
Misfolded Proteins Recognition Strategies of E3 Ubiquitin Ligases and Neurodegenerative Diseases
Upload
independentCategory
view
1download
0
Regulation of p53 by the Ubiquitin-conjugating EnzymesUbcH5B/C in Vivo*
Received for publication, March 25, 2004, and in revised form, July 16, 2004Published, JBC Papers in Press, July 26, 2004, DOI 10.1074/jbc.M403362200
Mark K. Saville‡§, Alison Sparks‡, Dimitris P. Xirodimas‡, Julie Wardrop‡,Lauren F. Stevenson‡, Jean-Christophe Bourdon, Yvonne L. Woods‡, and David P. Lane¶
From the Cancer Research UK, Cell Transformation Research Group, Department of Surgery and Molecular Oncology,Ninewells Hospital and Medical School, University of Dundee, Dundee DD1 9SY, Scotland
p53 levels are regulated by ubiquitination and 26 Sproteasome-mediated degradation. p53 is a substrate forthe E3 ligase Mdm2, however, the ubiquitin-conjugatingenzymes (E2s) involved in p53 ubiquitination in intactcells have not been defined previously. To investigatethe E2 specificity of Mdm2 we carried out an in vitroscreen using a panel of ubiquitin E2s. Of the E2s testedonly UbcH5A, -B, and -C and E2-25K support Mdm2-mediated ubiquitination of p53. The same E2s also sup-port Mdm2 auto-ubiquitination. Small interfering RNA-mediated knockdown of UbcH5B/C causes accumulationof Mdm2 and p53 in unstressed cells. We show that sup-pression of UbcH5B/C inhibits p53 ubiquitination anddegradation. Despite up-regulating the level of nuclearp53, UbcH5B/C knockdown does not on its own result inan increase in p53 transcriptional activity or sensitizep53 to activation by the therapeutic drugs doxorubicinand actinomycin D. We provide evidence that Mdm2 isresponsible, at least in part, for repression of the tran-scriptional activity of the accumulated p53. In MCF7cells levels of UbcH5B/C are reduced by doxorubicin andactinomycin D. This observation and the sensitivity ofp53 expression to levels of UbcH5B/C raise the possibil-ity that E2 regulation could be involved in signalingpathways that control the stability of p53. Our data in-dicate that UbcH5B/C are physiological E2s for Mdm2,which make a significant contribution to the mainte-nance of low levels of p53 and Mdm2 in unstressed cellsand that inhibition of p53 ubiquitination and degrada-tion by targeting UbcH5B/C is not sufficient to up-regu-late p53 transcriptional activity.
In normal unstressed cells the levels and activity of the p53tumor suppressor are kept low. p53 is stabilized, and its tran-scriptional activity is up-regulated following diverse stresses,including ionizing and UV radiation, genotoxic drugs, and theinappropriate activation of oncogenes. This regulates the ex-pression of multiple target genes and leads to cell cycle arrestor apoptosis, thus preventing damaged cells from proliferating(1, 2). Most tumor cells have escaped this p53 surveillance. p53is frequently inactivated by mutation. However, there are alsomany tumors expressing wild-type p53, which is inactivated byother mechanisms, including overexpression of Mdm2 and loss
of p14ARF. Activation of the endogenous p53 pathway is anattractive approach to treatment of tumors expressing wild-type p53 (3, 4). The precise mechanisms by which the activity ofp53 is regulated are thus the focus of much interest. Themaintenance of low levels of p53 in cells involves its ubiquiti-nation and consequent targeting for degradation by the 26 Sproteasome (5). Because ubiquitination is involved in many keyprocesses, there is also considerable general interest in compo-nents of the ubiquitination pathway as therapeutic targets fortreatment of diverse diseases. Clinical trials have been carriedout with the proteasome inhibitor PS-341. This displays anti-tumor activity and has been approved for therapy of patientswith multiple myeloma (6–8).
Ubiquitination of proteins occurs through the sequential ac-tions of three enzymes (9, 10). Initially ubiquitin is activated bythe ubiquitin-activating enzyme (E1).1 Ubiquitin is then trans-ferred from the E1 to a ubiquitin-conjugating enzyme (E2). Aubiquitin ligase (E3) then facilitates transfer of ubiquitin froman E2 to the substrate. There is one human ubiquitin E1 andmultiple ubiquitin E2s (10, 11). Mdm2 acts as an E3 ligase forp53 (12) and can promote its ubiquitination and degradation invivo (13, 14). Mdm2 also “auto-ubiquitinates” and is itself tar-geted for degradation by the proteasome (15). In addition toregulating p53 protein expression Mdm2 can inhibit p53 tran-scriptional activity by binding to its transactivation domain(16, 17), and it may directly repress basal transcription fromp53-responsive promoters (18). We have recently observed thatMdm2 can promote conjugation of the ubiquitin-like proteinNEDD8 (neural precursor cell-expressed developmentallydown-regulated) to p53 (19). This modification also repressesthe transcriptional activity of p53. The relative contribution toinhibition of p53 of Mdm2-dependent ubiquitin and NEDD8conjugation and transcriptional repression through direct bind-ing to p53 are not clear. Loss of Mdm2 in mice is embryoniclethal in a p53-dependent manner (20, 21). In transgenic adultmice, which express lower levels of Mdm2 than wild-type mice,the transactivation and apoptotic activities of p53 are en-hanced (22). Agents that bind to Mdm2 and block its associa-tion with p53 increase both the levels and transcriptional ac-tivity of p53 in vivo. These include high affinity Mdm2 bindingpeptides, the Mdm2-specific antibody 3G5 (23), and the Nutlinfamily of small molecules (24). These and other observationsindicate that Mdm2 is a key negative regulator of p53 (25, 26).
* The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby marked“advertisement” in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
‡ Recipients of postdoctoral fellowships from Cancer Research UK.§ To whom correspondence should be addressed: Tel.: 44-(0)1382-496-
402; Fax: 44-(0)1382-496-361; E-mail: [email protected].¶ A Gibb fellow of Cancer Research UK.
1 The abbreviations used are: E1, ubiquitin-activating enzyme; E2,ubiquitin-conjugating enzyme; E3, ubiquitin-protein ligase; PKA, pro-tein kinase A; RNAi, RNA interference; RT, reverse transcriptase;siRNA, small interfering RNA; STIP, super thioredoxin insert protein;TRX, thioredoxin; PBS, phosphate-buffered saline; GST, glutathioneS-transferase; Ni-NTA, nickel-nitrilotriacetic acid; DMEM, Dulbecco’smodified Eagle’s medium; 6-FAM, 6-carboxyfluorescein; TAMRA,carboxytetramethylrhodamine.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 279, No. 40, Issue of October 1, pp. 42169–42181, 2004© 2004 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org 42169
by guest on July 8, 2016http://w
ww
.jbc.org/D
ownloaded from

Mdm2 is itself a target for p53 transcriptional regulation.Mdm2 is induced by p53 resulting in a negative feedback loop(27). The Mdm2 structural homologue MdmX is also an impor-tant regulator of p53. MdmX binds to p53 and inhibits itstranscriptional activity but does not itself ubiquitinate p53 incells (28), although a low level of E3 ligase activity has beenobserved in vitro (29). Mdm2 and MdmX heterodimerizethrough their ring fingers (30). Strikingly, ablation of MdmXalso results in p53-dependent embryonic lethality in mice(31, 32).
Although auto-ubiquitination of Mdm2 and Mdm2-mediatedubiquitination of p53 have been reconstituted in vitro usingpurified E2s (12, 33, 34), the E2 specificity of Mdm2 has notbeen investigated in detail, and the one or more ubiquitin E2sinvolved in regulating the Mdm2/p53 pathway in intact cellshave not been determined previously. Ubiquitin E2s consist ofa conserved core domain of �150 amino acids. They have anactive site cysteine residue, which forms a thiolester bond withubiquitin transferred from the E1. Ubiquitination, and by in-ference ubiquitin E2s, are involved in regulating many cellularprocesses. However, the precise physiological roles of many ofthe ubiquitin E2s are not well defined, particularly in mamma-lian cells. There is evidence that E2s have unique functions,presumably arising from their specificity for particular E3s.Genetic studies in Saccharomyces cerevisiae indicate that Ubc2(RAD6) is central to DNA repair, Ubc3 (CDC34) is required forthe G1-S transition, and Ubc4/5 are required for viability (10).
Regulation of E2 levels or activity could provide a mecha-nism to alter the stability of key proteins. Studies using rattissue extracts indicate that E2s can be rate-limiting for ubiq-uitination (35). The expression of S. cerevisiae Ubc4 and -5 isgrowth- and heat shock-regulated (36). The levels of specificmammalian E2s have also been reported to be regulated by thecell cycle and by agents, including interferons � and �, insulin,insulin-like growth factor 1, amyloid-�, and herpes simplexvirus (37–45). E2 activity can be influenced by phosphoryla-tion. CDC34 and its homologue Ubc3b are substrates for pro-tein kinase CK2 (46), and UbcH1 (HHR6) is phosphorylated byCDK1 and -2 (47).
In this study we identified ubiquitin E2s involved in theMdm2/p53 pathway in intact cells. A panel of E2s were testedfor their ability to support the ubiquitin ligase activity of Mdm2in vitro. We show that there is specificity in E2 usage by Mdm2.Of the E2s tested only members of the UbcH5 family (A–C) andE2-25K are able to support the ubiquitin ligase activity ofMdm2. We observed that the pattern of Mdm2-mediated ubiq-uitination of p53 in vitro is dependent on the E2 used. UsingsiRNA-mediated knockdown we show that UbcH5B/C make asignificant contribution to the maintenance of low levels ofMdm2 and p53 in unstressed cells. In contrast, suppression ofUbcH5A or E2-25K has little effect on expression of Mdm2 andp53 in the cell lines examined. Levels of UbcH5B/C proteinexpression were found to be higher than those of UbcH5A,possibly accounting for the preferential usage of UbcH5B/C inintact cells. The balance of activities of the Mdm2/p53 pathwayis such that the p53 accumulated following UbcH5B/C knock-down is not transcriptionally active, the level of Mdm2 beingsufficient to maintain transcriptional repression of p53. Wefind that expression of UbcH5B/C is reduced by the p53 stabi-lizing therapeutic drugs doxorubicin and actinomycin D. Be-cause p53 expression is sensitive to the levels of UbcH5B/C, itis conceivable that E2 regulation could play some part in sig-naling to the p53 pathway.
EXPERIMENTAL PROCEDURES
Antibodies—The antibodies used were 4B2 for Mdm2; DO-1 for p53unless otherwise indicated; H-81 for CDC34 (Santa Cruz Biotechnology);
UBC4 (C-15) for UbcH5B/C (Santa Cruz Biotechnology), which was pre-cleared by absorption with GST-UbcH5A; Ab-1 for p21 (Oncogene); L27for CD20 (BD Biosciences); and Ab-1 for actin (Calbiochem). A rabbitpolyclonal anti-E2-25K antibody was purchased from Affiniti Research, arabbit anti-ubiquitin antiserum was purchased from Sigma and a sheepanti-UbcH5A polyclonal antiserum raised against purified full-lengthUbcH5A was provided by Dr. R. T. Hay. This was affinity-purified andthen precleared by absorption with GST-UbcH5B and -C. Where indi-cated, cross-reactivity was removed by incubating antibodies (15–30 �g)three times for 4 h with GST-E2s (�50 �g) bound to glutathione-Sepha-rose beads in 0.5 ml of 5% dried milk, 0.1% Tween 20 in PBS.
