
Rice histone deacetylase genes display specific expression patterns and developmental functions
6
Rice histone deacetylase genes display specific expression patterns and developmental functions Yongfeng Hu a , Fujun Qin a , Limin Huang a , Qianwen Sun a , Chen Li a , Yu Zhao a , Dao-Xiu Zhou a,b, * a National Key Laboratory for Crop Genetic Improvement, Huazhong Agricultural University, 430070 Wuhan, China b Institut de Biotechnologie des Plantes, Université Paris sud 11, 91405 Orsay, France article info Article history: Received 28 July 2009 Available online 5 August 2009 Keywords: Chromatin Epigenetic modification Histone acetylation Acetyltransferase Rice development abstract Histone deacetylases (HDAC) are important in plant gene expression. Here we show that the expression of rice HDAC genes is both tissue/organ-specific, and most of them are responsive to drought or salt stres- ses. Over-expression of several rice HDACs did not produce any visible phenotype, whereas down-regu- lation of a few HDAC genes affected different developmental aspects. Specifically, down-regulation of HDA703 by amiRNA reduced rice peduncle elongation and fertility, while inactivation of a closely related homolog HDA710 by RNAi affected vegetative growth. HDA704 RNAi altered plant height and flag leaf morphology. Down-regulation of HDT702 led to the production of narrowed leaves and stems. These data suggest that rice HDAC genes may have divergent developmental functions compared with closely related homologs in Arabidopsis. Ó 2009 Elsevier Inc. All rights reserved. Histone modifications including acetylation, methylation, phos- phorylation, ubiquitination and others are shown to play an impor- tant role in chromatin function and gene regulation. Specific patterns of histone modifications establish the so-called ‘‘histone code” that determines open or repressed states of the cognate DNA sequences. Histone acetylation/deacetylation appears as a key switch for inter-conversion between permissive and repressive states of chromatin domains for gene expression. The homeostatic balance of nucleosomal histone acetylation is maintained by antag- onistic action of histone acetyltransferases (HAT) and histone deacetylases (HDAC). Plant HDAC can be grouped into four classes. Among them, three have primary homology to three yeast HDAC groups: RDP3, HDA1 and SIR2 [1]. The fourth group known as the HD2 class is found in plants only [2]. During the last years, HDAC function has been most studied in Arabidopsis. Inactivation of Arabidopsis HD1 (AtHD1/HDA19) by expressing an antisense construct and by a T-DNA insertion mutation (athd1-t1) induces pleiotropic developmental abnormalities [3,4]. In addition, AtHD1/HDA19 is shown to form an antagonistic couple with the histone acetyltransferase GCN5 involved in the regulation of several pathways including photomorphogenic seedling develop- ment [5]. Recent data show that AtHD1/HDA19 is involved in expres- sion of genes responding to jasmonic acid and ethylene signaling [6]. Two other Arabidopsis RPD3 type genes AtRPD3A and AtRPD3B are shown to play an important role in plant growth, as down-regulation of the genes delays flowering [7]. Treatment of wild type seedlings with the specific HDAC inhibitor, Trichostatin-A (TSA), and mutations in HDA18 induce ectopic production of root hairs [8]. Furthermore, HDAC genes are found to be involved in epigenetic regulation, as AtH- DA6 is shown to be involved in RNA interference-mediated DNA methylation [9]. In addition, AtHDA6 and HDT2 are involved in nucleolar dominance in allopolyploid hybrids [10]. Rice genome contains at least 19 HDAC genes (Table 1). The function of most of them has not been studied, except for a few genes [11–13]. Here we report expression analysis and functional studies by transgenics of several rice HDAC genes. Our data sug- gests that individual rice HDAC genes have specific development functions that may be divergent from the Arabidopsis homologs. Materials and methods Plant materials. The rice variety Zhonghua11 (Oryza sativa spp. japonica) and Minghui63 (Oryza sativa spp. indica) were used for transgenic plant production in this study. Expression profile analysis of rice HDAC genes. The data of 33 rice tissues or organs were extracted from the CREP database (http:// crep.ncpgr.cn) for HDAC expression analysis. For HDAC responses to various abiotic stress, the data were extracted from GEO data- base (GSE6901, http://www.ncbi.nlm.nih.gov/geo) [14]. Expression values corresponding to each gene were analysed using the Cluster 3.0 program, and visualized by the Treeview program [15]. Construction of binary vector. To generate the RNAi constructs, the cDNA fragments were amplified with gene-specific primers 0006-291X/$ - see front matter Ó 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.bbrc.2009.07.162 Abbreviations: HDAC, histone deacetylase; HAT, histone acetyltransferase; GFP, green fluorescence protein; amiRNA, artificial microRNA * Corresponding author. Address: Institut de biotechnologie des Plantes, Univer- sité Paris sud 11, 91405 Orsay, France. Fax: +33 1 96153324. E-mail address: [email protected] (D.-X. Zhou). Biochemical and Biophysical Research Communications 388 (2009) 266–271 Contents lists available at ScienceDirect Biochemical and Biophysical Research Communications journal homepage: www.elsevier.com/locate/ybbrc
Transcript of Rice histone deacetylase genes display specific expression patterns and developmental functions

Biochemical and Biophysical Research Communications 388 (2009) 266–271
Contents lists available at ScienceDirect
Biochemical and Biophysical Research Communications
journal homepage: www.elsevier .com/locate /ybbrc
Rice histone deacetylase genes display specific expression patternsand developmental functions
Yongfeng Hu a, Fujun Qin a, Limin Huang a, Qianwen Sun a, Chen Li a, Yu Zhao a, Dao-Xiu Zhou a,b,*
a National Key Laboratory for Crop Genetic Improvement, Huazhong Agricultural University, 430070 Wuhan, Chinab Institut de Biotechnologie des Plantes, Université Paris sud 11, 91405 Orsay, France
a r t i c l e i n f o
Article history:Received 28 July 2009Available online 5 August 2009
Keywords:ChromatinEpigenetic modificationHistone acetylationAcetyltransferaseRice development
0006-291X/$ - see front matter � 2009 Elsevier Inc. Adoi:10.1016/j.bbrc.2009.07.162
Abbreviations: HDAC, histone deacetylase; HAT, hisgreen fluorescence protein; amiRNA, artificial microR
* Corresponding author. Address: Institut de biotecsité Paris sud 11, 91405 Orsay, France. Fax: +33 1 961
E-mail address: [email protected] (D.-X. Zho
a b s t r a c t
Histone deacetylases (HDAC) are important in plant gene expression. Here we show that the expressionof rice HDAC genes is both tissue/organ-specific, and most of them are responsive to drought or salt stres-ses. Over-expression of several rice HDACs did not produce any visible phenotype, whereas down-regu-lation of a few HDAC genes affected different developmental aspects. Specifically, down-regulation ofHDA703 by amiRNA reduced rice peduncle elongation and fertility, while inactivation of a closely relatedhomolog HDA710 by RNAi affected vegetative growth. HDA704 RNAi altered plant height and flag leafmorphology. Down-regulation of HDT702 led to the production of narrowed leaves and stems. These datasuggest that rice HDAC genes may have divergent developmental functions compared with closelyrelated homologs in Arabidopsis.
