
Inferring Skill from Tests of Programming Performance: Combining Time and Quality

This article was downloaded by: [Natural History Museum], [Dr David M. Williams]On: 01 November 2011, At: 02:08Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Diatom ResearchPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/tdia20
INFERRING DIATOM PHYLOGENY: ACLADISTIC PERSPECTIVEJohn P. Kociolek a , Edward C. Theriot b & David M. Williams ca Diatom Collection, Departments of Invertebrate Zoology andGeology, California Academy of Sciences, Golden Gate Park, SanFrancisco, CA, 94118, U.S.A.b Diatom Herbarium, Academy of Natural Sciences of Philadelphia,19th & The Parkway, Philadelphia, Pennsylvania, 19103, U.S.A.c Department of Botany, British Museum (Natural History), CromwellRoad, London, SW7 SBD, U.K.
Available online: 31 Oct 2011
To cite this article: John P. Kociolek, Edward C. Theriot & David M. Williams (1989): INFERRINGDIATOM PHYLOGENY: A CLADISTIC PERSPECTIVE, Diatom Research, 4:2, 289-300
To link to this article: http://dx.doi.org/10.1080/0269249X.1989.9705077
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representationthat the contents will be complete or accurate or up to date. The accuracy of anyinstructions, formulae, and drug doses should be independently verified with primarysources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand, or costs or damages whatsoever or howsoever caused arising directly orindirectly in connection with or arising out of the use of this material.

Diatom Research (1989) Volume 4 (2), 289-300.
INFERRING DIATOM PHYLOGENY: A CLADISTIC PERSPECTIVE John P. Kociolek
Diatom Collection, Departments of Invertebrate Zoology and Geology, California Academy of Sciences, Golden Gate Park,
San Francisco, CA 94118, U.S.A.
Edward C. Theriot Diatom Herbarium, Academy of Natural Sciences of Philadelphia., 19th & The Parkway, Philadelphia, Pennsylvania, 19103, U.S.A.
David M. Williams
Department of Botany, British Museum (Natural History), Cromwell Road, London SW7 SBD, U.K.
The true phylogeny of a lineage can never be known, only estimated. Phylogenetic Systematics or Cladistics represents one approach to inferring genealogical relationships of diatoms. The assumptions of the cladistic approach and its associated methodology are outlined, and examples are presented for the diatoms. The relevance of recognising monophyletic groups to phylogenetic reconstruction is discussed.
INTRODUCTION
‘The most difficult as well as most fascinating problem in connection with any group is its phylogeny. The data upon which we base opinions concerning phylogeny are never sufficient but such opinions usually stimulate research and are necessary to progress.’ (Coulter cited in Lotsy 1907).
‘Phylogeny, e.g. reconstruction of what has happened in the past, is no science but a product of phantastic [sic] speculations . . .’ (Lotsy 1916, p. 140).
The phylogeny of any group is a truth that can never be known entirely and, therefore, some may argue Lotsy may have been justified in his lament of 1916. And as Coulter points out we will always be without sufficient data to know the TRUE phylogeny. Thus all we can do is attempt to make some estimate of that phylogeny. This does not, however, make the endeavour hollow or uninteresting. The job of the systematist is, in our view, to identify homologous characteristics and with these data produce an hypothesis about phylogeny. In producing such an estimate we assume that similarities are a guide to relationships. Put another way, we must assume the similarities between two varieties of the same species are due to common genealogy and not the result of two taxa from vastly different lineages which have independently evolved nearly the same morphologies.
Reconstruction of diatom phylogeny has recently received renewed attention. Appreciation of cytological details and life cycles as well as the synthesis of over 50 years of electron microscopical observations have provided a wide range of characters on which to base hypotheses of relatedness. Several approaches have been applied or supported in the recent literature as means to the end of phylogeny estimation. One approach that has begun to receive considerable attention is known as Phylogenetic Systematics or Cladistics. The attention this approach has received is evidenced by the large number of botanists, zoologists and protistologists that have applied the method, and
Dow
nloa
ded
by [
Nat
ural
His
tory
Mus
eum
], [
Dr
Dav
id M
. Will
iam
s] a
t 02:
08 0
1 N
ovem
ber
2011

290 J.P. KOCIOLEK, E.C. THERIOT AND D.M. WILLIAMS
examples exist from the diatom literature (Williams 1985, Williams & Round 1986, 1987, 1988, Kociolek & Stoermer 1986, 1987, 1988, 1989, Theriot & Kociolek 1986, Theriot et al. 1987). In this paper we explore the necessary assumptions, methodology and expected results of cladistic analysis.
PHYLOGENETIC SYSTEMATICS
Two assumptions are necessary for cladistic analysis, namely, 1) there are natural taxa in the world and, 2) there is descent with modification (evolution). New taxa originate through evolution and through time a genealogy of taxa develops. Cladistic analysis attempts to estimate genealogical history on the basis of shared (homologous) derived character states. Such character states are termed synapomorphies (syn = together, shared; apomorphy = derived character). The basis for establishing relationships on synapomorphic character states alone stems from the work of Hennig (1965, 1966), who suggested that as lineages evolve, evolutionary novelties arise which are found in the descendant taxa. Subsequent evolution in this same lineage may result in the origin of other novelties found in the most recently evolved forms only (Figs la, b). Thus all descendants have the novelties first evolved, while only a subset of these taxa possess the novelties evolved later. These hierarchical levels (or nested sets) of taxa are defined by the shared derived features.
