
The organic osmolytes betaine and proline are transported by a shared system in early...
12
JOURNAL OF CELLULAR PHYSIOLOGY 210:266–277 (2007) The Organic Osmolytes Betaine and Proline Are Transported by a Shared System in Early Preimplantation Mouse Embryos MOHAMED-KHEIR IDRIS ANAS, 1 MARY-ANNE HAMMER, 1 MICHAEL LEVER, 4 JO-ANN L. STANTON, 5 AND JAY M. BALTZ 1,2,3 * 1 Hormones, Growth and Development Program, Ottawa Health Research Institute, Ottawa, Ontario, Canada 2 Departments of Obstetrics and Gynecology (Division of Reproductive Medicine), University of Ottawa, Ottawa, Ontario, Canada 3 Departments of Cellular and Molecular Medicine, University of Ottawa, Ottawa, Ontario, Canada 4 Biochemistry Unit, Canterbury Health Laboratories, Christchurch, New Zealand 5 Department of Anatomy and Structural Biology, University of Otago Medical School, Dunedin, New Zealand Betaine and proline protect preimplantation mouse embryos against increased osmolarity and decreased cell volume, implying that they may function as organic osmolytes. However, the transport system(s) that mediates their accumulation in fertilized eggs and early embryos was unknown, and previously identified mammalian organic osmolyte transporters could not account for their transport. Here, we report that there is a single saturable transport component shared by betaine and proline in 1-cell mouse embryos. A series of inhibitors had nearly identical effects on both betaine and proline transport by this system. In addition, K i values for reciprocal inhibition of betaine and proline transport were 100–300 mM, similar to K m values (200–300 mM) for their transport, and both had similar maximal transport rates (V max ). The K i values for inhibition of betaine and proline transport by dimethylglycine were similar (2 mM), further supporting transport of both substrates by a single transport system. Finally, betaine and proline transport each required Na þ - and Cl . These data were consistent with a single, Na þ - and Cl -requiring, betaine/proline transport system in 1-cell mouse embryos. While betaine was only transported by a single saturable system, we found an additional, less conspicuous proline transport route that was betaine-insensitive, Na þ -sensitive, and inhibited by alanine, leucine, cysteine, and methionine. Furthermore, we showed that betaine, like proline, is present in the mouse oviduct and thus could serve as a physiological substrate. Finally, accumulation of both betaine and proline increased with increasing osmolarity, consistent with a possible role as organic osmolytes in early embryos. J. Cell. Physiol. 210: 266 – 277, 2007. ß 2006 Wiley-Liss, Inc. Regulation of cell volume is a fundamental property of cells that animal cells accomplish by regulating intracellular osmolyte concentrations. Acute volume perturbations are alleviated by quickly adjusting in- organic ion concentrations (Hallows and Knauf, 1994), but cells that are normally subject to high osmolarity instead require ‘‘organic osmolytes,’’ small uncharged organic compounds, to provide long-term intracellular osmotic support (Yancey et al., 1982; Garcia-Perez and Burg, 1991; Lang et al., 1998). This avoids the severe metabolic disruption caused by the substantially increased intracellular ionic strength that would be required to balance high osmolarity using inorganic ions. While some organic osmolytes are generated intracellularly, most are imported from the external environment by specific transporters. In mammals, four organic osmolyte transporters, all of which are stimu- lated by hypertonicity, have previously been identified. These are the Na þ /myo-inositol transporter (SMIT), taurine/b-amino acid transporter (TAUT), betaine/ GABA transporter (BGT1), and System A amino acid transporter (ATA2) (Garcia-Perez and Burg, 1991; Kwon and Handler, 1995; Pastor-Anglada et al., 1996; Alfieri et al., 2001). The requirement for organic osmolytes, and for these organic osmolyte transporters, is well established in cells subjected to hypertonicity, such as in the kidney. However, a range of cells not normally subject to osmotic stress also use organic osmolytes (Lang et al., 1998), and thus organic osmo- lytes and osmolyte transporters are likely to function in ‘‘housekeeping’’ cell volume regulation as well as in response to hypertonic conditions. There is strong evidence that mammalian embryos use organic osmolytes as osmoprotective compounds during the earliest stages of their development. Mouse preimplantation (PI) embryos are acutely sensitive to increased osmolarity, but in vitro embryo development from fertilized eggs could be rescued if any of several organic compounds were added to the culture medium, implying that these compounds might function as organic osmolytes (Van Winkle et al., 1990c; Lawitts ß 2006 WILEY-LISS, INC. Contract grant sponsor: Canadian Institutes of Health Research (to JMB); Contract grant number: MOP62730; Contract grant sponsor: The Lalor Foundation (to MKIA and JMB); Contract grant sponsor: NERF, Foundation for Research, Science and Technology of New Zealand (to JLS); Contract grant sponsor: Health Research Council and National Heart Foundation of New Zealand (to ML). *Correspondence to: Jay M. Baltz, Moses and Rose Loeb Research Centre, Ottawa Health Research Institute, 725 Parkdale Avenue, Ottawa, ON, Canada K1Y 4E9. E-mail: [email protected] Received 14 June 2006; Accepted 17 August 2006 DOI: 10.1002/jcp.20872
Transcript of The organic osmolytes betaine and proline are transported by a shared system in early...

JOURNAL OF CELLULAR PHYSIOLOGY 210:266–277 (2007)
The Organic Osmolytes Betaine and ProlineAre Transported by a Shared System in Early
Preimplantation Mouse Embryos
MOHAMED-KHEIR IDRIS ANAS,1 MARY-ANNE HAMMER,1 MICHAEL LEVER,4
JO-ANN L. STANTON,5 AND JAY M. BALTZ1,2,3*1Hormones, Growth and Development Program,
Ottawa Health Research Institute, Ottawa, Ontario, Canada2Departments of Obstetrics and Gynecology (Division of Reproductive
Medicine), University of Ottawa, Ottawa, Ontario, Canada3Departments of Cellular and Molecular Medicine,
University of Ottawa, Ottawa, Ontario, Canada4Biochemistry Unit, Canterbury Health Laboratories,
Christchurch, New Zealand5Department of Anatomy and Structural Biology,
University of Otago Medical School, Dunedin, New Zealand
Betaine and proline protect preimplantation mouse embryos against increased osmolarity and decreased cell volume, implying thatthey may function as organic osmolytes. However, the transport system(s) that mediates their accumulation in fertilized eggs andearly embryos was unknown, and previously identified mammalian organic osmolyte transporters could not account for theirtransport. Here, we report that there is a single saturable transport component shared by betaine and proline in 1-cell mouseembryos. A series of inhibitors had nearly identical effects on both betaine and proline transport by this system. In addition, Ki valuesfor reciprocal inhibition of betaine and proline transport were �100–300 mM, similar to Km values (�200–300 mM) for theirtransport, and both had similar maximal transport rates (Vmax). The Ki values for inhibition of betaine and proline transport bydimethylglycine were similar (�2 mM), further supporting transport of both substrates by a single transport system. Finally, betaineand proline transport each required Naþ- and Cl�. These data were consistent with a single, Naþ- and Cl�-requiring, betaine/prolinetransport system in 1-cell mouse embryos. While betaine was only transported by a single saturable system, we found an additional,less conspicuous proline transport route that was betaine-insensitive, Naþ-sensitive, and inhibited by alanine, leucine, cysteine,and methionine. Furthermore, we showed that betaine, like proline, is present in the mouse oviduct and thus could serve as aphysiological substrate. Finally, accumulation of both betaine and proline increased with increasing osmolarity, consistent with apossible role as organic osmolytes in early embryos. J. Cell. Physiol. 210: 266–277, 2007. � 2006 Wiley-Liss, Inc.
Regulation of cell volume is a fundamental propertyof cells that animal cells accomplish by regulatingintracellular osmolyte concentrations. Acute volumeperturbations are alleviated by quickly adjusting in-organic ion concentrations (Hallows and Knauf, 1994),but cells that are normally subject to high osmolarityinstead require ‘‘organic osmolytes,’’ small unchargedorganic compounds, to provide long-term intracellularosmotic support (Yancey et al., 1982; Garcia-Perez andBurg, 1991; Lang et al., 1998). This avoids the severemetabolic disruption caused by the substantiallyincreased intracellular ionic strength that would berequired to balance high osmolarity using inorganicions. While some organic osmolytes are generatedintracellularly, most are imported from the externalenvironment by specific transporters. In mammals, fourorganic osmolyte transporters, all of which are stimu-lated by hypertonicity, have previously been identified.These are the Naþ/myo-inositol transporter (SMIT),taurine/b-amino acid transporter (TAUT), betaine/GABA transporter (BGT1), and System A aminoacid transporter (ATA2) (Garcia-Perez and Burg, 1991;Kwon and Handler, 1995; Pastor-Anglada et al., 1996;Alfieri et al., 2001). The requirement for organicosmolytes, and for these organic osmolyte transporters,is well established in cells subjected to hypertonicity,such as in the kidney. However, a range of cells notnormally subject to osmotic stress also use organicosmolytes (Lang et al., 1998), and thus organic osmo-
lytes and osmolyte transporters are likely to function in‘‘housekeeping’’ cell volume regulation as well as inresponse to hypertonic conditions.
