
The distribution and topographical organization in the thalamus of anterogradely-transported...
28
Exp Brain Res (1985) 58:227254 _F_. , _'mental Bran Research Springer-Verlag 1985 The distribution and topographical organization in the thalamus of anterogradely-transported horseradish peroxidase after spinal injections in cat and raccoon A.D. Craig* and H. Burton Departments of Anatomy and Neurobiology,and Physiology and Biophysics,Washington UniversitySchoolof Medicine, St. Louis, MO 63110, USA Summary. The distribution of anterogradely-trans- ported horseradish peroxidase (HRP) was examined in the rostral mesencephalon and thalamus of cats and raccoons that had received injections of HRP in the cervical and/or lumbosacral enlargements of the spinal cord. Labeling was consistently observed in a large number of loci. All regions previously iden- tified as targets of spinomesencephalic or spino- thalamic fibers were included. Evidence of topo- graphical organization was obtained in several regions. Adjacent fields of labeling were often separ- able on the basis of the distribution, appearance and topographical organization of the labeling. Subject to the methodological constraints imposed by the pos- sibilities of transneuronal and/or collateral labeling, we conclude that a wide variety of loci in the thalamus receive direct spinal input. The organiza- tion of these projections suggests that each terminal region may be associated with different aspects of spinal cord function. Key words: Anterograde transport - Horseradish peroxidase - Spinothalamic tract Introduction The mammalian spinothalamic tract (STT) is gener- ally associated with the functions of crude touch, pain and temperature sensation on the basis of clinical observations following partial transection. Yet, phy- siological evidence indicates that spinothalamic cells in rat, cat and monkey can respond to a wide variety * Present address: Physiologisches Institut der Universitfit, R6nt- genring 9, D-8700 Wfirzburg, Federal Republicof Germany Offprint requests to: A.D. Craig (address see footnote) of exteroceptive, proprioceptive, and enteroceptive stimuli (see Willis 1983). Complementary observa- tions obtained with anatomical retrograde tracing methods in these species indicate that STF cells of origin are widely distributed throughout the spinal gray matter (Carstens and Trevino 1978; Giesler et al. 1979; Willis et al. 1979). Therefore, full considera- tion should be given to the possibility that the STT contains elements functionally associated with each of the many aspects of spinal cord function. How- ever, this possibility is difficult to reconcile with the previous anatomical evidence regarding the terminal projection of the STT. It has generally been agreed that these occur in three main regions in the cat: the posterior group caudal to the ventrobasal thalamus, the ventrolateral nucleus, and the intralaminar nuclei of medial thalamus (Boivie 1971; Jones and Burton 1974; Berkley 1980). Evidence of anatomical organi- zation within these projections that might suggest a functionally-relevant parcellation has not been reported. While certain general differences in the location of the cells of origin of STI" projections to medial or lateral thalamus have been found, there is apparently considerable overlap. Similarly, phy- siological studies of these three regions have not revealed the functional role of their STT inputs, and few correlations have been observed between the location and/or response characteristics of physiologi- cally identified STT cells and the destination of the projecting axon of such cells in medial or lateral thalamus (Applebaum et al. 1979). These considera- tions recently led Berkley (1980) to speculate that functionally-relevant fiber-sorting processes might not occur within the STT projection to the dience- phalon. However, the considerable degree of func- tional and structural organization within the spinal cord itself (e.g. Brown 1981) suggests that the distribution and organization of the STT deserves further analYSiS (Boivie and Perl 1975).
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The distribution and topographical organization in the thalamus of anterogradely-transported...