Plasmids and Synthetic siRNA Duplexes—To generate constructs forbacterial expression of GST-E2s, the following IMAGE clones wereobtained from the UK MRC Human Genome Mapping Project resourcecenter: HHR6A (3854960), HHR6B (2178494), UbcH2 (1908854),UbcH5B (2169015), UbcH5C (3854578), and UbcH8 (3838534). UbcH5AcDNA was donated by Dr. R. T. Hay. These were used as templates forPCR, and the products were cloned into the BamHI and EcoRI sites ofpGEX-2T with the exception of HHR6A, which was cloned into the SmaIand EcoRI sites. pT7-7-Mdm2 used for bacterial expression of humanMdm2 was described previously (34). For expression of His6-taggedhuman p53 in bacteria, a PCR product was cloned into the SphI site ofpT7-7-SphI, which was generated as described previously (48). pGEX-2TK-ubiquitin used for bacterial expression of GST-ubiquitin was do-nated by Dr. K. Madura. For expression in cell lines, a transcriptionallyinactive mutant of human p53 (His-273) was cloned into the EcoRI andXhoI sites of pcDNA3. Plasmids used to express human Mdm2 andHis6-ubiquitin have been described previously (15). pCMVCD20 wasdonated by Dr. R. Watson.
siRNA target sequences were chosen according to the criteria of El-bashir et al. (49). The following target sequences were inserted into thepSUPER vector (50), which was obtained from Dr. R. Bernards: CDC34,CACCTACTACGAGGGCGGC; UbcH5A, GGTGGAGTCTTCTTTCTCA;UbcH5B, CAGATTACCCCTTCAAACC; UbcH5C, CAGACAGAGATAAG-TACAA; UbcH5B/C, CAGTAATGGCAGCATTTGT; UbcH5B/C (2), GAT-CACAGTGGTCGCCTGC; UbcH5B/C (3), TGGACTCAGAAGTATGCCA;and E2-25K, TAGTGGCCTTGTCTTCAAA. pSUPER-UbcH5B/C targetsidentical sequences in UbcH5B and -C. pSUPER-UbcH5B/C (2) andUbcH5B/C (3) target sequences in UbcH5C, which differ from those inUbcH5B, by two and one nucleotide, respectively, and were found tosuppress both UbcH5B and -C mRNA. Nonspecific control duplex IX,Mdm2 SMART pool (M-003279-00), and synthetic siRNA duplexes, whichtarget the same sequences as pSUPER-UbcH5A, UbcH5B/C, and E2-25K,were purchased from Dharmacon.
Gel Electrophoresis and Western Blotting—Cells were washed twicewith PBS at 4 °C. Cell extracts were prepared by direct lysis intoSDS-urea electrophoresis sample buffer: 100 mM Tris, pH 6.8, 4% SDS,8 M urea, 20% glycerol, 20 mM EDTA, 0.014% bromphenol blue. DNAwas sheared by passage through a 25-gauge needle, and protein con-centrations were measured using the BCA protein assay (Pierce). Di-thiothreitol was added to a final concentration of 100 mM, samples wereheated for 5 min at 95 °C, and proteins were resolved by SDS-PAGE.Gels were transferred onto nitrocellulose for 16 h, at 25 mA, or for 1 hat 150 mA, and membranes were processed as described in Midgley etal. (34). Membranes probed for ubiquitin were boiled in de-ionizedwater prior to blocking to expose epitopes in ubiquitin. Peroxidase-conjugated secondary antibodies were supplied by Jackson Immuno-Research Laboratories and used at a dilution of 1/10,000. Bound anti-bodies were detected by enhanced chemiluminescence (AmershamBiosciences) or using SuperSignal West Dura Extended Duration Sub-strate (Pierce).
Expression and Purification of Recombinant Proteins—Untagged hu-man Mdm2 was prepared from bacterial inclusion bodies as describedpreviously (34). Human E1 was purified from recombinant baculovirus-infected insect cells by affinity chromatography on ubiquitin-Sepharoseas described in Desterro et al. (51). Human GST-HHR6A, HHR6B,UbcH2 (52), UbcH5A, UbcH5B, UbcH5C, and UbcH8 were expressed inbacteria and purified as described previously (53). GST-ubiquitin con-taining a protein kinase A (PKA) consensus site between the ubiquitinand the tag was expressed in bacterial, purified, and phosphorylated byPKA as described in Tongaonkar et al. (54). GST was cleaved from E2sand ubiquitin by thrombin digestion prior to use in ubiquitinationassays, and free thrombin was removed by incubation with benzami-dine-Sepharose. UbcH6, -7, and -10 and E2-25K proteins were suppliedby Affiniti Research, and CDC34 was donated by Dr. R. T. Hay. His6-p53 was expressed in bacteria. Following induction with isopropyl-1-thio-�-D-galactopyranoside at room temperature p53 in the soluble frac-tion was partially purified on a heparin-Sepharose column as described
p53 Regulation by UbcH5B/C42170
by guest on July 8, 2016http://w
ww
.jbc.org/D
ownloaded from

previously (55). Fractions containing full-length p53 were further pu-rified on Ni2�-NTA-agarose.
In Vitro Ubiquitination Assays—Reactions contained 50 mM Tris, pH7.6, 5 mM MgCl2, 2 mM ATP, 10 mM creatine phosphate, 3.5 units/mlcreatine kinase, and 0.6 unit/ml inorganic pyrophosphatase, 20 �M
[32P]ubiquitin, and human E1 (50 nM), E2s (1.5 �M), and where indi-cated p53 (1 �M). Reactions were initiated by the addition of humanMdm2 (50–100 nM). Following a 2-h incubation at 37 °C, reactions wereterminated by the addition of SDS-urea sample buffer. Products wereresolved by SDS-PAGE. [32P]Ubiquitin was detected by phosphorimag-ing of dried gels, and p53 was detected by Western blotting using DO-1.
Cell Culture and Transfection—MCF7 breast tumor cells and U2-OSosteosarcoma cells were cultured in DMEM and H1299 lung carcinomacells in RPMI. Media were supplemented with 10% fetal calf serum andgentamycin (50 �g/ml), and cells were kept at 37 °C, 5% CO2 in ahumidified atmosphere. Unless otherwise indicated MCF7 and U2-OScells were transfected with 1.5 �g of pSUPER construct per 10-cm dish.Transfection of these cells lines with plasmids was performed withFuGENE 6 transfection reagent (Roche Applied Science) following themanufacturer’s instructions. Cells were washed after 16 h, and freshmedium was added. Unless otherwise indicated H1299 cells were trans-fected with 7 �g of pSUPER constructs per 10-cm dish. Transfection ofthis cell line was performed using the calcium phosphate method es-sentially as described in Xirodimas et al. (15). Transfection with singlesiRNA synthetic duplexes (100 nM) or SMART pools (200 nM) wascarried out using Oligofectamine (Invitrogen) according to the manu-facturer’s instructions.
CD20 Enrichment of Transfected Cells—MCF7 cells in 10-cm disheswere transfected with pSUPER constructs and pCMVCD20 (1 �g). Atthe indicated time after transfection, cells were detached by washingwith 3 mM EDTA in PBS. Cells were resuspended thoroughly in DMEM,10% FCS, and incubated for 20 min at 4 °C with magnetic DynabeadsPan Mouse IgG (Dynal Biotech) coated with anti-CD20 antibody (1 �107 beads and 1 �g of antibody per dish). The beads were isolated witha magnet, and the selected cells were washed twice with PBS beforelysis for RNA extraction or Western blotting.
Immunofluorescence—Cells were seeded onto NUNC Permanoxslides. 36 h after transfection with synthetic siRNA duplexes, cells werefixed with ice-cold methanol-acetone and incubated with primary anti-bodies followed by fluorescence isothiocyanate-conjugated donkey anti-mouse secondary antibodies (Jackson ImmunoResearch) as describedpreviously (15).
Purification of His6-tagged Ubiquitin Conjugates—MCF7 cells wereseeded onto 10-cm dishes and transfected with pSUPER constructs andHis6-ubiquitin (1 �g) using FuGENE 6. 64 h later, cells were harvestedby direct lysis into 8 M urea, 100 mM Na2HPO4/NaH2PO4, 10 mM Tris,pH 8.0, 10 mM �-mercaptoethanol, 5 mM imidazole. Purification ofHis6-ubiquitin conjugates using Ni2�-NTA-agarose beads was carriedout as described in Xirodimas et al. (15).
Quantitative Detection of �-Galactosidase—Cells in 24-well plateswere washed twice with PBS and lysed on the plate with 150 �l ofpassive lysis buffer (Promega). 150 �l of substrate solution containing80 �g/ml chlorophenol red �-D-galactopyranoside (Roche Applied Sci-ence) 0.5 mM MgCl2, 23 mM �-mercaptoethanol in 0.1 M sodium phos-phate buffer, pH 7.5, was added to 15 �l of extract in a 96-well plate.The assay was quantitated by absorbance measurements at a wave-length of 590 nm in a plate reader. Luciferase activities were measuredusing the Berthold microplate luminometer LB 96V and used to controlfor transfection efficiency.
RNA Preparation and Real-time RT-PCR—Total RNA was extractedusing RNeasy columns (Qiagen) according to the manufacturer’s instruc-tions, including an on-column DNase treatment step. RNA (300–600 ng)was incubated with random hexamers and Superscript II reverse tran-scriptase (Invitrogen) to generate cDNA. Real-time PCR was carried outwith an ABI Prism 7700 sequence detector using the following protocol:50 °C for 2 min, 95 °C for 10 min, and 40 cycles of 95 °C for 15 s and 60 °Cfor 1 min. For ubiquitin E2s, primers and 6-FAM/TAMRA-labeled probeswere as used in Okamoto et al. (56). Other specific probes and primerswere as follows: p53 primers: 5�-CAGCCAAGTCTGTGACTTGCA-3�, 5�-GTGTGGAATCAACCCACAGCT-3�, probe: 6-FAM-TCCCCTGCCCTCA-ACAAGATGTTTTGCC-TAMRA; Mdm2 primers: 5�-CAAGTTACTGTGT-ATCAGGCAGGG-3�, 5�-TCTGTTGCAATGTGATGGAAGG-3�, probe: 6-FAM-TTGGAAATGCACTTCAT-TAMRA; p21 primers: 5�-GACTCTCA-GGGTCGAAAACGG-3�, 5�-GCGGATTAGGGCTTCCTCTT-3�, probe: 6-FAM-CTACCACTCCAAACGCCGGCTGATCT-TAMRA; BAX primers:5�-ACTCCCCCCGAGAGGTCTT-3�, 5�-GCAAAGTAGAAAAGGGCGAC-AA-3�, probe: 6-FAM-AGCTGACATGTTTTCTGACGGCAACTTCAACT-TAMRA; PIG3 primers: 5�-CCGGGCGGACTTAATGC-3�, 5�-CTCAAGT-
CCCAAAATGTTGCTG-3�, probe: 6-FAM-AGACAAGGCCAGTATGACC-CACCTCCAGGAG-TAMRA. Probes and primers for actin werepurchased from Applied Biosystems.
