
Apoptosis-Inducing Galactolipids from a Cultured Marine Diatom, Phaeodactylum tricornutum

Resting stages and ecology of the planktonic diatom Aulacoseira skvortzowii in Lake
Baikal
David H. Jewson1
Freshwater Laboratory, University of Ulster, Traad Point, Ballyronan, Co. Derry BT45 6LR, United Kingdom
Nick G. Granin, Andre A. Zhdanov, Ludmila A. Gorbunova, Nina A. Bondarenko andRuslan Yu. GnatovskyLimnology Institute, Russian Academy of Sciences Siberian Division, 6644033, Irkutsk, P.O. Box 4199, Russia
Abstract
Populations of the planktonic diatom Aulacoseira skvortzowii in Lake Baikal developed below 4uC, withmortality increasing rapidly at temperatures above 6.5uC. Resting spores were produced before the temperaturerise associated with summer stratification. The main cue for sporulation was a decline in phosphate concentrationbelow 15–20 mg L21 P-PO4. If phosphate declined after the onset of stratification, sporulation was poor. Inculture, all cells sporulated when phosphate limited but only 15% did so when nitrate limited. Also, sporeformation was diameter dependent, with most narrow cells switching to size regeneration. This affectedpopulation dynamics, with high biomasses developing in the south and middle basins but only rarely in the northbasin, because phosphate did not always fall below the induction threshold necessary for sporulation and sizeregeneration, leading to poor recruitment. In culture, germination occurred when spores were placed in newmedia, with stored reserves sufficient to complete two to three divisions, even in the dark. This helped populationsre-establish when resuspended by wave action from coastal sediments where they lay dormant during summer.
Lake Baikal is the world’s oldest and deepest lake andone of the few places where diatom resting spores are foundin an open lake environment (Edlund et al. 1996; Likhosh-way et al. 2004). Spores were first reported in the lake 70 yrago by Skvortzow (1937). Later, Skabichevsky (1953, 1960)gave a detailed description of their formation but thetaxonomy of the species was uncertain until Edlund et al.(1996) proposed that Lake Baikal’s spore-forming popula-tion was morphologically distinct from the widely distrib-uted Aulacoseira (Melosira) islandica (O. Mull.) Simonsenand should be considered a separate species, which theynamed Aulacoseira skvortzowii Edlund, Stoermer & Taylor.Recent molecular studies have suggested that A. skvortzowiiand another Aulacoseira species occurring in Lake Baikal,Aulacoseira baicalensis (Meyer) Wislouch (Babanazarova etal. 1996; Popovskaya et al. 2002), are the closest amongliving forms to extinct taxa of the genus and may have
separated from each other within the lifetime of the lake(i.e., less than 25 million yr) (Sherbakova et al. 1998; Edgarand Theriot 2004). However, this does not necessarily meanthey are endemic, because spore-forming species similar toA. skvortzowii have been discovered in other Asian sites,including high mountain lakes (Genkal and Bondarenko2001, 2004), Lake Khanka/Xingkai (Genkal and Schure2000), and River Amur (Likhoshway et al. 2004). As aresult, Popovskaya et al. (2002) concluded that thesystematic status of this complex needs additional research.Spores are also found in fossil deposits (Edlund et al. 1996;Likhoshway et al. 2004), where their apparent greaterabundance may be an artifact due to the better preservationof their thicker cell wall. In extant diatom populations,spores have been found in several other Aulacoseira speciesand some other genera but they are generally relatively rarein freshwaters (Edlund et al. 1996). The commonestoccurrence of diatom resting spores is in temperate marineneritic habitats (McQuoid and Hobson 1996; Kuwata andTakahashi 1999; McQuoid and Godhe 2004) and one of theaims of this study was to investigate whether sporeformation occurred in a similar niche in Lake Baikal.
Studies of Lake Baikal phytoplankton populationsstretch back over a century, with the foundations ofseasonal change laid down by Skabichevsky (1929) andYasnitsky (1930). Other early studies of A. skvortzowiiand A. islandica in Russian lakes have been reviewed bySkabichevsky (1960) and Lund (1966). After 1946, system-atic long-term studies of phytoplankton populations werestarted (see Anitpova in Kozhov 1963; Bondarenko 1999;Popovskaya 2000) that found large interannual variationsin cell numbers of A. skvortzowii and A. baicalensis. Otherrecent studies have also looked at spatial variation ofphytoplankton pigments (Fietz et al. 2005; Heim et al.
1 To whom correspondence should be addressed. Presentaddress: 96 Desertmartin Road, Magherafelt, Co. Derry BT455HE, United Kingdom ([email protected]).
AcknowledgmentsWe thank staff at the Limnology Institute, especially M. A.
Grachev, Ye. Likhoshway, N. Guselnikova, M. N. Shimaraev, V.Tsekhanovsky, A. Yakushin, L. Olbolkina, V. A. Olbolkin, O.Molozhavaya, L. Levin, A. Kuzmina, N. Melnik, L. Dor-oschenko, and the crews of the Verischagin and Titov for theirhelp. We are grateful to anonymous reviewers for helpfulcomments and to S. F. Lowry, M. Sturm, R. Kipfer, A. Mackay,R. Flower, D. Ryves, B. Goddeeris, and R. W. Battarbee forassistance during this work.
We also thank The Royal Society, the Russian Academy ofSciences Siberian Division, NATO, INTAS and NERC, who haveall supported parts of this work.
Limnol. Oceanogr., 53(3), 2008, 1125–1136
E 2008, by the American Society of Limnology and Oceanography, Inc.
1125
2005) and species (Kozhova 1987; Bondarenko et al. 1996),including the distribution of cells near thermal bars(Likhoshway et al. 1996).
An important step toward gaining a fuller understandingof diatom population dynamics, reducing uncertainty overtaxonomy, and investigating speciation requires a betterknowledge of the function and adaptation of morpholog-ical features and how they can lead to niche separation. Todo this, we need to know more about the underlyingecological pressures for selection throughout the life cycle.Therefore, information from a multidisciplinary study overan 11-yr period (1994 to 2005) was used to investigate thisin A. skvortzowii. In particular, the ways in whichreproductive and dormant stages were adapted to theenvironment in Lake Baikal, where the lake is frozen for 4to 6 months each year but the water column remainsoxygenated throughout its great depth (1,636 m) (Kozhov1963; Weiss et al. 1991; Kozhova and Izmest’eva 1998),mainly due to differences in density between layers that canlead to instability and deep water mixing (Weiss et al. 1991;Shimaraev et al. 1993; Wuest et al. 2005).