Exp Brain Res (1985) 58:227254 _F_. , _'mental Bran Research �9 Springer-Verlag 1985
The distribution and topographical organization in the thalamus of anterogradely-transported horseradish peroxidase after spinal injections in cat and raccoon
A.D. Craig* and H. Burton
Departments of Anatomy and Neurobiology, and Physiology and Biophysics, Washington University School of Medicine, St. Louis, MO 63110, USA
Summary. The distribution of anterogradely-trans- ported horseradish peroxidase (HRP) was examined in the rostral mesencephalon and thalamus of cats and raccoons that had received injections of HRP in the cervical and/or lumbosacral enlargements of the spinal cord. Labeling was consistently observed in a large number of loci. All regions previously iden- tified as targets of spinomesencephalic or spino- thalamic fibers were included. Evidence of topo- graphical organization was obtained in several regions. Adjacent fields of labeling were often separ- able on the basis of the distribution, appearance and topographical organization of the labeling. Subject to the methodological constraints imposed by the pos- sibilities of transneuronal and/or collateral labeling, we conclude that a wide variety of loci in the thalamus receive direct spinal input. The organiza- tion of these projections suggests that each terminal region may be associated with different aspects of spinal cord function.
Key words: Anterograde transport - Horseradish peroxidase - Spinothalamic tract
Introduction
The mammalian spinothalamic tract (STT) is gener- ally associated with the functions of crude touch, pain and temperature sensation on the basis of clinical observations following partial transection. Yet, phy- siological evidence indicates that spinothalamic cells in rat, cat and monkey can respond to a wide variety
* Present address: Physiologisches Institut der Universitfit, R6nt- genring 9, D-8700 Wfirzburg, Federal Republic of Germany
Offprint requests to: A.D. Craig (address see footnote)
of exteroceptive, proprioceptive, and enteroceptive stimuli (see Willis 1983). Complementary observa- tions obtained with anatomical retrograde tracing methods in these species indicate that STF cells of origin are widely distributed throughout the spinal gray matter (Carstens and Trevino 1978; Giesler et al. 1979; Willis et al. 1979). Therefore, full considera- tion should be given to the possibility that the STT contains elements functionally associated with each of the many aspects of spinal cord function. How- ever, this possibility is difficult to reconcile with the previous anatomical evidence regarding the terminal projection of the STT. It has generally been agreed that these occur in three main regions in the cat: the posterior group caudal to the ventrobasal thalamus, the ventrolateral nucleus, and the intralaminar nuclei of medial thalamus (Boivie 1971; Jones and Burton 1974; Berkley 1980). Evidence of anatomical organi- zation within these projections that might suggest a functionally-relevant parcellation has not been reported. While certain general differences in the location of the cells of origin of STI" projections to medial or lateral thalamus have been found, there is apparently considerable overlap. Similarly, phy- siological studies of these three regions have not revealed the functional role of their STT inputs, and few correlations have been observed between the location and/or response characteristics of physiologi- cally identified STT cells and the destination of the projecting axon of such cells in medial or lateral thalamus (Applebaum et al. 1979). These considera- tions recently led Berkley (1980) to speculate that functionally-relevant fiber-sorting processes might not occur within the STT projection to the dience- phalon. However, the considerable degree of func- tional and structural organization within the spinal cord itself (e.g. Brown 1981) suggests that the distribution and organization of the STT deserves further analYSiS (Boivie and Perl 1975).
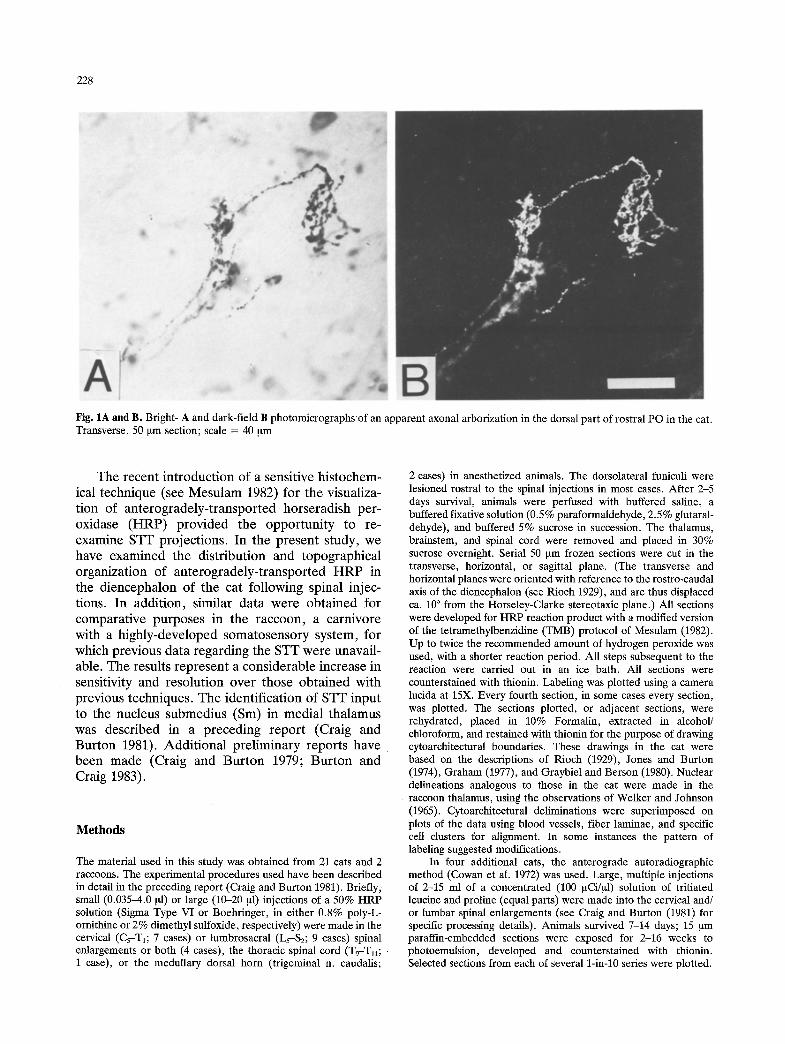
228
Fig. 1A and B. Bright- A and dark-field B photomicrographs of an apparent axonal arborization in the dorsal part of rostral PO in the cat. Transverse, 50 ~tm section; scale = 40 gm
The recent introduction of a sensitive histochem- ical technique (see Mesulam 1982) for the visualiza- tion of anterogradely-transported horseradish per- oxidase (HRP) provided the opportunity to re- examine STT projections. In the present study, we have examined the distribution and topographical organization of anterogradely-transported HRP in the diencephalon of the cat following spinal injec- tions. In addition, similar data were obtained for comparative purposes in the raccoon, a carnivore with a highly-developed somatosensory system, for which previous data regarding the STT were unavail- able. The results represent a considerable increase in sensitivity and resolution over those obtained with previous techniques. The identification of STT input to the nucleus submedius (Sm) in medial thalamus was described in a preceding report (Craig and Burton 1981). Additional preliminary reports have been made (Craig and Burton 1979; Burton and Craig 1983).
Methods
The material used in this study was obtained from 21 cats and 2 raccoons. The experimental procedures used have been described in detail in the preceding report (Craig and Burton 1981). Briefly, small (0.035-4.0 ~tl) or large (10-20 ~tl) injections of a 50% HRP solution (Sigma Type VI or Boehringer, in either 0.8% poly-L- ornithine or 2% dimethyl sulfoxide, respectively) were made in the cervical (Cr-T1; 7 cases) or lumbrosacral (Lr-S2; 9 cases) spinal enlargements or both (4 cases), the thoracic spinal cord (Tg-Tn; �9 1 case), or the medullary dorsal horn (trigeminal u. caudalis;
2 cases) in anesthetized animals. The dorsolateral funiculi were lesioned rostral to the spinal injections in most cases. After 2-5 days survival, animals were perfused with buffered saline, a buffered fixative solution (0.5% paraformaldehyde, 2.5% glutaral- dehyde), and buffered 5% sucrose in succession. The thalamus, brainstem, and spinal cord were removed and placed in 30% sucrose overnight. Serial 50 ~tm frozen sections were cut in the transverse, horizontal, or sagittal plane. (The transverse and horizontal planes were oriented with reference to the rostro-caudal axis of the diencephalon (see Rioch 1929), and are thus displaced ca. 10 ~ from the Horseley-Clarke stereotaxic plane.) All sections were developed for HRP reaction product with a modified version of the tetramethylbenzidine (TMB) protocol of Mesnlam (1982). Up to twice the recommended amount of hydrogen peroxide was used, with a shorter reaction period. All steps subsequent to the reaction were carried out in an ice bath. All sections were counterstained with thionln. Labeling was plotted using a camera lucida at 15X. Every fourth section, in some cases every section, was plotted. The sections plotted, or adjacent sections, were rehydrated, placed in 10% Formalin, extracted in alcohol/ chloroform, and restained with thionin for the purpose of drawing cytoarchitectural boundaries. These drawings in the cat were based on the descriptions of Rioch (1929), Jones and Burton (1974), Graham (1977), and Graybiel and Berson (1980). Nuclear delineations analogous to those in the cat were made in the raccoon thalamus, using the observations of Welker and Johnson (1965). Cytoarchitectural deliminations were superimposed on plots of the data using blood vessels, fiber laminae, and specific cell clusters for alignment. In some instances the pattern of labeling suggested modifications.
In four additional cats, the anterograde autoradiographic method (Cowan et al. 1972) was used. Large, multiple injections of 2-15 ml of a concentrated (100 ~tCi/ixl ) solution of tritiated leucine and protine (equal parts) were made into the cervical and/ or lumbar spinal enlargements (see Craig and Burton (1981) for specific processing details). Animals survived 7-14 days; 15 ~tm paraffin-embedded sections were exposed for 2-16 weeks to photoemnlsion, developed and counterstained with thionin. Selected sections from each of several 1-in-10 series were plotted.
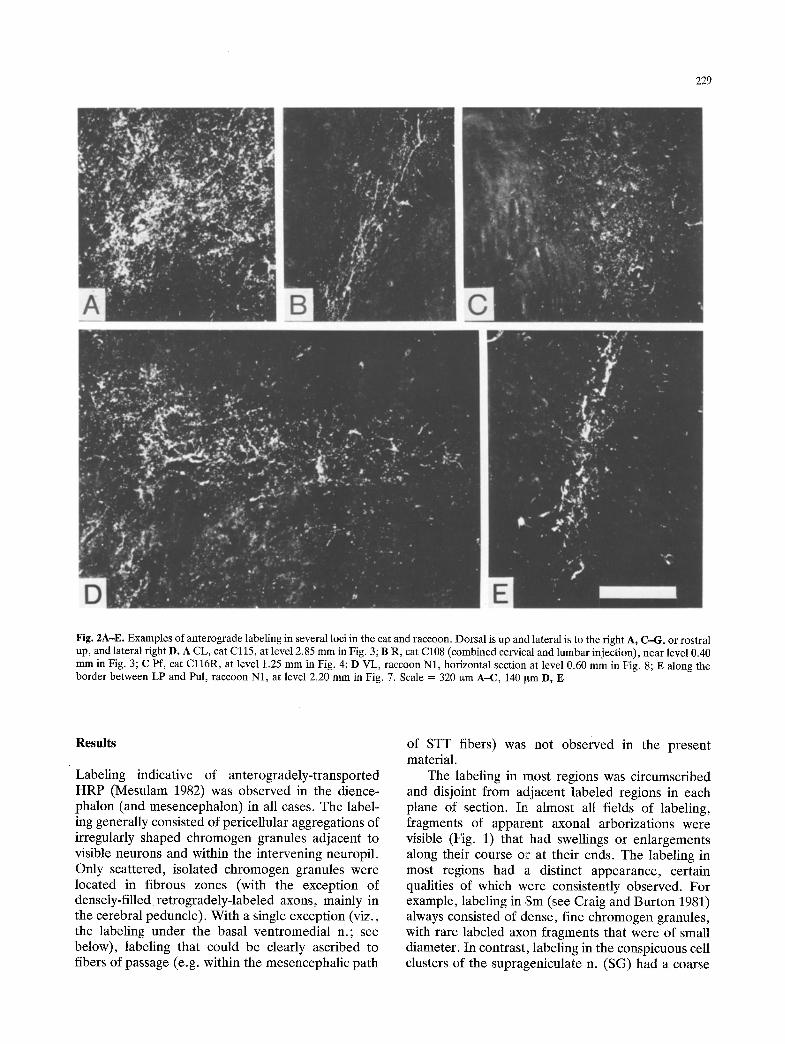
229
Fig. 2A-E. Examples of anterograde labeling in several loci in the cat and raccoon. Dorsal is up and lateral is to the right A, C-G, or rostral up, and lateral right D. A CL, cat Cl15, at level 2.85 mm in Fig. 3; B R, cat C108 (combined cervical and lumbar injection), near level 0.40 mm in Fig. 3; C Pf, cat Cl16R, at level 1.25 mm in Fig. 4; D VL, raccoon N1, horizontal section at level 0.60 mm in Fig. 8; E along the border between LP and Pul, raccoon N1, at level 2.20 mm in Fig. 7. Scale = 320 Ixm A-C, 140 ~m D, E
Results
Labeling indicative of anterogradely-transported H R P (Mesulam 1982) was observed in the dience- phalon (and mesencephalon) in all cases. The label- ing generally consisted of pericellular aggregations of irregularly shaped chromogen granules adjacent to visible neurons and within the intervening neuropil. Only scattered, isolated chromogen granules were located in fibrous zones (with the exception of densely-filled retrogradely-labeled axons, mainly in the cerebral peduncle). With a single exception (viz., the labeling under the basal ventromedial n.; see below), labeling that could be clearly ascribed to fibers of passage (e.g. within the mesencephalic path
of STT fibers) was not observed in the present material.
The labeling in most regions was circumscribed and disjoint f rom adjacent labeled regions in each plane of section. In almost all fields of labeling, fragments of apparent axonal arborizations were visible (Fig. 1) that had swellings or enlargements along their course or at their ends. The labeling in most regions had a distinct appearance, certain qualities of which were consistently observed. For example, labeling in Sm (see Craig and Burton 1981) always consisted of dense, fine chromogen granules, with rare labeled axon fragments that were of small diameter. In contrast, labeling in the conspicuous cell dusters of the suprageniculate n. (SG) had a coarse
230
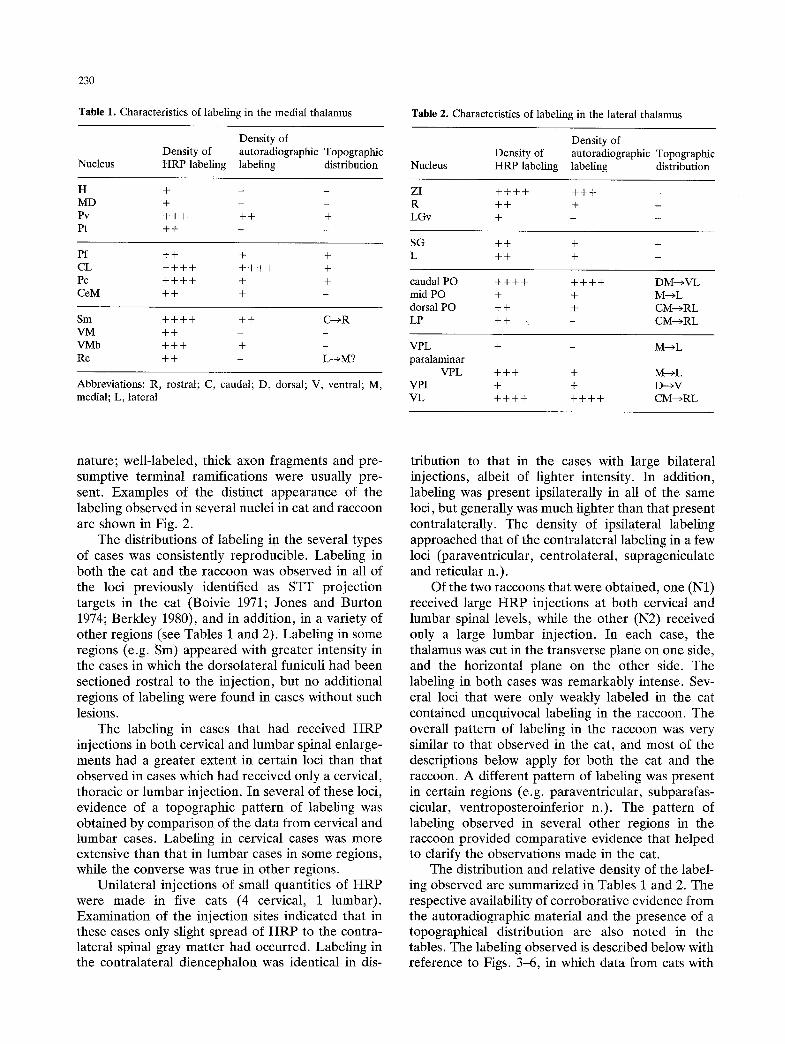
Table 1. Characteristics of labeling in the medial thalamus
Densi ty of Densi ty of autoradiographic Topographic
Nucleus H R P labeling labeling distribution
H -[ - - -
MD + - - Pv + + + + + + Pt + + - -
Pf + + + + CL + + + + + + + + + Pc + + + + + + CeM + + + -
Sm + + + + + + C ~ R VM + + - - VMb + + + + -
Re + + - L ~ M ?
Abbreviations: R, rostral; C, caudal; D, dorsal; V, ventral; M, medial; L, lateral
Table 2. Characteristics of labeling in the lateral thalamus
Densi ty of Densi ty of autoradiographic Topographic
Nucleus HRP labeling labeling distribution
ZI + + + + + + + R + + + LGv + -
SG + + + L + + +
caudal PO + + + + + + + + DM---~VL mid PO + + M---~L dorsal PO + + + CM---~RL
LP + + , - C M - * R L
VPL + - M----~L paralaminar
VPL + + + + M-*L VPI + + D---~V VL + + + + + + + + C M - * R L
nature; well-labeled, thick axon fragments and pre- sumptive terminal ramifications were usually pre- sent. Examples of the distinct appearance of the labeling observed in several nuclei in cat and raccoon are shown in Fig. 2.
The distributions of labeling in the several types of cases was consistently reproducible. Labeling in both the cat and the raccoon was observed in all of the loci previously identified as STT projection targets in the cat (Boivie 1971; Jones and Burton 1974; Berkley 1980), and in addition, in a variety of other regions (see Tables 1 and 2). Labeling in some regions (e.g. Sm) appeared with greater intensity in the cases in which the dorsolateral funiculi had been sectioned rostral to the injection, but no additional regions of labeling were found in cases without such lesions.
The labeling in cases that had received HRP injections in both cervical and lumbar spinal enlarge- ments had a greater extent in certain loci than that observed in cases which had received only a cervical, thoracic or lumbar injection. In several of these loci, evidence of a topographic pattern of labeling was obtained by comparison of the data from cervical and lumbar cases. Labeling in cervical cases was more extensive than that in lumbar cases in some regions, while the converse was true in other regions.
Unilateral injections of small quantities of HRP were made in five cats (4 cervical, 1 lumbar). Examination of the injection sites indicated that in these cases only slight spread of HRP to the contra- lateral spinal gray matter had occurred. Labeling in the contralateral diencephalon was identical in dis-
tribution to that in the cases with large bilateral injections, albeit of lighter intensity. In addition, labeling was present ipsilaterally in all of the same loci, but generally was much lighter than that present contralaterally. The density of ipsilateral labeling approached that of the contralateral labeling in a few loci (paraventricular, centrolateral, suprageniculate and reticular n.).
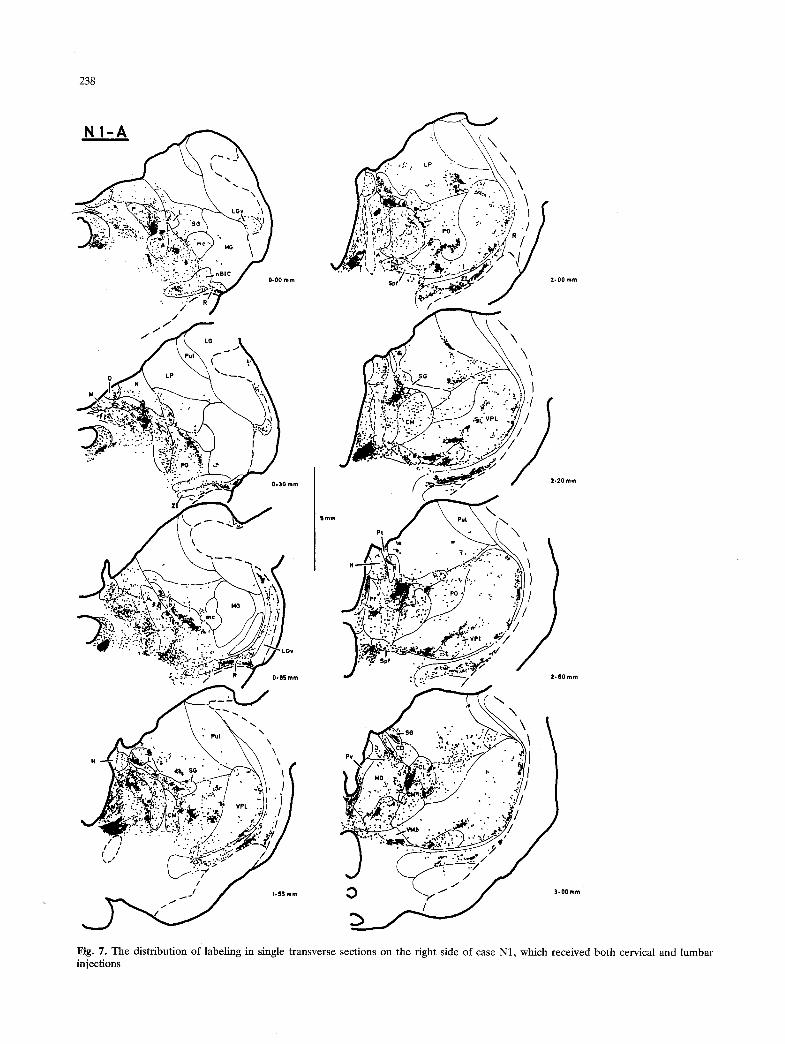
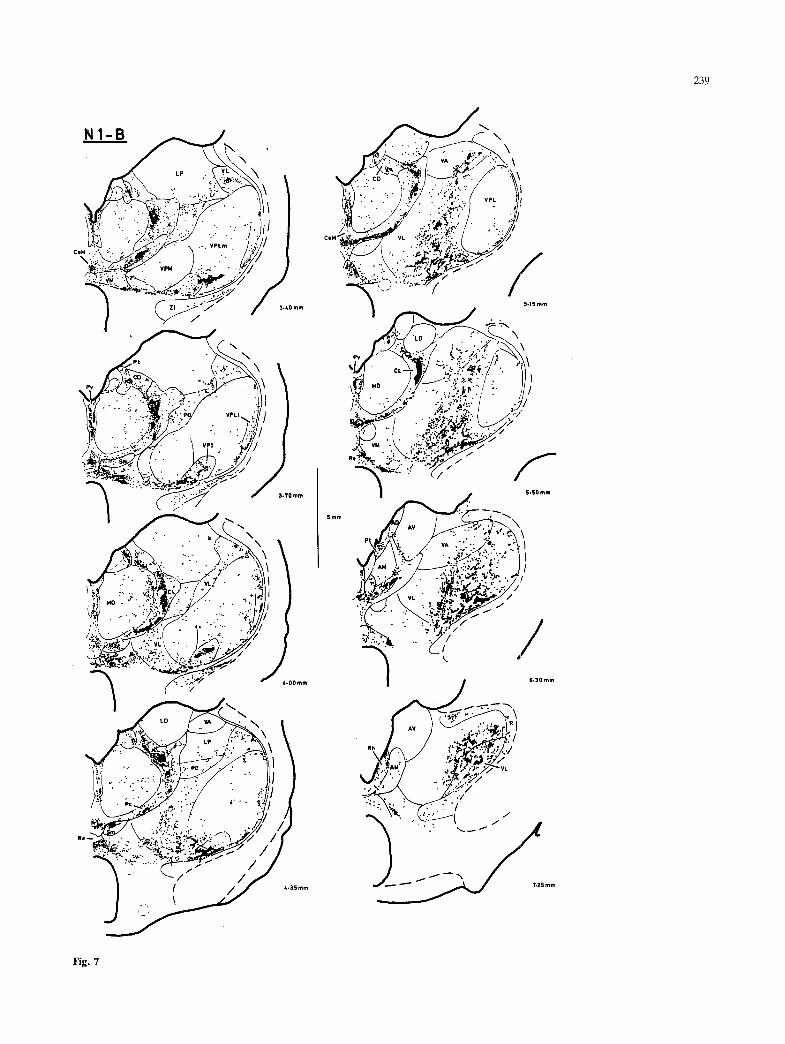
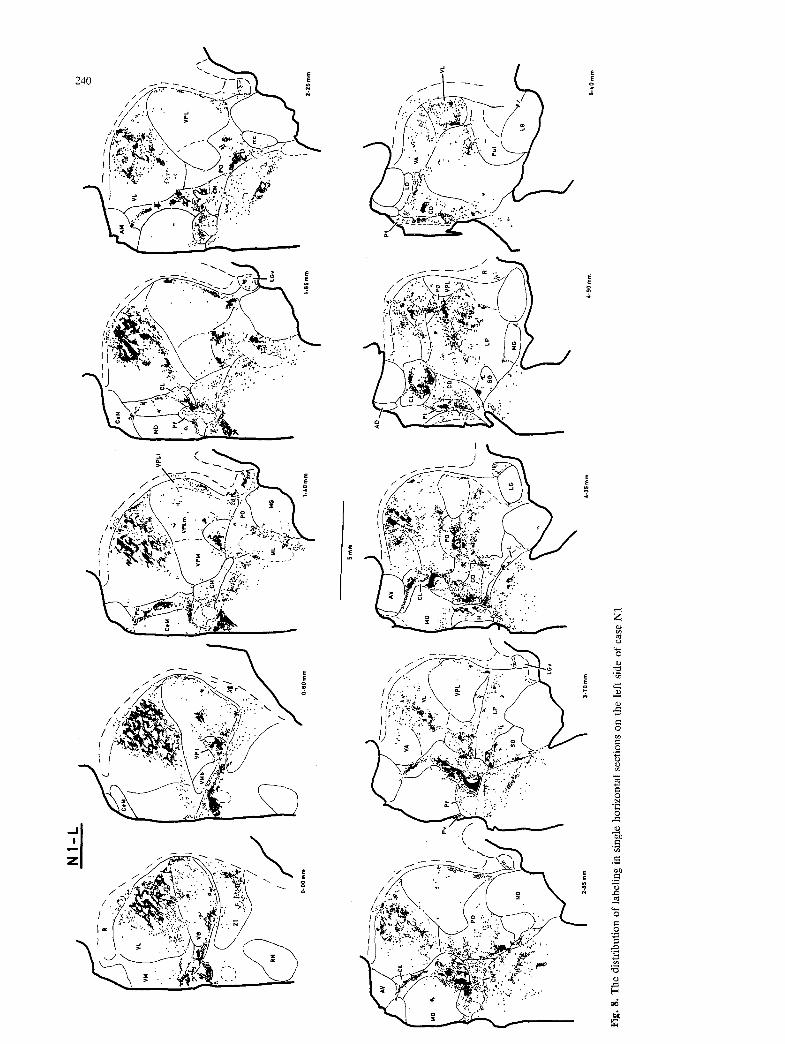
Of the two raccoons that were obtained, one (N1) received large HRP injections at both cervical and lumbar spinal levels, while the other (N2) received only a large lumbar injection. In each case, the thalamus was cut in the transverse plane on one side, and the horizontal plane on the other side. The labeling in both cases was remarkably intense. Sev- eral loci that were only weakly labeled in the cat contained unequivocal labeling in the raccoon. The overall pattern of labeling in the raccoon was very similar to that observed in the cat, and most of the descriptions below apply for both the cat and the raccoon. A different pattern of labeling was present in certain regions (e.g. paraventricular, subparafas- cicular, ventroposteroinferior n.). The pattern of labeling observed in several other regions in the raccoon provided comparative evidence that helped to clarify the observations made in the cat.
The distribution and relative density of the label- ing observed are summarized in Tables 1 and 2. The respective availability of corroborative evidence from the autoradiographic material and the presence of a topographical distribution are also noted in the tables. The labeling observed is described below with reference to Figs. 3-6, in which data from cats with

