
Ultrastructure of in vitro oocyte maturation in buffalo ( Bubalus bubalis)

ELSEVIER
RATE OF TRANSPORT AND DEVELOPMENT OF PREIMPLANTATION EMBRYO IN THE SUPEROVULATED BUFFALO (Bubalus bubalis)
A.K. Misra, R. Kasiraj, M. Mutha RaoJ N.S. Rangareddy, R.S. Jaiswal and H.C. Pant
Sabarmati Ashram Gaushala, Lali-387120, Kaira, India 1Buffalo Breeding Centre, Nekarikallu- 522615, India
Received for publication : June 18, 1997 Accepted' Apr i l 22, 1998
ABSTRACT
This study was designed to ascertain the rate of transport and development of preimplantation embryo in the superovulated buffalo in order to determine the optimum time for their nonsurgical collection. Eighteen Murrah-type buffalo were superovulated with 600 mg NIH-FSH-P1. Luteolysis was induced by administration of PGF2ct at 72 (PG+72) and 84 h (PG+84) after initiating gonadotrophin treatment and fixed-time AI was done beginning at 36 h post PG+72 administration and at 12-h intervals thereafter, upto 72 h. Six control buffalo received treatment similar to experimental group except that in place of FSH they received normal saline. For embryo collection, experimental animals were humanely killed at 6-h intervals corresponding to 156 (n=2), 162 (n=2), 168 (n=2), 174 (n=3), 180 (n=3), 186 (n=3) and 192 h(n=3)after PG+72 treatment, whereas the control animals were humanely killed at 156 (n=2), 174 (n=2) and 192 h (n=2). Superovulated buffalo had higher number of ovulations than untreated controls (8.78±5.00 vs 0.67±0.51) and total ova/embryos recovered was 4.11±2.46 and 0.674-0.51, respectively. The high estradiol-1713 (E2) levels with its prolonged rise may, by leading to reverse peristalsis in the oviduct with a consequent loss of some embryos in the peritoneal cavity, be one of the reasons for our inability to recover nearly 84/158 ova/embryos in the superovulated buffalo. In superovulated animals, nearly all the ova/embryos reached the uterus between 168 and 174 h post PG+72 treatment or about 134 h (circa 5.5d) after the onset of superovulatory estrus, suggesting that the ideal time for non-surgical embryo collection in the buffalo is between Days 7 to 8 after PG+72 treatment or Days 5.5 to 6.0 of the superovulated cycle (estrus=Day 0). Embryo development of superovulated buffalo showed considerable variation as various stages of embryos (8 cell to expanded blastocyst) were recovered from the same donor buffalo, and the rate of development appeared to be 24 to 36 h faster than in cattle. © 1998 by Elsevier Science Inc.
Key words : buffalo, superovulation, embryo development and transport
Acknowledgments We thank Pro£ Maarten Drost, University of Florida, for a critical review of this manuscript. Thanks are also due to Dr. VM Mehta, Dean, College of Veterinary Science and Animal Husbandry, Anand, for providing facilities for hormone analysis. Financial assistance from the Department of Biotechnology, Govt. of India, under the National Science and Technology Project on Embryo Transfer, is gratefully acknowledged.
Theriogenology 50:637449, 1998 0093-69 tX/98/$19.00 © 1998 by Elsevier Science Inc. PII SO093-691X(98)OO168-X

638 Theriogenology
INTRODUCTION
Limited information concerning the transport rate of ova/embryos in superovulated buffalo suggests that ova/embryos remain in oviduct for varying lengths ranging between 74 and 100-h (29) and reach the uterus around Days 4 to 5 after the onset of estrus (13, 27, 28, 29). Additionally, few studies on the development of preimplantation embryo in superovulated buffalo indicate a faster rate of development than in cattle. Except for one report (29) suggesting 60 to 72-h faster rate of development of buffaio embryos, all other reports indicate 24 to 36-h faster rate of development (10, 16, 27, 38). The development of bubaiine embryos from compact morula stage to blastoeyst is reported to be very fast (13,29): however, there are conflicting reports regarding the hatching of blastoeyst: on Day 5 (29), Days 6 to 6.5 (38), Days 6 to 7 (1,16,49) and Days 7 to 7.5 (10). This is in contrast to the development of cattle embryos which hatch on Days 8 to 9 (30).
The transport rate of embryos is greatly influenced by the periovulatory steroid profile. Thus, understanding of estradiol-17f3 (E2) and progesterone (P4) levels during this period in relation to embryo recovery is of significance for the collection of most of the embryos from the donors.
In view of these conflicting claims the aim of this study was to assess the development and transport rate of embryos in superovulated buffaloes in order to determine the optimum time for nonsurgical collection of embryos. Since behavioral expression ofestrus in buffalo is weak, it is very difficult to judge the time of onset or duration of standing estrus in buffalo. In the present
study, an attempt was made to work out the time of embryo collection in relation to PGF2ot treatment during superovulation in buffalo.
