
Role of mitogen activated protein kinases in skin tumorigenicity of patulin
Upload
independentCategory
view
2download
0
ORIGINAL ARTICLE
V. S. Thakur Æ B. Shankar Æ S. ChatterjeeS. Premachandran Æ K. B. Sainis
Role of tumor-derived transforming growth factor-b1 (TGF-b1) insite-dependent tumorigenicity of murine ascitic lymphosarcoma
Received: 28 September 2004 / Accepted: 7 December 2004 / Published online: 23 February 2005� Springer-Verlag 2005
Abstract An ascitic lymphosarcoma (LS-A) of Swissmice that regressed spontaneously on subcutaneous(s.c.) transplantation was investigated for the mechan-ism of its progressive growth and host mortality on in-traperitoneal (i.p.) transplantation. In vitro studiesindicated significant inhibition of LS-A proliferationseeded at higher cell density (>104/ml). Culture super-natants of LS-A caused bi-modal growth effects, theearly supernatants (24 h) caused stimulation and the late(72 h) supernatants inhibited LS-A proliferation. The72-h supernatants also suppressed T and B cell responseto mitogens in a dose-dependent manner. Pan anti-transforming growth factor-b antibody abrogated theinhibitory effects of supernatants. The supernatantscontained both latent as well as bio-active form oftransforming growth factor-b1 (TGF-b1) as determinedby ELISA. Mice bearing i.p. ascites tumor had elevatedserum TGF-b1, hemoglobulinemia, splenic lymphope-nia, impaired response of the T cells to mitogen andreduced expression of transferrin receptor (CD71) on thebone marrow cells. However, mice which rejected s.c.transplants, did not show significant changes in theseparameters. Our studies indicated profound influence ofsite of tumor growth on tumor progression and hostimmune system mediated by tumor-derived TGF-b1. Itis possible that human tumors which secrete TGF-b1may exhibit similar patho-physiological effects in thehost depending on the anatomical site of the tumor.
Keywords TGF-b Æ Immunosuppression ÆSite-dependent tumor growth Æ Hemoglobulinemia ÆLymphopenia Æ Transferrin receptor (CD71)
Introduction
The site of tumor growth and its tissue microenviron-ment is an important factor in tumor progression, inva-sion, metastasis, host immune response and response totherapeutic treatments. Preclinical studies have showndifferences in response to chemotherapy when the sametumor was implanted at several anatomical sites [8, 33,37] suggesting profound influence of organ microenvi-ronments in the control of tumor growth. Differentialorgan microenvironmental effects are also reported in thedevelopment of anti-tumor immunity by the host. Usingorthotopic and ectopic transplantation of pancreatictumor expressing human MUC1 transgene, the organsite was shown to be critical in the development of anti-MUC1 immune response in mice [20]. Understanding thehost–tumor interactions of a particular tumor at differentanatomical sites is, therefore, important so as to developeffective therapeutic treatments for primary as well asmetastatic tumor growths.
Tumors which express immunogenic epitopes can eli-cit anti-tumor immune responses and often undergorejection in syngeneic/autochthnous host and it is acumulative result of the action of a number of cellular andhumoral mediators [24]. Cytokines elaborated by thestromal tissue, the cells of immune system and the tumoritself can modify tumor microenvironment and hencehost response to tumors. Many human tumors secretecytokines that act as autocrine tumor growth factors.These include interleukin-6 in hematological tumors suchas myeloma or hairy cell leukemia and renal cell carci-noma, tumor necrosis factor (TNF) in leukemia or neu-roblastoma, interleukin-10 (IL-10) in lymphoma [5] andtransforming growth factor-b (TGF-b) in case of carci-noma [1, 36]. The spectrum of tumor-derived cytokineshas been shown to be different at different anatomical
V. S. Thakur Æ B. Shankar Æ S. Chatterjee Æ S. PremachandranK. B. Sainis (&)Radiation Biology Health Sciences Division,Bioscience Group, Modular Laboratories,Bhabha Atomic Research Centre,Trombay, Mumbai, 400 085 IndiaE-mail: [email protected].: +91-22-25593638/+91-22-25505326Fax: +91-22-25505326/+91-22-25505151
Cancer Immunol Immunother (2005) 54: 837–847DOI 10.1007/s00262-004-0656-z

sites of tumor [14]. These cytokines may cause bothgeneralized and specific inhibition of immune responses[6]. Earlier, we have reported that an ascitic lymphosar-coma (LS-A) tumor in Swiss mice, showed spontaneousregression on s.c transplantation and induced anti-tumorimmunity that could be transferred adoptively by tumor-immune CD4+ T cells [35]. The anti-tumor immune re-sponse against LS-A tumor was not observed in micetransplanted by i.p. route and even a very small dose ofLS-A tumor inoculum was lethal to the mice. The presentstudies were undertaken to examine the mechanismunderlying i.p. site-dependent tumorigenicity of LS-Acells in the syngeneic Swiss mice and to ascertain the roleof tumor-derived cytokines, if any.
Materials and methods
Chemicals
RPMI 1640 medium, sodium bicarbonate, 2-mercapto-ethanol, fetal calf serum, anti-mouse CD71, anti-mouse-IgG-PE, pan TGF-b antibody and anti-rabbit-IgG-PEwere obtained from Sigma Chemical Company, USA.TGF-b1, TGF-b2, TGF-a, Oncostatin M (OSM), Leu-kemia inhibitory factor (LIF) were purchased fromBoehringer Mannheim, Germany. Concanavalin A (ConA) was procured from CalBiochem, USA. LPS waspurchased from Difco Laboratories, USA. Purified ratanti-mouse, human, pig TGF-b1 monoclonal antibody(Rat IgG2a, j; clone A75-2.1) and biotinylated rat anti-mouse, human, pig monoclonal antibody (Rat IgG2a, j,clone A75-3.1) were procured from BD PharMingen,USA. 3H-thymidine (specific activity: 6.5 Ci/mM) wasobtained from Board of Radiation and Isotope Tech-nology, Mumbai, India. Penicillin and streptomycinwere purchased from reputed local manufacturers.