injections in only the cervical or only the lumbar spinal cord are presented in the transverse and horizontal planes at closely-spaced (ca. 250 ~tm) intervals. Figures 7-9 illustrate the labeling pattern observed in the raccoon. Labeling in the hypothalamus is not presented, due to the difficulties of interpretation caused by the large number of retrogradely-labeled neurons present.
Rostral mesencephalon
The observations made in the rostral mesencephalon in the present study generally concur with the recent study of Wiberg and Blomqvist (1983), in that the densest labeling occurred in the posterior pretectal n., the central gray and the n. of Darkschewitsch. The labeling in the latter in the present material was noteworthy in that it extended rostrally to the ventromedial aspect of the fasciculus retroflexus, where it lay adjacent to a dense cluster of retro- gradely-labeled cells in the subfascicular area (see Peschanski and Mantyh 1983). In addition, diffuse labeling occurred in all pretectal nuclei, particularly in the anterior pretectal n. (A), where a topographic pattern was observed. Labeling in A in cervical cases was located in the caudomedial portion, and in lumbar cases in the rostrolateral portion (cf. Figs. 3 and 4, level 0.00 mm).
Medial thalamus
Dorsal nuclei: H, MD, Pv, Pt. Labeling appeared in each of these nuclei, but no labeling occurred in the rhomboid n. and anterior group of nuclei. It was most dense in the paraventricular (Pv) and paratae- nial (Pt) nuclei, moderate in the mediodorsal n. (MD), and light in the habenula (H). Labeling in H was more apparent in the raccoon (Fig. 7), and it was denser in lumbar than in cervical cases; it was always concentrated in the caudal part of the lateral habenula n. (e.g. Fig. 3, 1.75 mm; Fig. 6, 4.25 mm).
Labeling was observed in three portions of MD. Sparse, scattered labeling occurred in the ventral and medial portions of MD (Fig. 3, 2.20-3.05 mm; Fig. 4,
231
1.70-1.95 mm). Dense labeling occurred in the most caudolateral portion of MD, which has been included in CL in the present study and is described in detail below. Moderately dense labeling appeared in the dorsolateral aspect of MD, dorsomedial to CL (Fig. 3, 2.85-3.60 mm; Fig. 4, 1.70-2.70 mm). While in the cat the cytoarchitectural characteristics of this latter region were not distinct, a separable nucleus could be delineated clearly in both planes of section in the raccoon that has been denoted the centrodor- sal n. (CD, Fig. 7). Labeling extended through most of CD in case N1 (Fig. 8, 4.35 mm) but was present only at its middle level in case N2 (Fig. 9, 4.25 mm).
Labeling in Pv was present throughout its lon- gitudinal extent, but was most dense at middle levels, where it was concentrated over the ventrolateral half of the nucleus (e.g. Fig. 3, 2.60 mm; Fig. 4, 1.95 mm; Fig. 6, 3.60 mm). Intense labeling was also located in a small cluster of neurons at the caudal end of Pv that extended vertically, adjoining Pf (only partially illus- trated in Fig. 4, 1.25 mm, adjacent to labeling in Pf). Labeling was considerably denser in lumbar than in cervical cases and was slightly more ventrolaterally located. Labeling in Pv often extended underneath the habenula toward Pt. Labeling in Pt appeared in scattered bursts that were most dense in its ventral portion.
In the raccoon, the area denoted as Pv was confined to the midline through much of its length (Fig. 7). It was not contiguous with Pt, except at its most rostral pole; and, unlike the cat, labeling did not extend ventral to the habenula between the separate fields of label in Pv and Pt.
Intralaminar nuclei: Pf, CL, Pc, CeM. Moderately dense labeling was distributed throughout the para- fascicular n. (Pf) along the medial, lateral and rostral aspects of the fasciculus retroflexus (e.g. Fig. 3, 1.75 ram; Fig. 4, 1.25-1.60 mm). Small patches of dense label appeared within regions of fine labeling (Fig. 2C). Labeling in dorsolateral Pf was generally less dense and had a different appearance than that in neighboring CL. Labeling in Pf in lumbar cases was distributed differently from that in cervical cases, but a topographic pattern could not be discerned. Label- ing in medullary dorsal horn cases was present only in lateral Pf.
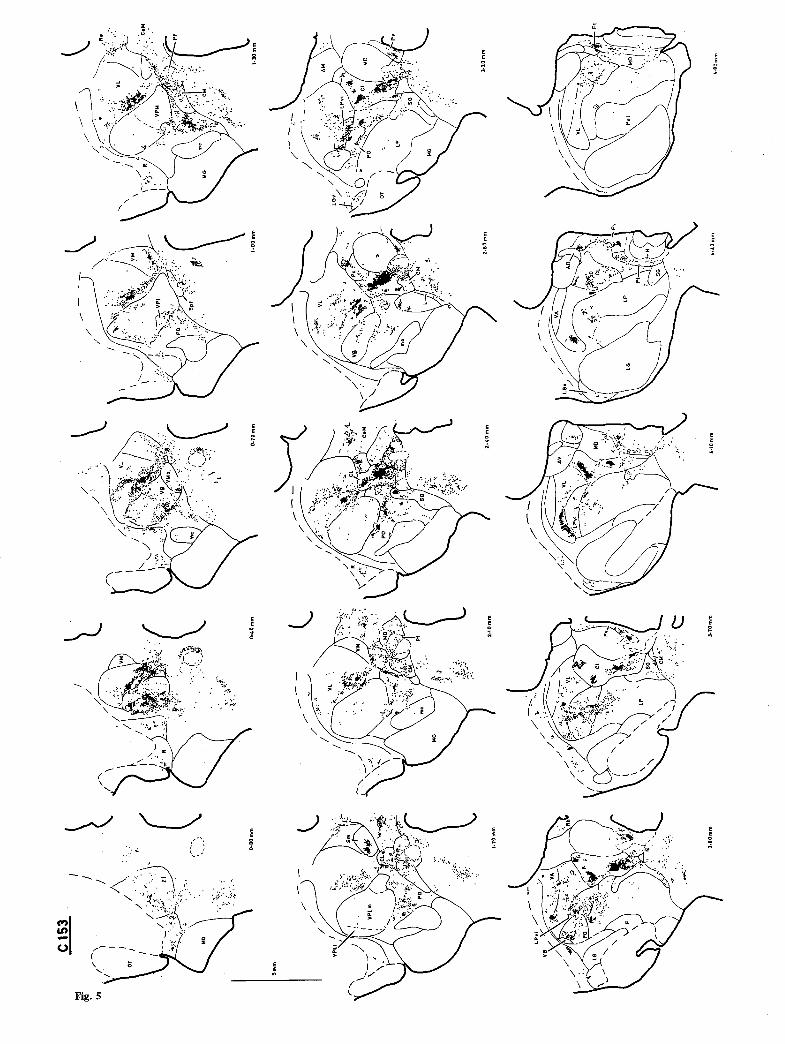
Fig. 3. The distribution of anterograde-HRP labeling in single 50 ~tm transverse sections in case Cl15, which had received a large (20 gl) HRP injection in the cervical enlargement (C4-T 0. Numbers indicate distance rostral to the most caudal level shown. (This figure and Figs. 4, 5, 7 and 9 were included in a preliminary symposium report (Burton and Craig 1983) and are reproduced here with permission.)
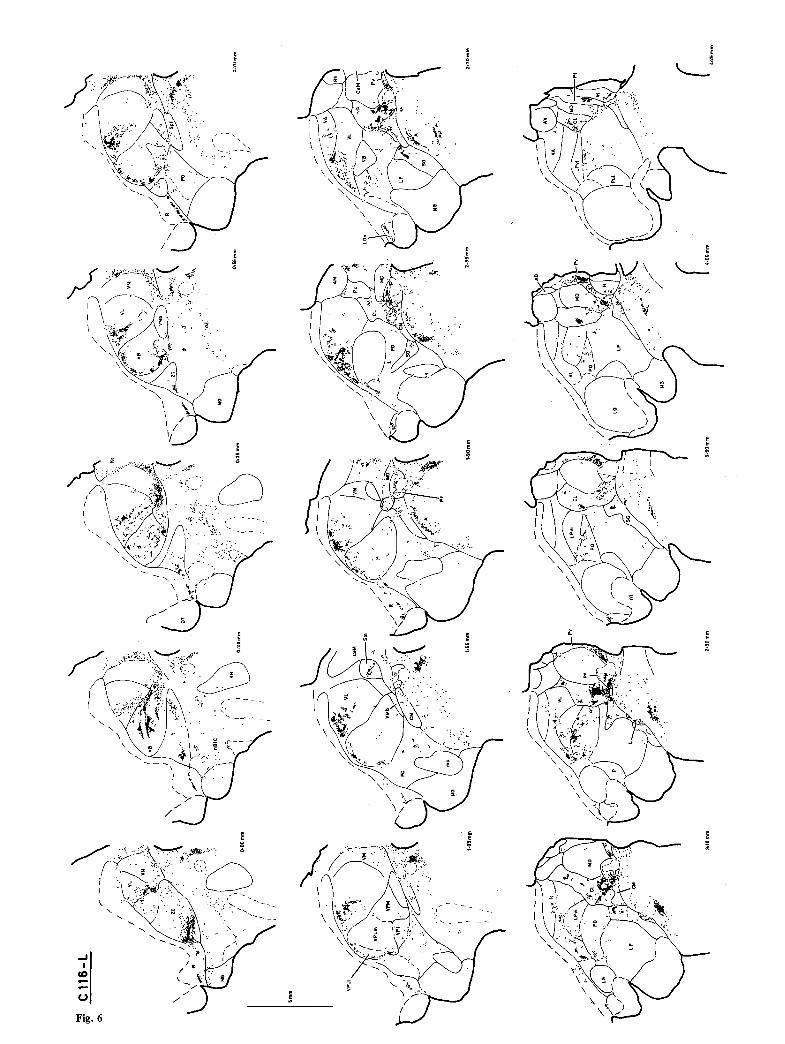
Fig. 4. The distribution of labeling in single transverse sections on the right side of case Cl16, which had received a large (10 ~tl) HRP injection in the lumbar enlargement (L5--$3)

�9 "
"',Z.
-,"-
:d~.
',.
; ,':
.ZW
" ..-
,. ...
. ..
. ,~
,. ("
~"
,..:
" ..
...
"'"
\ ":
:,"'
; .:
::,
�9
.Is;.?
�9 .:;+._
_,~.~%~
.... ..., ........
. ,
,.,.,. ,, ...... ..
, ,,
~,,~..::,~.
-
<"
�9 :.,
~"
"*'-~
i~("
-."~
" "~
. ~
i
Y " :
,.a.:
- �9
. . .
'.~:,
" "
."
. ...
. ';<
','
: ~.~
'.:
:.
::..
:,
""
..
,"~;
". .... ,i;."
~ t.-
"~:.
. '.
,.
'. ..
.. .
~
- ."~.,
.~.
. .A ~,
...
,:
. .~.
~ ";,t;
,,
~ ,.
, ,,
....
~ ,,
.,,:
~ ~:~,
\ ,
-, ,,
.-~
'-
.,
,,.
.+-"
,iX
-
�9
~ ,
,' ..
.Y
..
. �9
..,
,~,.
."'
~:
.... �9
",~ "
: ~,~
: ....
"~;~
. ".:
. ",
.~.:"
.a
" ~"
:, :j:
."..':
.,'t
,:"\,.
~'-"
~;.
' x"
;';
~ <~
~'" ...
.. ,,
. ~:
: .
, ,.
,'~!
~..-.
...
,..~
.,.~..~
:. ,
. ,_
'..~.;?
. ..,,
.<, ....
~;
:i~:.:
:'i.-~
,
....
..
"-,~
., --
. ,.:~
;.. ,..
, ~.
,..
:,....
..
..
..
..
,r
...
:. ~-
~,
,. .
~i
~
.... ,,,~,,
, ............
,,:,
~ ,,..,~,,:
: ...... ~
" -,
..
..
,...,.
.. .,
...
. ..
, ..
.
~...:~
:.'.~
i. ~
~.
,:,~
~.:~
~
�9 ~
<.~:
~,-.
~ :.
o-
i:;l
:~i...~ ~ I ;~ ~
\X.~Ag
~ ~- ~'~
~L~'x
~._;, ~
i~ ~
~ '=, ~i~i\ ". ~--
%-.
~'~'.;Y~ ~
:~.~
,c',';~
\~,~
..
. -".
�9
il. ~
" .~.~
~.
. ."
' -.'~
" "
":"
>~:~X
>..~
.,. '
�9 I
�9 : .~..
\ "
..-~
+~':
~---
--~.
~-.~
~i
�9
< \v".---~h
~ .---~,--~
.A.~
\ ,, ,".
.~'X
..,~
...:
\
.,,..,,..., ~
~&
"--
\ ~
.... �9
...~
..d
....
....
....
. �9 .
h~.
..
..
..
.
~ .,~.
.~,
�9
.,.
,,.
~. ~
..-...~..~
, .
~.>:,, .....
i~.~..
":,
".~;
."'.
'~ .....
-~""
~,.','#'.:~,!'.."~
.,'..'.,.~'...,~,'.".~,.'UY
".,~
'."
,'....'~..,".
, .',,..:, .,"r.. ....
.,. ....
..
\ ~-~.:<:,".,,~
, ..':. ~'.,',:,
. .c,.,,..". ......
'. ~,~
.~.
' ..'~:,..,. 1 �9
. ...
. ,:,' ~:...
\w,
....
...
\ ~
"\
~.z'".
~"
'h~;
'". .....
'. ~
. Y: ..
....
..
" "
" ~"
'" ...... .~."
\ ~
X.~,
.....
~ .
., .
..
..
..
.
..... . ....
\..
,.
.,., .
..
..
..
-~...
' "
:';'! ! ":
~'....
..,":~: :<:'
\ "
~ WT".
:.
~'.,
..
,,i'i.:! ~
\~i.,
" ' i'~;''"
< -A '.~-:~.':~;".Z..~, .,.:,~
~"
~
~,,~.. ~:," ,.
l "\ 7 ~;.,.!'
' A
'~"
" ;"
"
"""""
"'" .... "
\ ~
�9 "
X':. ""
\ -~
.... "..<
|

