
Anti-La/SSB antibodies transported across the placenta bind apoptotic cells in fetal organs targeted...
8
ARTHRITIS & RHEUMATISM Vol. 46, No. 6, June 2002, pp 1572–1579 DOI 10.1002/art.10316 © 2002, American College of Rheumatology Anti-La/SSB Antibodies Transported Across the Placenta Bind Apoptotic Cells in Fetal Organs Targeted in Neonatal Lupus Hai B. Tran, 1 Peter J. Macardle, 1 Jenny Hiscock, 1 Dana Cavill, 1 John Bradley, 1 Jill P. Buyon, 2 and Tom P. Gordon 1 Objective. To determine whether La and/or Ro epitopes on apoptotic cells in fetal organs that are targeted in neonatal lupus syndrome (NLS) are acces- sible for binding by autoantibodies in vivo, we traced the fate of transplacental autoantibodies in a murine pas- sive transfer model. Methods. Pregnant mice at day 15 of gestation (E15) were injected intraperitoneally with human anti- Ro/La–positive sera or control sera, and transplacental transfer of human autoantibodies was tested by enzyme- linked immunosorbent assay with recombinant anti- gens. Multiple cryostat sections at the level of the heart of E17 fetuses were visualized simultaneously for hu- man IgG binding and apoptosis (TUNEL) under confo- cal microscopy. Serial paraffin sections of E17 and E19 fetuses were examined for histologic evidence of inflam- mation. Results. Human IgG anti–52-kd Ro, anti–60-kd Ro, and anti-La autoantibodies were transported effi- ciently into the fetal circulation. Human IgG–apoptotic cell complexes were detected in the heart (atrial trabec- ulae and atrioventricular node), skin, liver, and newly forming bone of fetuses from mothers injected with anti-Ro/La sera but not control sera. The IgG binding was fetal-specific and organ-specific; transplacental au- toantibodies did not bind to apoptotic cells in the fetal thymus, lung, brain, or gut. The complexes were not associated with an inflammatory reaction. Injection of mothers with affinity-purified anti-La autoantibodies (but not anti-Ro/La Ig depleted of anti-La) revealed an identical location of IgG binding to apoptotic cells in the fetuses. Conclusion. This is the first study to demonstrate that transplacental anti-La autoantibodies bind specif- ically to apoptotic cells in selected fetal organs in vivo, similar to the organ involvement in NLS. We hypothe- size that additional factors are required to promote proinflammatory clearance of IgG–apoptotic cell com- plexes and subsequent tissue damage. Neonatal lupus syndrome (NLS) is a model of passively acquired autoimmunity in which damage to the neonatal heart, skin, and liver is presumed to arise from the transplacental passage of autoantibodies directed against Ro/SSA and La/SSB ribonucleoprotein antigens (1–5). Congenital heart block (CHB) is the most severe manifestation of NLS and is characterized in postmor- tem studies by inflammation and connective tissue re- placement of the fetal conduction system (6,7). The role of anti-Ro/La autoantibodies in the selective fetal tissue injury is poorly understood (5,8–10). Animal models have recently provided strong evidence that the neonatal cardiac conduction defects are mediated specifically by anti-Ro/La autoantibodies, while the maternal heart is spared. Mice actively immu- nized with components of the Ro/La complex delivered pups with various conduction disturbances (11), and electrocardiographic abnormalities were also detected in pups from mice injected with human anti-Ro/La auto- antibodies from mothers of children with CHB (12). However, these studies did not examine the heart or other organs for deposition of maternal autoantibodies and histologic features of tissue damage. Recent investigations have focused on apoptotic Supported by grants from the Australian National Health and Medical Research Council, the National Institute of Arthritis and Musculoskeletal and Skin Diseases (AR-42455), and the Australian International Postgraduate Research Scholarship. 1 Hai B. Tran, MSc, Peter J. Macardle, PhD, Jenny Hiscock, PhD, Dana Cavill, BSc (Hons), John Bradley, MD, PhD, Tom P. Gordon, MD, PhD: Flinders Medical Centre, Flinders University, Adelaide, Australia; 2 Jill P. Buyon, MD: Hospital for Joint Diseases, New York University School of Medicine, New York, New York. Address correspondence and reprint requests to Tom P. Gordon, MD, PhD, Department of Immunology, Allergy and Arthritis, Flinders Medical Centre, Bedford Park, 5042, SA, Australia. E-mail: [email protected]. Submitted for publication October 16, 2001; accepted in revised form February 7, 2002. 1572
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Anti-La/SSB antibodies transported across the placenta bind apoptotic cells in fetal organs targeted...

ARTHRITIS & RHEUMATISMVol. 46, No. 6, June 2002, pp 1572–1579DOI 10.1002/art.10316© 2002, American College of Rheumatology
Anti-La/SSB Antibodies Transported Across the Placenta BindApoptotic Cells in Fetal Organs Targeted in Neonatal Lupus
Hai B. Tran,1 Peter J. Macardle,1 Jenny Hiscock,1 Dana Cavill,1 John Bradley,1
Jill P. Buyon,2 and Tom P. Gordon1
Objective. To determine whether La and/or Roepitopes on apoptotic cells in fetal organs that aretargeted in neonatal lupus syndrome (NLS) are acces-sible for binding by autoantibodies in vivo, we traced thefate of transplacental autoantibodies in a murine pas-sive transfer model.
Methods. Pregnant mice at day 15 of gestation(E15) were injected intraperitoneally with human anti-Ro/La–positive sera or control sera, and transplacentaltransfer of human autoantibodies was tested by enzyme-linked immunosorbent assay with recombinant anti-gens. Multiple cryostat sections at the level of the heartof E17 fetuses were visualized simultaneously for hu-man IgG binding and apoptosis (TUNEL) under confo-cal microscopy. Serial paraffin sections of E17 and E19fetuses were examined for histologic evidence of inflam-mation.
