![Near-Membrane [Ca2+] Transients Resolved Using the Ca2+ Indicator FFP18](https://static.fdokumen.com/doc/165x107/631286873ed465f0570a4533/near-membrane-ca2-transients-resolved-using-the-ca2-indicator-ffp18.jpg)
Near-Membrane [Ca2+] Transients Resolved Using the Ca2+ Indicator FFP18

Real Time Measurements of Water Flow in AmphibianGastric GlandsMODULATION VIA THE EXTRACELLULAR Ca2�-SENSING RECEPTOR*
Received for publication, November 14, 2006, and in revised form, February 9, 2007 Published, JBC Papers in Press, March 15, 2007, DOI 10.1074/jbc.M610585200
Andrea Gerbino‡1, Gregorio Fistetto‡2, Matilde Colella‡1, Aldebaran M. Hofer§3, Lucantonio Debellis‡1,Rosa Caroppo‡1, and Silvana Curci§4
From the ‡Dipartimento di Fisiologia Generale ed Ambientale, Universita’ di Bari, 70126 Bari, Italy and the §Veterans Affairs BostonHealthcare System and the Department of Surgery, Harvard Medical School and Brigham and Women’s Hospital,West Roxbury, Massachusetts 02132
The mechanisms for the formation of the osmotic gradientdriving water movements in the gastric gland and its modula-tion via the extracellular Ca2�-sensing receptor (CaR) wereinvestigated. Real time measurements of net water flux in thelumen of single gastric glands of the intact amphibian stomachwere performed using ion-selective double-barreled microelec-trodes.Watermovement wasmeasured by recording changes inthe concentration of impermeant TEA� ions ([TEA�]gl) withTEA�-sensitive microelectrodes inserted in the lumen of indi-vidual gastric glands. Glandular K� (K�
gl) and H� (pHgl) werealso measured by using K�- and H�-sensitive microelectrodes,respectively. Stimulationwith histamine significantly decreased[TEA]gl, indicating net water flow toward the gland lumen. Thisresponse was inhibited by the H�/K�-ATPase inhibitor, SCH28080. Histamine also elicited a significant and reversibleincrease in [K�]gl that was blocked by chromanol 293B, ablocker of KCQN1 K� channels. Histamine failed to induce netwater flow in the presence of chromanol 293B. In the “restingstate,” stimulation of CaR with diverse agonists resulted in sig-nificant increase in [TEA]gl. CaR activation also significantlyreduced histamine-induced water secretion and apical K�
transport. Our data validate the strong link between histamine-stimulated acid secretion and water transport. We also showthat cAMP-dependent [K�]gl elevation prior to the onset of acidsecretion generates the osmotic gradient initially driving waterinto the gastric glands and thatCaRactivation inhibits this proc-ess, probably through reduction of intracellular cAMP levels.
Water transport is a process of vital importance for the phys-iology of the digestive system. Significant amounts of water are
secreted/absorbed every day in the GI tract, and the molecularmechanisms involved are well characterized for most segments(1). In the case of the stomach, however, although much inter-est has been centered on the ion transport mechanisms linkedto acid or alkaline secretion, the molecular mechanisms ofwater transport are not fully understood. Most of the informa-tion in this field derives from rather early reports where watertransport was measured either in amphibian or mammalianstomach by monitoring change in weight (2) or volume (3–5),by gravimetric (6) or radioactive methods (7, 8), or with fluo-rescent indicators in membrane vesicles (9).In general, it is believed that in the stomach, as in other organ
systems, water movement occurs secondarily to osmotic driv-ing forces created by active ion transport (6, 10) and/or tohydrostatic pressure differences (2, 11). Since gastric water flow(which can amount to up to 2 liters/day in humans (1, 12)) isproportional to acid secretion, it was previously assumed thatsecreted H� and Cl� might create the osmotic driving force forwater movement (6). Later, Berglindh et al. (13) proposed the“osmotic swelling” model in which the efflux of K� from theacid-secreting cells into the canaliculi and into the gland lumenwould create a gradient for the osmotic flow of water. To date,however, a clear correlation between the transport of specificions and gastric water movement has yet to be established.More importantly, the temporal dynamics of gastric water flowhave never been defined.Here we studied how the osmotic gradient driving water into
the lumen of the glands in the initial phases of acid secretion isgenerated using an approach that allows real time monitoringof fluid movement in single gastric glands. Water movementwas measured by recording changes in the concentration ofimpermeant ionswith ion-sensitivemicroelectrodes inserted inthe lumen of gastric glands in the intact perfused amphibianstomach. Our data show a direct correlation between timecourse of transport of specific ions, namely K�, and watermovement in the secretory state.In this study, we also investigated how the activation of an
unusual G-protein-coupled receptor, the extracellular Ca2�-sensing receptor (CaR),5 influences water secretion in thestomach. CaR was first described in the parathyroid gland (14),where it serves as a detector for changes in extracellular [Ca2�],
* The costs of publication of this article were defrayed in part by the paymentof page charges. This article must therefore be hereby marked “advertise-ment” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1 Supported by Cofin, MURST, FIRB Grant RBIN04PHZ7 (Rome, I) and by Finan-ziamenti di Ateneo (Bari, I).
2 Supported by a doctoral fellowship awarded jointly from the University ofBari and the European Community (FSE).
3 Supported by the Department of Veterans Affairs and the Brigham SurgicalGroup.
4 To whom correspondence should be addressed: Veterans Affairs BostonHealthcare System and the Dept. of Surgery, Harvard Medical School,Brigham and Women’s Hospital, 1400 VFW Pkwy., West Roxbury, MA02132. Tel.: 617-323-7700 (ext. 35902); Fax: 857-203-5592; E-mail:[email protected].
5 The abbreviations used are: CaR, Ca2�-sensing receptor; OCs, oxyntopepticcells.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 282, NO. 18, pp. 13477–13486, May 4, 2007Printed in the U.S.A.
MAY 4, 2007 • VOLUME 282 • NUMBER 18 JOURNAL OF BIOLOGICAL CHEMISTRY 13477
by guest on August 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from

amino acids, and other polyvalent cations. The receptor hassince been identified inmany other cell types, including epithe-lial cells of the alimentary tract (for reviews, see Refs. 15 and 16).The formation and secretion of gastric juice, which is com-
posed mostly of water, hydrochloric acid (145 mmol/liter inhumans), and pepsinogen, is under the control of hormonal andneural pathways and is essentially mediated by gastrin, hista-mine, and acetylcholine (17–21). In addition, we previouslyshowed that changes in the concentration of external free Ca2�
([Ca2�]ext), working via CaR, can influence the secretory func-tion of gastric mucosa (22). We found that stimulation withcarbachol, which mobilizes intracellular Ca2� in the oxynto-peptic cells (OCs), resulted in a substantial local increase in theextracellular [Ca2�] at the luminal face and a comparabledepletion at the serosal aspect of amphibian acid-secreting cells(23). The increase in [Ca2�] in the gastric gland lumen is due toactivation of the plasma membrane Ca2�-ATPase, which ishighly expressed at the apical membrane of these cells, where itco-localizes with CaR.In the amphibian gastric mucosa, carbachol is a well known
agonist of bicarbonate (24–26) and pepsinogen secretion (27).We previously found that changes in [Ca2�]ext secondary tocarbachol-induced increases in intracellular [Ca2�] were suffi-cient andnecessary to elicit both alkaline andpepsinogen secre-tion, independently of intracellular [Ca2�] changes (22).Recent reports show that CaR may also be involved in the
modulation of fluid secretion in different tissues. Lactatingmammary gland can “sense” extracellular Ca2� and adjustsmilk secretion as well as the secretion of parathyroid hormone-related protein and water in response to changes in [Ca2�]ext.This intricate set of responses defines a homeostatic systemthat helps to match milk production to the availability of Ca2�
(28). In the kidney, CaR regulates renal Ca2� excretion andinfluences the transepithelial movement of water and otherelectrolytes (29, 30). Finally, CaR, which is expressed along theentire gastrointestinal tract, seems to be involved in the modu-lation of fluid transport in the colon (31).We found here that stimulation of CaR results in consider-
able attenuation of gastric water secretion under resting condi-tions. This modulation becomes less important, however, dur-ing maximal rates of acid secretion.
MATERIALS AND METHODS
Tissue and Solutions—The experiments were performed ongastric fundus mucosa of Rana esculenta in accordance withthe Italian guidelines for animal experiments. Frogs were sacri-ficed by decapitation followed by destruction of the spinal cordand brain. The isolated mucosa was mounted horizontallybetween two halves of a top-open Lucite chamber (aperture 0.2cm2) with the serosal side facing up. The connective tissue layerwas further removed with sharpened watchmaker forcepsunder directmicroscopic observation in order to expose a num-ber of glands for impalement with microelectrodes. Both theserosal and mucosal surfaces were constantly superfused withoxygenated Ringer’s solution at room temperature. Fast fluidexchange in the chamber was achieved within seconds from ashock-free, electronically controlled eight-way manifold.