RESULTS
The Ubiquitin Ligase Activity of Mdm2 Is Specifically Sup-ported by UbcH5A, -B, and -C and E2-25K in Vitro—Mdm2-mediated ubiquitination of p53 has previously been reconsti-tuted in vitro with the E2s UbcH5A and -B (12, 34). However,the ubiquitin E2 specificity of Mdm2 has not been investigatedin detail. To address this we screened a range of characterizedE2s for the ability to support the ubiquitin ligase activity ofMdm2. Purified E2s were matched for protein concentrationand tested for functionality by their ability to form thiolesterswith 32P-labeled ubiquitin (data not shown). We first looked atMdm2 auto-ubiquitination in the absence of p53. Followingincubations with purified E1, [32P]ubiquitin and the indicatedE2s, proteins were resolved by SDS-PAGE under reducing
FIG. 1. In vitro specificity of Mdm2 for ubiquitin E2s. A, Mdm2auto-ubiquitination. In vitro ubiquitination assays were carried outwith E1, 32P-labeled ubiquitin, and the indicated E2s in the presence orabsence of Mdm2. Gels were run under reducing conditions, and [32P]u-biquitin was detected by phosphorimaging. B, Mdm2-mediated ubiq-uitination of p53. In vitro ubiquitination assays were carried out withp53 and the indicated E2s (where s is for sugar) in the presence orabsence of Mdm2. p53 was detected by Western blotting with themonoclonal antibody DO-1.
p53 Regulation by UbcH5B/C 42171
by guest on July 8, 2016http://w
ww
.jbc.org/D
ownloaded from

conditions (Fig. 1A). Ubiquitinated Mdm2 migrated as a veryhigh molecular weight species near the top of the gel. Theidentity of this species was confirmed by immunodepletionusing a mixture of anti-Mdm2 antibodies (data not shown). Ofthe E2s tested, only UbcH5A, -B, and -C and E2-25K were ableto support auto-ubiquitination of Mdm2. E2-25K has not pre-viously been shown to support the ubiquitin ligase activity ofMdm2. Typically, the incorporation of ubiquitin into Mdm2 was7- to 8-fold higher with UbcH5 family members than withE2-25K. Mdm2-mediated ubiquitination of p53 was also ob-served exclusively with members of the UbcH5 family andE2-25K (Fig. 1B). In these assays p53 was detected by Westernblotting using the anti-p53 antibody DO-1. In contrast to arecent report (57) we did not observe Mdm2-mediated ubiquiti-nation of p53 with HHR6A or -B. The pattern of p53 ubiquiti-nation was similar with UbcH5A, -B, and -C and consistedpredominantly of relatively low molecular weight ubiquiti-nated species. E2-25K supported predominantly high molecu-lar weight ubiquitination of p53. These observations show thatthere is specificity for E2 usage by Mdm2, that the E2s whichsupport Mdm2 auto-ubiquitination can also support Mdm2-mediated ubiquitination of p53, and that Mdm2 can displaydifferent biochemical properties dependent on the E2 thatis used.
Members of the UbcH5 family are highly homologous.UbcH5B and -C are 97% identical at the amino acid level, andwith a difference of only four amino acids, it is likely thatUbcH5B and -C have similar activities. UbcH5A is more diver-gent, being 89 and 88% identical to UbcH5B and -C, respec-tively, at the amino acid level (58, 59). UbcH5A and UbcH5B/Chave been observed to have different biochemical properties invitro (60).
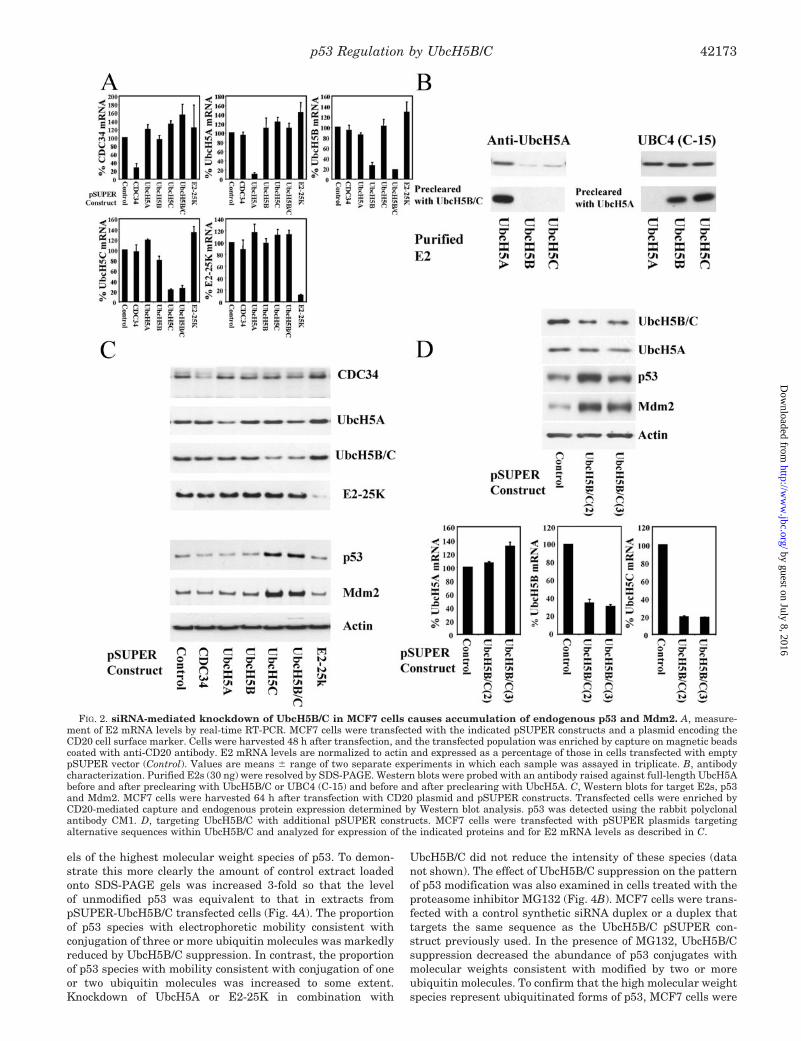
siRNA-mediated Knockdown of UbcH5B/C Causes Accumu-lation of p53 and Mdm2 Protein—To investigate the involve-ment of UbcH5 family members and E2-25K in regulation ofp53 and Mdm2 expression in intact cells, we used RNA inter-ference (RNAi) to knockdown specific E2s. Target sequenceswere inserted into the pSUPER vector, which directs the syn-thesis of small interfering RNA (siRNA) (50). We targeted theE2s able to support the ubiquitin ligase activity of Mdm2 invitro and as a negative control CDC34 with which no ubiquitinligase activity of Mdm2 was observed. Due to their high degreeof homology and the consequent likelihood of functional redun-dancy between UbcH5B and -C, we generated single pSUPERconstructs that would suppress both UbcH5B and -C. We em-ployed MCF7 human breast adenocarcinoma cells, which ex-press wild-type p53 and Mdm2. In these cells p53 levels andactivity are sensitive to agents that disrupt the p53-Mdm2interaction, therapeutic drugs, and p14ARF. These cells wereco-transfected with pSUPER constructs and a plasmid encod-ing the CD20 cell surface marker. Cells were harvested after48 h, and the transfected population of cells was captured usinganti-CD20 antibody bound to magnetic beads. For each of theE2s targeted several sequences were tested. pSUPER plasmidswere selected that gave 75% or greater knockdown of theirtarget mRNA (Fig. 2A).
To facilitate detection of siRNA-mediated knockdown of E2proteins, anti-UbcH5 antibodies were characterized using pu-rified recombinant E2s (Fig. 2B). A UbcH5A-selective antibodywas generated by preclearing a polyclonal antiserum raisedagainst purified UbcH5A by adsorption of cross-reacting anti-bodies with GST-UbcH5B and -C. A UbcH5B/C-selective anti-body was generated by preclearing UBC4 (C-15) anti-peptidepolyclonal by absorption of cross-reacting antibodies with GST-UbcH5A. To date we have been unable to generate an antibodythat can distinguish between the highly homologous E2s
UbcH5B and -C. Transfection of MCF7 cells with the pSUPERconstructs specifically reduced expression of their target pro-teins (Fig. 2C). CDC34, UbcH5A, UbcH5B/C, and E2-25K pro-tein expression was reduced by a similar extent. Knockdown ofUbcH5C reduced the total level of UbcH5B/C immunoreactiv-ity, whereas this was unaffected by UbcH5B knockdown. Thissuggests that in MCF7 cells the level of UbcH5C protein ex-pression is higher than that of UbcH5B. Knockdown of UbcH5Cor UbcH5B/C resulted in the accumulation of endogenous p53and Mdm2 protein (Fig. 2C). To confirm that the accumulationof p53 and Mdm2 was not due to an off-target effect of thesiRNA, two additional pSUPER constructs that targetUbcH5B/C were tested. These pSUPER constructs reduced themRNA and protein levels of UbcH5B/C and caused accumula-tion of p53 and Mdm2 protein (Fig. 2D). To investigate thegenerality of these observations, the effects of ubiquitin E2knockdown were also determined in U2-OS human osteosar-coma cells, which express wild-type p53 and Mdm2, and inH1299 human lung carcinoma cells, which express Mdm2 andare p53 null. In these cell lines transfection with pSUPERconstructs specifically reduced expression of the target proteins(Fig. 3A). Knockdown of UbcH5B/C caused accumulation ofendogenous p53 and Mdm2 protein in U2-OS cells and of en-dogenous Mdm2 in H1299 cells. To confirm that UbcH5B/C arerequired for Mdm2-dependent degradation of p53, the effect ofUbcH5B/C knockdown on the ability of ectopically expressedMdm2 to promote degradation of p53 was examined. U2-OScells were transfected with a transcriptionally inactive p53mutant (His-273) to avoid the deleterious effects associatedwith prolonged expression of wild-type p53. Overexpression ofMdm2 promoted degradation of exogenous p53 (Fig. 3B).UbcH5B/C knockdown caused accumulation of ectopically ex-pressed Mdm2 and attenuated Mdm2-dependent p53 degrada-tion. These data indicate that UbcH5B/C are required for themaintenance of low levels of p53 and Mdm2 in a range of celllines. In MCF7 cells this principally involves UbcH5C.UbcH5B/C knockdown caused accumulation of Mdm2 in a p53null cell line showing that UbcH5B/C can regulate Mdm2 in-dependently of p53.
In the cell lines examined, no effect on p53 and Mdm2 ex-pression was observed following knockdown of the other E2stargeted, including UbcH5A and E2-25K. Knockdown ofUbcH5A or E2-25K in combination with UbcH5B/C, by co-transfection with pSUPER plasmids, did not increase p53 andMdm2 levels more than suppression of UbcH5B/C alone (datanot shown). In vitro UbcH5A supported the ubiquitination ofp53 equally as well as UbcH5B/C. To investigate whether dif-ferences in E2 usage in vivo could be related to differences inprotein expression, protein levels of UbcH5 family memberswere determined by Western blotting using recombinant pro-teins as standards. The level of protein expression ofUbcH5B/C was 3- to 5-fold higher than that of UbcH5A in thecell lines examined (Fig. 3C).
