
Aged garlic extract attenuates gentamicin induced renal damage and oxidative stress in rats
Upload
independentCategory
view
0download
0
lsevier.com/locate/lifescie
Life Sciences 78 (20
Reactive oxygen species scavenging capacity of different
cooked garlic preparations
Jose Pedraza-Chaverrı *, Omar N. Medina-Campos, Rosaura Avila-Lombardo,
Alma Berenice Zuniga-Bustos, Marisol Orozco-Ibarra
Facultad de Quımica, Edificio B, Segundo Piso, Laboratorio 209, Departamento de Biologıa, Universidad Nacional Autonoma de Mexico (UNAM),
Ciudad Universitaria, 04510, Mexico, D.F., Mexico
Received 4 March 2005; accepted 16 May 2005
Abstract
It was studied if the ability of aqueous garlic extracts to scavenge superoxide anion (O2&�), hydrogen peroxide (H2O2), and hydroxyl radical
(OH&) is altered in the following aqueous preparations: (a) extracts of boiled garlic cloves (BG), (b) extracts of microwave-treated garlic cloves
(MG), and (c) extracts of pickled garlic (PG), and heated extracts of (a) garlic powder (HGP) and (b) raw garlic (HRG). The data were compared
with the unheated raw garlic (RG) or with the unheated garlic powder (GP). Extracts of GP and RG scavenged O2&�, H2O2, and OH& in a
concentration-dependent way. The reactive oxygen species scavenging capacity was not decreased in the aqueous garlic extracts except in MG and
HRG (for O2&�) and in HGP and PG (for H2O2). The heating before or after garlic cutting was unable to eliminate the capacity of the extracts to
scavenge H2O2, O2&�, and OH&.
D 2005 Elsevier Inc. All rights reserved.
Keywords: Raw garlic; Pickled garlic; Microwave-treated garlic; Boiled garlic; Superoxide anion; Hydrogen peroxide; Hydroxyl radical
Introduction
Garlic (Allium sativum) has been cultivated since ancient
times and used as a spice and condiment for many centuries
(Block, 1985). Ninety-five percent of the sulfur in intact garlic
cloves is found in two classes of compounds in similar
abundance: the S-alkylcysteine sulfoxides and the g-glu-
tamyl-S-alkylcysteines (Lawson, 1998). The most abundant
sulfur compound in garlic is alliin (S-allylcysteine sulfoxide),
which is present at 10 mg/g fresh garlic or 30 mg/g dry weight
(Lawson, 1998). When garlic cloves are cut, crushed, or
chopped (or when the powder of dried cloves becomes wet in a
non-acid solution), the cysteine sulfoxides, which are odorless,
are very rapidly converted to a new class of compounds, the
thiosulfinates, which are responsible for the odor of freshly
chopped garlic. The formation of thiosulfinates takes place
when the cysteine sulfoxides, which are located only in the
clove mesophyll storage cells, come in contact with the enzyme
allinase or alliin lyase (E.C. 4.4.1.4), which is located only in
0024-3205/$ - see front matter D 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.lfs.2005.05.075
* Corresponding author. Tel./fax: +52 55 5622 3515.
E-mail address: [email protected] (J. Pedraza-Chaverrı).
the vascular bundle sheath cells. Allinase is active at pH 4–5.8,
but is immediately inhibited at acidic pH values below 3.5 or
by cooking. Furthermore, microwave heating destroys allinase
activity in 1 min (Song and Milner, 1999). Due to the
abundance of alliin, the main thiosulfinate formed upon
crushing garlic is allicin (Lawson, 1998). The half-life of
allicin at room temperature is 2–16 h; however, in crushed
garlic (or in garlic juice) it is 2.4 days (Lawson, 1998).
During the past years, there has been a growing awareness of
the potential medicinal uses of garlic (Banerjee et al., 2002b;
Pedraza-Chaverri et al., 1998). The antioxidant properties of
garlic are well documented (Banerjee et al., 2002b; Dillon et al.,
2002; Rahman, 2003; Pedraza-Chaverri et al., 2000; Sener et
al., 2003, 2005; Gedik et al., 2005). Diet with 2% of garlic
powder decreased the ischemia and reperfusion-induced
arrhythmias (Rietz et al., 1993) and the acute renal failure and
oxidative stress induced by gentamicin (Pedraza-Chaverri et al.,
2000). Garlic feeding also decreased lipid peroxidation and
prevented the decrease in glutathione peroxidase activity in red
blood cells of mice treated with adriamycin (Thabrew et al.,
2000). In addition, chronic administration of raw garlic
homogenate protects heart against oxidative damage induced
06) 761 – 770
www.e
J. Pedraza-Chaverrı et al. / Life Sciences 78 (2006) 761–770762
by ischemia and reperfusion (Mukherjee et al., 2002). Fur-
thermore, it has been found that aqueous extract of raw garlic
scavenges hydroxyl radical (OH&) (Prasad et al., 1996; Kim et
al., 2001) and superoxide anion (O2&�) (Kim et al., 2001),
inhibits lipid peroxidation (Prasad et al., 1996; Yin and Cheng,
1998a), Cu2+-induced lipoprotein oxidation (Pedraza-Chaverri
et al., 2004b), and the formation of lipid hydroperoxides (Prasad
et al., 1996; Kim et al., 2001). Furthermore, aqueous extracts of
0
25
50
75
100
125
150
0 0.3 0.7 2.6 3.1 84.1 101.5
b
b
bb
b
b b
[GP], mg/mL
0
25
50
75
100
125
b
bb
b
bb
0
25
50
75
100
125
a bb
b
b
bb b
0
25
50
75
100
125
0 0.1 0.4 1 2 4 8 15 20
b
bb
b b
[MG], mg/mL
0 0.1 0.4 1 2 4 8 15 20
[BG], mg/mL
0 0.1 0.3 0.6 1.5 3 15 30 60
[NDGA], µg/mL
A
C
E
G
O2.- SCAVENGIN
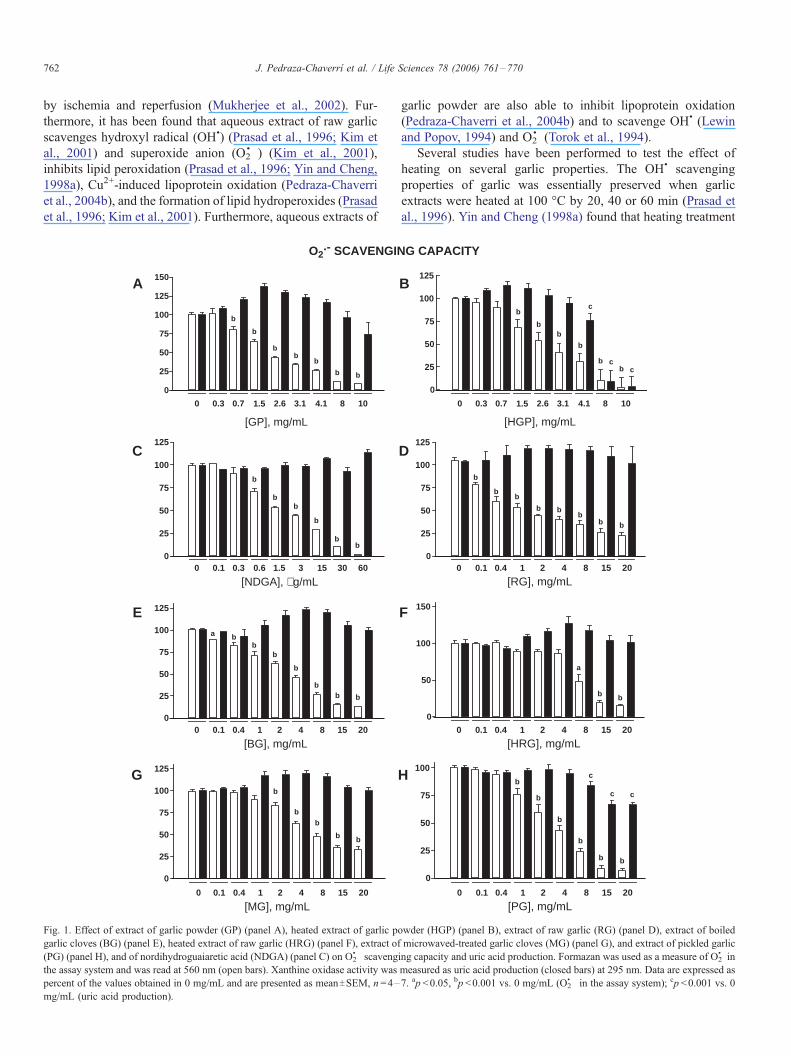
Fig. 1. Effect of extract of garlic powder (GP) (panel A), heated extract of garlic p
garlic cloves (BG) (panel E), heated extract of raw garlic (HRG) (panel F), extract of
(PG) (panel H), and of nordihydroguaiaretic acid (NDGA) (panel C) on O2&� scaveng
the assay system and was read at 560 nm (open bars). Xanthine oxidase activity was
percent of the values obtained in 0 mg/mL and are presented as meanTSEM, n =4–
mg/mL (uric acid production).