A
A \
Fig. la, b. (a). Divergence of taxa A and B, with evolutionary novelties 1 and 2 developed in B. (b). Later in the history of this lineage taxa C and D have evolved and share novelties 3 and 4.
All features can be homologous at some level in the systematic hierarchy and discovery of its appropriate level is a fundamental part of cladistic analysis. Stevens (1984, p. 396) defines homology as, ‘A similarity in structure in a group of organisms that delimits that group and that was found in the common ancestor.’ For example, it has been suggested that there are three major groups of
Dow
nloa
ded
by [
Nat
ural
His
tory
Mus
eum
], [
Dr
Dav
id M
. Will
iam
s] a
t 02:
08 0
1 N
ovem
ber
2011

CLADISTICS 291
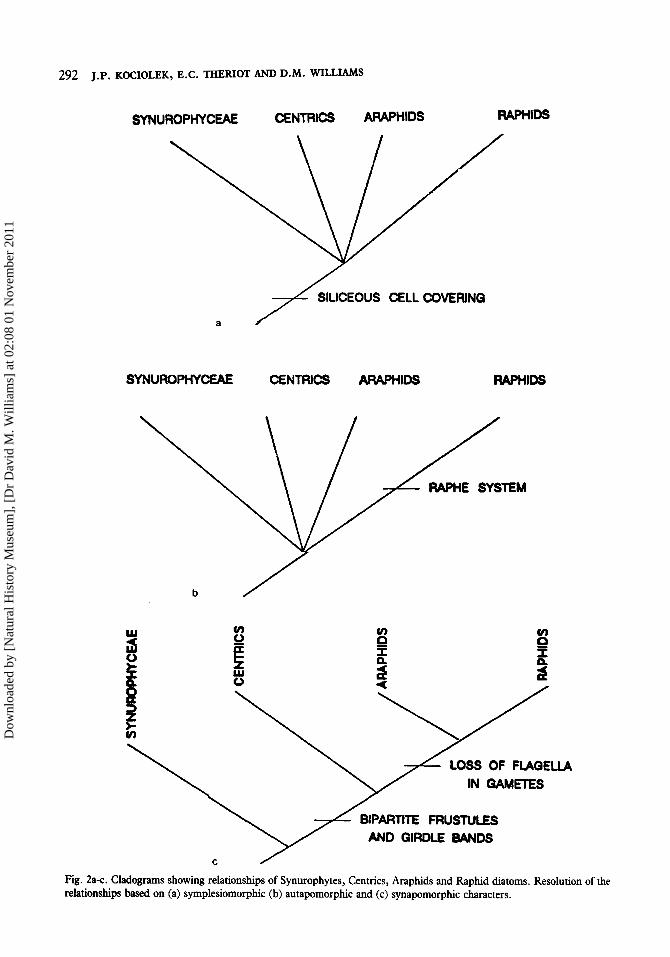
diatoms; centrics, araphids and raphids (cf. Smith 1872). The Synurophycean Chrysophytes have been suggested by some (Round & Crawford 1981, Kociolek 1986, Cavalier-Smith 1986) to be the group most closely allied to the diatoms (that is, the sister group of the diatoms). We might ask, ‘What are the relationships between these four taxa (centric diatoms, araphid diatoms, raphid diatoms and Synurophyceae)?’ One of the characters we could choose to help define the relationships is ‘Cell covering of silica.’ This feature is found in all the groups (Fig. 2a), and therefore cannot help resolve relationships between the four taxa. Such shared but primitive features are termed ‘symplesiomorphies’ and are considered uninformative to the question at hand. Another character we could possibly choose is, ‘Presence of a raphe system.’ This feature is derived, but unique to the raphid group (Fig. 2b). Such derived but unique features, termed ‘autapomorphies,’ define terminal taxa, but do not help resolve the broader question of interrelationships among them. Two characters that may help resolve this question of phylogenetic relationships are ‘Presence of bipartite frustules and girdle bands’ and ‘Loss of flagella in gametes.’ The former defines the group comprised of the centrics, araphids and raphids (diatoms), while the latter defines the araphids and raphids (pennate diatoms). These characters are both shared and derived and, as shown in figure 2c, do provide resolution of the relationships of the four taxa. Note that characters can be synapomorphic at one level of hierarchy and symplesiomorphic at another; thus, the level of hierarchy at which phylogenetic questions are being posed must necessarily be explicitly stated.
If all observed features were real homologies, then the true hypothesis of descent would be obvious; this occurs rarely if at all. The condition of a feature being incongruent with a particular phylogenetic hypothesis is termed ‘homoplasy’, which is a more general term than ‘reversal’ ‘convergence’ or ‘parallelism’. The term homoplasy is useful because, in some cases, there is ambiguity as to whether character incongruency is due to double gains of an apomorphic feature (e.g. convergence, Fig. 3a) or a gain and a loss (e.g. reversal, Fig. 3b). The concept of homoplasy includes both types of character distribution patterns.