Results. Human IgG anti–52-kd Ro, anti–60-kdRo, and anti-La autoantibodies were transported effi-ciently into the fetal circulation. Human IgG–apoptoticcell complexes were detected in the heart (atrial trabec-ulae and atrioventricular node), skin, liver, and newlyforming bone of fetuses from mothers injected withanti-Ro/La sera but not control sera. The IgG bindingwas fetal-specific and organ-specific; transplacental au-toantibodies did not bind to apoptotic cells in the fetal
thymus, lung, brain, or gut. The complexes were notassociated with an inflammatory reaction. Injection ofmothers with affinity-purified anti-La autoantibodies(but not anti-Ro/La Ig depleted of anti-La) revealed anidentical location of IgG binding to apoptotic cells in thefetuses.
Conclusion. This is the first study to demonstratethat transplacental anti-La autoantibodies bind specif-ically to apoptotic cells in selected fetal organs in vivo,similar to the organ involvement in NLS. We hypothe-size that additional factors are required to promoteproinflammatory clearance of IgG–apoptotic cell com-plexes and subsequent tissue damage.
Neonatal lupus syndrome (NLS) is a model ofpassively acquired autoimmunity in which damage to theneonatal heart, skin, and liver is presumed to arise fromthe transplacental passage of autoantibodies directedagainst Ro/SSA and La/SSB ribonucleoprotein antigens(1–5). Congenital heart block (CHB) is the most severemanifestation of NLS and is characterized in postmor-tem studies by inflammation and connective tissue re-placement of the fetal conduction system (6,7). The roleof anti-Ro/La autoantibodies in the selective fetal tissueinjury is poorly understood (5,8–10).
Animal models have recently provided strongevidence that the neonatal cardiac conduction defectsare mediated specifically by anti-Ro/La autoantibodies,while the maternal heart is spared. Mice actively immu-nized with components of the Ro/La complex deliveredpups with various conduction disturbances (11), andelectrocardiographic abnormalities were also detected inpups from mice injected with human anti-Ro/La auto-antibodies from mothers of children with CHB (12).However, these studies did not examine the heart orother organs for deposition of maternal autoantibodiesand histologic features of tissue damage.
Recent investigations have focused on apoptotic
Supported by grants from the Australian National Health andMedical Research Council, the National Institute of Arthritis andMusculoskeletal and Skin Diseases (AR-42455), and the AustralianInternational Postgraduate Research Scholarship.
1Hai B. Tran, MSc, Peter J. Macardle, PhD, Jenny Hiscock,PhD, Dana Cavill, BSc (Hons), John Bradley, MD, PhD, Tom P.Gordon, MD, PhD: Flinders Medical Centre, Flinders University,Adelaide, Australia; 2Jill P. Buyon, MD: Hospital for Joint Diseases,New York University School of Medicine, New York, New York.
Address correspondence and reprint requests to Tom P.Gordon, MD, PhD, Department of Immunology, Allergy and Arthritis,Flinders Medical Centre, Bedford Park, 5042, SA, Australia. E-mail:[email protected].
Submitted for publication October 16, 2001; accepted inrevised form February 7, 2002.
1572

cells in the developing fetus as potential targets forbinding of passively transferred maternal autoantibod-ies. The Ro and La antigens translocate to the surfaceblebs of cultured apoptotic keratinocytes, a commontarget in NLS (13). Anti-Ro/La autoantibodies bind invitro to the surface of nonpermeabilized apoptotic hu-man fetal cardiocytes and subsequently promote thesecretion of the proinflammatory cytokine tumor necro-sis factor � by cocultured macrophages (14,15). Al-though opsonization of fetal apoptotic cells by maternalautoantibodies remains an attractive hypothesis for theinitiation of tissue injury in NLS, the formation ofIgG–apoptotic cell complexes in selective fetal tissueshas not yet been demonstrated in vivo.
We have recently identified apoptotic cells in theconduction system of the unmanipulated mouse fetalheart and demonstrated that clustering of La antigennear the surface of apoptotic bodies occurs in vivo underphysiologic conditions (16). Accordingly, the presentstudy was initiated to assess proof of the concept and toexamine whether La and/or Ro epitopes on apoptoticcells are accessible for binding by autoantibodies in vivo.To accomplish this goal, we used a murine passivetransfer model in which the fate of injected anti-Ro/Laautoantibodies, which are actively transported across theplacenta, can be traced in fetal tissues known to havehigh rates of apoptosis. We report that anti-La autoan-tibodies bind to apoptotic cells in the developing heart,liver, bone, and skin. The formation of IgG–apoptoticcell complexes is organ-specific and fetal-specific, andexcept for bone, corresponds to the organ involvementof NLS.
MATERIALS AND METHODS
Human sera. Sera containing anti-Ro and anti-Laprecipitins on counterimmunoelectrophoresis were obtainedfrom 2 female patients with primary Sjogren’s syndrome (17)and a mother whose baby had CHB (18). All sera were positivefor anti–52-kd Ro, anti–60-kd Ro, and anti-La on immunoblot(INNO-LIA; Innogenetics, Ghent, Belgium). Additional serawere collected from individual patients with the followingdiagnoses: primary Sjogren’s syndrome with monospecificanti–60-kd Ro, systemic lupus erythematosus with high-titeranti–double-stranded DNA (anti-dsDNA), systemic sclerosiswith anticentromere autoantibodies, and drug-induced lupuserythematosus with antihistone autoantibodies. Normal hu-man sera (NHS) were collected from 3 healthy donors.
Affinity purification of human anti-La autoantibodies.Soluble recombinant fusion mouse La-GST protein preparedas described previously (19) was coupled to cyanogenbromide–activated Sepharose 4B (Pharmacia, Uppsala, Swe-den). The Ig fractions positive for anti–52-kd Ro, anti–60-kdRo, and anti-La were isolated from the sera of 2 patients with
primary Sjogren’s syndrome by caprylic acid precipitation (20).Ig dissolved in phosphate buffered saline (PBS) was loadedonto the La-GST immunoabsorbent column and the unboundflowthrough fraction passed over the column 3 times.
