
Stealing to Survive: Crime and Income Shocks in 19th Century France

Journal of Plankton Research Vol.18 no.4 pp.597-613. 1996
Can freshwater planktonic ciliates survive on a diet ofpicoplankton?
Karel Simek, Miroslav Macek, Jakob Pern thaler1, Vera Straskrabova andRoland Psenner1
Hydrobiological Institute CAS, Na sddkdch 7, 37005 Ceske Budejovice, CzechRepublic and 'Institute of Zoology, University of Innsbruck, Technikerstrasse 25,A-6020 Innsbruck, Austria
Abstract. Ciliate picoplanktivory was studied in two different freshwater systems with abundant auto-trophic picoplankton (APP): the eutrophic Rimov Reservoir (South Bohemia) during the late summerof 1993 and the oligo- to mesotrophic lake Piburger See (Tyrol) during May 1994. Picoplankton weresized by an image-analysis system and species-specific grazing rates of ciliates on bacteria and APPwere determined using fluorescently labelled prey. According to the grazing data (as the mean uptakerate of picoplankton ind.~' h1), ciliates were split into three ecological groups in order of their decreas-ing picoplanktivory and increasing significance of nanophytoplankton in their diet, (i) Highly efficientfine suspension feeders consisting of vorticellids (4200 bacteria, 560 APP), four oligotrichs andCyclidium sp. (380-1580 bacteria, 57-210 APP), with < 7% of individuals ingesting algae >2 u.m. (ii)Less efficient fine suspension feeders, coarse filter feeders and detritophages consisting ofCinetochilum margaritaceum, Pelagostrombidium fallax, Cyrtolophosis mucicola and Coleps spp.(60-173 bacteria, 2-27 APP), with 7-32% of individuals ingesting algae >2 n.m. (iii) Raptorial feedersconsisting of four prostomatids belonging to the genera Urotricha and Balanion, with a negligibleimportance of picoplanktivory (8-61 bacteria, 0.2-14 APP). With the exception of one prostomatid.>58% of individuals ingested algae >2 (j.m. Grazing data for the six species from the first group wereconverted into organic carbon. Assuming a 35 % gross growth efficiency, the calculated potential doub-ling times were between 29 (Hatteria grandinella) and 43 h (an unidentified oligotrich), except for themixotrophic Pelagohalteria viridis (118 h). The doubling times estimated from changes in ciliate abun-dance for four out of the six species were very close to the calculated values, indicating that this group ofheterotrophic ciliates can meet all of its carbon requirements by feeding exclusively on picoplankton.
Introduction
Ciliates are an important component of the protistan plankton in freshwaters (e.g.Finlay et al., 1988; Beaver and Crisman, 1989). Recent reports indicate that mostplanktonic ciliate communities, at least in meso- to eutrophic freshwater lakes andreservoirs, are numerically dominated by species <20-35 |xm (Beaver andCrisman, 1989; Miiller et al., 1991) or temporarily even smaller species (Miiller,1989; Simek e/a/., 1995).
Small pelagic ciliates can consume prey ranging in size from nanophytoplankton(especially prostomatids and haptorids; Miiller et al., 1991) to picoplankton (mostscuticociliates and oligotrichs, e.g. Sanders et al., 1989; Sherr et al., 1991). Ciliategrazing can thus significantly contribute to the control of biomass and productionof both autotrophic (APP) and heterotrophic picoplankton (e.g. Simek et al., 1990,1995; Sherr et al., 1991; Simek and StraSkrabovd, 1992; Iriberri et al., 1993). How-ever the question of whether picoplankton at natural concentrations might besufficient as the sole food source for ciliate growth has not yet been clearlyaddressed. Bacterial abundances typical for most freshwater and marine ecosys-tems, i.e. 1-5 x 106 cells ml*1, are often not considered to be sufficient for ciliate
© Oxford University Press 597
by guest on July 7, 2013http://plankt.oxfordjournals.org/
Dow
nloaded from