� 2009 Elsevier Inc. All rights reserved.
Histone modifications including acetylation, methylation, phos- shown to play an important role in plant growth, as down-regulation
phorylation, ubiquitination and others are shown to play an impor-tant role in chromatin function and gene regulation. Specificpatterns of histone modifications establish the so-called ‘‘histonecode” that determines open or repressed states of the cognateDNA sequences. Histone acetylation/deacetylation appears as akey switch for inter-conversion between permissive and repressivestates of chromatin domains for gene expression. The homeostaticbalance of nucleosomal histone acetylation is maintained by antag-onistic action of histone acetyltransferases (HAT) and histonedeacetylases (HDAC). Plant HDAC can be grouped into four classes.Among them, three have primary homology to three yeast HDACgroups: RDP3, HDA1 and SIR2 [1]. The fourth group known as theHD2 class is found in plants only [2].During the last years, HDAC function has been most studied inArabidopsis. Inactivation of Arabidopsis HD1 (AtHD1/HDA19) byexpressing an antisense construct and by a T-DNA insertion mutation(athd1-t1) induces pleiotropic developmental abnormalities [3,4]. Inaddition, AtHD1/HDA19 is shown to form an antagonistic couplewith the histone acetyltransferase GCN5 involved in the regulationof several pathways including photomorphogenic seedling develop-ment [5]. Recent data show that AtHD1/HDA19 is involved in expres-sion of genes responding to jasmonic acid and ethylene signaling [6].Two other Arabidopsis RPD3 type genes AtRPD3A and AtRPD3B are
ll rights reserved.
tone acetyltransferase; GFP,NAhnologie des Plantes, Univer-53324.u).
of the genes delays flowering [7]. Treatment of wild type seedlingswith the specific HDAC inhibitor, Trichostatin-A (TSA), and mutationsin HDA18 induce ectopic production of root hairs [8]. Furthermore,HDAC genes are found to be involved in epigenetic regulation, as AtH-DA6 is shown to be involved in RNA interference-mediated DNAmethylation [9]. In addition, AtHDA6 and HDT2 are involved innucleolar dominance in allopolyploid hybrids [10].
Rice genome contains at least 19 HDAC genes (Table 1). Thefunction of most of them has not been studied, except for a fewgenes [11–13]. Here we report expression analysis and functionalstudies by transgenics of several rice HDAC genes. Our data sug-gests that individual rice HDAC genes have specific developmentfunctions that may be divergent from the Arabidopsis homologs.
Materials and methods
Plant materials. The rice variety Zhonghua11 (Oryza sativa spp.japonica) and Minghui63 (Oryza sativa spp. indica) were used fortransgenic plant production in this study.
Expression profile analysis of rice HDAC genes. The data of 33 ricetissues or organs were extracted from the CREP database (http://crep.ncpgr.cn) for HDAC expression analysis. For HDAC responsesto various abiotic stress, the data were extracted from GEO data-base (GSE6901, http://www.ncbi.nlm.nih.gov/geo) [14]. Expressionvalues corresponding to each gene were analysed using the Cluster3.0 program, and visualized by the Treeview program [15].
Construction of binary vector. To generate the RNAi constructs,the cDNA fragments were amplified with gene-specific primers

Table 1Rice HDAC genes studied (+) or not studied (�) by transgenics in this work.
Gene Loc. No. GenBank Over-expression RNAi AmiRNA
HDA701 LOC_Os01g40400 NM_192536 � �HDA702 (OsHDAC1) LOC_Os06g38470 AK068051 �a +b +HDA703 (OsHDAC3) LOC_Os02g12350 AK120027 �a � +HDA704 LOC_Os07g06980 AK111557 + +HDA705 LOC_Os08g25570 AK111861 � �HDA706 (OsHDAC6) LOC_Os06g37420 AK100812 +c +HDA707 LOC_Os01g12310 NM_188519 � �HDA709 LOC_Os11g09370 AK064282 + +HDA710 (OsHDAC2) LOC_Os02g12380 AK103097 �a � +HDA711 LOC_Os04g33480 AK066667 + +HDA712 LOC_Os05g36920 AK102786 + +HDA713 LOC_Os07g41090 AK111892 � �HDA714 (OsHDAC10) LOC_Os12g08220 AK072557 �e �HDA716 LOC_Os05g36930 AK068179 � �SRT701 (OsSRT1) LOC_Os04g20270 AK060337 +f +f
SRT702 (OsSir2b) LOC_Os12g07950 AK067069 +d +HDT701 LOC_Os05g51830 AK072845 + +HDT702 LOC_Os01g68104 XM_463594 + +
a Over-expression of these genes is studied in Jang et al. [13].b Knock-out of this gene is studied in Chung et al. [11].c Localized in chloroplasts [22].d localized in mitochondria [22].e Localized in both chloroplasts and mitochondria [22].f Studied in Huang et al. [12].