After washing with an excess of PBS, the bound anti-LaIg fraction was eluted with 0.1M glycine containing 0.5M NaCl,pH 2.3, and neutralized with 2M Tris, pH 7.5. Both eluted(affinity-purified) and unbound fractions were dialyzed againstPBS, and Ig levels were measured by nephelometry. Theaffinity-purified anti-La contained �90% of the anti-La activ-ity of the starting serum, as assessed by recombinant Laenzyme-linked immunosorbent assay (ELISA) (data notshown). The unbound fraction (anti-La autoantibody–depletedfraction) retained �90% anti–60-kd Ro and anti–52-kd Roactivity, but contained �5% anti-La activity, by ELISA.
Murine model of passive transfer of human autoanti-bodies. Protocols of procedures on animals were approved bythe Flinders University Animal Welfare Committee. Time-mated pregnant BALB/c mice (7–8 weeks old) were injectedintraperitoneally on day 15 of gestation (E15) with humanserum (0.5 ml), affinity-purified anti-La autoantibodies (450�g in 0.5 ml of PBS), or anti-Ro/La autoantibodies depleted ofanti-La (0.5 ml). Twenty-eight dams were injected with 1 of thefollowing: sera positive for anti–52-kd Ro, anti–60-kd Ro, andanti-La (n � 9); serum monospecific for anti–60-kd Ro (n �2); sera containing anti-dsDNA (n � 1), anticentromere (n �1) or antihistone (n � 1) antibodies; NHS (n � 9); affinity-purified human anti-La autoantibodies (n � 3); or humananti-Ro/La Ig depleted of anti-La autoantibodies (n � 2).
The E15 time point for injection was chosen to opti-mize the chance of antibody binding in utero to apoptotic cellsin the fetal heart conducting tissue. In our previous study onBALB/c mice (full-term is at E19), apoptotic cells were clearlydemonstrated at E15 and E17 in the atrioventricular (AV)node, AV bundle, and sinoatrial (SA) node (16). Furthermore,most of the transfer of human IgG across the murine placentatakes place after E13, reaching a maximum level during the lastdays of gestation (21).
To examine human IgG binding to mouse tissue, 21pregnant mice were killed at E17 by cervical dislocation. Thefetuses were removed, decapitated, and the fetal bodies wereimmediately snap-frozen in tissue-embedding medium(Sakura, Tokyo, Japan) at the freezing point of isopentane(APS Ajax Finechem, Auburn, New South Wales, Australia) inliquid nitrogen. Sections (5 �m thick) were cut on a cryostat(Leica Instruments, Nussloch, Germany) in the frontal plane.At the levels between the AV and SA nodes, �70 serialsections were collected.
Sections were mounted on glass slides that had beenprecoated with APES (Sigma, St. Louis, MO), fixed andpermeabilized with acetone:methanol (3:1) at �20°C for 3minutes, air-dried, and stored at �80°C. In additional experi-ments to exclude postfixation artifactual binding of humanIgG, sections were fixed for 10 minutes at room temperature in4% paraformaldehyde rather than acetone:methanol, and per-meabilized with 0.05% Triton X-100 in PBS for 15 minutesbefore staining for human IgG.
To examine for histologic evidence of leukocyte infil-tration and inflammation, 15 fetuses (E17) and 15 pups (on theday of delivery) from anti-Ro/La–injected dams were killed asabove, embedded in paraffin, and serial sections were cut and
ORGAN-SPECIFIC BINDING OF ANTI-LA/SSB IGG TO FETAL APOPTOTIC CELLS 1573
stained with hematoxylin and eosin. The fetal AV node, AVbundle, and SA node were identified in serial sections asdescribed previously (16).
ELISA. Sera from mothers and their fetuses (1:250dilution) were tested for anti–52-kd Ro, anti–60-kd Ro, andanti-La autoantibodies by ELISA, as described previously (22).Polysorb microtiter plates (Nunc, Roskilde, Denmark) werecoated with the recombinant human (h) or mouse (m) antigens6His-hLa, 6His-mLa, 6His-hRo52, 6His-mRo52, 6His-hRo60,MBP-mRo60 and with control recombinant proteins (MBPand 6His-HEL) in 0.03M carbonate buffer, pH 9.6, overnightat 4°C. Bound human IgG autoantibodies were detected withalkaline phosphatase–labeled anti-human IgG (Sigma).
Dual staining for TUNEL and human IgG in tissuesections. Frozen tissue sections were washed with 0.1% Tween20 in PBS (PBST), and a TUNEL assay was used forfluorescein-labeling of fragmented DNA in apoptotic cells(Promega, Madison, WI). Sections were washed twice inPBST, blocked for 30 minutes with serum-free protein block-ing solution (Dako, Glostrup, Denmark), and incubated for 60minutes at room temperature with a 1:1,000 dilution of Cy3-conjugated donkey anti-human IgG (Jackson ImmunoRe-search, West Grove, PA). After 3 washes in PBST, coverslipswere added using glycerol phosphate mountant. Multiplesections (�10 from each fetus) were screened at the levelbetween the SA and the AV node.
Confocal microscopy. Sections double stained forTUNEL and human IgG were viewed with a Leica microscopeand then with a Bio-Rad MRC 1024 confocal scanning lasersystem (Bio-Rad, Hemel Hempstead, UK) installed on anOlympus AX70 microscope (Olympus, Tokyo, Japan)equipped with a krypton argon laser. Differential visualizationof the fluorophores was achieved using a 488-nm excitationfilter and a 522-nm emission filter for fluorescein isothiocya-nate and a 568-nm excitation filter and a 585-nm emission filterfor Cy3. Sections were scanned sequentially to collect green orred fluorescence using a 20� objective for low-power imagesdemonstrating the position of TUNEL–Cy3 staining. A 100�objective lens and 3� zoom were used to localize IgG fluores-cence at the cellular level. Images of 512 � 512 pixels werefalse-colored and merged using Lasersharp software (Bio-Rad). TUNEL-labeled cells of special interest were scannedwith a 100� objective lens and 3� zoom at 0.4-�m intervals inthe vertical axis to localize IgG within the cell.