MATERIALS AND METHODS
Twenty-four not lactating, multiparous, cyclic Murrah-type buffalo (Bubaius bubaiis), aged 6 to 7 yr and weighing between 450 and 550 kg, were used as donors. The animals were housed in shaded half-wail shed and were maintained under optimum management conditions. This experiment was conducted in the winter season (ambient temperature: 18-24°C) which is also the normal breeding season for buffalo. After balancing for age, weight and parity, the animals were divided into 2 groups of 6 (control) and 18 (experimental). Estrus was synchronized by injecting 2 doses each of 15 mg luprostiol (PGF2ot analogue, Prosolvin, Intervet, Holland) intramuscularly at an interval of 11 d (PG1 and PG2 treatments). A day before initiating superovulatory treatment between Days 9 and I l of the estrous cycle (estrus = Day 0), presence of a corpus luteum (CL) was ascertained in the ovary of each buffalo by rectal palpation. Commercially prepared 600 mg NIH-FSH-Pl(Folltropin, Vetrepharm, Canada) was used im for inducing superovulation in experimental animals in a twice-daily descending dose schedule for 5 d. Luteolysis was induced by administration of 15 mg, im luprostiol at 72 (PCJ+72) and 84 h (PG+84) after initiating gonadotrophin treatment. All animals were inseminated with frozen-thawed semen from fertile bulls at preset times beginning at 36 h post PG+72 administration and at 12-h interval thereafter upto 72 h. In the control group, all 6 animals were treated with normal saline (vehicle) in place

Theriogenology 639
of Folltropin between Days 9 and 11 of the synchronized estrus, ieceived PG+72 and PG+84 injections and were inseminated as per schedule described for the experimental group.
Animals of the experimental group were killed humanely at 6-h intervals corresponding to 156 (n=2), 162 (n=2), 168 (n=2), 174 (n=3), 180 (n=3), 186 (n=3) and 192 (n=3) h after PG+72 treatment, while the control animals were sacrificed at 156 (n=2), 174 (n=2) and 192 (n=2) h. Immediately after slaughter, genital organs (including ovaries) were removed and transported to the laboratory in a tray containing normal saline. Ovaries were examined for the presence of actual number of CL and anovulatory follicles > 10 mm in diameter. Various parts of the genital organs were classified as: proximal oviduct (half of the oviduct towards infundibulum), distal oviduct (half of the oviduct next to the uterus), proximal uterine horn (half of the uterine horn towards oviduct) and distal uterine horn (half of the uterine horn towards body of uterus). For embryo collection, the utero-tubul junction of each uterine horn was ligated, and the proximal portion of each horn was flushed by transcervical introduction of a 20-g Foley catheter stiffened with a styler, and the balloon of the catheter was located equidistant from the uterine body and utero-tubal junction. The balloon was inflated with air, and the proximal horn was initially flushed with 20 mL of the flushing medium (Dulbecco's Phosphate Buffered Saline; D-PBS; Sigma, St.Louis, MO, USA) supplemented with 0.1% bovine serum albumin (BSA, fraction V, Sigma) and later with an additional 60 mL of flushing medium. After completing flushing of the proximal horn, the catheter balloon was deflated and the catheter retracted to ensure placement of the balloon at the intercomual junction to flush the distal horn. Simultaneously, the proximal horn was clamped with the help of 8 inch long artery forceps placed just ahead of the balloon for flushing the proximal horn; then the distal horn was flushed similarly to the proximal. Each oviduct was ligated in the middle and then cut just proximal to the ligature. Each proximal oviduct was then flushed by introducing the flushing medium (20 mL) into the ampulla through the ostium with a blunt needle connected to a syringe. The ligature was then opened, and each distal oviduct was flushed by inserting the blunt needle at this end. The washings for the distal oviduct were collected from the utero-tubal junction, which was cut open just prior to flushing. Effluent of each portion of the uterine horns and oviducts was transferred into separate 100-x 20-mm flat-bottom grid dish (Becton and Dickinson, New Jersey, USA). Embryos were isolated using a stereozoom microscope (SMZ-2T, Nikon, Japan) at x 20 to 30 magnification. The recovered embryos were transferred to a small Petridish (35-x10-mm, Becton and Dickinson) containing holding medium (D-PBS with 0.4% BSA) using a micropipette. The embryos were evaluated at x 60 magnification and classified as 8-cell, 16-ce11, early morula, morula, compact morula, early blastocyst, blastocyst, expanded blastocyst, hatched blastocyst, retarded embryo and degenerated embryo, unfertilized ovum and empty zona, or zona pieces.
Samples of jugular venous blood were collected from 6 of 18 superovulated buffalo for measurement of plasma estradiol-1715 (E2) and progesterone (P4). An indwelling catheter was inserted into the jugular vein on the day before sampling began (37). Blood samples were collected at 24 h post PG+72 administration and at 8-h intervals thereafter, upto 72 h and then at 12-h intervals upto 144 h. All blood samples (circa 10 mL) were collected into heparinized tubes (6 to 10 units of heparin/mL blood), centrifuged immediately for 10 min at 1000 x g, and the plasma stored at -20 °C until assayed.

