences 220 (2009) 34–44 39
dim Ker L=codim Im L ¼ 2n. Therefore, L is a Fredholm mapping ofindex zero. It is easy to prove that P and Q are two projectors, andIm P=ker L, Im L=ker Q=Im ðI � QÞ. By using the Arzela-Ascoli theo-rem, we know that for every bounded subset X 2 X;KpðI � QÞN arerelatively compact on X in X, i.e. N is L-compact on X.
Consider the operator equation
Lx ¼ kNx; k 2 ð0;1Þ;
that is
_miðtÞ ¼ �kaimiðtÞ þ kPnj¼1
eijhijR tij
0 fijðsÞpjðt � sÞds� �
þ kaiðtÞ;
_pkðtÞ ¼ �kckpkðtÞ þ kdkR sji
0 gkðsÞmkðt � sÞdsþ kbkðtÞ;ðA:1Þ
where i; k ¼ 1;2; . . . ;n:First, we prove that the solution of Eq. (2.3) is bounded. Denote
li ¼ kðRnj¼1sups2RhijðsÞ þ aþi Þ. By using (A.1), we obtain
�li 6_miðtÞ þ kaimiðtÞ 6 li: ðA:2Þ
Multiplying both sides of (A.2) by ekaitand integrating the inequalityabove from 0 to uðu P 0Þ, we obtain
�li
Z u
0ekaitdt 6 miðuÞekaiu �mið0Þ 6 li
Z u
0ekaitdt:
Hence
miðuÞP �li
Z u
0e�kaiðu�tÞdt þmið0Þe�kaiu
and
miðuÞ 6 li
Z u
0e�kaiðu�tÞdt þmið0Þe�kaiu;
for u P t P 0. So we have
jmiðuÞj 6 mið0Þ �li
kai
��������þ li
kai:
By similar argument, we obtain piðtÞ is also bounded.Next, we shall prove the conditions of Lemma 4 is satisfied. By
the A2, we know thatZ tij
0fijðsÞpiðt � sÞds
is uniformly convergent. Therefore, the following iterated integralZ T
0miðtÞ
Z tij
0fijðsÞpjðt � sÞdsdt
can be changed integrating order.Suppose that ðm1ðtÞ;m2ðtÞ; . . . ;mnðtÞ; p1ðtÞ; p2ðtÞ; . . . ; pnðtÞÞ
> 2 Xis any periodic solution of system (2.3) for a certain k 2 ð0;1Þ. Mul-tiplying both sides of
_miðtÞ ¼ �kaimiðtÞ þ kXn
j¼1
ei;jhij
Z tij
0fijðsÞpjðt � sÞds
� �þ kaiðtÞ
ðA:3Þ
by miðtÞ and integrating from 0 to T, we obtain
0 ¼ �Z T
0kaim2
i ðtÞdt þZ T
0kXn
j¼1
eijmiðtÞhij
Z tij
0fijðsÞpjðt � sÞds
� �dt
þZ T
0kaiðtÞmiðtÞdt:
From ðA1Þ; ðA2) and noting that ðR T
0 jpjðt � sÞj2dtÞ1=2 ¼R T0 jpjðtÞj
2dt� �1=2
¼ kpjk, we have
ai
Z T
0m2
i ðtÞdt ¼Z T
0
Xn
j¼1
eijmiðtÞhij
Z tij
0fijðsÞpjðt � sÞds
� �dt
þZ T
0miðtÞaiðtÞdt
¼Z T
0
Xn
j¼1
eijmiðtÞ hij
Z tij
0fijðsÞpjðt � sÞds
� �� hijð0Þ
� dt
þZ T
0
Xn
j¼1
miðtÞeijhijð0Þdt þZ T
0miðtÞaiðtÞdt
6
Xn
j¼1
eijMijkpjkkmik þffiffiffiTp Xn
j¼1
eijhijð0Þ þ aþi
!kmik;
ðA:4Þthat is
kmik 61ai
Xn
j¼1
eijMijkpjk þffiffiffiTp
ai
Xn
j¼1
eijhijð0Þ þ aþi
!: ðA:5Þ
By similar argument, we obtain
kpik 6di
cikmik þ
ffiffiffiTp
cibþi : ðA:6Þ
It follows (A.5) and (A.6) that
Cy 6 s; ðA:7Þ
where C ¼ In C12
C21 In
� �; In is a unit matrix and C12¼