The control Ringer’s solution had the following composition:102.4 mmol/liter Na�, 4.0 mmol/liter K�, 1.4 mmol/liter Ca2�,0.8 mmol/liter Mg2�, 91.4 mmol/liter Cl�, 17.8 mmol/literHCO3
� and 11mmol/liter glucose. It was gassedwith 5%CO2 inO2 and had a pH of 7.36.Tissues were maintained in resting state by serosal addition
of cimetidine (a histamine H2 receptor blocker used to preventacid secretion) or stimulated by serosal perfusion with hista-mine. All chemicals were of reagent grade and purchased fromFarmitalia Carlo Erba (Milan, Italy), Sigma, Fluka Chemie AG(Buchs, Switzerland), Alexis Biochemicals (Lausen, Switzer-land), or SmithKline Beecham (Baranzate, Italy).Transepithelial and Extracellular Ion Measurements—The
transepithelial potential difference (Vt) was measured with amodel 610C high impedance differential electrometer(Keithley, Cleveland, OH) using two flowing boundary calomelhalf-cells filledwith 2.7mol/liter KCl solution and connected toeach bath solution downstream of the tissue. The serosal bathwas connected to ground. The lumen of the gastric gland waspunctured by lowering the ion-sensitive (TEA�, K�, H�, orCl�) double-barreled microelectrode, mounted on a Leitzmicromanipulator, perpendicular to the surface of an exposedgland under oblique (45°) observation through a stereomicro-scope at �50 magnification (Wild, Heerbrugg, Switzerland).The correct positioning of the microelectrode tip in the glandlumen was established by the following criteria: (i) the nearidentity of the glandular luminal potential (Vgl) with the tran-sepithelial potential (Vt); (ii) the near identity of the electricalresistance recorded between the microelectrode referencechannel and serosal bath macroelectrode with the transepithe-lial resistance. All measurements were performed with a modelFD 223 dual channel electrometer (World Precision Instru-ments, NewHaven, CT) and recorded on a strip chart recorder(Kipp & Zonen, Delft, The Netherlands).TEA�-sensitive Microelectrodes—Double-barreled TEA�-
sensitive microelectrodes were constructed as described previ-ously for pH-sensitive microelectrodes (24, 32). Briefly, twopieces of filament-containing aluminum silicate glass tubing ofdifferent diameter (1.5-mm outer diameter and 1.0-mm innerdiameter and 1.1-mmouter diameter and 0.75-mm inner diam-eter) (Hilgenberg, Malsfeld, Germany) were fixed in paralleland melted together by first twisting and then untwisting themat melting point before they were pulled in a PE2 vertical puller(Narishige, Tokyo, Japan). Then the back of the thin channelwas closed, and the thick channel was silanized for 180 s indimethyldichlorosilane vapor (Fluka Chemie AG) and baked inthe oven. The shank of the thick channel was back-filled with asmall amount of the ligand mixture (World Precision Instru-ments), and its shaft was later filled with a solution containing160 mmol/liter KCl and 1 mmol/liter TEA�.
For all microelectrodes, the reference channel contained 500mmol/literKCl, and anAg/AgClwirewas inserted. The averageslope and resistance of the electrodes were 57.5 � 0.4 mV perdecade change in [TEA�] (n � 26) and 180 � 4 gigaohms(selective channel) and 125� 3megaohms (reference channel).All microelectrodes were calibrated in the upper half-chamberbefore each puncture and, if the micropuncture was successful,also after the puncture by flushing the chamber with a HEPES-
Water Transport in the Gastric Gland Lumen
13478 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 18 • MAY 4, 2007
by guest on August 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from
buffered Ringer’s solution containing different TEA� concen-trations (0.8, 1.0, and 1.5 mmol/liter). Although ion-selectivemicroelectrodes respond to ionic activities and not to concen-trations, calibrating solutions and results are reported as con-centrations with the assumption that the activity coefficientdoes not change significantly during measurements (see Ref.33). The slope ofmicroelectrodes was not affected by pH (rang-ing between 4.0 and 8.0) or by spermine and Ca2�.pH-sensitiveMicroelectrode—The tip of the selective channel
was back-filled with H� ligand (Hydrogen Ionophore II, Mix-tureA/Mixture B; Fluka, Buchs, Switzerland) and the shaft witha HEPES-buffered Ringer’s solution with pH 7.0. Average slopeand resistance were 55.6 � 0.4 mV/pH unit (n � 23), 292 � 29gigaohms (selective channel), and 187 � 19 megaohms (refer-ence channel). All microelectrodes were calibrated usingHEPES-buffered Ringer’s solutions with pH values between 6.8and 7.8.K�-sensitive Microelectrode—The K� ligand (K�-selective
liquid ion exchanger, Mixture A; Fluka) was inserted in the tipof the selective channel, and the shaft was filled with a HEPES-buffered Ringer’s solution containing 4.0 mmol/liter K�. K�
microelectrodes were calibrated in HEPES-buffered Ringer’ssolutions containing 0.8, 4.0, and 10.0 mmol/liter K�. The sen-sitivity was 52.0 � 0.6 mV (n � 13) for a 10-fold change in K�
concentration.Cl�-sensitive Microelectrodes—The Cl� ligand (Cl�-selec-
tive ion exchanger, Mixture A; Fluka) was inserted in the tip ofthe selective channel while the shaft was filled with HEPES-buffered Ringer’s solution containing 91.4 mmol/liter Cl�. Cl�microelectrodes were calibrated in Ringer’s solutions contain-ing 50, 91.4, or 130 mmol/l Cl�. Average slope was 56.7 � 0.9mV (n � 4).Acid Secretion Measurements—Tissues were mounted verti-
cally between twohalves of a Lucite chamber having an exposedarea of 0.64 cm2. Each half-chamber consisted of a circular fluidcanal of 2.5-ml total volume filled with modified Ringer’s solu-tion that was constantly recirculated by means of a bubble lift.The control Ringer’s solution on the serosal side was gassedcontinually with 5% CO2 in O2 (pH 7.36).The luminal solution was unbuffered and had the following
composition: 102.4 mmol/liter Na�, 4.0 mmol/liter K�, 91.4mmol/liter Cl�, 15 mmol/liter isethionate, 7 mmol/liter man-nitol, and 11mmol/liter D-glucose. To prevent accumulation ofCO2, this solution was gassed with 100% O2 that was passedthrough a bottle containing Ba(OH)2 solution (50 mmol/liter).Acid secretion was measured with the pH-stat method (Radi-ometer, Copenhagen, Denmark). The titration procedure wasactivated every 10 min using 5 mmol/liter NaOH as the titrant,respectively. The transepithelial potential (Vt) was monitoredwith a voltmeter using two calomel half-cells connected to eachbath solution. HCl secretion was stimulated, adding 500 �mol/liter histamine to the serosal solution (no cimetidine).Data Analysis and Statistics—Mean values are expressed �
S.E. of n individual micropuncture recordings. The significanceof the observations was evaluated by Student’s t test for pairedor unpaired data as appropriate, and p � 0.05 denoted a statis-tical difference.