garlic powder are also able to inhibit lipoprotein oxidation
(Pedraza-Chaverri et al., 2004b) and to scavenge OH& (Lewin
and Popov, 1994) and O2&�(Torok et al., 1994).
Several studies have been performed to test the effect of
heating on several garlic properties. The OH& scavenging
properties of garlic was essentially preserved when garlic
extracts were heated at 100 -C by 20, 40 or 60 min (Prasad et
al., 1996). Yin and Cheng (1998a) found that heating treatment
0 0.3 0.7 2.6 3.1 84.1 101.5
0
25
50
75
100
125
b
bb
b
bb c
c
c
[HGP], mg/mL
0
25
50
75
100
125
b
bb
b bb
b b
0
50
100
150
b
a
b
0 0.1 0.4 1
0
25
50
75
100
2 4 8 15 20
b
b
b
b
b b
c
c c
[PG], mg/mL
0 0.1 0.4 1 2 4 8 15 20
[HRG], mg/mL
0 0.1 0.4 1 2 4 8 15 20
[RG], mg/mL
B
D
F
H
G CAPACITY
owder (HGP) (panel B), extract of raw garlic (RG) (panel D), extract of boiled
microwaved-treated garlic cloves (MG) (panel G), and extract of pickled garlic
ing capacity and uric acid production. Formazan was used as a measure of O2&�in
measured as uric acid production (closed bars) at 295 nm. Data are expressed as
7. ap <0.05, bp <0.001 vs. 0 mg/mL (O2&� in the assay system); cp <0.001 vs. 0

Table 1
O2&� scavenging capacity of nordihydroguaiaretic acid (NDGA) and extracts of
garlic powder (GP) and heated garlic powder (HGP)
Garlic extract or standard IC50, mg/mL
NDGA (7) 0.0031T0.00014
GP (4) 2.5160T0.4640a
HGP (4) 2.6610T0.3171a
Data are meanTSEM. ap <0.001 vs. NDGA. Number of determinations are in
parentheses.
Table 2
O2&� scavenging capacity of nordihydroguaiaretic acid (NDGA), extract of raw
garlic (RG), extract of boiled garlic cloves (BG), heated extract of raw garlic
(HRG), extract of microwave-treated garlic cloves (MG), and extract of pickled
garlic (PG)
Garlic extract or standard IC50, mg/mL
NDGA (7) 0.0031T0.00014
RG (7) 1.9270T0.5620a
BG (7) 2.4910T0.3000b
HRG (5) 7.8370T0.8088c,d
MG (6) 7.7100T0.7401c,d
PG (5) 2.8370T0.4422b
Data are meanTSEM. ap <0.05, bp <0.01, cp <0.001 vs. NDGA, dp <0.001 vs.
RG. Number of determinations are in parentheses.
J. Pedraza-Chaverrı et al. / Life Sciences 78 (2006) 761–770 763
(100 -C for 15 min in an oven) of chopped garlic reduced the
ability to inhibit lipid peroxidation. In contrast, Shobana and
Naidu (2000) found that the capacity of garlic to inhibit lipid
peroxidation was not affected by boiling (30 min at 100 -C). We
recently showed that garlic’s ability to prevent in vitro Cu2+-
induced lipoprotein oxidation in human serum is preserved in
heated garlic (Pedraza-Chaverri et al., 2004b). However, to our
knowledge, there are no studies exploring if the heating of garlic
cloves affects its ability to scavenge O2&�, hydrogen peroxide
(H2O2), and OH&, or if the heating of aqueous extract of raw
garlic or garlic powder affects their ability to scavenge O2&� or
H2O2. In the present paper we studied if the ability of aqueous
garlic extracts to scavenge O2&�, H2O2, and OH
& is altered in the
following aqueous preparations: (a) extracts of boiled garlic
cloves (BG), (b) extracts of microwave-treated garlic cloves
(MG), and (c) extracts of pickled garlic (PG), and heated extract
of (a) garlic powder (HGP) or (b) of raw garlic (HRG). The data
were compared with the unheated raw garlic (RG) or with the
unheated garlic powder (GP).
Materials and methods
Materials and reagents
Bulbs of garlic were obtained in a local market. Garlic
powder was obtained from McCormick (Mexico City, Mexico).
Xanthine oxidase, xanthine, nitroblue tetrazolium (NBT),
xylenol orange, butylated hydroxytoluene, thiobarbituric acid,
and 2-deoxy-d-ribose, deferoxamine mesylate, nordihydro-
guaiaretic acid (NDGA), sodium pyruvate, dimethylthiourea
(DMTU), and tetramethoxypropane were purchased from
Sigma Chemical Co (St. Louis, Mo., USA).
Preparation of aqueous extracts of garlic
Extract of garlic powder (GP)
Garlic powder was weighted (0.3 g), dissolved, and stirred
with 3 mL of distilled water for 10 min. This solution was
centrifuged at 20,124 �g for 10 min at 4 -C. The supernatant
was recovered and used at the indicated final concentrations.
Heated extract of garlic powder (HGP)
The procedure was similar to the previous one except that
the mixture was boiled for 30 min before the centrifugation
step. The amount of water evaporated was replaced at the end
of the heating. The supernatant was recovered and used at the
indicated final concentrations.
Extract of raw garlic (RG)
Garlic cloves were peeled off, weighted, chopped, and
homogenized with distilled water in a Polytron (Model
PT2000, Brinkmann, Switzerland). This homogenate was
centrifuged at 20,124 �g for 10 min at 4 -C. The supernatant
was recovered and used at the indicated final concentrations.