Y. Hu et al. / Biochemical and Biophysical Research Communications 388 (2009) 266–271 267
and then sequentially cloned into the vector pDS1301 with KpnI/BamHI and SacI/SpeI [16]. For construction of over-expression vec-tors, HDAC full-length cDNA were amplified by RT-PCR. The PCRproducts (digested by KpnI/BamHI) were inserted into the binaryvector pU1301, which was modified based on pCAMIA1301 (CAM-BIA) and contains a maize ubiquitin promoter [12]. The HDAC1-3artificial microRNAs (amiHDAC1-3) were constructed as described[17]. AmiHDAC1-3 sequences (50-TTCCATGTTACTTGCTGCAGC,50-TGTTTCAGGAATTCGCTCTGC, 50-TTGATCTTCATCTGGCTCACG)were engineered into the miR319a precursor in vector RS300 bysite-directed mutagenesis and the resulting artificial microRNAswere cloned into pU1301 (digested by KpnI/BamHI)
Reverse transcription and real time PCR. For gene expression anal-ysis, total RNA isolation and RT-PCR were performed as describedpreviously [18]. Real time PCR analysis was performed usinggene-specific primers and SYBR Premix Ex TaqTM (Takara, Dalian)on a real time PCR 7500 system (Applied Biosystems). The riceactin gene was used as internal control. The relative expressionsof each gene in transgenic plants and wild type were calculatedusing the 2�DDCt methods as described [19].
RNA gel blot analysis. Fifteen micrograms of total RNA was trans-ferred onto the Nylon membrane (Hybond-XL, Amersham). Genespecific-probes were labeled with 32P-dCTP and hybridized to theRNA blots. The fragments for probes were digested from theover-expression vectors with appropriate restriction enzymes.
Protein gel blot analysis. Histone proteins were extracted from thewild type and transgenic rice leaves (2 g) with the buffer as described[20]. The histone pellet was separated by SDS–PAGE, detected withthe acetylated H4 antibody purchased from Millipore, and peroxi-dase (HRP)-conjugated goat anti-rabbit IgG as secondary antibody.
Light microscopy. Flag leaves were harvested from matureplants. The procedures of dehydration, clearing, infiltration andembedding were carried out as described [21]. The microtome sec-tions (10 lm) were mounted on glass slides for imaging.
Results and discussion
Expression profiles of rice HDAC genes
To study the expression profiles of rice HDAC genes, weanalyzed the Collection of Rice Expression Profiles (CREP) that is
established by a serial Affimetrix microarray analysis of transcriptsfrom various organs at different developmental stages (http://crep.ncpgr.cn). Several genes were found to have a specific expressionpattern. For instance, HDA710 was more expressed in germinatingand young seedlings as well as in stamens, whereas HDA703 washighly expressed in calli and in imbibed seeds. HAD714 andHDA706 were found to be mainly expressed in shoots and leaves,whereas HDA716 showed a strong expression in developing endo-sperm and germinating seeds. The leaf/shoot specificity of HDA714is consistent with its protein localization in chloroplasts and mito-chondria [22], suggesting that this HDAC may play a role in plantspecific functions. Three groups of closely related genes (group 1:HDA705, HDA702, HDA709, HDA710 and HDA711; group 2:HDA704 and HDA713; group 3: HDT701 and HDT702) showed avery similar expression pattern with a relatively high level indeveloping panicles and calli. However, the expression patternsof the two members of the Sir2-like HDACs, SRT701 and SRT702were found to be different. The two proteins are likely to have dis-tinct functions, as SRT701 (OsSRT1) is nuclear localized and isshown to be involved in histone H3K9 deacetlyation required fortransposon repression in rice [12], whereas SRT702 is recentlyshown to be localized in the mitochondria [22].