K.Simek el al.
growth (Fenchel, 1980,1986; Rivier etal., 1985; Stoecker, 1988) and only specificenvironments, such as the chemocline, oxycline, etc., were suggested to have suf-ficiently high bacterial abundances for bacterivorous ciliates (Fenchel et al., 1990).On the other hand, some authors consider 1-3 x 106 bacteria ml"1 to be sufficient forthe growth of at least several ciliate species which seem to be voracious bacterialfeeders (Sherr and Sherr, 1987; Sherr et al., 1989; Simek and StraSkrabova", 1992).A potential role of APP in the diet of ciliates has recently been suggested by sev-eral authors (e.g. Stockner, 1988; Sherr et al., 1991; Weisse, 1993). However, it hasnot yet been sufficiently quantified under natural food concentration in fresh-waters which proportion of the carbon requirements of fine suspension-feedingciliates can be met by bacterioplankton and APP.
Preliminary results showed that in the eutrophic Rimov Reservoir, picocyano-bacteria might significantly contribute to the diet of pelagic ciliate communities(dominated by oligotrichs) and that picoplankton as a sole food source might covermost of their carbon requirements (Simek et al., 1995). Here, we present grazingrates of dominant ciliate taxa on bacteria and APP in two systems of differenttrophic state where abundant APP populations occurred, i.e. eutrophic RimovReservoir and the oligo-mesotrophic lake Piburger See. The role of both pico-plankton food items in the diet of particular ciliate taxa was quantified as well asthe proportion of individuals within the respective ciliate taxa ingesting particleslarger than picoplankton (i.e. >2 (Jim). Following these criteria, the natural pelagicciliate communities were split into three ecological groups according to thepreferred size of food particles ingested and the proportion of their organic carbonrequirements that might be covered exclusively from picoplanktivory.
Method
Study sites and sampling
The following study sites were investigated: (i) the eutrophic Rfmov Reservoir(RIM; South Bohemia, Czech Republic, 470 m a.s.l., area 2.06 km2, maximumdepth 43 m, mean depth 16.5 m, dimictic) during 16 August-23 September 1993;(ii) the oligo- to mesotrophic Piburger See (PIB; Tyrol, Austria, 913 m a.s.l., area0.13 km2, maximum depth 24.6 m, mean depth 13.7 m, dimictic) during 4 May-1 June 1994. For a more detailed characterization of both study sites, see Simekand StraSkrabova' (1992) and Sommaruga and Psenner (1995), respectively. Spe-cific features of the dynamics of ciliates, heterotrophic flagellates, bacteria, APP,phyto- and zooplankton during the study periods have already been reported(Simek et al., 1995; Pernthaler et al., 1996).
Samples were taken at the deepest points of both study sites with a 21 Friedingersampler (RIM) or a 5 1 Schindler-Patalas sampler (PIB) three times a week fromtwo layers: the epilimnion (a mixed sample from 0.5,1 and 1.5 m depths) and themetalimnion (a mixed sample from the thermocline and 0.5 m above and belowthis depth) characterized by a concurrent strong decrease of both temperature(PIB) and oxygen (RIM). For a detailed description of the sampling procedures inRIM and PIB, see Simek et al. (1995) and Pernthaler et al. (1996), respectively.Since the dominant ciliate taxa in both systems were species <50 pxa (Table I),
598
by guest on July 7, 2013http://plankt.oxfordjournals.org/
Dow
nloaded from
Diet of pelagic ciliates on picoplankton
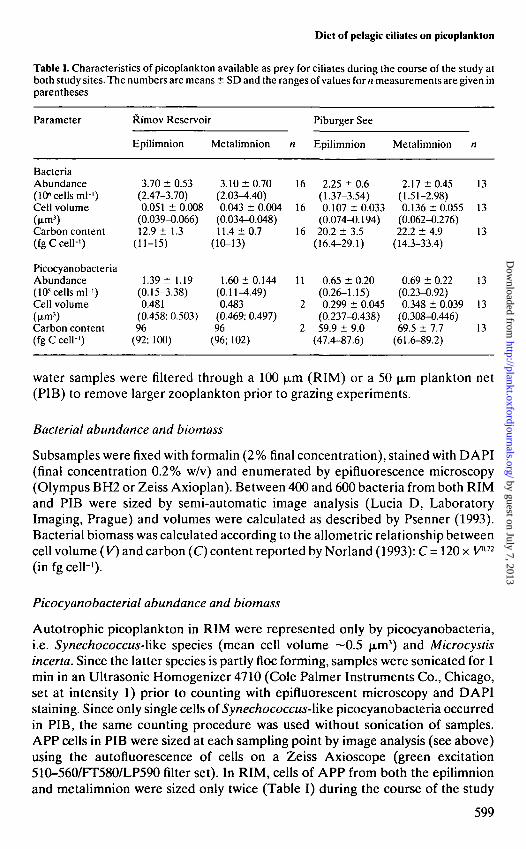
Table I. Characteristics of picoplankton available as prey for ciliates during the course of the study atboth study sites. The numbers are means ± SD and the ranges of values for n measurements are given inparentheses
Parameter
BacteriaAbundance(106 cells ml-')Cell volume(u-m')Carbon content(fgCcell-1)
PicocyanobacteriaAbundance(105 cells ml-1)Cell volume(M-m3)Carbon content(fg C cell-')
Ri'mov Reservoir
Epilimnion
3.70 ± 0.53(2.47-3.70)0.051 ± 0.008
(0.039-0.066)12.9 ± 1.3
(11-15)
1.39 ± 1.19(0.15-3.38)0.481
(0.458; 0.503)96
(92; 100)
Metalimnion
3.10 ± 0.70(2.03-4.40)0.043 ± 0.004
(0.034-0.048)11.4 ±0 .7
(10-13)
1.60 ±0.144(0.11^».49)0.483
(0.469; 0.497)96
(96; 102)
n
16
16
16
11
2
2
Piburger See
Epilimnion
2.25 ± 0.6(1.37-3.54)0.107 ± 0.033
(0.074-0.194)20.2 ± 3.5
(16.4-29.1)
0.65 ± 0.20(0.26-1.15)0.299 ± 0.045
(0.237-0.438)59.9 ± 9.0
(47.4-87.6)
Metalimnion
2.17 ± 0.45(1.51-2.98)0.136 ± 0.055
(0.062-0.276)22.2 ± 4.9
(14.3-33.4)
0.69 ± 0.22(0.23-0.92)0.348 ± 0.039
(0.308-0.446)69.5 ± 7.7
(61.6-89.2)
n
13
13
13
13
13
13
water samples were filtered through a 100 |xm (RIM) or a 50 |xm plankton net(PIB) to remove larger zooplankton prior to grazing experiments.
Bacterial abundance and biomass
Subsamples were fixed with formalin (2% final concentration), stained with DAPI(final concentration 0.2% w/v) and enumerated by epifluorescence microscopy(Olympus BH2 or Zeiss Axioplan). Between 400 and 600 bacteria from both RIMand PIB were sized by semi-automatic image analysis (Lucia D, LaboratoryImaging, Prague) and volumes were calculated as described by Psenner (1993).Bacterial biomass was calculated according to the allometric relationship betweencell volume (V) and carbon (C) content reported by Norland (1993): C = 120 x V-72
(in fg cell"1).
Picocyanobacterial abundance and biomass
Autotrophic picoplankton in RIM were represented only by picocyanobacteria,i.e. Synechococcus-like species (mean cell volume ~0.5 u.m3) and Microcystisincerta. Since the latter species is partly floe forming, samples were sonicated for 1min in an Ultrasonic Homogenizer 4710 (Cole Palmer Instruments Co., Chicago,set at intensity 1) prior to counting with epifluorescent microscopy and DAPIstaining. Since only single cells of Synechococcus-\ike picocyanobacteria occurredin PIB, the same counting procedure was used without sonication of samples.APP cells in PIB were sized at each sampling point by image analysis (see above)using the autofluorescence of cells on a Zeiss Axioscope (green excitation510-560/FT580/LP590 filter set). In RIM, cells of APP from both the epilimnionand metalimnion were sized only twice (Table I) during the course of the study
599
by guest on July 7, 2013http://plankt.oxfordjournals.org/
Dow
nloaded from

K.Simek el al.
using a fine-scale eyepiece micrometer. Cell volumes of APP were transformed tocarbon using a conversion factor of 200 fg C ixnr3 (Weisse, 1993).
Protargol staining
Ciliate samples were fixed directly (RIM) or post-fixed (PIB) with Bouin's fixative(5% final concentration). In the latter case, acid Lugol's solution was used as afixative, the samples were sedimented and concentrated 10 times, then the Bouin'sfixative was added. Ten to 40 millilitres of a sample were filtered onto a nitro-cellulose membrane filter (1.2 \im pore size; Millipore, USA), mounted in agar,fixed with formalin and protargol stained at 60°C according to Skibbe's (1994)modification of the method of Montagnes and Lynn (1987).
Ciliate determination, abundance and grazing
The use of protargol staining (above) and live sample observation in parallel withfluorescent microscopy allowed us to determine most of the ciliates to the level ofgenus and, in many cases, to species (Table II). Nevertheless, usually between 5and 25% of ciliates could not be identified. Since it is difficult to identify ciliates inthe fluorescent microscope, we used additional criteria for determination, such assize and position of nuclei, presence, size and shape of chloroplasts in mixotrophicciliates (especially in PIB), and the way in which prey were arranged in the foodvacuoles, so we were able to distinguish the dominant taxa determined with theprotargol technique and live observation (Table II). Moreover, most of the domi-nant ciliate species in RIM and PIB have been isolated, or at least shortly kept incultures and identified; these reference cultures were used to confirm determi-nations with protargol staining and fluorescent microscopy. Genus and speciesidentifications are based on publications by Small and Lynn (1985), Foissner et al.,(1991,1992,1994) and references therein.
Two different kinds of prey were used to measure protozoan grazing on pico-plankton: fluorescently labelled bacterioplankton (FLB) and fluorescentlylabelled Synechococcus-Wkz cyanobacteria (FLC). For details of the concentrationprocedure for bacterioplankton from RIM, see Simek and StraSkrabova (1992).Bacteria from PIB were concentrated using an Amicon hollow-fibre filter appar-atus as described by Sommaruga and Psenner (1993).
A round-shaped Synechococcus sp. isolated from RIM (mean cell volume ± SD:0.48 ± 0.23 u.m3) and a rod-shaped Synechococcus sp. (Culture Collection,Institute of Botany, Tfebofi; mean cell volume ± SD: 0.32 ± 0.13 |im3) were grownin cultures and harvested by centrifugation. The former species of Synechococcuswas used as labelled prey in RIM and the latter one in PIB (see Table I for APPmean cell sizes). Both bacterial and cyanobacterial prey were fluorescentlylabelled according to the protocol of Sherr and Sherr (1993).
For grazing experiments, 500 ml samples were dispensed into acid-soaked andrinsed 11 flasks. To avoid possible interference of the labelled food particles, ciliateuptake rates were determined separately for the two different tracers used—FLBand FLC. For details of the grazing experiments, see Simek et al. (1995) andPernthaler et al. (1996). Thirty (RIM) to 50 (PIB) ml subsamples for protozoanenumeration and tracer ingestion determinations were taken at 0, 3,6,10, 20 and
600
by guest on July 7, 2013http://plankt.oxfordjournals.org/
Dow
nloaded from