RESULTS
Transfer of human anti-Ro/La autoantibodiesacross the murine placenta. Preliminary experimentsconfirmed efficient transport of IgG from both normaland autoantibody-containing human sera across themurine placenta. Levels of human IgG in fetal mousesera were determined from a standard curve in which anormal mouse serum mixed with increasing amounts ofhuman IgG was coated onto ELISA plates, and humanIgG was detected with alkaline phosphatase–labeledanti-human IgG diluted 1:1,000. Sera from 8 fetusesfrom anti-Ro/La–injected mothers and 8 fetuses from
NHS-injected mothers gave mean � SD values of hu-man IgG of 234 � 67 mg/liter and 184 � 8 mg/liter,respectively (P � 0.33 by Mann-Whitney U test). Fetallevels of human IgG ranged from 50% to 75% ofmaternal levels.
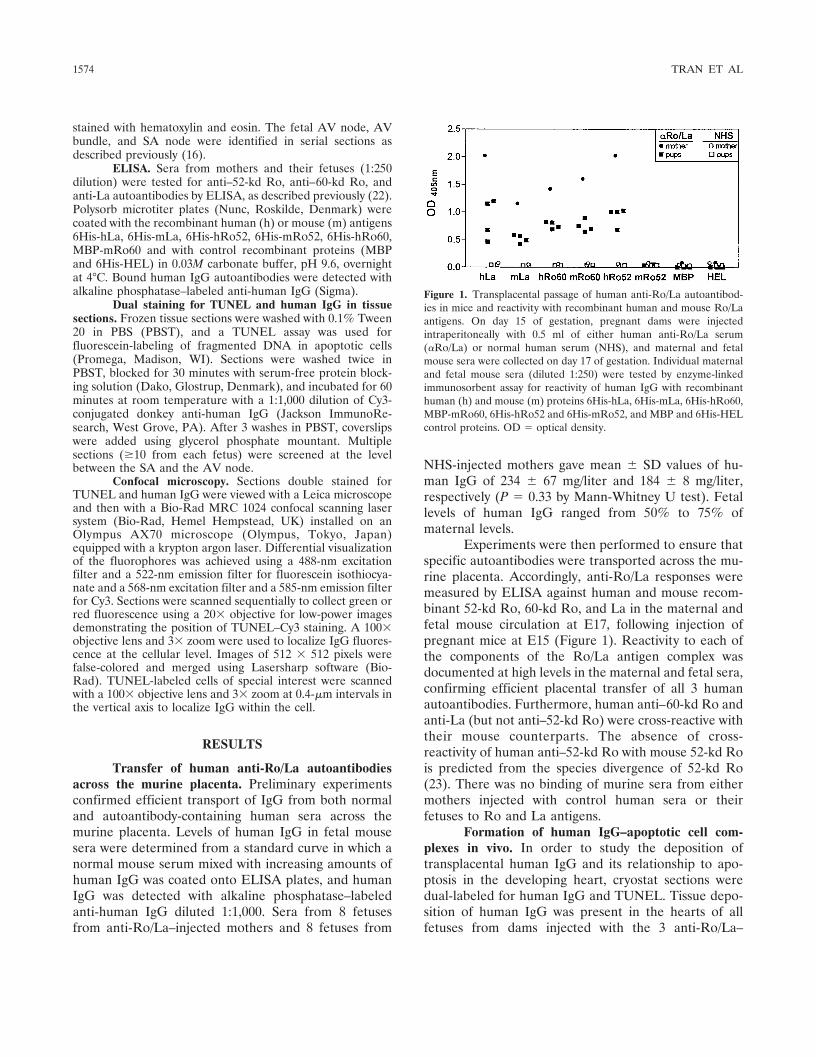
Experiments were then performed to ensure thatspecific autoantibodies were transported across the mu-rine placenta. Accordingly, anti-Ro/La responses weremeasured by ELISA against human and mouse recom-binant 52-kd Ro, 60-kd Ro, and La in the maternal andfetal mouse circulation at E17, following injection ofpregnant mice at E15 (Figure 1). Reactivity to each ofthe components of the Ro/La antigen complex wasdocumented at high levels in the maternal and fetal sera,confirming efficient placental transfer of all 3 humanautoantibodies. Furthermore, human anti–60-kd Ro andanti-La (but not anti–52-kd Ro) were cross-reactive withtheir mouse counterparts. The absence of cross-reactivity of human anti–52-kd Ro with mouse 52-kd Rois predicted from the species divergence of 52-kd Ro(23). There was no binding of murine sera from eithermothers injected with control human sera or theirfetuses to Ro and La antigens.
Formation of human IgG–apoptotic cell com-plexes in vivo. In order to study the deposition oftransplacental human IgG and its relationship to apo-ptosis in the developing heart, cryostat sections weredual-labeled for human IgG and TUNEL. Tissue depo-sition of human IgG was present in the hearts of allfetuses from dams injected with the 3 anti-Ro/La–
Figure 1. Transplacental passage of human anti-Ro/La autoantibod-ies in mice and reactivity with recombinant human and mouse Ro/Laantigens. On day 15 of gestation, pregnant dams were injectedintraperitoneally with 0.5 ml of either human anti-Ro/La serum(�Ro/La) or normal human serum (NHS), and maternal and fetalmouse sera were collected on day 17 of gestation. Individual maternaland fetal mouse sera (diluted 1:250) were tested by enzyme-linkedimmunosorbent assay for reactivity of human IgG with recombinanthuman (h) and mouse (m) proteins 6His-hLa, 6His-mLa, 6His-hRo60,MBP-mRo60, 6His-hRo52 and 6His-mRo52, and MBP and 6His-HELcontrol proteins. OD � optical density.
1574 TRAN ET AL
positive sera. These deposits were observed in the fetalatrial trabeculae and occasionally in the vicinity of theAV node. They were also detected in several othertissues (see below).