Methods
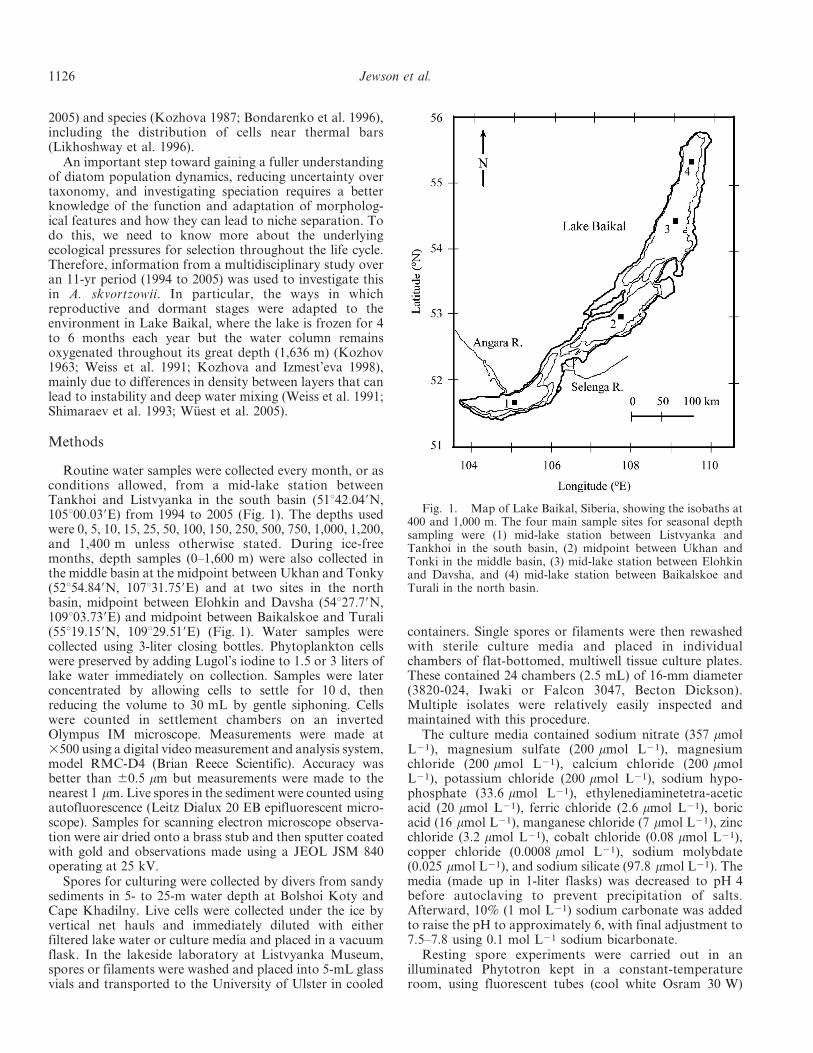
Routine water samples were collected every month, or asconditions allowed, from a mid-lake station betweenTankhoi and Listvyanka in the south basin (51u42.049N,105u00.039E) from 1994 to 2005 (Fig. 1). The depths usedwere 0, 5, 10, 15, 25, 50, 100, 150, 250, 500, 750, 1,000, 1,200,and 1,400 m unless otherwise stated. During ice-freemonths, depth samples (0–1,600 m) were also collected inthe middle basin at the midpoint between Ukhan and Tonky(52u54.849N, 107u31.759E) and at two sites in the northbasin, midpoint between Elohkin and Davsha (54u27.79N,109u03.739E) and midpoint between Baikalskoe and Turali(55u19.159N, 109u29.519E) (Fig. 1). Water samples werecollected using 3-liter closing bottles. Phytoplankton cellswere preserved by adding Lugol’s iodine to 1.5 or 3 liters oflake water immediately on collection. Samples were laterconcentrated by allowing cells to settle for 10 d, thenreducing the volume to 30 mL by gentle siphoning. Cellswere counted in settlement chambers on an invertedOlympus IM microscope. Measurements were made at3500 using a digital video measurement and analysis system,model RMC-D4 (Brian Reece Scientific). Accuracy wasbetter than 60.5 mm but measurements were made to thenearest 1 mm. Live spores in the sediment were counted usingautofluorescence (Leitz Dialux 20 EB epifluorescent micro-scope). Samples for scanning electron microscope observa-tion were air dried onto a brass stub and then sputter coatedwith gold and observations made using a JEOL JSM 840operating at 25 kV.
Spores for culturing were collected by divers from sandysediments in 5- to 25-m water depth at Bolshoi Koty andCape Khadilny. Live cells were collected under the ice byvertical net hauls and immediately diluted with eitherfiltered lake water or culture media and placed in a vacuumflask. In the lakeside laboratory at Listvyanka Museum,spores or filaments were washed and placed into 5-mL glassvials and transported to the University of Ulster in cooled
containers. Single spores or filaments were then rewashedwith sterile culture media and placed in individualchambers of flat-bottomed, multiwell tissue culture plates.These contained 24 chambers (2.5 mL) of 16-mm diameter(3820-024, Iwaki or Falcon 3047, Becton Dickson).Multiple isolates were relatively easily inspected andmaintained with this procedure.
The culture media contained sodium nitrate (357 mmolL21), magnesium sulfate (200 mmol L21), magnesiumchloride (200 mmol L21), calcium chloride (200 mmolL21), potassium chloride (200 mmol L21), sodium hypo-phosphate (33.6 mmol L21), ethylenediaminetetra-aceticacid (20 mmol L21), ferric chloride (2.6 mmol L21), boricacid (16 mmol L21), manganese chloride (7 mmol L21), zincchloride (3.2 mmol L21), cobalt chloride (0.08 mmol L21),copper chloride (0.0008 mmol L21), sodium molybdate(0.025 mmol L21), and sodium silicate (97.8 mmol L21). Themedia (made up in 1-liter flasks) was decreased to pH 4before autoclaving to prevent precipitation of salts.Afterward, 10% (1 mol L21) sodium carbonate was addedto raise the pH to approximately 6, with final adjustment to7.5–7.8 using 0.1 mol L21 sodium bicarbonate.
Resting spore experiments were carried out in anilluminated Phytotron kept in a constant-temperatureroom, using fluorescent tubes (cool white Osram 30 W)
Fig. 1. Map of Lake Baikal, Siberia, showing the isobaths at400 and 1,000 m. The four main sample sites for seasonal depthsampling were (1) mid-lake station between Listvyanka andTankhoi in the south basin, (2) midpoint between Ukhan andTonki in the middle basin, (3) mid-lake station between Elohkinand Davsha, and (4) mid-lake station between Baikalskoe andTurali in the north basin.
1126 Jewson et al.

with an irradiance of 10–15 mmol m22 s21 and 8 : 16 light :dark (LD) cycles, unless otherwise stated. Cell numbers inmicrotiter plate chambers were counted once or twice dailyon an inverted microscope (Olympus, model IM), whichminimized disturbance. The experiments on the effect oftemperature were also carried out with an illuminated,cooled incubator (Gallenkamp INF-781-T). Growth rateestimates at each new temperature were only started afterone division had been completed and then rates wereaveraged for the next three or four doublings. Replicates(15 to 25) were made at each temperature. The mortalityrates were estimated from five replicates for each temper-ature and based on visual observation of degeneration ofcell contents and failure of further division.
Irradiances were measured with Li-Cor Ll-185B and Ll-190SB quantum sensors (Li-Cor). Silicate, phosphate (error61.5%), and nitrate (error 64%) were measured by standardcolorimetric methods (Stroganov and Buzinova 1980; Wetzeland Likens 1991). Temperature profiles were made withconductivity–temperature–depth models AML-12 (AppliedMicrosystems) and SBE-25 (Sea-Bird Electronics).