Animals
Inbred Swiss albino male mice, 8–10 weeks old andweighing between 23–25 g were reared in the animalhouse facility of the Bhabha Atomic Research Centre(BARC). Inbred Swiss mice were obtained by breedingamong littermates for more than 40 generations. Thesyngenicity was routinely checked by reciprocal skingrafts, which survived indefinitely. The guidelines issuedby the Institutional Animal Ethics Committee ofBhabha Atomic Research Centre, Government of India,regarding the maintenance and dissection of experi-mental animals were followed.
Tumor
These studies were carried out on an ascitic lympho-sarcoma (LS-A), which had originated spontaneously ininbred Swiss mice [35]. The tumor had arisen sponta-
neously in a mouse in the Swiss mouse colony of ourResearch Centre. The mouse showed splenomegaly andlymph node enlargement but no ascites. The asciticvariant, LS-A, arose following serial i.p. passages of theoriginal tumor cells of the spleen. The tumor wasmaintained by serial i.p. transplantation of 106 cells/animal, twice a month, into naı̈ve mice.
Medium
All the cell suspensions were prepared in RPMI 1640medium. The medium was supplemented with sodiumbicarbonate (2 g/l), 2- mercaptoethanol (5·10�5 M),10% heat-inactivated fetal calf serum, penicillin (100 U/ml) and streptomycin (100 lg/ml). This is referred to ascomplete medium.
Tumor cell suspension
Ascitic lymphosarcoma cells were obtained ascepticallyfrom peritoneal lavage of mice bearing 10–15-day-oldascitic tumor. The cells were washed twice in completemedium by centrifugation. Viability was determined bytrypan blue dye exclusion method. It was usually>95%.
Generation of LS-A culture supernatants
Ascitic lymphosarcoma cells (2·106) were cultured in10 ml complete medium at 37�C in a humidified 95% air/5% CO2 atmosphere, using tissue culture flasks (Falcon).Cell-free supernatants were collected at the end of 24-hand 72-h by centrifugation. Aliquots of supernatantswere stored at �70�C and were used within 3 weeks.
Measurement of effect of culture supernatantson in vitro proliferation of LS-A cells
The effect of LS-A-derived culture supernatants onproliferation of LS-A cells was determined using pro-liferation assay based on the incorporation of 3H-thy-midine [7] with minor modifications. LS-A cells (104/ml)were seeded in six replicates in 0.2 ml complete mediumusing 96 well microplates (Nunc, Denmark) in presenceor absence of culture supernatants. The cultures wereincubated in a CO2 incubator at 37�C for 72 h and werepulsed with 3H-thymidine (1 lCi/well) for additional16 h. The cultures were harvested using a cell harvester(Dynatech; USA) on fibre glass filter papers. The scin-tillation fluid used was prepared using 4 g PPO and 0.3 gPOPOP (Sigma Chemical Company, USA) in 1 L oftoluene (Sarabhai Chemical Company, India). Radio-activity was measured using a liquid scintillation counter(LKB Wallac; Model: 1217 Rack beta).
838

Effect of anti TGF-b antibody on tumor growth
Ascitic lymphosarcoma cells were grown in presence oftumor-derived culture supernatants in presence or ab-sence of pan TGF-b antibody (10 lg/ml) and theirgrowth was assessed as mentioned earlier.
Sensitivity of LS-A tumor cells to recombinant cytokines
The sensitivity of LS-A cells to a spectrum of re-combinant cytokines such as TGF-b1, TGF-b2, TGF-a,OSM, and LIF was determined by proliferation assay.LS-A cells (104/ml) were treated with different concen-trations of cytokines in a total volume of 0.2 ml andcellular proliferation was determined by the method of3H-thymidine incorporation as described earlier.
Estimation of TGF-b1 in culture supernatants and seraof mice bearing s.c. and i.p. tumors
Mice were injected with 106 LS-A cells by either s.c. ori.p. route (3 mice/group). Naı̈ve, normal mice served ascontrol. Blood samples were collected from retro-orbitalplexus on different days (5, 10, 15 and 20); post tumortransplantation and sera were separated. The ELISA forTGF-b1 measures only the active form of the cytokineand acidification would add the latent amount to the un-acidified sample. Serum samples and culture superna-tants stored at �70�C, were acidified with 1 N HCl andanalyzed for TGF-b1 content by sandwich ELISA.Briefly, 96 well ELISA plates were coated with purifiedrat anti-mouse, human, pig TGF-b1 monoclonal anti-body. Recombinant human TGF-b1 (from CHO cells)was used as a standard. Biotinylated, rat anti-mouse,human, pig TGF-b1, monoclonal antibody was used asdetection antibody. Avidin-Horse radish peroxidase wasused as an enzyme conjugate. The substrate used was3,3¢, 5,5¢ tetramethylbenzidine (TMB). The reaction wasstopped by addition of 1 N H2SO4 and optical densitymeasurements were made on an automated ELISAreader (MediSpec Instruments India Ltd) at 450 nm.
Staining for intracellular TGF-b
Detection of intracellular TGF-b was carried out by themethod of Garba et al. [10] with minor modifications.LS-A cells were fixed with 4% formaldehyde for 10 minin ice. Cells were washed with PBS and treated with 10%FCS for 20 min in ice to block non-specific binding ofthe antibody. The cells were then incubated with panTGF-b antibody (1 lg/106 cells) for 30 min in ice. Afterextensive washing with PBS, cells were labeled with anti-rabbit-IgG-PE. The cells treated only with the secondaryantibody served as negative control. Twenty thousandcells were acquired for each sample on flow cytometer(FACS Vantage, Becton Dickinson, USA). The per cent
cells positive for intracellular TGF-b were calculatedusing CellQuest software (Becton Dickinson Immuno-cytometry Systems, USA).