In order to know whether rice HDAC genes are regulated by abi-otic stresses, we analyzed the microarray data obtained from ricetreated with drought, salt and cold stresses [14]. Two genes(HDA703 and HDA710) are induced, while nine others (i.e.HDA701, HDA702, HDA704, HDA705, HDA706, HDA712, HDA714,HDA716, HDT701 and HDT702) are clearly repressed by droughtand salt (Fig. 1). Two additional genes (HDA709 and SRT702) are in-duced by drought, but repressed by salt. However, cold treatmentseemed to have less impact on rice HDAC gene expression. Abscisicacid (ABA) is the major plant hormone in water stress signaling andregulates water balance and osmotic stress tolerance [23]. It hasbeen shown that ABA down-regulates the expression of AtHD2C(also known as AtHDT3) [24]. In a recent report it is shown thatHDT701, HTD702, SRT701 and SRT702 are also repressed by ABAtreatment [25]. The present analysis revealed that the expressionof most of the rice HDAC genes is responsive to drought and saltstresses, with a large proportion of the genes being repressed. Pre-vious results show that Arabidopsis HDAC (i.e. AtHD1/HAD19, AtH-DT3) are involved in stress responses [6,24]. Abiotic stresses
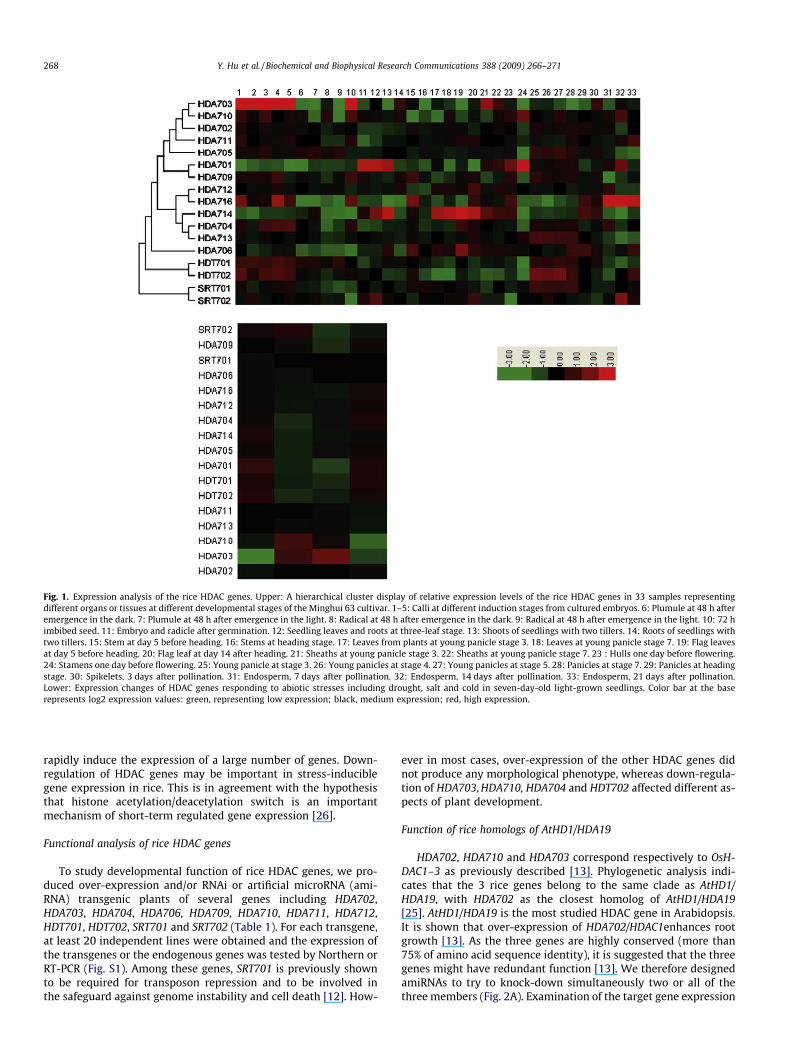
Fig. 1. Expression analysis of the rice HDAC genes. Upper: A hierarchical cluster display of relative expression levels of the rice HDAC genes in 33 samples representingdifferent organs or tissues at different developmental stages of the Minghui 63 cultivar. 1–5: Calli at different induction stages from cultured embryos. 6: Plumule at 48 h afteremergence in the dark. 7: Plumule at 48 h after emergence in the light. 8: Radical at 48 h after emergence in the dark. 9: Radical at 48 h after emergence in the light. 10: 72 himbibed seed. 11: Embryo and radicle after germination. 12: Seedling leaves and roots at three-leaf stage. 13: Shoots of seedlings with two tillers. 14: Roots of seedlings withtwo tillers. 15: Stem at day 5 before heading. 16: Stems at heading stage. 17: Leaves from plants at young panicle stage 3. 18: Leaves at young panicle stage 7. 19: Flag leavesat day 5 before heading. 20: Flag leaf at day 14 after heading. 21: Sheaths at young panicle stage 3. 22: Sheaths at young panicle stage 7. 23 : Hulls one day before flowering.24: Stamens one day before flowering. 25: Young panicle at stage 3. 26: Young panicles at stage 4. 27: Young panicles at stage 5. 28: Panicles at stage 7. 29: Panicles at headingstage. 30: Spikelets, 3 days after pollination. 31: Endosperm, 7 days after pollination. 32: Endosperm, 14 days after pollination. 33: Endosperm, 21 days after pollination.Lower: Expression changes of HDAC genes responding to abiotic stresses including drought, salt and cold in seven-day-old light-grown seedlings. Color bar at the baserepresents log2 expression values: green, representing low expression; black, medium expression; red, high expression.
268 Y. Hu et al. / Biochemical and Biophysical Research Communications 388 (2009) 266–271
rapidly induce the expression of a large number of genes. Down-regulation of HDAC genes may be important in stress-induciblegene expression in rice. This is in agreement with the hypothesisthat histone acetylation/deacetylation switch is an importantmechanism of short-term regulated gene expression [26].
Functional analysis of rice HDAC genes
To study developmental function of rice HDAC genes, we pro-duced over-expression and/or RNAi or artificial microRNA (ami-RNA) transgenic plants of several genes including HDA702,HDA703, HDA704, HDA706, HDA709, HDA710, HDA711, HDA712,HDT701, HDT702, SRT701 and SRT702 (Table 1). For each transgene,at least 20 independent lines were obtained and the expression ofthe transgenes or the endogenous genes was tested by Northern orRT-PCR (Fig. S1). Among these genes, SRT701 is previously shownto be required for transposon repression and to be involved inthe safeguard against genome instability and cell death [12]. How-
ever in most cases, over-expression of the other HDAC genes didnot produce any morphological phenotype, whereas down-regula-tion of HDA703, HDA710, HDA704 and HDT702 affected different as-pects of plant development.