Results
Morphology—A. skvortzowii usually formed short fila-ments of one to four vegetative cells (Fig. 2c) but longer
filaments up to and above eight cells also occurred. Restingspores were formed endogenously, i.e., a single spore wasformed within the parent cell. Figure 2d shows twoadjacent daughter cells where each has undergone an initialdivision to form a rounded epivalve. The other curvedhypovalve was then formed after another division(Fig. 2b,e). The majority of cell contents were withdrawninto the spore. Any remaining chloroplasts (Fig. 2d)degenerated once the cross-walls were complete (Fig. 2e).The spore then became densely packed with storageproducts (Fig. 2e).
A. skvortzowii also formed resting cells, which containedrounded plastids but showed no observable change to thecell wall or accumulation of storage products (see uppercell, Fig. 2e). This mostly occurred in cells that sank intoaphotic conditions below the convective mixing zone butwas also observed in some cells that failed to form spores(Fig. 2e, left cell).
Growth and spore formation in culture—The optimumtemperature for normal vegetative cell division in A.skvortzowii was between 3.5uC and 6.5uC in irradiances of65 mmol m22 s21 and photoperiods of 8 : 16 LD (Fig. 3).The mean doubling time of 13 isolates at this temperaturewas 2 d, but one clone achieved a sustained doubling timeof 1.7 d. At temperatures above 6.5uC, growth rates rapidlydeclined and mortality increased, with little or no growthabove 15uC (Fig. 3). Cells seemed particularly vulnerableduring division, with cells lengthening but failing to formthe cross-walls of new daughter cells. Their chloroplastsand other contents then deteriorated, so that 30% of cellsdied during division at 9.5uC and 88% at 15uC (Fig. 3).
Spores began forming in cultures within 2 d of expo-nential growth ceasing. In phosphate-limited cultures, up to72% of cells completed spore formation (both morpholog-ical changes and accumulation of storage products) within9 d. In nitrogen-limited cultures, only 15% of cells
Fig. 2. Light and scanning electron micrographs of Aulaco-seira skvortzowii. Light micrographs are all the same magnifica-tion. (a) Parent valve still attached to a spherical initial cell, (b)scanning electron micrograph of resting spore from surfacesediment at the mid-lake station in the south basin (scale10 mm), (c) normal vegetative cells, (d) formation of roundedepivalve in adjacent daughter cells, (e) two resting spores and oneresting cell, (f) germinated spore with four cells in the filament andretaining rounded epivalves. Panels a, d, and e: Mid lake station inthe south basin on 18 May 1996. Panels c and f were from cultures.
Fig. 3. The effect of temperature on growth rates andmortality of Aulacoseira skvortzowii from Lake Baikal. Verticalbars are means 6 SD. Mortality is based on those cells failing tocomplete division (see text).
Resting stages in Baikal 1127

produced spores, with formation peaking after 5 d and thenceasing. None of these accumulated storage products to thedegree seen in phosphate-limited cultures. The majority ofnitrate-limited cells looked unhealthy and soon died,although some did show slight wall thickening but didnot accumulate any reserve products.
Timing of spore production—A. skvortzowii cell concen-trations were measured at selected depth intervals down to1,400 m at a mid-lake site between Tankhoi and List-vyanka in the south basin (see site 1, Fig. 1) every month,or as conditions allowed, from 1994 to 2005. Depthchanges in concentration were also made at four othersites in the south basin for shorter periods but they showedsimilar seasonal changes, so only significant differences arenoted in the following description. Typical depth distribu-tions of cells and spores of A. skvortzowii during 1 yr (1997)are shown in Fig. 4. Ice formation started on 22 Januaryand by 13 March 1997 was 0.6 m thick, with a snowcovering that varied between 0.1 and 0.2 m. Underneaththe ice, there was convective mixing in the 0- to 5-m layer,which had a temperature of 0.05uC (Fig. 5a). Cellconcentration was then 78 3 106 cells m22 (Fig. 6). The
concentrations of cells in Fig. 6 are expressed per squaremeter to take account of changes in the depth of mixing. By13 April, cells were being mixed down to 50 m and by 22April to 72 m, when cell numbers reached 5,420 3 106 cellsm22 (Fig. 6). Ice breakup began on 30 April but mixingdepth continued to increase, so by 22 May it was 95 m andcell concentration reached a maximum of 29,200 3 106 cellsm22 (Figs. 4a, 6). Up to this point, the net populationincrease of A. skvortzowii was equivalent to a doubling timeof 7 to 8 d. However, by 22 May, there were rising losses ofcells into deeper layers (see 50 to 250 m, Fig. 4a) as thedepth of mixing increased (Fig. 5a) but convectionweakened. Even so, concentrations at 250 m were less than1% of those at the surface.
On 22 May 1977, the first spores were found in smallnumbers (3% of the population) but by 06 June nearly allcells (95%) in the mixing zone down to 100 m hadsporulated. The temperature of this layer was now 3.0uC(Fig. 5a). Two weeks later (20 June) the whole watercolumn became isothermal (3.5–3.6uC, Fig. 5a), whichmeant that convective mixing declined, so spores andremaining cells began to sink rapidly (Fig. 4b) and, becauseof the additional effect of downwelling at this site, werefound throughout the water column. In the surface layers,
Fig. 4. Depth profiles of cell concentrations of Aulacoseiraskvortzowii at mid-lake station between Listvyanka and Tankhoiin the south basin of Lake Baikal during 1997 (a) during anincreasing population 13 March, 22 April, and 22 May; and (b) adecreasing population 06 June, 20 June, 07 July, and 14 August.
Fig. 5. Temperature depth profiles of the upper 150 m at themid-lake station between Listvyanka and Tankhoi in the southbasin of Lake Baikal from March 1997 to February 1998 (a)during warming and (b) during cooling.
1128 Jewson et al.

temperatures continued to rise as summer stratificationbegan and by August, no spores were found in the upper150 m, although some were still present in deeper, coolerlayers (cf. Figs. 4, 5). However, even these had disappearedby 09 September, so no live cells or spores were found atany depth in the water column. Similar seasonal patternswere found in other years (Fig. 6), with spores and cellsabsent or very rare in offshore surface waters (0–150 m)during summer stratification.
Resting spores reappeared in coastal areas after latesummer or autumn storms. They germinated if nutrientswere available (see below) and were then dispersed offshoreby currents and gyres. However, in most years, further netpopulation increase offshore was not observed because cellswere light limited by the deep vertical mixing (over 100 m,Fig. 5b) that occurred in late autumn. Net populationincrease started again with the onset of winter stratificationand reduced vertical mixing (Fig. 5b).