ðuijÞn�n;uij ¼�eijMij=ai;C21 =diag f�d1=c1;�d2=c2; . . . ;�dn=cnÞ;y ¼ ðkm1k; km2k; . . . ; kmnk; kp1k; kp2k; . . . ; kpnkÞ
>; s ¼ ðs1; s2; . . . ; s2nÞ>,
si ¼ffiffiffiTp
Rnj¼1eijhijð0Þ þ aþi
� �=ai,snþi ¼
ffiffiffiTp
bþi =ci;1 6 i 6 n. Applicationof Lemma 3 yields
y 6 C�1s , ðr1; r2; . . . ; r2nÞ>;
which implies that kmik and kpik are bounded: kmik 6 ri andkpik 6 rnþi; i ¼ 1; . . . ;n: It is not difficult to check that there existt�i ; t
�nþi 2 ½0; T� such that
miðt�i Þ 6 ri=ffiffiffiTp
; piðt�nþiÞ 6 rnþi=ffiffiffiTp
; i ¼ 1; . . . ; n:
BecauseRþ1
0 fijðsÞpiðt � sÞds is uniformly convergent, multiplyingboth sides of (A.3) by _miðtÞði ¼ 1;2; . . . ; nÞ and integrating from 0to T, we obtain
k _mik2 ¼ kRnj¼1
Z T
0eij _miðtÞhij
Z tij
0fijðsÞpjðt � sÞds
� �dt
� kZ T
0ai _miðtÞmiðtÞdt þ k
Z T
0aiðtÞ _miðtÞdt
6 Rnj¼1eijMij
Z T
0j _miðtÞj
Z þ1
0fijðsÞpjðt � sÞdsdt
þ Rnj¼1eijhijð0Þ
Z T
0j _miðtÞjdt þ
Z T
0aij _miðtÞmiðtÞjdt
þ kaþiZ T
0
_miðtÞdt
6 Rnj¼1eijMijk _mikkpjk þ
ffiffiffiTp
Rnj¼1eijhijð0Þk _mik
þffiffiffiTp
aþi k _mik þ aik _mikkmik: ðA:8ÞTherefore k _mik 6 Rn
j¼1eijMij kpjk þffiffiffiTpðRn
j¼1eijhijð0Þ þ aþi Þ þ aikmik:It is easy check that
miðtÞ ¼ miðt�Þ þZ T
t�_miðtÞdt 6 ri=
ffiffiffiTpþ
ffiffiffiTpk _mik
6 1=ffiffiffiTpþ ai
ffiffiffiTp� �
ri þffiffiffiTp
Rnj¼1eijMijrnþj
þ T Rnj¼1eijhijð0Þ þ aþi
� �, ri=
ffiffiffiTpþ fi
and

40 Y. Wang et al. / Mathematical Biosciences 220 (2009) 34–44
piðtÞ 6 rnþi=ffiffiffiTpþ
ffiffiffiTpk _pik 6 rnþi=
ffiffiffiTpþ
ffiffiffiTpðckpik þ dkmik þ bþi ÞÞ
6 rnþi=ffiffiffiTpþ
ffiffiffiTp
dri þffiffiffiTp
cirnþi þffiffiffiTp
bþi , rnþi=ffiffiffiTpþ fnþi:
Let ni ¼ ri=ffiffiffiTpþ fi þ 1 and nnþj ¼ rnþj=
ffiffiffiTpþ fnþj þ 1(0 < 1� 1Þ. We
take X ¼ fx ¼ ðm1ðtÞ; . . . ;mnðtÞ;p1ðtÞ; . . . ; pnðtÞÞ> 2 X; jmiðtÞj< ni;
jpnþiðtÞj < nnþig.Obviously, the condition (1) of Lemma 4 is satisfied. When
x 2 @XT
ker L=@XT
R2n; x is a constant vector in R2n withjmiðtÞj ¼ ni; jpiðtÞj ¼ nnþi. Then, we have miðQNxÞi ¼ �aim2
i þmiPn
j¼1eijhijðpiÞ þmi �ai, and
piðQNxÞnþi ¼ �cip2i þ piui þ pi
�bi:
We claim that there exists some i (1 6 i 6 n) such that
miðQNxÞi < 0 or piðQNxÞnþi < 0:
If miðQNxÞi P 0 and piðQNxÞnþi P 0, then, we obtain
aim2i 6 mi
Xn
j¼1
eijhijðpiÞ þmiaþi
¼ mi
Xn
j¼1
eij½hijðpjÞ � hijð0Þ� þmi
Xn
j¼1
eijhijð0Þ þmiIaþi
6 jmijXn
j¼1
eijMijjpjj þ jmijXn
j¼1
eijhijð0Þ þ jmijaþi :
Which implies
ni 61ai
Xn
j¼1
eijMijnnþi þ1ai
Xn
j¼1
eijhijð0Þ þ aþi
!