Heated extract of raw garlic (HRG)
The procedure was similar to the previous one except that
the homogenate was boiled for 30 min before the centrifugation
step. The amount of water evaporated was replaced at the end
of the heating. The supernatant was recovered and used at the
indicated final concentration.
Extract of boiled garlic cloves (BG)
Unpeeled garlic cloves were boiled in water for 30 min.
After this time, garlic cloves were peeled off and the aqueous
extract was prepared as described before (extract of raw garlic).
The supernatant was recovered and used at the indicated final
concentrations.
Extract of microwave-treated garlic cloves (MG)
Unpeeled garlic cloves were submitted to microwave
heating for 30 s (1100 W). After this time, garlic cloves
were peeled off and the aqueous extract was prepared as
described before (extract of raw garlic). When allinase is
inactivated by heating, the cascade of thiosulfinate formation
is blocked from alliin, and allicin and its derivates can not be
formed. It has been shown that as little as 60 s of microwave
heating (600 W) can totally destroy allinase enzyme activity
whereas microwave heating for 30 s (600 W) inhibits 90% of
allinase activity compared with unheated garlic (3). The
supernatant was recovered and used at the indicated final
concentrations.
Extract of pickled garlic (PG)
Garlic cloves were peeled off carefully to avoid allinase
activation and put in an aqueous solution of vinegar (1 :1, v/v)
and then heated to the boiling point for 30 min. The
experiments with pickled garlic were performed the same
day. The aqueous extract was prepared as described before
(extract of raw garlic). The supernatant was recovered and used
at the indicated final concentrations.
J. Pedraza-Chaverrı et al. / Life Sciences 78 (2006) 761–770764
O2&� scavenging assay
Xanthine–xanthine oxidase system was used to determine
the O2&� scavenging capacity of the extracts. O2
&� in the assay
system and xanthine oxidase activity were measured as NBT
reduction and uric acid production, respectively (Bielski et al.,
1980), using a DU-64O series Beckman spectrophotometer.
This system is useful to test for O2&� scavenging capacity only
0
25
50
75
100b
b
0 0.1 0.3 0.8 1.5 2.6 4.6
0 0.1 0.3 0.8 1.5 2.6 4.6
0 0.1 0.3 0.8 1.5 2.6 4.6
0 0.1 0.3 0.8 1.5 2.6 4.6
b
bb
[GP], mg/mL
0
25
50
75
100 b
b
b
bb b
[Pyruvate], mg/mL
0
25
50
75
100b
b
b
b
b
[BG], mg/mL
0
25
50
75
100 b
b
b
b b b
[MG], mg/mL
H2O2 SCAVENGING CAP
A
C
E
G
B
D
F
H
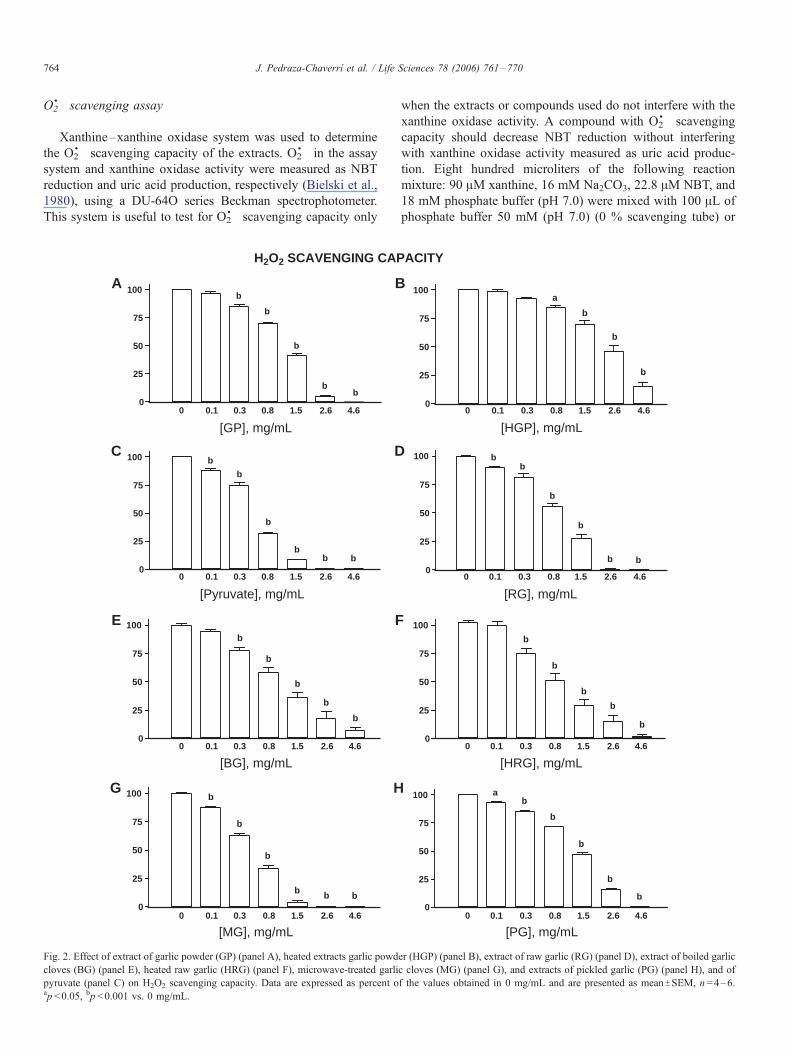
Fig. 2. Effect of extract of garlic powder (GP) (panel A), heated extracts garlic powd
cloves (BG) (panel E), heated raw garlic (HRG) (panel F), microwave-treated garli
pyruvate (panel C) on H2O2 scavenging capacity. Data are expressed as percent oap <0.05, bp <0.001 vs. 0 mg/mL.
when the extracts or compounds used do not interfere with the
xanthine oxidase activity. A compound with O2&� scavenging
capacity should decrease NBT reduction without interfering
with xanthine oxidase activity measured as uric acid produc-
tion. Eight hundred microliters of the following reaction
mixture: 90 AM xanthine, 16 mM Na2CO3, 22.8 AM NBT, and
18 mM phosphate buffer (pH 7.0) were mixed with 100 AL of
phosphate buffer 50 mM (pH 7.0) (0 % scavenging tube) or
0 0.1 0.3 0.8 1.5 2.6 4.6
0 0.1 0.3 0.8 1.5 2.6 4.6
0 0.1 0.3 0.8 1.5 2.6 4.6
0 0.1 0.3 0.8 1.5 2.6 4.6
0
25
50
75
100a
b
b
b
[HGP], mg/mL
0
25
50
75
100 bb
b
b
b b
[RG], mg/mL
0
25
50
75
100
b
b
b
b
b
[HRG], mg/mL
0
25
50
75
100 ab
b
b
b
b
[PG], mg/mL
ACITY
er (HGP) (panel B), extract of raw garlic (RG) (panel D), extract of boiled garlic
c cloves (MG) (panel G), and extracts of pickled garlic (PG) (panel H), and of
f the values obtained in 0 mg/mL and are presented as meanTSEM, n =4–6.

Table 3
H2O2 scavenging capacity of pyruvate and of garlic powder (GP) and heated
extract of garlic powder (HGP)
Garlic extract or standard IC50, mg/mL
Pyruvate (6) 0.4856T0.0222
GP (4) 0.8509T0.0067a
HGP (4) 1.7646T0.0974a,b
Data are meanTSEM. ap <0.001 vs. pyruvate; bp <0.001 vs. GP. Number of
determinations are in parentheses.