Environmental cue for spore formation—In cultureexperiments (see above), reduced phosphate concentrationwas the most effective cue for spore formation. In the lake,phosphate declined to undetectable concentrations in everyyear studied (mainly due to growth of other diatoms andpicoplankton). Nitrate also declined to below detectionlimits but it did so after phosphate (Fig. 7), usually whentemperatures were already rising. In contrast, silicatedeclined only in those years with high diatom biomass,i.e., 1994, 1997, and 2000 (e.g., Fig. 7a). However, as wellas changes in concentration, an important additional factorwas the timing of nutrient decline relative to the onset of
summer stratification, which is shown by comparingdifferent conditions in 1995, 1996, and 1997. In 1995, thediatom population was very low, so there was littlereduction in silicate (Fig. 7a), but phosphate concentrationfell to 6 mg L21 in the upper 15 m by 11 May because ofgrowth of picoplankton. By 04 June, 80% of A. skvortzowiicells had sporulated (Fig. 7c). In contrast, in the following
Fig. 6. Changes in cell concentration of Aulacoseira skvort-zowii over 11 yr (1994–2004) at the mid-lake station betweenListvyanka and Tankhoi in the south basin of Lake Baikal.Results are integrated per unit area (0–250 m). (a) Vegetative cellsand (b) resting spores.
Fig. 7. Seasonal changes from 1995 to 1997 at the mid-lakestation between Listvyanka and Tankhoi in the south basin ofLake Baikal. (a) Silicate concentration and nitrate concentrationat 5 m, (b) orthophosphate and temperature at 5 m, and (c)vegetative cells and proportion of cells producing resting spores.
Resting stages in Baikal 1129

year (1996) sporulation was poor because phosphateconcentrations were still high (31 mg L21 on 26 June) bythe time stratification started (Fig. 7b) and concentrationsonly decreased to 10 mg L21 by 07 July. By then, thetemperature at the surface was 11.4uC and only 0.5% ofcells formed spores between 06 and 26 June. The finalnumber of spores represented only 1.5% of peak cellconcentrations in June. At three other sites in the southbasin there were even fewer spores produced. The followingyear, 1997, was a high biomass year for diatoms and thiscaused an early decline in silicate during late April andearly May (Fig. 7a). However, phosphate concentrationdecreased even earlier, from 25 to 7 mg L21 between 01April and 22 April (Figs. 7b, 8). The first spores were foundon 22 May (3%) and by 03 June nearly the wholepopulation (95%) had sporulated (Fig. 7c). Overall, fromfield measurements, a decline below 15–20 mg L21 P-PO4
appeared to be the likely threshold for inducing sporeformation but only if it occurred before isothermal mixing,as in 1995 and 1997. If the threshold was reached whentemperatures were rising, as in 1996, then spore productionwas poor.
Resting spores were also produced in low concentrationsunder the ice in winter. These periods coincided with timesof high phytoplankton biomass when phosphate concen-trations became temporarily depleted. A. skvortzowiipopulations were not generally large enough under the iceto affect nutrient changes themselves, but other planktonicdiatoms (A. baicalensis, Cyclotella baicalensis, Stephano-discus meyeri, Nitzschia acicularis, and Synedra acus) andpicoplankton did form dense populations that coulddeplete the nutrients. One such period occurred on 29March 2000, when there was convective mixing down to20 m. The majority of cells were in this layer, but, justunder the ice, some cells became isolated in a separatecirculation (Granin et al. 1999) where phosphate concen-trations decreased to 9 mg L21 P-PO4. Spores were thenproduced that sank through the convective layer but were
held up at the thermocline at 25 m, where they made uponly a small proportion of the population (1% or 534spores L21). The other cells did not sporulate becausephosphate was available in the convective layer.
Spatial variation between basins—Figure 9 shows theconcentration of spores and vegetative cells along themiddle of the lake between 30 May and 06 June 2000.Sporulation was complete in the south basin (90–100% ofcells) but still in progress in the middle basin (20%). In thenorth basin, populations were low (1 million cells m22) andrestricted to one small area between Elohkin and Davsha(site 3, Fig. 1). In other years, there was also considerablespatial variation in the success of sporulation. In the middlebasin, the number of cells sporulating at the time ofisothermal mixing was 0%, 6%, 20%, 99%, 43%, 90%, and62% on 11 June 1998, 03 June 1999, 31 May 2000, 20 June2001, 03 June 2002, 15 June 2003, and 20 June 2004respectively. In the north basin, no cells or spores werefound in 1998, 1999, and 2003 at any depth (0–850 m). Evenin those years when cells were present in the north basin,concentrations were low and spore production was poor.For example, by 18 July 2001, when the water column wasjust past isothermal conditions at 3.8uC (Fig. 10a), nearly amonth after the south basin (Fig. 5a), only 14% of cells hadsporulated of a total of 12 million cells m22. A few cellsremained in the top 10 m, including a few spores (79 L21),but the main crop sank into deeper water (Fig. 10b) withoutsporulating. In 2002, there were no cells or spores in thesurface layers but below 500 m some cells were found(nearly 20 3 106 cells and spores m22) that were probablythe remainder of the crop from 2001. The low sporeproduction in the north was because the phosphateconcentration remained just above the induction thresholdat 23 mg L21 P-PO4 into July. During the years of this study,the phosphate concentrations in the mixed layer (ahead ofthe onset of stratification) on 13 June 1998, 05 June 1999, 01June 2000, 17 June 2001, 06 June 2002, and 16 June 2003were 31, 20, 18, 18, 15, and 22 mg L21 P-PO4 respectively.Thus, phosphate concentrations were at or just above thethreshold for both spore and size regeneration (see below).
Fig. 8. Depth profiles of phosphate concentration (PO4-P mgL21) at the mid-lake station between Listvyanka and Tankhoi inthe south basin of Lake Baikal on 22 April and 22 May 1997.
Fig. 9. Changes in surface cell concentrations of vegetativecells and resting spores of Aulacoseira skvortzowii along themiddle of the lake from 31 May to 06 June 2000.
1130 Jewson et al.

Diameter dependence, resting, and auxospore formation—Production of resting spores was diameter dependent. Cellswith diameters at and above 10 mm nearly all producedspores in those years when the phosphate threshold wasreached ahead of stratification, but fewer narrower cells didso. For example, in 1994 only 40% of cells narrower than7 mm produced spores and in 1995 and 2004 it fell to 10%.In cultures of cells with diameters below 9 mm, motilegametes and auxospores (see Fig. 2a) were readily pro-duced when exponential growth ceased, so the narrowercells were mostly switching to size regeneration linked tosexual reproduction. In the lake, although size-regeneratedauxospores and initial cells were commonly seen, motilegametes were much more difficult to identify in preservedsamples. However, they were observed once in live fieldsamples (02 June 1996, 7 km from Lystvyanka) when thecrop had reached its spring peak (Fig. 5a). The diameterrange of oocytes in Fig. 11 was estimated from valvesremaining attached to auxospores and initial cells. Eachyear the age class lost in size regeneration was replaced by adecline in diameter of broader cells.
Size regeneration followed the decline in phosphateconcentration, so was mostly in late May and early June inthe south basin (Fig. 12). However, resting spores and size-regenerating auxospores were also found during periods oflow phosphate under the ice, e.g., 13 March 1997 and 29March 2000 at the mid-lake site and 11 April 2000 at4.5 km from Listvyanka. After size regeneration, many
Fig. 10. Depth profiles at the mid-lake station betweenElohkin and Davsha in the north basin of Lake Baikal during2001. (a) Temperature between June and October and (b) cellconcentrations of Aulacoseira skvortzowii (L21) in June and July.