640 Theriogenology
Plasma E2 was analyzed by a direct double-antibody 1251 radioimmunoassay method (Diagnostic Products Corporation, Los Angeles, CA, USA), according to the manufacturer after validation in the laboratory. The standard curve was linear in the range of 5 to 500 pg/mL. The assay sensitivity was 1.4 pg/mL. The intra- and inter-assay coefficients of variation were 8.5 and 15.1%, respectively.
Plasma P4 was analyzed by a direct solid-phase 125I radioimmunoassay method (Coat-A-Count TKPG, Diagnostic Products Corporation), in 100-~L samples in duplicate, according to the manufacturer after validation in the laboratory. The standard curve was linear in the range of 0.1 to 40ng/mL. The assay sensitivity, defined as twice the standard deviation from the buffer control, was 0.05ng/mL. The intra- and inter-assay coefficients of variation were 7.8 and 11.2%, respectively.
The differences in the total and viable embryo recovery due to differing endocrine profile were assessed by Chi-square test (50). Correlation analysis was used to determine the relationship between circulating concentration of E2 and the E2:P4 ratio with recovery of total embryos.
RESULTS
The 18 superovulated buffalo had a total of 158 ovulations; however, only 74 ova/embryos (46.8%) were recovered. The right ovary had more ovulations than the left ovary. Accordingly, more (P > 0.025) embryos were collected from right side of the genital tract than the iett (Table 1). Of the control animals only 4 ovulated, and all 4 unfertilized ova were recovered.
Tablel. Superovulatory response and embryo recovery in buffalo slaughtered between 156 and 192 hours after PG+72 treatment Mean ± SD).
Attributes Experimental group
No. of animals
No. of ovulations (CL)
Ovulations in :
right ovary
left ovary
Anovulatory follicles
Total ova / embryos recovered
Ova / embryos recovered from
oviduct
uterus
Ova / embryos recovered from
right side of genital tract
left side of genital tract
18
8.78 + 5.00
4.83 ± 2.54
3.94 ± 3.60
2.11 ±2.07
4.11 + 2.46
0.83 + 1.56
3.28 ± 2.67
2.72 ± 1.78
1.39 ± 1.56
Theriogenology 641
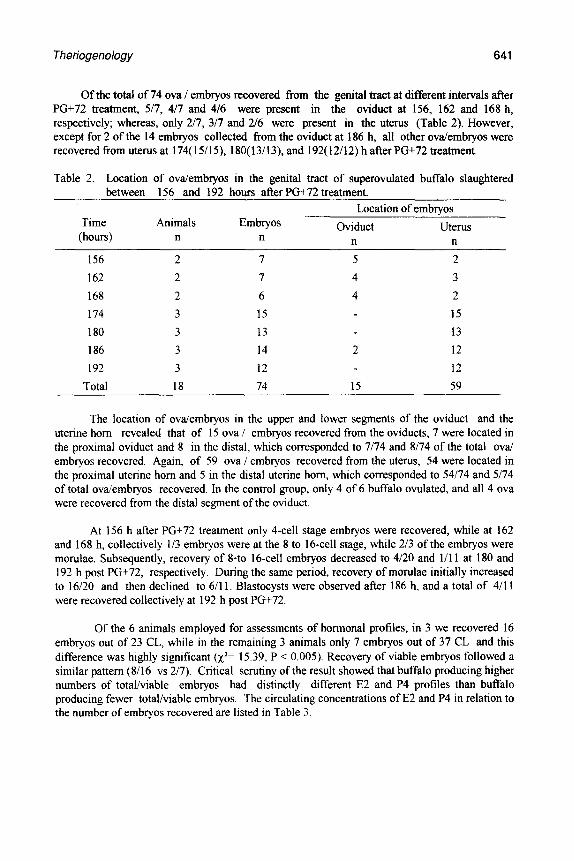
Of the total of 74 ova / embryos recovered from the genital tract at different intervals after PG+72 treatment, 5/7, 4/7 and 4/6 were present in the oviduct at 156, 162 and 168 h, respectively; whereas, only 2/7, 3/7 and 2/6 were present in the uterus (Table 2). However, except for 2 of the 14 embryos collected from the oviduct at 186 h, all other ova/embryos were recovered from uterus at 174(15/15), 180(13/13), and 192(12/12) h after PG+72 treatment.
Table 2. Location of ova/embryos in the genital tract of superovulated buffalo slaughtered between 156 and 192 hours after PG+72 treatment.
Location of embryos Time Animals Embryos Oviduct Uterus (hours) n n n n
156 2 7 5 2
162 2 7 4 3
168 2 6 4 2
174 3 15 15
t80 3 13 13
186 3 14 2 12
192 3 12 12
Total 18 74 15 59
The location of ova/embryos in the upper and lower segments of the oviduct and the uterine horn revealed that of 15 ova / embryos recovered from the oviducts, 7 were located in the proximal oviduct and 8 in the distal, which corresponded to 7/74 and 8/74 of the total ova/ embryos recovered. Again, of 59 ova / embryos recovered from the uterus, 54 were located in the proximal uterine horn and 5 in the distal uterine horn, which corresponded to 54/74 and 5/74 of total ova/embryos recovered. In the control group, only 4 of 6 buffalo ovulated, and all 4 ova were recovered from the distal segment of the oviduct.