Diet of pelagic ciliates on picoplanklon
Table II. Mean cell volume (1 SD in parentheses) of the major ciliate taxa found in the ftfmov Reservoir(RIM) and Piburger See (PIB)
Taxa
OligotrichidaPelagohalteria viridisHalteria grandinellaStrobilidium hexakinetumOligotrichous ciliate on
SiaurastrumPelagostrombidium fallax
PeritrichidaVorticella aquadulcis complex
CyrtolophosididaCyrtolophosis mucicola
ScuticociliatidaCinetochilum margaritaceumCyclidium sp.
ProstomateaUrotricha spp. 1 (U.furcata >
90%)Urotricha spp. 2 (U.furcata >
60%)Urotricha sp.. smallBalanion planclonicumColeps sp.Coleps spetai
Abbreviation
PVHGSH
OSPF
VO
CM
CICY
UR1
UR2UNBACOCS
Studysite
PIBRIMRIM
RIMPIB
RIM
RIM
RIMRIM
RIM
PIBRIMPIBRIMPIB
Cell volume(urn1)
520728601230
153024 600
8940
2050
32901070
930
1167450
111428503050
(2870)(1380)(690)
(1130)(11000)
(4940)
(1590)
(1400)(510)
(690)
(1108)(210)(498)
(1890)(1826)
n
226235
99
4535
33
146
10646
234
16072
2248732
30 min after tracer addition, and fixed by adding 0.5% of alkaline Lugol's solution,followed during 2-3 s by 2% borate-buffered formalin (final concentration) andseveral drops of 3% sodium thiosulphate to clear the Lugol's colour (Sherr andSherr, 1993).
Ciliate grazing rates were usually determined in the time series from 3-10 minsubsamples. The time series from 10-30 min subsamples were used only whenciliate assemblages were dominated by species with a low capture efficiency onpicoplankton. Twenty to 40 ml aliquots were stained with DAPI, filtered through 2(xm pore size black Poretics filters and inspected via epifluorescence microscopy.At least 30 ciliates were inspected for FLB and FLC ingestion in each sample.
Owing to the low number of ciliate individuals of each taxon inspected forgrazing per sample, all grazing data of single taxa of unambiguously identifiedciliates were pooled to calculate individual grazing rates at the genus or specieslevel. To estimate mean cell volumes, all ciliates inspected for tracer uptake inpreserved samples were measured and the volumes were calculated by approxi-mation to prolate spheroids.
Estimation of doubling times of fine suspension-feeding ciliates
Based on their picoplankton consumption. For the six species that showed exclus-ive or predominant consumption of picoplankton in both study sites (Table III),
601
by guest on July 7, 2013http://plankt.oxfordjournals.org/
Dow
nloaded from

Tab
le II
I. M
ean
graz
ing
rate
s (C
R ±
1 S
D) a
nd c
lear
ance
rate
s (C
R) o
f cili
ates
on
bact
eria
(B
Arr).
pic
ocya
noba
cter
ia (
APP
). C
/?A
PP/C
/?B
An
ratio
, and
the
prop
ortio
n of
algi
vory
. i.e
. per
cen
t of
indi
vidu
als
taki
ng u
p ph
ytop
lank
ton
larg
er th
an 2
p,m
Gra
zing
and
cle
aran
ce r
ates
on
Bac
teri
a
GR
BA
CT
(bac
teri
a ce
ll ' h
Hig
hly
effic
ient
fin
e su
spen
sion
fee
ders
Pel
agoh
alte
ria
viri
dis
Hal
teri
a gr
andi
nell
aSt
robi
lidi
umhe
xaki
netu
mO
ligot
rich
ous
cilia
te o
nSt
aura
stru
mC
ycli
dium
sp.
Vor
tice
lla
aqua
dulc
isco
mpl
ex
799
± 46
915
80 ±
1110
380
± 27
0
440
± 61
0
470
± 35
042
00 i
3800
Les
s ef
ficie
nt f
ine
susp
ensi
on f
eede
rs, c
oars
eP
elag
oslr
ombi
dium
fall
axC
ynol
opho
sis
muc
icol
aC
olep
s sp
.C
olep
s sp
etai
Cin
etoc
hilu
mm
arga
rita
ceum
Rap
tori
al f
eede
rsU
rolr
icha
spp
. 1
(U.fu
rcat
a> 9
0%)
Uro
tric
ha s
pp. 2
(U.fu
rcal
a >
60%
)U
rolr
icha
sp.
, sm
all
Bal
anio
n pl
anct
onic
um
136
± 10
7
173
± 12
463
±8
762
±8
557
±8
8
23 ±
64
10 ±
21
61 ±
108
8 ±
12
CRB
ACT
') (n
lcel
l'h
353
465
111
131
139
1240
filte
r fe
eder
s.62 51 19 28 19 7 5 18 4
pico
plan
kton
n 114
118 72 29 30 23
AP
P
G/?
AP
P
(AP
P ce
ll-1 h
-1)
57 ±
25
210
± 15
676
± 5
7
90 ±
121
80 +
45
560
± 63
0
detr
itop
hage
s19 115 65 17 76 151 87 49 122
16 +
21
27 +
37
8±
112
± 3
7±
12
2±
7
2±
4
14 ±
29
0.2
± 1
C/?
AP
P
(nl
cell
-
851
1180 426
504
448
31
50
239
151 45 33 39 11 30 78 3
n•h
-)
112
117 27 16 16 10 14 30 22 15 30 83 74 23 99
Ra
tio
C/?
AP
P/C
/?1
)AC
T
2.4
2.5
3.8
3.9
3.2
2.5
3.9
3.0
2.4
1.2 2.3
1.7 6.0
4.4
0.8
Ob
serv
ed
alg
ivo
ry
Ing
esti
on
of
alga
e >2
n.m
(%)
0 2 3 7 0 3 18 7 9 12 32 62 58 8 66
Tot
al c
iliat
esin
spec
ted
(")
226
235 99 45 46 33 33 146 87 32 106
234
161 72 221
by guest on July 7, 2013http://plankt.oxfordjournals.org/Downloaded from