Fig. 11. Relative size distribution of diameters of Aulacoseiraskvortzowii at the mid-lake station between Listvyanka andTankhoi in the south basin of Lake Baikal. (a) May/Junepopulations of 1994–1997 (3,500 measurements), (b) parent cellsattached to auxospores on 100 filaments from 04 June 1997 and 28March 2000 and 150 size-regenerated initial cells, and (c) under-icetotal population 22 April 1997 and resting spores on 20 June 1997.
Fig. 12. Seasonal change in concentrations of vegetative cellsof Aulacoseira skvortzowii; auxospores plus initial cells and restingspores in 1997 at the mid-lake station between Listvyanka andTankhoi in south basin of Lake Baikal.
Resting stages in Baikal 1131

spherical initial cells became packed with storage productssimilar to the resting stages.
Settling and live cells in sediment—At the peak of themain growth period in May and June, cells of A.skvortzowii were usually dispersed across the wholesouthern basin. Filaments and spores then sank out ofthe surface waters (Fig. 4b) and gradually disappearedfrom the water column in summer, with no live sporesfound in layers down to 250 m in offshore waters duringAugust and September. Although rates of sporulation wereoften high, many spores failed to survive and many emptyvalves were found shortly after their production. Forexample, on 05 June 2000 the number of empty spores (2.763 106 m22) exceeded live spores (2.45 3 106 m22).
Coastal sediments were investigated as possible sites forsurvival in two bottom transects during 1994. The firstsurvey looked at inshore shallow sediments in the southbasin between 0 and 20 m at Bolshoi Koty and at 25 m atnearby Cape Khadilny, where coarser sediments extendeddeeper. In the surface layer (1 cm) of sand at 5 m depth, theconcentration of live spores (i.e., with cell contents that stillautofluoresced) on 19 July 1994 was 823 mL21 of sediment.This increased to 6,700 at 10 m; 8,420 at 20 m; and 10,800at 25 m. When expressed per unit area, there were 8 millionlive spores per square meter in the surface layer of sedimentat 5 m and 108 million m22 at 25 m. At this time, therewere no spores detectable in the upper layers of the watercolumn in the south basin.
In a survey of surface layers (0–0.5 cm) of deeper,muddy sediments in the middle basin, from 200 m down tonear the deepest point in the lake at 1,642 m, duringAugust 1994, live spores were found in very low concen-trations at 200 m (120 mL21), 310 m (30 mL21), and580 m (60 mL–1) but none at any other depths (275, 810,1,185, 1,365, 1,410, or 1,600 m). The surviving spores in thedeep, muddy sediments were all of recent origin, as theirchloroplasts fluoresced brightly under autofluorescence(typical of cells of less than 1 yr, as autofluorescence fadeswith time). No live cells of other diatom species were foundin the deeper sediments.
Factors controlling germination (excystment)—Germina-tion experiments were carried out on spores collected on 21July 1994 from surface sand at 20 m depth at Bolshoi Kotyand 25 m at Cape Khadilny. The spores had been in thesediment for approximately 2 months. Initially spores wereput in irradiances of 15 to 17 mmol m22 s21 on a 12 : 12 LDcycle at 4uC but they stopped developing and broke up. Thereason for this is unknown, because division of normalvegetative cells of A. skvortzowii occurred up to 65 mmolm22 s21 on 8 : 16 LD cycles. The experiment was repeated ata lower irradiance of 12 mmol m22 s21 and all sporesgerminated. Germination began with spore lengthening.Sidewalls of new daughter valves were first laid down andthen the cross-walls formed (Fig. 1f), so the first two cellshad one curved parent valve and one normal valve. Aftertwo or three divisions (i.e., four to eight cells), the cellscontaining the original curved resting spore valves wereusually lost. In an experiment in which eight spores were
followed individually, the first began to lengthen after 4.7 dand the last one after 9.7 d (mean 6.3 d). In six of the spores,the first division (i.e., presence of a cross-wall) occurred inless than a day after lengthening began; the other two took38 h and 48 h. Growth rates over five or six subsequentdoublings were similar in all isolates at an average of 1.8 dper doubling (both in cell numbers and filament length), witha narrow range between 1.5 and 2.1 d.
To test longevity, some spores were kept in the dark (inunsealed flasks to prevent anoxia developing) at 4uC for1 yr (until 30 May 1995). By that time only 31% of sporesstill had cell contents and less than half of these could begerminated. When put in the light, the first of the viablespores started to lengthen after 4 d and the last after 10 d.The first cell division was in 6 d. Thus, times to startlengthening were similar for spores whether kept in thedark for 2 or 12 months. After 2 yr, spores kept in the darkcould not be germinated. However, spores that were kept indim light but nutrient-limited media were still nearly allviable after 2 yr (June 1995 to July 1997).
In the lake, newly formed spores were often foundgerminating (forming up to two or four cells) once theysank into deeper, nutrient-rich water. To test whether lightor nutrients or both were involved in triggering this, sporesthat had been stored in the dark in nutrient-deficient mediawere divided into two groups; one group was placed in newmedia and illuminated while the other was also put intonew media but kept in the dark (Fig. 13). The spores in thelight germinated most quickly, taking 1.5 d to startlengthening compared with 5 d for those in the dark. Theinitial growth rates in both were then similar and some ofthe fastest recorded, doubling in less than a day for the firsttwo to three divisions. Then there was a marked difference.
Fig. 13. Excystment (germination) of resting spores ofAulacoseira skvortzowii. The time is taken from the point whenspores were placed into new media in the light and dark. Theincrease in the number of cells per filament is an average of fivespore germinations in the light and six in the dark.
1132 Jewson et al.

The cells in the light continued to grow and divide rapidlybut at a rate (doubling time 1.5 to 2.0 d) that was similar togrowth rates obtained in other culture experiments. In theother group, the spores in the dark managed to form fourto six cells but then stopped lengthening or dividing(Fig. 13), i.e., the spores had sufficient reserves for two orthree divisions.