UbcH5B/C Knockdown Attenuates p53 Ubiquitination with-out Causing Gross Changes in the General Pattern of UbiquitinConjugates—To investigate the effect of E2 suppression on theubiquitination of endogenous p53, MCF7 cells were transfectedwith pSUPER plasmids and harvested by direct lysis into SDS-urea electrophoresis sample buffer. This denaturing buffer in-hibits de-ubiquitinating enzymes and thus preserves ubiquitin-protein conjugates (61). A ladder of high molecular weight p53species was detected in control samples. This ladder resultsfrom conjugation of multiple ubiquitin or ubiquitin-like pro-teins to p53. Using pSUPER constructs that target two differ-ent sequences in UbcH5B/C, to control for off-target effects, weobserved that UbcH5B/C knockdown reduced the relative lev-
p53 Regulation by UbcH5B/C42172
by guest on July 8, 2016http://w
ww
.jbc.org/D
ownloaded from
els of the highest molecular weight species of p53. To demon-strate this more clearly the amount of control extract loadedonto SDS-PAGE gels was increased 3-fold so that the levelof unmodified p53 was equivalent to that in extracts frompSUPER-UbcH5B/C transfected cells (Fig. 4A). The proportionof p53 species with electrophoretic mobility consistent withconjugation of three or more ubiquitin molecules was markedlyreduced by UbcH5B/C suppression. In contrast, the proportionof p53 species with mobility consistent with conjugation of oneor two ubiquitin molecules was increased to some extent.Knockdown of UbcH5A or E2-25K in combination with
UbcH5B/C did not reduce the intensity of these species (datanot shown). The effect of UbcH5B/C suppression on the patternof p53 modification was also examined in cells treated with theproteasome inhibitor MG132 (Fig. 4B). MCF7 cells were trans-fected with a control synthetic siRNA duplex or a duplex thattargets the same sequence as the UbcH5B/C pSUPER con-struct previously used. In the presence of MG132, UbcH5B/Csuppression decreased the abundance of p53 conjugates withmolecular weights consistent with modified by two or moreubiquitin molecules. To confirm that the high molecular weightspecies represent ubiquitinated forms of p53, MCF7 cells were
FIG. 2. siRNA-mediated knockdown of UbcH5B/C in MCF7 cells causes accumulation of endogenous p53 and Mdm2. A, measure-ment of E2 mRNA levels by real-time RT-PCR. MCF7 cells were transfected with the indicated pSUPER constructs and a plasmid encoding theCD20 cell surface marker. Cells were harvested 48 h after transfection, and the transfected population was enriched by capture on magnetic beadscoated with anti-CD20 antibody. E2 mRNA levels are normalized to actin and expressed as a percentage of those in cells transfected with emptypSUPER vector (Control). Values are means � range of two separate experiments in which each sample was assayed in triplicate. B, antibodycharacterization. Purified E2s (30 ng) were resolved by SDS-PAGE. Western blots were probed with an antibody raised against full-length UbcH5Abefore and after preclearing with UbcH5B/C or UBC4 (C-15) and before and after preclearing with UbcH5A. C, Western blots for target E2s, p53and Mdm2. MCF7 cells were harvested 64 h after transfection with CD20 plasmid and pSUPER constructs. Transfected cells were enriched byCD20-mediated capture and endogenous protein expression determined by Western blot analysis. p53 was detected using the rabbit polyclonalantibody CM1. D, targeting UbcH5B/C with additional pSUPER constructs. MCF7 cells were transfected with pSUPER plasmids targetingalternative sequences within UbcH5B/C and analyzed for expression of the indicated proteins and for E2 mRNA levels as described in C.
p53 Regulation by UbcH5B/C 42173
by guest on July 8, 2016http://w
ww
.jbc.org/D
ownloaded from
co-transfected with His6-tagged ubiquitin and pSUPER plas-mids. His6-tagged conjugates were isolated on Ni2�-agaroseand analyzed by Western blotting for p53. The electrophoretic
mobility of high molecular weight p53 species in whole celllysates from cells treated with MG132 was similar to the mo-bility of ubiquitinated p53 species isolated on Ni2�-agarose(Fig. 4C). The presence of these p53 species in the Ni2�-agar-ose-purified material was dependent on transfection with His6-ubiquitin indicating that they contain ubiquitin. Consistentwith observations made with whole cell lysates, UbcH5B/Csuppression reduced the relative amount of conjugates contain-ing three or more ubiquitin molecules but increased the inten-sity of conjugates containing one or two ubiquitin molecules.The mono- and di-ubiquitinated species of p53 present afterUbcH5B/C suppression could represent ubiquitination of p53that is supported by residual UbcH5B/C, because the knock-down is incomplete. E3 up-regulation following UbcH5B/Cknockdown may also be a contributory factor.
Human UbcH5A, -B, and -C are closely related to S. cerevi-siae Ubc4 and Ubc5. Simultaneous deletion of these E2s inS. cerevisiae results in a severe reduction in high molecularweight ubiquitin conjugates (36). We examined the effect of E2knockdown on the general pattern of ubiquitination in MCF7cells. The transfection efficiency was sufficient to detect a re-duction in UbcH5B/C expression in SDS-urea whole cell lysates(Fig. 4D, top panel). There were no dramatic changes in thepattern or level of conjugates detected by Western blot analysisusing an anti-ubiquitin antibody (Fig. 4D, middle panel).
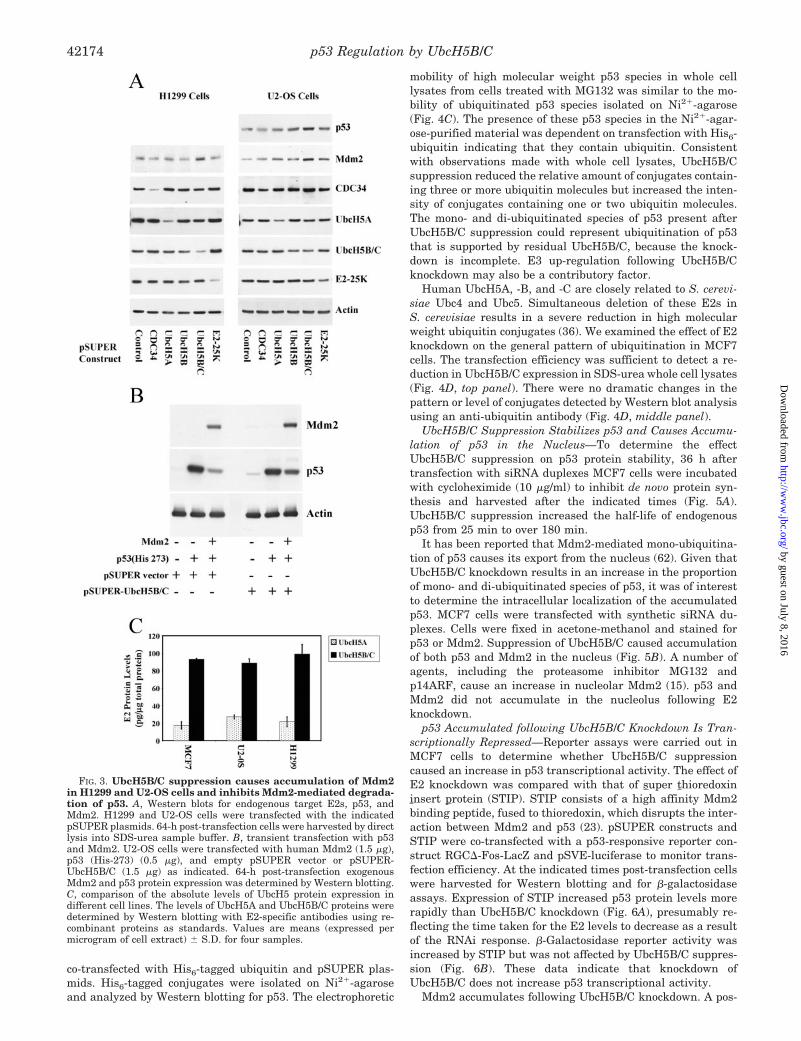
UbcH5B/C Suppression Stabilizes p53 and Causes Accumu-lation of p53 in the Nucleus—To determine the effectUbcH5B/C suppression on p53 protein stability, 36 h aftertransfection with siRNA duplexes MCF7 cells were incubatedwith cycloheximide (10 �g/ml) to inhibit de novo protein syn-thesis and harvested after the indicated times (Fig. 5A).UbcH5B/C suppression increased the half-life of endogenousp53 from 25 min to over 180 min.
It has been reported that Mdm2-mediated mono-ubiquitina-tion of p53 causes its export from the nucleus (62). Given thatUbcH5B/C knockdown results in an increase in the proportionof mono- and di-ubiquitinated species of p53, it was of interestto determine the intracellular localization of the accumulatedp53. MCF7 cells were transfected with synthetic siRNA du-plexes. Cells were fixed in acetone-methanol and stained forp53 or Mdm2. Suppression of UbcH5B/C caused accumulationof both p53 and Mdm2 in the nucleus (Fig. 5B). A number ofagents, including the proteasome inhibitor MG132 andp14ARF, cause an increase in nucleolar Mdm2 (15). p53 andMdm2 did not accumulate in the nucleolus following E2knockdown.
p53 Accumulated following UbcH5B/C Knockdown Is Tran-scriptionally Repressed—Reporter assays were carried out inMCF7 cells to determine whether UbcH5B/C suppressioncaused an increase in p53 transcriptional activity. The effect ofE2 knockdown was compared with that of super thioredoxininsert protein (STIP). STIP consists of a high affinity Mdm2binding peptide, fused to thioredoxin, which disrupts the inter-action between Mdm2 and p53 (23). pSUPER constructs andSTIP were co-transfected with a p53-responsive reporter con-struct RGC�-Fos-LacZ and pSVE-luciferase to monitor trans-fection efficiency. At the indicated times post-transfection cellswere harvested for Western blotting and for �-galactosidaseassays. Expression of STIP increased p53 protein levels morerapidly than UbcH5B/C knockdown (Fig. 6A), presumably re-flecting the time taken for the E2 levels to decrease as a resultof the RNAi response. �-Galactosidase reporter activity wasincreased by STIP but was not affected by UbcH5B/C suppres-sion (Fig. 6B). These data indicate that knockdown ofUbcH5B/C does not increase p53 transcriptional activity.
Mdm2 accumulates following UbcH5B/C knockdown. A pos-
FIG. 3. UbcH5B/C suppression causes accumulation of Mdm2in H1299 and U2-OS cells and inhibits Mdm2-mediated degrada-tion of p53. A, Western blots for endogenous target E2s, p53, andMdm2. H1299 and U2-OS cells were transfected with the indicatedpSUPER plasmids. 64-h post-transfection cells were harvested by directlysis into SDS-urea sample buffer. B, transient transfection with p53and Mdm2. U2-OS cells were transfected with human Mdm2 (1.5 �g),p53 (His-273) (0.5 �g), and empty pSUPER vector or pSUPER-UbcH5B/C (1.5 �g) as indicated. 64-h post-transfection exogenousMdm2 and p53 protein expression was determined by Western blotting.C, comparison of the absolute levels of UbcH5 protein expression indifferent cell lines. The levels of UbcH5A and UbcH5B/C proteins weredetermined by Western blotting with E2-specific antibodies using re-combinant proteins as standards. Values are means (expressed permicrogram of cell extract) � S.D. for four samples.