2 3 3
, / ' ' ~ ' " .".,:7 " - . ', .~ / I ~ ' ' " ~ T J t
- ' . L - �9 . ~ .
.f'. ,;- -." ~.:-'::
/ / /
J
s~, / ~.#~- / ::'.~"..-
�9 ,~<,..,,: ~ . . j ~ , ~: ~L'..'->:." . . . . . . / " C l M : . : . . . , ' . " . . . . ~ . :"
%,i<~:-..~. >---
Fig. 4
f - ~ x .I
" - ~ . ->--~.< <,.~ / K ") W , , ..
~ , , 5 . , .. ". 7
'n~i~ ' - - " , w c , .
\
.,, I~~/ " ~ J / , , ' j ~
I
(q'f.. - ; -, I ~ "%C ~ "Y '%, ."
" "o.... "" ;7
- j /.,,r ~ / , . - ~ ' " ,,,', / /
i ,
2 . 3 5 m m
\
-% 3 . 1 S m m
\\-
. . . ~ /
,,,.. VL
/ /
"X
/
1 . O O m m
\ \
, /
l , ~ . :~ : O L . : :~s , : - .~ ; " , .....
.~ /..- . .'I/ ,,./>-~ \ ,J~ .~ , lV_~., h,
~..--/ , x: ; ' .~ WY" I' :-~- .---:"1.
\ ~ " ~ - - s . O o m
":":'.~:" i ..... ~ . �9 : 3 . 6 o . .
�9 . \ \
. . -~ .~ . . . ~ -" ~ . . . " /
.(.-.T'.TL... i ..... ' .....
~ ^:. I
~ 2~y ./.,<;
\
\
I I/
C 1
53
t l
/. ~''''
I I ,_/: ":...:.~'~:...!
/ ...,-1
~ %/~W.":-~/'_.,I_ \
,-~.. d.
, ,
.~.--.-.<.W....,.'.-'.
"-~":
~ .... I
,-..-
,,'
~:,
: ....
",,-,:', ..
..
..
.
" .--'
-,-.
.,
J'"
....
i"
~. ~
.....
.,,'.,..
zy.__.~<i-.,;>.~.:~.i_~:.
":'-
--""
I ,..~"?;'
- ....... ,~ {,
' -,,-~L~,'. ,
..
..
.
',...>
,~, . ..:
,.:....,. ,,,,..: ...
.~
--
- '
,-,.
...,,;.,.,
,..
- ..:,,, ,<
. .....
-:.
,...
,...
. ....
�9 '.
,,~
,,,
:~:." ....
"''~c~
:."
spf
.....-
~ ..
....
:-.
. ;,
~
,,::}
\ �9
m
m
~
�9
mm
~
|.
$O
mm
/"-"
.....
:~/:"
/ �9 ';"--' :<"
~o,
" :.'::,i}
"
~'"\
/I1",
~- ~!~:;~ ~,,-,\~"
-,,-
- ),~
,' ,,,
~o'
~-."~
"-"."
,~
,x,.
~'--
, ",
.
%.-
.,
'Of
.'."
. \
..,
~."
--,
',-""
'.".'.
-x
",
-'..
/
k '
~..~
..'"
'!-".
~"
- " .
..
..
',.
.....
"'
' ..
..
..
..
.
,o
" ...
....
::~
\
~ ~ ~r
..
..
..
..
..
..
..
..
..
-.,
..
..
..
..
..
.
,
7 ~
"~:.
::'"
~::/
'.'.
~,'%
~.: 2"
""J,
"~'~
"--'I
' ,
.,
..,..
1 .
~ ....,,..:,
,,
q......,.
,,.~,
~._.
?,'
[ \
~~
~'
"..
" . ;W~"
L ~ ': .'-
" ~"'~'
W-.+:"" -
:""
I,?
Om
m
:. ~ ~
,o W
"...~
.::;"
>~
~ ~
I/'';"
,_,
,:,::.Q
'k,
',)" ~.(
,
" ": ";L.--
:~
x.\
\ .
;..:
3.w
~
3.70
ram
~
- m
m
~ 6.
.~0
mm
6.
co m
m
I " '" " '" "" :'~:.;" '
" ~ ( . - i::.~:' \ ' : " ~
\ \
.. y . . o.
I ~ "" . " "'~
-_L_.I '~
(;.~~..<U_b t, ,, ~... I .... ) ~ S . . . ' . , . , ' "
�9 . , . . . - . . . - - - - , ~ , . . > . . Y - - - ~ ~ . . . . . . < ~ i " " " " " " ~ ' " " " \ ~ . 2 ~ ',:'.':.".~ ...,X �9 " ~ ~ ' ~ " ' : "; "
" ' " " , ~ / ~ ~ , y ' . . ~ i " ' ~ . . . . . . �9 "
6 \~;\ ~I ~

236
The densest labeling in the present material was located in the centrolateral n. (CL; Fig. 2A). In those portions of CL where the labeling was most intense, it completely surrounded all counterstained neurons. The labeling was unevenly distributed amongst the several neuronal clusters of CL (Rioch 1929). Certain clusters contained dense labeling in each case, whereas adjoining clusters often had little or no labeling. In addition, the appearance of the labeling often differed in adjacent cell clusters. The coarsest labeling, in which large-diameter labeled axon frag- ments were always visible, was usually associated with clusters of large neurons in CL; labeling that had a finer, granular character was found in clusters of small neurons. Labeling in cases with injections at both cervical and lumbar spinal levels was clearly greater in its extent within CL than that in cases with injections at either level alone, indicating that some degree of topographic segregation occurs. Labeling in several portions of CL was observed predomi- nantly following HRP injections in either one or the other spinal level (cf. Figs. 3 and 4).
Three particular labeled portions of CL were distinct in all cases. A cluster of large cells appeared ventrolateral to the rostral portion of CM that was continuous rostromedially with the remainder of CL (see Fig. 34 of Hendry et al. 1979). Coarse labeling occurred in this cluster in both cervical and lumbar cases (e.g. Fig. 3, 2.20 mm; Fig. 4, 1.80 mm; Fig. 5, 3.20 ram; Fig. 6, 2.70 mm). Dense, coarse labeling was present in each case in the large-celled lateral cluster at the middle rostrocaudal levels of CL (see Fig. 11 of Jones and Burton 1974; also Itoh and Mizuno 1977). Labeling of lumbar origin was gener- ally confined to the rostral, dorsal and caudo-ventral aspects of this cluster (Fig. 4, 2.70-3.45 mm; Fig. 6, 3.60 mm), while labeling of cervical origin was concentrated in the center of this portion of CL (Fig. 3, 3.60-4.00 mm; Fig. 5, 4.10 mm). Finally, the most intense labeling in CL in these cases was distributed within its most caudo-medial aspect, where CL adjoins both PF and MD (Fig. 3, 2.00-2.60 mm; Fig. 4, 1.70-2.35 mm; Fig. 5, 2.40-3.50 mm; Fig. 6, 2.70-3.60 mm). A considerable portion of this label- ing occurred within a cytoarchitecturally differenti- able part of the lateral aspect of MD, as it has previously been identified (see esp. Rioch 1929). This labeling was continuous in terms of density,
appearance, and distribution with that within CL. In the raccoon, this region was more easily discrimin- able from MD in both transverse (Fig. 7, 3.00 ram) and horizontal (Fig. 8, 2.85 mm) sections.
Labeling in the paracentral n. (Pc) and the centromedial n. (CeM) occurred throughout their rostrocaudal extent in dense bursts. Large diameter labeled axon fragments within these bursts were oriented along the mediolateral gradient of Pc. In cases with combined cervical and lumbar injections, the labeling extended almost continuously ros- trocaudally throughout Pc, but in cases with injec- tions at either level alone the labeling appeared as separate patches (Fig. 5, 2.40-3.20 mm; Fig. 6, 2.35-3.10 mm). This pattern was particularly clear in the raccoon in horizontal sections (Fig. 8, 1.40 mm; Fig. 9, 2.30 mm).
Finally, in contrast to the dense labeling in all other intralaminar nuclei, the centre median (CM) generally contained only fine, isolated chromogen granules that were diffusely scattered throughout CM as observed in each plane of section. With rare exceptions, these did not occur in clusters. However, small bursts were present in the raccoon (Fig. 7, 2.00 mm).
Ventral nuclei: Sin, Re, VM, VMb. Labeling in the ventral portion of medial thalamus was most dense in the n. submedius (Sm), as detailed in the preceding report (Craig and Burton 1981). It was essentially confined to the dorsal part of Sm; no label occurred in the recently delineated anterior part of Sm (Craig et al. 1982). Labeling in Sm was topographically distributed rostrocaudally, with trigeminal labeling located most caudally, and cervical (Fig. 3, 3.40-4.00 mm; Fig. 5, 1.70 mm) and lumbar (Fig. 4, 2.50-2.90 mm; Fig. 6, 1.55 mm) labeling located successively more rostrally.
Labeling in the ventromedial n. (VM) was con- centrated in the ventral aspect of its caudal half, where it extended mediolaterally from CeM to the basal part of VM (VMb) (e.g. Fig. 3, 2.40 mm; Fig. 4, 1.70 mm). The labeling was of moderate density and it was fairly uniformly distributed, except for occasional patches of greater density (e.g. Fig. 4, 1.80 mm). The most caudal, interventricular portion of VM was differentiated neither by the distribution nor by the appearance of the labeling.
Fig. 5. The distribution of labeling in single horizontal sections in case C153, in which a large injection in the cervical enlargement was made. Numbers indicate distance dorsal to the most ventral section shown
Fig. 6. The distribution of labeling in single horizontal sections in the left side of case Cl16 (lumbar injection)

237
Labeling in VM extended rostromedially along its border with the n. reuniens (Re; Fig. 3, 3.05-4.85 mm; Fig. 4, 1.95-4.05 ram). This labeling was of moderate density caudally; it became sparser at rostral levels. Occasionally the labeling extended well into Re (e.g. Fig. 4, 2.35 and 3.60 mm). Labeling in lumbar cases was much denser than that observed in cervical cases, and in some cases seemed to be slightly more medially placed along the VM/Re border (e.g. Fig. 5, 1.30 mm; Fig. 6, 0.70 mm).
At the caudo-lateral extent of VM, a dense horizontal band of labeling was concentrated over a region of lightly-stained cells just ventral to VMb (e.g. Fig. 3, 2.20 mm; Fig. 4, 1.70 mm). The labeling frequently extended dorsally into VMb, especially in medullary dorsal horn cases. The continuity of the labeling in this locus with both the labeling in VM and that in the ventrolateral n. (VL) was clearest in horizontal sections (Fig. 5, 0.40--1.00 ram; Fig. 6, 0.35-0.55 mm), in which well-labeled axon fragments were usually observed coursing rostrally towards VM or VL. Thus, in this particular instance, fibers of passage may have contributed significantly to the labeling observed. However, labeling in the neuropil of this locus was in each case considerably denser than in the rostrally adjacent regions (e.g. Fig. 6, 0.35 mm). In the raccoon, labeling occurred in two additional nuclei in the ventral portion of medial thalamus that were unlabeled in the cat, the sub- parafascicular n. (Spf; Fig. 7, 2.00-2.60 mm), and the rhomboid n. (Rh; Fig. 7, 7.25 mm).
Lateral thalamus
Ventral thalamic nuclei: ZI, R, LGv. Labeling oc- curred in three of the nuclei of the ventral thalamus, i.e. the zone incerta (ZI), the reticular n. (R) and the ventral subnucleus of the lateral geniculate (LGv), while no labeling appeared in the subthalamic n. The labeling in each of these nuclei was much denser in lumbar than in cervical cases.
The dense labeling in the caudal half of ZI seemed to define, in transverse sections, a single horizontal "plane" through the middle of the nucleus (e.g. Fig. 4, 0.75 mm). Occasional retrogradely- labeled cells were also present in ZI, but these were located caudo-medially, separate from the antero- gradely-labeled field.
In both transverse and horizontal sections (e.g. Fig. 4, 0.00 mm; Fig. 6, 0.00 mm; Fig. 7, 1.55 mm), the labeling in ZI was tenuously continuous with labeling in the cando-ventral aspect of R. The labeling in R generally had a coarse appearance; labeled axon fragments oriented along the dorso-
ventral path of the neuropil were often present (Fig. 2B). The labeling did not extend into the internal capsule. In horizontal sections, labeling in R appeared as repetitive patches over cellular regions, which were separated by fibrous zones (Fig. 6, 0.70 mm). Occasional labeling was observed in the rostral portion of R (Fig. 3, 4.20 ram; Fig. 7, 7.25 mm). Labeling in R also extended into the ventral part of the lateral geniculate (LGv), where light, fine label- ing was observed (e.g. Fig. 5, 3.20 mm; Fig. 6, 2.70 mm). Labeling in LGv was confined to the anterior and ventral portions that adjoin R.
Caudal nuclei: SG, L, inc. Several distinct patterns of labeling were observed in the caudal portion of lateral thalamus that often coincided with cytoar- chitectural differences. The labeling in the posterior group (PO) is described separately below. Light labeling was distributed throughout the supragenicu- late n. (SG), but moderately dense labeling was confined to the discrete clusters of darkly-stained cells that are characteristic of this nucleus. The labeling completely filled some neuronal clusters, but only a portion of others. Fine, diffuse labeling gradually became more dense rostrally in the dorsal extension of SG (Graybiel and Berson 1980; Fig. 3, 2.00 mm; Fig. 4, 1.45 mm). Label was present within occasional strands of cells that extended dorsally from SG, then laterally along the dorsal edge of thalamus, and finally ventrally to form a pocket of cells in dorsomedial LP (e.g. Fig. 3, 1.40 and 2.00 ram; Fig. 4, 1.45 mm). However, the labeling in this dorsalmost region otherwise did not occur within distinct cytoarchitectural zones, and it continued rostrally into MD and LP, i.e. beyond the area that could be identified as SG in either the transverse or the horizontal plane. Moderately dense labeling also occurred in an elongated cluster of deeply-stained cells (n. limitans) that extended rostrally and dorsally along the meso-diencephalic boundary to the region of the habenula (Fig. 3, 0.40-1.75 mm).
In general, labeling did not appear in the mag- nocellular portion of the medial geniculate (mc). Labeling in the vicinity of mc occurred in the medially adjacent PO, similar to previous descrip- tions of STT terminations in this region (Jones and Burton 1974; Berkley 1980). The labeling in PO only occasionally infiltrated the mc, and isolated labeled axon fragments were infrequently observed near large somata in inc.
Nuclei adjacent to the ventral tier: PO, LP. Labeling in the posterior group (PO) extended from its caudal pole through the rostral portion of PO (which lies dorsal to the ventroposteromedial (VPM) and ven-
~'g.
~ -
-
,.,
, =
~ :.'
9....
,,,P
x
..
..
"
' "7
+" .-'
,-.
- ~i
~ ~ .
+:
~, ~
-,?.>,
-o "
,' T"
r ..
.,
:!
-.
..
.,
+,
'~
%
.:::
~'-
,
":"
.,..
,.-
..
..
.
:."
., ~.
�9
g
, "~
:+-+
...,
. ,.
,.
....
.k?~
,.
.. ,
~.
...
..,<.
.-~,
.,
.~
.'
',"~.
.C~-
. �9
,'.,.:
,~
:,+.,e
--".;.
, �9
....+
~ \
...~.
....
,~."
~ "
�9 =
. �9
"v,.
....
�9
..
..
: ~,
~
."
.:
' ~
. \
......
. ..~
'.-
:
\ ..
..:.
.,..
. ~
. .
. ....
~
+-"
, ~
- ::
.~;:
~
�9 ~
)..~
' ,,
"~
'.-,
,,
..,.
. ,Z
,>:
"+
.-...
,,+,~
..:_~
.-'~
:<
.... :
~,;
..-
+ .,.
�9
-,
, ...
..
. ...
.. :,
, ..,+
- ~
....
...
i...
~
, ~.
. ....
+,,:..~
+. ,~, �
9 :,..
,;..>, .
.~~.
.,
7 .
,~
~.
,
�9 -.
,.: ....
. ~~~.
.,+,
,-.,
~.
,.:.
..,.
�9
...
. ~
�9
.~.~
: :
....
�9
,
.,,
.;~
...,
. ~,
. ,
...,
.
.,.
::...
-:
..
+-
+"
~ =,
+ ,
M
+ "-+ ,
v
o -
%
~\ \
~.~"
f ...
.. ~~%
.~..c
. .:.
- +
�9
,~..
t...
. ~.
,. :.,
~-
~
.~..
~
,,.-.-
>. .
..
..
..
0
* ~
" "+
i'.
""
" 0
~.+
. *d
=.
. :..
q
"7 "
Ne,
+"
- -
"I"'
"
....
":
. "-
I.l,, tl
M..
_~
..k
:= M
.
n g
~ g
.~.
; -,
; ,.
E g,"
e~
t,..a
o,I
( l-'-c.;.f,-<.. \
~ \
_ \ ,',
,~,r,~..~,.,~-'.,\ I
,-7 ~,-,,..k' ",-",-.':
~~:,f~".~. ". -.-- ~ '
"-~.
�9 -~.' I
I '~
~'~ ~
, "
"~
i~ "~'
""- ~::" \
~, \
~~
g\
\
~ : ,.:
:-~.--.-0 �9 ~ .~\',
~" \-
-.",....~J:2~ ,,
,,. ...:-.-....: ....
, .
..
..
.
~-, ...,,,.-.:,
.,,,,, ,..
-,~- i,~-
t~.kr-~ �9
�9 ":'.,~:-.:r..~-~.~.~,,x /I
\.~., I
- ""~'~.'r'-"t.'.,'~,:
/ "-<_
I .
%
'/."" "
. .."..,,"~... '
- "
~ .."
:: ~b..',
"-'.." .-,..
r �9
~,,'" .... "'"%.
~ .'..,
-,: .... I
"~, .
..
.
.- "..
�9 ,.~/., -
. ~,--;
.. ~-c-......;-,"
. ,~:
...~ ~-:.-
., ~
, .~
{-~<--
..
..
----
_ ..-- ~
-'--._-~ ~ "~
/f,,~ -
..z_";2~\ ~
"~
',
"~
"~
-
" -'~.""~
"~
/,,~,
2- ..
" .... ~
\ \
. .
~.
'-
"i'
- ....:.,.,
. .'Fl.~,
!:-:., �9 "
�9 .-.~
.,... -
�9 .
..:~.-.,.. ,,
~;
~
;.. ,~, :-~;~..,
,,.
�9 .~..: ..;,s,,.~ "
"( "J
X
i' "
~
..
..
,~t k..-,
. ~.-
..-.~,....'.'. ~,
: "
..,: .--~-.'..-'..~-',~1i"..
"" .~'t.':~.':
.-~ "~
: "
" ,.",.. "..'..'.
'" .
"~'. ,'-'...'::
'" ~
-' :".
"' X,
.t �9
.'...'z ,
" ~.
:, ,,
..,:" ,
..... ,
,,~,~,J ,.
..
..
..
.
.., ~.....:..
... :~
:~ ...:
. -,,-
~1 ~!:"..-.....:..~.~,-:.,~.~ _
~.i: ": ~i~
- ~":-~.
~,, ~
~ :"; ' ]~:~:b '~---,
I
N1
-L
I>J
~"
~/
j
C/
,7"
"...:.-
," ~ ~'.,,'-Ii ,~ ,,"
�9
\~I
�9 ..
",~;
.,'.
e~\
\
-', ;":"
i'lL: :~
----
""
'~1'
x~
,~ PO
"'
0.00
mm
~"
'~
1,~0
mm
~
2-25
mm
r
~,~
,~
.::?,~',--..... ~
...,..~.~,~;~r.~.~...'~,
I /%
~. ~:
- \
.~;:~
.~,~,:
:~:"~
I ~
:
~ .. ..
~ ~
~, ~..."
.':.
. m.:"
\ ~,
" .~,,,. ~. %.,.
'..-
. ~..
. ,
7" ":y.---'~'.'~"."':~: ,.
..,.
, .
..
.
~" .
~..~.-.
- ...: ........
"'.:.,~i.:.~.~i :.."~ .... "
,
~i~.,, /
- ..~,,.~;~..~
~I ~~"
.:,~
"-:"
?":~
, �9 I 1
[.
~-~
~ :.
,:~ ~
.~c.
"~'~'-
*"':~"
~::" ''
~<""
:
-::-
..,. ..
.. .,
,.~
'LO
y ~'
~""
';:~
IX
:': "
'"."""":-~-:-, "L;"";' ~''
I
Fig
. 8.
The
dis
trib
utio
n of
lab
elin
g in
sin
gle
hori
zont
al s
ecti
ons
on t
he l
eft
side
of
case
N1
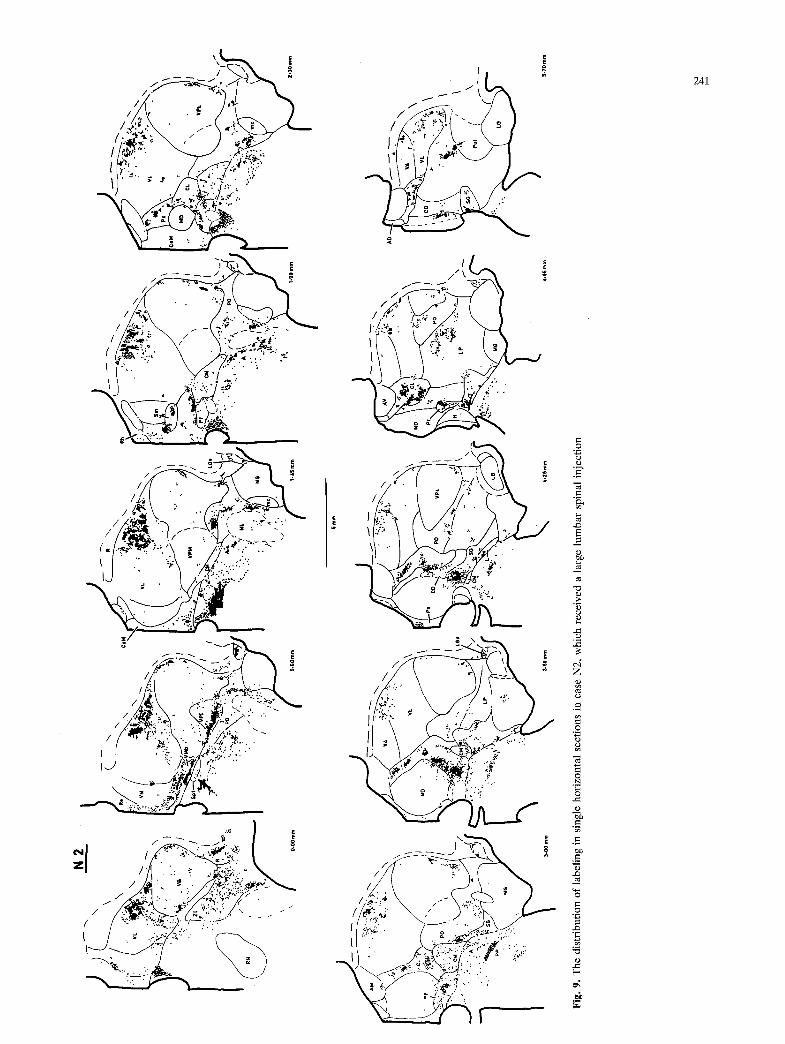
N2
.f,"/~
.,~ \\
"-
I ~
::,~
, _
,, "
" ,
9 ~
~I:.
~:-~
_.~:
'~,-
~:
71
~....
..~-<v,,.. �9
..
'- +".
.+'-'.-
.'~:.~
"--I'.:
;/ ~-
I :'
- ~':
-.:\,~
o '.~
,,,:X
;~.~
:,,.\
\ /
) ~-
!::~::
.. .':~
I :;:".'.r
�9
..
.
~"
. ~,
,...+
.---.~
,
LLV
,'- , .':~,,~,,~'
o.oo.. I
"-,, ::,,,..~.!~:'~~
~C
~\
g
~
~ ~
..
�9 ~.
: ~
"~\-r
...-'? ~
"~..
'-
,~-~
k.
r .~
..~"~
~.~
r +..
.-:~:.:..
~ .~
' IL~
Z~-
....~.
_~"/"
~,,
I, ::
~-~
~, "
." ~
, I
~,,
...~
.
�9
I' ,
-~w~,,.':....:
~A.
~.,A F_! J?-",: .~,~',>~\-
:~
::: .:: ::
... . .- .'.
,: :: "
,~
,.~r
":":::'
,.':'",.
-,q~M
, : :!
:.~
,. /
-f/
-..:.~:
,~-':
( J"
I
l ...
.. i
f -
'
....
.
i~~~'~"
";"
" ~
" " "*"
\ ~
~~
~
~"
l ~" ~Y.'~ :". ii~ " ~x'q
""-~ -~
..
..
..
i.oo
mm
~
3-ew
,, "
":a,,!
Fig.
9.
The
dis
trib
utio
n of
lab
elin
g in
sin
gle
hori
zont
al s
ecti
ons
in c
ase
N2,
whi
ch r
ecei
ved
a la
rge
lum
bar
spin
al i
njec
tion
~+.,
.
.-, y
.%
I'."..~ .:.",..., Y~..
"
"~"~'"
" -~
x
,qf
.......
~"-%~" "
h~-
"----'\l .' l
�9
~.:.
.~.'~
/
I A
JJ