Function of rice homologs of AtHD1/HDA19
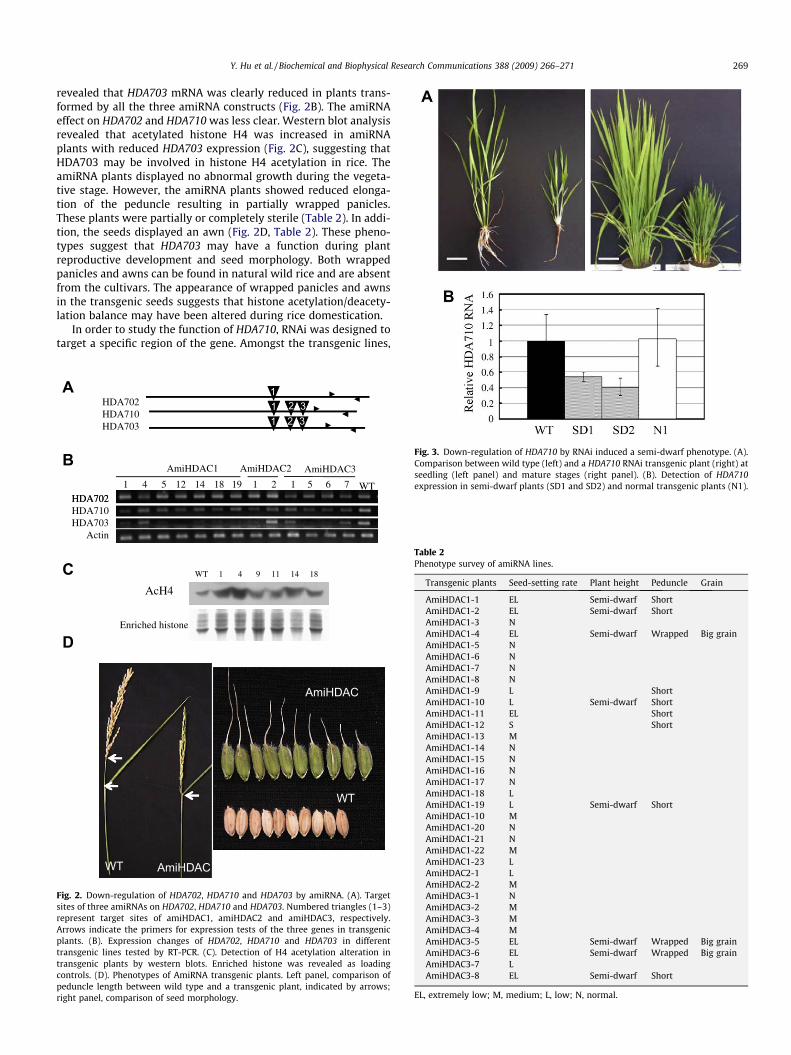
HDA702, HDA710 and HDA703 correspond respectively to OsH-DAC1–3 as previously described [13]. Phylogenetic analysis indi-cates that the 3 rice genes belong to the same clade as AtHD1/HDA19, with HDA702 as the closest homolog of AtHD1/HDA19[25]. AtHD1/HDA19 is the most studied HDAC gene in Arabidopsis.It is shown that over-expression of HDA702/HDAC1enhances rootgrowth [13]. As the three genes are highly conserved (more than75% of amino acid sequence identity), it is suggested that the threegenes might have redundant function [13]. We therefore designedamiRNAs to try to knock-down simultaneously two or all of thethree members (Fig. 2A). Examination of the target gene expression
Y. Hu et al. / Biochemical and Biophysical Research Communications 388 (2009) 266–271 269
revealed that HDA703 mRNA was clearly reduced in plants trans-formed by all the three amiRNA constructs (Fig. 2B). The amiRNAeffect on HDA702 and HDA710 was less clear. Western blot analysisrevealed that acetylated histone H4 was increased in amiRNAplants with reduced HDA703 expression (Fig. 2C), suggesting thatHDA703 may be involved in histone H4 acetylation in rice. TheamiRNA plants displayed no abnormal growth during the vegeta-tive stage. However, the amiRNA plants showed reduced elonga-tion of the peduncle resulting in partially wrapped panicles.These plants were partially or completely sterile (Table 2). In addi-tion, the seeds displayed an awn (Fig. 2D, Table 2). These pheno-types suggest that HDA703 may have a function during plantreproductive development and seed morphology. Both wrappedpanicles and awns can be found in natural wild rice and are absentfrom the cultivars. The appearance of wrapped panicles and awnsin the transgenic seeds suggests that histone acetylation/deacety-lation balance may have been altered during rice domestication.
In order to study the function of HDA710, RNAi was designed totarget a specific region of the gene. Amongst the transgenic lines,
HDA702HDA710
121 2 23
A
AmiHDAC1 AmiHDAC2 AmiHDAC3
WTHDA702
HDA710HDA703
1 4 5 12 14 118 19 2 1 5 6 7
211
22
233
B
HDA702HDA710HDA703
Actin
WT 1 4 9 11 14 18CAcH4
Enriched histone
D
AmiHDAC
WT
AmiHDACWT
Fig. 2. Down-regulation of HDA702, HDA710 and HDA703 by amiRNA. (A). Targetsites of three amiRNAs on HDA702, HDA710 and HDA703. Numbered triangles (1–3)represent target sites of amiHDAC1, amiHDAC2 and amiHDAC3, respectively.Arrows indicate the primers for expression tests of the three genes in transgenicplants. (B). Expression changes of HDA702, HDA710 and HDA703 in differenttransgenic lines tested by RT-PCR. (C). Detection of H4 acetylation alteration intransgenic plants by western blots. Enriched histone was revealed as loadingcontrols. (D). Phenotypes of AmiRNA transgenic plants. Left panel, comparison ofpeduncle length between wild type and a transgenic plant, indicated by arrows;right panel, comparison of seed morphology.