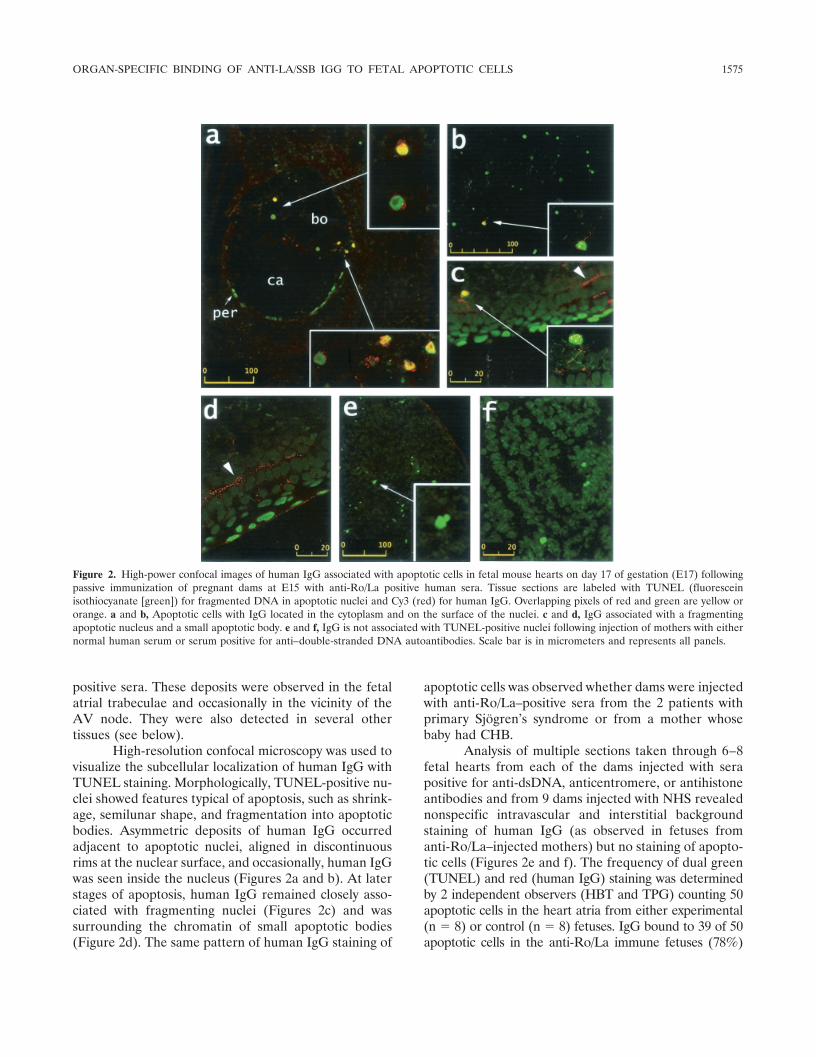
High-resolution confocal microscopy was used tovisualize the subcellular localization of human IgG withTUNEL staining. Morphologically, TUNEL-positive nu-clei showed features typical of apoptosis, such as shrink-age, semilunar shape, and fragmentation into apoptoticbodies. Asymmetric deposits of human IgG occurredadjacent to apoptotic nuclei, aligned in discontinuousrims at the nuclear surface, and occasionally, human IgGwas seen inside the nucleus (Figures 2a and b). At laterstages of apoptosis, human IgG remained closely asso-ciated with fragmenting nuclei (Figures 2c) and wassurrounding the chromatin of small apoptotic bodies(Figure 2d). The same pattern of human IgG staining of
apoptotic cells was observed whether dams were injectedwith anti-Ro/La–positive sera from the 2 patients withprimary Sjogren’s syndrome or from a mother whosebaby had CHB.
Analysis of multiple sections taken through 6–8fetal hearts from each of the dams injected with serapositive for anti-dsDNA, anticentromere, or antihistoneantibodies and from 9 dams injected with NHS revealednonspecific intravascular and interstitial backgroundstaining of human IgG (as observed in fetuses fromanti-Ro/La–injected mothers) but no staining of apopto-tic cells (Figures 2e and f). The frequency of dual green(TUNEL) and red (human IgG) staining was determinedby 2 independent observers (HBT and TPG) counting 50apoptotic cells in the heart atria from either experimental(n � 8) or control (n � 8) fetuses. IgG bound to 39 of 50apoptotic cells in the anti-Ro/La immune fetuses (78%)
Figure 2. High-power confocal images of human IgG associated with apoptotic cells in fetal mouse hearts on day 17 of gestation (E17) followingpassive immunization of pregnant dams at E15 with anti-Ro/La positive human sera. Tissue sections are labeled with TUNEL (fluoresceinisothiocyanate [green]) for fragmented DNA in apoptotic nuclei and Cy3 (red) for human IgG. Overlapping pixels of red and green are yellow ororange. a and b, Apoptotic cells with IgG located in the cytoplasm and on the surface of the nuclei. c and d, IgG associated with a fragmentingapoptotic nucleus and a small apoptotic body. e and f, IgG is not associated with TUNEL-positive nuclei following injection of mothers with eithernormal human serum or serum positive for anti–double-stranded DNA autoantibodies. Scale bar is in micrometers and represents all panels.
ORGAN-SPECIFIC BINDING OF ANTI-LA/SSB IGG TO FETAL APOPTOTIC CELLS 1575

versus 0 of 50 fetuses from mothers injected with NHS.Maternal hearts from anti-Ro/La–injected dams showedonly intravascular and interstitial background staining ofhuman IgG (data not shown).
A postmortem or fixation artifact in our modelwas ruled out based on the following. First, the fetuseswere killed and immediately snap frozen. Second, hu-man IgG deposition was also detected in tissue sectionsfixed directly in paraformaldehyde, a cross-linking re-agent which does not remove lipid (data not shown).Third, IgG did not bind to fetal apoptotic cells afterpassive immunization of mothers with human sera re-acting with other nuclear antigens. Fourth, IgG bound toapoptotic cells in selective organs, yet spared otherorgans with high rates of apoptosis (see below).
To determine whether binding of apoptotic cellsby human IgG in vivo is associated with an inflammatoryresponse, multiple sections of paraffin-embedded fe-tuses (E17) and pups (on the day of delivery) from damsinjected at E15 with anti-Ro/La sera were examined bylight microscopy. There was no evidence of leukocyteinfiltration or inflammation in the vicinity of the AVnode, AV bundle, SA node, atria, or ventricles (data notshown).