Staining for transferrin receptor (CD71) on bonemarrow cells of tumor bearing mice
Mice were injected with 106 LS-A cells either by s.c. ori.p. route. Naı̈ve, normal mice served as control. Mar-row cells were obtained from the femur bone of normalmice or s.c. and i.p. tumor (15-day-old tumor) bearingmice (3 mice/group). Labeling of CD71 on bone marrowcells was carried out according to the method of Scho-maker et al. [30]. The cells were washed with PBS andwere labeled with 1 lg/106 cells of anti-mouse CD71 for30 min in ice. The isotype control used was mouse IgG1(MOPC-21) at the same concentration. After extensivewashing, the cells were further treated with anti-mouse-IgG-PE for 30 min in ice. Twenty thousand cells wereacquired for each sample on flow cytometer. The per-centage of cells positive for transferrin receptor and themean fluorescence intensity at 585 nm were calculatedusing CellQuest software.
Estimation of blood hemoglobin and total nucleatedcells in the spleen
Mice were injected with LS-A cells (106/animal) by s.c.or i.p. route. Naı̈ve, normal mice served as controlgroup. Three mice were used per group per time point.Blood samples were collected, under anesthesia indi-vidually from the retro-orbital plexus on different days(5, 10, 15, 20 and 23), following tumor transplantation.Hemoglobin content in the heparinized blood sampleswas estimated by Drabkin’s method [2]. Spleens werealso dissected out from these animals on different days(5, 10, 15, 20 and 23) following tumor transplantationand cell suspensions were prepared individually in themedium. Total nucleated cells per spleen were enumer-ated by using methylene blue:acetic acid diluent.
Lymphocyte proliferation assay
The effect of LS-A-derived culture supernatants on thefunction of normal T cells and B cells and those ob-tained from s.c. and i.p. LS-A tumor bearing mice wasdetermined using polyclonal mitogens. Spleen cells wereobtained from either normal mice or mice bearing 15-day-old s.c. and i.p. tumors (3 mice/group). Red bloodcells were lysed by treatment with 0.01 M Tris–NH4Cl(0.83%) solution (pH 7.2). Viability was determined bytrypan blue dye exclusion method. Spleen cells werestimulated with either concanavalin A (2.5 lg/ml per 106
cells) or E. coli lipopolysaccharide (LPS) at 50 lg/ml per106 cells, in a total volume of 0.2 ml complete medium.The cultures were pulsed with 3H-thymidine (1 lCi/well)for 16 h at the end of 72 h and 48 h, respectively.
839

Statistical analyses
The data were analyzed by applying Student’s t test. A Pvalue of 0.05 and less was considered significant.
Results
LS-A growth in vitro
The effect of initial cell density on LS-A proliferation invitro is shown in Fig. 1. Optimal 3H-thymidine incor-poration at the end of 72 h was observed with initial celldensities of 103–104/ml, above which significant(P<0.01) inhibition of 3H-thymidine incorporation wasobserved. Total inhibition of 3H-thymidine incorpora-tion was observed when 106 LS-A cells/ml were initiallyseeded and cultured for 72 h (Fig. 1). It was possiblethat the tumor cells seeded at higher densities did growinitially. However, at the end of 72 h when 3H-thymi-dine was added, no dividing cells remained due to theinhibitory action of some tumor-derived soluble factor.
To ascertain this possibility, LS-A cells were seeded atthe initial density of 2·105/ml. Culture supernatants wereobtained at two different time points. The effects of earlysupernatant (24 h) and late supernatant (72 h) weretested on in vitro proliferation of LS-A cells. It was
observed that the early supernatant caused significant(P<0.01) stimulation of proliferation (Fig. 2a) up to 1:4dilution. At higher dilution (1:8), 3H-thymidine incor-poration decreased but remained higher than the control(Fig. 2a). Figure 2b shows the effect of late supernatants(72 h) on LS-A proliferation. At lower dilution (1:2),significant inhibition (P<0.01) of proliferation of LS-Acells was observed. This inhibition decreased at higherdilution (1:8) of the supernatant and 3H-thymidineincorporation was almost at the level of the control(Fig. 2b).
Immunosuppressive effects of LS-A supernatants
The effect of LS-A culture supernatants on the mitogen-induced stimulation of host lymphocytes is shown inFig. 3. Polyclonal mitogens viz. con A and LPS stimu-lated splenic T and B lymphocytes respectively, of nor-mal mice (Fig. 3). When the spleen cells were co-culturedwith 24-h supernatant in presence of con A, there was nosignificant suppression of T cell proliferation. However,co-culture with 72-h supernatant significantly (P<0.01),suppressed T cell proliferation. A similar trend of sup-pression was observed in the case of LPS-induced pro-liferation of B cells co-cultured with 72-h culturesupernatant (Fig. 3).
This immunosuppressive effect was also seen in vivo.Figure 4a shows lymphocyte counts in spleens of normalmice and mice bearing i.p. or s.c. tumors. In normalmice, the lymphocyte count per spleen did not changesignificantly with time. In case of s.c. and i.p. tumorbearing mice, there was an initial increase in lymphocytecount, probably due to immune response mounted bythe host. In tumor bearing mice, this initial surge in thelymphocyte count declined only by day 23, after tumortransplantation. On the contrary, i.p. tumor bearingmice showed sharp decline in lymphocyte count withtime. Severe lymphopenia was observed in these mice onday 23, a time point when the mice had full-blownascites.
The functional capability of splenic lymphocytes ofs.c. and i.p. tumor bearing mice was evaluated ex vivo.Fifteen days after tumor transplantation, the spleen cellswere stimulated with con A. Lymphocytes from micebearing s.c. tumors showed about 30% reduction in 3H-thymidine incorporation as compared to control mice.Strikingly, lymphocytes of i.p. tumor bearing miceexhibited 95% reduction (P<0.01) in response to con Aas compared to control mice (Fig. 4b).