There is strong evidence that mammalian embryosuse organic osmolytes as osmoprotective compoundsduring the earliest stages of their development. Mousepreimplantation (PI) embryos are acutely sensitive toincreased osmolarity, but in vitro embryo developmentfrom fertilized eggs could be rescued if any of severalorganic compounds were added to the culture medium,implying that these compounds might function asorganic osmolytes (Van Winkle et al., 1990c; Lawitts
� 2006 WILEY-LISS, INC.
Contract grant sponsor: Canadian Institutes of Health Research(to JMB); Contract grant number: MOP62730; Contract grantsponsor: The Lalor Foundation (to MKIA and JMB); Contractgrant sponsor: NERF, Foundation for Research, Science andTechnology of New Zealand (to JLS); Contract grant sponsor:Health Research Council and National Heart Foundation of NewZealand (to ML).
*Correspondence to: Jay M. Baltz, Moses and Rose Loeb ResearchCentre, Ottawa Health Research Institute, 725 Parkdale Avenue,Ottawa, ON, Canada K1Y 4E9. E-mail: [email protected]
Received 14 June 2006; Accepted 17 August 2006
DOI: 10.1002/jcp.20872

and Biggers, 1991, 1992; Dawson and Baltz, 1997; Baltz,2001; Hadi et al., 2004). We previously screened a rangeof compounds for their ability to protect mouse embryodevelopment from fertilized eggs against increasedosmolarity, and identified glycine, betaine (N,N,N-trimethylglycine or glycine betaine), proline, and gluta-mine as the most effective, and thus probable organicosmolytes in PI embryos (Dawson and Baltz, 1997;Baltz, 2001).
We focused first on glycine, since early PI embryoswere known to transport glycine at a high rate, via theGLYT1 (slc6a9) transporter (Hobbs and Kaye, 1985;Van Winkle et al., 1988), which is a member of the Naþ-and Cl�-dependent neurotransmitter transporter (NTTor SLC6) family (Nelson, 1998; Hoglund et al., 2005).Also, a large amount of glycine is present in the oviduct,with mouse oviductal fluid containing 0.5–3.0 mMglycine (Guerin et al., 1995; Harris et al., 2005) andsimilarly high amounts in tubal fluid of other mammals.Recently, we established GLYT1 as an organic osmolytetransporter that has a major role in cell volume home-ostasis in early PI embryos, functioning at normalphysiological osmolarity (Steeves et al., 2003; Steevesand Baltz, 2005). Glycine was accumulated via GLYT1by 1-cell mouse embryos to very high intracellularconcentrations (20–50 mM), which increased withincreasing osmolarity over a wide range. Accumulationof glycine and GLYT1 activity were required for embryoviability when osmolarity was similar to that ofoviductal fluid in vivo, and also for maintenance ofnormal cell volume in 1-cell embryos. Thus, this novelmechanism, and not one of the previously identifiedmammalian organic osmolyte transporters, accountedfor the ability of glycine to act as an organic osmolyte inearly PI embryos. How the amount of glycine in early PIembryos is regulated by osmolarity has not beencompletely elucidated. Part of the regulation mayinvolve a modest stimulation of glycine uptake viaGLYT1 by increased osmolarity (Steeves et al., 2003).However, the main regulatory mechanism may becontrol over glycine efflux through the cell swelling-activated anion and organic osmolyte channel present inembryos (Kolajova and Baltz, 1999; Steeves and Baltz,2005) as well as most other cells.
Betaine was also extremely effective at protectingmouse embryo development against increased osmolar-ity (Biggers et al., 1993), with an EC50 of about 40 mM(Hammer and Baltz, 2002), comparable to glycine (about50 mM, (Dawson and Baltz, 1997)). Since GLYT1 acceptssarcosine (N-methylglycine) as a high-affinity sub-strate, we had originally proposed that transport viaGLYT1 could also account for the ability of betaine(N,N,N-trimethylglycine) to protect PI mouse embryosagainst increased osmolarity (Baltz, 2001). However, weunexpectedly found that GLYT1 in 1-cell mouseembryos did not transport betaine (Hammer and Baltz,2002). Furthermore, established mammalian organicosmolyte transporters shown to accept betaine(BGT1 and System A) were not present in early PImouse embryos (Van Winkle et al., 1988; Hammer andBaltz, 2002). Proline also effectively protected PImouse embryo development against increased osmolar-ity (Dawson and Baltz, 1997; Baltz, 2001). Sincebetaine and proline share a number of unrelatedtransport mechanisms in animals, plants, and bacteria(Stevens and Wright, 1985; MacMillan et al., 1999;Waditee et al., 2002), we hypothesized that proline andbetaine may be substrates of a single putative organicosmolyte transporter in embryos. The objective of this
study was to begin to characterize betaine and prolinetransport in early PI mouse embryos.
MATERIALS AND METHODSChemicals and media
All chemicals were obtained from Sigma (St. Louis, MO)unless otherwise noted, and were embryo tested grade or cellculture grade. Media were based on KSOM embryo culturemedium (Lawitts and Biggers, 1993), modified by omittingglutamine and replacing bovine serum albumin with polyvinylalcohol (1 mg/ml; average MW 30-70K) as the macromolecularcomponent of the medium. The osmolarity of this modifiedKSOM was about 250 mOsM. For experiments in whichdifferent osmolarities were used, osmolarity was adjustedby adding appropriate amounts of the trisaccharide D(þ)raffinose, which is neither transported nor metabolized bymammalian cells, as previously described (Dawson and Baltz,1997). Osmolarities were confirmed using a vapor–pressureosmometer (model 5520; Wescor, Logan, UT). This actuallydetermines osmolality (osmoles/kg solvent), but for dilutesolutions like those used here, the two are essentiallyequivalent, and we use the more convenient units of osmolarity(OsM) here. Modified HEPES–KSOM (pH 7.4) was used forflushing zygotes from the oviduct (Lawitts and Biggers, 1993).Where specified, Naþ in the medium was replaced with choline(mainly) or Kþ (where choline salts were not available), and Cl�
was replaced with gluconate. Betaine and amino acid stocks(10–100 mM) were kept at �208C, and added directly to themedium as specified for each experiment.
Embryos
One-cell embryos were obtained from female CF1 mice(Charles River Canada, St-constant, PQ, Canada) that hadbeen superovulated by intraperitoneal injection of 5 IU ofequine chorionic gonadotropin (eCG) followed�47.5 h later by5 IU of human chorionic gonadotropin (hCG) and immediatelymated overnight with BDF1 males. The mated female micewere sacrificed by cervical dislocation 20–24 h after adminis-tration of human chorionic gonadotropin, and the fertilizedeggs were removed from the excised oviducts by flushing withHEPES–KSOM supplemented with 300 mg/ml hyaluronidase.Animal protocols were approved by the Animal Care Commit-tee of the Ottawa Health Research Institute. The eggs werewashed four times in HEPES–KSOM, and fertilized eggsshowing two pronuclei were selected. Where indicated, theselected embryos were maintained according to standardtechniques in microdrop cultures under mineral oil (Sigma orAldrich Chemical Co., Milwaukee, WI) at 378C in 5% CO2/airuntil used.
Measurement of 3H-labeled compounds in embryos3H-betaine ([methyl-3H] betaine, 85 Ci/mmol, 1 mCi/ml) and
3H-proline (L-[2,3-3H] proline, �45 Ci/mmol, 1 mCi/ml) wereobtained from Amersham Biosciences (Arlington Heights, IL).3H-proline is a standard product and was ordered as needed.3H-betaine was custom-synthesized by Amersham in a 10 mCibatch using 3H methyl iodide, and was 99.4% pure betaine byHPLC (analyzed by Amersham). Stocks were stored at �808Cuntil needed. The 3H-proline stock was in 2% EtOH in water,and was added directly to culture medium. The 3H-betainestock was in 50% EtOH in water. Aliquots of betaine stock wereconcentrated by room-temperature evaporation immediatelybefore addition to medium, so that the final EtOH content was�2%. Standard curves for 3H-betaine and 3H-proline wereconstructed for each set of experiments by serial dilutions ofthe radiolabeled compounds. Since initial experiments indi-cated that betaine transport into 1-cell mouse embryos waslinear with time up to 90 min for concentrations up to 1 mM andproline transport was linear for at least 30 min (data notshown), 30 min and 10 min uptake times were routinely usedfor experiments with betaine and proline, respectively.