Apoptotic cells are bound by anti-La autoanti-bodies. We next investigated the specificity of humanautoantibodies bound to fetal mouse heart apoptoticcells. No cardiac deposition of human IgG was observedin fetuses from dams injected with an anti–60-kd Romonospecific serum despite the fact that anti–60-kd Roreactivity was detected in the fetal circulation (data notshown). Anti–52-kd Ro binding was unable to be testedspecifically in this model, since human anti–52-kd Roautoantibodies do not cross-react with mouse 52-kd Ro(Figure 1).
To investigate whether the anti-La component ofanti-Ro/La–positive serum bound to apoptotic cells, weimmunized mothers with the affinity-purified anti-La au-toantibodies or with the human anti-Ro/La Ig fractiondepleted of anti-La autoantibodies (450 �g of Ig perpregnant mouse), and examined for colocalization of IgGand TUNEL staining in the fetal hearts. In fetuses fromaffinity-purified anti-La autoantibody–injected dams, IgG–apoptotic cell complexes were seen in the identical loca-tions as observed following injection of human anti–52-kdRo/60-kd Ro/La–positive sera, including the fetal AV node(Figures 3a and b). Finally, no IgG binding was detected infetal hearts following injection of dams with the anti-Ro/La
Figure 3. Binding of human IgG (red) to apoptotic cells (TUNEL [bright green]) in the fetal mouse heart following passive transfer ofaffinity-purified anti-La autoantibodies (a and b) but not the anti-La–depleted Ig fraction, which retained anti–52-kd Ro and anti–60-kd Roreactivity (c). a, Low-power fluorescence micrograph of fetal mouse heart showing apoptotic cells (bright green). The dull green background labelingallows identification of the anatomical landmarks of the atrioventricular (AV) node containing 2 TUNEL-positive cells (boxed area labeled b). ias �interatrial septum; mv � mitral valve; tv � tricuspid valve; ivs � interventricular septum. b, Higher magnification view of the boxed area in a,showing anti-La human IgG binding to 1 of the 2 apoptotic cells (top) that were present in the vicinity of the AV node. Inset, Higher magnificationview of area indicated by the arrow, showing the close association of the human IgG to the TUNEL-positive bodies. c, TUNEL-positive cells in theatrial trabeculae of the fetal heart did not show binding of the human IgG depleted of anti-La. The dull red staining represents human IgG in thechamber of the fetal atrium. Inset, Higher magnification view of area indicated by the arrow.
1576 TRAN ET AL
Ig fraction, which had been depleted of anti-La autoanti-bodies but retained anti–52-kd Ro and anti–60-kd Roactivity (Figure 3c).
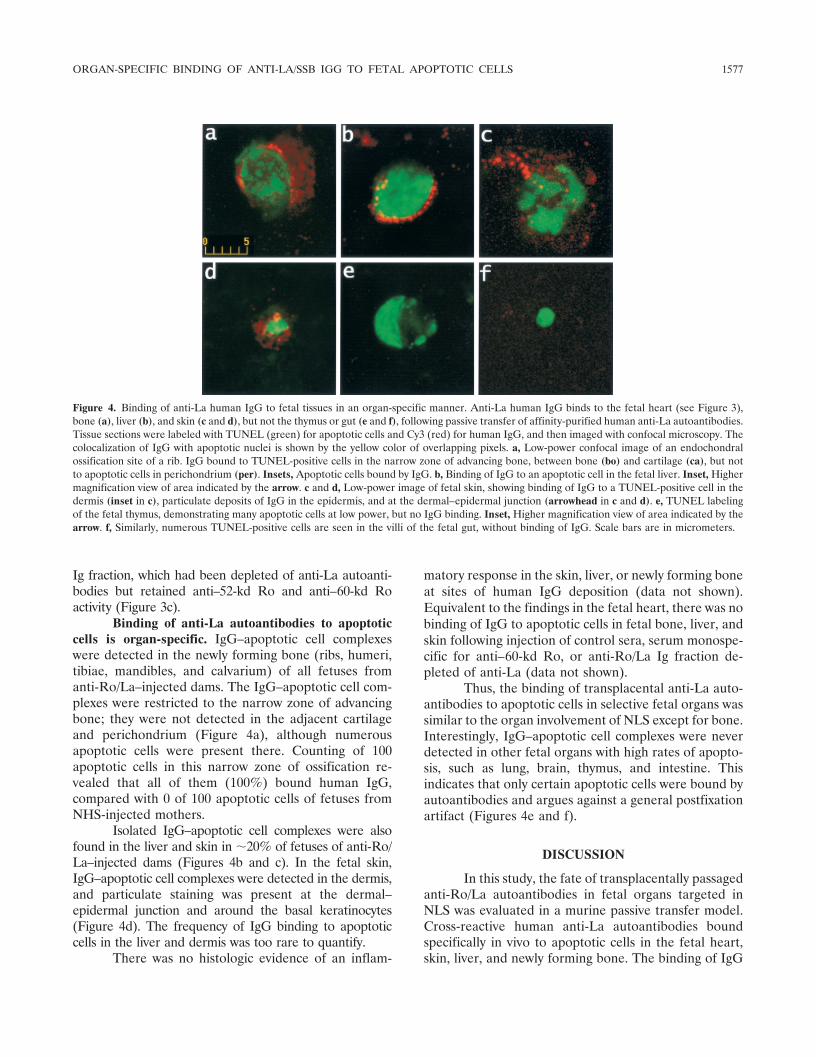
Binding of anti-La autoantibodies to apoptoticcells is organ-specific. IgG–apoptotic cell complexeswere detected in the newly forming bone (ribs, humeri,tibiae, mandibles, and calvarium) of all fetuses fromanti-Ro/La–injected dams. The IgG–apoptotic cell com-plexes were restricted to the narrow zone of advancingbone; they were not detected in the adjacent cartilageand perichondrium (Figure 4a), although numerousapoptotic cells were present there. Counting of 100apoptotic cells in this narrow zone of ossification re-vealed that all of them (100%) bound human IgG,compared with 0 of 100 apoptotic cells of fetuses fromNHS-injected mothers.