Identification of LS-A derived soluble factor
As a preliminary step towards identification of the sol-uble factor(s) present in the supernatant, different re-combinant cytokines were added to LS-A cultures andtheir effect on proliferation was measured. Fig. 5a showsthat TGF-b1 caused bi-modal effects on the growth of
Fig. 1 Effect of initial cell density on the growth of LS-A cells invitro. Cells were cultured for 3 days in complete medium in a 95%air/5% CO2 atmosphere before 3H thymidine (1 lCi/well) wasadded and they were cultured further for 16 h. Cellular prolifer-ation was estimated in terms of 3H-thymidine incorporation. Eachvalue represents mean CPM ± SE of six replicates. Twoindependent experiments were carried out. **P<0.01, as comparedto the optimal incorporation at 104/ml
840

LS-A, i.e., at lower concentration (0.01 ng/ml), signifi-cant (P<0.01) stimulation was observed but higherconcentrations of the cytokine (>10 ng/ml) significantly(P<0.01) suppressed LS-A proliferation. The isoformTGF-b2 caused inhibition of proliferation at all theconcentrations used (Fig. 5b). Other cytokines viz.TGF-a, LIF and OSM showed no significant effect onLS-A proliferation at any of the concentrations used(Fig. 5b). Incubation of LS-A cells with supernatants(72 h) treated with pan TGF-b antibody abrogatedproliferation inhibitory effect of these supernatants al-most completely (P<0.01; Fig. 5c), indicating that thesupernatant contained TGF-b as the growth-modulatingfactor.
The above results suggested the involvement of TGF-b in the bimodal action of LS-A culture supernatant.Furthermore, intracellular TGF-b was detected in LS-Acells obtained from i.p. tumor ascites using flowcytometry. About 58% of LS-A cells expressed TGF-b,whereas negative control cells showed less than 5%staining (Fig. 6).
Estimation of TGF-b1 in LS-A supernatants andsera of tumor bearing mice
Figure 7 shows the amounts of active and latent formof TGF-b1 present in the early (24 h) and the late
(72 h) supernatants. To estimate total TGF-b1 (activeplus latent), supernatants were acidified to release thebioactive form. Both, un-acidified and acidified su-
Fig. 2 Proliferation of LS-A cells (1·104/ml) in vitro in thepresence of different dilutions of culture supernatants. a Earlysupernatant (24-h culture) and b late supernatant (72-h culture).Proliferation of cells was measured as explained in Fig. 1. Eachvalue represents mean CPM ± SE of six replicates. Fourindependent experiments were carried out and one representativeexperiment is shown. **P<0.01, as compared to 3H-thymidineincorporation in control group (cells grown in complete medium)
Fig. 3 Effect of 24-h and 72-h culture supernatants on the responseof normal T and B cells to polyclonal mitogens. Spleen cells (1·106/ml) were stimulated with con A (2.5 lg/ml) or LPS (50 lg/ml) in0.2 ml complete medium and cultured for 72 h and 48 h,respectively before the addition of 1 lCi of 3H-thymidine per well.Each value represents mean CPM ± SE of six replicates. Fourindependent experiments were carried out and one representativeexperiment is shown. **P<0.01, as compared to mitogen treatedcontrol. Filled rectangle cells only, line shaded rectangle cells +mitogen, gray shaded rectangle cells + mitogen + 24 h sup, openrectangle cells + mitogen + 72 h sup
841

pernatants were analyzed for TGF-b1 content usingELISA. The acidified supernatants (24 h and 72 h)contained higher quantity of TGF-b1 as compared totheir respective non-acidified counterparts, whichindicated the presence of both the active and latentform of TGF-b1 in the supernatants. Further, the 72-hsupernatants contained almost 20 times more activeTGF-b1 as compared to the 24-h supernatants(Fig. 7).
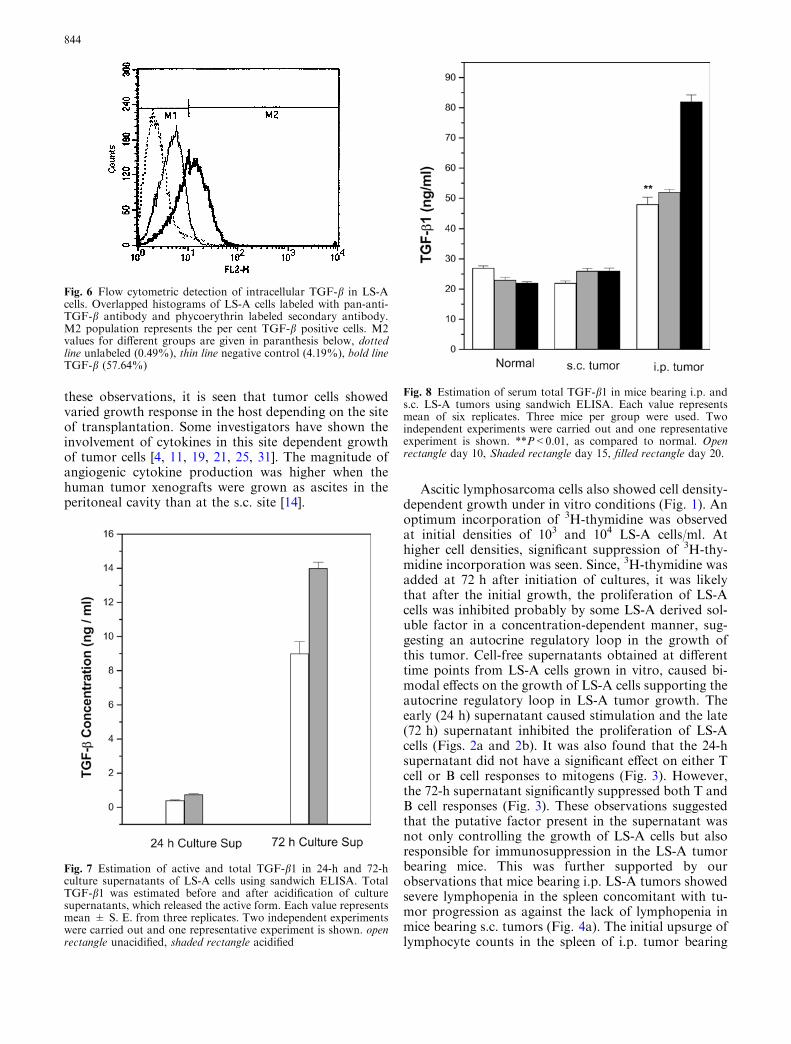
Transforming growth factor-b1 levels were deter-mined in the acidified sera of mice bearing LS-A tumorsat the two anatomical sites (i.p. and s.c.) at different timepoints after tumor transplantation. Serum TGF-b1 levelgradually increased in mice bearing i.p. ascites (Fig. 8).These levels were significantly (P<0.01) higher than thatof normal mice or those bearing s.c. tumors, at all thetime points. However, TGF-b1 levels did not increasesignificantly during s.c. tumor growth as a function oftime and they were comparable to that of normal mice(Fig. 8).