Diet of pelagic ciliates on picoplankton
doubling times were calculated based on the amount of bacterial and APP carbonconsumed. The amount of organic carbon consumed by individual ciliates was cal-culated from grazing rates multiplied by the amount of organic carbon per foodparticle (bacteria or APP). Since species-specific cell volumes and grazing rates ofciliates on both food items were not significantly different in the epilimnion andmetalimnion (r-test, P < 0.05), the data from both layers were pooled to calculatedoubling times by using the means for all inspected individuals of the respectivespecies or taxon. As grazing rates on FLB and FLC were measured separately, wereceived two sets of data characterizing the respective amount of organic carboningested. The sum of carbon in both food items and the cell volumes of ciliates wereused to calculate potential doubling times of different ciliate taxa. Cell volumes ofciliates were transformed to carbon using a conversion factor of 140 fg C |inr3
recommended for formalin-preserved samples (Putt and Stoecker, 1989). To con-vert the carbon data into potential doubling times of the typical picoplankton-feeding ciliates, we used three values of gross growth efficiency (GGE) for bacteri-vorous ciliates, ranging from very low (19%; Turley etal., 1986), to medium (35%)and high (50%; reported by Fenchel, 1987).
Based on changes in cell number. The growth rate of ciliates was estimated in 500ml of a 100 (xm screened water sample incubated in the dark at in situ temperaturefor 1-2 days (RIM) or in 500 ml of a 50 u.m screened water sample exposed in whitebottles in situ for 2-3 days (PIB). For details, see Macek et al. (1996). Numbers ofciliates were evaluated by the quantitative protargol staining method (Montagnesand Lynn, 1987). Population growth rates (JJL) for the dominant species were calcu-lated from changes in abundance, assuming exponential growth: (x = (In N, - InN0)/t, where N, and No are the numbers of ciliates at the start and end of the exper-iment, and / is the duration of the experiment. Doubling time (T) was calculatedaccording to the formula T - In 21 \L.
Results and discussion
Characteristics of picoplankton available as prey for ciliates
The studied systems are different considering their size, altitude, and especiallytheir trophy. The chosen periods were also different: the second half of the summerphytoplankton peak in RIM (water temperatures in the epi- and metalimnionwere between 15.3 and 22.2°C, and 13.8 and 20.2°C, respectively) and the late stageof the spring phytoplankton peak in PIB (water temperatures in the epi- andmetalimnion were between 13.5 and 16.5°C, and 9.6 and 14.6°C, respectively).While chlorophyll a concentrations in the epilimnion and metalimnion of RIMwere in the range from 7 to 112 (xg I"1 during the study period (Simek et al., 1995),these values in PIB were mostly between 2 and 4 jxg I"1 (Pernthaler et al., 1996).Nevertheless, numbers and cell sizes of picoplankton available as prey for ciliatesdiffered by a factor of less than three at both sites (Table I).
In general, there were only minor differences between the epilimnetic and meta-limnetic water layers at both study sites. When comparing RIM with PIB, bacterialabundances were on average >50% higher in RIM, but the mean cell
603
by guest on July 7, 2013http://plankt.oxfordjournals.org/
Dow
nloaded from

K.Simek et al.
volumes of bacteria showed an opposite trend and were more than twice as large inPIB. Bacterial carbon content per cell in RIM was 50% smaller than in PIB. As aresult, the total amount of bacterial carbon available was very similar at both studysites with minor oscillations between 30 and 60 (xg I"1 (Figure 1). While the bacterialabundances are typical for these sites as previously reported by Simek et al. (1990)and Simek and Straskrabova (1992) for RIM, and by Sommaruga and Psenner(1993, 1995) for PIB, bacterial cell volumes, compared with those in the abovereports, seem to be rather low in RIM, but high in PIB (Pernthaler et al., 1996).Since bacterioplankton at both sites were actively growing with doubling timesmostly between 2 and 4 days (Simek et al., 1995; Pernthaler et al., 1996), a possibleexplanation for the small bacterial cell volumes in RIM might be a very stronggrazing impact of protozoa dominated by heterotrophic flagellates, as also noted inthe summer of 1990 by Straskrabova et al. (1993). As shown by Simek andChrzanowksi (1992) and Simek et al. (1994), both flagellates and ciliates in fresh-waters prefer larger cells when feeding on bacterioplankton. On average >90% ofbacterial production in RIM and roughly 70% of bacterial production in PIB wereconsumed by protozoa (Simek et al., 1995; Pernthaler et al., 1996). On the otherhand, the unusual increase in bacterial cell volumes, especially in the metalimnionof PIB, seemed to be affected by an increased proportion of long rod-shapedbacteria (J.Pernthaler et al., in preparation).
There are more differences apparent in parameters of APP (Table I, Figure 1).Their mean abundances in RIM were >2-fold higher and mean cell volumes were~50% larger than in PIB. Generally, however, APP abundances at both study sitesfit well to the range of values reported by Weisse (1993, and references therein) forlakes and reservoirs of corresponding trophy. The mean abundances in RIM werecalculated from 11 sampling dates out of 16, i.e. when it was possible to use thetracer technique. Owing to the very low number of APP in RIM during the last fivesampling dates (<1.7 x 104 cells ml"1; Simek et al., 1995), grazing rates on FLC werenot measured for this period. Compared to the relatively stable amount of organiccarbon available in bacterial prey, the dynamics of APP was remarkably different.In RIM, the amount of organic carbon in APP constituted between 16 and 41% oftotal organic carbon in picoplankton during the first half of the study (correspond-ing absolute values were between 9 and 27 |xg C I"1), while its proportion fell tonegligible levels during the second half of the study (Figure 1). An opposite trendwith slowly increasing proportions of organic carbon in APP (correspondingabsolute values were between 2 and 9 u,g C H) was found in PIB, although itsproportion of total picoplankton carbon never exceeded 17%.
Ciliate grazing rates on picoplankton
Total ciliate numbers, the proportion of the respective species and the populationdynamics of the dominant ciliate taxa in RIM and PIB have been described else-where (Simek et al., 1995; Macek etal., 1996). The mean cell volumes of the 15 mostabundant ciliate taxa (Table II) ranged from 450 u.m3 for small Urotricha sp. to24 600 jjim3 for Pelagostrombidium fallax. Each specimen of these taxa wasinspected for the uptake of FLB and FLC to calculate species-specific grazing rateson both food items. For statistical reasons, only those species or taxa were used for
604
by guest on July 7, 2013http://plankt.oxfordjournals.org/
Dow
nloaded from