Table 4
H2O2 scavenging capacity of pyruvate and extract of raw garlic (RG), extract of
boiled garlic cloves (BG), heated extract of raw garlic (HRG), extract of
microwave-treated garlic cloves (MG), and extract of pickled garlic (PG)
Garlic extract or standard IC50, mg/mL
Pyruvate (6) 0.4856T0.0222
RG (4) 0.6416T0.0421BG (4) 0.8226T0.0772a
HRG (4) 0.7330T0.0596a
MG (6) 0.4592T0.0263
PG (4) 0.9618T0.0175a,b
Data are meanTSEM. ap <0.01 vs. pyruvate, bp <0.001 vs. RG. Number of
determinations are in parentheses.
J. Pedraza-Chaverrı et al. / Life Sciences 78 (2006) 761–770 765
with 100 AL of different concentrations of the garlic extracts.
The reaction was started by the addition of 100 AL of xanthine
oxidase (168 U/L). Optical density was registered both at 295
nm (for uric acid production) and 560 nm (for O2&� in the
assay system). Scavenging percent was obtained from the
optical densities at 560 nm. NDGA was used as standard for
O2&� scavenging in this assay (unpublished data from our
laboratory).
Determination of H2O2 by the ferrous ion oxidation-xylenol
orange (FOX) assay
A solution of 75 AM H2O2 was mixed (1 :1 v/v) with
water (0% scavenging tube) or with different concentrations
of garlic extracts and incubated for 30 min at room temper-
ature. After this, H2O2 was measured by the method
described by Long et al. (1999). Briefly, 9 volumes of 4.4
mM butylated hydroxytoluene in HPLC-grade methanol were
mixed with 1 volume of 1 mM xylenol orange and 2.56 mM
ammonium ferrous sulfate in 0.25 M H2SO4 to give the
‘‘working’’ FOX reagent. Ninety microliters of the H2O2-
sample solutions were pippeted in 1.5 mL Eppendorf tubes
and mixed with 0.01 mL of HPLC-grade methanol immedi-
ately followed by the addition of 0.9 mL of FOX reagent,
vortexed for 5 s and then incubated at room temperature for
10 min. The tubes were centrifuged for 15,000 �g for 10 min
and absorbance at 560 nm was read against methanol blank.
The concentration of H2O2 was calculated from a standard
curve prepared with increasing H2O2 concentrations. Pyruvate
was used as standard for H2O2 scavenging (Salahudeen et al.,
1991).
OH & scavenging assay
The ability of garlic extracts to scavenge OH& was
conducted in the Fe3+–EDTA–H2O2-deoxyribose system
(Halliwell et al., 1987). Nine hundred microliters of the
following reaction mixture: 0.2 mM ascorbic acid, 0.2 mM
FeCl3, 0.208 mM EDTA, 1 mM H2O2, 0.56 mM 2-deoxy-d-
ribose, and 20 mM phosphate buffer (pH 7.4) were mixed
with 100 AL of distilled water (0% scavenging tube) or with
100 AL of different concentrations of garlic extracts.
Hydroxyl radicals were generated by incubating the mixture
at 37 -C for 60 min. The iron salt (FeCl3) was mixed with
EDTA before addition to the reaction mixture. The extent of
2-deoxy-d-ribose degradation by the formed OH& was
measured directly in the aqueous phase by the thiobarbituric
acid test. DMTU was used as standard for OH& scavenging
(Walker and Shah, 1988).
Statistics
Data are expressed as meanTSEM. The data were compared
against 0% scavenging tube using one way analyses of variance
and Dunnett post-test. O2&� and H2O2 scavenging capacity was
expressed as 50% of inhibitory concentration (IC50) value,
which denotes the concentration of the extract and standard
(mg/mL) required to give a 50% reduction in scavenging
capacity relative to the tube without extract or standard. The
lower the IC50 value the higher is the scavenging capacity of the
compound. OH& scavenging capacity was expressed as percent
of the tube without extract. The IC50 values and the percent data
were compared by one way analysis of variance and Bonferroni
t post-test. P <0.05 was considered significant.
Results
O2&� scavenging capacity of garlic extracts
Fig. 1 shows the effect of garlic extracts (panels A, B, and
D–H) and of NDGA (panel C) on O2&� in the assay system and
uric acid production. Data are expressed as percent of O2&� in
the assay system and uric acid production, and were compared
against the tube without garlic extract or NDGA (0 Ag/mL).
Garlic extracts and NDGA decreased concentration-depend-
ently O2&� in the assay system. GP and HGP decreased
concentration-dependently O2&� in the assay system starting at
0.7 and 1.5 mg/mL, respectively (Panels A and B). GP was
unable to affect uric acid production indicating that the
decrease in O2&� in the assay system was not secondary to the
inhibition of xanthine oxidase. In contrast, HGP decreased uric
acid production at 4.1, 8 and 10 mg/mL suggesting that the
decrease in O2&� in the assay system is affected partially (at 4.1
mg/mL) or completely (at 8 and 10 mg/mL) by the decrease in
xanthine oxidase activity. However, it is clear that at 1.6–3.1
mg/mL, HGP decreases O2&� in the assay system without
affecting uric acid production. Panel C shows that the standard
NDGA decreased O2&� in the assay system starting on 0.6 Ag/