Tumor-induced hemoglobulinemia
Hemoglobin levels were measured in peripheral bloodsamples of normal and tumor (i.p. or s.c) bearing mice.Both normal mice and s.c. tumor bearing mice hadcomparable blood hemoglobin profile (Fig. 9a). Theirlevels also remained steady with time. Interestingly, micebearing i.p. tumors developed severe hemoglobulinemia,concomitant with tumor progression. On day 23, a sig-nificant reduction (62.5%) as compared to hemoglobinlevel of corresponding normal mice was observed. Sinceone of the reasons for hemoglobulinemia could be thereduced uptake of iron carrier transferrin due to reducedexpression of transferrin receptor (CD71) on hemato-poietic erythroid progenitor cells, CD71 expression onbone marrow cells was determined using flow cytometry.The percentage of cells expressing CD71 was not sig-nificantly different among normal, s.c. and i.p. tumorbearing mice (Fig.9b, Table 1). However, the mean flu-orescence intensity which indicates the transferrinreceptor density was significantly reduced (P<0.01) onthe bone marrow cells of i.p. tumor bearing mice ascompared to normal or s.c. tumor bearing mice (Fig. 9b,Table 1).
Discussion
Earlier, we have shown that LS-A cells transplanted byi.p. route formed ascites and caused mortality in therecipient mice at the lowest dose (102 cells/animal) used.However, solid tumor formed by ectopic s.c. transplan-tation of LS-A cells, regressed spontaneously even whena much higher dose of 106 cells/animal was used. Theseanimals subsequently developed anti-tumor immunity
Fig. 4 a Total nucleated cell count per spleen taken at differenttime points following i.p. and s.c. LS-A transplantation. Each valuerepresents mean for nine mice obtained from data of threeexperiments each comprising of three mice per time point. opensquare normal,filled circle s.c. tumor, filled triangle i.p. tumor. bResponse of splenic lymphocytes obtained from i.p. and s.c. tumorbearing mice to con A. Spleen cells were taken 15 days after tumortransplantation and were stimulated with con A (2.5 lg/ml/106
cells/ml) and cultured for 72 h before the addition of 1 lCi of 3H-thymidine per well. Each value represents mean CPM ± SE of sixreplicates. Three mice per group were used in each experiment andtwo such independent experiments were carried out. One represen-tative experiment is shown. **P<0.01 as compared to mitogentreated cells of normal mice. filled rectangle Cells, open rectanglecells + con A
842

[35]. Similar site-dependent growth of tumors has beenshown by other investigators using different tumormodels. Ectopic transplantation of human pancreatic
tumor cells (AsPC-1) into nude mice, caused slowgrowth of tumor [34] AK-5, a rat histiocytoma that grewas ascites on i.p. transplantation, regressed spontane-ously after s.c. transplantation [15]. Human colon car-cinoma cells (KM12) metastasized only following cecalwall transplantation but not when transplanted at thesubcutaneous site [22] indicating organ microenviron-ment as a decisive factor in tumor progression. Anti-tumor responses against human MUC1 antigen ex-pressed as transgene by pancreatic cancer cell line syn-geneic to C57BL/6 mice, were less effective at theorthotopic site than at the ectopic s.c. site [20]. From
Fig. 5 Identification of the tumor-derived cytokine of LS-A cells.Effect of a recombinant TGF-b1 b r-TGFa (open square), r-TGF-b2 (filled circle), r- LIF (open triangle), and r-OSM (Down filledtriangle) on the proliferation of LS-A cells in vitro. Untreated LS-Acells incorporated 223778±8419 cpm. c Effect of pan-anti TGF-bantibody on proliferation of LS-A cells in presence of 72-h culturesupernatants. Each value represents mean CPM ± SE of sixreplicates. Two independent experiments were carried out and onerepresentative experiment is shown. **P<0.01, as compared torespective controls
843
these observations, it is seen that tumor cells showedvaried growth response in the host depending on the siteof transplantation. Some investigators have shown theinvolvement of cytokines in this site dependent growthof tumor cells [4, 11, 19, 21, 25, 31]. The magnitude ofangiogenic cytokine production was higher when thehuman tumor xenografts were grown as ascites in theperitoneal cavity than at the s.c. site [14].