For determination of the intracellular content of radiola-beled compounds in embryos, 10–15 embryos were removedfrom the 3H-betaine- or 3H-proline-containing medium andimmediately washed several times in ice-cold HEPES–KSOM
BETAINE AND PROLINE TRANSPORT IN EMBRYOS 267
Journal of Cellular Physiology DOI 10.1002/jcp

of the same osmolarity as the radiolabeled compound-contain-ing medium. The embryos were then placed in scintillationvials, and 4 ml of scintillation fluid (Scintiverse BD; FisherScientific, Pittsburgh, PA) were added to each vial. 3H wasdetected using a model 2200CA TriCarb liquid scintillationcounter (Packard Instrument Co., Downer’s Grove, IL), witheach sample counted for 5 min. Background was measured bytaking a small volume of the final wash drop and determiningthe residual counts. Background values were subtracted fromthe counts obtained for the embryo samples. The total amountof each compound taken up by embryos was calculated usingthe standard curves and transport rates were expressed asmolar amount per-embryo per-min (by dividing by totalincubation time), while total accumulation was expressed asmolar amount per embryo. Where specified, the soluble andinsoluble 3H fractions accumulated by embryos were separatedby trichloroacetic acid (TCA) precipitation as previouslydescribed (Borland and Tasca, 1974).
For experiments in which the external concentrations oflabeled compounds were varied, the concentration of radi-olabeled betaine or proline was kept constant at 1 mM, whilethe total concentration was adjusted to the desired valuesby adding appropriate amounts of the unlabeled com-pounds. Specific activities used to calculate intracellularconcentrations were adjusted to account for this proportionalmixing.
Estimation of kinetic parameters and data analysis
To estimate the kinetics of betaine and proline transport, therates of total transport were measured as a function ofsubstrate concentration. Non-specific transport rates weredetermined in the presence of 5–10 mM unlabeled substrate,and subtracted from the total transport rates to yield saturabletransport. The resulting saturable transport rate data werethen fit by nonlinear least-squares regression to the Michae-lis–Menten equation in the form rate¼Vmax[S]/(Kmþ [S]),where [S] is the concentration of the substrate (i.e., proline orbetaine). The kinetic parameters Km and Vmax were obtainedfrom the curve fitting.
Inhibition constants (Ki) were estimated using Dixon plots(reciprocal of the average transport rate vs. inhibitor concen-tration) at three different substrate concentrations. For Dixonplots, points were included only if the specific transport wassignificantly above background (determined by t-test). Datawere fit by linear regression. The intersections of the two linesrepresenting the lowest substrate concentrations (steepestslopes) with the line of the highest substrate concentration(lowest slope) were taken to provide an estimate of the range ofKi.
For kinetic parameters (Km, Vmax, Ki), it was not practicalto construct replicate Michaelis–Menten nor Dixon plots toobtain the three or more independent measurements of eachkinetic parameter that would be required for statisticalcomparison between parameters, owing to the large numberof embryos and length of time needed for each set ofmeasurements (data in Figs. 1, 2, and 5). Therefore, onlysingle curves were constructed (with 3–5 replicate measure-ments at each point), and the calculated kinetic parameterstaken to be estimates.
One set of experiments (‘‘ABC testing,’’ discussed below;data in Fig. 5) involved comparing Ki values estimated fromDixon plots with Km values for betaine and proline transport.Since it was not practical to obtain replicate measurements topermit statistical analysis, we instead determined Km fromlinear regressions on Lineweaver-Burke plots (rate�1 vs.concentration�1), where the x-intercept is Km
�1. We assumedthat the inverse of any value within the range where the 95%confidence interval (CI) intercepted the concentration-1 axiswas not different from the estimated Km.
For experiments in which the ability of a range of substancesto inhibit uptake of proline or betaine was determined (data inFigs. 3 and 6), a substrate concentration of 1 mM was used,while the concentrations of potentially inhibitory compoundswere at least 5 mM (5–20 mM, as specified). As proposed byVan Winkle et al. (1990a,b), this low substrate concentration isemployed for two reasons. First, this increases the chance that
high affinity transport components will be detected in thepresence of high-capacity, low-affinity transport while stillallowing any lower affinity transport to be detected. Second,some amino acid transport systems, including in preimplanta-tion embryos, have Km values near 1 mM, and would besaturated at much higher substrate concentrations. Highconcentrations of potential inhibitors are employed to max-imize their effect, even if they are, for example, lower affinitycompetitive inhibitors. Extensive discussions of the rationalefor this type of design are found in articles by Van Winkle et al.(1990a,b).
Linear and nonlinear regressions, and determinations of CIfor linear regressions, were performed using SigmaPlot 8.2(SPSS, Chicago, IL). Mean values of independent treatmentgroups were compared by t-test (for two groups) or ANOVAfollowed by Tukey–Kramer Multiple Comparisons test(INSTAT, GraphPad, San Diego, CA). For data with one ortwo continuous independent variables (osmolarity, time),univariate ANOVA assuming a general linear model was usedto determine the significance of dependence of the independentvariable (intracellular concentration) on the independentvariable(s), performed with SPSS for Windows 13.0 (SPSS).In all cases, differences were considered to be statisticallysignificant when P< 0.05.
Measurement of endogenous betaine in oviducts
Oviducts were obtained from 129 strain female mice. Theywere mated with Swiss Webster strain males where indicated.Whole oviducts were excised. In one set of measurements,oviducts from mated females were processed for betainedetermination complete with embryos still in the oviduct. Ina second set of measurements, oviducts from unmated femaleswere processed complete with unfertilized eggs still in theoviduct. For some measurements, where noted, the embryosand cumulus cell mass were excised through the oviduct walland discarded, or oviductal fluid and eggs were flushed out ofthe oviduct with PBS (1 ml, using a 30 g needle and syringe)and discarded, before betaine was measured in the remainingoviduct. The betaine content of whole oviducts was measuredbecause it was impractical to obtain enough embryos oroviductal fluid to measure betaine in them directly.
Fig. 1. Total transport of betaine and proline in 1-cell mouse embryosas a function of concentration. The rate of transport of betaine andproline (indicated by key) as a function of their concentration (1 to1,000 mM) after subtraction of non-specific transport was fit bynonlinear least-squares regression to the Michaelis–Menten equationto obtain the curves shown. Kinetic parameters calculated from thesefits are given in the text. The non-specific transport (determined in thepresence of 5–10 mM unlabeled substrate, which should eliminate>90% of saturable transport) is shown in the Inset. Each pointrepresents the mean�SEM of at least three replicates.
268 ANAS ET AL.
Journal of Cellular Physiology DOI 10.1002/jcp
Each oviduct was blotted dry and weighed in a 1.5-mlmicrocentrifuge tube. Total betaine in the oviduct was thenmeasured using a method based on fluorescence derivatizationand HPLC (Storer et al., 2006; Storer and Lever, 2006). Briefly,50 ml of acetonitrile was added to each tube and the sampleshomogenized with a steel rod and followed by ultrasonication.Fifteen microliters of each extract was added to 250 ml ofextraction solvent (consisting of 10% methanol/90% acetoni-trile). Samples were then derivatized with 6-methoxy-2-naphthacyl triflate and betaine determined by HPLC withfluorescence detection. Data are reported as mg betaine/g wetweight of the oviduct. External standards (for calibration) andcontrols were treated similarly.
RESULTSKinetics of betaine and proline transport
in 1-cell mouse embryos
The rates of total betaine and proline transport in1-cell stage mouse embryos weredetermined as a functionof substrate concentration from 10 to 1,000 mM. Therewas a large component of non-saturable transport ofeach substrate (Fig. 1, inset). After non-saturable trans-port was subtracted, both betaine and proline displayedsaturable kinetics and were well fit by the Michaelis–Menten equation (Fig. 1), yielding estimated Km values
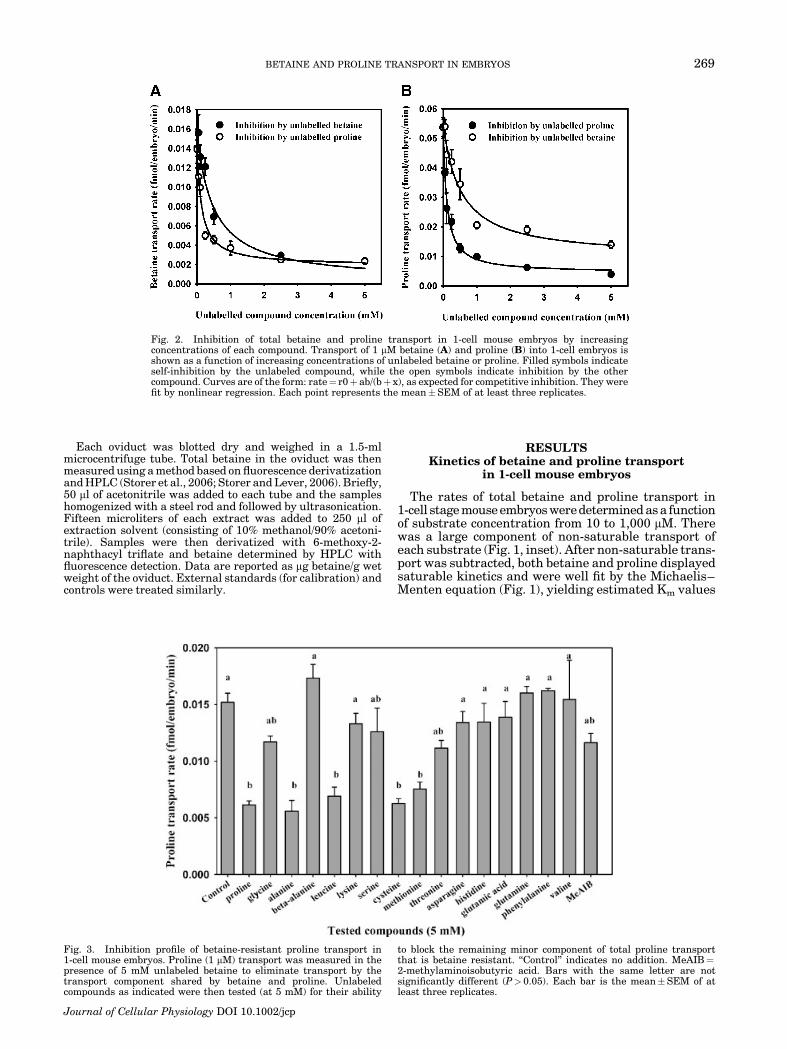
Fig. 2. Inhibition of total betaine and proline transport in 1-cell mouse embryos by increasingconcentrations of each compound. Transport of 1 mM betaine (A) and proline (B) into 1-cell embryos isshown as a function of increasing concentrations of unlabeled betaine or proline. Filled symbols indicateself-inhibition by the unlabeled compound, while the open symbols indicate inhibition by the othercompound. Curves are of the form: rate¼ r0þ ab/(bþ x), as expected for competitive inhibition. They werefit by nonlinear regression. Each point represents the mean�SEM of at least three replicates.