At 156 h after PG+72 treatment only 4-cell stage embryos were recovered, while at 162 and 168 h, collectively 1/3 embryos were at the 8 to 16-cell stage, while 2/3 of the embryos were morulae. Subsequently, recovery of 8-to 16-cell embryos decreased to 4/20 and 1/11 at 180 and 192 h post PG+72, respectively. During the same period, recovery ofmorulae initially increased to 16/20 and then declined to 6/11. Blastocysts were observed after 186 h, and a total of 4/1l were recovered collectively at 192 h post PG+72.
Of the 6 animals employed for assessments of hormonal profiles, in 3 we recovered 16 embryos out of 23 CL, while in the remaining 3 animals only 7 embryos out of 37 CL and this difference was highly significant C( 2= 15.39, P < 0.005). Recovery of viable embryos followed a similar pattern (8/16 vs 2/7). Critical scrutiny of the result showed that buffalo producing higher numbers of total/viable embryos had distinctly different E2 and P4 profiles than buffalo producing fewer total/viable embryos. The circulating concentrations of E2 and P4 in relation to the number of embryos recovered are listed in Table 3.

642 Thefiogenology
Table 3. Mean (±S.D.) concentrations of progesterone and estradiol-1713 in the jugular venous plasma of superovulated buffalo related to embryo recovery.
More embryos recovered (16/23) a Fewer embryos recovered (7/37) a Hours Day of pest cycle
PG+72 Progesterone Estradiol- 17[~ Progesterone Estradiol- 1713 (ng/mL) (pg/mL) (ng/mL) (pg/mL)
24 -1 0.36±0.12 14.00±8.65 0.43±0.16 23.33±8.49
32 -1 0.28±0.02 16.60-a:10.58 0.30±0.03 27.00±6.07
40 0 0.16±0.10 25.33±8.08 0.25±0.02 44.33±26.53
48 0 0.26+0.19 15.47±14.56 0.24±0.08 48.00±20.27
56 1 0.42±0.14 10.67+6.02 0.21±0.08 34.67±1.16
64 1 0.60±0.19 7.13±4.22 0.30i0.21 24.00±7.19
72 2 0 .78-q :0 .19 8 . 0 0 - ± 5 . 1 9 0.26±0.08 14.00~7.80
84 2 0.74±0.15 10.10±6.02 0.36±0.14 11.27±5.93
96 3 i .69±0.93 8.73±6.24 0.68±0.45 12.33±7.49
108 3 2.12±1.54 10.73±10.60 0.93±0.43 9.80i-6.26
120 4 2.57±1.23 5.37±5.74 ! .70±0.65 8.63±4.89
132 4 2.93±1.05 4.73±3.73 2.33±0.97 5.60±1.90
144 5 3.13±!.37 7.83±5.05 3.10±1.85 9.93±6.08
a = embryos/CL.
It is apparent that in the 3 buffalo producing more embryos, the mean E2 concentration was 14.00±8.65 pg/mL plasma at 24 h after PG+72 treatment. The value thereafter increased to the peak level of 25.33±8.08 pg/mL at 40 h, followed by a sharp decline to less than 10.0 pg/mL by 64 h. Duration of the rise in circulating E2 concentration considered as the interval between the values above 10.0 pg/mL was about 32 h. Contrarily, the remaining 3 buffalo which produced fewer total embryos had very high E2 levels (23.33±8.49 pg/mL) at 24 h post PG+72 and increased sharply to the peak value of 48.00±20.27 pg/mL at 48-h. The E2 concentration also remained high for a longer duration (circa 72 h) and returned very slowly to less than 10.0 pg/mL plasma at 108 h post PG+72. Apparently, both the peak E2 concentration and the duration of its rise were nearly double the corresponding values observed in animals which produced more total/viable embryos.

Theriogenology 643
2 6 o
2oo
0 1so
~" 100
M o r e to t=H/v iab le e m b r y o == --I-- LOllS t o t a l / v l a b l o e m b r y o l
5 0 -
o i n I n I I 1 r ~ " ~ - ~ ' 2 4 3 2 4 0 4 8 5 6 6 4 7 2 8 4 0 6 1 0 8 1 2 0 1 3 2 1 4 4
Hours post P G + 7 2
Figure 1. OeslTadiol-17B to progesterone ratio in the jugular venous plasma of superovulated buffalo in relation to recovew of total/viable embryos
The mean circulating concentration of P4 in the 3 buffalo which produced a high number of total and viable embryos increased from basal level of 0.16-~0.10 ng/mL at 40 h after PG+72 administration to 0.42±0.14 ng/mL at 56 h. Thereafter, the concentrations continued to increase sharply, reaching 1.69±0.93, 2.12+1.54 and 2.57±1.23 ng/mL at 96, 108 and 120 h, respectively (Table 3). In the remaining 3 buffalo which produced fewer number of total and viable embryos, the mean P4 concentration did not show any characteristic rise for upto 84 h after PG+72 (<0.36±0.14 ng/mL) treatment. Thereafter, the concentration increased gradually to 0.68-~0.45, 0.93~=0.43 and 1.70~:0.65 at 96, 108 and 120 h, respectively, suggesting a slower rate of P4 increase in these animals.