100
80-
22
60
O>
40 20
Aim
ov R
ese
rvoir
Epi
limnl
on
Pic
ocyanobacte
ria
Bacte
ria
Met
alim
nion
16
23
30
Au
gu
st
6
13
22
Se
pte
mb
er
100 20
100' 80 60 40 20
Pib
urg
erS
ee
Epi
limni
on
Met
alim
nion
1118
May
25
1Ju
ne
Fig
. 1.
Tim
e co
urse
of
orga
nic
carb
on a
vail
able
to
cili
ates
in
the
form
of
bact
eria
or
pico
cyan
obac
teri
a in
the
Ri'm
ov R
eser
voir
an
dP
ibur
ger
See
.
I- 2. 3
by guest on July 7, 2013http://plankt.oxfordjournals.org/Downloaded from

k.Simek el al.
calculations, from which at least 10 individuals were inspected for uptake of bothFLB and FLC (Table III). In seven out of 15 cases 106-235 individuals per taxonwere inspected, in three cases 72-99 individuals and in five cases 32-46 individuals.To document possible algivory, the content of food vacuoles was inspected for thepresence of chlorophyll a-containing particles >2 (xm (presumably algae; TableIII).
Generally, grazing rates on APP were roughly one order of magnitude lowerthan those on bacteria for most of the inspected ciliate taxa. Clearance rates forAPP, however, were significantly higher than for bacteria (Table III). The ciliatetaxa were split into three ecologically different groups partly based on publishedreviews (Fenchel, 1987; Sanders et al., 1989; Foissner et al., 1991,1994, and refer-ences therein), but preferentially by using our measured feeding rates on bacteriaand APP (Table III). The first group, i.e. highly efficient fine suspension feeders,was characterized by the highest grazing and clearance rates on both picoplanktonfood particles along with the lowest percentage (0-7%) of observed algivory(Table III, Figure 2). This assemblage was represented by four oligotrichous ciliatespecies, one scuticociliate, Cyclidium sp., and one peritrichous ciliate, Vorticellaaquadulcis complex. The highest species-specific grazing and clearance rates wereobserved in vorticellids (4200 bacteria and 560 APP cell"1 lr1, corresponding toclearance rates of 1240 and 3150 nl cell"1 lr1), followed by oligotrichs (380-1580bacteria and 57-210 APP cell1 h1, 111-456 and 423-1180 nl cell-' Ir1) andCyclidium sp. (470 bacteria and 80 APP cell"1 h-\ 139 and 448 nl cell"1 Ir1). Amongthe oligotrichs, two closely related species, Pelagohalteria viridis (PIB) and Halte-ria grandinella (RIM), showed the highest grazing rates on bacteria and APP inboth systems (Table III). The mean grazing rate of H.grandinella on bacteria inRIM (Table III) was significantly higher than the range of grazing rates reportedfor this ciliate by Sanders et al. (1989) in the eutrophic Lake Oglethorpe. Two verysmall oligotrichs (Table II), Strobilidium hexakinetum and an unidentified oligo-trich attached by a stalk to Staurastrum cells, grazed 380 and 440 bacteria cell"' lr1,and 76 and 90 APP cell"1 lr1, respectively.
While five taxa of the highly efficient fine suspension-feeding ciliates were foundto be abundant in the eutrophic RIM, only one species (P.viridis) matched thiscategory in the oligo-mesotrophic PIB. This is in agreement with the hypothesis ofBeaver and Crisman (1989) on the proportion of bacterivorous ciliates in lakes ofdifferent trophy. Among 226 inspected individuals of mixotrophic P.viridis, noneingested algal particles >2 u.m (Table III). Volume-specific clearance rates ofP. viridis were ~2-3 times lower than those calculated for other highly efficient finesuspension feeders (Figure 2), which should probably be compensated for by itsmixotrophy since this species (and similarly Coleps spetai, both from PIB) con-tained numerous zoochlorellae (see also Foissner, 1994). Our data, especially thevolume-specific clearance rates of two frequently confused species, H.grandinellaand P.viridis, can also provide a good ecological reason justifying the taxonomicseparation of these species (Foissner et al., 1991; Foissner, 1994). Almost no dataabout feeding rates of these species under natural picoplankton concentrationshave been reported, except for relatively low clearance rates of H.grandinella onmicrospheres and FLB (Sanders et al., 1989).
606
by guest on July 7, 2013http://plankt.oxfordjournals.org/
Dow
nloaded from

Diet of pelagic ciliates on picoplankton
VSCRon bacterioplankton (10 h )
Fig. 2. Comparison of volume-specific clearance rates (VSCR) of the 15 ciliate species from the RimovReservoir and Piburger See calculated from ingestion rates measured on bacterioplankton and pico-cyanobacteria. Inset: Detail for seven species with low VSCR. PV, Pelagohalteria viridis; HG, Halteriagrandinella; SH, Strobilidium hexakinetum; OS, oligotrich on Staurastrum; CY, Cyclidium sp.; VO,Vorticella aquadulcis complex; PF, Pelagostrombidium fallax; CM, Cyrtolophosis mucicola; CI, Cine-tochilum margaritaceum; CO, Coleps sp.; CS, Coleps spelai; UR1, Urotricha sp. 1: UR2, Urotricha sp. 2;UN, Urotricha sp., small; BA, Balanion planctonicum.
In the second group, we listed less efficient fine suspension feeders, coarse filterfeeders and detritophagous ciliate species (Fenchel, 1968; Foissner et al., 1991,1994). Not only the latter references, but also our measured grazing and clearancerates, indicate that picoplankton probably played only an additional, minor role inthe carbon requirements of these species. This group is characterized by low tomoderate grazing rates on bacteria and APP (57-173 bacteria cell"1 h 1 and 2-27APP cell"1 h"1; Table III). Except for a medium-sized oligotrich, P.fallax, all othermembers of the group are rather small species with cell volumes around 2000-3200\im\ Clearance rates, 19-62 nl cell"1 h"1 for bacteria and 33-239 nl cell"1 h1 for APP,were significantly lower compared to those found for species listed in the firstgroup. Volume-specific clearance rates on both food items (0.5-2 x 104 rr1; Figure2) for all but one species {Cyrtolophosis mucicola) were rather low. We observed aremarkably higher proportion of individual ciliates from this group that ingestedalgal particles >2 |xm (7-32%; Table III). From an ecological point of view, theclassification of the scuticociliate Cinetochilum margaritaceum as a detritophagousspecies seems to be adequate (reviewed by Foissner et al., 1994), as we frequentlyobserved this species with ingested single cells of Microcystis aeruginosa, whichcreated large floes during the study in RIM.
The third group, raptorial feeders, is represented by typical algivorous prosto-matids belonging to the genera Urotricha and Balanion (Miiller, 1991; Miiller andGeller, 1993). Correspondingly, this group can be characterized by the lowest cap-ture efficiency on both picoplankton food particles, by the highest proportion ofindividuals with ingested algae >2 u.m, and by very low values of clearance rates on
607
by guest on July 7, 2013http://plankt.oxfordjournals.org/
Dow
nloaded from