mL without affecting uric acid production. Table 1 shows that
the IC50 values for GP and HGP were not different between
them but they were higher than that of NDGA indicating that
J. Pedraza-Chaverrı et al. / Life Sciences 78 (2006) 761–770766
(1) the heating was unable to affect the scavenging capacity of
garlic extract and (2) NDGA is more effective to scavenge O2&�
than GP and HGP. Panels D–H show the effect of RG, BG,
HRG, MG, and PG on the above mentioned parameters. The
decrease in O2&� in the assay system was significative at 0.1
mg/mL for RG and BG, at 1 mg/mL for PG, at 2 mg/mL for
MG, and at 8 mg/mL for HRG. PG decreased uric acid
production at the higher concentrations: 8–20 mg/mL, how-
ever the decrease in O2&� in the assay system was clearly higher
0
25
50
75
100 aa
a
a
0 0.1 0.3 0.7 1.5 2.6
0 0.1 0.3 0.7 1.5 2.6
0 0.1 0.3 0.7 1.5 2.6
[GP], mg/mL
0
25
50
75
100
0 0.002 0.01 0.08 0.625 2.6
a
a
aa a
[DMTU], mg/mL
0
25
50
75
100a a
a aa
[BG], mg/mL
0
25
50
75
100 a
aa a a
[MG], mg/mL
OH. SCAVENGING
A
C
E
G
B
D
F
H
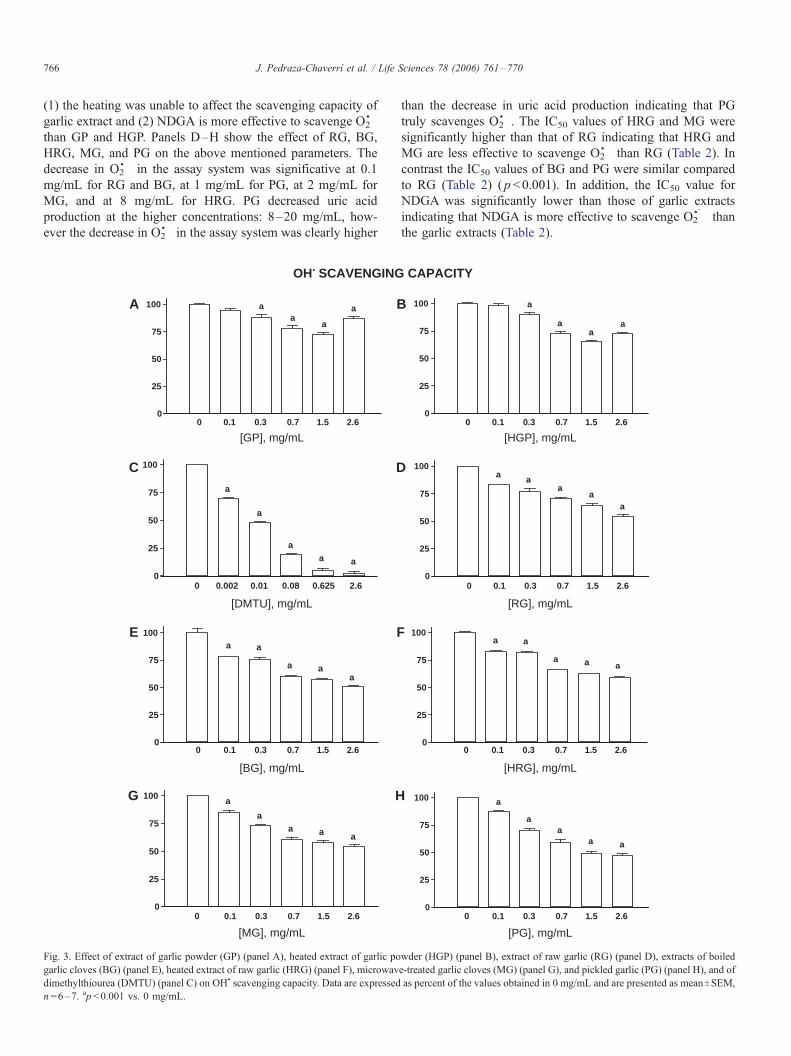
Fig. 3. Effect of extract of garlic powder (GP) (panel A), heated extract of garlic po
garlic cloves (BG) (panel E), heated extract of raw garlic (HRG) (panel F), microwav
dimethylthiourea (DMTU) (panel C) on OH& scavenging capacity. Data are expressed
n =6–7. ap <0.001 vs. 0 mg/mL.
than the decrease in uric acid production indicating that PG
truly scavenges O2&�. The IC50 values of HRG and MG were
significantly higher than that of RG indicating that HRG and
MG are less effective to scavenge O2&� than RG (Table 2). In
contrast the IC50 values of BG and PG were similar compared
to RG (Table 2) ( p <0.001). In addition, the IC50 value for
NDGA was significantly lower than those of garlic extracts
indicating that NDGA is more effective to scavenge O2&˙� than
the garlic extracts (Table 2).
0 0.1 0.3 0.7 1.5 2.6
0 0.1 0.3 0.7 1.5 2.6
0 0.1 0.3 0.7 1.5 2.6
0 0.1 0.3 0.7 1.5 2.6
0
25
50
75
100 a
aa
a
[HGP], mg/mL
0
25
50
75
100
aa
aa
a
[RG], mg/mL
0
25
50
75
100a a
a a a
[HRG], mg/mL
0
25
50
75
100 a
aa
a a
[PG], mg/mL
CAPACITY
wder (HGP) (panel B), extract of raw garlic (RG) (panel D), extracts of boiled
e-treated garlic cloves (MG) (panel G), and pickled garlic (PG) (panel H), and of
as percent of the values obtained in 0 mg/mL and are presented as meanTSEM,

Table 6
OH& scavenging capacity of dimethylthiourea (DMTU) and extract of raw
garlic (RG), extract of boiled garlic cloves (BG), heated extract of raw garlic
(HRG), extract of microwave-treated garlic cloves (MG), and extract of pickled
garlic (PG)
Extract or DMTU (2.61 mg/mL) OH& scavenging capacity (%)
DMTU (6) 95.93T0.29
RG (6) 42.94T1.22a
BG (7) 41.37T3.65a
HRG (7) 46.04T1.32a
MG (6) 44.89T0.76a
PG (6) 48.21T2.15a
Data are meanTSEM. ap <0.001 vs. DMTU. Number of determinations are in
parentheses.
J. Pedraza-Chaverrı et al. / Life Sciences 78 (2006) 761–770 767
H2O2 scavenging capacity of garlic extracts
Fig. 2 shows the ability of garlic extracts (Panels A, B, and
D–H) and pyruvate (panel C) to scavenge H2O2. Data were
expressed as percent of H2O2 in the assay mixture and were
compared against tube 0 mg/mL which does not contain
garlic extract or pyruvate. Garlic extracts and pyruvate
scavenged H2O2 in a concentration-dependent way. GP and
HGP scavenged H2O2 starting at 0.3 and 0.8 mg/mL,
respectively (Panels A and B). Panel C shows the effect of
the standard pyruvate which decreased H2O2 concentration
starting at 0.1 mg/mL. The IC50 value for HGP was
significantly higher than that of RG (Table 3) indicating that
heating decreased the H2O2 scavenging capacity of garlic
extract. IC50 for pyruvate was significantly lower that those
for GP and HGP indicating that pyruvate is more effective to
scavenge H2O2 than GP and HGP. Panels D–H show the
H2O2 scavenging capacity of RG, BG, HRG, MG, and PG.
The H2O2 scavenging capacity was significative at 0.1 mg/
mL for RG, MG, and PG, and at 0.3 mg/mL for BG and
HRG. The IC50 value of PG was significantly higher than that
of RG indicating that PG is less effective than RG to
scavenge H2O2. In contrast, the IC50 values of BG, HRG, and
MG were not different from that of RG (Table 4) ( p <0.001).
The IC50 value of pyruvate was similar than those of RG and
MG but lower than those of BG, HRG and PG indicating that
pyruvate is more effective to scavenge H2O2 than these three
last extracts.
Hydroxyl radical scavenging capacity of garlic extracts
Fig. 3 shows the ability of garlic extracts (Panels A, B, and
D–H) and DMTU (Panel C) to scavenge OH&. Data are
expressed as percent of OH& in the assay mixture and were
compared against the tube without garlic extract (0 mg/mL).
The garlic extracts and the standard DMTU scavenged OH& in a
concentration-dependent way. The OH& scavenging capacity of
GP and HGP was significative at 0.3 mg/mL (Panel A and B)
and that of DMTU was significative at 0.002 mg/mL (Panel C).