61ai
Xn
j¼1
eijMijnnþj þsiffiffiffiTp ðA:9Þ
By a similar argument, we obtain
nnþi 61ci
dini þ1ci
bþi 61ci
dini þsnþiffiffiffi
Tp ðA:10Þ
On the other hand,
n > r=ffiffiffiTpþ f > r=
ffiffiffiTp¼ P�1s=
ffiffiffiTp
where n ¼ ðn1; n2; . . . ; nnþmÞ, f ¼ ðf1; f2; . . . ; fnþmÞ and r ¼ ðr1;
r2; . . . ; rnþmÞ. It follows that there exists that some i or j such that
ni �1ai
Xn
j¼1
eijMijnnþj >siffiffiffiTp
or
nnþi �1ci
dini >snþiffiffiffi
Tp ;
which is contradiction with (A.9) and (A.10). Then, there exists somei (1 6 i 6 n) such that miðQNxÞi < 0 or piðQNxÞnþi < 0. Therefore,
kQNxk1 ¼Xn
i¼1
jðQNxÞij þXn
i¼1
jðQNxÞnþij > 0:
This indicates that the condition (2) of Lemma 4 is satisfied.Define Hðx; hÞ ¼ �hxþ ð1� hÞQNx; h 2 ½0;1�, where
x ¼ ðm1ðtÞ; . . . ;mnðtÞ; p1ðtÞ; . . . ; pnðtÞÞ> 2 R2n:
When x 2 Ker LT@X, we have
kHðx; hÞk1 ¼Xn
i¼1
jHðui; hÞj þXn
i¼1
jHðpi; hÞj
¼Xn
i¼1
j � hmi þ ð1� hÞðQNxÞij
þXn
n¼1
j � hpi þ ð1� hÞðQNxÞij > 0
that is Hðx; hÞ– 0: According to the invariant of homology, we have
degfJQNx;X\
Ker L;0 – 0;
where J : Im Q ! Ker L is an isomorphism. Therefore, accordingto the continuation theorem of Gaines and Mawhin, GN (2.3)has at least one T-periodical oscillation. The proof iscompleted.
Appendix B. Global attractivity of oscillation
Now, we will discuss the global attractivity of the periodic solu-tion of system (2.3). Under the assumptions of Theorem 4, system(2.3) has at least one T-periodic solution x�ðtÞ ¼ ðm�1ðtÞ;m�2ðtÞ;. . . ;m�nðtÞ; p�1ðtÞ; p�2ðtÞ; . . . ; p�nðtÞÞ
>.
Theorem 5. Assume that all conditions of Theorem 4 hold. Thesystem (2.3) has a unique T- periodic solution, which is globallyattractive, if
O ¼O11 O12
O21 O22
� �is a non-singular M-matrix, where O11 ¼ diag ð2a1�Pn
j¼1e1jM1j;2a2�Pn
j¼1e2jM2j; . . . ;2an�Pn
j¼1enjMnjÞ;O12 ¼ðv ijÞn�n;v ij¼�eijMij;O21 ¼ diag ð�d1;�d2; . . . ;�dnÞ;O22 ¼ diag ð2c1�d1;2c2�d2; . . . ;2cn�dnÞ.
Proof. Let zðtÞ ¼ ðm1ðtÞ �m�1ðtÞ;m2ðtÞ �m�2ðtÞ; . . . ;mnðtÞ �m�nðtÞ;p1ðtÞ � p�1ðtÞ; p2ðtÞ � p�2ðtÞ; . . . ; pnðtÞ � p�nðtÞÞ
>. Then,
ziðtÞ _ziðtÞ ¼ �aijziðtÞj2 �Xn
j¼1
eijziðtÞhij
�Z tij
0fijðsÞp�j ðt � sÞds
� �
þXn
j¼1
eijziðtÞhij
Z tij
0fijðsÞ½znþjðt � sÞ
�
þp�j ðt � sÞ�dso6 �aijziðtÞj2
þXn
j¼1
eijMij
Z þ1
0fijðsÞziðtÞznþjðt � sÞds 6 �aijziðtÞj2
þ 12
Xn
j¼1
eijMij jziðtÞj2 þZ þ1
0fijðsÞjznþjðt � sÞj2ds
� �;
znþiðtÞ _znþiðtÞ ¼ �cijznþiðtÞj2 þ di
Z sji
0giðsÞznþiðtÞziðt � sÞds
6 �cijznþiðtÞj2 þ12
di jznþiðtÞj2 þZ þ1
0giðsÞjziðt � sÞj2ds
� �;
for i ¼ 1;2; . . . ; n:
We define the following Lyapunov functionals:
ViðtÞ ¼ jziðtÞj2 þXn
j¼1
eijMij
Z þ1
0
Z t
t�sfijðsÞjznþjðuÞj2dsdu;
VnþiðtÞ ¼ jznþiðtÞj2 þ di
Z þ1
0
Z t
t�sgiðsÞjziðuÞj2dsdu:
Calculating the derivatives DViðtÞ and DVnþiðtÞ along the solution of(2.3) [27], and estimating it via the assumptions, we have
DViðtÞ ¼ 2ziðtÞ _ziðtÞþXn