Fig. 3. Down-regulation of HDA710 by RNAi induced a semi-dwarf phenotype. (A).Comparison between wild type (left) and a HDA710 RNAi transgenic plant (right) atseedling (left panel) and mature stages (right panel). (B). Detection of HDA710expression in semi-dwarf plants (SD1 and SD2) and normal transgenic plants (N1).
Table 2Phenotype survey of amiRNA lines.
Transgenic plants Seed-setting rate Plant height Peduncle Grain
AmiHDAC1-1 EL Semi-dwarf ShortAmiHDAC1-2 EL Semi-dwarf ShortAmiHDAC1-3 NAmiHDAC1-4 EL Semi-dwarf Wrapped Big grainAmiHDAC1-5 NAmiHDAC1-6 NAmiHDAC1-7 NAmiHDAC1-8 NAmiHDAC1-9 L ShortAmiHDAC1-10 L Semi-dwarf ShortAmiHDAC1-11 EL ShortAmiHDAC1-12 S ShortAmiHDAC1-13 MAmiHDAC1-14 NAmiHDAC1-15 NAmiHDAC1-16 NAmiHDAC1-17 NAmiHDAC1-18 LAmiHDAC1-19 L Semi-dwarf ShortAmiHDAC1-10 MAmiHDAC1-20 NAmiHDAC1-21 NAmiHDAC1-22 MAmiHDAC1-23 LAmiHDAC2-1 LAmiHDAC2-2 MAmiHDAC3-1 NAmiHDAC3-2 MAmiHDAC3-3 MAmiHDAC3-4 MAmiHDAC3-5 EL Semi-dwarf Wrapped Big grainAmiHDAC3-6 EL Semi-dwarf Wrapped Big grainAmiHDAC3-7 LAmiHDAC3-8 EL Semi-dwarf Short
EL, extremely low; M, medium; L, low; N, normal.
270 Y. Hu et al. / Biochemical and Biophysical Research Communications 388 (2009) 266–271
five showed reduced vegetative growth (Fig. 3). Among the threeclosely related genes, HDA702 and HDA710 showed a very similarexpression pattern (Fig. 1). Chung et al. [11] have isolated two T-DNA insertion alleles of HDA702/HDAC1, but failed to obtain homo-zygous mutants, suggesting that the gene may be essential forplant growth. However, in the heterozygous insertion plants, rootgrowth was affected [11]. The vegetative growth of HDA710 RNAiplants was reduced, which may be due to poorly developed roots(Fig. 3). The similar expression pattern and the down-regulationphenotypes of HDA702/HDAC1 and HDA710 suggest that both genesmay play a similar but important role in root and vegetativegrowth. Furthermore, the expression levels of the two genes may
HDA704rRNA
WT 1 2 3 4 5 6 7 8 9 10 11
A
0.6
0.9
1.2
1.5
rRNA
B
0
0.3
WT R2 R3 R9 R10
C
WT R2 WT R9 R2
Da b c
d e
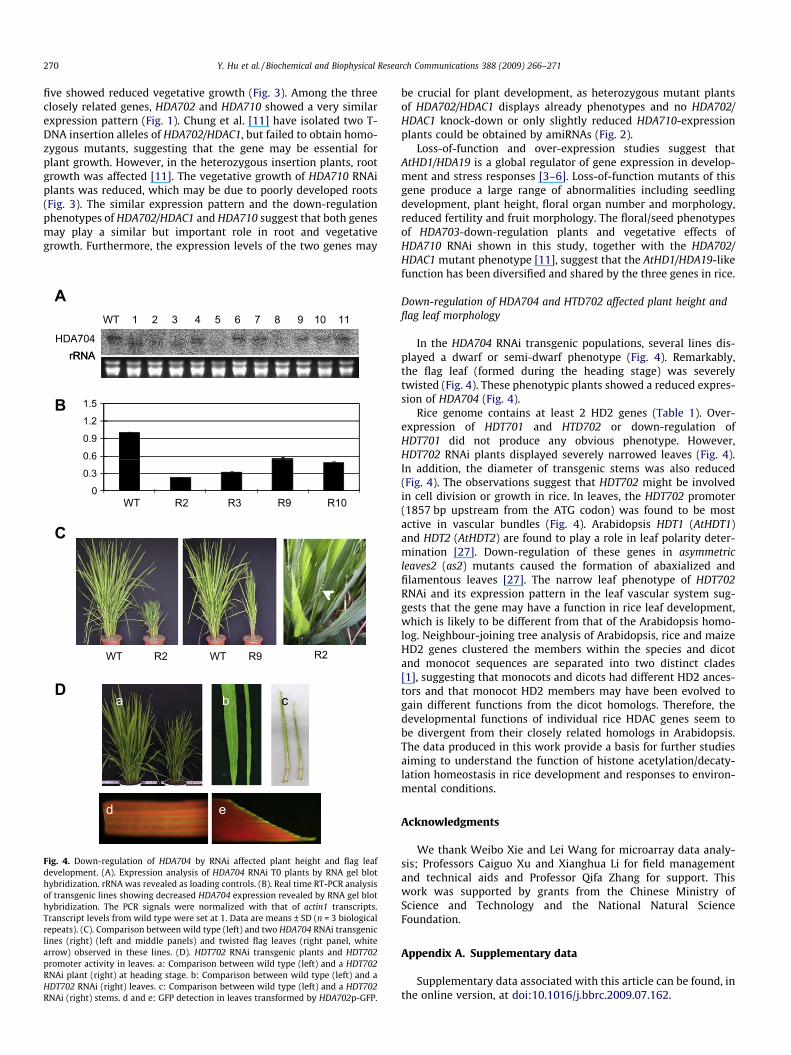
Fig. 4. Down-regulation of HDA704 by RNAi affected plant height and flag leafdevelopment. (A). Expression analysis of HDA704 RNAi T0 plants by RNA gel blothybridization. rRNA was revealed as loading controls. (B). Real time RT-PCR analysisof transgenic lines showing decreased HDA704 expression revealed by RNA gel blothybridization. The PCR signals were normalized with that of actin1 transcripts.Transcript levels from wild type were set at 1. Data are means ± SD (n = 3 biologicalrepeats). (C). Comparison between wild type (left) and two HDA704 RNAi transgeniclines (right) (left and middle panels) and twisted flag leaves (right panel, whitearrow) observed in these lines. (D). HDT702 RNAi transgenic plants and HDT702promoter activity in leaves. a: Comparison between wild type (left) and a HDT702RNAi plant (right) at heading stage. b: Comparison between wild type (left) and aHDT702 RNAi (right) leaves. c: Comparison between wild type (left) and a HDT702RNAi (right) stems. d and e: GFP detection in leaves transformed by HDA702p-GFP.