p53 Regulation by UbcH5B/C42174
by guest on July 8, 2016http://w
ww
.jbc.org/D
ownloaded from

sible explanation for the lack of p53 activation following knock-down of UbcH5B/C is that it is held inactive through binding toMdm2. To address this possibility we used STIP, which dis-rupts the interaction between p53 and Mdm2. Cells were firsttransfected with control or UbcH5B/C siRNA duplexes. After24 h the cells were replated and then transfected with emptythioredoxin vector (TRX) or STIP along with RGC�-Fos-LacZand pSVE-luciferase. 20 h later cells were harvested for West-ern blotting and for �-galactosidase assays. STIP up-regulated
p53 reporter activity in cells in which UbcH5B/C was sup-pressed (Fig. 6D). Reporter activity in UbcH5B/C siRNA- andSTIP-transfected cells was higher than in cells transfected withcontrol siRNA and STIP, correlating with the higher level ofp53 protein expression caused by UbcH5B/C knockdown, at thetime point examined (Fig. 6C). This suggests that the tran-scriptional activity of the p53 accumulated following suppres-sion of UbcH5B/C is repressed, at least in part, by Mdm2. STIPcan also probably target MdmX (63), and it is thus possible that
FIG. 4. Suppression of UbcH5B/C in MCF7 cells alters the pattern of p53 ubiquitination without altering the general pattern ofubiquitin conjugates. A, effect of UbcH5B/C knockdown on modification of endogenous p53. MCF7 cells were transfected with the indicatedpSUPER constructs and harvested 64 h later by direct lysis into SDS-urea sample buffer. Two different pSUPER-UbcH5B/C constructs(UbcH5B/C(2) and UbcH5B/C) were used to control for off-target effects. p53 protein expression was determined by Western blotting. To directlycompare the ratio of p53 conjugates to unmodified p53 under the different conditions, 3-fold more extract from control cells (Control 3�) was loadedso that levels of unmodified p53 were approximately equivalent. Where indicated MG132 (15 �M) was added for 4 h prior to harvesting. Lowerpanel: short exposure showing level of unmodified p53. Upper panel: extended exposure showing p53 conjugates. B, the effect of UbcH5B/Csuppression on p53 modification in the presence of MG132. MCF7 cells were transfected with the indicated synthetic siRNA duplex, 20 h aftertransfection MG132 (15 �M) was added, and cells were incubated for a further 4 h before harvesting. p53 protein expression was determined byWestern blotting. Lower panel: short exposure showing level of unmodified p53. Upper panel: extended exposure showing p53 conjugates. C,Ni2�-agarose purification of His6-ubiquitinated p53. MCF7 cells were transfected with His6-ubiquitin and pSUPER constructs as indicated.His6-ubiquitinated proteins were purified using Ni2�-NTA-agarose beads and analyzed by Western blotting for endogenous p53 (upper left panel).p53 conjugates present in total cell lysates (upper right panel) that co-migrate with ubiquitinated species of p53 are indicated with arrows. Theposition of a major band which is recognized non-specifically (NS) and of unmodified p53, which sticks to the beads independently of transfectionwith His6-ubiquitin (*) are also indicated. The level of unmodified p53 in total cell lysates was determined by Western blot analysis (lower panel).D, E2 suppression does not alter the general pattern of ubiquitin conjugates. MCF7 cells were transfected with the indicated pSUPER constructsand harvested 64 h later by direct lysis into SDS-urea sample buffer. Western blots were probed for UbcH5B/C and ubiquitin. Extracts from controland pSUPER UbcH5B/C transfected cells were those used in A.
p53 Regulation by UbcH5B/C 42175
by guest on July 8, 2016http://w
ww
.jbc.org/D
ownloaded from
MdmX could contribute to inactivation of the accumulated p53.The effect of UbcH5B/C knockdown on the mRNA levels of
endogenous p53 target genes was also investigated. We com-pared this to the effect of Mdm2 knockdown. MCF7 cells weretransfected with siRNA duplexes and harvested the indicatedtime after transfection (Fig. 7A). An increase in p53 proteinexpression was observed following suppression of Mdm2, whichoccurred within 24 h of transfection with Mdm2 siRNA. How-ever, p53 expression declined at longer time points, eventhough levels of Mdm2 remained low. UbcH5B/C knockdownresulted in a robust increase in p53 protein levels, which wassustained for at least 72 h. mRNA levels of endogenous p53target genes were determined by real-time RT-PCR 36 h aftertransfection of MCF7 cells with synthetic siRNA duplexes (Fig.7B). At this time point levels of p53 protein were maximalfollowing UbcH5B/C knockdown, and there was an increase inprotein expression of the CDK inhibitor p21 but levels of p53following suppression of Mdm2 were no longer elevated. Sup-pression of UbcH5B/C did not result in a significant increase inthe mRNA levels of the p53 target genes Mdm2, p21, or BAXconfirming that p53 transcriptional activity is not increased. Incontrast, up-regulation of p21 and BAX mRNA occurred follow-ing Mdm2 knockdown. Suppression of UbcH5B/C thus resultedin an increase in protein levels of p21 without an increase inp21 mRNA levels (Fig. 7, A and B). p21 is itself ubiquitinated
and undergoes proteasomal degradation (64). UbcH5B/C maythus also be involved in regulation of p21 protein stability.
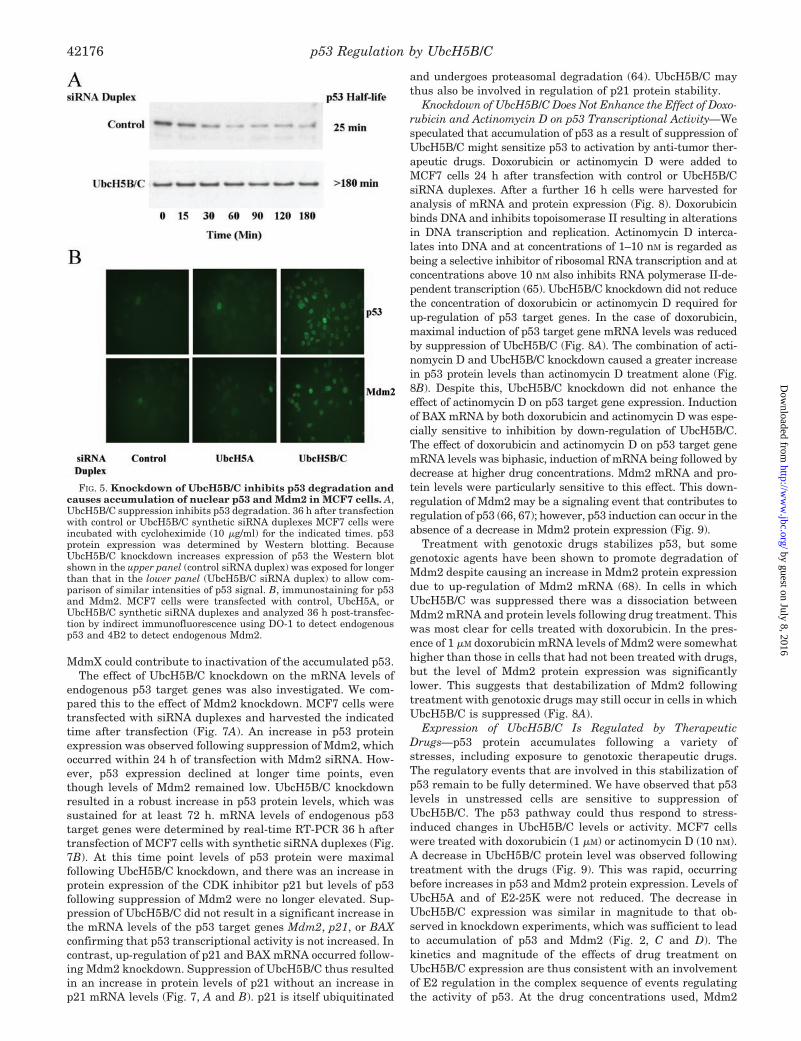
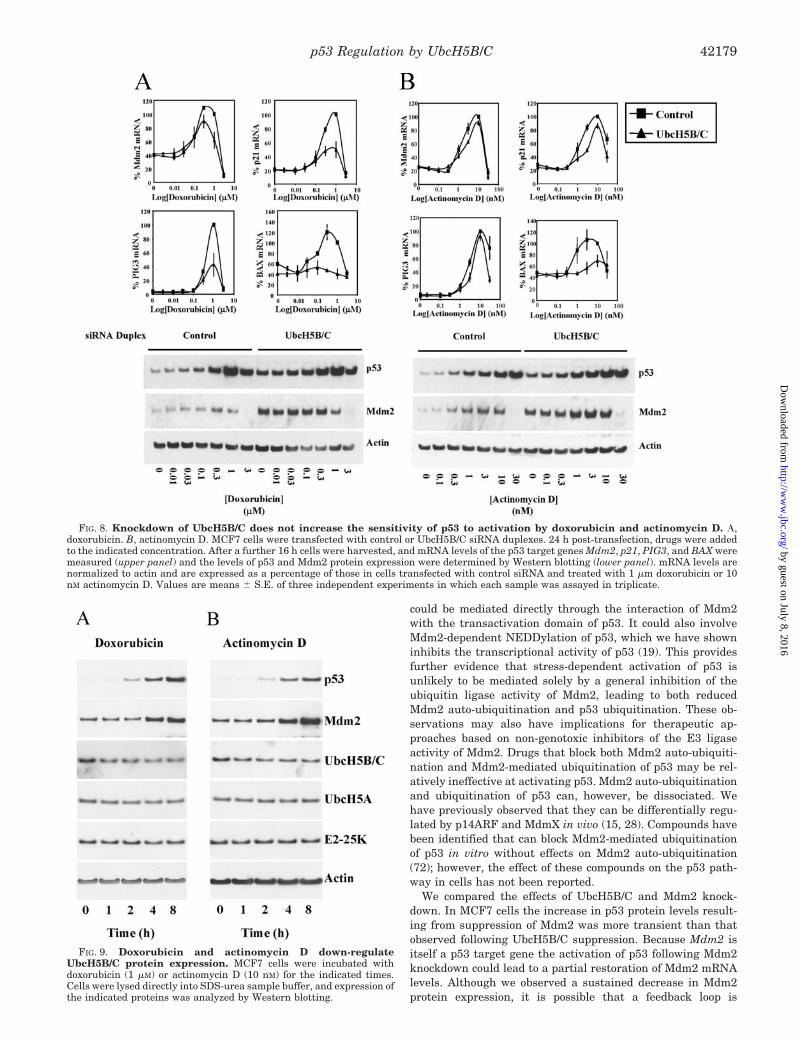
Knockdown of UbcH5B/C Does Not Enhance the Effect of Doxo-rubicin and Actinomycin D on p53 Transcriptional Activity—Wespeculated that accumulation of p53 as a result of suppression ofUbcH5B/C might sensitize p53 to activation by anti-tumor ther-apeutic drugs. Doxorubicin or actinomycin D were added toMCF7 cells 24 h after transfection with control or UbcH5B/CsiRNA duplexes. After a further 16 h cells were harvested foranalysis of mRNA and protein expression (Fig. 8). Doxorubicinbinds DNA and inhibits topoisomerase II resulting in alterationsin DNA transcription and replication. Actinomycin D interca-lates into DNA and at concentrations of 1–10 nM is regarded asbeing a selective inhibitor of ribosomal RNA transcription and atconcentrations above 10 nM also inhibits RNA polymerase II-de-pendent transcription (65). UbcH5B/C knockdown did not reducethe concentration of doxorubicin or actinomycin D required forup-regulation of p53 target genes. In the case of doxorubicin,maximal induction of p53 target gene mRNA levels was reducedby suppression of UbcH5B/C (Fig. 8A). The combination of acti-nomycin D and UbcH5B/C knockdown caused a greater increasein p53 protein levels than actinomycin D treatment alone (Fig.8B). Despite this, UbcH5B/C knockdown did not enhance theeffect of actinomycin D on p53 target gene expression. Inductionof BAX mRNA by both doxorubicin and actinomycin D was espe-cially sensitive to inhibition by down-regulation of UbcH5B/C.The effect of doxorubicin and actinomycin D on p53 target genemRNA levels was biphasic, induction of mRNA being followed bydecrease at higher drug concentrations. Mdm2 mRNA and pro-tein levels were particularly sensitive to this effect. This down-regulation of Mdm2 may be a signaling event that contributes toregulation of p53 (66, 67); however, p53 induction can occur in theabsence of a decrease in Mdm2 protein expression (Fig. 9).