Isolated IgG–apoptotic cell complexes were alsofound in the liver and skin in �20% of fetuses of anti-Ro/La–injected dams (Figures 4b and c). In the fetal skin,IgG–apoptotic cell complexes were detected in the dermis,and particulate staining was present at the dermal–epidermal junction and around the basal keratinocytes(Figure 4d). The frequency of IgG binding to apoptoticcells in the liver and dermis was too rare to quantify.
There was no histologic evidence of an inflam-
matory response in the skin, liver, or newly forming boneat sites of human IgG deposition (data not shown).Equivalent to the findings in the fetal heart, there was nobinding of IgG to apoptotic cells in fetal bone, liver, andskin following injection of control sera, serum monospe-cific for anti–60-kd Ro, or anti-Ro/La Ig fraction de-pleted of anti-La (data not shown).
Thus, the binding of transplacental anti-La auto-antibodies to apoptotic cells in selective fetal organs wassimilar to the organ involvement of NLS except for bone.Interestingly, IgG–apoptotic cell complexes were neverdetected in other fetal organs with high rates of apopto-sis, such as lung, brain, thymus, and intestine. Thisindicates that only certain apoptotic cells were bound byautoantibodies and argues against a general postfixationartifact (Figures 4e and f).
DISCUSSION
In this study, the fate of transplacentally passagedanti-Ro/La autoantibodies in fetal organs targeted inNLS was evaluated in a murine passive transfer model.Cross-reactive human anti-La autoantibodies boundspecifically in vivo to apoptotic cells in the fetal heart,skin, liver, and newly forming bone. The binding of IgG
Figure 4. Binding of anti-La human IgG to fetal tissues in an organ-specific manner. Anti-La human IgG binds to the fetal heart (see Figure 3),bone (a), liver (b), and skin (c and d), but not the thymus or gut (e and f), following passive transfer of affinity-purified human anti-La autoantibodies.Tissue sections were labeled with TUNEL (green) for apoptotic cells and Cy3 (red) for human IgG, and then imaged with confocal microscopy. Thecolocalization of IgG with apoptotic nuclei is shown by the yellow color of overlapping pixels. a, Low-power confocal image of an endochondralossification site of a rib. IgG bound to TUNEL-positive cells in the narrow zone of advancing bone, between bone (bo) and cartilage (ca), but notto apoptotic cells in perichondrium (per). Insets, Apoptotic cells bound by IgG. b, Binding of IgG to an apoptotic cell in the fetal liver. Inset, Highermagnification view of area indicated by the arrow. c and d, Low-power image of fetal skin, showing binding of IgG to a TUNEL-positive cell in thedermis (inset in c), particulate deposits of IgG in the epidermis, and at the dermal–epidermal junction (arrowhead in c and d). e, TUNEL labelingof the fetal thymus, demonstrating many apoptotic cells at low power, but no IgG binding. Inset, Higher magnification view of area indicated by thearrow. f, Similarly, numerous TUNEL-positive cells are seen in the villi of the fetal gut, without binding of IgG. Scale bars are in micrometers.
ORGAN-SPECIFIC BINDING OF ANTI-LA/SSB IGG TO FETAL APOPTOTIC CELLS 1577

anti-La was specific to the fetus and was not detected inthe maternal heart or skin. These findings are an exten-sion of previous work in which we demonstrated that Laautoantigen is clustered near the surface of apoptoticcells in fetal mouse cardiac conducting tissue and thatanti-La autoantibodies bind to the surface of culturedapoptotic human myocardiocytes (14–16).
In the murine model presented herein, epitopeson La molecules redistributed during apoptosis arerecognized in vivo by anti-La autoantibodies that aretransported across the placenta. Interestingly, transpla-cental anti-dsDNA autoantibodies did not bind to apo-ptotic cells in our model, suggesting that DNA remainson the inside of the apoptotic bodies and is not accessi-ble on the outside for binding by the autoantibodies. Thepresence of human IgG in the interstitium is consistentwith passage of IgG and other plasma proteins throughintercellular clefts between endothelial cells (24). Fromthe present study, it is not possible to determine whetherthe intracellular binding of anti-La is due to an activeprocess or the loss of membrane integrity seen in thelater stages of apoptosis. We would favor a process ofinternalization of complexes of anti-La and La antigenformed at the surface of the apoptotic cells, rather thannonspecific leakage through the membrane, since auto-antibodies directed against intracellular antigens (DNA,centromere, histone) did not bind apoptotic cells.
Since human anti–52-kd Ro autoantibodies donot cross-react with mouse 52-kd Ro, we were not ableto test for binding of anti–52-kd Ro to fetal apoptoticcells in this murine model. It remains unexplained whythe anti–60-kd Ro autoantibodies did not bind despitetransport across the placenta. However, it is of interestthat in another reported study, anti-La autoantibodies,but not anti–60-kd Ro autoantibodies, immunoprecipi-tated surface-biotinylated substrate in cultured cardio-cytes (15).
Although the IgG–apoptotic cell complexes weredetected in all fetal hearts following injection of thedams with anti-La autoantibodies, they were not associ-ated with histologically evident inflammation in thefetuses and pups. Under the experimental conditions ofthis study, placental transport of autoantibodies is notsufficient to produce lesions characteristic of NLS. Thisparallels clinical observations in which fewer than 5% ofchildren born to anti-Ro/La-positive mothers have heartblock (5). A reasonable hypothesis is that, analogous tothe murine model, cardiac IgG–apoptotic cell complexesmay be formed in most, if not all, fetuses from motherspositive for anti-Ro/La autoantibodies, but in the major-ity of cases, the complexes are cleared without perma-nent sequelae. Anti-Ro/La autoantibodies are necessary,
but not sufficient, to cause CHB. Coincident factorsduring a vulnerable period of fetal cardiac development,such as local hypoxia, mild viral infection, trauma, orspecific fetal genetic factors related to phagocytosis, mayincrease the apoptotic cell load and promote clearancevia inflammatory pathways, leading to tissue damageand CHB in rare cases.