be crucial for plant development, as heterozygous mutant plantsof HDA702/HDAC1 displays already phenotypes and no HDA702/HDAC1 knock-down or only slightly reduced HDA710-expressionplants could be obtained by amiRNAs (Fig. 2).
Loss-of-function and over-expression studies suggest thatAtHD1/HDA19 is a global regulator of gene expression in develop-ment and stress responses [3–6]. Loss-of-function mutants of thisgene produce a large range of abnormalities including seedlingdevelopment, plant height, floral organ number and morphology,reduced fertility and fruit morphology. The floral/seed phenotypesof HDA703-down-regulation plants and vegetative effects ofHDA710 RNAi shown in this study, together with the HDA702/HDAC1 mutant phenotype [11], suggest that the AtHD1/HDA19-likefunction has been diversified and shared by the three genes in rice.
Down-regulation of HDA704 and HTD702 affected plant height andflag leaf morphology
In the HDA704 RNAi transgenic populations, several lines dis-played a dwarf or semi-dwarf phenotype (Fig. 4). Remarkably,the flag leaf (formed during the heading stage) was severelytwisted (Fig. 4). These phenotypic plants showed a reduced expres-sion of HDA704 (Fig. 4).
Rice genome contains at least 2 HD2 genes (Table 1). Over-expression of HDT701 and HTD702 or down-regulation ofHDT701 did not produce any obvious phenotype. However,HDT702 RNAi plants displayed severely narrowed leaves (Fig. 4).In addition, the diameter of transgenic stems was also reduced(Fig. 4). The observations suggest that HDT702 might be involvedin cell division or growth in rice. In leaves, the HDT702 promoter(1857 bp upstream from the ATG codon) was found to be mostactive in vascular bundles (Fig. 4). Arabidopsis HDT1 (AtHDT1)and HDT2 (AtHDT2) are found to play a role in leaf polarity deter-mination [27]. Down-regulation of these genes in asymmetricleaves2 (as2) mutants caused the formation of abaxialized andfilamentous leaves [27]. The narrow leaf phenotype of HDT702RNAi and its expression pattern in the leaf vascular system sug-gests that the gene may have a function in rice leaf development,which is likely to be different from that of the Arabidopsis homo-log. Neighbour-joining tree analysis of Arabidopsis, rice and maizeHD2 genes clustered the members within the species and dicotand monocot sequences are separated into two distinct clades[1], suggesting that monocots and dicots had different HD2 ances-tors and that monocot HD2 members may have been evolved togain different functions from the dicot homologs. Therefore, thedevelopmental functions of individual rice HDAC genes seem tobe divergent from their closely related homologs in Arabidopsis.The data produced in this work provide a basis for further studiesaiming to understand the function of histone acetylation/decaty-lation homeostasis in rice development and responses to environ-mental conditions.
Acknowledgments
We thank Weibo Xie and Lei Wang for microarray data analy-sis; Professors Caiguo Xu and Xianghua Li for field managementand technical aids and Professor Qifa Zhang for support. Thiswork was supported by grants from the Chinese Ministry ofScience and Technology and the National Natural ScienceFoundation.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.bbrc.2009.07.162.

Y. Hu et al. / Biochemical and Biophysical Research Communications 388 (2009) 266–271 271
References
[1] R. Pandey, A. Muller, C.A. Napoli, D.A. Selinger, C.S. Pikaard, E.J. Richards,J. Bender, D.W. Mount, R.A. Jorgensen, Analysis of histoneacetyltransferase and histone deacetylase families of Arabidopsis thalianasuggests functional diversification of chromatin modification amongmulticellular eukaryotes, Nucleic Acids Res. 30 (2002) 5036–5055.
[2] A. Lusser, G. Brosch, A. Loidl, H. Haas, P. Loidl, Identification of maize histonedeacetylase HD2 as an acidic nucleolar phosphoprotein, Science 277 (1997)88–91.
[3] L. Tian, Z.J. Chen, Blocking histone deacetylation in Arabidopsis inducespleiotropic effects on plant gene regulation and development, Proc. Natl. Acad.Sci. USA 98 (2001) 200–205.
[4] L. Tian, M.P. Fong, J.J. Wang, N.E. Wei, H. Jiang, R.W. Doerge, Z.J. Chen,Reversible histone acetylation and deacetylation mediate genome-wide,promoter-dependent and locus-specific changes in gene expression duringplant development, Genetics 169 (2005) 337–345.
[5] M. Benhamed, C. Bertrand, C. Servet, D.X. Zhou, Arabidopsis GCN5, HD1, andTAF1/HAF2 interact to regulate histone acetylation required for light-responsive gene expression, Plant Cell 18 (2006) 2893–2903.