Comparison of the E2:P4 ratio revealed that except at 40 h post PG+72, the ratio was lower in buffalo which produced more embryos at all other intervals (Figure 1). A significant negative correlation was observed between the percentage of the total embryo recovery and plasma E2 concentration (r = - 0.87, P <0.05) and the E2:P4 ratio (r = - 0.87, P < 0.05) on Day 1 after superovulatory ¢strus.
DISCUSSION
The superovulatory response in this study (8.78+5.00) was similar (mean, 8.8) to that reported earlier (28) and was nearly twice that of the means of 3.6 to 4.3 ovulations reportod by others (1,2,14,26,31,48). This may be attributed to the greater purity of the FSH preparation employed (only 12.5% LH impurity), the induction of superovulation during the normal breeding season, and the good condition of the donors. In cattle, the superovulatory response is affectod by the impurity of FSH (21,33,34), plane of nutrition (41 ) and season (6,25).

644 Theriogenology
The anovulatory follicles in our study (2.11~:2.07) may have indicated an inadequate endogenous LH surge (47,53), However, as the administration of GnRH at the superovulatory estrus has failed to decrease anovulatory follicles in buffalo (38,52) and cattle (46,57), it remains to be determined if some of these follicles were biochemically aberrant (8).
Mean recovery of 4.11±2.46 total ova/embryos in our experiment was more than in earlier reports in swamp (9) and riverine (28) buffalo, despite the fact that in the later study the animals were slaughtered after superovulation. In our study, even after thorough flushing of the oviducts and uterine horns, only 74/158 ova/embryos were recovered. Others have reported even lower (9/44 to 22/67) (28) or only slightly higher (26/46) rates of (55) ova/embryo recovery in buffalo slaughtered after superovulation. Similar studies in superovulated cattle have also reported recovery rates of 22 to 63% for ova/embryos after slaughter (4, 21,36). Loss of nearly 84/158 of ova/embryos in our study may be due to one or more of the several factors that are known to be associated with poor recovery of ova/embryos in superovulated cattle and buffalo. These factors include inability of fimbria to trap ova from enlarged superovulated ovary (54), difficulties in locating hatched blastocysts (1) and premature entry of ova/embryos into the uterus resulting into their expulsion (28, 43).
In our study, while a part of this loss could be due to repeated manual handling of the reproductive tract (4 times over 48 h) for AI as well as for flushing of the uterus in 2 sections, failure of the fimbria to pick up the ova/embryos when ovulations coincide with very high E2 levels or a high E2 and P4 ratio may also have been an important contributory factor. This suggestion is based on our results that in the buffalo with greater concentrations and longer durations of a rise in circulating E2, lower concentrations of P4 and high E2/P4 ratios after PG+72 administration, the ova/embryo recovery rate was much lower (7/37) than in animals with low concentrations of E2, shorter durations in the E2 rise and lower E2/P4 ratios (16/23). This conclusion is further substantiated by significant (P < 0.05) negative correlations between the E2 concentration and between the E2:P4 ratio on Day 1 after superovulatory estrus and total embryo recovery.
The method of ovum pick-up and transport has not been studied in the buffalo, and it is also not well understood in cattle (19); however, the flow of tubal fluid (and muscle contractions) near the time of ovulation has been shown to be in the opposite direction of oocyte transport (51). Thus perhaps in buffalo with high E2 levels, a prolonged duration of rise in E2 and a high E2/P4 ratio some of the oocytes may not have been either picked up or may have been lost in the peritoneal cavity as a consequence of reversed peristalsis in the oviduct. According to Beilve and McDonald (3), in sheep at estrus, the bulk of the oviduct fluid passes from the ampulla into the peritoneal cavity, and this phenomenon may be an important factor contributing to the poor recovery of ova/embryos in superovulated buffalo. Further studies are warranted on ova/embryo transport in cyclic and superovulated buffalo in order to clarify the physiological mechanisms involved. These results are the first that we are aware of to demonstrate the effect of high E2 levels and or high E2:P4 ratio at the superovulatory estrus on the total embryo recovery rate in the buffalo.