Treatment with genotoxic drugs stabilizes p53, but somegenotoxic agents have been shown to promote degradation ofMdm2 despite causing an increase in Mdm2 protein expressiondue to up-regulation of Mdm2 mRNA (68). In cells in whichUbcH5B/C was suppressed there was a dissociation betweenMdm2 mRNA and protein levels following drug treatment. Thiswas most clear for cells treated with doxorubicin. In the pres-ence of 1 �M doxorubicin mRNA levels of Mdm2 were somewhathigher than those in cells that had not been treated with drugs,but the level of Mdm2 protein expression was significantlylower. This suggests that destabilization of Mdm2 followingtreatment with genotoxic drugs may still occur in cells in whichUbcH5B/C is suppressed (Fig. 8A).
Expression of UbcH5B/C Is Regulated by TherapeuticDrugs—p53 protein accumulates following a variety ofstresses, including exposure to genotoxic therapeutic drugs.The regulatory events that are involved in this stabilization ofp53 remain to be fully determined. We have observed that p53levels in unstressed cells are sensitive to suppression ofUbcH5B/C. The p53 pathway could thus respond to stress-induced changes in UbcH5B/C levels or activity. MCF7 cellswere treated with doxorubicin (1 �M) or actinomycin D (10 nM).A decrease in UbcH5B/C protein level was observed followingtreatment with the drugs (Fig. 9). This was rapid, occurringbefore increases in p53 and Mdm2 protein expression. Levels ofUbcH5A and of E2-25K were not reduced. The decrease inUbcH5B/C expression was similar in magnitude to that ob-served in knockdown experiments, which was sufficient to leadto accumulation of p53 and Mdm2 (Fig. 2, C and D). Thekinetics and magnitude of the effects of drug treatment onUbcH5B/C expression are thus consistent with an involvementof E2 regulation in the complex sequence of events regulatingthe activity of p53. At the drug concentrations used, Mdm2
FIG. 5. Knockdown of UbcH5B/C inhibits p53 degradation andcauses accumulation of nuclear p53 and Mdm2 in MCF7 cells. A,UbcH5B/C suppression inhibits p53 degradation. 36 h after transfectionwith control or UbcH5B/C synthetic siRNA duplexes MCF7 cells wereincubated with cycloheximide (10 �g/ml) for the indicated times. p53protein expression was determined by Western blotting. BecauseUbcH5B/C knockdown increases expression of p53 the Western blotshown in the upper panel (control siRNA duplex) was exposed for longerthan that in the lower panel (UbcH5B/C siRNA duplex) to allow com-parison of similar intensities of p53 signal. B, immunostaining for p53and Mdm2. MCF7 cells were transfected with control, UbcH5A, orUbcH5B/C synthetic siRNA duplexes and analyzed 36 h post-transfec-tion by indirect immunofluorescence using DO-1 to detect endogenousp53 and 4B2 to detect endogenous Mdm2.
p53 Regulation by UbcH5B/C42176
by guest on July 8, 2016http://w
ww
.jbc.org/D
ownloaded from
levels did not decrease prior to p53 induction. This indicatesthat, although down-regulation of Mdm2 protein expressionmay play some part in control of p53, it is not an obligatory stepfor induction of p53 in MCF7 cells.
DISCUSSION
In this study we show that UbcH5B/C are required for themaintenance of low levels of p53 and Mdm2 in unstressed cells.UbcH5B/C participate in p53 ubiquitination and in Mdm2-mediated degradation of p53 in vivo, indicating that they arephysiological E2s for Mdm2. Suppression of UbcH5B/C resultsin the accumulation of nuclear p53 but does not on its ownincrease p53 transcriptional activity. A reduction in UbcH5B/Cexpression occurs following treatment with doxorubicin andactinomycin D raising the possibility that E2 regulation mayparticipate in signaling to p53.
We examined the ubiquitin E2 specificity of the E3 ligaseMdm2. In vitro auto-ubiquitination of Mdm2 and Mdm2-de-pendent ubiquitination of p53 were supported by UbcH5A, -B,and -C and by E2-25K. The pattern of Mdm2-mediated p53ubiquitination was similar with UbcH5 family members, whichgave predominantly low molecular weight ubiquitinated spe-cies. This is consistent with the observation that UbcH5B sup-ports multiple mono-ubiquitination of p53 in vitro (69). Mdm2promoted the formation of high molecular weight p53-ubiquitin
conjugates in the presence of E2-25K. This may be related tothe propensity of E2-25K to form chains of ubiquitin. E2-25Kdisplays the unique property of mediating the formation ofunanchored Lys-48- to Gly-76-linked polyubiquitin chains invitro without the requirement for an E3 or acceptor substrate(70). Only a small proportion of the total ubiquitin added to invitro reactions was converted to unanchored polyubiquitinchains (data not shown). Therefore E2-25K may selectivelytransfer polyubiquitin chains to p53 in preference to singleubiquitin molecules, or E2-25K may efficiently extend chains ofubiquitin conjugated to p53.
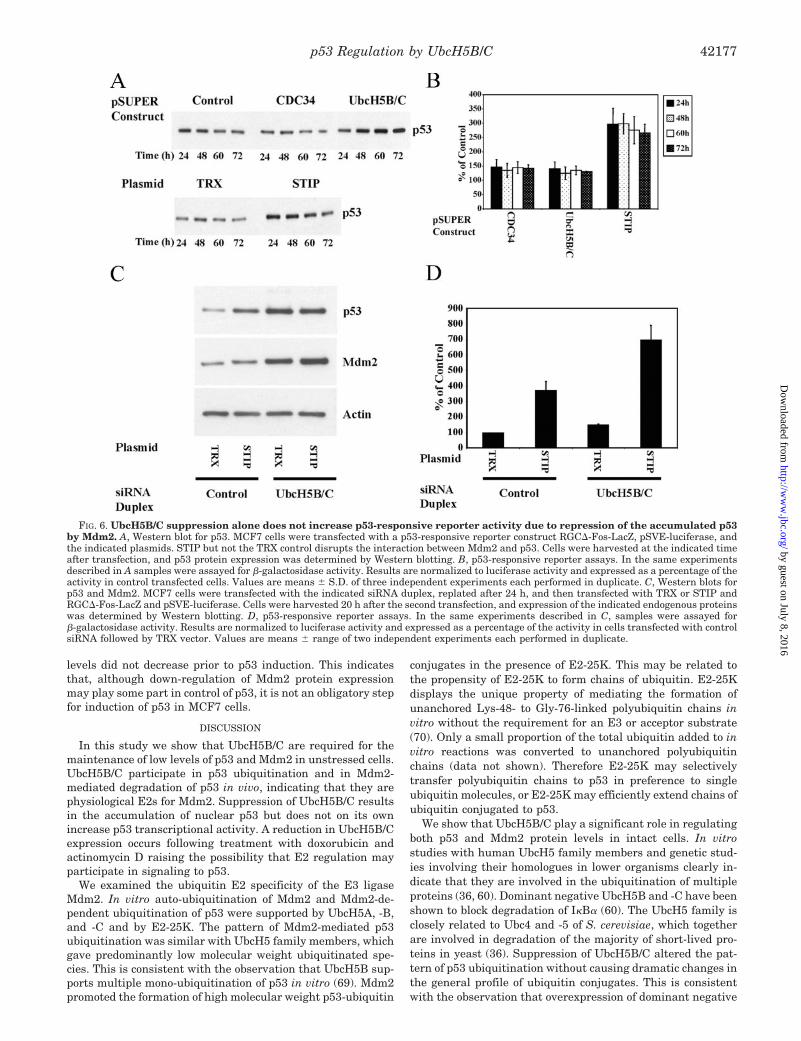
We show that UbcH5B/C play a significant role in regulatingboth p53 and Mdm2 protein levels in intact cells. In vitrostudies with human UbcH5 family members and genetic stud-ies involving their homologues in lower organisms clearly in-dicate that they are involved in the ubiquitination of multipleproteins (36, 60). Dominant negative UbcH5B and -C have beenshown to block degradation of I�B� (60). The UbcH5 family isclosely related to Ubc4 and -5 of S. cerevisiae, which togetherare involved in degradation of the majority of short-lived pro-teins in yeast (36). Suppression of UbcH5B/C altered the pat-tern of p53 ubiquitination without causing dramatic changes inthe general profile of ubiquitin conjugates. This is consistentwith the observation that overexpression of dominant negative
FIG. 6. UbcH5B/C suppression alone does not increase p53-responsive reporter activity due to repression of the accumulated p53by Mdm2. A, Western blot for p53. MCF7 cells were transfected with a p53-responsive reporter construct RGC�-Fos-LacZ, pSVE-luciferase, andthe indicated plasmids. STIP but not the TRX control disrupts the interaction between Mdm2 and p53. Cells were harvested at the indicated timeafter transfection, and p53 protein expression was determined by Western blotting. B, p53-responsive reporter assays. In the same experimentsdescribed in A samples were assayed for �-galactosidase activity. Results are normalized to luciferase activity and expressed as a percentage of theactivity in control transfected cells. Values are means � S.D. of three independent experiments each performed in duplicate. C, Western blots forp53 and Mdm2. MCF7 cells were transfected with the indicated siRNA duplex, replated after 24 h, and then transfected with TRX or STIP andRGC�-Fos-LacZ and pSVE-luciferase. Cells were harvested 20 h after the second transfection, and expression of the indicated endogenous proteinswas determined by Western blotting. D, p53-responsive reporter assays. In the same experiments described in C, samples were assayed for�-galactosidase activity. Results are normalized to luciferase activity and expressed as a percentage of the activity in cells transfected with controlsiRNA followed by TRX vector. Values are means � range of two independent experiments each performed in duplicate.
p53 Regulation by UbcH5B/C 42177
by guest on July 8, 2016http://w
ww
.jbc.org/D
ownloaded from

UbcH5B or -C does not change the pattern of ubiquitination inHeLa cells (60). UbcH5B/C may thus be required for the ubiq-uitination of a specific subset of proteins in vivo. The humangenome encodes for more E2s than that of S. cerevisiae andcontains several homologues of S. cerevisiae Ubc4/5 (11, 56).This provides the scope for both divergence and for greaterredundancy of E2 function in humans. Alternatively, ubiquiti-nation of p53 may be more sensitive to changes in UbcH5B/Clevels than the ubiquitination of the major UbcH5B/C sub-strates. Consistent with this the Km of UbcH5B for Mdm2-mediated ubiquitination of p53 is �3 �M (69), whereas theapparent Km of a rat homologue for stimulation of ubiquitina-tion of its major substrates in rat tissue extracts is in the regionof 50 nM (35).