RESULTS
TEA� as a Tool to Measure Water Transport in Single GastricGlands
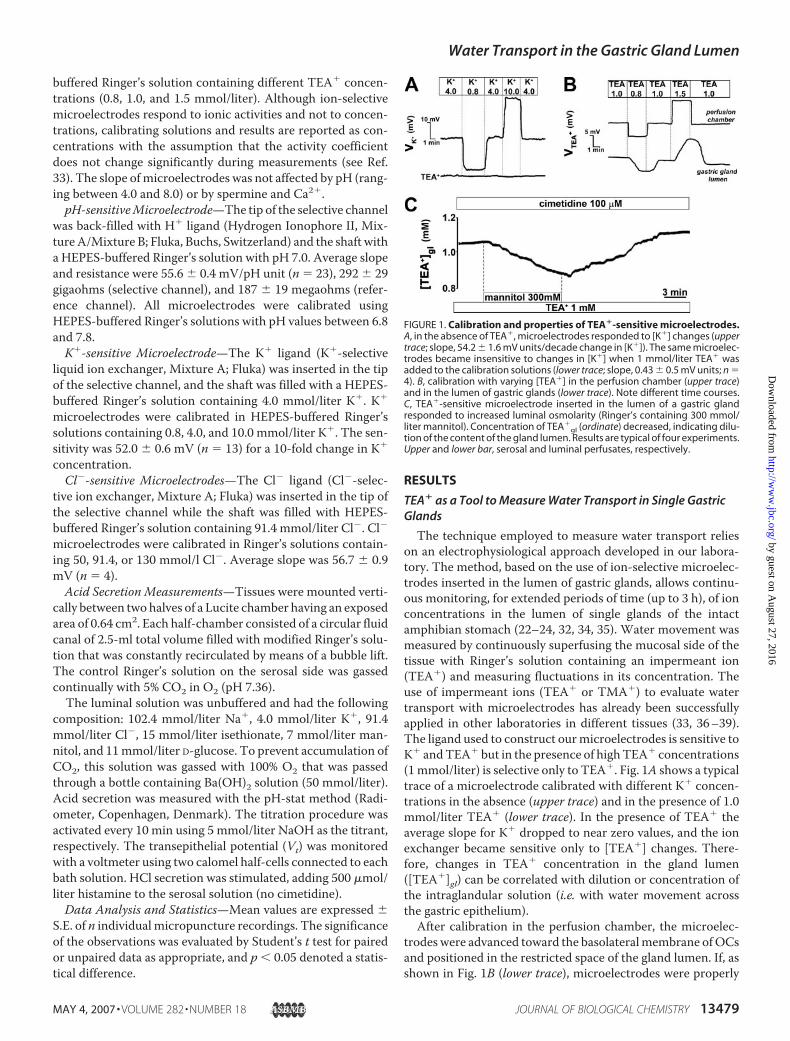
The technique employed to measure water transport relieson an electrophysiological approach developed in our labora-tory. The method, based on the use of ion-selective microelec-trodes inserted in the lumen of gastric glands, allows continu-ous monitoring, for extended periods of time (up to 3 h), of ionconcentrations in the lumen of single glands of the intactamphibian stomach (22–24, 32, 34, 35). Water movement wasmeasured by continuously superfusing the mucosal side of thetissue with Ringer’s solution containing an impermeant ion(TEA�) and measuring fluctuations in its concentration. Theuse of impermeant ions (TEA� or TMA�) to evaluate watertransport with microelectrodes has already been successfullyapplied in other laboratories in different tissues (33, 36–39).The ligand used to construct ourmicroelectrodes is sensitive toK� andTEA� but in the presence of highTEA� concentrations(1mmol/liter) is selective only to TEA�. Fig. 1A shows a typicaltrace of a microelectrode calibrated with different K� concen-trations in the absence (upper trace) and in the presence of 1.0mmol/liter TEA� (lower trace). In the presence of TEA� theaverage slope for K� dropped to near zero values, and the ionexchanger became sensitive only to [TEA�] changes. There-fore, changes in TEA� concentration in the gland lumen([TEA�]gl) can be correlated with dilution or concentration ofthe intraglandular solution (i.e. with water movement acrossthe gastric epithelium).After calibration in the perfusion chamber, the microelec-
trodeswere advanced toward the basolateralmembrane ofOCsand positioned in the restricted space of the gland lumen. If, asshown in Fig. 1B (lower trace), microelectrodes were properly
FIGURE 1. Calibration and properties of TEA�-sensitive microelectrodes.A, in the absence of TEA�, microelectrodes responded to [K�] changes (uppertrace; slope, 54.2 � 1.6 mV units/decade change in [K�]). The same microelec-trodes became insensitive to changes in [K�] when 1 mmol/liter TEA� wasadded to the calibration solutions (lower trace; slope, 0.43 � 0.5 mV units; n �4). B, calibration with varying [TEA�] in the perfusion chamber (upper trace)and in the lumen of gastric glands (lower trace). Note different time courses.C, TEA�-sensitive microelectrode inserted in the lumen of a gastric glandresponded to increased luminal osmolarity (Ringer’s containing 300 mmol/liter mannitol). Concentration of TEA�
gl (ordinate) decreased, indicating dilu-tion of the content of the gland lumen. Results are typical of four experiments.Upper and lower bar, serosal and luminal perfusates, respectively.
Water Transport in the Gastric Gland Lumen
MAY 4, 2007 • VOLUME 282 • NUMBER 18 JOURNAL OF BIOLOGICAL CHEMISTRY 13479
by guest on August 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from
positioned in the gland lumen (for details, see “Materials andMethods”) the time course of the response to changes in lumi-nal [TEA�] was much slower compared with when the tip wasin the luminal bath (time to peak was 141.0 � 11.8 s versus20.3� 0.9 s; n� 4, respectively). Average slopes of bath/gastricgland lumen were not significantly different. The slope ofmicroelectrodes was not affected by pH (ranging between 4.0and 8.0) or by spermine and Ca2�. Fig. 1C shows that when themicroelectrode tip was inserted in the lumen of a gastric gland,it was possible to record an immediate and reversible decreasein [TEA�]gl in response to increased osmolarity of the mucosalperfusate (300 mmol/liter mannitol), indicating entry of waterin the gland lumen.
Water Transport at Rest and after Stimulation: Correlationwith Acid Secretion
The H�/K�-ATPase located on the apical membrane of theacid-secreting cells exchanges H� for K�, and K� is recycledfrom the lumen into the cytoplasm via apical K� channels (40).These channels are necessary for the H�-K�-ATPase to func-tion, and in fact their inhibition results in extremely effectivereduction of acid secretion (41–45).TEA� is a known blocker of different types of K� channels
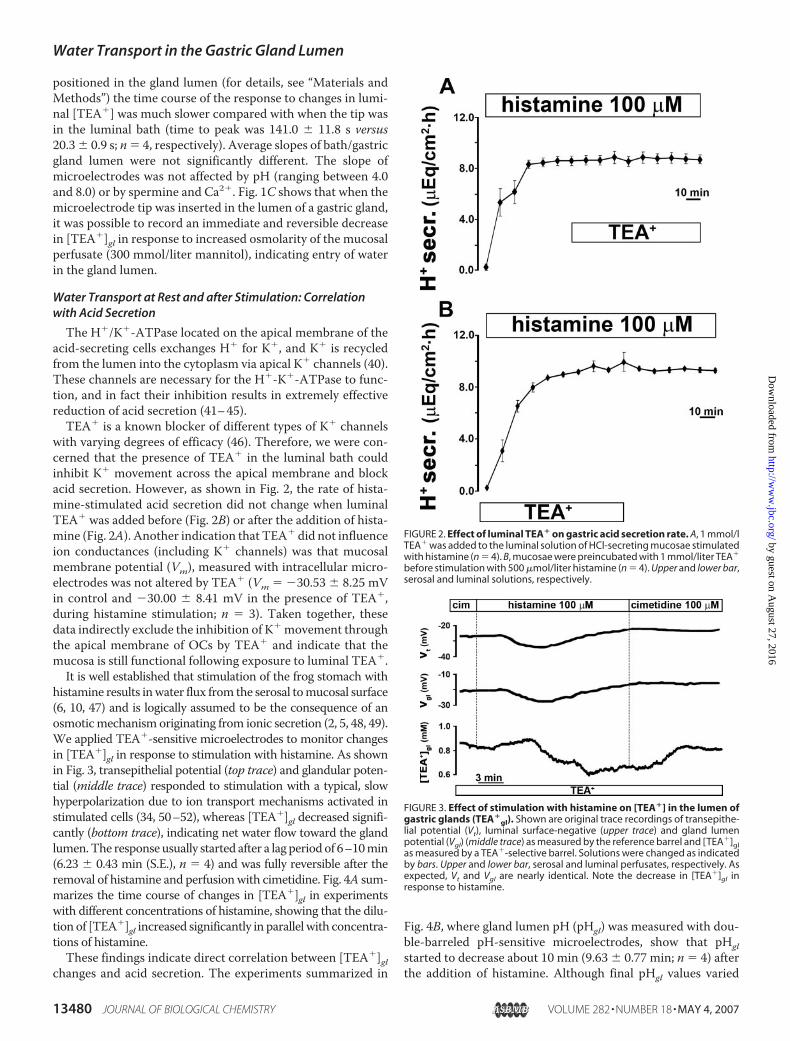
with varying degrees of efficacy (46). Therefore, we were con-cerned that the presence of TEA� in the luminal bath couldinhibit K� movement across the apical membrane and blockacid secretion. However, as shown in Fig. 2, the rate of hista-mine-stimulated acid secretion did not change when luminalTEA� was added before (Fig. 2B) or after the addition of hista-mine (Fig. 2A). Another indication that TEA� did not influenceion conductances (including K� channels) was that mucosalmembrane potential (Vm), measured with intracellular micro-electrodes was not altered by TEA� (Vm � �30.53 � 8.25 mVin control and �30.00 � 8.41 mV in the presence of TEA�,during histamine stimulation; n � 3). Taken together, thesedata indirectly exclude the inhibition of K� movement throughthe apical membrane of OCs by TEA� and indicate that themucosa is still functional following exposure to luminal TEA�.It is well established that stimulation of the frog stomach with
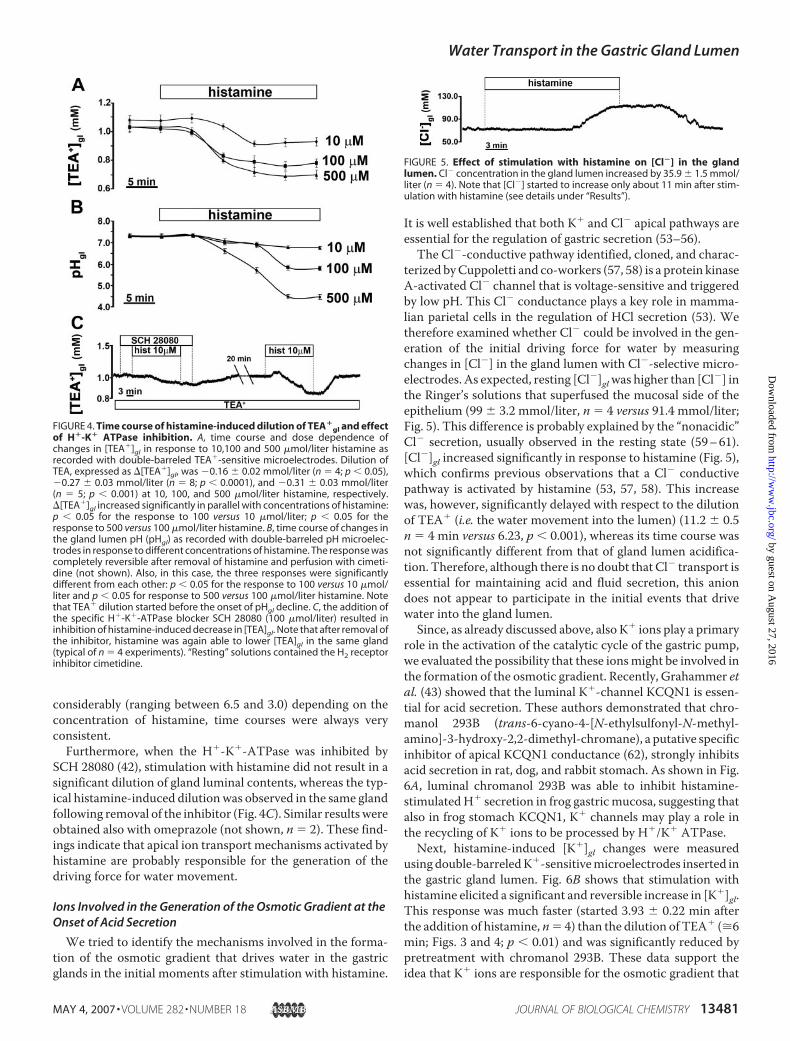