Concentrations of GP and HGP higher than 1.5 mg/mL
interfered with the assay (Panels A and B). Concentrations of
RG, BG, HRG, MG, and PG higher than 2.6 mg/mL interfered
with the assay (data not shown). Therefore, we were unable to
test higher concentrations of garlic extracts in our system and
thus to reach 50% of OH& scavenging capacity (with the
exception of PG) which prevented us to calculate IC50. Then,
the comparisons among the OH& scavenging capacity were
Table 5
OH& scavenging capacity of dimethylthiourea (DMTU) and extracts of garlic
powder (GP) and heated garlic powder (HGP)
Extract or DMTU (1.53 mg/mL) OH& scavenging capacity (%)
DMTU (6) 95.61T0.47
GP (6) 27.59T0.49a
HGP (6) 30.57T1.75a
Data are meanTSEM. ap <0.001 vs. DMTU. Number of determinations are in
parentheses.
made at the maximum concentration tested. Table 5 shows that
the OH& scavenging capacity was unaffected by the heating
(GP vs. HGP, p >0.05). In addition, the OH& scavenging
capacity of DMTU was significantly higher than those of GP
and HGP (Table 5). The OH& scavenging capacity of RG, BG,
HRG, MG and PG was significative at 0.1 mg/mL (Panels D–
H). The OH& scavenging capacity was not significantly
different among the extracts (Table 6) ( p >0.05). However,
the OH& scavenging capacity of DMTU was significantly
higher than those of RG, BG, HRG, MG, and PG (Table 6)
( p <0.001).
Discussion
Garlic is among the oldest of all cultivated plants and has
been used as a medicinal agent for thousands of years. Many
recent studies have demonstrated several pharmacologic effects
of garlic, such as antibacterial, antifungal, hypolipidemic,
hypoglycemic, antithrombotic, antihypertensive, and anti-
cancer properties (Block, 1985; Lawson, 1998; Song and
Milner, 1999; Banerjee et al., 2002a; Pedraza-Chaverri et al.,
1998; Dillon et al., 2002). It has been shown in many cases that
the protective effect of garlic is associated with its antioxidant
properties (Banerjee et al., 2002b). The method of garlic’s
preparation may influence its medicinal properties (Banerjee et
al., 2002a,b; Dillon et al., 2002; Rahman, 2003; Pedraza-
Chaverri et al., 2000; Rietz et al., 1993; Thabrew et al., 2000;
Mukherjee et al., 2002). In this context, several studies have
been performed to test the effect of heating on several garlic
properties including the antioxidant properties. The boiling of
garlic cloves by 15 min impairs significantly its ability to
inhibit cyclooxygenase activity (Ali, 1995) and thromboxane
B2 synthesis (Bordia et al., 1996). In addition, heating of garlic
cloves by 60 s in microwave reduces its anticancer properties
(Song and Milner, 1999). Interestingly when microwave
heating was applied 10 min after garlic crushing the anticancer
properties were preserved indicating that allinase activation is
necessary to generate anticancer compounds which are thermo-
stable (Song and Milner, 1999). In a similar way, the OH&
scavenging properties of garlic were essentially preserved
when garlic extracts were heated (Prasad et al., 1996). In
contrast, heating of garlic extracts by 10 min at 100 -C reduced
the bactericidal activity against Helicobacter pylori (Cellini et

J. Pedraza-Chaverrı et al. / Life Sciences 78 (2006) 761–770768
al., 1996), the antifungal activity (Yin and Cheng, 1998b), and
the ability to inhibit platelet aggregation (Ali et al., 1999). In
addition, it was recently shown that the ability of garlic extracts
to inhibit Cu2+-induced lipoprotein oxidation in human serum
was not affected by heating garlic cloves before cutting (by
boiling, microwaving, and pickling) or by the heating of
extracts of garlic powder or raw garlic showing that this
antioxidant property is not affected by the heating before or
after garlic cutting (Pedraza-Chaverrı et al., 2004b). In this
work it was studied the O2&�, OH&, and H2O2 scavenging
capacity of heated extracts of garlic powder and raw garlic and
aqueous extracts of boiled garlic cloves, microwave-treated
garlic cloves, and pickled garlic to test (a) if the scavenging
compound(s) are thermostable and (b) if allinase activity is
related with the antioxidant capacity of garlic extracts.
Chopping or crushing of garlic releases alliinase which rapidly
converts alliin (S(+)-alkyl-l-cysteine sulfoxide) to allicin
(dialkyl thiosulfinate) (Lawson, 1998). Allicin is the major
thiosulfinate compound found in crushed garlic but is quite
unstable and quickly is converted to several other sulfur
compounds, such as diallyl sulfide, diallyl disulfide, diallyl
trisulfide, ajoene, and polysulfide compounds (Block, 1985).
In this work we have confirmed the observation of Prasad
et al. (1996) who found that the OH& scavenging properties of
garlic were essentially preserved when extracts of garlic
powder were heated at 100 -C by 20, 40 or 60 min. In
addition, we found that the O2&� scavenging capacity of the
heated extract of garlic powder (HGP) is preserved (Fig. 1)
and that the H2O2 scavenging capacity of the above extract is
decreased, but no eliminated, compared to the unheated extract
of garlic powder (GP) (Fig. 2). Furthermore, the heating of
raw garlic extracts does not affect the OH& and H2O2
scavenging properties but it decreases the O2&� scavenging
properties. The above data suggest that the heating is not able
to eliminate the scavenging properties of aqueous extract of
garlic powder or raw garlic suggesting that the compound(s)
involved in these properties are thermostable. The in vitro
scavenging properties of extracts of garlic powder or raw
garlic are in agreement with the in vivo antioxidant properties
of these extracts found in several experimental models such as
gentamicin nephrotoxicity (Pedraza-Chaverri et al., 2000),
heart ischemia and reperfusion (Rietz et al., 1993; Mukherjee
et al., 2002), adriamycin toxicity (Thabrew et al., 2000),
quinolinic acid toxicity (Perez-Severiano et al., 2004a),
amyloid-beta peptide-induced neurotoxicity (Perez-Severiano
et al., 2004b), and cancer (reviewed in Khanum et al., 2004).
The H2O2 scavenging ability of garlic observed in the in vitro
assay in this work agrees with the decrease in H2O2
production in the kidney and liver of garlic-fed rats
(Pedraza-Chaverri et al., 2001).
We studied additional extracts in which the thermolabile
enzyme allinase was inhibited before garlic cutting (boiled
garlic, microwave-treated garlic cloves and pickled garlic). The
ability of these extracts to scavenge O2&�, H2O2, and OH& was
completely preserved with the following exceptions in which
the scavenging capacity was decreased but no eliminated: the
ability of microwave-treated garlic (MG) to scavenge O2&� (Fig.
1), and the ability of pickled garlic (PG) to scavenge H2O2
(Fig. 2). These data strongly suggest that the antioxidant
properties of garlic are not dependent of allinase activation.
Our data are essentially in agreement with previous findings in
which it was observed that the allinase activation is not related
to the ability of garlic to inhibit Cu2+-induced lipoprotein
oxidation in human serum (Pedraza-Chaverri et al., 2004b).