242
troposterolateral (VPL) nuclei) and into the adjacent ventral portion of the lateral posterior n. (LP). The labeling was unevenly distributed through PO, and three separable patterns emerged from the differ-
ential distribution of PO labeling in cervical and lumbar cases. First, dense labeling appeared in the caudal portion of PO adjacent to the medial lemnis- cus (ML; Fig. 3, 0.00-0.95 mm; Fig. 4, 0.00-0.75 mm; Fig. 5, 0.70-1.70 mm; Fig. 6, 0.55-1.90 mm). The labeling extended dorsally towards SG and rostro-medially towards CM. The strong rostro- medial orientation of this labeling was most apparent in horizontal sections in the raccoon (Fig. 8, 2.25 ram). Labeling in lumbar cases generally occurred ventrolateral to the portion of caudal PO labeled in cervical cases. Light, diffuse labeling extended rost- rally from these dense patches. At the caudal aspect of the ventroposterior nuclei (VP), a second region of moderately dense, patchy labeling occurred. In cervical cases, these patches were most frequently found medially, near the PO border with VPM and the medial part of VPL (VPLm; e.g. Fig. 3, 1.75-2.20 mm), where little labeling was present in lumbar cases (e.g. Fig. 4, 1.25-1.60 mm). In lumbar cases, labeling appeared more rostro-laterally near the PO border with the caudal aspect of the lateral part of VPL (VPL1; e.g. Fig. 4, 2.35-2.70 mm; Fig. 6, 1.55-2.35 mm). In both cases, isolated patches of labeling also occurred in the most lateral portion of PO, rostral to mc. The clusters of labeling at the caudal aspect of VP were especially numerous in the raccoon (Fig. 7, 1.55-2.20 mm).
Third, patches of moderately dense labeling occurred within a broad field of light, diffuse labeling in the portion of PO that extends rostrally, dorsal to VPM and VPL. These patches were often aligned, forming rods of labeling that extended rostro-later- ally (e.g. Fig. 5, 3.20 mm; Fig. 8, 3.70 mm). Well- labeled axon fragments (Fig. 1) were often present. Labeling in cervical cases was concentrated caudo- medially (Fig. 5, 3.20 mm), and labeling in lumbar cases generally occurred rostro-laterally (Fig. 6, 3.30 mm). Only infrequent labeling in this dorsal part of PO was observed in medullary dorsal horn cases.
Labeling extended from the dorsal part of PO into the adjacent rostral half of LP. The labeling within LP was generally light, with occasional denser regions. It formed a conical field, the peak of which reached the border between LP and the pulvinar (Pul; Fig. 3, 2.60-4.00 mm; Fig. 5, 3.50-4.80 mm). Moderately dense labeling occurred particularly within a thin vertical sheet of darkly-stained, medium-sized cells located between LP and Pul. In the raccoon, labeling in this distinct band of cells was
quite dense (Fig. 2E; Fig. 7, 2.00-3.00). The topogra- phy of labeling in LP paralleled that in the dorsal part of PO.
A second field of moderately dense labeling in rostral LP was observed in cervical cases (Fig. 3, 4.20-4.85 mm), but did not occur in lumbar cases. This region was distinguishable from adjoining por- tions of PO, VL and LP on the basis of its cytoar- chitecture in the cat, but not in the raccoon. The cells in this region appeared smaller and more tightly packed than those in the adjoining portion of LP; it was denoted the ventral intermediate part of LP (LPvi), consonant with the terminology of Rioch (1929).
Ventroposterior nuclei: VPM, VPI, VPL. Light label- ing was scattered throughout these nuclei; moder- ately dense labeling was concentrated ventrally. The light labeling, which consisted of small aggregations of chromogen granules and labeled axon fragments, occurred throughout VPLm in cervical cases (e.g. Fig. 3, 3.60 mm) and throughout VPL1 in lumbar cases (e.g. Fig. 4, 2.70 mm). Labeling was most often observed within the neuropil adjacent to the major fiber laminae separating VPM/VPLm and VPLm/ VPL1, particularly in the raccoon (Fig. 7, 2.20 mm and 4.00 mm). A burst of labeling occurred in a single duster of cells in dorso-medial VPLm in the cat that usually appeared to be partially encapsulated by an extension of the VPM/VPLm fiber lamina (Fig.10A). Labeling appeared in this locus in both cervical (Fig. 3, 2.60 mm) and lumbar (Fig. 4, 1.80 mm) cases, but was less dense in the latter. Labeling appeared in a similar cell duster at the dorso-lateral end of the VPLm/VPL1 lamina in lumbar cases (Fig. 10B).
In cases with combined cervical and lumbar injections, moderately dense labeling occurred within the entire medio-lateral extent of the ventral aspect of VP. The differential distribution of labeling between cases with injections at either level alone, and between the cat and the raccoon, indicated the presence of three separable fields. The most medial field was located ventral to VMb, and is described above. In the cat, a second field of lightly scattered labeling occurred in the ventroposteroinferior n. (VPI). Labeling was present throughout VPI in combined spinal cases. In cervical cases, it was generally confined to the dorsal half (e.g. Fig. 3, 2.85 mm), while in lumbar cases it appeared in the ventral half of VPI (e.g. Fig. 4, 1.95 mm). The labeling occurred in the neuropil of VPI, but not over the many longitudinal fiber bundles that penetrate the region.