Fig. 3. Inhibition profile of betaine-resistant proline transport in1-cell mouse embryos. Proline (1 mM) transport was measured in thepresence of 5 mM unlabeled betaine to eliminate transport by thetransport component shared by betaine and proline. Unlabeledcompounds as indicated were then tested (at 5 mM) for their ability
to block the remaining minor component of total proline transportthat is betaine resistant. ‘‘Control’’ indicates no addition. MeAIB¼2-methylaminoisobutyric acid. Bars with the same letter are notsignificantly different (P> 0.05). Each bar is the mean�SEM of atleast three replicates.
BETAINE AND PROLINE TRANSPORT IN EMBRYOS 269
Journal of Cellular Physiology DOI 10.1002/jcp
of �200 mM (213 mM obtained from least-squares fit) forbetaine and �400 mM (413 from fit) for proline. Theapparent Vmax was 3.2 fmol/embryo/min for betaine,but was much higher for proline, for which Vmax was8.8 fmol/embryo/min.
We examined the saturability of transport of bothcompounds in more detail, using a concentration of1 mM labeled substrate (see Materials and Methods)with increasing concentrations of unlabeled substratefor competition. About 85% of 3H betaine transport(Fig. 2A), and 90% of proline transport (Fig. 2B) in 1-cellmouse embryos was eliminated by 5 mM of the sameunlabeled compound. Thus, transport of 1 mM of bothbetaine and proline in 1-cell embryos were similarlysaturable.
To determine whether betaine and proline arecompetitive inhibitors of each other’s saturable trans-port, we measured the transport of each compound into1-cell mouse embryos in the presence of increasingconcentrations of the other unlabeled compound. Wefound that essentially all saturable 3H-betaine (1 mM)transport was also inhibitable by unlabeled proline,since the residual betaine transport rate in the presenceof either 5 mM unlabeled betaine or proline werenearly identical (Fig. 2A). However, only about 80% ofsaturable 3H-proline (1mM) transport was inhibitable by5 mM betaine (Fig. 2B), indicating the possible presenceof more than one component of proline transport in 1-cellmouse embryos.
Characteristics of the betaine-resistantcomponent of proline transport
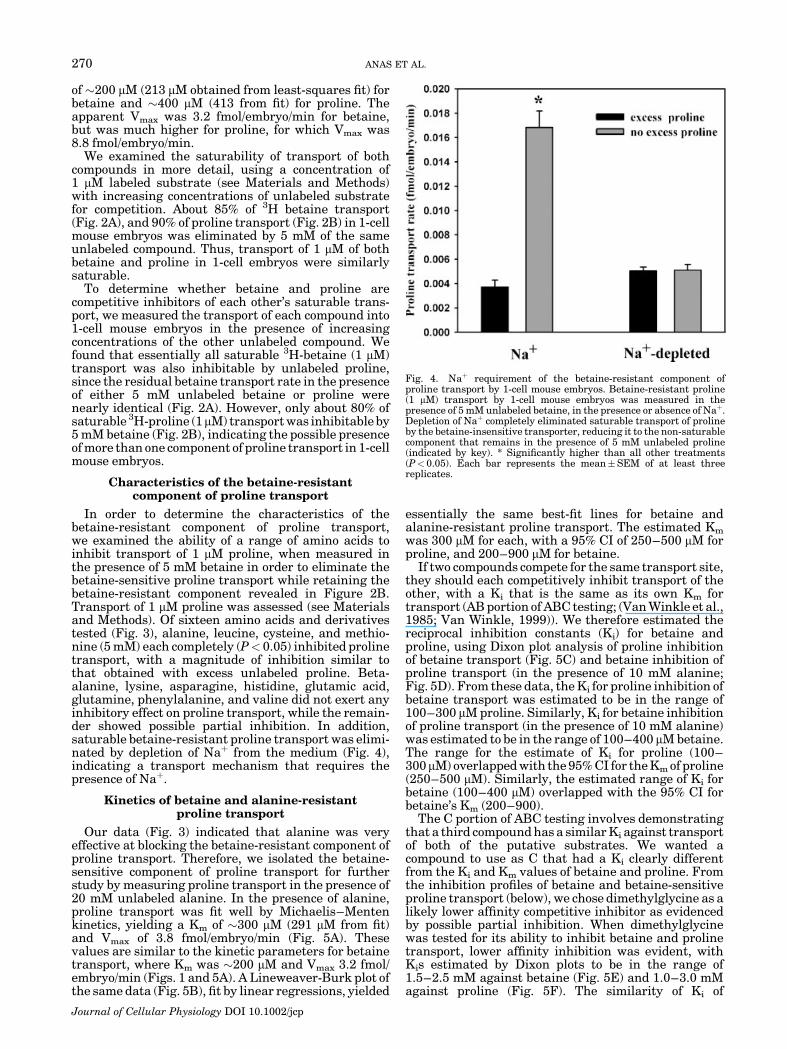
In order to determine the characteristics of thebetaine-resistant component of proline transport,we examined the ability of a range of amino acids toinhibit transport of 1 mM proline, when measured inthe presence of 5 mM betaine in order to eliminate thebetaine-sensitive proline transport while retaining thebetaine-resistant component revealed in Figure 2B.Transport of 1 mM proline was assessed (see Materialsand Methods). Of sixteen amino acids and derivativestested (Fig. 3), alanine, leucine, cysteine, and methio-nine (5 mM) each completely (P< 0.05) inhibited prolinetransport, with a magnitude of inhibition similar tothat obtained with excess unlabeled proline. Beta-alanine, lysine, asparagine, histidine, glutamic acid,glutamine, phenylalanine, and valine did not exert anyinhibitory effect on proline transport, while the remain-der showed possible partial inhibition. In addition,saturable betaine-resistant proline transport was elimi-nated by depletion of Naþ from the medium (Fig. 4),indicating a transport mechanism that requires thepresence of Naþ.
Kinetics of betaine and alanine-resistantproline transport
Our data (Fig. 3) indicated that alanine was veryeffective at blocking the betaine-resistant component ofproline transport. Therefore, we isolated the betaine-sensitive component of proline transport for furtherstudy by measuring proline transport in the presence of20 mM unlabeled alanine. In the presence of alanine,proline transport was fit well by Michaelis–Mentenkinetics, yielding a Km of �300 mM (291 mM from fit)and Vmax of 3.8 fmol/embryo/min (Fig. 5A). Thesevalues are similar to the kinetic parameters for betainetransport, where Km was �200 mM and Vmax 3.2 fmol/embryo/min (Figs. 1 and 5A). A Lineweaver-Burk plot ofthe same data (Fig. 5B), fit by linear regressions, yielded
essentially the same best-fit lines for betaine andalanine-resistant proline transport. The estimated Km
was 300 mM for each, with a 95% CI of 250–500 mM forproline, and 200–900 mM for betaine.
If two compounds compete for the same transport site,they should each competitively inhibit transport of theother, with a Ki that is the same as its own Km fortransport (AB portion of ABC testing; (Van Winkle et al.,1985; Van Winkle, 1999)). We therefore estimated thereciprocal inhibition constants (Ki) for betaine andproline, using Dixon plot analysis of proline inhibitionof betaine transport (Fig. 5C) and betaine inhibition ofproline transport (in the presence of 10 mM alanine;Fig. 5D). From these data, the Ki for proline inhibition ofbetaine transport was estimated to be in the range of100–300 mM proline. Similarly, Ki for betaine inhibitionof proline transport (in the presence of 10 mM alanine)was estimated to be in the range of 100–400 mM betaine.The range for the estimate of Ki for proline (100–300mM) overlapped with the 95% CI for the Km of proline(250–500 mM). Similarly, the estimated range of Ki forbetaine (100–400 mM) overlapped with the 95% CI forbetaine’s Km (200–900).