The antioxidant properties of allicin, the main component in
aqueous extract from raw garlic and garlic powder, may
explain, at least in part, the reactive oxygen species scavenging
capacity of extracts of raw garlic and garlic. It has been found
that allicin scavenges OH& and inhibits lipid peroxidation
(Prasad et al., 1995) and prevents the lung damage induced by
ischemia-reperfusion (Batirel et al., 2002). Furthermore, alliin,
the main component in extracts from boiled garlic cloves,
microwave-treated garlic cloves and pickled garlic, scavenges
OH& (Kourounakis and Rekka, 1991), H2O2 (Ide et al., 1996),
and inhibits lipid peroxidation (Ide et al., 1996) and LDL
oxidation (Ho et al., 2001). This may explain, at least in part,
the ability of boiled garlic, microwave-treated garlic, or pickled
garlic to scavenge reactive oxygen species. Interestingly, it has
been shown that another garlic compounds, such as S-allyl-
cysteine (Ide et al., 1997; Kim et al., 2001), N-acetyl-S-allyl-
cysteine (Ide et al., 1997), allixin (Ide et al., 1997), and S-
ethylcysteine, N-acetylcysteine, diallyl sulfide (Ou et al., 2003;
Pedraza-Chaverri et al., 2003b), diallyl disulfide (Ou et al.,
2003; Pedraza-Chaverri et al., 2003a; Koh et al., 2005), and S-
allylmercaptocysteine (Pedraza-Chaverrı et al., 2004a), which
may be present in some of the garlic extracts used in this work,
also have antioxidant properties.
Conclusion
In conclusion, the heating before or after garlic cutting was
unable to eliminate the capacity of the extracts to scavenge
O2&�, H2O2, and OH&. These data suggest that (a) the
compound(s) involved in the scavenging capacity of garlic
extracts are essentially heat stable and (b) the scavenging
capacity of the garlic extracts is not related to allinase activity.
Acknowledgements
Thisworkwas supported byCONACYT (GrantNo. 40009-M).
References
Ali, M., 1995. Mechanism by which garlic (Allium sativum) inhibits cyclo-
oxygenase activity. Effect of raw versus boiled garlic extract on the
synthesis of prostanoids. Prostaglandins, Leukotrienes, and Essential Fatty
Acids 53 (6), 397–400.
Ali, M., Bordia, T., Mustafa, T., 1999. Effect of raw versus boiled aqueous
extract of garlic and onion on platelet aggregation. Prostaglandins,
Leukotrienes, and Essential Fatty Acids 60 (1), 43–47.
Banerjee, S.K., Maulik, M., Mancahanda, S.C., Dinda, A.K., Gupta, S.K.,
Maulik, S.K., 2002a. Dose-dependent induction of endogenous antioxidants
in rat heart by chronic administration of garlic. Life Sciences 70 (13),
1509–1518.
Banerjee, S.K., Mukherjee, P.K., Maulik, S.K., 2002b. Garlic as an antioxidant:
the good, the bad and the ugly. Phytotherapy Research 17 (2), 97–106.

J. Pedraza-Chaverrı et al. / Life Sciences 78 (2006) 761–770 769
Batirel, H.F., Naka, Y., Kayano, K., Okada, K., Vural, K., Pinsky, D.J., Oz,
M.C., 2002. Intravenous allicin improves pulmonary blood flow after
ischemia-reperfusion injury in rats. The Journal of Cardiovascular Surgery
43 (2), 175–179.
Bielski, B.H.J., Shlue, G.G., Bajuk, S., 1980. Reduction of nitro blue
tetrazolium by CO2� and O2&� radicals. The Journal of Physical Chemistry
84, 830–833.
Block, E., 1985. The chemistry of garlic and onions. Scientific American 252
(3), 114–119.
Bordia, T., Mohammed, N., Thomson, M., Ali, M., 1996. An evaluation of
garlic and onion as antithrombotic agents. Prostaglandins, Leukotrienes,
and Essential Fatty Acids 54 (3), 183–186.
Cellini, L., Di Campli, E., Masulli, M., Di Bartolomeo, S., Allocati, N., 1996.
Inhibition of Helicobacter pylori by garlic extract (Allium sativum). FEMS
Immunology and Medical Microbiology 13 (4), 273–277.
Dillon, S.A., Lowe, G., Billington, D., Rahman, K., 2002. Dietary supplemen-
tation with aged garlic extract reduces plasma and urine concentrations of 8-
iso-prostaglandin F(2 alpha) in smoking and nonsmoking men and women.
The Journal of Nutrition 132 (2), 168–171.
Gedik, N., Kabasakal, L., Sehirli, O., Ercan, F., Sirvanci, S., Keyer-Uysal, M.,
Sener, G., 2005. Long-term administration of aqueous garlic extract (AGE)
alleviates liver fibrosis and oxidative damage induced by biliary obstruction
in rats. Life Sciences 76 (22), 2593–2606.
Halliwell, B., Gutteridge, J.M., Aruoma, O.I., 1987. The deoxyribose method: a
simple ‘‘test-tube’’ assay for determination of rate constants for reactions of
hydroxyl radicals. Analytical Biochemistry 165 (1), 215–219.
Ho, S.E., Ide, N., Lau, B.H., 2001. S-allyl cysteine reduces oxidant load in cells
involved in the atherogenic process. Phytomedicine 8 (1), 39–46.
Ide, N., Matsuura, H., Itakura, Y., 1996. Scavenging effect of aged garlic extract
and its constituents on active oxygen species. Phytotherapy Research 10,
340–341.
Ide, N., Nelson, A.B., Lau, B.H., 1997. Aged garlic extract and its constituents
inhibit Cu(2+)-induced oxidative modification of low density lipoprotein.
Planta Medica 63 (3), 263–264.
Khanum, F., Anilakumar, K.R., Viswanathan, K.R., 2004. Anticarcinogenic
properties of garlic: a review. Critical Reviews in Food Science and
Nutrition 44 (6), 479–488.
Kim, K.M., Chun, S.B., Koo, M.S., Choi, W.J., Kim, T.W., Kwon, Y.G.,
Chung, H.T., Billiar, T.R., Kim, Y.M., 2001. Differential regulation of NO
availability from macrophages and endothelial cells by the garlic
component S-allyl cysteine. Free Radical Biology & Medicine 30 (7),
747–756.
Koh, S.H., Kwon, H., Park, K.H., Ko, J.K., Kim, J.H., Hwang, M.S., Yum,
Y.N., Kim, O.H., Kim, J., Kim, H.T., Do, B.R., Kim, K.S., Kim, H.,
Roh, H., Yu, H.J., Jung, H.K., Kim, S.H., 2005. Protective effect of
diallyl disulfide on oxidative stress-injured neuronally differentiated
PC12 cells. Brain research. Molecular Brain Research 133 (2), 176–186.
Kourounakis, P.N., Rekka, E.A., 1991. Effect on active oxygen species of alliin
and Allium sativum (garlic) powder. Research Communications in
Chemical Pathology and Pharmacology 74 (2), 249–252.
Lawson, L.D., 1998. Garlic: a review of its medicinal effects and indicated
active compounds. In: Lawson, L.S., Bauer, R. (Eds.), Phytomedicines of
Europe: Chemistry and Biological Activity, ACS Symposium Series, 691.
American Chemical Society, Washington, D.C., pp. 176–209.
Lewin, G., Popov, I., 1994. Antioxidant effects of aqueous extract. 2nd
communication: inhibition of the Cu(2+)-initiated oxidation of low density
lipoproteins. Arzneimittel-Forschung 44 (5), 604–607.
Long, L.H., Evans, P.J., Halliwell, B., 1999. Hydrogen peroxide in
human urine: Implications for antioxidant defense and redox regu-
lation. Biochemical and Biophysical Research Communications 262 (3),
605–609.
Mukherjee, S., Banerjee, S.K., Maulik, M., Dinda, A.K., Talwar, K.K., Maulik,
S.K., 2002. Chronic garlic administration protects rat heart against
oxidative stress induced by ischemic reperfusion injury. BMC Pharmacol-
ogy 2 (1), 16 ([electronic resource]).