K.Simek el al.
both picoplankton food items (Figure 2, Table III). Thus, the data clearly indicatethe negligible role of picoplankton in the diet of these ciliates. The only exceptionto this pattern was the smallest Urotricha sp. (mean cell volume 450 (jun3), forwhich picoplanktivory might be of some significance. This ciliate showed muchhigher respective clearance rates on picoplankton than other prostomatids (Figure2, Table III) and also <10% of all individuals ingested algae >2 (xm. This findingseems to confirm the conclusions of several authors that especially small species ofUrotricha might be partly bacterivorous (Muller, 1989; Foissner et al., 1994). Therates of bacterivory of prostomatids in our study were generally 2-4 times lowerthan the grazing rates of a ciliate community dominated by U.furcata andB.planctonicum previously reported for PIB by Sommaruga and Psenner (1993).In the latter study, however, not species-specific grazing rates, but only mean graz-ing rates were calculated for the whole ciliate community where picoplanktivorousP.viridis occurred as well.
The role of picocyanobacteria in the diet of ciliates
Ciliates appear to be significant grazers of algal picoplankton, as recently reportedfrom both marine and freshwaters (Stockner, 1988; Rassoulzadegan et al., 1988;Weisse, 1993; and references therein). Our data in Table III and Figure 3 clearlydemonstrate the importance of picocyanobacteria as a potential carbon source forciliates. Assuming exclusive picoplanktivory for the six fine suspension feedersfrom RIM and PIB (i.e. that the sum of carbon in bacteria and APP equals 100% ofthe carbon ingested), which in fact holds for P. viridis and Cyclidium sp. as well, wecalculated that APP constituted 48-60% of the total prokaryote organic carboningested (Figure 3). The only exception from the above range was P. viridis, wherecarbon ingested from APP constituted only 20%. APP were not only ingested, butalso digested by ciliates, as we frequently observed autofluorescing APP cells atvarious degrees of digestion, especially in food vacuoles of H.grandinella, P. viridisand S.hexakinetum.
For all six highly efficient fine suspension feeders from both study sites thereexists, however, a large discrepancy between the proportion of APP biomass inboth pelagic environments and in the diet of the ciliates (cf. Figures 1 and 3). Sig-nificantly higher clearance rates for APP compared to those calculated for bacteriaindicate strong size selection towards larger picoplankton prey (Table III, Figure2). This trend holds for all but one {B.planctonicum) ciliate species independentlyof their feeding mode and capture efficiency on picoplankton (Table III, Figure 2).In most cases, our data indicate 2-4 times higher clearance rates on APP. It hasalready been reported from laboratory experiments that Cyclidium speciesisolated from RIM prefer larger bacterioplankton cells (Simek et al., 1994).
Doubling times of fine suspension-feeding ciliates based on their picoplanktivory
The question of whether ciliates can survive exclusively on pelagic bacteria hasrarely been addressed and conclusions are inconsistent (e.g. Fenchel, 1980; Sherrand Sherr, 1987). Several authors suggested that ciliates cannot survive in pelagicconditions exclusively on a diet of bacteria (Fenchel, 1980,1986; Rivier et al., 1985;
608
by guest on July 7, 2013http://plankt.oxfordjournals.org/
Dow
nloaded from

Diet of pelagic ciliates on picoplankton
100
C 40O
(6O 20
PV HG SH OS CY VO
Bacteria j Picocyanobacteria
Fig. 3. Proportion of organic carbon in bacteria and picocyanobacteria ingested in picoplankton by sixfine suspension-feeding ciliates from the Rimov Reservoir and Piburger See. PV, P.viridis; HG,H.grandinella; SH, S.hexakinetum; OS, oligotrich on Staurastrum; CY, Cyclidium sp.; VO,V.aquadulcis complex.
Stoecker, 1988), but the organic carbon available in APP has not been involved insuch considerations. The average concentrations of bacteria in RIM and PIB werein the range 2.2-3.7 x 106 ml"', and a significant amount of organic carbon wasavailable in the biomass of APP (Figure 1). Our values of volume-specific clear-ance rates for the six fine suspension-feeding species, from 0.7 to 1.6 x 10s Ir1 forbacteria and from 1.6 to 4.2 x 105 Ir1 for APP (Figure 2), indicate a very highcapture efficiency of these species on picoplankton. These values are —1-2 ordersof magnitude higher than those estimated by Fenchel (1980) for ciliates feeding onbacteria. On the other hand, the range of values agrees very well with thosepublished for several fine suspension-feeding ciliates from marine waters, whenFLB were used as prey (Sherr and Sherr, 1987).
High densities of APP in both systems raised a question of whether or not thecarbon consumed as picoplankton would be sufficient as a sole food source to sup-port the growth of the six predominantly picoplanktivorous ciliate species. If theyingested some algae, then very rarely (Table III) and moreover only cells <4 u,m,which were generally very scarce in the phytoplankton assemblages of both studysites (Simek et al., 1995; Pernthaler et al., 1996).
Species-specific grazing rates on both food items and cell volumes of the sixciliate species were converted into carbon units. Certainly, we are aware of prob-lems of this approach as it requires the use of various conversion factors. Anotherassumption in our protocol is the value of a GGE to convert carbon data intopotential doubling times of the typical picoplankton-feeding ciliates, since thisparameter is partly species specific (Fenchel, 1987) and also depends on the qualityof the prey and the growth rate and phase. Moreover, most of the values of GGEfor bacterivorous ciliates are based on laboratory cultures and not measured undernatural pelagic conditions. Therefore, to convert the carbon data into potential
609
by guest on July 7, 2013http://plankt.oxfordjournals.org/
Dow
nloaded from

K.Simek el al.
doubling times, we used three published values of GGE for bacterivorous ciliates(Figure 4), ranging from very low (i.e. 19%; Turley etal., 1986), to medium (35%;in the range reported by Fenchel, 1987) and high (50%; Fenchel, 1987). Themedium value, i.e. 35%, was used as the mean between both published extremes.Thus, the potential calculated doubling times for the five species from RIM (35%of GGE; Figure 4) would be in the range from 29 h (H.grandinella) to 43 h (for theunidentified oligotrich on Staurastrum). The corresponding potential doublingtimes using the highest and the lowest values of GGE (i.e. 50 and 19%) would be inthe range from 20 to 30 h and 53 to 79 h, respectively. These values fit very wellthose found for oligotrichs from Lake Constance estimated from changes in ciliateabundance with time (Miiller, 1989). The shortest potential doubling times wereestimated for H.grandinella, Cyclidium sp. and V.aquadulcis complex, i.e. forciliates with the lowest percentage of algivory (Figure 4). It has already beenreported that Cyclidium sp. could meet its carbon requirements exclusively frombacteria in RIM (Simek and Straskrabova', 1992). Only one species from our groupof highly efficient fine suspension feeders, P.viridis, showed significantly longercalculated doubling times than the others, i.e. 216,118 and 82 h at a GGE of 19,35and 50%, respectively (Figure 4). However, as already noted, this species is mixo-trophic and hence the calculated doubling times are probably underestimated.
To confirm our estimates of doubling times of five ciliate species based on pico-plankton uptake (H.grandinella, S.hexakinetum, P.viridis, V.aquadulcis complexand Cyclidium sp.), doubling times were independently measured from changes intheir abundance after the removal of larger zooplankton (Figure 4). For four out offive species (H.grandinella, S.hexakinetum, V.aquadulcis complex and Cyclidiumsp.), the values measured (41, 49, 23 and 21 h, respectively) matched the range ofvalues calculated on the basis of their picoplanktivory. Assuming exclusive pico-planktivory of these four species, the calculated corresponding values of GGEwould be 25,29,46 and 48%, respectively. The latter values have to be consideredcautiously, since these species, except for Cyclidium sp., were not exclusively pico-planktivorous, which might partly cause an overestimation of the GGE values.However, generally only few individuals of these species ingested algae >2 ^m(Table III). On the other hand, the very high calculated values of GGE forV.aquadulcis complex and Cyclidium sp. could be partly influenced by fastergrowth of these species in bottles compared to natural conditions (for details, seeMacek el al., 1996). Another potential bias of our data might be that ciliates couldhave consumed heterotrophic flagellates, which were dominated in RIM and PIBby small species of diameter 3-8 u,m (not shown). As this group of protozoa wasalways 1-2 orders of magnitude less abundant than APP at both study sites (Simeket al., 1995; Pernthaler et al., 1996), and the fine suspension-feeding ciliates wereobserved to ingest only particle <4 p.m, this source of error should be insignificant.
Since the subsamples from RIM were incubated after being screened through a100 (im mesh, some rotifers passed the net (Macek et al., 1996). This might be amore significant bias leading to underestimation of the growth rate of the fourciliate species from RIM calculated from the changes in cell numbers (Figure 4), assmall species of ciliates might be efficiently preyed upon by rotifers (Arndt, 1993;Sanders and Wickham, 1993).
610
by guest on July 7, 2013http://plankt.oxfordjournals.org/
Dow
nloaded from