CHARACTER POLARITY AND THE FORMULATION OF HYPOTHESES
Since cladistics attempts to estimate phylogeny based on shared, derived characters, polarity of character states is essential to the analysis. There exist three criteria on which to rationally base determinations of character polarity: fossils, ontogeny and outgroups. The first criterion is derived from the fossil record and suggests that character states observed in stratigraphically older taxa are primitve relative to character states found in stratigraphically younger taxa. Examples in the diatom literature may be found in Sims (1986) and Fryxell et al. (1986). ‘Tracing the evolution of valve outline in Sphynctolethus shows this to be broadly lanceolate in the Cretaceous, passing through elliptic, elongate ahd finally circular in the Oligocene’ (Sims 1986, p. 266) is an example of applying the stratigraphic criterion. Recently some systematists (Eldredge & Cracraft 1980; see also Kociolek 1986) have cautioned against the exclusive use of stratigraphic precedence in polarizing character states because a stratigraphically older taxon may have derived character states. A case in point is Mesodictyon Theriot & Bradbury which seems to have evolved early in the history of the freshwater Thalassiosiraceae (Krebs et al. 1987) but has highly derived loculate areolae.
The second method of determining character polarity is through ontogeny. The ontogeny criterion states, ‘Given an ontogenetic character transformation, from a character observed to be more general to a character to be less general, the more general character is primitive and the less general advanced’ (Nelson & Platnick 1981, p. 332), or more concisely, for a transformation of character states observed in ontogeny, primitive features are followed by more derived features. Although the general applicability of the ontogeny criterion is still being debated (e.g. Kluge 1985, Nelson 1985, De Querioz 1985, Kraus 1988, papers in Humphries 1988) it has been used to determine character polarity in many studies. Some have argued unicells such as diatoms may be without ontogenies (‘How can a single cell differentiate?’), however Kociolek & Williams (1987) have suggested ontogenetic information may be gained from unicells at two levels: during a
Dow
nloa
ded
by [
Nat
ural
His
tory
Mus
eum
], [
Dr
Dav
id M
. Will
iam
s] a
t 02:
08 0
1 N
ovem
ber
2011
292 J.P. KOCIOLEK, E.C. THERIOT A N D D.M. WILLIAMS
SYNUROPHYCEAE CENTRICS ARAPHIDS RAPHIDS
CELL COVERING
SYNUROPHYCEAE CENTRlCS ARAPHIDS RAPHIDS
tn tn tn
IN W R E S
AND GIRDLE BANDS
Fig. 2a-c. Cladograms showing relationships of Synurophytes, Centrics, Araphids and Raphid diatoms. Resolution of the relationships based on (a) symplesiomorphic (b) autapomorphic and (c) synapomorphic characters.
Dow
nloa
ded
by [
Nat
ural
His
tory
Mus
eum
], [
Dr
Dav
id M
. Will
iam
s] a
t 02:
08 0
1 N
ovem
ber
2011

CLADISTICS 293
0 1 1 0
A B C D
A
1
B
1
C 0
b / Fig. 3a, b. Examples of homoplasy. In both, upper case letters refer to taxa, 0 = primitive character state, 1 = derived character state. (a) shows the evolution of the derived feature in taxa B and C by ‘double gain’. (b) shows the evolution of the feature in the ancestor of the lineage B+C+D, and its secondary loss in D. Regardless of the mechanism, a minimum of 2 transformations must occur.
particular process or phase of the life cycle (valve morphogenesis, gametogenesis) and between stages of the life cycle (comparison of initial valves to ‘normal’ vegetative cells). An example in which the ontogeny criterion has been applied to diatoms is Round & Crawford’s (1981) comparison of scale-bearing initial cells with normal valves, which led them to conclude presence of scales is the ancestoral condition in diatoms. Other examples are presented in Kociolek & Williams (1987) .
The third method.by which character states may be polarized is known as the outgroup criterion. This is the most widely used method and considered by some cladists as the ‘best’ way to polarize characters. The outgroup criterion suggests that character states present in both the group under investigation (the ingroup) and closely allied sister taxa (the outgroups) must be primitive, because such character states were most likely present in the taxon ancestoral to both
Dow
nloa
ded
by [
Nat
ural
His
tory
Mus
eum
], [
Dr
Dav
id M
. Will
iam
s] a
t 02:
08 0
1 N
ovem
ber
2011

294 J.P. KOCIOLEK, E.C. THERIOT AND D.M. WILLIAMS
groups (Watrous & Wheeler 1981, Maddison et aZ. 1984). Alternatively, character states found in the ingroup but not the outgroups must have evolved subsequent to their divergence and thus are considered derived. Kociolek (1986) used outgroup analysis to determine character polarity of chromatophore type and sexuality in the diatoms. Implicit application of the outgroup criterion can be found in Theriot & Kociolek’s (1986) definition of Stephanodiscus, determination of polarity of rimoportule morphology (Theriot et al. 1987) and Mann’s (1986) attempt to polarize types of sexuality in the genus Nitzschza. In the last example, Mann tried to establish whether isogamy or anisogamy is primitive in the Bacillariaceae. He suggests that in Amphora sensu Zato exist ‘the closest relatives of the fibulate diatoms’ (Mann 1986, p. 202). Thus Amphora serves as the outgroup in Mann’s inherently cladistic approach. ‘Since Amphora species are isogamous . . .’ (Mann 1986, p. 202) it is concluded behavioural anisogamy is the derived condition within the Bacillariaceae. In the case where a hypothesis of outgroup relationships is not well corroborated, different outgroups may be used in repeated cladistic analyses (Donoghue & Cantino 1984). Occasionally, character states vary among several outgroup taxa as well as within the ingroup. This precludes direct application of the outgroup criterion. Maddison et al. (1984) have demonstrated that character polarity can be established after a tree is rooted with outgroup analysis. Use of multiple outgroups is always recommended.