Both UbcH5A and E2-25K were able to support the ubiquitinligase activity of Mdm2 in vitro; however, in the cell lines testedno up-regulation of Mdm2 or p53 was observed following sup-pression of these E2s. This dependence on UbcH5B/C ratherthan UbcH5A in vivo could reflect differences in expression ofthe E2s. In MCF7, U2-OS, and H1299 cells UbcH5B/C proteinexpression was higher than that of UbcH5A. This may gener-ally be the case, because previous studies have shown thatmRNA levels of UbcH5B/C are higher than those of UbcH5A ina range of tissues (56, 59). We cannot exclude the possibilitythat UbcH5A or E2-25K may participate in the regulation ofMdm2 and p53 under different conditions or in different cell
types. Overexpression of Mdm2 to moderate levels stimulatesprincipally multiple mono-ubiquitination of p53, however over-expression to higher levels has been reported to promote p53polyubiquitination (62). Polyubiquitinated rather than mono-ubiquitinated p53 may be targeted for proteasomal degrada-tion. Further studies are required to determine whether E2-25K plays a part in this polyubiquitination of p53. It is ofinterest to note that E2-25K is induced by low density lipopro-tein in human macrophages (71). Up-regulation of E2-25K un-der these circumstances is associated with a decrease in p53levels and with a decrease in the apoptotic potential of the cells,however, the direct causal linkage of these events has not beendemonstrated. We did not detect p53 ubiquitination withHHR6A or -B in vitro; however, these E2s have been found incomplexes with p53 in cells (57), and it remains to be deter-mined whether they could also be involved in p53 ubiquitina-tion in vivo.
Despite causing accumulation of nuclear p53, suppression ofUbcH5B/C did not on its own increase p53 transcriptionalactivity, and it did not enhance the sensitivity of p53 to acti-vation by doxorubicin or actinomycin D. Suppression ofUbcH5B/C resulted in an increase in the level of endogenousMdm2. Experiments involving the expression of STIP, whichdisrupts the interaction between Mdm2 and p53, indicated thatin cells in which UbcH5B/C are suppressed, the accumulatedp53 is repressed at least in part, by Mdm2. This repression
FIG. 7. Comparison of the effects of UbcH5B/C and Mdm2 suppression on p53 protein levels and on the mRNA levels of endogenousp53 target genes. A, the effects of Mdm2 and UbcH5B/C knockdown on p53 and p21 protein expression. MCF7 cells were transfected with siRNAduplexes and harvested at the indicated times. Levels of protein expression were determined by Western blotting. B, measurement of endogenousp53 target gene mRNA levels by real-time RT-PCR. MCF7 cells were transfected with synthetic siRNA duplexes. After 36 h protein expression wasanalyzed by Western blotting and mRNA levels of p53 and the p53 target genes Mdm2, p21, and BAX determined. mRNA levels normalized to actinare expressed as a percentage of the levels in cells transfected with control siRNA. Values are means � S.D. of three independent experiments inwhich each sample was assayed in triplicate.
p53 Regulation by UbcH5B/C42178
by guest on July 8, 2016http://w
ww
.jbc.org/D
ownloaded from
could be mediated directly through the interaction of Mdm2with the transactivation domain of p53. It could also involveMdm2-dependent NEDDylation of p53, which we have showninhibits the transcriptional activity of p53 (19). This providesfurther evidence that stress-dependent activation of p53 isunlikely to be mediated solely by a general inhibition of theubiquitin ligase activity of Mdm2, leading to both reducedMdm2 auto-ubiquitination and p53 ubiquitination. These ob-servations may also have implications for therapeutic ap-proaches based on non-genotoxic inhibitors of the E3 ligaseactivity of Mdm2. Drugs that block both Mdm2 auto-ubiquiti-nation and Mdm2-mediated ubiquitination of p53 may be rel-atively ineffective at activating p53. Mdm2 auto-ubiquitinationand ubiquitination of p53 can, however, be dissociated. Wehave previously observed that they can be differentially regu-lated by p14ARF and MdmX in vivo (15, 28). Compounds havebeen identified that can block Mdm2-mediated ubiquitinationof p53 in vitro without effects on Mdm2 auto-ubiquitination(72); however, the effect of these compounds on the p53 path-way in cells has not been reported.
We compared the effects of UbcH5B/C and Mdm2 knock-down. In MCF7 cells the increase in p53 protein levels result-ing from suppression of Mdm2 was more transient than thatobserved following UbcH5B/C suppression. Because Mdm2 isitself a p53 target gene the activation of p53 following Mdm2knockdown could lead to a partial restoration of Mdm2 mRNAlevels. Although we observed a sustained decrease in Mdm2protein expression, it is possible that a feedback loop is
FIG. 9. Doxorubicin and actinomycin D down-regulateUbcH5B/C protein expression. MCF7 cells were incubated withdoxorubicin (1 �M) or actinomycin D (10 nM) for the indicated times.Cells were lysed directly into SDS-urea sample buffer, and expression ofthe indicated proteins was analyzed by Western blotting.
FIG. 8. Knockdown of UbcH5B/C does not increase the sensitivity of p53 to activation by doxorubicin and actinomycin D. �,doxorubicin. B, actinomycin D. MCF7 cells were transfected with control or UbcH5B/C siRNA duplexes. 24 h post-transfection, drugs were addedto the indicated concentration. After a further 16 h cells were harvested, and mRNA levels of the p53 target genes Mdm2, p21, PIG3, and BAX weremeasured (upper panel) and the levels of p53 and Mdm2 protein expression were determined by Western blotting (lower panel). mRNA levels arenormalized to actin and are expressed as a percentage of those in cells transfected with control siRNA and treated with 1 �m doxorubicin or 10nM actinomycin D. Values are means � S.E. of three independent experiments in which each sample was assayed in triplicate.
p53 Regulation by UbcH5B/C 42179
by guest on July 8, 2016http://w
ww
.jbc.org/D
ownloaded from

achieved that limits p53 induction. As well as p53 degradationmediated by viral proteins, there are a number of reports ofMdm2-independent p53 degradation in the normal cellularcontext (73–75). Pirh2 and COP1 can stimulate the ubiquiti-nation and degradation of p53 independently of Mdm2 in mam-malian cells (76, 77). Pirh2 and COP1 are themselves p53target genes and may be up-regulated in response to Mdm2knockdown. It has been shown in U2-OS cells that knockdownof COP1 and Mdm2 in combination has a greater effect on p53stability than knockdown of Mdm2 alone (77). UbcH5 familymembers support ubiquitination of p53 in vitro mediated byPirh2 and COP1. If this is the case in vivo, then targeting ofUbcH5B/C may have a more robust effect on p53 stability thantargeting individual p53 E3 ubiquitin ligases.
p53 protein expression is sensitive to the level of UbcH5B/Cindicating that the activity of UbcH5B/C is close to rate-limit-ing for p53 degradation in the cell lines examined. Changes inthe level or activity of UbcH5B/C could thus be involved inregulating the p53 pathway. E2 regulation might be involved inresponses to cellular stress. It could participate in a generalstress response, which affects multiple UbcH5B/C substrates,however it may selectively affect particular pathways depend-ent for example, on the affinity of UbcH5B/C for the E3s in-volved. The p53 pathway appears to be tuned such that it issensitive to modulation of UbcH5B/C levels. We observed adecrease in UbcH5B/C expression following treatment with thetherapeutic drugs doxorubicin and actinomycin D. This pre-ceded induction of p53 by the drugs and its magnitude wassufficient to influence p53 levels. Consistent with an involve-ment of E2 down-regulation in signaling to p53, accumulationof p53 following exposure to doxorubicin and actinomycin D isassociated with attenuation of its ubiquitination and conse-quent protein stabilization (78). However, our data indicatethat the decrease in UbcH5B/C levels could not account for themagnitude of the increase in p53 levels following treatmentwith these drugs, and in addition we have demonstrated that areduction in E2 levels would not on its own lead to an increasein p53 transcriptional activity. It is possible that a reduction inUbcH5B/C expression could participate in regulation of p53,but additional signaling events are undoubtedly required toachieve full p53 activation. It has recently been shown that,under conditions where several genotoxic agents increaseMdm2 protein expression due to up-regulation of its mRNAlevels, Mdm2 protein stability is actually reduced through anincompletely defined mechanism (68). It has also been reportedthat genotoxic agents, including doxorubicin, promote protea-somal degradation of MdmX and that this has a requirementfor Mdm2 (79). These effects on Mdm2 and MdmX stabilityserve to regulate the ratio of these repressors to p53, a param-eter which our study confirms is a critical determinant of p53activity. This destabilization could involve stress-dependentmodification of Mdm2, which changes the sites or pattern ofMdm2 auto-ubiquitination or which selectively increases theefficiency of auto-ubiquitination through an increase in therate of transfer of ubiquitin to Mdm2 from the remainingUbcH5B/C. It could result from a stress-dependent decrease inMdm2 de-ubiquitinating activity or signaling events, whichenhance proteasomal recognition of Mdm2. Enhanced degrada-tion of Mdm2/MdmX could also result from the targeting ofMdm2 by another E2/E3. It is conceivable that a reduction inUbcH5B/C levels could participate in a switch from Mdm2/p53to Mdm2/MdmX degradation. Although UbcH5B/C make a ma-jor contribution to the maintenance of low levels of Mdm2 andp53 in unstressed cells, it remains a possibility that another E2may participate in Mdm2/MdmX degradation following cellstress. One possible model is that a correctly timed decrease in
UbcH5B/C expression could be involved in facilitating the in-teraction of Mdm2 with a distinct stress-activated E2, whichfavors Mdm2/MdmX degradation over p53 degradation. p53 isactivated by diverse stresses and is regulated by multiple sig-naling events (1, 80, 81). Further studies will be required todetermine the role of E2 regulation and of modulation of theinteraction between Mdm2 and E2s in the signaling pathwayscontrolling p53 and to determine the mechanism of the drug-dependent changes in expression of UbcH5B/C.
Acknowledgments—We are grateful to Drs R. T. Hay, K. Madura,R. Bernards, and R. Watson for providing us with reagents.
REFERENCES
1. Ryan, K. M., Phillips, A. C., and Vousden, K. H. (2001) Curr. Opin. Cell Biol.13, 332–337
2. Oren, M. (2003) Cell Death Differ. 10, 431–4423. Lain, S., and Lane, D. (2003) Eur. J. Cancer 39, 1053–10604. Woods, Y. L., and Lane, D. P. (2003) Hematol. J. 4, 233–2475. Yang, Y., Li, C. C., and Weissman, A. M. (2004) Oncogene 23, 2096–21066. Richardson, P. G., Barlogie, B., Berenson, J., Singhal, S., Jagannath, S., Irwin,
D., Rajkumar, S. V., Srkalovic, G., Alsina, M., Alexanian, R., Siegel, D.,Orlowski, R. Z., Kuter, D., Limentani, S. A., Lee, S., Hideshima, T., Essel-tine, D. L., Kauffman, M., Adams, J., Schenkein, D. P., and Anderson, K. C.(2003) N. Engl. J. Med. 348, 2609–2617
7. Voorhees, P. M., Dees, E. C., O’Neil, B., and Orlowski, R. Z. (2003) Clin. CancerRes. 9, 6316–6325
8. Lara, P. N., Jr., Davies, A. M., Mack, P. C., Mortenson, M. M., Bold, R. J.,Gumerlock, P. H., and Gandara, D. R. (2004) Semin. Oncol. 31, 40–46
9. Hershko, A., and Ciechanover, A. (1998) Annu. Rev. Biochem. 67, 425–47910. Pickart, C. M. (2001) Annu. Rev. Biochem. 70, 503–53311. Jones, D., Crowe, E., Stevens, T. A., and Candido, E. P. (2002) Genome Biology
http://genomebiology.com/2001/3/1/RESEARCH/000212. Honda, R., Tanaka, H., and Yasuda, H. (1997) FEBS Lett. 420, 25–2713. Haupt, Y., Maya, R., Kazaz, A., and Oren, M. (1997) Nature 387, 296–29914. Kubbutat, M. H., Jones, S. N., and Vousden, K. H. (1997) Nature 387, 299–30315. Xirodimas, D., Saville, M. K., Edling, C., Lane, D. P., and Lain, S. (2001)
Oncogene 20, 4972–498316. Momand, J., Zambetti, G. P., Olson, D. C., George, D., and Levine, A. J. (1992)
Cell 69, 1237–124517. Chen, J., Marechal, V., and Levine, A. J. (1993) Mol. Cell. Biol. 13, 4107–411418. Thut, C. J., Goodrich, J. A., and Tjian, R. (1997) Genes Dev. 11, 1974–198619. Xirodimas, D. P., Saville, M. K., Bourdon, J. C., Hay, R. T., and Lane, D. P.