Ascitic lymphosarcoma cells also showed cell density-dependent growth under in vitro conditions (Fig. 1). Anoptimum incorporation of 3H-thymidine was observedat initial densities of 103 and 104 LS-A cells/ml. Athigher cell densities, significant suppression of 3H-thy-midine incorporation was seen. Since, 3H-thymidine wasadded at 72 h after initiation of cultures, it was likelythat after the initial growth, the proliferation of LS-Acells was inhibited probably by some LS-A derived sol-uble factor in a concentration-dependent manner, sug-gesting an autocrine regulatory loop in the growth ofthis tumor. Cell-free supernatants obtained at differenttime points from LS-A cells grown in vitro, caused bi-modal effects on the growth of LS-A cells supporting theautocrine regulatory loop in LS-A tumor growth. Theearly (24 h) supernatant caused stimulation and the late(72 h) supernatant inhibited the proliferation of LS-Acells (Figs. 2a and 2b). It was also found that the 24-hsupernatant did not have a significant effect on either Tcell or B cell responses to mitogens (Fig. 3). However,the 72-h supernatant significantly suppressed both T andB cell responses (Fig. 3). These observations suggestedthat the putative factor present in the supernatant wasnot only controlling the growth of LS-A cells but alsoresponsible for immunosuppression in the LS-A tumorbearing mice. This was further supported by ourobservations that mice bearing i.p. LS-A tumors showedsevere lymphopenia in the spleen concomitant with tu-mor progression as against the lack of lymphopenia inmice bearing s.c. tumors (Fig. 4a). The initial upsurge oflymphocyte counts in the spleen of i.p. tumor bearing
Fig. 6 Flow cytometric detection of intracellular TGF-b in LS-Acells. Overlapped histograms of LS-A cells labeled with pan-anti-TGF-b antibody and phycoerythrin labeled secondary antibody.M2 population represents the per cent TGF-b positive cells. M2values for different groups are given in paranthesis below, dottedline unlabeled (0.49%), thin line negative control (4.19%), bold lineTGF-b (57.64%)
Fig. 7 Estimation of active and total TGF-b1 in 24-h and 72-hculture supernatants of LS-A cells using sandwich ELISA. TotalTGF-b1 was estimated before and after acidification of culturesupernatants, which released the active form. Each value representsmean ± S. E. from three replicates. Two independent experimentswere carried out and one representative experiment is shown. openrectangle unacidified, shaded rectangle acidified
Fig. 8 Estimation of serum total TGF-b1 in mice bearing i.p. ands.c. LS-A tumors using sandwich ELISA. Each value representsmean of six replicates. Three mice per group were used. Twoindependent experiments were carried out and one representativeexperiment is shown. **P<0.01, as compared to normal. Openrectangle day 10, Shaded rectangle day 15, filled rectangle day 20.
844

mice could be due to host sensitization by immunogenicLS-A tumor, which later failed to eliminate the tumor,possibly due to paracrine immunosuppressive effects ofthe putative LS-A-derived soluble factor. Further, a se-vere impairment of splenic T cells response to con A ini.p tumor bearing mice as against a marginal impairmentin s.c. tumor bearing mice (Fig. 4b), suggested animmunosuppressive role of LS-A-derived soluble factorin tumor bearing mice.
The nature of the soluble factor in LS-A culturesupernatant could be deduced from the response of
LS-A cells to different recombinant cytokines. r-TGF-b1showed bi-modal effect on tumor growth (Fig. 5a),similar to that seen with early and late supernatants ofLS-A cultures (Fig. 2a, b). r-TGF-b2 showed growthinhibition at all the concentrations used (Fig. 5b). Pro-liferation of LS-A cells was not affected by other cyto-kines r-TGF-a, r-LIF and r-oncostatin M (Fig. 5b).Further, pan anti-TGF-b antibody completely abolishedthe inhibitory effect of 72-h culture supernatant on LS-Acells which confirmed the role of TGF-b as the majorgrowth modulator present in the LS-A supernatants(Fig. 5c). TGF-b was also detected intracellularly(Fig. 6) in LS-A cells using flow cytometry providing anadditional evidence that this cytokine is released in thesupernatant. About 60% of LS-A cells were positive forTGF-b, indicating heterogeneity of LS-A tumor, whichwas seen earlier by karyotyping [17].
Both latent and active forms of the TGF-b1 weredetected in LS-A derived supernatant. The 24-h super-natant contained lower concentration (0.4 ng/ml) andthe 72-h supernatant contained higher concentration(9 ng/ml) of active TGF-b1 (Fig. 7). These concentra-
Table 1 Expression of CD71 on bone marrow cells of normal miceand mice bearing s.c. and i.p. tumors
Group Mean fluorescence intensity(Percentage of bone marrow cellspositive for transferrin receptor CD71)
Normalmice 169.20±5.0 (32.97±0.8)s.c. tumor bearing mice 151.61±4.8 (35.49±3.2)i.p. tumor bearing mice 120.05±1.4 (29.89±1.8)
Fig. 9 a Total hemoglobinlevels in peripheral blood takenat different time pointsfollowing i.p. and s.c. tumortransplantation. Each valuerepresents mean for nine miceobtained from pooling threeexperiments comprising of threemice per time point. filledsquare normal, open circle s.c.tumor, filled triangle i.p. tumor.b Flow cytometric profilesshowing CD71 expression onbone marrow cells of normalmice and mice bearing s.c. andi.p. tumors
845

tions of active TGF-b1 in the culture supernatants(Figs. 2a, b, respectively) matched with stimulatory andinhibitory effects respectively observed with r-TGF-b1(Fig. 5a).
The cytokines belonging to the TGF-b family aresecreted by many types of normal and malignant cells[26, 28]. Different isoforms identified as TGF-b1 toTGF-b 5 are constitutively produced in a latent formthat contains mature TGF-b and its NH2-terminalpropeptide called latency-associated peptide (LAP). TheTGF-b/LAP complex is noncovalently attached tomembers of latent TGF-b binding proteins (LTBPs) inmost cells forming the large latent complex [23, 29].However, several cell lines established from malignanttissues have been shown to secrete active form of TGF-b[3, 9, 16]. TGF-b is reported to play a key role inembryonic development, hematopoisis, wound healing,matrix formation, immune function as well as diseasestates including chronic inflammation, immune disor-ders and cancer [1, 27]. In the present study, we coulddetect a biologically active form of TGF-b1 in LS-Atumor-derived supernatants and its elevated serum levelsin progressive i.p. tumor bearing mice (Figs. 7, 8,respectively). We have observed decreased tumor celldensity in peritoneal ascites, at late stage of tumorgrowth and proliferation-inhibitory activity of cell-freesupernatants of ascites (Data not shown). High con-centration of TGF-b1 in i.p. transplanted mice led tohost mortality subsequently. The death of i.p. tumorbearing mice may be due to several reasons including (1)tumor load (2) immunosuppression and subsequentinfections (3) paracrine effects of TGF-b, etc.