Once homologies are recognized and character states polarized, the next step in a cladistic analysis is to generate a phylogenetic hypothesis or cladogram. The possibility may exist for producing a multitude of cladograms, as different characters may support different hypotheses. The arbitor for choosing between the total number of possible hypotheses is known as parsimony. The principle of parsimony suggests that the favoured hypothesis is the one which explains as much of the data as synapomorphies, and minimizes the amount of homoplasy (Humphries & Chappill 1988). Farris (1983) and Kluge (1984) provide detailed explanations for the application of parsimony to cladistic analysis. Unless parsimony is invoked as the arbitor for choosing between hypotheses, we are left with nothing but ‘fantastic speculations.’
Use of parsimony in deciding between competing hypotheses is demonstrated in the following (admittedly contrived) example. Through the history of phycological research alignment of the diatoms with other algal groups has been debated. Three opinions concerning the systematic position of the diatoms include:
‘Diatomaceae were long believed to be animals . . .but the discovery of the coupling [process] . . . leaves no doubt that they belong to the same order of beings as Desmidiaceae.’ (Berkeley 1857,
‘. . . the suggested relationship of Diatoms with Desmids is based merely on superficial
‘. . . the first diatom probably evolved from an early synurean.’ (Cavalier-Smith 1986, p. 340). To evaluate the relationship between diatoms, desmids and synurophytes a number of charac-
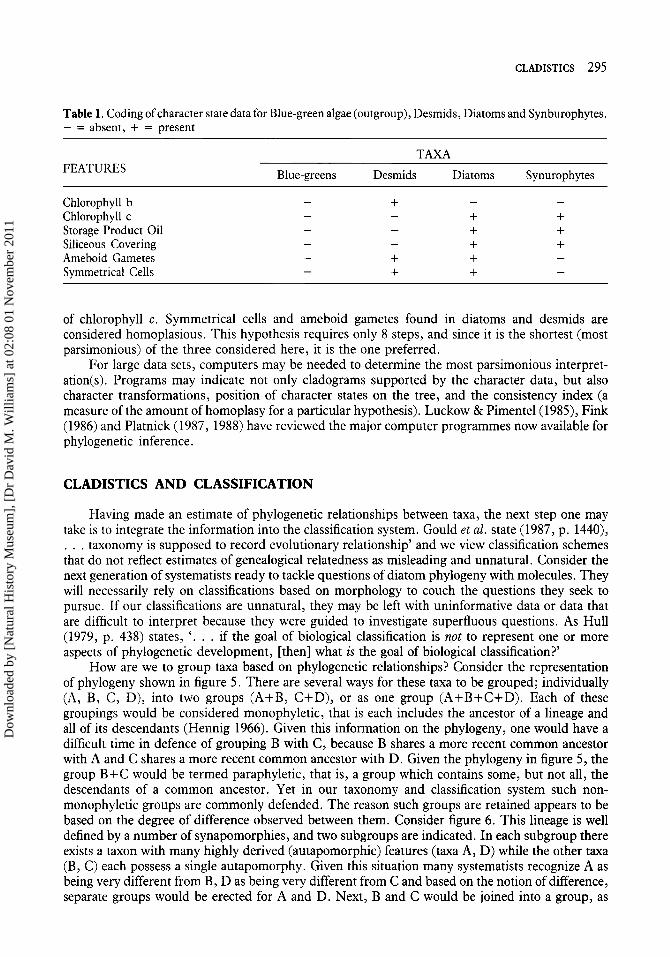
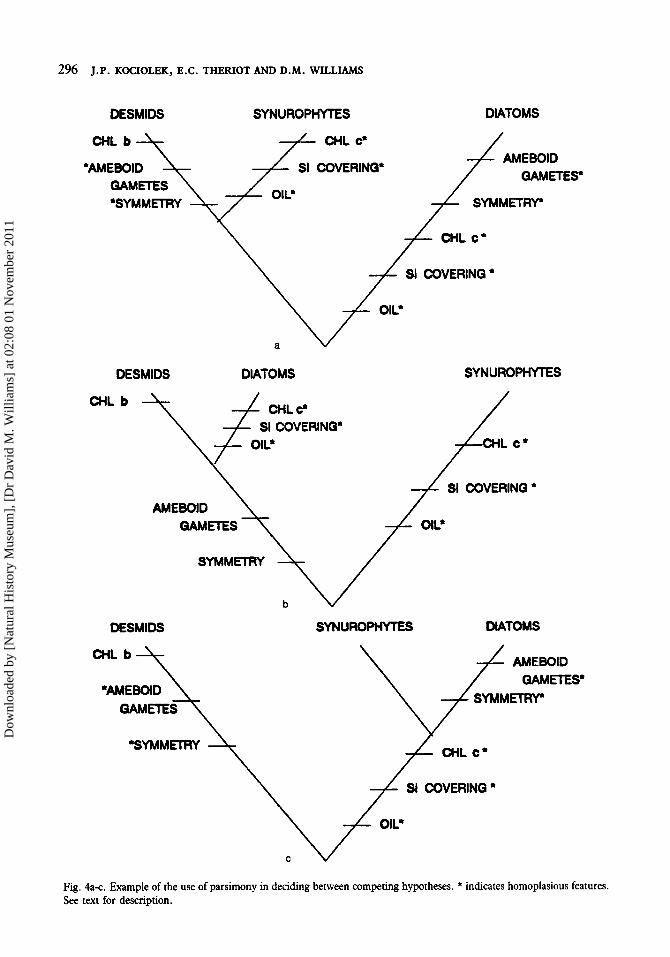
ters are used (Table 1). In this example character states are polarized using the outgroup method, with blue-green algae serving as the outgroup. Three hypotheses may be generated from the character state data (Figs 4a-c). The first hypothesis (Fig. 4a), not previously proposed in the literature, suggests desmids and synurophytes share a more recent common ancestor than either does with diatoms. In this hypothesis, all the data must be considered homoplasious (attributable to either evolutionary reversals or convergence), except presence of chlorophyll b which is autapo- morphic for desmids. The first hypothesis requires 11 evolutionary steps. The second hypothesis generated from the data in Table 1 (Fig. 4b) indicates diatoms and desmids share a more recent common ancestor with one another than either does with synurophytes. In this hypothesis the features ‘ameboid gametes’ and ‘symmetrical cells’ are considered synapomorphies and define the diatoms+desmids lineage. The features shared between diatoms and synurophytes are considered homoplasies. The second hypothesis requires 9 evolutionary steps and, since it is shorter, is preferred over the first. The third hypothesis (Fig. 4c) suggests diatoms and synurophytes share a more recent common ancestor than either does with desmids. Support for this hypothesis comes from the shared, derived features of siliceous cell coverings, oil as a storage product, and presence
pp. 125-126).
resemblance.’ (Fritsch 1935, p. 642).
Dow
nloa
ded
by [
Nat
ural
His
tory
Mus
eum
], [
Dr
Dav
id M
. Will
iam
s] a
t 02:
08 0
1 N
ovem
ber
2011
CLADISTICS 295
Table 1. Coding of character state data for Blue-green algae (outgroup), Desmids, Diatoms and Synburophytes. - = absent, + = present
FEATURES T A M
Blue-greens Desmids Diatoms Synurophytes
Chlorophyll b Chlorophyll c Storage Product Oil Siliceous Covering Ameboid Gametes Symmetrical Cells