histamine results inwater flux from the serosal tomucosal surface(6, 10, 47) and is logically assumed to be the consequence of anosmoticmechanism originating from ionic secretion (2, 5, 48, 49).We applied TEA�-sensitive microelectrodes to monitor changesin [TEA�]gl in response to stimulation with histamine. As shownin Fig. 3, transepithelial potential (top trace) and glandular poten-tial (middle trace) responded to stimulation with a typical, slowhyperpolarization due to ion transport mechanisms activated instimulated cells (34, 50–52), whereas [TEA�]gl decreased signifi-cantly (bottom trace), indicating net water flow toward the glandlumen.The response usually started after a lag periodof 6–10min(6.23 � 0.43 min (S.E.), n � 4) and was fully reversible after theremoval of histamine and perfusionwith cimetidine. Fig. 4A sum-marizes the time course of changes in [TEA�]gl in experimentswith different concentrations of histamine, showing that the dilu-tion of [TEA�]gl increased significantly in parallel with concentra-tions of histamine.These findings indicate direct correlation between [TEA�]gl
changes and acid secretion. The experiments summarized in
Fig. 4B, where gland lumen pH (pHgl) was measured with dou-ble-barreled pH-sensitive microelectrodes, show that pHglstarted to decrease about 10 min (9.63 � 0.77 min; n � 4) afterthe addition of histamine. Although final pHgl values varied
FIGURE 2. Effect of luminal TEA� on gastric acid secretion rate. A, 1 mmol/lTEA� was added to the luminal solution of HCl-secreting mucosae stimulatedwith histamine (n � 4). B, mucosae were preincubated with 1 mmol/liter TEA�
before stimulation with 500 �mol/liter histamine (n � 4). Upper and lower bar,serosal and luminal solutions, respectively.
FIGURE 3. Effect of stimulation with histamine on [TEA�] in the lumen ofgastric glands (TEA�
gl). Shown are original trace recordings of transepithe-lial potential (Vt), luminal surface-negative (upper trace) and gland lumenpotential (Vgl) (middle trace) as measured by the reference barrel and [TEA�]glas measured by a TEA�-selective barrel. Solutions were changed as indicatedby bars. Upper and lower bar, serosal and luminal perfusates, respectively. Asexpected, Vt and Vgl are nearly identical. Note the decrease in [TEA�]gl inresponse to histamine.
Water Transport in the Gastric Gland Lumen
13480 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 18 • MAY 4, 2007
by guest on August 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from
considerably (ranging between 6.5 and 3.0) depending on theconcentration of histamine, time courses were always veryconsistent.Furthermore, when the H�-K�-ATPase was inhibited by
SCH 28080 (42), stimulation with histamine did not result in asignificant dilution of gland luminal contents, whereas the typ-ical histamine-induced dilutionwas observed in the same glandfollowing removal of the inhibitor (Fig. 4C). Similar resultswereobtained also with omeprazole (not shown, n � 2). These find-ings indicate that apical ion transport mechanisms activated byhistamine are probably responsible for the generation of thedriving force for water movement.
Ions Involved in the Generation of the Osmotic Gradient at theOnset of Acid Secretion
We tried to identify the mechanisms involved in the forma-tion of the osmotic gradient that drives water in the gastricglands in the initial moments after stimulation with histamine.
It is well established that both K� and Cl� apical pathways areessential for the regulation of gastric secretion (53–56).The Cl�-conductive pathway identified, cloned, and charac-
terized byCuppoletti and co-workers (57, 58) is a protein kinaseA-activated Cl� channel that is voltage-sensitive and triggeredby low pH. This Cl� conductance plays a key role in mamma-lian parietal cells in the regulation of HCl secretion (53). Wetherefore examined whether Cl� could be involved in the gen-eration of the initial driving force for water by measuringchanges in [Cl�] in the gland lumen with Cl�-selective micro-electrodes. As expected, resting [Cl�]glwas higher than [Cl�] inthe Ringer’s solutions that superfused the mucosal side of theepithelium (99 � 3.2 mmol/liter, n � 4 versus 91.4 mmol/liter;Fig. 5). This difference is probably explained by the “nonacidic”Cl� secretion, usually observed in the resting state (59–61).[Cl�]gl increased significantly in response to histamine (Fig. 5),which confirms previous observations that a Cl� conductivepathway is activated by histamine (53, 57, 58). This increasewas, however, significantly delayed with respect to the dilutionof TEA� (i.e. the water movement into the lumen) (11.2 � 0.5n � 4 min versus 6.23, p � 0.001), whereas its time course wasnot significantly different from that of gland lumen acidifica-tion. Therefore, although there is no doubt that Cl� transport isessential for maintaining acid and fluid secretion, this aniondoes not appear to participate in the initial events that drivewater into the gland lumen.Since, as already discussed above, also K� ions play a primary
role in the activation of the catalytic cycle of the gastric pump,we evaluated the possibility that these ionsmight be involved inthe formation of the osmotic gradient. Recently, Grahammer etal. (43) showed that the luminal K�-channel KCQN1 is essen-tial for acid secretion. These authors demonstrated that chro-manol 293B (trans-6-cyano-4-[N-ethylsulfonyl-N-methyl-amino]-3-hydroxy-2,2-dimethyl-chromane), a putative specificinhibitor of apical KCQN1 conductance (62), strongly inhibitsacid secretion in rat, dog, and rabbit stomach. As shown in Fig.6A, luminal chromanol 293B was able to inhibit histamine-stimulatedH� secretion in frog gastricmucosa, suggesting thatalso in frog stomach KCQN1, K� channels may play a role inthe recycling of K� ions to be processed by H�/K� ATPase.