The amount of TGF b1 is very high in i.p. tumorbearing mice. Normally, one would expect a rapid he-patic elimination of this active cytokine. However thismay not be so in the progressively growing tumorbearing mice as, among others, hepatic function mayalso be compromised. Even in clinical situation, e.g.,patients with colorectal carcinoma, high levels of TGFb1 (53±12 ng/ml) have been observed before surgery.After tumor resection the level dropped to 36±6 ng/mlindicating elevated TGF b1 as a sign of progressivedisease [32]. In other carcinomas, an association wasfound between high tumor burden and elevated circu-lating plasma levels of TGF-b1 [1].
This cytokine is reported to impair a number of im-mune functions such as T cell growth, cytotoxicity and Tcell cytokine production necessary for potentiation ofCTL activity and antigen presentation [5]. It was alsoobserved that mice bearing i.p. tumors developed pro-gressive hemoglobulinemia which was absent in micetransplanted by s.c. route (Fig. 9a). This could be partlyexplained by the reduced expression of transferrinreceptor (CD71) on the bone marrow cells of i.p. tumorbearing mice as compared to that of s.c. tumor bearingmice (Fig. 9b). Transferrin receptor is a marker onproliferating cells and is involved in uptake of ironloaded transferrin which determines intracellular ironlevels [12]. Reduction in the expression of CD71 have
been shown in activated human T lymphocytes treatedwith TGF-b [13] under in vitro conditions. It is possiblethat LS-A tumor-derived TGF-b1 can impair uptake oftransferrin via transferrin receptor down-regulation onthe hemopoietic erythroid progenitors and thus con-tribute to hemoglobulinemia observed in i.p. tumorbearing mice. TGF-b1 or the tumor itself may enhancethe tumoricidal activity of macrophages [15, 18] but thismechanism may be more relevant to subcutaneouslytransplanted tumor.
To conclude, our studies identified TGF-b1 as theendogenous growth factor involved in LS-A growth. Itdisplayed bimodal growth effects on LS-A proliferationin vitro depending on the concentration. The cytokinecaused patho-physiological effects like lymphopenia,hemoglobulinemia, reduction in CD71 expression onbone marrow cells and impaired the host T and B cellfunction by paracrine effects. Tumor progression andmortality in i.p. transplanted mice could be attributed toelevated levels of serum TGF-b1. Further, the datasuggested that the tumorigenicity or immunogenicity isdependent on the route of transplantation. This is thefirst report showing tumor-derived TGF-b1 as a site-specific modulator of murine tumor of spontaneousorigin.
Acknowledgements The authors, thank Shri S. Santosh Kumar forcritically going through the manuscript and Shri N.S. Sidnalkar,also, Shri K.S. Munankar for their expert technical help in animalexperiments.
References
1. Akhurst RJ, Derynck R (2001) TGF-b signaling in cancer—a double-edged sword. Trends Cell Biol 11:S44–S51
2. Balasubramaniam P, Malathi A (1992) Comparative study ofhemoglobin estimated by Drabkin’s and Sahli’s methods. JPost Grad Med 38:8–9
3. Bodmer S, Srommer K, Frei K, Siepl C, de,Tribolet N, Heid I,Fontana A (1989) Immunosuppression and transforminggrowth factor beta in glioblastoma. Preferential production oftransforming growth factor beta2. J Immunol 143:3222–3229
4. Burbridge MF, Kraus-Berthier L, Naze M, Pierre A, Atassi G,Guilbaud N (1999) Biological and pharmacological character-ization of three models of human ovarian carcinoma estab-lished in nude mice: use of the CA 125 tumor marker to predictantitumor activity. Int J Oncol 15:1155–1162
5. Chouaib S, Asselin-Paturel C, Mami-Chouaib F, Caignard A,Blay JY (1997) The host-tumor immune conflict: from imm-nosuppression to resistance and destruction. Immunol Today18:493–497
6. Czarniecki W, Chiu HH, Wong GH, Mc Cabe SM, PalladinoMA (1988) Transforming growth factor beta 1 modulates theexpression of class II histocompatibility antigens on humancells. J Immunol 140:4217–4223
7. De Coteau JF, Knaus PI, Yankelev H, Reis MD, Lowsky R,Lodish HF, Kadin ME (1997) Loss of functional cell surfacetransforming growth factor beta (TGF-b) type I receptor cor-relate with insensitivity to TGF-b in chronic lymphocyte leu-kemia. Proc Natl Acad Sci USA 94:5877–5881
8. Donelli MG, Russo R, Garattini S (1975) Selective chemo-therapy in relation to the site of tumor transplantation. Int JCancer 32:78–88
846

9. Fischer JR, Darjes H, Schindel M, Drings P, Krammer PH(1994) Constitutive secretion of bioactive transforming growthfactor beta1 by small cell lung cancer cell lines. Eur J Cancer30A:2125–2129
10. Garba L, Frelinger JA (2001) Intracellular cytokine staining forTGF-beta. J Immunol Methods 258:193–198
11. HamiltonTC,YoungRC,LouieKG,Behrens BC,McKoyWM,GrotzingerKR (1984) Characterization of a xenografts model ofhuman ovarian carcinoma which produces ascites and intraab-dominal carcinomatosis in mice. Cancer Res 44:5286–5290
12. Hentze MW, Kuhn LC (1996) Molecular control of vertebrateiron metabolism: mRNA-based regulatory circuits operated byiron, nitric oxide and oxidative stress Proc Natl Acad Sci USA93:8175–8182