The C portion of ABC testing involves demonstratingthat a third compound has a similar Ki against transportof both of the putative substrates. We wanted acompound to use as C that had a Ki clearly differentfrom the Ki and Km values of betaine and proline. Fromthe inhibition profiles of betaine and betaine-sensitiveproline transport (below), we chose dimethylglycine as alikely lower affinity competitive inhibitor as evidencedby possible partial inhibition. When dimethylglycinewas tested for its ability to inhibit betaine and prolinetransport, lower affinity inhibition was evident, withKis estimated by Dixon plots to be in the range of1.5–2.5 mM against betaine (Fig. 5E) and 1.0–3.0 mMagainst proline (Fig. 5F). The similarity of Ki of
Fig. 4. Naþ requirement of the betaine-resistant component ofproline transport by 1-cell mouse embryos. Betaine-resistant proline(1 mM) transport by 1-cell mouse embryos was measured in thepresence of 5 mM unlabeled betaine, in the presence or absence of Naþ.Depletion of Naþ completely eliminated saturable transport of prolineby the betaine-insensitive transporter, reducing it to the non-saturablecomponent that remains in the presence of 5 mM unlabeled proline(indicated by key). * Significantly higher than all other treatments(P< 0.05). Each bar represents the mean�SEM of at least threereplicates.
270 ANAS ET AL.
Journal of Cellular Physiology DOI 10.1002/jcp

dimethylglycine against transport of both proline andbetaine further supports their transport by a singlesystem.
Inhibition profile of betaine andalanine-resistant proline transport
We tested the hypothesis that the inhibitionprofiles of betaine and alanine-resistant prolinetransport would be identical, as expected if these two
compounds share a single transport mechanism.A set of amino acids and amino acid derivativesincluding several proline derivatives and analogs werescreened for their ability to inhibit betaine transport andalanine-resistant proline transport in 1-cell mouseembryos, using 1 mM substrate and 5 mM of eachpotential inhibitor (see Materials and Methods).Sarcosine, 2-methylaminoisobutyric acid (MeAIB), andthe proline-related compounds pipecolic acid, proline
Fig. 5. Kinetic analysis of the shared component of betaine andproline transport in 1-cell mouse embryos. A: The rate of transport ofproline (in the presence of 20 mM alanine) and betaine (indicated bykey) as a function of their concentration (10–1,000 mM), aftersubtraction of non-specific transport. Data were fit by nonlinearleast-squares regression to the Michaelis–Menten equation to obtainthe curves shown. Kinetic parameters calculated from these fits aregiven in the text. B: Lineweaver-Burk plot. Lines were fit by linearregression to the data in (A) (excluding 10 mM) for betaine and proline(indicated by key). The lines are too close to be distinguishable,
however. The thin solid line is the 95% confidence interval (CI) forbetaine, and the thin dashed line is the CI for proline. Kms areestimated from 1/(x-intercept), while the range is similarly obtainedfrom the intercepts of the CI lines (see text). C–F: Dixon plot analysesof inhibition of: betaine transport by proline (C), proline (in thepresence of 10 mM alanine) transport by betaine (D), and betaine (E)and proline (in the presence of 10 mM alanine) (F) transport bydimethylglycine. Concentrations of each substrate used are given inthe inset keys. Ki values obtained from these plots are given in thetext. Each point is the mean�SEM of at least five replicates.
BETAINE AND PROLINE TRANSPORT IN EMBRYOS 271
Journal of Cellular Physiology DOI 10.1002/jcp

methylester, and dehydroproline effectively inhibited(P<0.05) saturable betaine transport (Fig. 6A) andalanine-resistant proline (Fig. 6B) transport to alevel similar to that inhibited by excess unlabeledbetaine. Dimethylglycine, leucine, phenylalanine, nor-leucine, and cysteine each significantly (P< 0.05) butincompletely inhibited betaine and proline transport.Glycine, methionine, and histidine had no significantinhibitory effect (Fig. 6). As can be readily seen bycomparing the patterns in Figure 6A,B, the inhibitionprofiles for betaine and alanine-resistant prolinetransport were very similar.
Naþ- and Cl�-dependence of betaineand alanine-resistant proline transport in
1-cell mouse embryos
To determine the ion sensitivity of betaine transportand the alanine-resistant component of proline trans-port by 1-cell mouse embryos, transport of betaine orproline (in the presence of 10 mM alanine) wasmeasured in the presence or absence of Naþ or Cl�.
Transport of betaine was reduced by 86% and that ofproline by 78% when Naþwas depleted from the medium(Fig. 7). Similarly, betaine transport was reduced by85% and proline by 77% in Cl�-depleted media. Thissuggests that the transport of both betaine andproline by 1-cell mouse embryos is mediated by a Naþ-and Cl�-requiring transport process.
Osmosensitivity of accumulation of betaine andproline by mouse embryos
If betaine and/or proline can function as organicosmolytes in early preimplantation mouse embryos,then they may be accumulated to higher levels whenexternal osmolarity is increased, as is the case forglycine, an established organic osmolyte in 1-cellembryos (Steeves et al., 2003). To test whether totalaccumulation of betaine or proline was stimulatedby increased osmolarity, 1-cell mouse embryos werecultured for 1–24 h in medium supplemented with 1 mMbetaine or 1 mM proline at 250, 310, or 350 mOsM andthe total accumulation of betaine and proline measured
Fig. 6. Inhibition profile of the shared component of betaine and proline transport in 1-cell mouseembryos. The rate of betaine (1 mM) transport (A) and proline (in the presence of 10 mM alanine) transport(B) by 1-cell embryos was measured. The indicated compounds (5 mM each) were assessed for their abilityto block transport. ‘‘Control’’ indicates no addition. MeAIB¼ 2-methylaminoisobutyric acid. Bars with thesame letter are not significantly different (P> 0.05). Each bar represents the mean�SEM of at least threereplicates.
272 ANAS ET AL.
Journal of Cellular Physiology DOI 10.1002/jcp

in embryos at 1, 4, 8, and 24 h (at the last time point, theyhad reached the 2-cell stage). The total amount ofbetaine and proline both depended significantly onexternal osmolarity, and an appreciable difference inaccumulated betaine or proline was evident at 24 h(Fig. 8A,B). A more detailed investigation of totalintracellular betaine or proline accumulated at 24 h asa function of osmolarity confirmed that significance of
the dependence (Fig. 8C,D). Accumulation dependedon osmolarity mainly between 250 and 300 mOsM,especially for proline, which reached a clear plateau bythat point, while betaine accumulation may increasesomewhat beyond 300 mOsM. As with similar data forglycine (Steeves et al., 2003), there may be a loss ofviability at 370 mOsM or higher affecting the amount ofaccumulation.
To determine whether the measured 3H accumulatedby embryos was associated with soluble compounds orinstead had been incorporated into macromolecules,embryos were cultured in the presence of 1 mM 3H-betaine or 3H-proline for 24 h in 310 mOsM medium, asabove, and the amount of TCA-soluble and precipitable3H measured. The soluble fraction of accumulatedbetaine was 3.5�0.4 pmol/embryo (mean�SEM) com-pared to an insoluble fraction of 0.42�0.06 pmol/embryo. For proline, the soluble fraction was 2.1�0.1pmol/embryo compared to 0.30�0.02 pmol/embryothat was insoluble. Thus, approximately 90% of 3Haccumulated by embryos from either betaine or prolineremained in soluble form.
Betaine content of oviducts
For betaine to be a physiologically relevant substrateof a putative betaine/proline transport system in 1-cellmouse embryos, it would have to be present in their invivo environment, as previously shown for proline. Sinceit was not technically feasible to measure betainedirectly in oviductal fluid, we instead measuredthe betaine content of whole mouse oviducts as anindication of its likely availability. In one set ofmeasurements, the betaine content of whole mouseoviducts was determined in oviducts from mated
Fig. 7. Naþ and Cl� requirement of the shared component of betaineand proline transport in 1-cell mouse embryos. Betaine (1 mM) andproline (1 mM, in the presence of 10 mM alanine) transport by 1-cellmouse embryos were measured in the presence or absence of Naþ orCl�. Depletion of Naþ or Cl� essentially eliminated saturabletransport of both betaine and proline. * Significantly lower thanrespective controls (P< 0.05). Each bar represents the mean�SEM ofthree replicates.
Fig. 8. Intracellular accumulation of betaine and proline in 1-cellmouse embryos. The total amounts of betaine or proline accumulatedin embryos cultured for 24 h from the 1-cell to 2-cell stage weremeasured. Labeled betaine or proline was present at a total externalconcentration of 1 mM during the entire incubation. A: Betaineaccumulated as a function of time at 250, 310, or 350 mOsM. Therewas significant dependence on both osmolarity (P¼0.003 by Uni-variate ANOVA) and time (P< 10�6). B: Proline accumulated as afunction of time at 250, 310, or 350 mOsM. There was significant
dependence on both osmolarity (P¼7� 10�5) and time (P<10�6).Curves fit in A and B are single exponentials rising to maxima of theform G[1� e�at] as previously described (Steeves et al., 2003).C: Betaine accumulated after 24 h as a function of osmolarity. Thedependence on osmolarity was significant (P¼0.004 by UnivariateANOVA). D: Proline accumulated after 24 h as a function ofosmolarity. The dependence on osmolarity was significant(P¼ 0.031). Each point represents the mean�SEM of at least four(A,B) or three (C,D) replicates.