of chlorophyll c. Symmetrical cells and ameboid gametes found in diatoms and desmids are considered homoplasious. This hypothesis requires only 8 steps, and since it is the shortest (most parsimonious) of the three considered here, it is the one preferred.
For large data sets, computers may be needed to determine the most parsimonious interpret- ation(s). Programs may indicate not only cladograms supported by the character data, but also character transformations, position of character states on the tree, and the consistency index (a measure of the amount of homoplasy for a particular hypothesis). Luckow & Pimentel(1985), Fink (1986) and Platnick (1987, 1988) have reviewed the major computer programmes now available for phylogenetic inference.
CLADISTICS AND CLASSIFICATION
Having made an estimate of phylogenetic relationships between taxa, the next step one may take is to integrate the information into the classification system. Gould et al. state (1987, p. 1440), . . . taxonomy is supposed to record evolutionary relationship’ and we view classification schemes that do not reflect estimates of genealogical relatedness as misleading and unnatural. Consider the next generation of systematists ready to tackle questions of diatom phylogeny with molecules. They will necessarily rely on classifications based on morphology to couch the questions they seek to pursue. If our classifications are unnatural, they may be left with uninformative data or data that are difficult to interpret because they were guided to investigate superfluous questions. As Hull (1979, p. 438) states, ‘. . . if the goal of biological classification is not to represent one or more aspects of phylogenetic development, [then] what is the goal of biological classification?’
How are we to group taxa based on phylogenetic relationships? Consider the representation of phylogeny shown in figure 5. There are several ways for these taxa to be grouped; individually (A, B, C, D), into two groups (A+B, C+D), or as one group (A+B+C+D). Each of these groupings would be considered monophyletic, that is each includes the ancestor of a lineage and all of its descendants (Hennig 1966). Given this information on the phylogeny, one would have a difficult time in defence of grouping B with C, because B shares a more recent common ancestor with A and C shares a more recent common ancestor with D. Given the phylogeny in figure 5, the group B+C would be termed paraphyletic, that is, a group which contains some, but not all, the descendants of a common ancestor. Yet in our taxonomy and classification system such non- monophyletic groups are commonly defended. The reason such groups are retained appears to be based on the degree of difference observed between them. Consider figure 6 . This lineage is well defined by a number of synapomorphies, and two subgroups are indicated. In each subgroup there exists a taxon with many highly derived (autapomorphic) features (taxa A, D) while the other taxa (B, C) each possess a single autapomorphy. Given this situation many systematists recognize A as being very different from B, D as being very different from C and based on the notion of difference, separate groups would be erected for A and D. Next, B and C would be joined into a group, as
Dow
nloa
ded
by [
Nat
ural
His
tory
Mus
eum
], [
Dr
Dav
id M
. Will
iam
s] a
t 02:
08 0
1 N
ovem
ber
2011
296 J.P. KOCIOLEK, E.C. THERIOT AND D.M. WILLIAMS
DESMIDS SYNUROPHYES DIATOMS
SI COVERING'
Si COVERING'
DESMIDS DIATOMS SY N UROPHYlES
CHLb
SI COVERING.
b
OESMIDS
*AMEBOID
\ *SYMMETRY
SYNUROPHYIES MATOMS
AMEBOID OAMETES'
SYMMETRY
CHL C*
Si COVERING'
C
Fig. 4a-c. Example of the use of parsimony in deciding between competing hypotheses. * indicates homoplasious features. See text for description.