Theriogenology 645
Nearly all the ova/embryos reached the uterus between 168 and 174 h post PG+72 treatment or about 134 h (circa 5.5 days) after the onset of superovulatory estrus. These observations are different from those of other researchers (1, 13), who reported the descent of the bubaline embryo into the uterus occurs around Day 4 (estrus = Day 0) but are similar to an earlier Bulgarian (28) and a recent Chinese report (55). Our results suggest that the ideal time for nonsurgical embryo collection in the buffalo is between Days 7 to 8 after PG+72 treatment or Days 5.5 to 6.0 of .the superovulated cycle (estrus = Day 0). Earlier investigators collected buffalo embryos on Day 7 (17) or between Days 5.5 to 7 (9) of the superovulated cycle. However, in subsequent studies nonsurgical collections performed around Day 5.5 have produced comparatively better recovery rates (11,38,39,42,47). In this respect, the buffhlo seems to be different from superovulated cattle, as in the latter species the ideal time for nonsurgical collection is considered to be between Days 6 and 8 after superovulatory estrus (15, 20, 22). Recovery of 2 embryos from oviduct at 186-h post PG+72 suggests that a small percentage of ova may remain in the oviducts of superovulated buffalo for a considerable time, as occurs in caRRie, in which a small percentage of ova/embryos were recovered on Day 7 or on Days 10 to 16 after superovulatory estrus.
Recovery of all 4 ova from the oviduct of control animals until 192 h post PG+72 suggests that the transport rate of ova/embryos in superovulated buffalo is hastened, as it is in cattle (18,24,43). In superovulated cattle this has been attributed to a higher blood P4 concentration at estrus (12) and a rapid rise in P4 secretion (7, 44, 45). In our study an earlier rise in P4 secretion in the superovulated cycle (after PG+72) in comparison with that of the nonsuperovulated cycle (after PG2) may have hastened tubal transport rate (37). However, the role of P4 and E2 on tubal transport rate of buffalo ova needs further study.
Our results indicate that embryo development in superovulated buffalo varies considerably as in cattle (40). Similarly to earlier reports (10, 29), we recovered various stages of embryos (8 cell to expanded blastocyst) from the same donor buffalo. In superovulated cattle, onsiderable time gap between the first and the last ovulation, ranging from 12 h (58) to 48 h (56), an altered tubal and uterine environment due to higher circulating concentrations of E2 and P4 (35), and changes in embryo quality (23) account for the variation in embryo development rates. Arguably, these factors may also be operative in superovulated buffalo. We have observed that a circulating concentration of E2 and duration of E2 rise were considerably higher in the superovulated cycle (37), and this may have affected the development of early embryos (47).
Our study further suggests that most of the early stage (4 cell) embryos eveloped to the 8-to 16-cell stage or to early morula stage by 168- h post PG+72. Further, by 180-h, most of the 8-tol6-cell embryos grew to morula or compact morula stage (total morula 7/9). Subsequently, recovery of 4/11 blastoeysts by 192 h indicates further development of morula to the blastocyst stage, as the recovery of morulae decreased from 16/20 to 6/11. Though zona pellucida pieces were recovered between 156 to 186 h post PG+72, we failed to recover even a single hatched blastocyst. This pattern of development of buffalo embryos closely resembles that in earlier reports, indicating 24 to 36 h faster growth of buffalo embryos than cattle (10, 16o 27, 38) embryos and differs with the reported hatching of all embryos after 117 h from the onset of estrus (29).

646 Thefiogenology
REFERENCES
1. Alexiev A, Vlahov K, Karaivanov Ch, Kacheva D, Polykhronov O, Petrov M, Nikolov N, Drogoev A and Radev P. Embryo transfer in buffaloes in Bulgari&Proc II World buffalo Congress 1988; II:591-595.
2. Ambrose JD, Singla SK, Jailkhani S, Prakash BS and Madan ML. Superovulation response in buffaloes (Bubalus bubalis) to different treatment regimens of Folltropin. Theriogenology 1991; 35:181 abstr.
3. Bellve A R, McDonald M F. Directional flow of fallopian tube secretion in the ewe at onset of the breeding season. J Reprod Fertil 1970; 22: 147-149.
4. Betteridge KJ (ed). Embryo Transfer in Farm Animals : A Review of Techniques and Applications. Canada Department of Agriculture Monograph 1977.
5. Betteridge K J, Eaglesome MD, Randall GCB, Mitchell D. Collection, description and transfer of embryos from cattle 10-16 days after estrus. J Reprod Fertil 1980; 59: 205-216.
6. Boland MP, Gordon I, McGovem H, Lynn G. Superovulatory response of cows and heifers in summer in Saudi Arabia. Theriogenology 1988; 29:227 abstr.
7. Booth WD, Newcomb R, Strange H, Rowson LEA, Sacher HB. Plasma oestrogen and progesterone in relation to superovulation and egg recovery in the cow. Vet Rec 1975; 97: 366-369.
8. Callesen H, Greve T, Hyttel P. Preovulatory endocrinology and oocyt¢ maturation in superovulated cattle. Theriogenology 1986; 25: 71-86.
9. Chantaraprateep P, Lohachit C, Kobayashi G, Techkumphu M, Vir~ul P, Kunavongla'it A, Prateep P. Ovarian responses to gonado-trophin stimulation in swamp buffalo (Bubalus bubalis). Buffalo Bull 1988; 7 : 82-86.