BETAINE AND PROLINE TRANSPORT IN EMBRYOS 273
Journal of Cellular Physiology DOI 10.1002/jcp

females, which contained 1-cell embryos. Oviductscontaining 1-cell embryos had a betaine content of60�6 mg/g wet weight (mean�SEM, N¼9). We alsoassessed the effect of removing and discarding thecumulus mass containing the embryos on the amount ofbetaine in the oviduct, but this did not significantlychange their measured betaine content (48� 5 mg/g wetweight; N¼ 8; one outlier at 133 mg/g was excluded;P¼0.16 vs. with 1-cell embryos by t-test). In a second setof measurements, betaine was determined in oviductsfrom unmated females, which contained unfertilizedeggs. Oviducts containing unfertilized eggs had a meanbetaine content of 34� 4 mg/g wet weight (mean�SEM,N¼ 8). Removing the eggs and oviductal fluid byflushing the oviductal lumen did not have a significanteffect on the amount of betaine in the remaining oviduct(28�2 mg/g betaine; N¼9; P¼0.22).
DISCUSSION
Early mouse embryos possess mechanisms for cellvolume regulation and osmoregulation. One novelmechanism is the glycine transporter GLYT1, whichfunctions at the earliest stages of preimplantationdevelopment (Van Winkle et al., 1990c; Steeves et al.,2003; Steeves and Baltz, 2005). However, previousevidence pointed to the presence of an additionalmechanism in 1-cell mouse embryos that acceptedbetaine rather than glycine (Biggers et al., 1993;Hammer and Baltz, 2002). This study was aimed atdemonstrating that a betaine transport mechanism ispresent in 1-cell embryos, and determining its kinetics,inhibition profile, and whether it likely also transportsproline. Based on the present results, we propose thatthere is a single significant transport pathway forbetaine in 1-cell mouse embryos that is also the majorcomponent of saturable proline transport, and thatthis mechanism may be a second organic osmolytetransporter in 1-cell mouse embryos.
While betaine appeared to be transported only by asingle significant saturable mechanism that alsoaccepted proline (as discussed in detail below), a portionof proline was also transported into 1-cell embryos via atleast one other route. The second route was betaine-resistant, but was blocked by alanine, leucine, cysteine,and methionine, and required the presence of Naþ. Itcould be distinguished from the betaine-sensitive com-ponent of proline transport not only by its resistance tocompetition by betaine, but also in its sensitivity toalanine and relative resistance to MeAIB inhibition. Theinhibition profile of betaine-resistant proline transportin 1-cell embryos does not match that of known aminoacid transport systems in early embryos. Its resistanceto serine, asparagine, and valine would appear to ruleout the B0 transporter that is significant in blastocysts(Van Winkle et al., 1990c). Resistance to serine, glycine,and lysine would appear to similarly rule out thetransport system B0,þ that becomes highly conspicuousby the blastocyst stage (Van Winkle et al., 1985, 1990c).Naþ-dependence and susceptibility to alanine andcysteine are consistent with transport by an ASC-typetransporter of the SLC1 gene family (Kanai andHediger, 2004), at least some of which can transportproline (Pinilla-Tenas et al., 2003), but the resistance toserine (which we confirmed in two independent sets ofexperiments, not shown) does not support such anassignment. It also appears to have a different inhibi-tory profile than the recently cloned B0AT2 transporterthat can accept proline (Broer et al., 2006). It is possiblethat the betaine-resistant component of proline trans-
port represents more than one system, complicatinginterpretation. In any case, future investigations wouldbe required to clarify its identity. For the present work,however, the partial elucidation of its inhibition profile(Fig. 3) allowed us to determine that alanine completelyblocked betaine-resistant proline transport. Couplingthis observation with the previous finding that thereis very little saturable alanine transport in 1-cellmouse embryos (Van Winkle et al., 1990c) implied thatmeasuring proline transport in the presence of excessalanine would allow isolation of proline transport by thesystem shared with betaine with minimal perturbationof the embryo.
The results presented here support the proposal thatthere is a transport system in 1-cell stage mouseembryos that is shared by betaine and proline. Con-sistent with this, Vmax values were similar for betaineand proline (when the betaine-resistant component ofproline transport is eliminated in the presence ofexcess alanine; Fig. 5A). Although similar maximaltransport rates do not necessarily indicate that twosubstrates share the same system, this finding isconsistent with the possibility that betaine and prolineare transported by the same mechanism which, whensaturated, transports each substrate at the samemaximal rate.
Both betaine and alanine-resistant proline transportwere eliminated to essentially the same extent by eitherNaþ or Cl� depletion (Fig. 7). While many transportersof amino acids and organic substrates are dependent onNaþ, only a fairly restricted set are also Cl�-requiring,supporting the hypothesis that betaine and prolineshare a transport mechanism. By analogy to glycinetransport via GLYT1, it would be expected that Naþ andCl� are co-transported into the embryo with betaine orproline, and then removed in an energy-requiringprocess wherein Naþ is extruded via Naþ, Kþ ATPaseactivity and Cl� efflux is driven by the membranepotential.
The observation that the inhibition profiles of betainetransport and alanine-resistant proline transport by 1-cell mouse embryos were nearly identical (Fig. 6)provided strong evidence for a single transport mechan-ism accepting both betaine and proline. This systemappears to prefer N-methylated compounds related toglycine, since it transports betaine (N,N,N-trimethyl-glycine), and was inhibited by N,N-dimethylglycine(with much lower affinity) and sarcosine (N-methylgly-cine), but not by glycine. Likewise, transport of bothbetaine and proline was sensitive to dehydroproline,proline methyl ester, and pipecolic acid, indicating thatthe transport mechanism for both accepts prolinederivatives.
The putative betaine/proline transport system in1-cell embryos had similar apparent affinities for bothcompounds, since Michaelis–Menten curves fitted to theconcentration-dependence of betaine and proline trans-port (Fig. 5A) each fell within the error bars of transportof both compounds, and non-linear least squares fits tothe Michaelis–Menten equation (Fig. 5A) and Line-weaver-Burk plots (Fig. 5B) both yielded similar valuesfor Km of betaine and proline of �200–300 mM. Inaddition, the 95% confidence intervals of the Line-weaver-Burk plots completely overlapped, indicatingthat the Kms for betaine and proline were not signifi-cantly different. The similar affinities imply that thistransport system would not show significant preferencebetween these substrates if they were present at similarconcentrations.
274 ANAS ET AL.
Journal of Cellular Physiology DOI 10.1002/jcp

The AB portion of an ABC test indicated a Km of�200 mM for betaine by non-linear least squares fit to aMichaelis–Menten plot, and 300 mM with a 95% CI of200–900 mM by Lineweaver-Burk plot, which overlapsthe Ki of 100–400 mM for betaine inhibition of prolinetransport estimated from Dixon plots. Similarly, the Km
for proline transport was �300 mM by both methods,with a 95% CI of 200-500 mM (by Lineweaver-Burk plot),which overlaps the Ki of 100–300 mM for prolineinhibition of betaine estimated from Dixon plots. There-fore, the AB portion of ABC testing gave resultsreasonably consistent with a shared route of transportof both compounds. In addition, part C of the ABCtesting, using dimethylglycine, revealed similar Kisagainst both betaine (1.5–2.5 mM) and proline (1.0–3.0 mM), supporting the hypothesis of shared betaine/proline transport in 1-cell embryo.
We could not determine whether the measured Km
and Ki values differed significantly from one another bystatistical tests (e.g., ANOVA) because the cost and timerequired to perform each of the relevant Km and Ki
measurements (Fig. 5A–F) in at least triplicate isprohibitive. However, the method we used—comparingthe estimated range for Ki to the 95% CI of Km from aLineweaver-Burk plot—allowed a semi-quantitativedetermination that the relevant Km and Ki pairs wereof similar magnitude. In addition, it was easily seen thatKis of dimethylglycine against both betaine and prolinewere approximately an order of magnitude greater.Thus, the findings of ABC testing are compatible withthe hypothesis that there is a route of transport sharedby betaine and proline in 1-cell mouse embryos. It shouldbe noted, however, that the current results cannot ruleout the presence of more than one transport mechanismexisting in 1-cell embryos for both betaine and prolinethat cannot be distinguished kinetically nor by inhibi-tion profile. Although unlikely, this possibility can onlybe ruled out conclusively by establishing the molecularidentity of the transporter, which will require futureinvestigations.