Dow
nloa
ded
by [
Nat
ural
His
tory
Mus
eum
], [
Dr
Dav
id M
. Will
iam
s] a
t 02:
08 0
1 N
ovem
ber
2011

CLADISTICS 297
Fig. 5 . Set of relationships between four taxa. See text for discussion.
they are considered to be more similar to one another than either is to members of the other groups (i.e. A or D). Note however that the similarities between B and C are based on the characteristics that define the entire lineage and, in fact, other than these symplesiomorphies there is no derived feature which supports grouping B with C. Put another way, B and C do not share together a unique genealogical history, and thus constitute an unnatural group. Such unnatural groups are unable to participate in natural processes (such as evolution) (Donoghue & Cantino 1988); therefore non-monophyletic groups should be abandoned.
Fig. 6 . Set of relationships between four taxa with synapomorphic characters plotted on cladogram. See text for discussion.
An example of a paraphyletic group is the centric lineage in Simonsen’s (1979) illustration of diatom relationships. Although Simonsen (p. 11) accepts the bipartition of the diatoms into centrics and pennates (implying each group is monophyletic), his figure 3 shows the Eupodiscaceae of the centrics to share a more recent common ancestor with pennates than with other centrics. This
Dow
nloa
ded
by [
Nat
ural
His
tory
Mus
eum
], [
Dr
Dav
id M
. Will
iam
s] a
t 02:
08 0
1 N
ovem
ber
2011

298 J.P. KOCIOLEK, E.C. THERIOT AND D.M. WILLIAMS
representation shows the centrics to be composed of some, but not all, descendants of a common ancestor and, thus, paraphyletic.
FINAL CONSIDERATIONS
Cladistics has existed as an explicit methodological research programme for just over 20 years and thus there are theoretical and practical matters still being debated. One point that remains unresolved is whether to incorporate each branching event into the classification. With the recognition of sister groups at equal taxonomic rank, it is easy to imagine the inadequacy of the Linnaean system of classification applied to a lineage of many taxa. Wiley (1981) has made some proposals which address this question. One view of the relationship between cladistics and classification doubts whether a strict adherence to monophyletic (sensu Hennig) groups is valuable, as it would mean the break up of well established taxonomic groups (Ashlock 1979, Cronquist 1987). However, the taxonomic imbalances that may result (e.g. monotypic families) from a strict adherence to recognizing monophyletic groups may actually be of great interest from the standpoint of evolutionary biology. Arbitrarily-balanced taxonomic schemes may actually hide pertinent or interesting questions.
Cladistics was often asserted to be a hypothetico-deductive method of phylogenetic analysis (e.g. Eldredge 8z Cracraft 1980). However, phylogenetic analysis is increasingly being understood as a problem of statistical inference. Statistical evaluation is necessary because inference implies probabilistic statements (e.g. a priori expectation of character transition in phylogenetic studies). One methodological question in phylogenetic analysis today is: how can we make probabilistic statements about character transition in the absence of a priori expectation of character evolution? Two authors have made significant contributions to systematics by asking the related question: what is the minimum set of assumptions about probability of character transition to make a parsimonious solution also the most likely solution? Sufficient assumptions have been demonstrated to produce coincidence between parsimony and certain kinds of likelihood analysis and, although the debate is far from over, the results are encouraging (Felsenstein 8z Sober 1986). Parsimony had been criticized by a host of taxonomists as an inappropriate criterion for judging phylogenetic hypotheses on the basis that evolution cannot be assumed to be parsimonious. However, it has been shown clearly that non-parsimonious evolution can still lead to acceptable phylogenetic hypotheses produced by the parsimony criterion (Felsenstein 1983, Sober 1983).
It has also been pointed out that the rational basis of cladistics is not new. The rationale behind cladistic analysis has been applied by systematists before Hennig , and examples exist from the diatom literature. For example, Cleve (1894) proposed that two features distinguish the genus Gomphoneis from Gomphunema, namely doubly punctate striae and longitudinal lines visible in valve view. Hustedt (1942) in his work on the diatoms from the Hawaiian Islands, described Gomphonema intermedium a species with doubly punctate striae but without longitudinal lines. Hustedt reasoned that the presence of doubly punctate striae in this Hawaiian species makes it intermediate (hence the specific epithet) between Gomphonema sensu strict0 and Gowhoneis, and therefore according to him only the presence of longitudinal lines define Gomphoneis. Hustedt identified the shared derived feature which defines the group Gomphonema intermedium + Gom- phoneis (doubly punctate striae) and the derived feature unique to Gomphoneis (longitudinal lines). Cronquist (1987, p. 48) states, ‘[Cladistics] represents a formulation and codification of something that good taxonomists have been doing informally for many years.’ He also notes the technique (p. 45) ‘provide[s] a useful (but not exclusive) way to consider data on which taxonomic schemes and phylogenetic hypotheses are based.’ On these points we agree. Cladistics is a means to estimate phylogeny and character evolution, which may ultimately guide us to establish a more natural system of classification.