10. Chantaraprateep P, Lohachit C, Techkumphu M, Kobayashi G, Virakul P, Kunayongkrit A, Prateep P, Limskul A. Early embryonic development in Thai Swamp buffalo (Bubalus bubalis). Theriogenology 1989; 31: 1131- 1139.
11. Cruz LC, Venturina H, Jha SS, Adriano F, Serra P, Duran PG, Smith OF, Lorenzo N, Faylon PS. Successful transfer of Murrah buffalo embryo into Phillipine Swamp buffalo recipients. Proc lIIrd World Buffalo Congress 1991; Ill : 586-590.
12. Donaldson LE, Ward DN, Glenn SD. Use of porcine follicle stimulating hormone ai~r chromatographic purification in superovulation of cattle. Theriogenology 1986; 25: 747-757.
13. Drost M Training Manual for Embryo Transfer in Water Buffaloes. FAO Animal Production and Health Paper 84, Food and Agriculture Organization of the United Nations, Rome, 1991.
14. Drost M, Alexiev A, Vlahov K, Karaivanov Ch, Cripe WS, Lconards AP, Kacheva D, Polihronov O, Nicolov N, Petrov M, Dragoev A. Successful nonsurgical embryo transfer in buffalo (Bubalus bubalis) in Bulgaria. Tberiogenology 1988; 30: 659-668.
15. Drost M, Brand A, Aarts MH. A device for nonsurgical recovery of bovine embryos. Theriogenology 1976; 6: 503-508.
16. Drost M and Elsden R P. Blastocyst development in the water buffalo (Bubalus bubalis). Theriogenology 1985; 23:191 abstr.
17. Drost M, Wright Jr JM, Cripe WS, Richter AR. Embryo transfer in water buffalo (Bubalus bubalis). Theriogenoiogy 1983; 20: 579- 584.
18. EI-Bana AA, Hafez ESE. Egg transport in beef cattle. J Anita Sci 1970; 30: 430-432.

Theriogenology 647
19. Ellington JE, The bovine oviduct and its role in reproduction : a review of the literature. Comell Vet 1991; 81: 313-328.
20. Elsden RP, Hailer JF, Seidel GE Jr. Nonsurgical recovery of bovine eggs. Theriogenology 1976; 6: 523-532.
21. Gonzales A, Lussier JG, Carruthers TD, Murphy BD and Mapletoft, RJ. Superovulation of beef heifers with Folltropin : a new FSH preparation containing reduced LH activity. Theriogenology 1990; 33: 519-529.
22. Greve T. (ed), Bovine Egg Transplantation in Denmark. Copenhagen : Carl Fr Mortensen, 1981.
23. Greve T, Callesan H, Hyttel P, Endocrine profiles and egg quality in the superovulated cow. Nord Vet-Med 1983; 35: 408-421.
24. Hafez ESE, Sugie T, Gordon I. Superovulation and related phenomena in the beef cow. I. superovulatory responses following PMSG and HCG injection. J Reprod Fertil 1963; 5: 359-379.
25. Hasler JF, McCauley AD, Schermerhorn EC, Foote RH. Superovulatory response of Holstein cows. Tberiogenology 1983; 19: 83-99.
26. Jain GC, Khanna S, Chopra SC. Efficacy of various superovulatory protocols on superovulation and embryo recovery in buffaloes. In : Seminar on Embryo Transfer and Associated Technologies in Livestock Improvement, Present Status and Future Needs, Indian Veterinary Research Institute, Izatnagar, 1992.
27. Jainudeen MR. Embryo transfer technology in the buffalo : a review. Presented to FAO / LAEA / ACIAR Research Coordination and Planning Meeting on Buffalo Productivity, CSIRO, Rockhampton, Queensland, Australia, 1989.
28. Karaivanov C, Kacbeva D, Petrov M, Vlahov K, Sapundjiev E. Superovutatory response of river buffalo (Bubalus bubalis). Theriogenology 1990; 33: 453-464.
29. Karaivanov C, Vlahov K, Petrov M, Kacheva D, Stojanova M, Alexiev A, Polihronov O, Danev A. Studies on preimplantation development of buffalo embryos. Theriogenology 1987; 28: 747-753.
30. Linder GM, Wright RW Jr. Bovine embryo morphology and evaluation. Theriogenology 1983; 20:407-416.
31. Madan ML, Singla SK, Singh C, Prakash BS, Jailkhani S. Embryo transfer technology in buffaloes : Endocrine responses and limitations. Proc II World Buffalo Congress New Delhi, 1988; III:195 - 211.
32. Mapletoft RJ. Bovine embryo transfer. In : Morrow DA (ed), Current Therapy in Theriogenology. Philadelphia : WE Saunders Co,1986; 54-58.
33. Mapletoft RJ, Murphy BD. Superovulation in the cow: effects of biological activity of gonadotrophins. International Workshop on Advanced Reproductive Technologies, Western College of Veterinary Medicine, University of Saskatchewan, Canada, 1993; 1-21.
34. Mapletoft R J, Wu M, Murphy BD. Superovulation of beef cattle with Folltropin. Proc 11 th lnt Cong Anim Reprod Artif Insem, 1988b; 173 abstr.