120 -
Diet of pelagic ciliales on picoplankton
SH
19% 35% 50% 19% 35% 50% 19% 35% 50%
Fig. 4. Potential doubling times of the six typical picoplankton-feeding ciliates. Values of gross growthefficiency for bacterivorous ciliates cover the whole range of published values, i.e. from the lowest(19%; Turley et al., 1986), to the medium (35%) and highest (50%; Fenchel, 1987). For all but one, anoligotrich on Staurastrum, the calculated values based on their picoplanktivory are compared withthose measured on the basis of changes in ciliate abundance with time, indicated by a thick horizontalline. The key is the same as in Figure 3.
For P.viridis, the discrepancy between the doubling times estimated fromchanges in ciliate cell numbers and those based on picoplanktivory (Figure 4) isunlikely to be explained by the consumption of heterotrophic flagellates. This spe-cies was observed to be the exclusive picoplanktivore in PIB (see Table III). Norotifers were observed before and after incubation of in situ exposed samplesscreened through a 50 u.m mesh. Apparently, the main source of the discrepancybetween doubling times estimated by the two different approaches was the mixo-trophy of this ciliate. In concordance, almost no growth of P. viridis was observed inbottles exposed at the lower light intensities in the metalimnion (Macek et al.,1996). However, it was impossible to quantify directly the contribution of autotro-phy to the diet of this ciliate. Therefore, using the same approach as above andassuming 35% as the most realistic estimate of the GGE, we calculated that auto-trophy would cover roughly 43% of P.viridis carbon requirements.
Bacterivory by oligotrich ciliates has been reported for many species (e.g.Fenchel and Jonsson, 1988; Pierce and Turner, 1992; and references therein) andpicoplanktivory in general was found to be prevailing for most oligotrichs <30 u.m(Rassoulzadegan et al., 1988). We conclude that our data clearly show the ability offive out of six picoplanktivorous species, dominated by oligotrichs, to meet allcarbon requirements exclusively from a picoplankton diet, especially when abun-dant APP populations occur. In the special case of the mixotrophic ciliate P. viridis,in PIB picoplanktivory may have supplied >50% of its carbon requirements.
611
by guest on July 7, 2013http://plankt.oxfordjournals.org/
Dow
nloaded from

K.Simek et a I.
Acknowledgements
We thank Wilhelm Foissner, Helga Miiller, John Stockner and Jaroslav Vrba forcomments on earlier versions of the manuscript, Wilhelm Foissner also for helpwith the identification of several ciliate species and Angela Schwarzenbacher forsizing of picoplankton. We also thank Helga Miiller for supplying the referencecultures of U.furcata and B.planctonicum, and Ivona Stupkov& for excellentlaboratory assistance. This study was supported by the Austrian Ministry ofResearch, EAST-WEST PROGRAM, 'Bacterial Grazing by Protozoa' (GZ45.281/2-IV/93), and partly by a CAS GA research grant no. 617102 and a GA CRresearch grant no. 204/94/1672.
ReferencesArndt.H. (1993) Rotifers as predators on components of the microbial web (bacteria, heterotrophic
flagellates, ciliates)—a review. Hydrobiologia, 255/256, 231-246.BeaverJ.R. and Crisman.T.L. (1989) The role of ciliated protozoa in pelagic freshwater ecosystems.
Microb. Ecol., 17,111-136.Fenchel.T. (1968) The ecology of marine microbenthos II. The food of marine benthic ciliates. Ophelia,
5,73-121.Fenchel.T. (1980) Suspension feeding in ciliated protozoa: Feeding rates and their ecological signifi-
cance. Microb. Ecol., 6, 13-25.Fenchel,T. (1986) Protozoan filter feeding. Prog. Protistol., 1, 65-113.Fenchel.T. (1987) Ecology of Protozoa: The Biology of Free-living Phagotrophic Prolists. Science
Technical Publishers, Madison, WI.Fenchel,T. and Jonsson.P.R. (1988) The functional biology of Strombidium sulcatum, a marine
oligotrich ciliate (Ciliophora, Oligotrichina). Mar. Ecol. Prog. Ser., 48,1-15.Fenchel.T., Kristensen,L.D. and Rasmussen.L. (1990) Water column anoxia: vertical zonation of
planktonic protozoa. Mar. Ecol. Prog. Ser., 62,1-10.Finlay,B.J., Clarke.K.J.. Cowling.A.J., Hindle.R.M., Rogerson.A. and Berninger.U.G. (1988) On the
abundance and distribution of protozoa and their food in a productive freshwater pond. Eur. J.Protofo/., 23,205-217.
Foissner, W. (1994) Progress in taxonomy of planktonic freshwater ciliates. Mar. Microb. Food Webs, 8,9-35.
Foissner,W., Blatterer,F.H., Berger,H. and Kohmann,F. (1991) Taxonomische und okologischerevision der Cilialen des Saprobiensystems—Band I. Cyrtophorida, Oligotrichida, Hypotrichia,Colpodea. Informationsberichte des Bayer. Landesamtes fur Wasserwirtschaft, 478 pp.
Foissner,W., Berger.H. and Kohmann,F. (1992) Taxonomische und okologische revision der Cilialendes Saprobiensystems—Band II. Peritrichia, Heterotrichida, Odontostomatida. Informationsberichtedes Bayer. Landesamtes fur Wasserwirtschaft, 502 pp.
Foissner,W., Berger.H. and Kohmann.F. (1994) Taxonomische und okologische revision der Ciliatendes Saprobiensystems—Band III. Hymenostomata, Prostomatida, Nassulida. Informationsberichtedes Bayer. Landesamtes fiir Wasserwirtschaft, 548 pp.
IriberriJ., Begona.A., Unanue.M., BarcinaJ. and Egea,L. (1993) Channelling of bacterioplankton pro-duction toward phagotrophic flagellates and ciliates under different seasonal conditions in a river.Microb. Ecol., 26,111-124.
Macek.M., Simek.K., PernthalerJ., Vyhn^lek.V. and Psenner,R. (1996) Growth rates of dominantplanktonic ciliates in two freshwater bodies of different trophic degree. J. Plankton Res., 18,463-481.
Montagnes,D.J.S. and Lynn,D.H. (1987) A quantitative protargol staining (QPS) for ciliates: methoddescription and test of its quantitative nature. Mar. Microb. Food Webs, 2,83-93.
Miiller.H. (1989) The relative importance of different ciliate taxa in the pelagic food web of LakeConstance. Microb. Ecol., 18, 261-274.
Miiller.H. (1991) Pseudobalanion planctonicum (Ciliophora, Prostomatida): Ecological significance ofan algivorous nanociliate in a deep, meso-eutrophic lake. J. Plankton Res., 13,247-262.
Miiller.H. and Geller.W. (1993) Maximum growth rates of aquatic ciliated protozoa: the dependenceon body size and temperature reconsidered. Arch. Hydrobiol., 126, 315-327.
Miiller.H., SchSne.A., Pinto-Coelho,R.M., Schweizer,A. and Weisse,T. (1991) Seasonal succession ofciliates in Lake Constance. Microb. Ecol., 21,119-138.
612
by guest on July 7, 2013http://plankt.oxfordjournals.org/
Dow
nloaded from