243
Labeling in the region of VPI in the raccoon was distinctly different from that in the cat. In addition to scattered labeling, five very dense, distinct bursts of label occurred within and/or bordering VPI; three of these are visible in Fig. 7, 2.20 mm, and the other two in Fig. 7, 3.40 and 4.00 mm, respectively. Compari- son of the labeling in cases N1 and N2 in transverse sections suggested that at least two of these bursts could comprise separate topographic representa- tions. (These are: the burst seen in Fig. 7, 3.40 mm; and, the burst on the dorsal edge of VPI in Fig. 7, 2.20 mm.)
A third pattern of labeling was observed in the ventral margin of VPL. Moderately dense labeling was concentrated in the ventromedial aspect of both VPL1 (Fig. 11A) and VPLm (Fig. l lB). Labeling usually continued dorsally from the ventromedial corner of VPL1 along the VPLm/VPL1 lamina for ca. 500 vm, but otherwise the labeling did not extend into VPL farther than ca. 200 ~tm. Labeling also occurred along the ventrolateral and lateral margin of VPLI. In general, the labeling occurred in the neuropil around small fusiform neurons. These neurons were oriented medio-laterally, were more lightly-stained than most VPL neurons, and often occurred in discrete clusters separated from VPL proper by fiber bundles. This region was most distinct in the ventral aspect of VPLm in the cat (Fig. 11B). The labeled cell clusters were often aligned with the major vessels that pass roughly medio- laterally along the ventral margin of VPL (Fig. 11C). In the raccoon, labeled clusters of small neurons were more common along the ventral and lateral margins of VPL1 (Fig. 7, 3.00 mm).
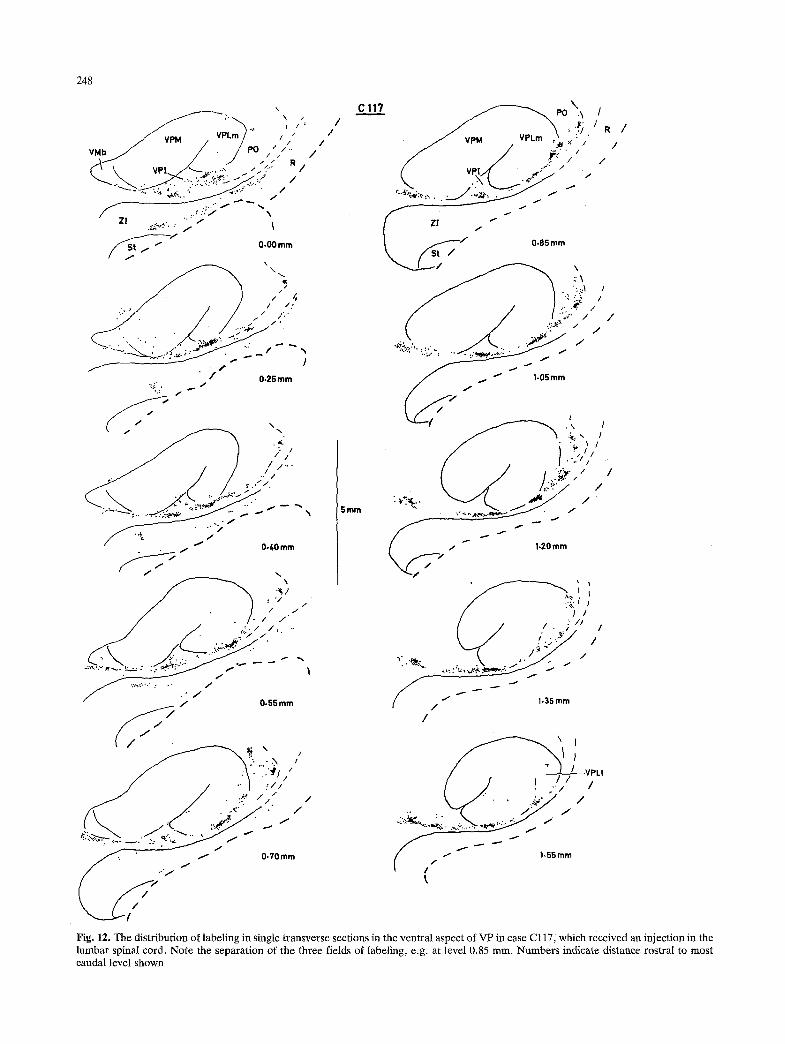
In cervical cases, labeling was concentrated in the ventral aspect of VPLm (Fig. 3, 2.60-3.60 mm), and did not extend laterally into VPL1. In lumbar cases, labeling occurred in the ventral aspect of VPLm only at its most caudal and most rostral levels, but it was present all along the ventral and lateral aspects of VPL1 (Fig. 4, 2.35-3.60 mm). Thus, labeling of lumbar origin formed a C-shaped field in the horizon- tal plane (Fig. 6, 0.20-1.55 mm), partially surround- ing the field of cervical labeling. This pattern is more dearly presented in Fig. 12, which illustrates the distribution of labeling of lumbar origin in the ventral aspect of VPL in greater detail. Labeling in lumbar cases was denser in the ventral portions of the discrete neuronal clusters in the ventral aspect of VPLm, whereas in cervical cases labeling was denser in the dorsal portions.
Rostral nuclei of the ventral tier: VL, VA. The densest labeling in lateral thalamus occurred in the ventrolat- eral n. (VL). Dense patches of coarse labeling (Fig.
2D) were observed in most of VL, except for the most dorsomedial portion. These were characteristi- cally interconnected, forming uneven rods of labeling that were oriented obliquely along the ventro-medial to dorso-lateral orientation of the neuropil in trans- verse sections (e.g. Fig. 3, 4.20 mm; Fig. 4, 5.00 mm), and along the caudo-medial to rostro-lateral orientation observed in horizontal sections (e.g. Fig. 5, 1.00 mm; Fig. 6, 0.70 mm). The labeling filled individual VL cell clusters, while intervening cell clusters contained only sparse labeling, if any, result- ing in the appearance of gaps between the dense rods of labeling (e.g. Fig. 5, 0.70 mm; Fig. 6, 0.55 and 2.35 mm). This grid-like pattern of labeling was most apparent in the raccoon (e.g. Fig. 8, 0.60 ram; Fig. 2D). The labeling generally became lighter and more diffuse dorso-medially in VL. Rostrally, the labeling was unevenly distributed in scattered, dense bursts near and within the caudo-ventral portion of the ventroanterior n. (VA).
The topographic organization of labeling in VL was clearest in horizontal sections (Fig. 5, 0.40-4.40 mm; Fig. 6, 0.00-3.30 mm; also, cf. Figs. 8 and 9). In cervical cases, labeling was concentrated in the caudo-medial portion of VL, while labeling in lumbar cases was located in the rostro-lateral portion. Thus, labeling of lumbar origin filled a caudally concave region that embraced the region in which labeling of cervical origin occurred. However, areas of probable interdigitation and/or overlap were also apparent. Isolated bursts of labeling sometimes occurred within the topographically inappropriate region of VL (e.g. Fig. 5, 2.10 and 2.80 mm; Fig. 6, 0.55 and 2.35 mm). In addition, at ventral levels in horizontal sections, an extended rod of labeling which curved rostro- laterally around the lumbar region of VL was regu- larly observed in cervical cases (Fig. 5, 0.70 mm). Finally, labeling occurred in both lumbar and cervical cases in the most caudal portion of ventromedial VL (e.g. Fig. 3, 3.75 mm; Fig. 4, 2.90 mm).
Discussion
The present results demonstrate that anterogradely- transported HRP can be observed within a large number of regions in the thalamus of the cat and the raccoon following spinal injections. The reproduci- bility, particularly for the cat, of both the distribution pattern and certain qualitative characteristics sug- gests that the labeling observed may correspond almost entirely to the terminal distribution of spinothalamic projections. This conclusion, however, is subject to several methodological constraints.
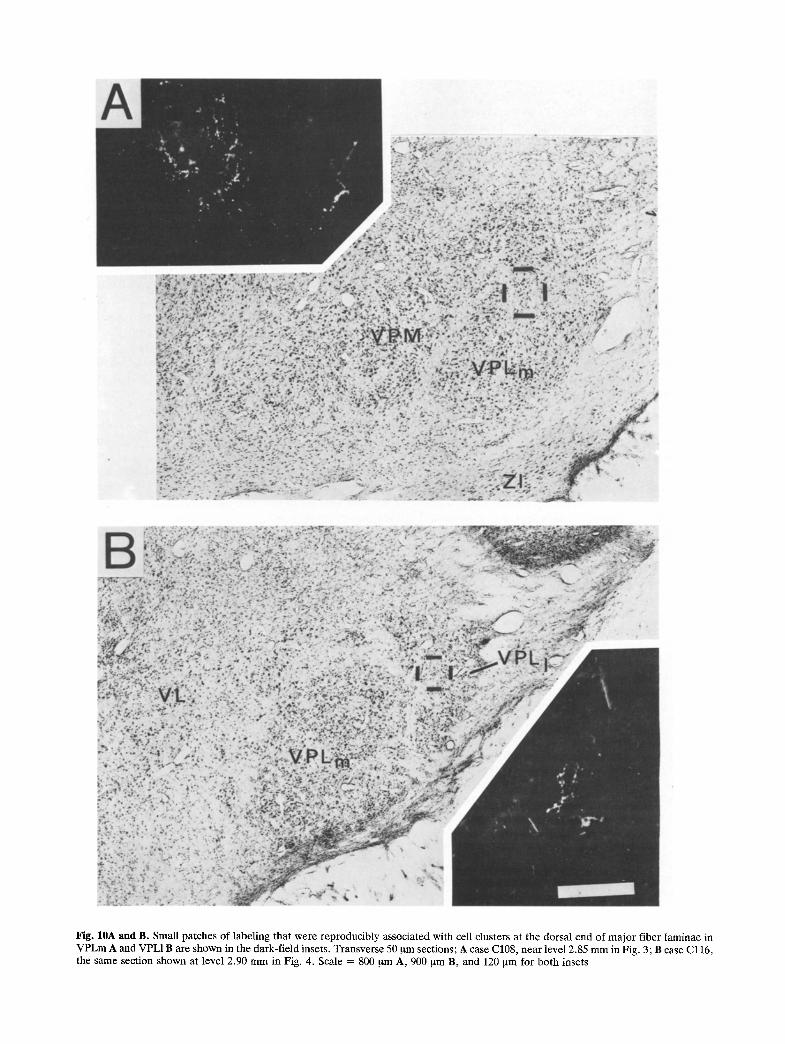
Fig. 10A and B. Small patches of labeling that were reproducibly associated with cell clusters at the dorsal end of major fiber laminae in VPLm A and VPL1B are shown in the dark-field insets. Transverse 50 gm sections; A case C108, near level 2.85 m m in Fig. 3; B case Cl16, the same section shown at level 2.90 m m in Fig. 4. Scale = 800 ~tm A, 900 ~na B, and 120 ~m for both insets

245
Methodological considerations
The anterograde HRP method is capable of demon- strating not only terminal projection fields, but also the path of the efferent fibers (Mesulam 1982). In the present material, however, only sparse labeling was observed in the identified locations of ascending spinal fibers (Boivie 1971). Labeling with characteris- tics consistently suggestive of the presence of labeled fibers of passage was noted in only one location. The circumscribed and disjoint nature of most fields of labeling, coupled with the location of the labeling over neuropil and the presence of well-labeled frag- ments of apparent axonal arborizations, supports the interpretation that these indicate terminal fields. These characteristics are consonant with the possibil- ity that, with the methodology and survival times used, transported HRP was widely dispersed within the ascending segments of axons, but rapidly accumulated without immediate degradation in the terminal axonal segments. Nonetheless, analogous to the difficulties of interpreting material obtained with the degeneration or autoradiographic methods, the identification of anterograde HRP labeling as "termi- nal" is inherently uncertain at the light microscopic level.
It has been demonstrated that transneuronal movement of HRP can occur (Hongo et al. 1981). It is conceivable that some of the observed labeling may have resulted from secondary anterograde transport within transneuronally-labeled cells, especially given the large amounts of HRP injected. However, se- veral incidental observations suggest that trans- neuronal HRP passage did not occur to a detectable degree in the present experiments: 1) despite consid- erable anterograde labeling in the dorsal column nuclei (DCN), the distribution and number of labeled neuronal somata did not differ from the pattern of retrograde labeling from spinal injections observed with less sensitive HRP methods (Burton and Loewy 1977); 2) no labeled somata were observed in regions of dense anterograde labeling in the thalamus (e.g. CL or VL); and 3) no anterograde labeling was observed in the projection targets of the latter nuclei (i.e. basal ganglia, pericruciate cortex). In contrast, transneuronal anterograde transport is detectable when HRP-bound and unconjugated wheat germ agglutinin is used (Mantyh and Pe- schanski 1983; unpublished observations). Nonethe- less, this possibility remains as well in the present experiments.
A serious ambiguity in the interpretation of the present data as spinothalamic and spinomesence- phalic projections arises from the possibility that anterograde labeling of thalamic projections of cells
with collateral projections to the spinal injection sites may have occurred (De Olmos and Heimer 1977). Neurons in many locations known to provide input to both the spinal cord and the diencephalon were heavily retrogradely-labeled in these experiments. Such regions include the rostral spinal cord (seg- ments C1_2), the DCN, the vestibular and deep cerebellar nuclei, various medullary and brain stem sites, the red nucleus, the hypothalamus and the cerebral cortex. However, collateral labeling from many of these regions seems unlikely. For example, the laminar segregation of cortical efferent cells suggests that thalamic collaterals of cortico-spinal axons may not exist (see Jones 1981). Similarly, rubro-thalamic cells overlap little in distribution and morphology with rubro-spinal cells (Cond6 and Cond6 1980). Different sets of neurons in the ves- tibular nuclei also appear to project to thalamus (Kotchabhakdi et al. 1980a) or to spinal cord (Kuy- pers and Maisky 1975). A small number of neurons in the deep cerebellar nuclei do have collateral projec- tions to both the thalamus and the spinal cord (Bharos et al. 1981), but the thalamic projection of these neurons overlaps only slightly (in VM; Sugimoto et al. 1981) with the labeling in the present data. Nonetheless, the possibility of thalamic collat- eral labeling via cells in the upper cervical spinal cord, brain stem and hypothalamus cannot be dismis- sed until the appropriate retrograde double labeling results are available.
Thus, it is clear that all regions identified in these experiments are in some manner involved with spinal cord function, but due to the above ambiguities it cannot be firmly concluded that all observed labeled fields represent direct spinal projections without corroborative evidence from autoradiographic or degeneration experiments. Spinothalamic (STT) ter- minations in PO, VL and CL have consistently been described in the cat in studies based on the autoradio- graphic or degeneration methods (Mehler 1969; Boivie 1971; Jones and Burton 1974; Berkley 1980). In addition, Boivie (1971) reported terminations in Pf, CeM and ZI. Labeling in CeM was also indicated without elaboration in the charts presented by Jones and Burton (1974) and Berkley (1980), and in H, VM, LP and (probably) Sm as well in Berkley's chart. Boivie (1971) identified terminal degeneration within the ventral margin of VPL in two cases, and Berkley (1980) reported consistently finding such evidence in her material, though she ascribed it to VPI. Summaries of the earlier literature have been made by Mehler (1969) and Boivie (1971); it is noteworthy that Nauta and Kuypers (1958) and Anderson and Berry (1959) reported an STT projec- tion to ZI, and that Getz (1952) reported projections

246

247
to LP, Pc and R. In the present material, labeling was observed in all of these loci, and in a number of additional loci as well. The present methodology appears to be more sensitive, andto provide a higher resolution of detail with less background labeling than the degeneration and autoradiographic methods (Craig and Burton 1979, 1981; Mesulam 1982). This fact alone may explain the greater extent of labeling in the present results, even though the possibility of collateral labeling complicates this conclusion. Yet, the foregoing observations and the presence of labeling in several of these novel regions (e.g. A, PP, ZI, Pv, Pf, CeM, Sm) in the present autoradio- graphic material together offer corroborative sup- port. Thus, the present results strongly indicate that the distribution of STT terminations is significantly greater in extent than previously appreciated. Further, it is reasonable to suggest that most, if not all, of the labeled fields presently identified represent the terminations of direct spinal projections.
Functional considerations
The large number of diverse STT projection targets revealed in the present material could be interpreted simply to suggest that the STT is a highly divergent and widely distributed system. Yet, the detailed anatomical structure and the topographic organiza- tion indicate that a considerable degree of anatomical and functional specificity is probably present. Thus, it seems reasonable to infer that each identified locus may receive input from a particular subset of spinal neurons, perhaps with anatomical differences analog- ous to those recognized for different sets of neurons in the visual system (e.g. Sherman and Spear 1982). Differences in the distribution of STT cells projecting to medial or to lateral thalamus have already been noted in several species (Carstens & Trevino 1978; Giesler et al. 1979; Willis et al. 1979). The observa- tion that marginal zone (lamina I) cells appear to be the exclusive source of STT input to Sm illustrates the degree of anatomical specificity possible within the spinal projections to diencephalon (Craig and Burton 1981).
Similarly, functional differences in the STT cells projecting to these sites may reasonably be expected. The available physiological evidence indicates that
several functional classes of STT cells exist (Willis 1983). STT cells have been identified in most studies only by antidromic activation from the caudal aspect of lateral thalamus, however, so little information is presently available regarding the projection targets of different classes of cells. In two recent studies, physiological differences have been reported in STT cells projecting to medial and/or lateral thalamus. Giesler et al. (1981) reported that many of the STT cells that projected only to medial thalamus had receptive field properties distinct from those of laterally-projecting cells. Craig and Kniffki (1982) found that lumbosacral lamina I STT cells in the cat responsive to innocuous cutaneous cooling project predominantly to medial thalamus, while those responsive to both cooling and noxious cutaneous stimulation bifurcate and project to both medial and lateral thalamus. Further work in this direction is clearly needed.
Analysis of the functional significance of the diverse STT projections without detailed knowledge of the functional characteristics of the cells of origin of each projection can only be based on the available knowledge of the connectivity and functional charac- teristics of the relevant regions (see review: Jones 1981). Certain previous attempts to infer the func- tional significance of STT projections, such as the association of pain with projections to the intralami- nar nuclei, illustrate the inherent limitations. The breadth of the functional associations possible on the basis of the present data emphasizes additional caution. For example, the dense STT projection to Pv and Pt could be of particular interest to some investigators in light of the strong limbic connections of these nuclei (Ottersen and BenAri 1979; Saper et al. 1979; Luiten et al. 1982), but little else is available to indicate the functional role of this projection. Thus, attention is drawn in the following paragraphs only to those portions of the motor and somatosen- sory systems for which these results offer pertinent new findings.
Probable motor-related projections
Some of the regions identified, such as VL, the intralaminar nuclei and the pretectal nuclei, are probably involved with motor-related aspects of
Fig. I1A-C. Labeling in the paralaminar port ion of VPL. A A t the ventromedial corner of VPL1; transverse section, case C108, near the level of section 2.70 ram, Fig. 4. B A t the ventromedial corner of VPLm; transverse section, case C l16R, near level 2.35 m m , Fig. 4. C Ventral to VPLm; horizontal section, case C l l 6 L , near level 0.20 mm, Fig. 6. Scale = A 1.0 mm, dark-field inset 180 ~tm; B 0.70 m m , dark- field 180 ~m; C 1.60 m m , dark-field 250 ~tm
248
\
.~ / / - L / / /
vMb ~ / . / F o , ' , - .
(~ t vf, i _J~,.v . . . . . . . . . . - '" / . "-~:.-.'~,:. ~ .. , , . / �9
" " ~ .,a,r : / ~ ' " ~ _ _ t I
f - - : / ,,:, ~ ' ~ ~'~ Z l .~:'~":" " !
S l / / ~ ' ~ ~ ' I O.OOmm
/ " 4 , . , . / / / / / .
'-.i" .... " ~
- ' - " ~ : ' ? = " J ' / ' - ' " - - - " I f . / / 0 .25mm , Q . . , ,,
/ / /
/ / . / y
�9 . . " . j
�9 "~, / f " 2 ~" O..m.
-% ) ; ' / /
/ i - / / "
.',,:..~..," . '
........... . ~ .. . . ~ .. ~"
~ '~:,,,.:.L;, .. /
" i
~ i O.55mm
..~3 ..." ~ ' : . . " , I" / / {i,. I ,'~' ir ' ::.' I I
: / : " ~ " 0.70 mm
/ /
/
C 1 1 7
5mm
~ P O ~ . \ /
f
/ 0 .85ram \
.$:~@ " / / ....~.(" I
�9 ":i' ' ~' I
. t ' ~ 1-05ram 1 "
( / ,
,r 1.20 mm
/ / "DI/
( I ., - .,;-" I / . .,:~' i
I I , , / ~ " I
f ~ / 7 ~ 1.35 mm
/
/ ' / I / I V/PLi �9 j ' /
.. . i : " i
�9 / 1.55mm
I,
Fig, 12. The distribution of labeling in single transverse sections in the ventral aspect of VP in case Cl17, which received an injection in the lumbar spinal cord. Note the separation of the three fields of labeling, e.g. at level 0.85 rnm. Numbers indicate distance rostral to most caudal level shown