CONCLUSIONS
The cladistic method is essentially an analysis of characters. It requires the systematist to be explicit in defining characters, their polarity and states, as well as the level of hierarchy that questions
Dow
nloa
ded
by [
Nat
ural
His
tory
Mus
eum
], [
Dr
Dav
id M
. Will
iam
s] a
t 02:
08 0
1 N
ovem
ber
2011

CLADISTICS 299
are being posed. In return, cladistics offers explicit, testable hypotheses about interrelationships of taxa and the evolution of characters which define those relationships. We believe application of such explicit approaches to questions of diatom phylogeny will, in Coulter’s words, ‘. . . stimulate research and are necessary to progress.’
ACKNOWLEDGMENTS
Support for JPK was provided by Grant BSR-87225 16 of the National Science Foundation.
REFERENCES
ASHLOCK, P.D. (1979). A evolutionary systematist’s view of classification. Systematic Zoology, 28, 441-450 BERKELEY, M. J. (1857). Introduction to Cryptogamic Botany. H. Bailliere, London CAVALIER-SMITH, T. (1986). The Kingdom Chromista: Origin and systematics. Progress in Phycological
CLEVE, P.T. (1 894). Synopsis of the naviculoid diatoms. Kongliga Svenskn vetenskapsakademiens handlingar,
CRONQUIST, A. (1987). A botanical critique of cladism. The Botanical Review, 53, 1-52 DE QUERIOZ, K. (1985). The ontogenetic method for determining character polarity and its relevance to
DONOGHUE, M.J. & CANTINO, P.D. f1984). The logic and limitations of the outgroup substitution
DONOGHUE, M. J. & CANTINO, P.D. (1988). Paraphyly, ancestors, and the goals of taxonomy: A botanical
ELDREDGE, N. & CRACRAFT, J. (1980). Phylogenetic Patterns and the Evolutionary Process. Columbia
FARRIS, J.S. (1983). The logical basis of phylogenetic analysis. In: Advances in Cladistics, Vol. 2 (N.I.
FELSENSTEIN, J. (1983). Parsimony in systematics: Biological and statistical issues. Annual Reoiezu of
FELSENSTEIN, J. & SOBER, E. (1986). Parsimony and likelihood: An exchange. Systcmatic Zoology, 35,
FINK, W.L. (1986). Microcomputers and phylogenetic analysis. Science, 234, 1135-1 139 FRITSCH, F.E. (1935). Structure and Reproduction of the Algae, Volume I. Cambridge University Press,
FRYXELL, G.A., SIMS, P.A. & WATKINS, T.P. (1986). Azpeitia (Bacillariophyceae): Related genera and
GOULD, S.J., GILINSKY, N.L. & GERMAN, S.Z. (1987). Asymmetry of lineages and the direction of
HENNIG, W. (1965). Phylogenetic systematics. Annual Review of Entomology, 10, 97-1 16 HENNIG, W. (1966). Phylogenetic Systematics. University of Illinois Press, Urbana HULL, D.L. (1979). The limits of cladism. Systematic Zoology, 28, 416-440 HUMPHRIES, C. J. (ed.) (1988). Ontogeny and Systematics. Columbia University Press, New York HUMPHRIES, C.J. & CHAPPILL, J.A. (1988). Systematics as science: A response to Cronquist. The
Botanical Review, 54, 129-144 HUSTEDT, F. (1942). Susswasser-Diatomeen des indomalayischen Archipels und der Hawaii-Islen. Znterna-
tionale Revue der gesamten Hydrobiologie und Hvdrographie, 42, 1-252 KLUGE, A.G. (1984). The relevance of parsimony to phylogenetic inference. In: Cladistics: Perspectives on
the Reconstruction of Evolutionary Histoiy (T. Duncan & T. Stuessy, eds.), 24-38. Columbia University Press, New York
Research, 4, 309-347
N y Foljd, 26, 1-194
phylogenetic systematics. Systematic Zoology, 34, 280-299
approach to cladistic analysis. Systematic Botany, 9, 192-202
defense of cladism. The Botanical Review, 54, 107-128
University Press, New York
Platnick & V.A. Funk, eds), 7-36. Columbia University Press, New York
Ecology and Systematics, 14, 313-333
617-626
Cambridge
Promorphology. Systematic Botany Monographs, 13, 1-74
evolutionary time. Science, 236, 1437-1441
KLUGE, A.G. (1985). Ontogeny and Phylogenetic Systematics. Cladistics, 1, 13-28 KOCIOLEK, J.P. (1986). Determining character polarity in diatoms: An alternative approach. Diatom
Research, 1, 189-192
Dow
nloa
ded
by [
Nat
ural
His
tory
Mus
eum
], [
Dr
Dav
id M
. Will
iam
s] a
t 02:
08 0
1 N
ovem
ber
2011

300 J.P. KOCIOLEK, E.C. THERIOT AND D.M. WILLIAMS
KOCIOLEK, J.P. & STOERMER, E.F. (1986). Phylogenetic relationships and classification of monoraphid diatoms based on phenetic and cladistic methodologies. Phycologia, 25, 297-303
KOCIOLEK, J.P. & STOERMER, E.F. (1987). Ultrastructure of Cyrnbella sinuata (Bacillariophyta) and its allies, and their transfer to Reimeria, gen. nov. Systematic Botany, 12, 461- 469
KOCIOLEK, J.P. & STOERMER, E.F. (1988). A preliminary investigation of the phylogenetic relationships of the freshwater, apical pore field-bearing cymbelloid and gomphonemoid diatoms (Bacillariophyceae).