j¼1
eijMij
Z þ1
0fijðsÞjznþjðtÞj2ds
�Xn
j¼1
eijMij
Z þ1
0fijðsÞjznþjðt� sÞj2ds
6�2aijziðtÞj2þXn
j¼1
eijMijjziðtÞj2þXn
j¼1
eijMijjznþjðtÞj2
6 �2aiþXn
j¼1
eijMij
!jziðtÞj2þ
Xn
j¼1
eijMijjznþjðtÞj2;

Y. Wang et al. / Mathematical Biosciences 220 (2009) 34–44 41
DVnþiðtÞ ¼ 2znþiðtÞ _znþiðtÞþdi
Z þ1
0giðsÞjziðtÞj2ds
�di
Z þ1
0giðsÞjziðt� sÞj2ds6 dijziðtÞj2þð�2ciþdiÞjznþiðtÞj2:
Let VðtÞ ¼ ðV1ðtÞ;V2ðtÞ; . . . ;V2nðtÞÞ>, by above inequalities, weobtain
V 0ðtÞ 6 �OfðtÞ ðB:1Þ
where O ¼ O11 O12
O21 O22
� �;O11 ¼ diag ð2a1 �
Pnj¼1e1jM1j;2a2�Pn
j¼1e2jM2j;. ..;2an�Pn
j¼1enjMnjÞ;O12¼ðv ijÞn�n;v ij¼�eijMij;O21¼ diagð�d1;�d2;.. .;�dnÞ;O22¼ diag ð2c1�d1;2c2�d2;.. .;2cn�dnÞ;fðtÞ¼ðjz1ðtÞj2;jz2ðtÞj2; ... ;jz2nðtÞj2Þ>;VðtÞ¼ðV1ðtÞ;V2ðtÞ;.. .;VnðtÞÞT . O
is a non-singular M-matrix, and by Lemma 3, we have
O�1V 0ðtÞ 6 �fðtÞ: ðB:2Þ
Then, by definition of the vector
~VðtÞ ¼ ð~V1ðtÞ; ~V2ðtÞ; . . . ; ~V2nðtÞÞ>,O�1VðtÞP 0;
we have
~V 0ðtÞ ¼ ð~V 01ðtÞ; ~V 02ðtÞ; . . . ; ~V 02nðtÞÞ> ¼ O�1V 0ðtÞ;
that is
~V 0iðtÞ 6 �jziðtÞj2; ~V 0nþiðtÞ 6 �jznþiðtÞj2; ðB:3Þ
i ¼ 1;2; . . . ;n: Integrating both sides of (B.3) from 0 to t results in
~ViðtÞ þZ t
0jziðtÞj2 6 ~Við0Þ < þ1;
~VnþiðtÞ þZ t
0jznþiðtÞj2 6 ~Vnþið0Þ < þ1:
HenceR t
0 jziðsÞj2ds 6 ~Við0Þ andR t
0 jznþiðsÞj2ds 6 ~Vnþið0Þ, which implyjziðtÞj2 and jznþiðtÞj2 are integrable on ½0;þ1Þ for i ¼ 1;2; . . . ;n. Bythe definition of ViðtÞ and VnþiðtÞ, we obtain
jziðtÞj2 6 ViðtÞ and jznþiðtÞj2 6 VnþiðtÞ;
namely, fðtÞ 6 VðtÞ. Then, fðtÞ 6 O~VðtÞ, which implies thatO�fðtÞ 6 ~VðtÞ 6 ~Vð0Þ < þ1. Hence jziðtÞj2 < þ1 and jznþiðtÞj2 <þ1, which mean
jmiðtÞ �m�i ðtÞj26 þ1 and jpiðtÞ � p�i ðtÞj
26 þ1:
Therefore, jm�i ðtÞj and jp�i ðtÞj are bounded, which implies that jmiðtÞjand jpiðtÞj are bounded. By the boundedness of j _miðtÞj and j _piðtÞj, jfijand jfnþiðtÞjði ¼ 1;2; . . . ;nÞ are uniformly continuous on ½0;þ1Þ. ByLemma 5,
limt!þ1jmiðtÞ �m�i ðtÞj ¼ 0 and limt!þ1jpiðtÞ � p�i ðtÞj ¼ 0:
Thus, the proof is completed. h
Appendix C. Globally and asymptotically periodic solution
Now, we give following theorem to ensure a globally asymp-totic periodic solution:
Theorem 6. Assume that all conditions of Theorem 4 hold. Thesystem (2.3) has a unique T- periodic solution, which is globally andasymptotically stable if O 2T and is a weakly column diagonallydominant matrices.
Proof. Consider the Lyapunov function:
VðtÞ ¼X2n
k¼1
VkðtÞ
Calculating the derivative DVðtÞ of V along the solution of (2.3), wehave
DVðtÞ ¼ DX2n
k¼1
VkðtÞ
6
Xn
i¼1
�2ai þXn
j¼1
eijMij
!z2
i ðtÞ þXn
j¼1
eijMijjznþjðtÞj2( )
þXn
i¼1
dijziðtÞj2 þ ð�2ci þ diÞjznþiðtÞj2n o
¼Xn
i¼1
�2ai þXn
j¼1
eijMij þ di
( )jziðtÞj2
þXn
i¼1
f�2ci þXn
j¼1
eijMij þ digjznþiðtÞj2 < 0
This means that the periodic solution of system (2.3) is globally andasymptotically stable. The proof is completed.