Diet of pelagic ciliates on picoplankton
Norland.S. (1993) The relationship between biomass and volume of bacteria. In Kemp.P., Sherr.B..Sherr,E. and Cole J. (eds). Handbook of Methods in Aquatic Microbial Ecology. Lewis Publishers,Boca Raton, FL, pp. 303-308.
PernthalerJ.. Simek.K., Sattler.B., Schwarzenbacher.A.. BobkovaJ. and Psenner.R. (1996) Short-term changes of protozoan control on autotrophic picoplankton in an oligo-mesotrophic lake. J.Plankton Res., 18.443-462.
Pierce,R.W. and TurnerJ.T. (1992) Ecology of planktonic ciliates in marine food webs. Rev. Aquat.Sci., 6,139-181.
Psenner,R. (1993) Determination of size and morphology of aquatic bacteria by automated imageanalysis. In Kemp.P., Sherr.B., Sherr.E. and ColeJ. (eds). Handbook of Methods in AquaticMicrobial Ecology. Lewis Publishers, Boca Raton, FL, pp. 339-345.
Putt,M. and Stoecker,D.K. (1989) An experimentally determined carbon: volume ratio for marine'oligotrichous' ciliates from estuarine and coastal waters. Limnol. Oceanogr., 34,1097-1103.
Rassoulzadegan.F., Laval-Peuto,M. and Sheldon,R.W. (1988) Partitioning of the food ration of marineciliates between pico- and nanoplankton. Hydrobiologia, 159,75-88.
Rivier,A., Brownlee.D.C, Sheldon,R.W. and Rassoulzadegan,F. (1985) Growth of microzooplankton:A comparative study of bacterivorous zooflagellates and ciliates. Mar. Microb. Food Webs, 1,51-60.
Sanders,R.W. and Wickham,S. A. (1993) Planktonic protozoa and metazoa: predation, food quality andpopulation control. Mar. Microb. Food Webs, 7,197-223.
Sanders,R.W., Porter,K.G., Bennet,S.J. and DeBiase,A.E. (1989) Seasonal patterns of bacterivory byflagellates, ciliates, rotifers, and cladocerans in a freshwater planktonic community. Limnol.Oceanogr., 34, 673-687.
Sherr.E.B. and Sherr,B.F. (1987) High rates of consumption of bacteria by pelagic ciliates. Nature, 325,710-711.
Sherr,E.B. and Sherr.B.F. (1993) Protistan grazing rates via uptake of fluorescently labeled prey. InKemp,P., Sherr.B., Sherr,E. and Cole,J. (eds), Handbook of Methods in Aquatic Microbial Ecology.Lewis Publishers, Boca Raton, FL, pp. 695-701.
Sherr,E.B., Rassoulzadegan.F. and Sherr,B.F. (1989) Bacterivory by pelagic choreotrichous ciliates incoastal waters of the NW Mediterranean Sea. Mar. Ecol. Prog. Ser., 55, 235-240.
Sherr,E.B., Sherr.B.F., Berman.T. and Hadas.O. (1991) High abundance of picoplankton-ingestingciliates during late fall in Lake Kinneret, Israel. J. Plankton Res., 13, 789-799.
Simek,K. and Chrzanowski,T.H. (1992) Direct and indirect evidence of size-selective grazing onpelagic bacteria by freshwater nanoflagellates. Appl. Environ. Microbiol., 58, 3715-3720.
Simek,K. and StraSkrabova,V. (1992) Bacterioplankton production and protozoan bacterivory in amesotrophic reservoir. J. Plankton Res., 14, 773-787.
Simek,K., Macek,M., Sed'a,J. and Vyhnalek.V. (1990) Possible food chain relationships betweenbacterioplankton protozoans and cladocerans in a reservoir. Int. Rev. Ges. Hydrobiol., 75,583-596.
Simek.K., VrbaJ. and Hartman,P. (1994) Size-selective feeding by Cyclidium sp. on bacterioplanktonand various sizes of cultured bacteria. FEMS Microb. Ecol., 14.157-168.
Simek.K., BobkovaJ., Macek,M., Nedoma,J. and Psenner.R. (1995) Ciliate grazing on picoplankton ina eutrophic reservoir during the summer phytoplankton maximum: a study at the species and com-munity level. Limnol. Oceanogr., 40,1077-1090.
Skibbe,O. (1994) An improved quantitative protargol stain for ciliates and other planktonic protists.Arch. Hydrobiol., 130, 339-347.
Small.E.B. and Lynn.D.H. (1985) Phylum Ciliophora Doflein, 1901. In Lee,J.J., Hutner.S.H. andBovee.E.C. (eds). An Illustrated Guide to the Protozoa. Society of Protozoologists, Lawrence, USA,pp. 393-575.
Sommaruga.R. and Psenner.R. (1993) Nanociliates of the order Prostomatida: their relevance in themicrobial food web of a mesotrophic lake. Aquat. Sci., 55,179-187.
Sommaruga,R. and Psenner,R. (1995) Trophic interactions within microbial food web in Piburger See(Austria). Arch. Hydrobiol., 132,257-278.
StocknerJ.G. (1988) Phototrophic picoplankton: An overview from marine and freshwaterecosystems. Limnol. Oceanogr., 33, 765-775.
Stoecker,D.K. (1988) Are marine planktonic ciliates suspension-feeders? J. Protozool., 35,252-255.StraSkrabova.V., KomarkovaJ., Macek,M., Sed'aJ-, Simek.K., VrbaJ. and Vyhnalek,V. (1993)
Microbial-algal-crustacean interactions in a reservoir under different fishstock. Arch. Hydrobiol.Beih. Ergebn. Limnol., 40,209-221.
Turley,C.M., Newe!l,R.C. and Robbins.D.B. (1986) Survival strategies of two small marine ciliates andtheir role in regulating bacterial community structure under experimental conditions. Mar. Ecol.Prog. Ser, 33,59-70.
Weisse,T. (1993) Dynamics of autotrophic picoplankton in marine and freshwater ecosystems. InJonesJ.G. (ed.), Advances in Microbial Ecology. Plenum Press, New York, Vol. 13, pp. 327-370.
Received on March 5, 1995; accepted on November 30, 1995
613
by guest on July 7, 2013http://plankt.oxfordjournals.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