Next, histamine-induced [K�]gl changes were measuredusing double-barreledK�-sensitivemicroelectrodes inserted inthe gastric gland lumen. Fig. 6B shows that stimulation withhistamine elicited a significant and reversible increase in [K�]gl.This response was much faster (started 3.93 � 0.22 min afterthe addition of histamine, n� 4) than the dilution of TEA� (�6min; Figs. 3 and 4; p � 0.01) and was significantly reduced bypretreatment with chromanol 293B. These data support theidea that K� ions are responsible for the osmotic gradient that
FIGURE 4. Time course of histamine-induced dilution of TEA�gl and effect
of H�-K� ATPase inhibition. A, time course and dose dependence ofchanges in [TEA�]gl in response to 10,100 and 500 �mol/liter histamine asrecorded with double-barreled TEA�-sensitive microelectrodes. Dilution ofTEA, expressed as �[TEA�]gl, was �0.16 � 0.02 mmol/liter (n � 4; p � 0.05),�0.27 � 0.03 mmol/liter (n � 8; p � 0.0001), and �0.31 � 0.03 mmol/liter(n � 5; p � 0.001) at 10, 100, and 500 �mol/liter histamine, respectively.�[TEA�]gl increased significantly in parallel with concentrations of histamine:p � 0.05 for the response to 100 versus 10 �mol/liter; p � 0.05 for theresponse to 500 versus 100 �mol/liter histamine. B, time course of changes inthe gland lumen pH (pHgl) as recorded with double-barreled pH microelec-trodes in response to different concentrations of histamine. The response wascompletely reversible after removal of histamine and perfusion with cimeti-dine (not shown). Also, in this case, the three responses were significantlydifferent from each other: p � 0.05 for the response to 100 versus 10 �mol/liter and p � 0.05 for response to 500 versus 100 �mol/liter histamine. Notethat TEA� dilution started before the onset of pHgl decline. C, the addition ofthe specific H�-K�-ATPase blocker SCH 28080 (100 �mol/liter) resulted ininhibition of histamine-induced decrease in [TEA]gl. Note that after removal ofthe inhibitor, histamine was again able to lower [TEA]gl in the same gland(typical of n � 4 experiments). “Resting” solutions contained the H2 receptorinhibitor cimetidine.
FIGURE 5. Effect of stimulation with histamine on [Cl�] in the glandlumen. Cl� concentration in the gland lumen increased by 35.9 � 1.5 mmol/liter (n � 4). Note that [Cl�] started to increase only about 11 min after stim-ulation with histamine (see details under “Results”).
Water Transport in the Gastric Gland Lumen
MAY 4, 2007 • VOLUME 282 • NUMBER 18 JOURNAL OF BIOLOGICAL CHEMISTRY 13481
by guest on August 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from

drives water in the gastric gland lumen at the onset of gastricsecretion. Control experiments showed that pretreatment withchromanol 293B effectively inhibited histamine-induced dilu-tion of [TEA�] in the gland lumen (Fig. 6C).
Finally, we also tested if, by analogy with the mammalianstomach (63), the Na�-K�-2Cl� cotransporter-1 might beinvolved in the regulation of fluid secretion. Experiments per-formed with the Na�-K�-2Cl� cotransporter-1 inhibitorbumetanide (100 �mol/liter) indicate that Na�-K�-2Cl�cotransporter-1 does not appear to be involved in the regula-tion of fluid, K�, or acid secretion in our model (data notshown).
Modulation of Water Transport by CaR
At Rest and after Stimulation—The extracellular CaR, whichis expressed along the entire gastrointestinal tract (31, 64),belongs to group C of the G protein-coupled receptor super-family and has been shown to modulate many of the knowneffects of extracellular Ca2� in a variety of tissues (15). Werecently found that CaR is located in the apical membrane ofOCs (23) and is involved in the modulation of alkali and pep-sinogen secretion (22). Since it has been shown that this recep-tor can influence fluid transport in different tissues (28, 30, 31),we examined its role in the modulation of water movement inthe gastric epithelium. Stimulation of CaRwith either spermine(a CaR agonist present endogenously in the gastrointestinaltract (65)) or elevation of luminal [Ca2�] to 2.0 mmol/liter (a“physiological” concentration that is able to activate the recep-tor (22, 66)) resulted in a significant and reversible increase in
[TEA�]gl (Fig. 7A). Fig. 7B summarizes the data obtained undervarious conditions. Besides spermine and high luminal [Ca2�],we also stimulated CaR with carbachol, which is able to indi-rectly stimulate CaR due to its ability to induce asymmetricalchanges in external [Ca2�] (i.e. elevating luminal [Ca2�] from1.4 to 2.0 mmol/liter while decreasing serosal [Ca2�] from 1.4to 1.0 mmol/liter) (23). Fig. 7B shows that carbachol elicited anincrease in [TEA�]gl. and that mimicking carbachol-inducedasymmetric fluctuations in extracellular [Ca2�] also resulted inan increase in [TEA�]gl, similar to that observed in response toother agonists. The latter response was entirely due to theaction of high luminal [Ca2�], since lowering basolateral [Ca2�]alone did not cause any significant change in [TEA�]gl (Fig. 7B).Thus, in nonstimulated frog gastric mucosa, activation of CaRresulted in a significant reduction of water in the gland lumen.Since water transport and gastric acid secretion are closely
correlated in frog stomach (Figs. 3 and 4), we investigated theaction of CaR on histamine-induced water transport (Fig. 8A).Stimulation with low histamine concentrations resulted in adilution of glandular solution that was not affected by the addi-tion of spermine in the luminal perfusate, whereas sperminestill elicited an increase in [TEA�]gl before and after histaminein the same gland. However, pretreatment with spermine elic-ited the typical increase in [TEA�]gl and significantly reducedhistamine-induced water secretion (Fig. 8B).Since we have previously demonstrated that in frog gastric
mucosa CaR is coupled with the classical pertussis toxin-sensi-tive G�i (22), it is likely that the effect of CaR on water move-ment is the result of decreased intracellular cAMP levels. Sincegastric acid secretion is a cAMP-dependent process (18, 67, 68),one could predict that activation of CaR might result in inhibi-tion of gastric acid secretion. To test this possibility, we per-
FIGURE 6. Effect of chromanol 293B on acid secretion, K�, and watertransport. A, luminal chromanol 293B (20 �mol/liter) significantly inhibitedhistamine-induced proton secretion (from 4.06 � 0.22 to 0.85 � 0.30 �eq/cm2�h, n � 4, p � 0.01) as measured with the pH-stat method. B, histamine ledto a reversible increase in [K�] in the gland lumen ([K�]gl]) as measured withdouble-barreled K�-sensitive microelectrodes. This increase was inhibited bypretreatment with chromanol 293B in the same gland (by 74.28 � 5.15%; n �4; p � 0.05). C, luminal chromanol 293B significantly inhibited also histamine-induced dilution of TEA�
gl as measured with TEA�-sensitive microelectrodes(typical of six experiments). Note that in the absence of chromanol 293B,histamine was able to dilute TEA� in the same gland (typical of four experi-ments). Vt and Vgl have been omitted for simplicity in B and C. “Resting” solu-tions contained the H2 receptor inhibitor cimetidine.
FIGURE 7. Effect of stimulation of CaR on [TEA�]gl in resting state. A, stim-ulation of CaR with luminal spermine (2 mmol/liter) or elevation of luminalCa2� (from 1.4 to 2 mmol/liter) led to concentration of TEA�
gl. Vt and Vgl havebeen omitted for simplicity. B, summary of changes in [TEA�]gl (�[TEA�]gl)after the following treatments: luminal spermine, reduction in serosalCa2� (from 1.4 to 1 mmol/liter), elevation of luminal Ca2�, simultaneousdecrease of serosal (to 1 mmol/liter) and increase of luminal Ca2� (to 2 mmol/liter), or stimulation with carbachol. The H2 receptor inhibitor cimetidine wasalways present in all solutions.