Remark 1. For the system (2.3), when delay kernels fijðsÞ , giðsÞ ared-functions, i.e. we take fijðsÞ ¼ dðs� siÞ and giðsÞ ¼ dðs� riÞ, thensystem (2.3) can be reduced to a fixed time-delay system (2.4)and Theorems 1–3 are hold.
Example 2. Here, we consider the repressilator with distributeddelay. When tij ¼ sji ¼ þ1, we have following equations for therepressilator:
_miðtÞ ¼ �aimiðtÞ þ e
1þðR þ1
0f ðsÞpjðt�sÞdsÞ2
þ aiðtÞ;
_pjðtÞ ¼ �cjpjðtÞ þ djRþ1
0 gðsÞmiðt � sÞdsþ bjðtÞ;i ¼ lacl; tetR; cl; j ¼ cl; lacl; tetR;
ðC:1Þ
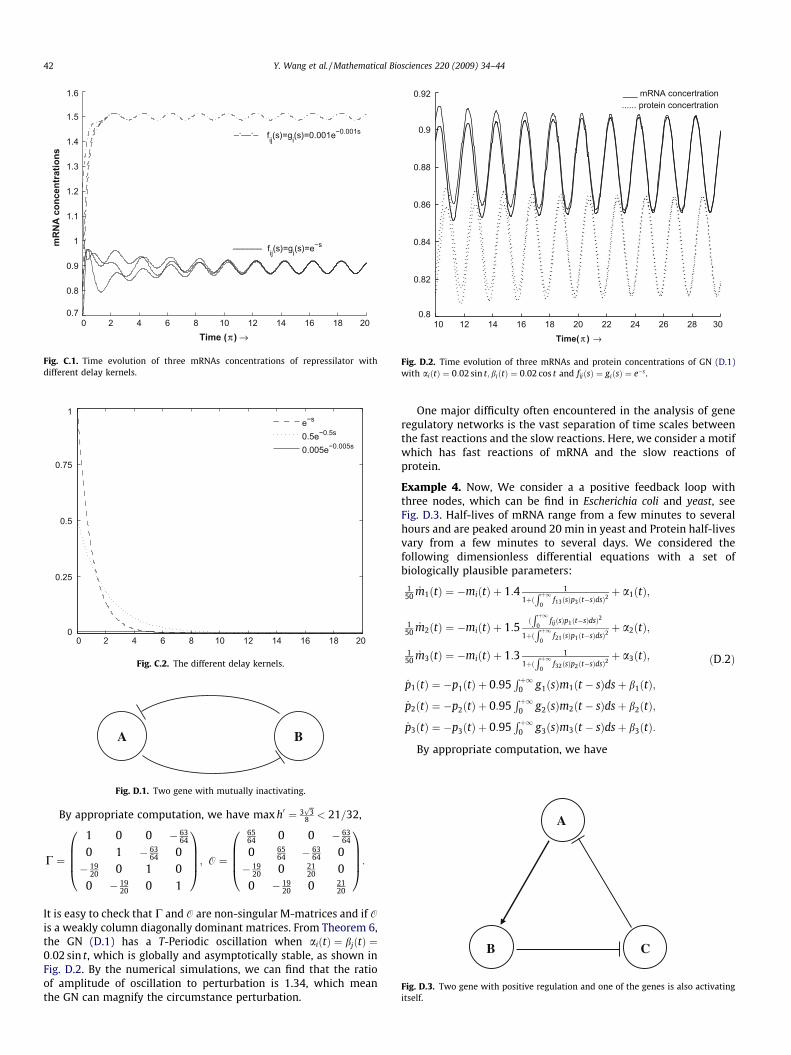
Conveniently, we select aiðtÞ ¼ 0:02 cos t, bjðtÞ ¼ 0:02þ 0:02sint andother parameters as same as in the Section 4. It is easy to show fromTheorem 6 that the repressilator has a T-Periodic oscillation, whichis globally and asymptotically stable. In Fig. C.1 , we plot the trajec-tories of mRNAs of the repressilator with different delay kernels(see Fig. C.2), and find that the average concentration of mRNAand protein are relate to the distributed delays.
Remark 2. A frequently used model for distributed time delays inbiological applications is to choose for the Gamma kernel due tomathematical difficulties. In this paper, we set a weak delay kernel,i.e. an exponential kernel.
Appendix D. Another examples
Complex cellular behaviors can be seen as a result of interac-tions of numerous intracellular or extracellular biomolecules. Tofigure out cellular behaviors, it is therefore important to investi-gate the topology of cellular circuits and corresponding dynamicalcharacteristics [38]. Here, we study dynamical of two motif whichcan be find in databases [37].