249
spinal cord function. This does not, however, neces- sarily imply that they receive spinal input concerned solely with proprioceptive sensory inflow or kinaesthetic spinal activity (Meyers and Snow 1982).
The present data indicate that STT input to VL extends throughout most of this nucleus, rather than being confined to a small area as previously reported (Boivie 1971; Jones and Burton 1977). This projec- tion thus overlaps considerably the region of cerebel- lar terminations in lateral thalamus (Hendry et al. 1979; Sugimoto et al. 1981). The grid-like array of labeling in VL suggests that cerebellar and spinal inputs may be intricately interlaced; this possibility is consistent with the somewhat patchy nature of cere- bellar terminations in VL (Sugimoto et al. 1981; Asanuma et al. 1983). ST1? input to VL does appear to be most dense caudoventrally; such input may in part be responsible for somatic sensitivity in VL (see Nyquist 1975) and the caudal aspect of motor cortex (Pappas and Strick 1981).
The present results demonstrate that STT input to VL is topographically organized. The pattern observed agrees with the topography inferred from previous observations of the thalamo-cortical con- nectivity of VL (Strick 1973; Sakai 1982). The lack of spinal input to the most dorsomedial aspect of VL is consistent with the projection of this region to the facial part of motor cortex in the cat (Strick 1973), and/or to the rostral portion of the forelimb part of motor cortex in the raccoon (Sakai 1982).
The caudal extent of dense VL labeling was fairly sharply delimited at the rostral pole of VP in this material, suggesting that overlap with input from the DCN within a border zone (Berkeley 1980) may be minimal (see also Asanuma et al. 1983). There may, however, be some overlap with inputs from the vestibular nuclei (Raymond et al. 1974; Kotch- abhakdi et al. 1980a) and nucleus z (Grant et al. 1973) in this region.
A pattern of non-overlapping convergence with cerebellar input has previously been inferred to occur in CL (Itoh and Mizuno 1977; Hendry et al. 1979; Sugimoto et al. 1981); but, in the present material labeling occurred over a much larger extent of CL, implying direct convergence. Convergence of STr input with ascending inputs from the pretectum and superior colliculus (Graham 1977; Itoh 1977), sub- stantia nigra (Hendry et al. 1979), periphypoglossal nuclei (Kotchabhakdi et al. 1980b) and vestibular nuclei (Kotchabhakdi et al. 1980a) also appear likely within both CL and Pc. However, the present observations support the view that CL is an aggregate of several differentiable cell clusters (e.g. Rioch 1929; Jones and Burton 1974); both the pattern of input convergence and output divergence may be
organized according to these clusters (see also Sato et al. 1979). Thus, the use of anterograde double-
�9 labeling techniques will be necessary to study these possibilities. The cytoarchitecturally ambiguous ("murky"; Graybiel and Berson 1980) region dorsal to CL in the cat, which in the raccoon could be delimited as CD, may be an additional component of the intralaminar nuclei; this region is also reported to receive cerebellar input (Sugimoto et al. 1981) and to project to the caudate n. (Sato et al. 1979). Boivie (1979) has recently documented a similar parcellated STI" distribution within CL in the monkey in degen- eration material. Comparison with primate data also suggests that the region of labeling observed in caudo-medial (MD-like) CL may correspond to the STT input to the portion of CL associated with paralamellar MD in the monkey (Mehler 1969; Boivie 1979), which is connected with motor and premotor cortical fields (Akert et al. 1979).
Physiological data are consistent with these anatomical connections in indicating that CL neurons are involved with visuomotor behavior (Schlag-Rey and Schlag 1977). Furthermore, cortical areas 5 and 7, which receive input from both CL and LP-Pul (Jones and Leavitt 1974; Graybiel and Berson 1980) are functionally associated in part with visuo-somatic integration (Hyvfirinen 1982). Yet, other physiologi- cal evidence indicates that some neurons in the intralaminar nuclei respond to noxious stimulation (Casey et al. 1974; Dong et al. 1978; Nakahama et al. 1981). Some spinothalamic cells that project to the region of CL can be activated by noxious stimulation in the monkey (Applebaum et al. 1979; Giesler et al. 1981); but, there is also a substantially greater STT input to CL from lamina I cells in the primate (Willis et al. 1979) than in the cat (Carstens and Trevino 1978; Craig and Burton 1981). Meyers and Snow (1982) have recently described a population of STT cells in the intermediate zone of the cat that project to the intralaminar nuclei; about half responded to deep natural stimulation, but a few responded to noxious cutaneous stimulation. A parcellated organi- zation within CL would imply that such inputs may be functionally segregated. Nonetheless, the role of nociceptive input in visual orientation remains to be determined (Nakahama et al. 1981).
Probable sensory-related projections
A role in the sensory aspects of spinal cord function seems likely for several of the loci labeled in these experiments, such as Sm, PO, LP and VP. The dense, topographically organized STT projection to the dorsal portion of Sm (Sma) received particular

250
attention in these studies. Analogous projections were observed in the rat and the monkey (Craig and Burton 1981); the STT projection to the subparacen- traln, in the prosimian Galago (Pearson and Haines 1980) may also be analogous. As detailed in the previous report (Craig and Burton 1981), Smd appears to receive STT input exclusively from lamina I cells of the spinal and medullary dorsal horn. Whether the projection to Stud is similarly function- ally specific remains to be determined. It was sug- gested previously that Smd may be involved in the affective aspects of nociception, on the basis of the available physiological descriptions of lamina I cells, as well as consideration of the relevant clinical observations and recordings from a limited number of Sma neurons. Recent evidence indicates that most lamina I cells that project in the STIr to medial thalamus in the cat are indeed nociceptive-specific neurons, but some respond specifically to innocuous cooling, while yet others respond to both modalities (Craig and Kniffki 1982). Thus, the analysis of the specificity and the functional significance of this STT projection requires further examination.
The present data indicate that, while spinal input surrounds VPM and VPL (cf. Rockel et al. 1972; Jones and Burton 1974; Berkley 1980), several regions can be separated on the basis of the distribu- tion and topographical organization of the labeling. These may constitute multiple somatic representa- tions in the lateral thalamus in addition to those already inferred to exist within VP (Andersson et al. 1966; Landgren et al. 1965; Dykes 1982; Kaas 1983).
The labeling within PO in these experiments conformed only partially with the medial, intermedi- ate and lateral partitions used previously (Jones and Powell 1971), in that much of the labeling occurred in the medial portion. These data, however, suggest subdivisions within PO that occur in a rostro-caudal sequence. The rostral portion of PO that lies dorsal to VP is also separable on the basis of its projection to cortical area 5a in the cat (Tanji et al. 1978; Hendry et al. 1979). The topographical gradient of the somatosensory representation (SIII) in this corti- cal area may be parallel to that of the post-cruciate (SI) somatosensory representation (Darian-Smith et al. 1966), which suggests that within dorsal PO a topography parallel to that in VPL may be observ- able. This is consistent with the present observations. Furthermore, comparable physiological results have recently been obtained (Craig et al. 1984). In con- trast, the caudal portion of PO projects t o the anterior ectosylvian gyrus (Heath and Jones 1971), and may be the source of input to one of the recently defined somatic representations adjacent to the sec- ond somatosensory area (Clemo and Stein 1982;
Burton et al. 1982). The observation that these cortical fields are crudely topographic is consistent with the relatively weak topographical STT organiza- tion observed in caudal PO. Most physiological studies of PO have concentrated on this caudal portion, but these have not yet resulted in clear indications of its role in somatosensory processing (e.g. Curry 1972; Berkley 1973; Nyquist and Greenhoot 1974; Guilbaud et al. 1977).
There is little previous evidence of STT input to LP (Getz 1952; Berkley 1980). Most of the presently observed labeling was considered an extension of that within the dorsal part of PO, which has an ambiguous border with LP; yet, it clearly overlapped with the region of visual-related inputs to LP (e.g. Graybiel and Berson 1980; Updyke 1983). This projection may provide a basis for observations of visuo-somatic convergence in LP (Avancini et al. 1977). Two labeled fields in LP may be separable entities: the distinct band of cells along the LP/Pul border, and the region denoted as LPvi in the cat. The latter area may be largely equivalent to the region defined as the lateral intermediate n., pars oralis (LIo), by Graybiel and Berson (1980), which reportedly is associated with cortical area 5 but does not receive a visual-related input. Both of these particular areas have been included recently in a heterogeneous 'shell' region of the LP complex (Updyke 1983).
The present data distinguish two regions within the ventral aspect of VPL, i.e. VPI, and the ventral, paralaminar margin of VPL. The nature of the possible somatic representation in VPI is not clear; input from other pathways has not been established, and little physiological evidence is available. Moreover, VPI may not be homologous across different species. In the present data, input to VPI in the cat appeared to be rather simply organized, while in the raccoon it was complex. In the cat and the dog, VPI may project to the neighborhood of cortical area 4 (Larsen and Asanuma 1979; Tanaka et al. 1983). In the raccoon, it has been reported that VPI is the thalamic source of input to SII (Herron 1983). Furthermore, VPI in the monkey appears to receive little or no STT input (Boivie 1979; Berkley 1980; Burton and Craig 1983), to project to the insula (Burton and Jones 1976), and to contain neurons predominantly responsive to Pacinian receptors (see Dykes 1982).
In contrast, the labeling observed in the ventral margin of VPL (or, paralaminar VPL) appeared in both cat and raccoon in an equivalent pattern. The distinct topographic pattern of the labeling in para- laminar VPL suggests that this region contains a separate somatosensory representation. Recent phy-

251
siological evidence in the cat indicates that nocicep- tive neurons are localized within this portion of VPL (Craig and Burton 1979; Kniffki and Mizumura 1983; Honda et al. 1983), while only low-threshold neurons are generally found in the remainder of VPL (Golov- chinsky et al. 1981). Furthermore, neurons within paralaminar VPL have receptive fields that are topographically discontinuous from the somatotopic representation in VPL proper (Craig and Burton 1979). Spinal cells in both lamina I and V project to this area of thalamus in the cat (Carstens and Trevino 1978). The lamina I cells that project to this area of thalamus are almost all nociceptive neurons (Craig and Kniffki 1982). In the monkey, nociceptive neurons are also present (though not exclusively) in the ventral aspect of caudal VPL (Kenshalo et al. 1980), where a dense STY input is found (Boivie 1979; Berkley 1980; Burton and Craig 1983). A similar small-celled nociceptive representation may exist in man (Hassler 1960; Halliday and Logue 1972).
Finally, stark differences between the patterns of STY input to VPL in the primate and the carnivore have been emphasized previously (Boivie 1971; Jones and Burton 1974; Berkley 1980), but the present data indicate certain similarities. In the cat, patches of labeling in VPL had a consistent relation- ship with certain cell clusters. In the monkey, Boivie (1979) has noted that the distribution of STT termi- nal clusters displays a distinct and constant pattern. He also noted that scattered fibers and terminals are more frequently observed.near the borders between VPM, VPLm and VPL1, similar to the present observations in the carnivore. Furthermore, dense STY input is found in the ventral aspect of VPL in the monkey, which as noted above may be analogous to paralaminar VPL in the carnivore. Thus, STT input to VPL in the carnivore and primate may not be as different as has recently been thought; however, the data are insufficient to conclude that they have a homologous pattern. Both Boivie (1979) and Berkley (1980) have observed that there seems to be a differential relationship between the spino-cervico- thalamic pathway and the STT in primate and carnivore. It should also be noted that a considerable number of spinal lamina IV cells project directly to thalamus in the primate, while these are rare in the cat (Willis et al. 1979; Carstens and Trevino 1978). This distinction alone may account for the major remaining difference between the STY input to VPL in the monkey and the cat.
Conclusions
It is reasonable to conclude from these data that the STT distributes to a vastly larger number of loci than
has been previously appreciated. Yet, the differentia- tion of several neuronal subregions and the topo- graphic organization of the input in many areas suggest that it is likely that considerable anatomical and functional specificity is inherent within this broad distribution. Only general functional associations can be made at the present time for most loci; additional data regarding the functional characteristics of the STT cells projecting to each target are necessary for further analysis. It seems prospective to suggest that each of the separate loci identified by these projec- tions may subserve a unique role with respect to the variety of separable and integrated aspects of spinal cord function. The present observations support the assumption that spinal connections with more rostral portions of the neuraxis are at least equally well- organized as the spinal cord itself.
Acknowledgements. The authors wish to thank R. Freund and E. Tallone for photographic help; D. Craig, C. Dawson and M. Derrick for typing the manuscript; Drs. E.G. Jones, J.L. Price and K. J. Berkley for offering criticism of an early draft, and Dr. J. Boivie for reviewing the final draft. For many valuable discussions and support during the preparation of this manuscript, sincere appreciation is expressed to Dr. K.-D. Kniffki. This work was supported by NIH grants NS07057 and NS09809.
Abbreviations
A anterior pretectal nucleus; AD anterodorsal n.; AM anterome- dial n.; AV anteroventral n.; CeM centromedial n.; CD centrodor- sal n. (raccoon); CL centrolateral n.; CM centre median; H habenula; L n. limitans; LD laterodorsal n.; LG lateral geniculate; LGv lateral geniculate, ventral subnucleus; LP lateral posterior n.; LPvi lateral posterior n., ventral intermediate part; M medial pretectal n.; me medial geniculate, magnocellular subnucleus; MD mediodorsal n.; MG medial geniculate; ML medial lemniscus; N pretectal nucleus of the optic tract; nBIC n. of the brachium of the inferior colliculus; O olivary pretectal n.; OT optic tract; P posterior nucleus of Rioch; Pc paracentral n. ; Pf parafascicular n.; PO posterior group of thalamus; PP posterior pretectal n.; Pt parataenial n.; Pul ptflvinar; Pv paraventricular n. of thalamus; R reticular n.; Re n. reuniens; Rh rhomboid n.; RN red nucleus; SG suprageniculate n.; S m n . submedius; SN substantia nigra; Spf subparafascicular n.; Tg mesencephalic tegmentum; VA ventroan- terior n.; VP ventroposterior thalamus (i.e. VPM, VPI, and VPL); VL ventrolateral n.; VM ventromedial n.; VMb ventromedial n., basal part; VPI ventroposteroinferior n.; VPL1 ventroposterola- teral n., lateral part; VPLm ventroposterolateral n., medial part; VPM ventroposteromedial n.; ZI zona incerta.
References
Akert K, Hartmann-von Monakow K, Kfinzle H (1979) Projection of precentral motor cortex upon nucleus medialis thalami in the monkey. Neurosci Lett 11:103-106
Anderson FD, Berry CM (1959) Degeneration studies of long ascending fiber systems in the cat brain stem. J Comp Neurol 111:195-229