Discussion
During summer stratification, temperatures in the surfacelayers of Lake Baikal become lethal for A. skvortzowii(Fig. 3). Also, nutrients decline to undetectable concentra-tions (Fig. 7a,b), mainly because of picoplankton growth(Nagata et al. 1994; Belykh and Sorokovikova 2003; Fietz etal. 2005). Therefore, a resting stage helps A. skvortzowiisurvive these unfavorable conditions but successful sporu-lation depends on having sufficient time to switch resourcesand prepare for dormancy. Cultures with high growth rates(doubling every 2 d) took 9 d to complete the morphologicalchanges and build up storage products. In the lake, wheredoubling times during spring mixing could be more thanthree times longer, sporulation sometimes took up to 3 to 4weeks to complete. Thus, for successful sporulation, an earlysignal of environmental change is required. McQuoid andHobson (1996) summarized the wide range of cues used bydiatoms for resting spore production, including those inmarine coastal waters, where nitrate is frequently important(Kuwata et al. 1993; Kuwata and Takahashi 1999).However, although low nitrate did initiate some sporeproduction in cultures of A. skvortzowii, low phosphate wasmore effective, with nearly all cells successfully formingspores. In the lake, phosphate reached the thresholdconcentration for sporulation ahead of stratification in 9of 11 yr in the south basin (e.g., Fig. 7b). In contrast, thenitrate decline was usually later and occurred whentemperatures were already rising (Fig. 7). At other times ofthe year, such as under the ice, spore formation alsooccurred when there was a decline in phosphate concentra-tion during periods of high phytoplankton biomass (e.g.,March 1997 and April 2000). The resting spores either sankto the bottom inshore or were held up at the thermoclineoffshore. Armand and Zielinski (2001) discussed a similarsituation in winter resting stages in Rhizosolenia in theSouthern Ocean, where they suggested that heavily silicifiedsingle cells did not sink to the sediment but only to thebottom of the mixing layer or pycocline.
Resting spores germinated if they settled into deeper,nutrient-rich water, where they then died because of lack oflight. Therefore, no live cells or spores were generally foundin the water column by September. This contrasted with theother major endemic members of the open-water phyto-plankton (A. baicalensis, C. baicalensis, Cyclotella minuta),which could be found in cooler subsurface layers (Jewsonunpubl. data). An alternative site for dormancy is thesediments, where settling and resuspension of cells is wellknown in lakes (Lund, 1955; Jewson 1992a; Schelske et al.1995) and now is also being shown to be important forcoastal marine populations (McQuoid 2002; McQuoid et al.2002; Ishikawa and Furuya 2004). McQuoid and Godhe
(2004), in a study of recruitment in a fjord, stressed theimportance in seasonal studies of including both sedimentand planktonic ‘‘life history stages,’’ concluding that ‘‘abenthic resting stage offers an extra level of insurance forsurvival of a species.’’ Investigations of Baikal coastalsediments (0–25 m) in summer found high concentrationsof spores at 10 to 25 m. These depths remain cooler andcan still be reached by wave action (Rzheplinsky andSorokina 1977), as the maximum wavelengths in LakeBaikal approach 66 m in 20 to 25 m s21 wind speeds. Insuch high winds, the depth of bottom disturbance (l/2)could reach 33 m in localized areas but more generally,sediments down to 20 m are more likely to be the mainzone for resuspension during winds of 10 to 15 m s21. Thisarea is approximately 2,380 km2 or about 7% of the lakebottom (see Kozhov 1963). However, not all this area issuitable, as much of it is rocky, but, even so, sandy coastalsediments and embayments represent a large area ofpotential ‘‘seed bank.’’
A big advantage of using coastal sediments fordormancy in Baikal is the relative narrowness (20 to40 km) of the lake compared to its length (636 km). Thisincreases the chances for both deposition and resuspension.In one study of coastal processes during late spring(Likhoshway et al. 1996), filaments trapped in thecirculation on the shoreward side of a thermal bar werefound in higher concentrations in currents descending overthe sediments, which would increase their likelihood ofsettling. After storms in autumn, A. skvortzowii popula-tions can sometimes build up quickly in the favorable lightclimate inshore, doubling every 2 d, if nutrient availabilityallows germination (Fig. 13). Currents and gyres (Shimar-aev et al. 1994) then carry cells offshore, allowing them torecolonize much wider areas, but, because of the poorerlight climate caused by vertical mixing to over 100 m, netpopulation increase slows and cell division may be assistedby using stored reserves (Fig. 13). These reserves may alsohelp longevity but in this study most spores stored in thedark lost their viability after 1 yr. This is less than the 2 yrfound in stored spores of some marine species (Hollibaughet al. 1981; McQuoid and Hobson 1996). However, nearlyall spores of A. skvortzowii were still viable and in goodcondition after 2 yr when stored in low light but nutrient-poor media to prevent germination. It is possible that suchconditions might occur in shallow coastal sediments andhelp survival over much longer periods.
Long-term studies of planktonic diatoms in the southbasin of Baikal show large annual variations in peaknumbers (Antipova cited in Kozhov 1963; Bondarenko1999; Popovskaya 2000), with densities of A. skvortzowiioften having a periodicity of 3 or 4 yr.
An important contributory factor to this variability isthe cyclic process of diameter reduction and size regener-ation, which is often overlooked in diatom populationstudies (see Jewson 1992a,b). In most centric diatomspecies, it is a clocking mechanism allowing the buildupof numbers asexually over several years, with a switch ofresources to sexual reproduction once cells are below agiven size threshold (usually less than half the maximumcell diameter). To ensure correct seasonal timing, an
Resting stages in Baikal 1133

environmental cue is also required once the cells are belowthis threshold. In A. skvortzowii populations, low phos-phate concentrations that induced sporulation in wider-diameter cells above 10 mm also caused a switch innarrower cells to gametogenesis and size regeneration(Fig. 11). In A. skvortzowii motile gametes and oocyteswere often observed in cultures once exponential growthceased. However, although we are still uncertain about theextent of sexual reproduction in the natural system, we cansee the impact of the clocking mechanism (size declinefollowed by size regeneration) on population dynamicsbecause the narrower-diameter cells are usually the mostabundant size class (see Fig. 11) but size-regenerated cellsrepresent only 1–2% of the total. This causes a marked shiftin size distribution of the population of A. skvortzowii(Fig. 11) between April and the end of growth in May, withloss of smaller-diameter cells. Thus, the most abundant sizeclass does not contribute any further to vegetative division.By the following year, cells in the narrowest-diameter sizerange are replaced because the mean diameter of largersize classes decreases. The size threshold for inducing sizeregeneration is about 9 mm and is reached about 3 or 4 yrafter size regeneration takes place, under normal growingconditions. This is equivalent to between 24 and 32divisions. Overall, the size distribution of the wholepopulation of A. skvortzowii in Baikal is made up ofoverlapping year classes. This can result in cycles inabundance of 3 to 4 yr that are largely influenced by thereproductive success of earlier years (e.g., 1994 to 2000).However, this underlying pattern is not always seenbecause the population can also be influenced by otherfactors governing population dynamics, especially grazingbut including nutrients, parasitism, ice conditions, andsporulation success.
Although low phosphate appears to be a reliable cue toinitiate sporulation and size regeneration, the studies of A.skvortzowii in the north basin show the limitations of thisadaptation, particularly the risk arising from reliance onother algae to reduce the concentration before tempera-tures rise. In the north basin, few spores were found(Fig. 9) because the decline in phosphate concentration wasoften insufficient to reach the required threshold formorphological change. In an earlier study, Goldman etal. (1996) had also found no evidence of nutrient limitationof the spring crop in the north basin. Phosphateconcentration does decline in the north basin but it isoften after isothermal mixing, when temperatures are rising(e.g., Fig. 3). Thus, with the timing of the declineunreliable, A. skvortzowii is on the edge of its survivallimits there. In particular, the resulting low recruitmentmakes it difficult for the population to sustain itself.Instead, it seems likely that the intermittent appearance ofpopulations in the north basin results from reseeding fromthe central basin. Shimaraev et al. (2003) have calculatedthat the water exchange between the north and centralbasins is about 240 km3 yr21 (about 2% to 3%) andbetween the southern and central basins is 90–130 km3
yr21. These estimates are for water at all depths and notjust surface waters containing phytoplankton, but it doesshow that although the importance in any one year is
probably low, over longer periods it may be sufficient forwider dispersal and survival of the population.