Example 3. We consider a motifs of only two genes produced themutually inactivating, such as genes Cdc28 and Sic1 in yeast, andGal3 and Gal80 in S. cerevisiae galactose regulation, see Fig. D.1.The following dimensionless differential equations are the kineticsof motif with small perturbation:
_miðtÞ ¼ �aimiðtÞ þP2j¼1
eijhijR tij
0 fijðsÞpjðt � sÞds� �
þ aiðtÞ;
_piðtÞ ¼ �cipiðtÞ þ diR sji
0 giðsÞmiðt � sÞdsþ biðtÞ;ðD:1Þ
where hijðxÞ ¼ 1=ð1þ x2Þ. We select a set of biologically plausibleparameters as ai ¼ ci ¼ 1;di ¼ 0:95; e11 ¼ 0; e12 ¼ 1:5; e21 ¼ 1:5;e22 ¼ 0; fijðsÞ ¼ giðsÞ ¼ e�s; tij ¼ sji ¼ þ1.
0 2 4 6 8 10 12 14 16 18 200.7
0.8
0.9
1
1.1
1.2
1.3
1.4
1.5
1.6
Time (π ) →
mR
NA
con
cent
ratio
ns
fij(s)=gi(s)=e−s
_.__._ fij(s)=gi(s)=0.001e−0.001s
______
Fig. C.1. Time evolution of three mRNAs concentrations of repressilator withdifferent delay kernels.
0 2 4 6 8 10 12 14 16 18 200
0.25
0.5
0.75
1e−s
0.5e−0.5s
0.005e−0.005s
Fig. C.2. The different delay kernels.
BA
Fig. D.1. Two gene with mutually inactivating.
10 12 14 16 18 20 22 24 26 28 300.8
0.82
0.84
0.86
0.88
0.9
0.92
Time( π ) →
___ mRNA concertration ...... protein concertration
Fig. D.2. Time evolution of three mRNAs and protein concentrations of GN (D.1)with aiðtÞ ¼ 0:02 sin t; biðtÞ ¼ 0:02 cos t and fijðsÞ ¼ giðsÞ ¼ e�s .
CB
A
Fig. D.3. Two gene with positive regulation and one of the genes is also activatingitself.
42 Y. Wang et al. / Mathematical Biosciences 220 (2009) 34–44
By appropriate computation, we have max h0 ¼ 3ffiffi3p
8 < 21=32,
C ¼
1 0 0 � 6364
0 1 � 6364 0
� 1920 0 1 0
0 � 1920 0 1
0BBB@
1CCCA; O ¼
6564 0 0 � 63
64
0 6564 � 63
64 0� 19
20 0 2120 0
0 � 1920 0 21
20
0BBB@
1CCCA:
It is easy to check that C and O are non-singular M-matrices and if Ois a weakly column diagonally dominant matrices. From Theorem 6,the GN (D.1) has a T-Periodic oscillation when aiðtÞ ¼ bjðtÞ ¼0:02 sin t, which is globally and asymptotically stable, as shown inFig. D.2. By the numerical simulations, we can find that the ratioof amplitude of oscillation to perturbation is 1.34, which meanthe GN can magnify the circumstance perturbation.
One major difficulty often encountered in the analysis of generegulatory networks is the vast separation of time scales betweenthe fast reactions and the slow reactions. Here, we consider a motifwhich has fast reactions of mRNA and the slow reactions ofprotein.
Example 4. Now, We consider a a positive feedback loop withthree nodes, which can be find in Escherichia coli and yeast, seeFig. D.3. Half-lives of mRNA range from a few minutes to severalhours and are peaked around 20 min in yeast and Protein half-livesvary from a few minutes to several days. We considered thefollowing dimensionless differential equations with a set ofbiologically plausible parameters:
150
_m1ðtÞ ¼ �miðtÞ þ 1:4 11þðR þ1
0f13ðsÞp3ðt�sÞdsÞ2
þ a1ðtÞ;
150
_m2ðtÞ ¼ �miðtÞ þ 1:5ðR þ1
0fijðsÞp1ðt�sÞdsÞ2
1þðR þ1
0f21ðsÞp1ðt�sÞdsÞ2
þ a2ðtÞ;
150
_m3ðtÞ ¼ �miðtÞ þ 1:3 11þðR þ1
0f32ðsÞp2ðt�sÞdsÞ2
þ a3ðtÞ;
_p1ðtÞ ¼ �p1ðtÞ þ 0:95Rþ1
0 g1ðsÞm1ðt � sÞdsþ b1ðtÞ;_p2ðtÞ ¼ �p2ðtÞ þ 0:95
Rþ10 g2ðsÞm2ðt � sÞdsþ b2ðtÞ;
_p3ðtÞ ¼ �p3ðtÞ þ 0:95Rþ1
0 g3ðsÞm3ðt � sÞdsþ b3ðtÞ:
ðD:2Þ
By appropriate computation, we have

Y. Wang et al. / Mathematical Biosciences 220 (2009) 34–44 43
C ¼
1 0 0 0 0 � 147160
0 1 0 � 6364 0 0
0 0 1 0 � 273320 0
� 1920 0 0 1 0 0
0 � 1920 0 0 1 0
0 0 � 1920 0 0 1
0BBBBBBBBB@
1CCCCCCCCCA
and
O ¼
173032 0 0 0 0 � 1470
32
0 162532 0 � 1475
32 0 00 0 1835
32 0 � 136532 0
� 1920 0 0 21
20 0 00 � 19
20 0 0 2120 0
0 0 � 1920 0 0 21
20
0BBBBBBBBB@
1CCCCCCCCCA:
It is easy to check that C and O are non-singular M-matrices. FromTheorem 5 that the GN (D.2) has a T-Periodic oscillation withaiðtÞ ¼ 0:02 sin t;bjðtÞ ¼ 0:02 cos t (aiðtÞ ¼ 0:05 sin t; bjðtÞ ¼ 0:05 cos t),which is globally attractive, as shown in Figs. D.4 and D.5respectively.