Water Transport in the Gastric Gland Lumen
13482 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 18 • MAY 4, 2007
by guest on August 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from
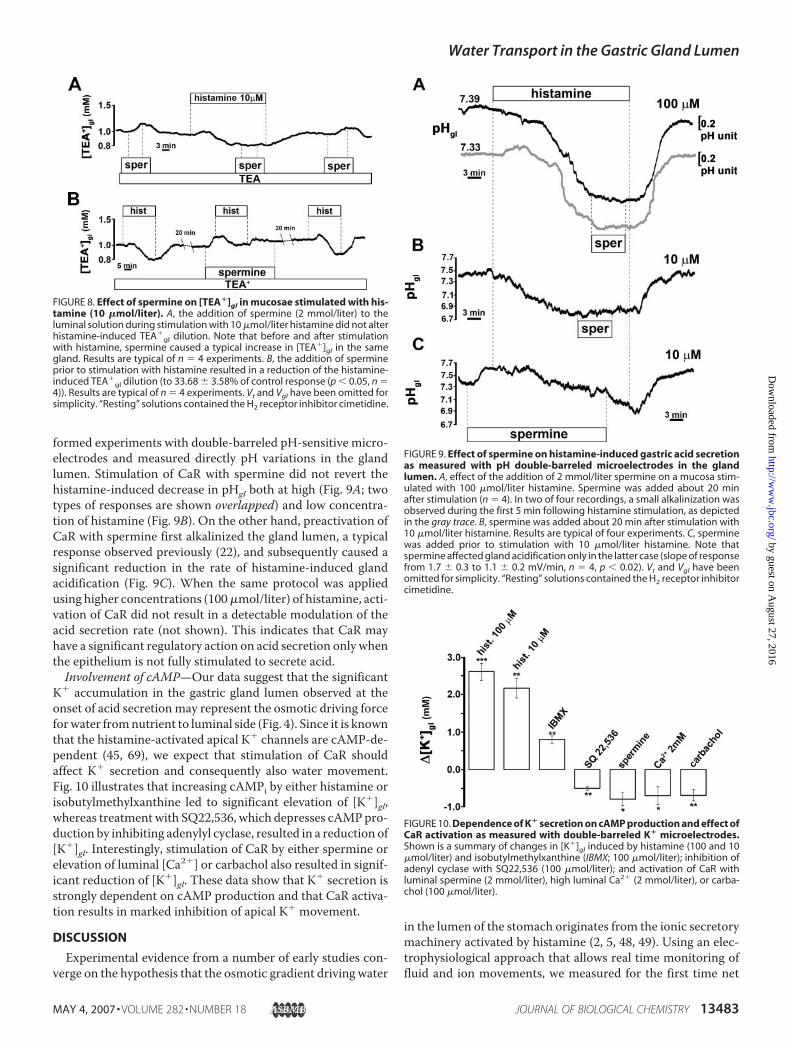
formed experiments with double-barreled pH-sensitive micro-electrodes and measured directly pH variations in the glandlumen. Stimulation of CaR with spermine did not revert thehistamine-induced decrease in pHgl both at high (Fig. 9A; twotypes of responses are shown overlapped) and low concentra-tion of histamine (Fig. 9B). On the other hand, preactivation ofCaR with spermine first alkalinized the gland lumen, a typicalresponse observed previously (22), and subsequently caused asignificant reduction in the rate of histamine-induced glandacidification (Fig. 9C). When the same protocol was appliedusing higher concentrations (100�mol/liter) of histamine, acti-vation of CaR did not result in a detectable modulation of theacid secretion rate (not shown). This indicates that CaR mayhave a significant regulatory action on acid secretion only whenthe epithelium is not fully stimulated to secrete acid.Involvement of cAMP—Our data suggest that the significant
K� accumulation in the gastric gland lumen observed at theonset of acid secretion may represent the osmotic driving forceforwater fromnutrient to luminal side (Fig. 4). Since it is knownthat the histamine-activated apical K� channels are cAMP-de-pendent (45, 69), we expect that stimulation of CaR shouldaffect K� secretion and consequently also water movement.Fig. 10 illustrates that increasing cAMPi by either histamine orisobutylmethylxanthine led to significant elevation of [K�]gl,whereas treatmentwith SQ22,536, which depresses cAMPpro-duction by inhibiting adenylyl cyclase, resulted in a reduction of[K�]gl. Interestingly, stimulation of CaR by either spermine orelevation of luminal [Ca2�] or carbachol also resulted in signif-icant reduction of [K�]gl. These data show that K� secretion isstrongly dependent on cAMP production and that CaR activa-tion results in marked inhibition of apical K� movement.
DISCUSSION
Experimental evidence from a number of early studies con-verge on the hypothesis that the osmotic gradient driving water
in the lumen of the stomach originates from the ionic secretorymachinery activated by histamine (2, 5, 48, 49). Using an elec-trophysiological approach that allows real time monitoring offluid and ion movements, we measured for the first time net
FIGURE 8. Effect of spermine on [TEA�]gl in mucosae stimulated with his-tamine (10 �mol/liter). A, the addition of spermine (2 mmol/liter) to theluminal solution during stimulation with 10 �mol/liter histamine did not alterhistamine-induced TEA�
gl dilution. Note that before and after stimulationwith histamine, spermine caused a typical increase in [TEA�]gl in the samegland. Results are typical of n � 4 experiments. B, the addition of spermineprior to stimulation with histamine resulted in a reduction of the histamine-induced TEA�
gl dilution (to 33.68 � 3.58% of control response (p � 0.05, n �4)). Results are typical of n � 4 experiments. Vt and Vgl have been omitted forsimplicity. “Resting” solutions contained the H2 receptor inhibitor cimetidine.
FIGURE 9. Effect of spermine on histamine-induced gastric acid secretionas measured with pH double-barreled microelectrodes in the glandlumen. A, effect of the addition of 2 mmol/liter spermine on a mucosa stim-ulated with 100 �mol/liter histamine. Spermine was added about 20 minafter stimulation (n � 4). In two of four recordings, a small alkalinization wasobserved during the first 5 min following histamine stimulation, as depictedin the gray trace. B, spermine was added about 20 min after stimulation with10 �mol/liter histamine. Results are typical of four experiments. C, sperminewas added prior to stimulation with 10 �mol/liter histamine. Note thatspermine affected gland acidification only in the latter case (slope of responsefrom 1.7 � 0.3 to 1.1 � 0.2 mV/min, n � 4, p � 0.02). Vt and Vgl have beenomitted for simplicity. “Resting” solutions contained the H2 receptor inhibitorcimetidine.
FIGURE 10. Dependence of K� secretion on cAMP production and effect ofCaR activation as measured with double-barreled K� microelectrodes.Shown is a summary of changes in [K�]gl induced by histamine (100 and 10�mol/liter) and isobutylmethylxanthine (IBMX; 100 �mol/liter); inhibition ofadenyl cyclase with SQ22,536 (100 �mol/liter); and activation of CaR withluminal spermine (2 mmol/liter), high luminal Ca2� (2 mmol/liter), or carba-chol (100 �mol/liter).