Ou, C.C., Tsao, S.M., Lin, M.C., Yin, M.C., 2003. Protective action on human
LDL against oxidation and glycation by four organosulfur compounds
derived from garlic. Lipids 38 (3), 219–224.
Pedraza-Chaverri, J., Tapia, E., Medina-Campos, O.N., de los Angeles
Granados, M., Franco, M., 1998. Garlic prevents hypertension induced
by chronic inhibition of nitric oxide synthesis. Life Sciences 62 (6),
PL71–PL77.
Pedraza-Chaverri, J., Maldonado, P.D., Medina-Campos, O.N., Olivares-
Corichi, I.M., Granados-Silvestre, M.A., Hernandez-Pando, R., Ibarra-
Rubio, M.E., 2000. Garlic ameliorates gentamicin nephrotoxicity:
relation to antioxidant enzymes. Free Radical Biology & Medicine 29
(7), 602–611.
Pedraza-Chaverri, J., Granados-Silvestre, M.D., Medina-Campos, O.N., Mal-
donado, P.D., Olivares-Corichi, I.M., Ibarra-Rubio, M.E., 2001. Post-
transcriptional control of catalase expression in garlic-treated rats. Molec-
ular and Cellular Biochemistry 216 (1–2), 9–19.
Pedraza-Chaverri, J., Gonzalez-Orozco, A.E., Maldonado, P.D., Barrera, D.,
Medina-Campos, O.N., Hernandez-Pando, R., 2003a. Diallyl disulfide
ameliorates gentamicin-induced oxidative stress and nephropathy in rats.
European Journal of Pharmacology 473 (1), 71–78.
Pedraza-Chaverri, J., Maldonado, P.D., Barrera, D., Ceron, A., Medina-
Campos, O.N., Hernandez-Pando, R., 2003b. Protective effect of diallyl
sulfide on oxidative stress and nephrotoxicity induced by gentamicin in rats.
Molecular and Cellular Biochemistry 254 (1–2), 125–130.
Pedraza-Chaverrı, J., Barrera, D., Maldonado, P.D., Chirino, Y.I., Macıas-
Ruvalcaba, N.A., Medina-Campos, O.N., Castro, L., Salcedo, M.I.,
Hernandez-Pando, R., 2004a. S-allylmercaptocysteine scavenges hydroxyl
radical and singlet oxygen in vitro and attenuates gentamicin-induced
oxidative and nitrosative stress and renal damage in vivo. BMC Clinical
Pharmacology 4 (1), 5 ([electronic resource]).
Pedraza-Chaverri, J., Gil-Ortiz, M., Albarran, G., Barbachano-Esparza, L.,
Menjivar, M., Medina-Campos, O.N., 2004b. Garlic’s ability to prevent in
vitro Cu2+-induced lipoprotein oxidation in human serum is preserved in
heated garlic: effect unrelated to Cu2+-chelation. Nutrition Journal 3 (1), 10
([electronic resource]).
Perez-Severiano, F., Rodriguez-Perez, M., Pedraza-Chaverri, J., Maldonado,
P.D., Medina-Campos, O.N., Ortiz-Plata, A., Sanchez-Garcia, A., Villeda-
Hernandez, J., Galvan-Arzate, S., Aguilera, P., Santamaria, A., 2004a. S-
Allylcysteine, a garlic-derived antioxidant, ameliorates quinolinic acid-
induced neurotoxicity and oxidative damage in rats. Neurochemistry
International 45 (8), 1175–1183.
Perez-Severiano, F., Salvatierra-Sanchez, R., Rodriguez-Perez, M., Cuevas-
Martinez, E.Y., Guevara, J., Limon, D., Maldonado, P.D., Medina-Campos,
O.N., Pedraza-Chaverri, J., Santamaria, A., 2004b. S-Allylcysteine prevents
amyloid-beta peptide-induced oxidative stress in rat hippocampus and
ameliorates learning deficits. European Journal of Pharmacology 489 (3),
197–202.
Prasad, K., Laxdal, V.A., Yu, M., Raney, B.L., 1995. Antioxidant activity of
allicin, an active principle in garlic. Molecular and Cellular Biochemistry
148 (2), 183–189.
Prasad, K., Laxdal, V.A., Yu, M., Raney, B.L., 1996. Evaluation of hydroxyl
radical-scavenging property of garlic. Molecular and Cellular Biochemistry
154 (1), 55–63.
Rahman, K., 2003. Garlic and aging: new insights into an old remedy. Ageing
Research Reviews 2 (1), 39–56.
Rietz, B., Isensee, H., Strobach, H., Makdessi, S., Jacob, R., 1993.
Cardioprotective actions of wild garlic (Allium ursinum) in ischemia and
reperfusion. Molecular and Cellular Biochemistry 119 (1–2), 143–150.
Salahudeen, A.K., Clark, E.C., Nath, K.A., 1991. Hydrogen peroxide-induced
renal injury. A protective role for pyruvate in vitro and in vivo. The Journal
of Clinical Investigation 88 (6), 1886–1893.
Sener, G., Satyroglu, H., Ozer Sehirli, A., Kacmaz, A., 2003. Protective effect
of aqueous garlic extract against oxidative organ damage in a rat model of
thermal injury. Life Sciences 73 (1), 81–91.
Sener, G., Sehirli, O., Ipci, Y., Ercan, F., Sirvanci, S., Gedik, N., Yegen, B.C.,
2005. Aqueous garlic extract alleviates ischaemia-reperfusion-induced
oxidative hepatic injury in rats. The Journal of Pharmacy and Pharmacology
57 (1), 145–150.
Shobana, S., Naidu, K.A., 2000. Antioxidant activity of selected Indian
spices. Prostaglandins, Leukotrienes, and Essential Fatty Acids 62 (2),
107–110.

J. Pedraza-Chaverrı et al. / Life Sciences 78 (2006) 761–770770
Song, K., Milner, J.A., 1999. Heating garlic inhibits its ability to suppress 7, 12-
dimethylbenz(a)anthracene-induced DNA adduct formation in rat mam-
mary tissue. The Journal of Nutrition 129 (3), 657–661.
Thabrew, M.I., Samarawickrema, N.A., Chandrasena, L.G., Jayasekera, S.,
2000. Protection by garlic against adriamycin induced alterations in the
oxido-reductive status of mouse red blood cells. Phytotherapy Research 14
(3), 215–217.
Torok, B., Belagyi, J., Rietz, B., Jacob, R., 1994. Effectiveness of garlic on the
radical activity in radical generating systems. Arzneimittel-Forschung 44
(5), 608–611.
Yin, M.C., Cheng, W.S., 1998a. Antioxidant activity of several Allium
members. Journal of Agricultural and Food Chemistry 46 (10), 4097–4101.
Yin, M.C., Cheng, W.S., 1998b. Inhibition of Aspergillus niger and
Aspergillus flavus by some herbs and spices. Journal of Food Protection
61 (1), 123–125.
Walker, P.D., Shah, S.V., 1988. Evidence suggesting a role for hydroxyl radical
in gentamicin-induced acute renal failure in rats. The Journal of Clinical
Investigation 81 (2), 334–341.
Copyright © 2022 FDOKUMEN




