0 2 4 6 8 10 12 14 16 18 20 22 24
0.4
0.5
0.6
0.7
0.8
0.9
1
1.1
Time (π) →
____ mRNA concertration...... protein concertrationα =0.02sint β =0.02cost
Fig. D.4. Time evolution of three mRNAs and protein concentrations of GN (D.2)with aiðtÞ ¼ 0:02 sin t;bjðtÞ ¼ 0:02 cos t and fijðsÞ ¼ giðsÞ ¼ e�s .
0 2 4 6 8 10 12 14 16 18 20 22 24
0.4
0.5
0.6
0.7
0.8
0.9
1
1.1
1.2
1.3
Time ( π ) →
____ mRNA concertration....... protein concertrationα=0.05sint β=0.05cost
Fig. D.5. Time evolution of three mRNAs and protein concentrations of GN (D.2)with aiðtÞ ¼ 0:05 sin t and bjðtÞ ¼ 0:05 cos t and fijðsÞ ¼ giðsÞ ¼ e�s .
Noting that the conditions are conservative, the globally at-tracted orbit can exist a large parameter space than we obtain .
Appendix E. Discussion
A cellular system is generally characterized with significanttime delays in gene regulation, in particular, for the transcription,translation, diffusion, and active transport. Moreover, periodic per-turbations are widespread in the external environment(e.g. dailylight-dark cycle and Moon’s gravitational)and internal circum-stance (e.g. cell division cycle or cellular motility). Such time delaysand perturbation may affect the dynamics of the entire biologicalsystem, both qualitatively and quantitatively.
In this paper, we derive new criteria for evaluating the globalstability of periodic oscillation for gene networks with small per-turbation, distributed delay, and lager time scales reaction. We re-sults relay on the of Lipschtiz conditions of Hill function, topologyof gene regulation networks and delay kernels. By numerical stim-ulation, we find that the amplitude of gene oscillation relay on theamplitude of perturbation regulatory functions and time scales,and the average of mRNA and protein contractions relay on the dif-ferent delay kernels and the size of changes of the environment. Inparticular, our method based on the proposed model transformsthe original network into matrix analysis problem, thereby notonly significantly reducing the computational complexity but alsomaking analysis of periodic oscillation tractable for even large-scale nonlinear networks.
References
[1] P. Smolen, D.A. Baxter, J.H. Byrne, Mathematical modeling of gene networks,Neuron 26 (3) (2000) 567–580.
[2] P. Smolen, D.A. Baxter, J.H. Byrne, Modeling transcriptional control in GeneNetworks methods, recent results, and future directions, Bulletin ofMathematical Biology 62 (2000) 247–292.
[3] M.B. Elowitz, S. Leibler, A synthetic oscillatory network of transcriptionalregulators, Nature 403 (2000) 335–338.
[4] T.S. Gardner, C.R. Cantor, J.J. Collins, Construction of a genetic toggle switch inEscherichia Coli, Nature 403 (2000) 339–342.
[5] D. McMillen, N. Kopell, J. Hasty, and J. Collins, Synchronizing genetic relaxationoscillators by intercell signaling, Natl. Acad. Sci., U.S.A., 99 (2002) 679-684.
[6] R. Wang, L. Chen, Synchronizing genetic oscillators by signaling molecules, JBiol Rhythms 20 (2005) 257–269.
[7] C. Li, L. Chen, K. Aihara, synchronization of coupeled nonidentical geneticoscillators, Phys. Bio. 3 (2006) 37–44.
[8] C. Li, L. Chen, K. Aihara, Stochastic synchronization of genetic oscillatornetworks, BMC Syst. Biol. 1 (2007), doi:10.1186/1752-0509-1-6.
[9] L. Chen, K. Aihara, Stability of genetic regulatory networks with time delay,IEEE Trans. Circuits Syst. I. 49 (2002) 602–608.
[10] C. Li, L. Chen, Stability of Genetic Networks With SUM Regulatory Logic Lur’eSystem and LMI Approach, IEEE Trans. Circuits Syst. I. 53 (2006) 2451–2458.