KOCIOLEK, J.P. & STOERMER, E.F. (1989). Phylogenetic relationships and evolutionary history of the
KOCIOLEK, J.P. & WILLIAMS, D.M. (1987). Unicell ontogeny and phylogeny: Examples from the
KRAUS, F. (1988). An empirical evaluation of the use of the ontogeny polarization criterion in phylogenetic
KREBS, W.N., BRADBURY, J.P. & THERIOT, E.C. (1987). Neogene and Quaternary lacustrine diatom
LOTSY, J.P. (1907). Vortruge ziber botanische Stammesgeschichte. Erster Band: Algen und Pike. Gustav Fisher,
LOTSY, J.P. (1916). Evolution by Means of Hybridization. M. Nijhoff, The Hague LUCKOW, M. & PIMENTEL, R.A. (1985). An empirical comparison of numerical Wagner computer
MADDISON, W.P., DONOGHUE, M.J. & MADDISON, D.R. (1984). Outgroup analysis and parsimony.
MANN, D.G. (1986). Methods of sexual reproduction in Nitzschia: Systematic and evolutionary implications
NELSON, G. (1985). Outgroups and ontogeny. Cladistics, 1, 29-45 NELSON, G. & PLATNICK, N.I. (1981). Systematics and Biogeography: Cladistics and Vicariance. Columbia
PLATNICK, N.I. (1987) An empirical comparison of microcomputer parsimony programs. Cladistics, 3, 121-
PLATNICK, N.I. (1988). Programs for quicker relationships. Nature, 335, 310 ROUND, F.E. & CRAWFORD, R.M. (1981). The lines of evolution of the Bacillariophyta I. Origin.
SIMONSEN, R. (1979). The diatom system: ideas on phylogeny. Bacillaria, 2, 9-71 S I M S , P.A. (1986). Sphynctolethus Hanna, Ailuretta gen. nov. and evolutionary trends within the Hemiaulo-
SMITH, H.L. (1872). Conspectus of the families and the genera of the Diatomaceae. The Lens, 1, 72-93 SOBER, E. (1983). Parsimony in statistics: philosophical issues. Annual Review of Ecology and Systematics,
STEVENS, P F. (1284). Homology and phylogeny: morphology and systematics. Systematic Botany, 9 , 395- 409
THERIOT, E. & KOCIOLEK, J.P. (1986). Two new Pliocene species of Cyclostephanos (Bacillariophyceae) with comments on the classification of the freshwater Thalassiosiraceae. Journal of Phycology, 22, 121- 128
THERIOT, E., STOERMER, E.F. & HAKANSSON, H. (1987). Taxonomicinterpretationoftherimoportula of freshwater genera in the centric diatom family Thalassiosiraceae. Diatom Research, 2, 25 1-268
WATROUS, L.E. & WHEELER, Q.D. (1981). The outgroup comparison method of character analysis. Systematic Zoology, 30, 1-1 1
WILEY, E.O. f 1981). I’hylogenetics. The Theory and Practice of Phylogenetic Systematics. J. Wiley & Sons, New York
WILLIAMS, D.M. (1985). Morphology, taxonomy and inter-relationships of the ribbed araphid diatoms from the genera Diatoma and Meridion (Diatomaceae: Bacillariophyta). Bibliotheca Diatomologica, 8, 1- 238
WILLIAMS, D.M. & ROUND, F.E. (1986). Revision of the genus Synedra Ehrenb. Diatom Research, 1,
WILLIAMS, D.M. & ROUND, F.E. (1987). Revision of the genus Fragilaria. Diatom Research, 2, 267-288 WILLIAMS, D.M. & ROUND, F.E. (1988). Phylogenetic systematics of Synedra. In: Proceedings of the 9th
International Diatom Symposium W. E. Round, ed.), 303-315. Biopress, Bristol & Koeltz Scientific Books, Koenigstein
Journal Of Phycology, 24, 377-385
diatom genus Gomphoneis. Phycologia (in press)
diatoms. Cladistics, 3, 274- 284
inference. Systematic Zoology, 37, 1106-41
biochronology, western U.S.A. Palaios, 2, 505-513
Jena
programs. Cladistics, 1, 47-66
Systematic Zoology. 33, 83-103
(Notes for a monograph of the Bacillariaceae 3). Diatom Research, 1, 193-2203
University Press, News York
144
Proceedings of the Royal Society of London, Series B , 211, 237-260
ideae. Diatom Research, 1, 241-269
14, 335-357
313-339
Dow
nloa
ded
by [
Nat
ural
His
tory
Mus
eum
], [
Dr
Dav
id M
. Will
iam
s] a
t 02:
08 0
1 N
ovem
ber
2011
Copyright © 2022 FDOKUMEN




