The properties of the proposed betaine/proline trans-port system in 1-cell mouse embryos do not correspond toany amino acid transporters previously shown to bepresent in mouse embryos (Baltz, 2001; Van Winkle,2001; Hammer and Baltz, 2002). The requirement forNaþ and Cl� may indicate that the embryonic betaine/proline transporter is likely a member of the sameneurotransmitter transporter (NTT, SLC6) family asGLYT1, since all known mammalian Naþ- and Cl�-dependent transporters of organic compounds lie in thisfamily (Nelson, 1998; Chen et al., 2004; Hoglund et al.,2005). Within this gene family, there are at least twopossible candidates for the transport system identifiedfunctionally here: the brain proline transporter (PROT;slc6a7) and the intestinal imino acid transporter(IMINO; slc6a20 or XT3S1). Both have been clonedand their transport properties characterized.
PROT is a purportedly brain-specific proline trans-porter with some properties in common with thetransport activity described here, including effectiveinhibition by sarcosine and pipecolic acid (Fremeauet al., 1992; Shafqat et al., 1995). Whether PROT acceptsbetaine has not been reported. In contrast to the presentfindings, however, histidine is reported to be an effectiveinhibitor of PROT while MeAIB is not, and its reportedaffinity for proline in other cells (Km of about 10 mM) ishigher than that of the betaine/proline transport systemin embryos. In addition, PROT is reportedly inhibited bythe peptide des-Tyr-Leu-enkephalin (GGFL; (Fremeau
et al., 1996; Galli et al., 1999)), but we found thatGGFL did not significantly inhibit betaine transportinto 1-cell mouse embryos (100 mM GGFL used; data notshown).
At least one isoform of IMINO was recently deter-mined to correspond to the previously ‘‘orphan’’transporter slc6a20 or XT3S1 (Kowalczuk et al., 2005;Takanaga et al., 2005), which is expressed in a variety oftissues. Its Km for proline is about 100–200 mM, and italso accepts betaine with a Km of about 200 mM (Stevensand Wright, 1985, 1987; Kowalczuk et al., 2005;Takanaga et al., 2005). MeAIB, sarcosine, dehydropro-line, proline methyl ester, and pipecolic acid were allfound to be substrates of IMINO. Moreover, IMINOwas poorly inhibited by leucine, phenylalanine, andcysteine, and was resistant to glycine, methionine, andhistidine (Stevens and Wright, 1985; Kowalczuk et al.,2005; Takanaga et al., 2005). These properties aresimilar to those of the betaine/proline transport activityin 1-cell mouse embryos described here. Further work,however, is needed to determine the molecularidentity of the betaine/proline transporter in oocytes.In particular, genetic knockout mouse models, which toour knowledge are not yet available for either transpor-ter, could help determine whether one of them isresponsible for betaine and proline transport inembryos.
We have proposed that the betaine/proline transpor-ter in 1-cell mouse embryos can function as an organicosmolyte transporter. If so, two predictions would bethat both betaine and proline should be accumulated tohigh levels by early mouse embryos, and that theamount accumulated will depend on external osmolar-ity, similar to what was shown for glycine accumulationvia GLYT1 (Steeves et al., 2003). Here, we found thatincreasing external osmolarity from 250 mOsM throughabout 350 mOsM caused a moderate increase inaccumulation of both betaine and proline in 1-cell mouseembryos over the course of culture for 24 h to the 2-cellstage (Fig. 8). Almost all of the accumulated betaine- andproline-associated 3H label remained in the solublefraction after 24 h. This is similar to what was previouslyshown for glycine (Steeves et al., 2003). Such differentialaccumulation that depends on osmolarity is a keyproperty of organic osmolytes in many cell types, andhas been conclusively shown for glycine in early mouseembryos (Steeves et al., 2003). Thus, our results supportthe idea that both betaine and proline can serve asorganic osmolytes in 1-cell mouse embryos. However,their importance in this area relative to glycine, which isaccumulated to higher levels and shows a strongerdependence on osmolarity (Steeves et al., 2003), remainsto be determined.
For glycine accumulation, GLYT1 is responsible forglycine uptake, but osmotic control of an efflux pathway,likely to be the swelling-activated anion/organic osmo-lyte channel, plays the major role in determining thetotal amount of glycine that is retained by embryos(Kolajova and Baltz, 1999; Steeves and Baltz, 2005).This swelling-activated channel in 1-cell embryos isalso permeable to betaine and proline (Baltz, 2001).The increase in glycine accumulation with increasedosmolarity is not dependent on protein synthesis,indicating that, in contrast to the previously knownmammalian organic osmolyte transporters, de novotransporter synthesis is not required for osmoregulatedglycine accumulation (Steeves et al., 2003). Thus, it ispossible that the amount of betaine or proline accumu-lated by embryos may be similarly regulated mainly by
BETAINE AND PROLINE TRANSPORT IN EMBRYOS 275
Journal of Cellular Physiology DOI 10.1002/jcp

osmotic control of their efflux, rather than through anysignificant, osmotically-stimulated increase in the rateof betaine and proline uptake.
In addition to serving as organic osmolytes, there areother possible roles for betaine and proline in embryos.Proline is, of course, incorporated into proteins,although the level of proline accumulated by embryos,at least when only proline was present (Fig. 8), wouldseem excessive for that purpose alone. Proline can alsoserve as a metabolic substrate (Adams and Frank, 1980),although whether it is metabolized in PI embryos, and ifso, by which pathways, has not been elucidated. Betaine,in addition to its well-established role as an organicosmolyte in a number of tissues including kidney, canalso serve as a methyl donor for various biochemicalmethylation reactions, a role mainly characterized inliver (Selhub, 1999).
For either betaine or proline to be physiologicalsubstrates of a proposed betaine/proline transporter in1-cell mouse embryos, these compounds must be presentin the in vivo environment of the embryo. It haspreviously been found that mouse oviductal fluidcontains about 140 mM proline (Guerin et al., 1995),equivalent to 1–3% of total amino acids (Baltz, 2001).Proline is also present at significant concentrations intubal fluid of cow and rabbit, to a lesser extent in sheep,but little in pig (Guerin et al., 1995; Elhassan et al.,2001). There was no information available about betainein oviducts. Therefore, we measured betaine in murineoviducts, and found that they contained measurableamounts of betaine. In two separate sets of experiments,the betaine content of intact whole oviducts was found tobe 60 mg/g wet weight (after fertilization) and 34 mg/g(before fertilization). Given a molecular weight ofbetaine of 117 D, this yields an average concentrationin the range of 300–500mM. Although this is the averageconcentration of betaine in whole oviducts, rather thanthe concentration in oviductal fluid, betaine may diffuseor be transported into oviductal fluid. Betaine concen-trations near this value would essentially saturate theembryo betaine transporter that was identified here.
Removing embryos or eggs and oviductal fluid fromthe oviducts slightly decreased betaine content of theremaining oviductal tissue, but in neither case was thedecrease significant. This indicated that a prohibitivelylarge number of embryos or amount of oviductal fluidwould be required to measure endogenous betaine inembryos or fluid directly, which we therefore have notattempted. The lower amount of betaine measured inoviducts containing unfertilized eggs compared to thosecontaining fertilized embryos may indicate an increasein betaine synthesis or betaine accumulation in oviduc-tal tissue at the time of fertilization. Although theamount of oviductal fluid or number of embryos thatwould be needed for direct determination of theirbetaine content made such measurements impractical,we infer that both oviductal fluid and embryos are likelyto contain betaine since it is clearly present in oviducts,especially after fertilization, and 1-cell embryos have atransport system that allows its intracellular accumula-tion. Thus, both betaine and proline should be taken upby 1-cell mouse embryos and may function as organicosmolytes through their accumulation via the betaine/proline transporter.
ACKNOWLEDGMENTS
We thank Chris McEntyre for excellent technicalassistance and performing the measurements of tissuebetaine contents reported here.
LITERATURE CITED
Adams E, Frank L. 1980. Metabolism of proline and the hydroxyprolines. AnnuRev Biochem 49:1005–1061.
Alfieri RR, Petronini PG, Bonelli MA, Caccamo AE, Cavazzoni A, Borghetti AF,Wheeler KP. 2001. Osmotic regulation of ATA2 mRNA expression and aminoacid transport System A activity. Biochem Biophys Res Commun 283:174–178.
Baltz JM. 2001. Osmoregulation and cell volume regulation in the preimplanta-tion embryo. In: Schatten GP, editor. Current Topics in Developmental Biology.San Diego: Academic Press. p 55–106.
Biggers JD, Lawitts JA, Lechene CP. 1993. The protective action of betaine on thedeleterious effects of NaCl on preimplantation mouse embryos in vitro. MolReprod Dev 34:380–390.
Borland RM, Tasca RJ. 1974. Activation of a Naþ-dependent amino acid transportsystem in preimplantation mouse embryos. Dev Biol 36:169–182.
Broer A, Tietze N, Kowalczuk S, Chubb S, Munzinger M, Bak LK, Broer S. 2006.The orphan transporter v7-3 (slc6a15) is a Naþ-dependent neutral amino acidtransporter (B0AT2). Biochem J 393:421–430.
Chen NH, Reith ME, Quick MW. 2004. Synaptic uptake and beyond: The sodium-and chloride-dependent neurotransmitter transporter family SLC6. PflugersArch 447:519–531.