[11] D. Bratsun, D. Volfson, L.S. Tsimring, and J. Hasty, Delay-induced stochasticoscillations in gene regulation, Natl. Acad. Sci., U.S.A., (2005)14593-14598.
[12] K. Rateitschak, O. Wolkenhauer, Intracellular delay limits cyclic changes ingene expression, Mathematical Biosciences 205 (2007) 163–179.
[13] M.B. Elowitz, A.J. Levine, E.D. Siggia, P.S. Swain, Stochastic gene expression in asingle cell, Science 297 (2002) 1183–1186.
[14] J. Paulsson, Summing up the noise in gene networks, Nature 427 (2004) 415–418.
[15] J.M. Raser, E.K. O’Shea, Noise in gene expression, Origins, consequences, andcontrol, Science 309 (2005) 2010–2013.
[16] W.J. Blake, M. Kaern, C.R. Cantor, J.J. Collins, Noise in eukaryotic geneexpression, Nature 422 (2003) 633–637.
[17] M. Kaern, T.C. Elston, W.J. Blake, J.J. Collins, Stochasticity in gene expression:From theories to pheno-types, Nature Reviews Genetics 6 (2005) 451–464.
[18] C. Li, L. Chen, K. Aihara, Transient resetting: A novel mechanism for synchronyand its biological examples, PLoS Comp Biol 2 (2006) e103.
[19] C.H. Yuh, H. Bolouri, E.H. Davidson, Genomic cis-regulatory logic, Experimentaland computational analysis of a sea urchin gene, Science 279 (1998) 1896–1902.
[20] N.E. Buchler, U. Gerland, T. Hwa, On schemes of combinatorial transcriptionlogic, Proc. Natl. Acas. Sci. USA 100 (2003) 5136–5141.
[21] S. Kalir, S. Mangan, and U. Alon, A coherent feed-forward loop with a SUMinput function prolongs flagella expression in Escherichia coli, Molecular Syst.Biol. (2005) 10.1038/msb4100010.
[22] J.K. Hale, S.M. Verduyn Lunel, introdunction to Functional DifferentialEquations, fourth ed., Springer, Ne York, 1993.

44 Y. Wang et al. / Mathematical Biosciences 220 (2009) 34–44
[23] J. Hasty, N. Dolnik, V. Rottschaer, J.J. Collins1, Synthetic Gene Network forEntraining and Amplifying Cellular Oscillations, PRL. V88 (14) (2002) 148101.
[24] A. Berman, R.J. Plemmons, Nonnegative Matrices in The Mathematical Science,Academic, New York, 1979.
[25] Q.K. Song, J. Cao, Global exponential stability of bidirectional associativememory neural networks with distributed delays, Journal of Computationaland Applied Mathematics 202 (2007) 266–279.
[26] T. Burton, Stability and Periodic Solution of Ordinary Differential Equation andFunctional Differential Equations, Academic, Orlando, FL, 1985.
[27] J.K. Hale, Theory of Functional Differential Equations, Springer, New York, 1977.[28] J.C. Dunlap, Molecular bases for circadian clocks, Cell 96 (1999) 271–290.[29] J.K. Yisraeli, S. Sokol, D.A. Melton, A two-step model for the localization of
maternal mRNA in Xenopus oocytes, involvement of microtubules andmicrofilaments in the translocation and anchoring of Vg1 mRNA,Development 108 (1990) 289–298.
[30] M.L. King, Molecular basis for cytoplasmic localization, Dev. Genet. 19 (1996)183–189.
[31] J. Sabry, T. O’Connor, M.W. Kirschner, Axonal transport of tubulin in Ti1Pioneer neurons in situ, Neuron 14 (1995) 1247–1256.
[32] C. Jacobs, L. Shapiro, Microbial asymmetric cell division, localization of cell fatedeterminants, Curr. Opin. Gen. Dev. 8 (1998) 386–391.
[33] J. Chen, X. Chen, Special Matrices, Tsinghua University Press, Beijing, 2001.277.
[34] R.E. Gaines, J.L. Mawhin, Coincidence Degree and Nonlinear DifferentialEquations, Springer, Berlin, 1977.
[35] K. Gopalsamy, Stability and Oscillation in Delay Equation of PopulationDynamics, Kluwer, Dordreeht, 1992.
[36] C.W. Eurich, A. Thiel, L. Fahse, Distributed delays stabilize ecological feedbacksystems, Phys. Rev. Lett. 94 (15) (2005) 158104.
[37] V. Chickarmane, C. Troein, U.A. Nuber, H.M. Sauro, C. Peterson, Transcriptionaldynamics of the embryonic stem cell switch. PLoS Comput Biol 2(9): e123.DOI: 10. 1371/journal.pcbi.0020123.
[38] U. Alon, Network motifs: theory and experimental approaches, NatureReviews Genetics 8 (2007) 450–461.
Copyright © 2022 FDOKUMEN


