Water Transport in the Gastric Gland Lumen
MAY 4, 2007 • VOLUME 282 • NUMBER 18 JOURNAL OF BIOLOGICAL CHEMISTRY 13483
by guest on August 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from

water flux within the lumen of gastric glands of the intactamphibian stomach, providing direct evidence for the stronglink between histamine-induced net water flow and acidsecretion.We showed that histamine-induced water flux precedes, to a
great extent (�3min), the onset of acid secretion. This time lagcannot be explained by different response times of the two dif-ferent types ofmicroelectrodes/resins employed (i.e.TEA� andpHmicroelectrodes) to measure water flow and acid secretion.The response time of the two microelectrodes was in fact vir-tually the same: 20.34 � 0.35 s (n � 35) for TEA� and 21.07 �0.38 s (n � 20) for pHmicroelectrodes. These findings indicatethat the osmotic gradient driving water in the gland lumen hasto originate from ion movements taking place just before theinitiation of acid secretion, which rules out H�. Other possiblecandidates are Cl� and K�. Stimulation of the oxyntopepticcells to secrete acid results in an extensive rearrangement of theapical membrane consisting in the fusion of the cytoplasmictubulovesicular compartment with the apical membrane andconsequent translocation of the H�/K� ATPase, in parallelwith K�- and Cl�-conductive pathways (53, 55, 57, 70–72).Both ions are essential for maintenance of acid secretion (13,54, 70, 72).Chloride ions are involved in the formation of the osmotic
gradient in a large number of tissues, from salivary glands (73)to choroid plexus epithelium (74) to cholangiocytes (75).Recently, parchorin, a novel protein with significant homologyto the family of chloride intracellular channels, has been clonedand characterized in the apical membrane of rabbit parietalcells (76). Parchorin is known to be involved in the formation ofion gradients for water movement in various tissues, such asmammary glands, pancreas, prostate, testis, and kidney (77).Although evidence based on cellular distribution and changesin expression of this protein are suggestive of a role for par-chorin in Cl� and water transport in the stomach, functionaldata are still lacking.Logdson and Machen (78) found that Cl� is not required
either for the initiation of H� secretion or for the maintenanceof stimulated morphology of frog stomach. Villegas andSananes (79) have shown that replacement of chloride by sul-fate in the solutions bathing frog stomach does not affect watertransport, whereas it inhibits histamine-induced acid secretion.The fact that the histamine-induced increase in [Cl�]gl is
significantly delayed with respect to water secretion favors theidea that Cl� is not the initiator of the osmotic driving force. Itis likely, however, that Cl� is indispensable for the potentiationof the gradient during later stages of the secretory response,when basolateral and apical influx and efflux mechanisms arefully activated.In contrast, a number of observations in this study show that
movement of K� from oxyntopeptic cells to the gland lumen isnecessary for initiation of water secretion in frog stomach. Wefound that the first event recorded following stimulation withhistamine is the increase in [K�]gl (about 4 min after stimula-tion; Fig. 5). Our data are consistent with the observation byLogdson and Machen (78) that removal of K� from solutionsresults in an occluded morphology of the gland lumen in stim-ulated gastric glands, indicating a drastic reduction in intrag-
landular hydrostatic pressure. The fact that chromanol 293B, ablocker of KCQN1 channels, strongly inhibits the histamine-induced increase in [K�]gl (Fig. 6) and the dilution of the glandlumen further supports the idea that K� is the promoter of theosmotic gradient driving water in the gland lumen, at least atthe onset of gastric secretion. To date, three different K� chan-nels (KCNQ1, Kir2.1, and Kir4.1), identified in the apical mem-brane of parietal cells (43–45, 80, 81) are potential candidatesfor apical recycling of K.These data lead to the identification of a specific sequence of
ion transport events activated in response to histamine. First,elevation in intracellular cAMP results in opening of apical K�
channels (45) and consequent exit of K� into the gland lumen.As to the counterion involved in this very first phase, we suggestthat a possible candidate might be HCO3
�. This idea derivesfrom our observations both in this (Fig. 9A, gray plot) and inprevious studies (24) that in the 6–10 min before pHgl starts todecline, a transient and small but significant alkalinization ofthe gland lumen was measured in 50% of the gland lumina.Second, as a result of K� andHCO3
� efflux, an osmotic gradientsufficient to drive water into the gland lumen is created. Third,the presence of K� in the gland lumen activates the H�/K�
ATPase, and H� moves into the lumen accompanied by Cl�ions. The resulting decrease in pHgl activates pH-sensitive api-cal Cl� channels (57), which further increases [Cl�]gl andresults in potentiation of the osmotic gradient. We can specu-late that at this stage, water transport becomes fully activated,and the hydrostatic pressure rises to push the content of thegland lumen into the greater lumen of the stomach. Watersecretionmay then account for the observed formation of chan-nels of fluid at the opening of gastric pits (82), ultimately facil-itating the flow of acid and pepsin through the mucus layercoating the luminal surface of the stomach.We have recently demonstrated that activation of the extra-
cellular CaR in the “resting” stomach (in the presence of H2receptor inhibitors) elicits alkaline and pepsinogen secretion infrog stomach from OCs (22). We show here that in the restingstate, CaR activation leads to reduction in gland lumen watercontent. A reduction of water fluxwould be functionally appro-priate for theweak alkaline secretion elicited byCaR to be effec-tive. An alkalinization of the gland lumenmight be important toprevent premature activation of pepsinogen to pepsin (knownto be favored by acidic pH (83)), which may damage the oxyn-topeptic cells. Since CaR is functionally coupled to a pertussistoxin-sensitive G�i in the OCs (22), activation of the receptorshould decrease the availability of cellular cAMP. Our datashow that activation of CaR in the resting state gives rise to atransient reduction of [K�]gl similar to that elicited bySQ22,536, awell known inhibitor of cAMPproduction, indicat-ing that the K� conductance involved is cAMP-dependent (seealso Fig. 10). These findings imply that there is a basal rate ofcAMP-dependentK� extrusion toward the gland lumen,whichin turn is responsible for basal water movement under restingconditions.The scenario is different when mucosae are stimulated with
histamine. After the acid-secretory mechanism is fully opera-tional, activation of the pertussis toxin-sensitive G�i-coupledpathway by CaR is probably insufficient to affect the physiolog-
Water Transport in the Gastric Gland Lumen
13484 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 18 • MAY 4, 2007
by guest on August 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from

ical progression of water and acid secretion (Figs. 8A and 9, Aand B). On the other hand, when intracellular cAMP levels arenot too high (i.e. at low histamine concentrations or duringbasal turnover of adenylyl cyclase), preincubation with CaRagonists and the consequent engagement of the G�i pathwaymay be sufficient to counteract the stimulatory action of cAMPon water and acid secretion (Figs. 8B and 9C). Our data reveal asubtle equilibrium between the stimulatory activity of G�s andantagonism by G�i; both serve to influence intracellular cAMPlevels and thereby fine tune the secretion of water and acidunder resting conditions.
Acknowledgments—We thank Lorenzo Iacovone, Mirangela Calder-aro, and Teresa Cuna for excellent assistance during experiments.
REFERENCES1. Masyuk, A. I., Marinelli, R. A., and LaRusso, N. F. (2002)Gastroenterology
122, 545–5622. Moody, F. G., and Durbin, R. P. (1969) Am. J. Physiol. 217, 255–2613. Villegas, L., and Sananes, L. (1974) Am. J. Physiol. 226, 1338–13434. Villegas, L., Sananes, L., and Michelangeli, F. (1971) Am. J. Physiol. 220,
1282–12885. Rehm,W. S., Schlesinger, H., andDennis,W.H. (1953)Am. J. Physiol. 175,
473–4866. Durbin, R. P. (1979) Am. J. Physiol. 236, E63–E697. Villegas, L. (1982) Biochim. Biophys. Acta. 685, 249–2528. Wang, K. S., Komar, A. R.,Ma, T., Filiz, F.,McLeroy, J., Hoda, K., Verkman,
A. S., and Bastidas, J. A. (2000) Am. J. Physiol. 279, G448–G4539. Priver, N. A., Rabon, E. C., and Zeidel, M. L. (1993) Biochemistry 32,
2459–246810. Rehm,W. S., Butler, C. F., Spangler, S. G., and Sanders, S. S. (1970) J. Theor.
Biol. 27, 433–45311. Finkelstein, A. (1987) in Distinguished Lectures Series of the Society of
General Physiologists, Vol. 4, pp. 1–228, John Wiley & Sons, Inc., NewYork
12. Zhang, E. B. (1996) in Gastrointestinal, Hepatobiliary, and NutritionalPhysiology (Zhang, E. B., Sitrin, M. D., and Black, D. B., eds) pp. 91–118,Lippincott-Raven, Philadelphia
13. Berglindh, T., Dibona, D. R., Ito, S., and Sachs, G. (1980) Am. J. Physiol.238, G165–G176
14. Brown, E.M., Gamba, G., Riccardi, D., Lombardi,M., Butters, R., Kifor, O.,Sun, A., Hediger, M. A., Lytton, J., and Hebert, S. C. (1993) Nature 366,575–580
15. Hofer, A.M., and Brown, E.M. (2003)Nat. Rev.Mol. Cell. Biol. 4, 530–53816. Hofer, A. M. (2005) J. Cell Sci. 118, 855–86217. Hersey, S. J., and Sachs, G. (1995) Physiol. Rev. 75, 155–18918. Yao, X., and Forte, J. G. (2003) Annu. Rev. Physiol. 65, 103–13119. Schubert, M. L. (2005) Curr. Opin. Gastroenterol. 21, 636–64320. Lloyd, K. C., and Soll, A. H. (1994) J. Gastroenterol. 29, Suppl. 7, 77–7921. Sachs, G., Zeng, N., and Prinz, C. (1997) Annu. Rev. Physiol. 59, 243–25622. Caroppo, R., Gerbino, A., Fistetto, G., Colella, M., Debellis, L., Hofer,
A. M., and Curci, S. (2004) J. Cell Biol. 166, 111–11923. Caroppo, R., Gerbino, A., Debellis, L., Kifor, O., Soybel, D. I., Brown, E.M.,
Hofer, A. M., and Curci, S. (2001) EMBO J. 20, 6316–632624. Debellis, L., Caroppo, R., Fromter, E., and Curci, S. (1998) J. Physiol.
(Lond.) 513, 235–24125. Flemstrom, G., and Garner, A. (1982) Am. J. Physiol. 242, G183–G19326. Flemstrom, G. (1977) Am. J. Physiol. 233, E1–E1227. Ruiz, M. C., Acosta, A., Abad, M. J., and Michelangeli, F. (1993) Am. J.