Dawson KM, Baltz JM. 1997. Organic osmolytes and embryos: Substrates of theGly and beta transport systems protect mouse zygotes against the effects ofraised osmolarity. Biol Reprod 56:1550–1558.
Elhassan YM, Wu G, Leanez AC, Tasca RJ, Watson AJ, Westhusin ME. 2001.Amino acid concentrations in fluids from the bovine oviduct and uterus and inKSOM-based culture media. Theriogenology 55:1907–1918.
Fremeau RT Jr, Caron MG, Blakely RD. 1992. Molecular cloning and expressionof a high affinity L-proline transporter expressed in putative glutamatergicpathways of rat brain. Neuron 8:915–926.
Fremeau RT Jr, Velaz-Faircloth M, Miller JW, Henzi VA, Cohen SM, Nadler JV,Shafqat S, Blakely RD, Domin B. 1996. A novel nonopioid action ofenkephalins: Competitive inhibition of the mammalian brain high affinityL-proline transporter. Mol Pharmacol 49:1033–1041.
Galli A, Jayanthi LD, Ramsey IS, Miller JW, Fremeau RT Jr, DeFelice LJ. 1999.L-proline and L-pipecolate induce enkephalin-sensitive currents in humanembryonic kidney 293 cells transfected with the high-affinity mammalian brainL-proline transporter. J Neurosci 19:6290–6297.
Garcia-Perez A, Burg MB. 1991. Role of organic osmolytes in adaptation of renalcells to high osmolality. J Membr Biol 119:1–13.
Guerin JF, Gallois E, Croteau S, Revol N, Maurin F, Guillaud J, Menezo Y. 1995.Techniques de rOcolte et aminogrammes des liquides tubaire et folliculairechez les femelles domestiques. Revue Med Vet 146:805–814.
Hadi T, Hammer MA, Algire C, Richards T, Baltz JM. 2004. Similar effects ofosmolarity, glucose, and phosphate on cleavage past the 2-cell stage inmouse embryos from outbred and f1 hybrid females. Biol Reprod 72:179–187.
Hallows KR, Knauf PA. 1994. Principles of cell volume regulation. In: Strange K,editor. Cellular and Molecular Physiology of Cell Volume Regulation. BocaRaton: CRC Press. p 3–29.
Hammer MA, Baltz JM. 2002. Betaine is a highly effective organic osmolyte butdoes not appear to be transported by established organic osmolyte transportersin mouse embryos. Mol Reprod Dev 62:195–202.
Harris SE, Gopichandran N, Picton HM, Leese HJ, Orsi NM. 2005. Nutrientconcentrations in murine follicular fluid and the female reproductive tract.Theriogenology 64:992–1006.
Hobbs JG, Kaye PL. 1985. Glycine transport in mouse eggs and preimplantationembryos. J Reprod Fertil 74:77–86.
Hoglund PJ, Adzic D, Scicluna SJ, Lindblom J, Fredriksson R. 2005. Therepertoire of solute carriers of family 6: Identification of new human and rodentgenes. Biochem Biophys Res Commun 336:175–189.
Kanai Y, Hediger MA. 2004. The glutamate/neutral amino acid transporterfamily SLC1: Molecular, physiological and pharmacological aspects. PflugersArch 447:469–479.
Kolajova M, Baltz JM. 1999. Volume-regulated anion and organic osmolytechannels in mouse zygotes. Biol Reprod 60:964–972.
Kowalczuk S, Broer A, Munzinger M, Tietze N, Klingel K, Broer S. 2005.Molecular cloning of the mouse IMINO system: An Naþ- and Cl�-dependentproline transporter. Biochem J 386:417–422.
Kwon HM, Handler JS. 1995. Cell volume regulated transporters of compatibleosmolytes. Curr Opin Cell Biol 7:465–471.
Lang F, Busch GL, Volkl H. 1998. The diversity of volume regulatorymechanisms. Cell Physiol Biochem 8:1–45.
Lawitts JA, Biggers JD. 1991. Optimization of mouse embryo culture media usingsimplex methods. J Reprod Fertil 91:543–556.
Lawitts JA, Biggers JD. 1992. Joint effects of sodium chloride, glutamine, andglucose in mouse preimplantation embryo culture media. Mol Reprod Dev31:189–194.
Lawitts JA, Biggers JD. 1993. Culture of preimplantation embryos. MethodsEnzymol 225:153–164.
MacMillan SV, Alexander DA, Culham DE, Kunte HJ, Marshall EV, Rochon D,Wood JM. 1999. The ion coupling and organic substrate specificities ofosmoregulatory transporter ProP in Escherichia coli. Biochim Biophys Acta1420:30–44.
Nelson N. 1998. The family of Naþ/Cl- neurotransmitter transporters.J Neurochem 71:1785–1803.
Pastor-Anglada M, Felipe A, Casado FJ, Ferrer-Martinez A, Gomez-Angelats M.1996. Long-term osmotic regulation of amino acid transport systems inmammalian cells. Amino Acids 11:135–151.
Pinilla-Tenas J, Barber A, Lostao MP. 2003. Transport of proline andhydroxyproline by the neutral amino-acid exchanger ASCT1. J Membr Biol195:27–32.
Selhub J. 1999. Homocysteine metabolism. Annu Rev Nutr 19:217–246.Shafqat S, Velaz-Faircloth M, Henzi VA, Whitney KD, Yang-Feng TL, Seldin MF,
Fremeau RT Jr. 1995. Human brain-specific L-proline transporter: Molecularcloning, functional expression, and chromosomal localization of the gene inhuman and mouse genomes. Mol Pharmacol 48:219–229.
276 ANAS ET AL.
Journal of Cellular Physiology DOI 10.1002/jcp

Steeves CL, Baltz JM. 2005. Regulation of intracellular glycine as an organicosmolyte in early preimplantation mouse embryos. J Cell Physiol
Steeves CL, Hammer MA, Walker GB, Rae D, Stewart NA, Baltz JM. 2003. Theglycine neurotransmitter transporter GLYT1 is an organic osmolyte transpor-ter regulating cell volume in cleavage-stage embryos. Proc Natl Acad Sci USA100:13982–13987.
Stevens BR, Wright EM. 1985. Substrate specificity of the intestinal brush-border proline/sodium (IMINO) transporter. J Membr Biol 87:27–34.
Stevens BR, Wright EM. 1987. Kinetics of the intestinal brush border proline(Imino) carrier. J Biol Chem 262:6546–6551.
Storer MK, Lever M. 2006. Aracyl triflates for preparing fluorescent and UVabsorbing derivatives of unreactive carboxylates, amines and other metabo-lites. Anal Chim Acta 558:319–325.
Storer MK, McEntyre CJ, Lever M. 2006. Separation of cationic aracylderivatives of betaines and related compounds. J Chromatogr A 1104:263–271.
Takanaga H, Mackenzie B, Suzuki Y, Hediger MA. 2005. Identification ofmammalian proline transporter SIT1 (SLC6A20) with characteristics ofclassical system imino. J Biol Chem 280:8974–8984.
Van Winkle LJ. 1999. Transport Kinetics. In: Van Winkle LJ, editor.Biomembrane Transport. San Diego: Academic Press. p 65–131.
Van Winkle LJ. 2001. Amino acid transport regulation and early embryodevelopment. Biol Reprod 64:1–12.
Van Winkle LJ, Christensen HN, Campione AL. 1985. Naþ-dependent transportof basic, zwitterionic, and bicyclic amino acids by a broad-scope system inmouse blastocysts. J Biol Chem 260:12118–12123.
Van Winkle LJ, Haghighat N, Campione AL, Gorman JM. 1988. Glycinetransport in mouse eggs and preimplantation conceptuses. Biochim BiophysActa 941:241–256.
Van Winkle LJ, Campione AL, Farrington BH. 1990a. Development of systemB0,þ and a broad-scope Na(þ)-dependent transporter of zwitterionic aminoacids in preimplantation mouse conceptuses. Biochim Biophys Acta 1025:225–233.
Van Winkle LJ, Campione AL, Gorman JM. 1990b. Inhibition of transport systemb0,þ in blastocysts by inorganic and organic cations yields insight into thestructure of its amino acid receptor site. Biochim Biophys Acta 1025:215–224.
Van Winkle LJ, Haghighat N, Campione AL. 1990c. Glycine protects pre-implantation mouse conceptuses from a detrimental effect on development ofthe inorganic ions in oviductal fluid. J Exp Zool 253:215–219.
Waditee R, Hibino T, Tanaka Y, Nakamura T, Incharoensakdi A, Hayakawa S,Suzuki S, Futsuhara Y, Kawamitsu Y, Takabe T, Takabe T. 2002. Functionalcharacterization of betaine/proline transporters in betaine-accumulatingmangrove. J Biol Chem 277:18373–18382.
Yancey PH, Clark ME, Hand SC, Bowlus RD, Somero GN. 1982. Living withwater stress: Evolution of osmolyte systems. Science 217:1214–1222.
BETAINE AND PROLINE TRANSPORT IN EMBRYOS 277
Journal of Cellular Physiology DOI 10.1002/jcp




