13. Kehrl JH, Wakefield LM, Roberts AB, Jakowlew S, Alvarez-Mon M, Derynck R, Sporn MB, Fauci AS (1986) Productionof transforming growth factor b by human T lymphocytes andits potential role in the regulation of T cell growth. J Exp Med163:1037–1050
14. Keyes KA, Mann L, Teicher B, Alvarez E (2003) Site-depen-dent angiogenic cytokine production in human tumor xeno-grafts. Cytokine 21:98–104
15. Khar A (1993) Characterization of spontaneous immunologicalrejection in a histiocytoma. I. Role of NK cell and anti tumorantibodies. Int J Oncol 3:393–398
16. Kloen P, Jennings CL, Gebhardt MC, Springfield DS, MankinHJ (1994) Expression of transforming growth factor beta(TGF-b) receptors. TGF-b1 and TGF-b2 production andautocrine growth control in osteosarcoma cells. Int J Cancer58:440–445
17. Kumar M, Seshadri M, Poduval TB, Chauhan PS, SundaramK (1987) Cytogenetic studies on murine lymphosarcoma. AdvBiosci 6:35–44
18. Liaw FD, Denkins YM, Collins JH, Van Arsdall M, Chang JL,Chakrabarty S, Nguyen D, Kruzel E, Fidler IJ (2002) Type-1transforming growth factor-beta differentially modulatestumoricidal activity of murine peritoneal macrophages againstmetastatic variants of the B16 murine melanoma. J Exp TherOncol 2:286–297
19. Mesiano S, Ferrara N, Jaffe RB (1998) Role of vascularendothelial growth factor in ovarian cancer: Inhibition ofascites formation by neutralization. Am J Pathol 153:1249–1256
20. Morikane K, Tempero R, Sivinski CL, Kitajima S, Gendler SJ,Hollingsworth MA (2001) Influence of organ site and tumorcell type on MUC1-specific tumor immunity. Int Immunol13:233–240
21. Morris PC, Scholten V (1996) Osmotic lysis of tumor spill inovarian cancer: a murine model. Am J Obstet Gynecol175:1489–1492
22. Nakajima M, Morikawa K, Fabra A, Bucana CD, Fidler IJ(1990) Influence of organ environment on extracellular matrixdegradative activity and metastasis of human colon carcinomacells. J Natl Cancer Inst 82:1890–1898
23. Oklu R, Hasketh R (2000) The latent transforming growthfactor beta binding protein (LTBP) family. Biochem. J352:601–610
24. Papac RJ (1996) Spontaneous regression of cancer. CancerTreat Rev 22:395–423
25. Paterson C, Davies M, Stone A, Huntley S, Smith E, Pring M,Eveson JW, Robinson CM, Parkinson EK, Prime SS (2002)TGF-beta 1 acts as a tumor suppressor of human malignantkeratinocytes independently of Smad 4 expression and ligand-induced G(1) arrest. Oncogene 21:1616–1624
26. Roberts B, Flanders KC, Kondaiah P, Thompson NL, Ob-berghen- Schilling E, Wakfield L, Rossi P, de Cambrugghe B,Heine U, Sporn MB (1988)Transforming growth factor beta:biochemistry and roles in embriogenesis, tissue repair andremodeling and carcinogenesis. Recent Prog Horm Res 44:157–168
27. Roberts B, Flanders KC, Heine UI, Jakowlew S, Kondaiah P,Kim SJ, Sporn MB (1990) Transforming growth factor-beta:multifunctional regulator of differentiation and development.Philos Trans R Soc Lond B Biol Sci 327:145–154
28. Rodeck U, Bosller A, Graeven U, Fox FE, Nowell PC, KnabbeK, Kari C (1994) Transforming growth factor beta productionand responsiveness in normal human melanocytes and mela-noma cells. Cancer Res 54:575–581
29. Saharinen J, Hyytiainen M, Taipale J, Keski-Oja J (1999) La-tent transforming growth factor beta binding proteins(LTBPs)– structural extracellular matrix proteins for targetingTGF beta action. Cytokine Growth Factor Rev 10:99–117
30. Schomaker SJ, Clemo FA, Amacher DE(2002) Analysis of ratbone marrow by flow cytometry following in vivo exposure tocyclohexanone oxime or daunomycin HCl. Toxicol ApplPharmacol 185:48–54
31. Schwarz RE, McCarty TM, Peralta EA, Diamond DJ, Ellen-horn JD (1999) An orthotopic in vivo model of human pan-creatic cancer. Surgery 126:562–567
32. Shim KS, Kim KH, Han WS, Park EB (1999) Elevated serumlevels of transforming growth factor-beta1 in patients withcolorectal carcinoma: its association with tumor progressionand its significant decrease after curative surgical resection.Cancer 85:554–561
33. Staroselsky N, Fan D, O’Brian CA, Bucana CD, Gupta KP,Fidler IJ (1990) Site-dependent differences in response of theUV-2237 murine fibrosarcoma to systemic therapy with adri-amycin. Cancer Res 50:7775–7782
34. Tan MH, Chu TM (1985) Characterization of the mousetumorigenic and metastatic properties of a human pancreatictumor cell line (AsPC-1) implanted orthotopically into nudemice. Tumor Biol 6:89–98
35. Thakur VS, Seshadri M, Shah DH, Poduval TB (1997)Adoptive immunotherapy of murine lymphosaroma: compar-ative study using allo-immune or tumor-immune T cells. Ind JExp Biol 35:581–588
36. Vanky F, Nagy N, Hising C, Sjovall K, Larson B, Klein E(1997) Human ex vivo carcinoma cells produce transforminggrowth factor-b and thereby can inhibit lymphocyte function invitro. Cancer Immnol Immunother 43:317–323
37. Wilmanns C, Fan D, O’Brian CA, Bucana CD, Fidler IJ (1992)Orthotopic and ectopic organ environments differentiallyinfluence the sensitivity of murine colon carcinoma cells todoxorubicin and 5-fluorouracil. Int J Cancer 52:98–104
847
Copyright © 2022 FDOKUMEN




