35. Maurer RR, Ecbternkamp SE. Hormonal asynchrony and embryonic development Theriogenology 1982; 17:11-22.
36. Mc Gowan MR, Braithwaite M, Jochle W, Mapletoft RJ. Superovulation of beef heifers with Pergonal (HMG): a dose response trial. Theriogenology 1985; 24: 173-184.

648 Theriogenology
37. Misra AK. Studies on Improving Certain Aspects of Embryo Biotechnology in Buffalo. PhD thesis, CS Azad University of Agriculture and Technology, Kanpur, 1996.
38. Misra AK, Joshi BV, Agrawala PL, Kasiraj R, Sivaiah S, Rangareddi NS, Siddiqui MU. Multiple ovulation and embryo transfer in Indian buffaloes (Bubalus bubalis). Theriogenology 1990; 33:1131-1142.
39. Misra AK, Joshi BV, Kasiraj R, Sivaiah S, Rangareddi NS. Improved superovulatory regimen for buffalo. Theriogenology 1991; 35:245 abstr.
40. Moor NW. The control of time of estrus and ovulation and the induction of superovulation in cattle. Aust J Agri Res 1975; 26: 295-304.
41. Murphy MG, Boland MP, Roche JF. Pattern of follicular growth and resumption of ovarian activity in post-partum beef suckler cows. J Reprod Fertil 1990; 90: 523-533.
42. Mutha Rao M, Kasiraj R, Rangareddy NS, Jaiswal RS, Misra AK. A study on repeated superovulations and nonsurgical recovery of embryo in buffaloes. Proc IVth World Buffalo Congress, 1994; III: 498-500.
43. Newcomb R, Rowson LEA, Trounson AO. The entry of superovulated eggs into the uterus. In: Rowson LEA (ed), Egg Transfer in Cattle. Luxembourg: Commission European Communities, 1976.
44. Saumande J. Concentrations of luteinizing hormone, oestradiol-I 7 beta and progesterone in the plasma of heifers treated to induce superovulation. J Endocrinol 1980; 84: 425- 437.
45. Saumande J, Tamboura D, Chupin D. Changes in milk and plasma concentration of progesterone in cows after treatment to induce superovulation and their relationship with number of ovulation and of embryos collected. Theriogenology 1985; 23: 719-731.
46. Savage NC, Howell W, Mapletoft RJ. Superovulation in the cow using estradiol-17b or GnRH in conjunction with FSH-P. Theriogenology 1987; 27:383-395.
47. Schallenberger E, Wagner HG, Papa R, Hartl P, Tenhumberg H. Endocrinological evaluation of the induction of superovulation with PMSG in water buffalo. (Bubalus bubalis) Theriogenology 1990; 34: 379-392.
48. Sharifuddin W, Jainudeen MR. Superovulation and nonsurgical collection of ova in the water buffalo (Bubalus bubalis). Proc X Int Congr Anim Reprod Artif Insem, 1984; lI: 240-242.
49. Singla SK, Madan ML, Manik RS, Ambrose JD, Chauhan MS. Fertilization and early embryo development pattern in superovulated buffaloes. Proc 12th Int Congr Anita Reprod 1992; 2: 817-819.
50. Steel RGD, Torrie JH. Principles and Procedures of Statistics. New York : Mc Graw Hill Book Co, 1960.
51. Stone SL, Hammer CD, Biochemistry and physiology of oviductal secretions. Gynecol Invest 1975; 6: 234-252.
52. Taneja M, Totey SM, Pawshe CH, Singh G, Ali A, Talwar GP. Ultrasonographic monitoring of ovarian follicular dynamics in superovulated buffaloes. Proc of the III World Buffalo Congress 1991;III:581-585.
53. Taneja VK, Nanda SK, Datta TK, Bhat PN. Embryo transfer in buffaloes : present status and future needs. Proc of II World Buffalo Congress, 1988;II: 603-609.
54. Ullah N, Wright RW Jr, Mehmood A, Baig SM. Endocrine profile in relation to ovarian response, recovery rate and quality of embryos in Nili-Ravi buffaloes treated with FSH. Buffalo J 1992; I: 47-56.

Theriogenology 649
55. Wang ZK, Wu TG, Yu DQ, Wang Z, Wang KH, Lei C, Lui ZH. Effect of LH on superovulation of Swamp buffalo in China. Theriogenology 1994; 41:331 abstr.
56. Wise T, Vernon M, Maurer RR. Endocrine and ionic changes in preovulatory bovine follicles, J Anim Sci 1985; 61 (Suppl.1):368 abstr.
57. Wubishet A, Graves CN, Spahr SL, Kesler D J, Favero RJ. Effects of GnRH treatment on superovulatory responses of dairy cows. Theriogenology 1986; 25:423-427.
58. Yadav MC, Walton JS, Laslie KE. Timing of the onset and duration of ovulation in superovulated beef heifers. Theriogenology 1986b; 26: 509-521.
Copyright © 2022 FDOKUMEN




