Not all cells of A. skvortzowii formed resting sporesunder stress; a few formed resting cells (see Fig. 2). Thisduality is also found in a marine planktonic diatomChaetocerus pseudocurvisetus, for which Kuwata andTakahashi (1999) showed that the option of forming eitherresting cells or spores can be beneficial at different times ofthe year in fluctuating nitrate conditions, with the trade-offthat resting cells begin growth more quickly than spores butthey do not survive as long when dormant in the watercolumn. In Baikal, resting cells rather than spores weremostly formed when cells were trapped in the dark belowthe euphotic zone. The resting cells are a short-term optionin aphotic conditions, with the big advantage that they donot switch on again if they sink into deeper water, wherethe higher nutrient concentration causes spores to germi-nate and then usually die. Essentially, it means that restingcells of A. skvortzowii are formed after a change inconditions, whereas resting spores are initiated by a cuethat precedes change. Thus the latter gives time for layingdown storage products, which can be used for both survivalduring dormancy and also as an aid to cell division aftergermination (Fig. 13).
In summary, A. skvortzowii has a life cycle suited to theenvironmental conditions in Lake Baikal. In particular, theresting spore helps A. skvortzowii survive periods of lethaltemperatures and low nutrients during summer by lyingdormant in shallow coastal sediments, where it can avoidthe problems of dissolution that occur in open waters. A.skvortzowii then relies on resuspension and dispersal bycurrents and gyres to recolonize the offshore waters eachyear. A disadvantage of spore formation is the relativelylong time needed for production, which means that it isnecessary to have an environmental cue well in advance ofany change in conditions. A. skvortzowii relies mainly onlow phosphate concentration. If phosphate does not declineor only does so once stratification is underway, then fewerspores are produced (Fig. 7c) and there is also lessrecruitment through size regeneration. This makes popula-tions less able to sustain themselves, which is presently thecase in the north basin. However, from paleoecologicalstudies, we know that there have been times when A.skvortzowii was abundant there for long periods (Bradburyet al. 1994; Bangs et al. 2000), suggesting that the decline ofphosphate concentration in the north occurred later. It isimportant to realize that this critical relationship withphosphate would be missed by the correlation analysespresently used by many studies for investigating significantvariables. The results from the north basin also show thebenefit for long-term survival of living in a complex habitatlike Lake Baikal, which stretches over 636 km and threebasins, i.e., there are likely to be refuge areas with the rightconditions somewhere. It is only through long-termmultidisciplinary studies that we can begin to understandfully the ecological niches of individual species and howthey interact with other phytoplankton in the samecommunity. Thus, although A. skvortzowii can be foundgrowing offshore together with other species in spring, forthe rest of the year it has a quite different ecology.
1134 Jewson et al.

References
ARMAND, L. K., AND U. ZIELINSKI. 2001. Diatom species of thegenus Rhizsolenia from Southern Ocean sediments andtaxonomic notes. Diatom Res. 16: 259–294.
BABANAZAROVA, O. V., Ye. V. LIKHOSHWAY, AND D. Yu.SHERBAKOV. 1996. On the morphological variability ofAulacoseira baicalensis and Aulacoseira islandica (Bacillario-phyta) of Lake Baikal, Russia. Phycologia 35: 113–123.
BANGS, M., AND oTHERS. 2000. Climate change in Lake Baikal:Diatom evidence in an area of continuous sedimentation. Int.J. Earth Sci. 2: 251–259.
BELYKH, O. I., AND E. G. SOROKOVIKOVA. 2003. Autotrophicpicoplankton in Lake Baikal: Abundance, dynamics anddistribution. Aquat. Ecosyst. Health Manage. 6: 251–261.
BONDARENKO, N. A. 1999. Floral shift in the phytoplankton ofLake Baikal, Siberia: Recent dominance of Nitzschia acicu-laris. Plankton Biol. Ecol. 46: 18–23.
———, N. E. GUSELNIKOVA, N. F. LOGACHEVA, AND G. V.POMAZKINA. 1996. Spatial distribution of phytoplankton inLake Baikal, spring 1991. Freshwat. Biol. 35: 517–523.
BRADBURY, P. J., Ye. V. BEZRUKOVA, G. P. CHERNYAEVA, S. M.COLMAN, G. KHURSEVICH, J. W. KING, AND Ye. V. LIKOSHWAY.1994. A synthesis of post-glacial diatom records from LakeBaikal. J. Paleolimnol. 10: 213–252.
EDGAR, M. S., AND E. C. THERIOT. 2004. Phylogeny of Aulacoseira(Bacillariophyta) based on molecules and morphology. J.Phycol. 40: 772–788.
EDLUND, M. B., E. F. STOERMER, AND C. M. TAYLOR. 1996.Aulacoseira skvortzowii sp. nov. (Bacillariophyta), a poorlyunderstood diatom from Lake Baikal, Russia. J. Phycol. 32:165–175.
FIETZ, S., G. KOBANOVA, L. IZMEST’EVA, AND A. NICKLISCH. 2005.Regional, vertical and seasonal distribution of phytoplanktonand photosynthetic pigments in Lake Baikal. J. Plankton Res.27: 793–810.
GENKAL, S. I., AND N. A. BONDARENKO. 2001. Floral study of algae(Centrophyceae, Bacillariophyta) in some lakes of Pribaikalieand Zabaikalie Biol. Inland Waters 1: 3–10 (in Russian).
———, AND ———. 2004. Electron microscopic studies ofBacillariophyta in the plankton of mountain lakes of theLena basin I. Centrophyceae. Bot. J. 89: 1588–1596 (inRussian).
———, AND L. A. SCHURE. 2000. New data on the flora ofBacillariophyta in Lake Khanka (Primorskiy Province,Russia) Algology 10: 278–281 (in Russian).
GOLDMAN, C. R., J. J. ELSER, R. C. RICHARDS, J. E. REUTER, J. C.PRISCU, AND A. L. LEVIN. 1996. Thermal stratification,nutrient dynamics, and phytoplankton productivity duringthe onset of spring phytoplankton growth in Lake Baikal,Russia. Hydrobiologia 331: 9–24.
GRANIN, N. G., AND oTHERS. 1999. Turbulent mixing in the waterlayer just below the ice and its role in development of diatomicalgae in Lake Baikal Doklady Akad. Nauk 366: 835–839 (inRussian).
HEIM, B., H. OBERHAENSLI, S. FIETZ, AND H. KAUFMANN. 2005.Variation in Lake Baikal’s phytoplankton distribution andfluvial input assessed by SeaWiFs satellite data. Glob. Planet.Change 46: 9–27.