[6] C. Zhou, L. Zhang, J. Duan, B. Miki, K. Wu, HISTONE DEACETYLASE19 is involvedin jasmonic acid and ethylene signaling of pathogen response in Arabidopsis,Plant Cell 17 (2005) 1196–1204.
[7] K. Wu, K. Malik, L. Tian, D. Brown, B. Miki, Functional analysis of a RPD3histone deacetylase homologue in Arabidopsis thaliana, Plant Mol. Biol. 44(2000) 167–176.
[8] C.R. Xu, C. Liu, Y.L. Wang, L.C. Li, W.Q. Chen, Z.H. Xu, S.N. Bai, Histoneacetylation affects expression of cellular patterning genes in the Arabidopsisroot epidermis, Proc. Natl. Acad. Sci. USA 102 (2005) 14469–14474.
[9] W. Aufsatz, M.F. Mette, J. van der Winden, M. Matzke, A.J. Matzke, HDA6, aputative histone deacetylase needed to enhance DNA methylation induced bydouble-stranded RNA, EMBO J. 21 (2002) 6832–6841.
[10] O. Pontes, R.J. Lawrence, M. Silva, S. Preuss, P. Costa-Nunes, K. Earley, N. Neves,W. Viegas, C.S. Pikaard, Postembryonic establishment of megabase-scale genesilencing in nucleolar dominance, PLoS One 2 (2007) e1157.
[11] P.J. Chung, Y.S. Kim, J.S. Jeong, S.H. Park, B.H. Nahm, J.K. Kim, The histonedeacetylase OsHDAC1 epigenetically regulates the OsNAC6 gene that controlsseedling root growth in rice, Plant J. (2009).
[12] L. Huang, Q. Sun, F. Qin, C. Li, Y. Zhao, D.X. Zhou, Down-regulation of a SILENTINFORMATION REGULATOR2-related histone deacetylase gene, OsSRT1,induces DNA fragmentation and cell death in rice, Plant Physiol. 144 (2007)1508–1519.
[13] I.C. Jang, Y.M. Pahk, S.I. Song, H.J. Kwon, B.H. Nahm, J.K. Kim, Structure andexpression of the rice class-I type histone deacetylase genes OsHDAC1–3:
OsHDAC1 overexpression in transgenic plants leads to increased growth rateand altered architecture, Plant J. 33 (2003) 531–541.
[14] M. Jain, A. Nijhawan, R. Arora, P. Agarwal, S. Ray, P. Sharma, S. Kapoor, A.K.Tyagi, J.P. Khurana, F-box proteins in rice, genome-wide analysis,classification, temporal and spatial gene expression during panicle and seeddevelopment, and regulation by light and abiotic stress, Plant Physiol. 143(2007) 1467–1483.
[15] M.B. Eisen, P.T. Spellman, P.O. Brown, D. Botstein, Cluster analysis and displayof genome-wide expression patterns, Proc. Natl. Acad. Sci. USA 95 (1998)14863–14868.
[16] M. Dai, Y. Hu, Y. Zhao, H. Liu, D.X. Zhou, A WUSCHEL-LIKE HOMEOBOX generepresses a YABBY gene expression required for rice leaf development, PlantPhysiol. 144 (2007) 380–390.
[17] R. Schwab, S. Ossowski, M. Riester, N. Warthmann, D. Weigel, Highly specificgene silencing by artificial microRNAs in Arabidopsis, Plant Cell 18 (2006)1121–1133.
[18] Y. Zhao, Y. Hu, M. Dai, L. Huang, D.X. Zhou, The WUSCHEL-related homeoboxgene WOX11 is required to activate shoot-borne crown root development inrice, Plant Cell 21 (2009) 736–748.
[19] K.J. Livak, T.D. Schmittgen, Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method, Methods 25 (2001)402–408.
[20] M. Tariq, H. Saze, A.V. Probst, J. Lichota, Y. Habu, J. Paszkowski, Erasure of CpGmethylation in Arabidopsis alters patterns of histone H3 methylation inheterochromatin, Proc. Natl. Acad. Sci. USA 100 (2003) 8823–8827.
[21] M. Dai, Y. Hu, Q. Ma, Y. Zhao, D.X. Zhou, Functional analysis of riceHOMEOBOX4 (Oshox4) gene reveals a negative function in gibberellinresponses, Plant Mol. Biol. 66 (2008) 289–301.
[22] P.J. Chung, Y.S. Kim, S.H. Park, B.H. Nahm, J.K. Kim, Subcellular localizationof rice histone deacetylases in organelles, FEBS Lett. 583 (2009) 2249–2254.
[23] L. Xiong, K.S. Schumaker, J.K. Zhu, Cell signaling during cold, drought, and saltstress, Plant Cell 14 (Suppl.) (2002) S165–S183.
[24] S. Sridha, K. Wu, Identification of AtHD2C as a novel regulator of abscisic acidresponses in Arabidopsis, Plant J. 46 (2006) 124–133.
[25] W. Fu, K. Wu, J. Duan, Sequence and expression analysis of histonedeacetylases in rice, Biochem. Biophys. Res. Commun. 356 (2007) 843–850.
[26] D.X. Zhou, Regulatory mechanism of histone epigenetic modifications inplants, Epigenetics 4 (2009) 15–18.
[27] Y. Ueno, T. Ishikawa, K. Watanabe, S. Terakura, H. Iwakawa, K. Okada,C. Machida, Y. Machida, Histone deacetylases and ASYMMETRIC LEAVES2are involved in the establishment of polarity in leaves of Arabidopsis,Plant Cell 19 (2007) 445–457.




