252
Andersson SA, Landgren S, Wolsk D (1966) The thalamic relay and cortical projection of group I muscle afferents from the forelimb of the cat. J Physiol (Lond) 183:576-591
Applebaum AE, Leonard RB, Kenshalo DR Jr, Martin RF, Willis WD (1979) Nuclei in which functionally identified spinothalamic tract neurons terminate. J Comp Neurol 188: 575-586
Asanuma C, Thach WT, Jones EG (1983) Distribution of cerebel- lar terminations and their relation to other afferent termina- tions in the ventral lateral thalamic region of the monkey. Brain Res Rev 5:237-265
Avanzini G, Spreafico R, Broggi G, Giovannini P, Franceschetti S (1977) Topographic distribution of visual and somesthetic unitary responses in the PUL-LP complex of the cat. Neurosci Lett 4:135-143
Berkley KJ (1973) Response properties of cells in the ventrobasal and posterior group nuclei of the cat. J Neurophysiol 36: 940-952
Berldey KJ (1980) Spatial relationships between the terminations of somatic sensory and motor pathways in the rostral brain- stem of cats and monkeys. I. Ascending somatic sensory inputs to lateral diencephalon. J Comp Neurol 193:283-317
Bharos TB, Kuypers HGJM, Lemon RN, Muir RB (1981) Divergent collaterals from deep cerebellar neurons to thalamus and tectum, and to medualla oblongata and spinal cord: retrograde fluorescent and electrophysiological studies. Exp Brain Res 42:399-410
Boivie J (1971) The termination of the spinothalamic tract in cat: an experimental study with silver impregnation methods. Exp Brain Res 12:331-353
Boivie J (1979) An anatomical reinvestigation of the termination of the spinothalamic tract in the monkey. J Comp Neurol 186: 343-370
Boivie JJG, Perl ER (1975) Neural substrates of somatic sensa- tion. In: Guyton AC, Hunt CC (eds) Physiology Series One, Vol III, Neurophysiology. University Park Press, Baltimore
Brown AG (1981) Organization in the spinal cord. Springer, Berlin Heidelberg New York, pp 238
Burton H, Craig AD (1983) Spinothalamic projections in cat, raccoon and monkey: a study based on anterograde transport of horseradish peroxidase. In: Macchi G, Rustioni A, Spreafico R (eds) Somatosensory integration in the thalamus. Elsevier, Amsterdam, pp 17-41
Burton H, Jones EG (1976) The posterior thalamic region and its cortical projection in new world and old world monkeys. J Comp Neurol 168:249-302
Burton H, Loewy AD (1977) Projections to the spinal cord from medttllary somatosensory relay nuclei. J Comp Neurol 173: 773-792
Burton H, Mitchell G, Brent D (1982) Second somatic sensory area in the cerebral cortex of cats: somatotopic organization and cytoarchitecture. J Comp Neurol 210:109-135
Carstens E, Trevino DL (1978) Laminar origins of spinothalamic projections in the cat as determined by the retrograde transport of horseradish peroxidase. J Comp Neurol 182: 151-166
Casey KL, Keene JJ, Morrow T (1974) Btdboreticular and medial thalamie unit activity in relation to aversive behavior and pain. In: Bonica JJ (ed) Advances in neurology. Raven, New York, pp 197-205
Clemo HR, Stein BE (1982) Somatosensory cortex: a "new" representation. Brain Res 235:162-168
Cond6 F, Cond6 H (1980) Demonstration of a rubrothalamic projection in the cat, with some comments on the origin of the rubrospinal tract. Neuroscience 5:789-802
Cowan WM, Gottlieb DI, Hendrickson AE, Price JL, Woolsey TA (1972) The autoradiographic demonstration of axonal
connections in the central nervous system. Brain Res 37: 21-51
Craig AD, Burton H (1979) Spinothalamic terminations in the ventroposterolateral nucleus of the cat. Soc Neurosci Abst 5: 705
Craig AD, Burton H (1981) Spinal and medullary lamina I projection to nucleus submedius in medial thalamus: a possible pain center. J Neurophysiol 45:443-466
Craig AD, Dunst R, Kniffki K-D (1984) Topographic distribution of somatosensory neurones dorsal to the cat's ventroposterior nuclei. Pflfigers Arch 400:R16
Craig AD, Kniffki K-D (1982) Lumbosacral lamina I cells projecting to medial and/or lateral thalamus in the cat. Soc Neurosci Abst 8:95
Craig AD, Wiegand SJ, Price JL (1982) The thalamo-cortical projection of the nucleus submedius in the cat. J Comp Neurol 206:28-48
Curry MJ (1972) The exteroceptive properties of neurones in the somatic part of the posterior group (PO). Brain Res 44: 439-462
Darian-Smith I, Isbister J, Mok H, Yokota T (1966) Somatic sensory cortical projection areas excited by tactile stimulation of the cat: a triple representation. J Physiol (Lond) 182: 671-689
De Olmos J, Heimer L (1977) Mapping of collateral projections with the HRP-method. Neurosci Lett 6:107-114
Dong WK, Ryu H, Wagman IH (1978) Nociceptive responses of neurons in medial thalamus and their relationship to spinothalamic pathways. J Neurophysiol 41:1592-1613
Dykes RW (1982) Parallel processing of cutaneous information in the somatosensory system of the cat. J Can Sci Neurol 9:9-19
Getz B (1952) The termination of spinothalamic fibers in the cat as studied by the method of terminal degeneration. Acta Anat 16:271-290
Giesler GJ Jr, Menetrey D, Basbaum AI (1979) Differential origins of spinothalamic tract projections to medial and lateral thalamus in the rat. J Comp Neurol 184:107-126
Giesler GJ Jr, Yezierski RP, Gerhart KD, Willis WD (1981) Spinothalamic tract neurons that project to medial and/or lateral thalamic nuclei: evidence for a physiologically novel population of spinal cord neurons. J Neurophysiol 46: 1285-1308
Golovchinsky V, Kruger L, Saporta SA, Stein BE, Young DW (1981) Properties of velocity-mechanosensitive neurons of the cat ventrobasal thalamic nucleus with special reference to the concept of convergence. Brain Res 209:355-374
Graham J (1977) An autoradiographic study of the efferent connections of the superior colliculus in the cat. J Comp Neurol 17:629-654
Grant G, Boivie J, Silfvenius H (1973) Course and termination of fibers from the nucleus z of the medulla oblongata. An experimental light microscopical study in the cat. Brain Res 55:55-70
Graybiel AM, Berson DM (1980) Histochemical identification and afferent connections of subdivisions in the lateralis posterior- pulvinar complex and related thalamic nuclei in the cat. Neuroscience 5:1175-1238
Guilbaud G, Caille D, Besson JM, Benelli G (1977) Single unit activities in ventral posterior and posterior group thalamic nuclei during nociceptive and non-nociceptive stimulation in the cat. Arch Ital Biol 115:38-56
Halliday AM, Logue V (1972) Painful sensations evoked by electrical stimulation in the thalamus. In: Somjen GG (ed) Neurophysiology studied in man. Excerpta Medica, Amster- dam, pp 221-230
Hassler R (1960) Die zentralen Systeme des Schmerzes. Acta Neurochir 8:353-423

253
Heath CJ, Jones EG (1971) An experimental study of ascending connections from the posterior group of thalamic nuclei in the cat. J Comp Neurol 141:397-426
Hendry SHC, Jones EG, Graham J (1979) Thalamic relay nuclei for cerebellar and certain related fiber systems in the cat. J Comp Neurol 185:679-714
Herron P (1983) The connections of cortical somatosensory areas I and II with separate nuclei in the ventroposterior thalamus in the raccoon. Neuroscience 8:243-257
Honda CN, Mense S, Perl ER (1983) Neurons in ventrobasal region of cat thalamus selectively responsive to noxious mechanical stimulation. J Neurophysiol 49:662-673
Hongo T, Kudo N, Yamashita M, Ishizuka N, Mannen H (1981) Transneuronal passage of intraaxonally injected horseradish peroxidase (HRP) from group Ib and II fibers into the secondary neurons in the dorsal horn of the cat spinal cord. Biomed Res 2:722-727
Hyvfirinen J (1982) Posterior parietal lobe of the primate brain. Physiol Rev 62:1060-1129
Itoh K (1977) Efferent projections of the pretectum in the cat. Exp Brain Res 30:89-105
Itoh K, Mizuno N (1977) Topographical arrangement of thalamocortical neurons in the centrolateral nucleus (CL) of the cat with special reference to the spino-thalamo-motor cortical path through the CL. Exp Brain Res 30:471-480
Jones EG (1981) Functional subdivision and synaptic organization of the mammalian thalamus. In: Porter R (ed) Neurophysiol- ogy IV, International review of physiology. University Park Press, Baltimore (vol 25, pp 173-245)
Jones EG, Burton H (1974) Cytoarchitecture and somatic sensory connectivity of thalamic nuclei other than the ventrobasal complex in the cat. J Comp Neurol 154:395-432
Jones EG, Leavitt RY (1974) Retrograde axonal transport and the demonstration of non-specific projections to the cerebral cortex and striatum from thalamic intralaminar nuclei in the rat, cat, and monkey. J Comp Neurol 154:349-378
Jones EG, Powell TPS (1971) An analysis of the posterior group of thalamie nuclei on the basis of its afferent connections. J Comp Neurol 143:185-216
Kaas JH (1983) What, if anything, is SI? Organization of first somatosensory area of cortex. Physiolog Rev 63:206-231
Kenshalo DR Jr, Giesler GJ Jr, Leonard RB, Willis WD (1980) Response of neurons in primate ventral posterior lateral nucleus to noxious stimuli. J Neurophysiol 43:1594-1614
Kniffki K-D, Mizumura K (1983) Responses of neurons in VPL and VPL-VL region of the cat to algesic stimulation of muscle and tendon. J Neurophysiot 49:649-661
Kotchabhakdi N, Rinvik E, Walberg F, Yingchareon K (1980a) The vestibulothalamic projection in the cat studied by retro- grade axonal transport of horseradish peroxidase. Exp Brain Res 40:405-418
Kotchabhakdi N, Rinvik E, Yingchareon K, Walberg F (1980b) Afferent projections to the thalamus from the perihypoglossal nuclei. Brain Res 187:457-461
Kuypers HGJM, Maisky VA (1975) Retrograde axonal transport of HRP from spinal cord to brain stem cell groups in the cat. Neurosci Lett 1:9.14
Landgren S, Nordwall A, Wangstr6m C (1965) The location of the thalamic relay in the spino-cervico-lemniscal path. Acta Physiol Scand 65:164-175
Larsen KD, Asanuma H (1979) Thalamic projections to the feline motor cortex studied with horseradish peroxidase. Brain Res 172:209.215
Luiten PGM, Kuipers F, Schuitmaker H (1982) Organization of diencephalic and brainstem afferent projections to the lateral septum in the rat. Neurosci Lett 30:2tl-216
Mantyh PW, Peschanski M (1983) The use of wheat germ
agglutinin-horseradish peroxidase conjugates for studies of anterograde axonal transport. J Neurosci Meth 7:117-128
Mehler WR (1969) Some neurological species differences - a posterior. Ann NY Acad Sci 167:424-468
Mesulam M-M (ed) (1982) Tracing neural connections with horseradish peroxidase. Wiley, New York, pp 251
Meyers DER, Snow PJ (1982) The responses to somatic stimuli of deep spinothalamic tract cells in the lumbar spinal cord of the cat. J Physiol (Lond) 329:355-371
Nakahama H, Shima K, Aya K, Kisara K, Sakurada S (1981) Antinociceptive action of morphine and pentazocine on unit activity in the nucleus centralis lateralis, nucleus ventralis lateralis and nearby structures of the cat. Pain 10:47-56
Nanta WJtt, Kuypers HGJM (1958) Some ascending pathways in the brain stem reticular formation. In: Jasper HH et al. (eds) Reticular formation of the brain. Little, Brown & Co., Boston, pp 3-30
Nyquist JK (1975) Somatosensory properties of neurons of thalamic nucleus ventralis lateraiis. Exp Neurol 48:123-135
Nyquist JK, Greenhoot JH (1974) Unit analysis of nonspecific thalamic responses to high-intensity cutaneous input in the cat. Exp Neurol 42:609-622
Ottersen OP, Ben-Aft Y (1979) Afferent connections to the amygdaloid complex of the rat and cat. I. Projections from the thalamus. J Comp Neurol 187:401-424
Pappas CL, Strick PL (1981) Physiological demonstration of multiple representation in the forelimb region of the cat motor cortex. J Comp Neurol 200:481-490
Pearson JC, Haines DE (1980) Somatosensory thalamus of a prosimian primate (Galago senegalensis). I. Configuration of nuclei and termination of spinothalamic fibers. J Comp Neurol 190:533-558
Peschanski M, Mantyh PW (1983) Efferent connections of the subfascicular area of the mesodiencephalic junction and its possible involvement in stimulation-produced analgesia. Brain Res 263:181-190
Raymond J, Sans A, Marty R (1974) Projections thaiamiques des noyaux vestibulaires: etude histologique chez le chat. Exp Brain Res 20:273-283
Rioch DM (1929) Studies on the diencephalon of Carnivora. Part I. The nuclear configuration of the thalamus, epithalamus, and hypothalamus of the dog and cat. J Comp Neurol 49: 1-119
Rockel AJ, Heath CJ, Jones EG (1972) Afferent connections to the diencephaion in the marsupial phalanger and the question of sensory convergence in the "posterior group" of the thalamus. J Comp Neurol 145:105-130
Sakai ST (1982) The thalamic connectivity of the primary motor cortex (MI) in the raccoon. J Comp Neurol 204:238-252
Saper CB, Swanson LW, Cowan WM (1979) Some efferent connections of the rostral hypothalamus in the squirrel monkey (Saimiri scittreus) and cat. J Comp Neurol 184: 205-242
Sato M, Itoh K, Mizuno N (1979) Distribution of thalamocandate neurons in the cat as demonstrated by horseradish peroxidase. Exp Brain Res 34:143-153
Schlag-Rey M, Schlag J (1977) Visual and presaccadic neuronal activity in thalamic internal medullary lamina of cat: a study of targeting. J Neurophysiol 40:156-173
Sherman SM, Spear PD (1982) Organization of visual pathways in normal and visually deprived cats. Physiolog Rev 62:738-855
Strick PL (1973) Light microscopic analysis of the cortical projec- tion of the thalamic ventrolateral nucleus in the cat. Brain Res 55:1-24
Sugimoto T, Mizuno N, Itoh K (1981) An autoradiographic study on the terminal distribution of cerebellothalamic fibers in the cat. Brain Res 215:29-47

254
Tanaka D Jr, Sakai ST, Gorska T (1983) Corticothalamic projec- tions from postcruciate area 4 in the dog. J Comp Neuro1214: 17-31
Tanji DG, Wise SP, Dykes RW, Jones EG (1978) Cytoarchitec- ture and thalamic connectivity of third somatosensory area of cat cerebral cortex. J Neurophysiol 41:268284
Updyke BV (1983) A reevaluation of the functional organization and cytoarchitecture of the feline lateral posterior complex, with observations on adjoining cell groups. J Comp Neurol 219:143-181
Welker WI, Johnson JI Jr (1965) Correlation between nuclear morphology and somatotopic organization in ventro-basal complex of the raccoon's thalamus. J Anat 99:761-790
Wiberg M, Blomqvist A (1983) The spinomesencephalic tract in the cat: its cells of origin and termination pattern as demon- strated by the intraaxonal transport method. Brain Res 291: 1-18
Willis WD (1983) The spinothalamic tract. In: Rosenberg RN (ed) The clinical neurosciences. Section V, Willis WD (ed) Neurobiology. Churchill-Livingstone, Edinburgh, pp 325-356
Willis WD, Kenshalo DR Jr, Leonard RB (1979) The cells of origin of the primate spinothalamic tract. J Comp Neuro1188: 543-574
Received January 2, 1984 / Accepted December 14, 1984




