(2004) Cell 118, 83–9720. Jones, S. N., Roe, A. E., Donehower, L. A., and Bradley, A. (1995) Nature 378,
206–20821. Montes de Oca Luna, R., Wagner, D. S., and Lozano, G. (1995) Nature 378,
203–20622. Mendrysa, S. M., McElwee, M. K., Michalowski, J., O’Leary, K. A., Young,
K. M., and Perry, M. E. (2003) Mol. Cell. Biol. 23, 462–47223. Bottger, A., Bottger, V., Sparks, A., Liu, W. L., Howard, S. F., and Lane, D. P.
(1997) Curr. Biol. 7, 860–86924. Vassilev, L. T., Vu, B. T., Graves, B., Carvajal, D., Podlaski, F., Filipovic, Z.,
Kong, N., Kammlott, U., Lukacs, C., Klein, C., Fotouhi, N., and Liu, E. A.(2004) Science 303, 844–848
25. Freedman, D. A., Wu, L., and Levine, A. J. (1999) Cell Mol. Life Sci. 55, 96–10726. Michael, D., and Oren, M. (2003) Semin. Cancer Biol. 13, 49–5827. Lane, D. P., and Hall, P. A. (1997) Trends Biochem. Sci. 22, 372–37428. Stad, R., Little, N. A., Xirodimas, D. P., Frenk, R., van der Eb, A. J., Lane,
D. P., Saville, M. K., and Jochemsen, A. G. (2001) EMBO Rep. 2, 1029–103429. Badciong, J. C., and Haas, A. L. (2002) J. Biol. Chem. 277, 49668–4967530. Tanimura, S., Ohtsuka, S., Mitsui, K., Shirouzu, K., Yoshimura, A., and
Ohtsubo, M. (1999) FEBS Lett. 447, 5–931. Parant, J., Chavez-Reyes, A., Little, N. A., Yan, W., Reinke, V., Jochemsen,
A. G., and Lozano, G. (2001) Nat. Genet. 29, 92–9532. Finch, R. A., Donoviel, D. B., Potter, D., Shi, M., Fan, A., Freed, D. D., Wang,
C. Y., Zambrowicz, B. P., Ramirez-Solis, R., Sands, A. T., and Zhang, N.(2002) Cancer Res. 62, 3221–3225
33. Fang, S., Jensen, J. P., Ludwig, R. L., Vousden, K. H., and Weissman, A. M.(2000) J. Biol. Chem. 275, 8945–8951
34. Midgley, C. A., Desterro, J. M., Saville, M. K., Howard, S., Sparks, A., Hay,R. T., and Lane, D. P. (2000) Oncogene 19, 2312–2323
35. Rajapurohitam, V., Bedard, N., and Wing, S. S. (2002) Am. J. Physiol. 282,E739–E745
36. Seufert, W., and Jentsch, S. (1990) EMBO J. 9, 543–55037. Wing, S. S., and Banville, D. (1994) Am. J. Physiol. 267, E39–E4838. Wing, S. S., and Bedard, N. (1996) Biochem. J. 319, 455–46139. Shang, F., Gong, X., and Taylor, A. (1997) J. Biol. Chem. 272, 23086–2309340. Rajapurohitam, V., Morales, C., El-Alfy, M., Lefrancois, S., Bedard, N., and
Wing, S. (1999) Dev. Biol. 212, 217–22841. Bastians, H., Topper, L. M., Gorbsky, G. L., and Ruderman, J. V. (1999) Mol.
Biol. Cell 10, 3927–394142. Nyman, T. A., Matikainen, S., Sareneva, T., Julkunen, I., and Kalkkinen, N.
(2000) Eur. J. Biochem. 267, 4011–401943. Song, S., Kim, S. Y., Hong, Y. M., Jo, D. G., Lee, J. Y., Shim, S. M., Chung,
C. W., Seo, S. J., Yoo, Y. J., Koh, J. Y., Lee, M. C., Yates, A. J., Ichijo, H., andJung, Y. K. (2003) Mol. Cell 12, 553–563
44. Gu, H., and Roizman, B. (2003) Proc. Natl. Acad. Sci. U. S. A. 100, 8963–896845. Liu, Q., Shang, F., Guo, W., Hobbs, M., Valverde, P., Reddy, V., and Taylor, A.
p53 Regulation by UbcH5B/C42180
by guest on July 8, 2016http://w
ww
.jbc.org/D
ownloaded from

(2004) Exp. Eye Res. 78, 197–20546. Semplici, F., Meggio, F., Pinna, L. A., and Oliviero, S. (2002) Oncogene 21,
3978–398747. Sarcevic, B., Mawson, A., Baker, R. T., and Sutherland, R. L. (2002) EMBO J.
21, 2009–201848. Midgley, C. A., Owens, B., Briscoe, C. V., Thomas, D. B., Lane, D. P., and Hall,
P. A. (1995) J. Cell Sci. 108, 1843–184849. Elbashir, S. M., Harborth, J., Lendeckel, W., Yalcin, A., Weber, K., and Tuschl,
T. (2001) Nature 411, 494–49850. Brummelkamp, T. R., Bernards, R., and Agami, R. (2002) Science 296,
550–55351. Desterro, J. M., Rodriguez, M. S., Kemp, G. D., and Hay, R. T. (1999) J. Biol.
Chem. 274, 10618–1062452. Kaiser, P., Seufert, W., Hofferer, L., Kofler, B., Sachsenmaier, C., Herzog, H.,
Jentsch, S., Schweiger, M., and Schneider, R. (1994) J. Biol. Chem. 269,8797–8802
53. Desterro, J. M., Thomson, J., and Hay, R. T. (1997) FEBS Lett. 417, 297–30054. Tongaonkar, P., and Madura, K. (1998) Anal. Biochem. 260, 135–14155. Hansen, S., Hupp, T. R., and Lane, D. P. (1996) J. Biol. Chem. 271, 3917–392456. Okamoto, Y., Ozaki, T., Miyazaki, K., Aoyama, M., Miyazaki, M., and Naka-
gawara, A. (2003) Cancer Res. 63, 4167–417357. Lyakhovich, A., and Shekhar, M. P. (2003) Mol. Cell. Biol. 23, 2463–247558. Scheffner, M., Huibregtse, J. M., and Howley, P. M. (1994) Proc. Natl. Acad.
Sci. U. S. A. 91, 8797–880159. Jensen, J. P., Bates, P. W., Yang, M., Vierstra, R. D., and Weissman, A. M.
(1995) J. Biol. Chem. 270, 30408–3041460. Gonen, H., Bercovich, B., Orian, A., Carrano, A., Takizawa, C., Yamanaka, K.,
Pagano, M., Iwai, K., and Ciechanover, A. (1999) J. Biol. Chem. 274,14823–14830
61. Mimnaugh, E. G., Bonvini, P., and Neckers, L. (1999) Electrophoresis 20,418–428
62. Li, M., Brooks, C. L., Wu-Baer, F., Chen, D., Baer, R., and Gu, W. (2003)Science 302, 1972–1975
63. Bottger, V., Bottger, A., Garcia-Echeverria, C., Ramos, Y. F., van der Eb, A. J.,
Jochemsen, A. G., and Lane, D. P. (1999) Oncogene 18, 189–19964. Bloom, J., and Pagano, M. (2004) Cell Cycle 3, 138–14065. Ochs, R. L. (1998) Methods Cell Biol. 53, 303–32166. Blattner, C., Sparks, A., and Lane, D. (1999) Mol. Cell. Biol. 19, 3704–371367. Ma, Y., Yuan, R., Meng, Q., Goldberg, I. D., Rosen, E. M., and Fan, S. (2000)
Mol. Cell. Biol. Res. Commun. 3, 122–12868. Stommel, J. M., and Wahl, G. M. (2004) EMBO J. 23, 1547–155669. Lai, Z., Ferry, K. V., Diamond, M. A., Wee, K. E., Kim, Y. B., Ma, J., Yang, T.,
Benfield, P. A., Copeland, R. A., and Auger, K. R. (2001) J. Biol. Chem. 276,31357–31367
70. Haldeman, M. T., Xia, G., Kasperek, E. M., and Pickart, C. M. (1997) Biochem-istry 36, 10526–10537
71. Kikuchi, J., Furukawa, Y., Kubo, N., Tokura, A., Hayashi, N., Nakamura, M.,Matsuda, M., and Sakurabayashi, I. (2000) Arterioscler. Thromb. Vasc. Biol.20, 128–134
72. Lai, Z., Yang, T., Kim, Y. B., Sielecki, T. M., Diamond, M. A., Strack, P., Rolfe,M., Caligiuri, M., Benfield, P. A., Auger, K. R., and Copeland, R. A. (2002)Proc. Natl. Acad. Sci. U. S. A. 99, 14734–14739
73. Fuchs, S. Y., Adler, V., Buschmann, T., Yin, Z., Wu, X., Jones, S. N., and Ronai,Z. (1998) Genes Dev. 12, 2658–2663
74. Bech-Otschir, D., Kraft, R., Huang, X., Henklein, P., Kapelari, B., Pollmann,C., and Dubiel, W. (2001) EMBO J. 20, 1630–1639
75. Asher, G., Lotem, J., Sachs, L., Kahana, C., and Shaul, Y. (2002) Proc. Natl.Acad. Sci. U. S. A. 99, 13125–13130
76. Leng, R. P., Lin, Y., Ma, W., Wu, H., Lemmers, B., Chung, S., Parant, J. M.,Lozano, G., Hakem, R., and Benchimol, S. (2003) Cell 112, 779–791
77. Dornan, D., Wertz, I., Shimizu, H., Arnott, D., Frantz, G. D., Dowd, P.,O’Rourke, K., Koeppen, H., and Dixit, V. M. (2004) Nature 429, 86–92
78. Isaacs, J. S., Saito, S., and Neckers, L. M. (2001) J. Biol. Chem. 276,18497–18506
79. Kawai, H., Wiederschain, D., Kitao, H., Stuart, J., Tsai, K. K., and Yuan, Z. M.(2003) J. Biol. Chem. 278, 45946–45953
80. Brooks, C. L., and Gu, W. (2003) Curr. Opin. Cell Biol. 15, 164–17181. Meek, D. W., and Knippschild, U. (2003) Mol. Cancer Res. 1, 1017–1026
p53 Regulation by UbcH5B/C 42181
by guest on July 8, 2016http://w
ww
.jbc.org/D
ownloaded from

Stevenson, Jean-Christophe Bourdon, Yvonne L. Woods and David P. LaneMark K. Saville, Alison Sparks, Dimitris P. Xirodimas, Julie Wardrop, Lauren F.
in VivoRegulation of p53 by the Ubiquitin-conjugating Enzymes UbcH5B/C
doi: 10.1074/jbc.M403362200 originally published online July 26, 20042004, 279:42169-42181.J. Biol. Chem.
10.1074/jbc.M403362200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/279/40/42169.full.html#ref-list-1
This article cites 80 references, 35 of which can be accessed free at
by guest on July 8, 2016http://w
ww
.jbc.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