HOLLIBAUGH, J. T., D. R. L. SIBERT, AND W. H THOMAS. 1981.Observations on the survival and germination of restingspores of three Chaetoceros (Bacillariophyceae) species. J.Phycol. 17: 1–9.
ISHIKAWA, A., AND K. FURUYA. 2004. The role of diatom restingstages in the onset of spring bloom in the East China Sea.Mar. Biol. 145: 633–639.
JEWSON, D. H. 1992a. Size reduction, reproductive strategy andthe life cycle of a centric diatom. Phil. Trans. R. Soc. Lond. B.336: 191–213.
———. 1992b. Life cycle of a Stephanodiscus sp. (BACILLAR-IOPHYTA). J. Phycol. 28: 856–866.
KOZHOV, M. 1963. Lake Baikal and its life. Junk.KOZHOVA, O. M. 1987. Phytoplankton of Lake Baikal: Structural
and functional characteristics. Arch. Hydrobiol. Beih. Er-gebn. Limnol. 25: 19–37.
———, AND L. R. IZMEST’EVA. 1998. Lake Baikal, evolution andbiodiversity. Backhuys.
KUWATA, A., T. HAMA, T., AND M. TAKAHASHI. 1993. Ecophys-iological characterisation of two life forms, resting spores andresting cells, of a marine planktonic diatom, Chaetocerospseudocurvisetus, formed under nutrient depletion. Mar. Ecol.Prog. Ser. 102: 245–255.
———, AND M. TAKAHASHI. 1999. Survival and recovery of restingspores and resting cells of the marine planktonic diatomChaetoceros pseudocurvisetus under fluctuating nitrate condi-tions. Mar. Biol. 134: 471–478.
LIKHOSHWAY, Y. V., A. YE. KUZMINA, T. G. POTYEMKINA, V. L.POTYEMKIN, AND M. N. SHIMARAEV. 1996. The distribution ofdiatoms near a thermal bar in Lake Baikal. J. Great LakesRes. 22: 5–14.
———, M. V. USOLTSEVA, G. I. POPOVSKAYA, S. S. VOROBYOVA,AND T. V. NIKOLINA. 2004. Spores of Aulacoseira (Bacillar-iophyta) from recent water bodies and deposits of differentages. Bot. J. 89: 60–72 (in Russian).
LUND, J. W. G. 1955. Further observations on the seasonal cycleof Melosira italica (Ehr.) Kutz. subsp. subarctica O. Mull. J.Ecol. 43: 90–102.
———. 1966. Znachenie turbulentnosti vody v priodichnostirazvitiya nekotorykh presnovodnyh vidov roda Melosira(Algae). Bot. Zh. Kyyiv. 51: 176–187.
MCQUOID, M. R. 2002. Pelagic and environmental controls on thespatial distribution of a viable diatom propagule bank on theSwedish west coast. J. Phycol. 38: 881–893.
———, AND A. GODHE. 2004. Recruitment of coastal planktonicdiatoms from benthic versus pelagic cells: Variations in bloomdevelopment and species composition. Limnol. Oceanogr. 49:1123–1133.
———, ——— , AND K. NORDBERG. 2002. Viability of phyto-plankton resting stages in the sediments of a coastal Swedishfjord. Eur. J. Phycol. 37: 191–201.
———, AND L. A. HOBSON. 1996. Diatom resting stages. J. Phycol.32: 889–902.
NAGATA, T., AND oTHERS. 1994. Autotrophic picoplankton insouthern Lake Baikal—abundance, growth and grazingmortality during summer. J. Plankton Res. 16: 945–959.
POPOVSKAYA, G. I. 2000. Ecological monitoring of phytoplanktonin Lake Baikal. Aquat. Ecosyst. Health Manage. 3: 215–225.
———, S. I. GENKAL, AND YE. V. LIKHOSHWAY. 2002. Diatoms ofthe plankton of Lake Baikal: Atlas and Key. Novosibirsk:Nauka (in Russian).
RZHEPLINSKY, G. V., AND A. I. SOROKINA. 1977. Atlas of wave andwind action in Lake Baikal. Gidrometeoizdat (in Russian).
SCHELSKE, C. L. H., H. J. CARRICK, AND F. J. ALDRIDGE. 1995. Canwind-induced resuspension of meroplankton affect phyto-plankton dynamics? J. North Am. Benthol. Soc. 14: 616–630.
SHERBAKOVA, T. A., S. V. KIRILCHIK, YE. V. LIKHOSHWAY, AND M.A. GRACHEV. 1998. Phylogenetic relationships of the Baika-lian Aulacoseira diatoms inferred from comparison of partial18S rRNA sequences. Mol. Biol. 32: 612–616.
SHIMARAEV, M. N., N. G. GRANIN, AND A. A. ZHADANOV. 1993.Deep ventilation of Lake Baikal waters due to spring thermalbars. Limnol. Oceanogr. 38: 1068–1072.
Resting stages in Baikal 1135

———, V. I. VERBOLOV, N. G. GRANIN, AND P. P. SHERSTYANKIN.1994. Physical limnology of Lake Baikal: A review. In M. N.Shimaraev and S. Okuda [eds.], Baikal International Centrefor Ecological Research, Irkutsk-Okayama.
———, AND oTHERS. 2003. Inter-basin water exchange in LakeBaikal. Vodniye Res. 30: 678–681.
SKABICHEVSKY, A. P. 1929. On the biology of Melosira baicalensis(K. Meyer) Wisl. Rus. Hydrobiol. Zh. 8: 93–114 (in Russian).
———. 1953. On spore formation of Melosira islandica O. Mull.Dokl. Akad. Nauk. USSR 92: 671–674 (in Russian).
———. 1960. Planktonic freshwater diatoms of the USSR.Systematics, biology and distribution. Moscow State Univer-sity (in Russian).
SKVORTZOW, B. W. 1937. Bottom diatoms from Olkhon Gate ofBaikal Lake, Siberia. Philipp. J. Sci. 62: 293–377.
STROGANOV, N. S., AND N. S. BUZINOVA. 1980. Practical manual onhydrochemistry. MGU, Moscow (in Russian).
WEISS, R. F. E., E. C. CARMACK, AND V. M. KOROPALOV. 1991.Deep water renewal and biological production in Lake Baikal.Nature 349: 665–669.
WETZEL, R. G., AND G. E. LIKENS. 1991. Limnological analyses.Springer Verlag.
WUEST, A., T. A. RAVENS, N. G. GRANIN, O. KOCSIS, M.SCHURTER, AND M. STURM. 2005. Cold intrusions in LakeBaikal: Direct observational evidence for deep-water renewal.Limnol. Oceanogr. 50: 184–196.
YASNITSKY, V. N. 1930. Results of observations of the plankton ofBaikal in the area of the Biological Station in 1926–1928Izvestyia Biol-Geogr. Inst. Irkutsk Univ. 4: 191–238 (inRussian).
Received: 12 February 2007Amended: 24 October 2007
Accepted: 9 December 2007
1136 Jewson et al.
Copyright © 2022 FDOKUMEN




