Physiol. 265, G934–G94128. VanHouten, J., Dann, P.,McGeoch, G., Brown, E.M., Krapcho, K., Neville,
M., and Wysolmerski, J. J. (2004) J. Clin. Invest. 113, 598–60829. Chattopadhyay, N., Baum, M., Bai, M., Riccardi, D., Hebert, S. C., Harris,
H. W., and Brown, E. M. (1996) Am. J. Physiol. 271, F736–F743
30. Brown, E. M. (2000) Annu. Rev. Nutr. 20, 507–53331. Hebert, S. C., Cheng, S., and Geibel, J. (2004) Cell Calcium 35, 239–24732. Debellis, L., Iacovelli, C., Fromter, E., and Curci, S. (1994) Pflugers Arch.
428, 655–66333. Nicholson, C. (1993) J. Neurosci. Methods 48, 199–21334. Debellis, L., Curci, S., and Fromter, E. (1992) Pflugers Arch. 422, 253–25935. Debellis, L., Papini, E., Caroppo, R., Montecucco, C., and Curci, S. (2001)
Am. J. Physiol. 281, G1440–G144836. Dietzel, I., Heinemann, U., Hofmeier, G., and Lux, H. D. (1980) Exp. Brain
Res. 40, 432–43937. Nicholson, C., and Phillips, J. M. (1981) J. Physiol. (Lond.) 321, 225–25738. Orkand, R. K., Dietzel, I., and Coles, J. A. (1984) Neurosci. Lett. 45,
273–27839. Chebabo, S. R., Hester, M. A., Jing, J., Aitken, P. G., and Somjen, G. G.
(1995) J. Physiol. (Lond.) 487, 685–69740. Sachs, G., Chang, H. H., Rabon, E., Schackman, R., Lewin, M., and Sacco-
mani, G. (1976) J. Biol. Chem. 251, 7690–769841. Wallmark, B., Larsson, H., and Humble, L. (1985) J. Biol. Chem. 260,
13681–1368442. Wallmark, B., Briving, C., Fryklund, J., Munson, K., Jackson, R., Mendlein,
J., Rabon, E., and Sachs, G. (1987) J. Biol. Chem. 262, 2077–208443. Grahammer, F., Herling, A.W., Lang, H. J., Schmitt-Graff, A.,Wittekindt,
O. H., Nitschke, R., Bleich, M., Barhanin, J., and Warth, R. (2001) Gastro-enterology 120, 1363–1371
44. Fujita, A., Horio, Y., Higashi, K., Mouri, T., Hata, F., Takeguchi, N., andKurachi, Y. (2002) J. Physiol. (Lond.) 540, 85–92
45. Malinowska, D. H., Sherry, A. M., Tewari, K. P., and Cuppoletti, J. (2004)Am. J. Physiol. 286, C495–C506
46. Stanfield, P. R. (1983) Rev. Physiol. Biochem. Pharmacol. 97, 1–6747. Durbin, R. P., Frank, H., and Solomon, A. K. (1956) J. Gen. Physiol. 39,
535–55148. Heinz, E., and Obrink, K. J. (1954) Physiol. Rev. 34, 643–67349. Powell, D. W., and Hirschowitz, B. I. (1964) Am. J. Physiol. 207, 868–87250. Debellis, L., Curci, S., and Fromter, E. (1990) Am. J. Physiol. 258,
G631–G63651. Ueda, S., and Okada, Y. (1989) Biochim. Biophys. Acta. 1012, 254–26052. Okada, Y., and Ueda, S. (1984) J. Physiol. (Lond.) 354, 109–11953. Cuppoletti, J., and Sachs, G. (1984) J. Biol. Chem. 259, 14952–1495954. Reenstra, W. W., and Forte, J. G. (1990) Am. J. Physiol. 259, G850–G85855. Wolosin, J. M., and Forte, J. G. (1984) Am. J. Physiol. 246, C537–C54556. Wallmark, B., Stewart, H. B., Rabon, E., Saccomani, G., and Sachs, G.
(1980) J. Biol. Chem. 255, 5313–531957. Cuppoletti, J., Baker, A. M., and Malinowska, D. H. (1993) Am. J. Physiol.
264, C1609–C161858. Malinowska, D. H., Kupert, E. Y., Bahinski, A., Sherry, A. M., and Cuppo-
letti, J. (1995) Am. J. Physiol. 268, C191–C20059. Hogben, C. A. (1955) Am. J. Physiol. 180, 641–64960. Forte, J. G., and Machen, T. E. (1975) J. Physiol. (Lond.) 244, 33–5161. Forte, J. G., Machen, T. E., and Obrink, K. J. (1980)Annu. Rev. Physiol. 42,
111–12662. Bleich, M., Briel, M., Busch, A. E., Lang, H. J., Gerlach, U., Gogelein, H.,
Greger, R., and Kunzelmann, K. (1997) Pflugers Arch. 434, 499–50163. McDaniel, N., Pace, A. J., Spiegel, S., Engelhardt, R., Koller, B. H., Seidler,
U., and Lytle, C. (2005) Am. J. Physiol. 289, G550–G56064. Brown, E. M., and MacLeod, R. J. (2001) Physiol. Rev. 81, 239–29765. Fujiwara, K., Masuyama, Y., and Kitagawa, T. (1996)Histochem. Cell Biol.
106, 465–47166. Quinn, S. J., Ye, C. P., Diaz, R., Kifor, O., Bai,M., Vassilev, P., and Brown, E.
(1997) Am. J. Physiol. 273, C1315–C132367. Chew, C. S., Hersey, S. J., Sachs, G., and Berglindh, T. (1980)Am. J. Physiol.
238, G312–G32068. Ochi, Y., Horie, S., Maruyama, T., Watanabe, K., and Yano, S. (2005) Life
Sci. 77, 2040–205069. Heitzmann, D., Grahammer, F., von Hahn, T., Schmitt-Graff, A., Romeo,
E., Nitschke, R., Gerlach, U., Lang, H. J., Verrey, F., Barhanin, J., andWarth, R. (2004) J. Physiol. (Lond.) 561, 547–557
70. Forte, J. G., and Yao, X. (1996) Trends Cell Biol. 6, 45–4871. Demarest, J. R., Loo, D. D., and Sachs, G. (1989) Science 245, 402–404
Water Transport in the Gastric Gland Lumen
MAY 4, 2007 • VOLUME 282 • NUMBER 18 JOURNAL OF BIOLOGICAL CHEMISTRY 13485
by guest on August 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from

72. Malinowska, D. H., Cuppoletti, J., and Sachs, G. (1983)Am. J. Physiol. 245,G573–G581
73. Nakahari, T., Steward,M. C., Yoshida, H., and Imai, Y. (1997) Exp. Physiol.82, 55–70
74. Zeuthen, T. (1994) J. Physiol. 478, 203–21975. Bijvelds, M. J., Jorna, H., Verkade, H. J., Bot, A. G., Hofmann, F., Agellon,
L. B., Sinaasappel, M., and de Jonge, H. R. (2005) Am. J. Physiol. 289,G870–G879
76. Nishizawa, T., Nagao, T., Iwatsubo, T., Forte, J. G., and Urushidani, T.(2000) J. Biol. Chem. 275, 11164–11173
77. Mizukawa, Y., Nishizawa, T., Nagao, T., Kitamura, K., and Urushidani, T.(2002) Am. J. Physiol. 282, C786–C795
78. Logsdon, C. D., and Machen, T. E. (1982) Am. J. Physiol. 242,G388–G399
79. Villegas, L., and Sananes, L. (1968) Am. J. Physiol. 214, 997–100080. Dedek, K., and Waldegger, S. (2001) Pflugers Arch. 442, 896–90281. Lambrecht, N. W., Yakubov, I., Scott, D., and Sachs, G. (2005) Physiol.
Genomics 21, 81–9182. Holm, L., and Flemstrom, G. (1990) J. Intern. Med. 732, (suppl.) 91–9583. Richter, C., Tanaka, T., and Yada, R. Y. (1998) Biochem. J. 335, 481–490
Water Transport in the Gastric Gland Lumen
13486 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 18 • MAY 4, 2007
by guest on August 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from

Debellis, Rosa Caroppo and Silvana CurciAndrea Gerbino, Gregorio Fistetto, Matilde Colella, Aldebaran M. Hofer, LucantonioMODULATION VIA THE EXTRACELLULAR Ca2+-SENSING RECEPTOR
Real Time Measurements of Water Flow in Amphibian Gastric Glands:
doi: 10.1074/jbc.M610585200 originally published online March 15, 20072007, 282:13477-13486.J. Biol. Chem.
10.1074/jbc.M610585200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/282/18/13477.full.html#ref-list-1
This article cites 82 references, 22 of which can be accessed free at
by guest on August 27, 2016
http://ww
w.jbc.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