Anti-La, but not human anti-Ro, autoantibodiesbound to the fetal skin and gave rise to particulatestaining of both the dermal–epidermal junction and thebasal layers of the epidermis. In contrast to the fetalheart, the epidermal IgG deposits were not coincidentwith TUNEL staining, although IgG–apoptotic cell com-plexes were occasionally detected in the fetal dermis.Particulate epidermal staining of IgG has been reportedin human skin–grafted mice after injection of purifiedhuman anti-Ro, but the binding of anti-La autoantibod-ies was not tested in this model (25,26). Deposition oftransplacental anti-La autoantibodies in the fetal skinwas not associated with histologic evidence of inflamma-tion, consistent with a requirement for environmentalfactors, such as ultraviolet irradiation which augmentsthe physiologic apoptosis, or genetic factors for skindisease to occur (27).
One of the most intriguing observations of thisstudy was the organ specificity of anti-La–apoptotic cellcomplex formation, with transplacental anti-La autoan-tibodies binding to apoptotic cells in the fetal heart(atrial trabeculae and AV node), skin, and liver. Thispattern overlaps with the organ involvement observed inNLS, in which CHB and skin rash are the two classicalmanifestations, while hepatic involvement has beenrarely reported but may be underestimated (4,5,9). Wealso detected binding of apoptotic cells by anti-Laautoantibodies in the narrow zone of ossification ofnewly forming bone, but we are unaware of reports ofskeletal damage in NLS. Since binding of anti-La auto-antibodies to apoptotic cells is a prominent feature inthe fetal bones, investigation of babies with NLS forsubtle skeletal changes may be warranted.
Although there was a striking distribution of theimmune complexes in organs known to be targeted inNLS, and a conspicuous absence of IgG binding in otherfetal organs with high rates of apoptosis, such as thymus,lung, brain, and intestine, a biologic explanation is notapparent at this time. La antigen is redistributed to thesurface of apoptotic cells irrespective of their tissueorigin (16), yet the binding of anti-La is organ-specific inthe passive transfer model. We suggest that binding ofapoptotic cells by transplacentally passaged anti-La au-toantibodies may be influenced by other factors, such asthe kinetics of apoptotic cell clearance, the level of tissue
1578 TRAN ET AL

vascularization, and the integrity of the blood–tissuebarrier in selected organs.
In conclusion, transplacental human anti-La au-toantibodies have been demonstrated to bind specificallyto apoptotic cells in selective fetal mouse organs in vivo,similar to the organ involvement in neonatal lupussyndrome. These observations support a role for opso-nization of fetal apoptotic cells by maternal autoantibod-ies in mediating tissue injury in NLS, although addi-tional factors are required for disease expression.Further studies are under way in the passive transfermodel, investigating environmental and/or genetic fac-tors that may shift the clearance of IgG–apoptotic cellcomplexes toward proinflammatory pathways and pre-dispose the fetus to lesions characteristic of NLS.
ACKNOWLEDGMENTS
We thank Dr. Dzung D. Nguyen, Ms Dimitra Berou-kas, Ms Nhuan T. Nguyen, Ms Michel Lewis, and Ms TheresaFischer for technical assistance. Advisory comments by Dr.Sally Waterman are appreciated.
REFERENCES
1. Franco BL, Weston WL, Peebles C, Forstot SL, Pheanuphak P.Autoantibodies directed against sicca syndrome antigens in neo-natal lupus. J Am Acad Dermatol 1981;4:67–72.
2. Scott JS, Maddison PJ, Taylor PV, Esscher E, Scott O, SkinnerRD. Connective tissue disease, antibodies to ribonucleoprotein,and congenital heart block. N Engl J Med 1983;4:209–12.
3. Reed DR, Lee LA, Harmon C. Autoantibodies to SS-A/Ro ininfants with congenital heart block. J Pediatr 1983;103:889–91.
4. Lee LA. Neonatal lupus erythematosus. J Invest Dermatol 1993;100:9S–13S.
5. Buyon JL. Neonatal lupus syndromes. In: Lahita RG, editor.Systemic lupus erythematosus. 3rd ed. San Diego: Academic Press;1999. p. 337–59.
6. Ho YS, Esscher E, Anderson RH, Michaelsson M. Anatomy ofcongenital heart block and relation to maternal anti-Ro antibod-ies. Am J Cardiol 1986;58:291–4.
7. Tseng C, Friedman D, Buyon JP. Spectrum of cardiac histopathol-ogy in cases of autoimmune-associated congenital heart block(CHB) obtained from the Research Registry for Neonatal Lupus.Arthritis Rheum 1997;40 Suppl 9:S333.
8. Ristic AD, Maisch B. Cardiac rhythm and conduction distur-bances: what is the role of autoimmune mechanisms? Herz2000;25:181–8.
9. Dorner T, Feist E, Pruss A, Chaoui R, Goldner B, Hiepe F.Significance of autoantibodies in neonatal lupus erythematosus.Int Arch Allergy Immunol 2000;123:58–66.
10. Garcia S, Campos-de-Carvalho AC. Neonatal lupus syndrome: theheart as a target of the immune system. An Acad Bras Cienc2000;72:83–9.
11. Miranda-Carus ME, Boutjdir M, Tseng CE, DiDonato F, ChanE-K, Buyon JP. Induction of antibodies reactive with SSA/Ro-
SSB/La and development of congenital heart block in a murinemodel. J Immunol 1998;161:5886–92.
12. Mazel JA, El-Sherif N, Buyon JL, Boutjdir M. Electrocardio-graphic abnormalities in a murine model injected with IgG frommothers of children with congenital heart block. Circulation1999;99:1914–8.
13. Casciola-Rosen LA, Anhalt G, Rosen A. Autoantigens targeted insystemic lupus erythematosus are clustered in two populations ofsurface structures on apoptotic keratinocytes. J Exp Med 1994;179:1317–30.
14. Miranda ME, Tseng C-E, Rashbaum W, Ochs RL, Casiano AA,DiDonato F, et al. Accessibility of SSA/Ro and SSB/La antigens tomaternal autoantibodies in apoptotic human fetal cardiac myo-cytes. J Immunol 1998;161:5061–9.
15. Miranda-Carus M-E, Askanase AC, Clancy RM, DiDonato F,Chou T-M, Libera MR, et al. Anti-SSA/Ro and anti-SSB/Laautoantibodies bind the surface of apoptotic fetal cardiocytes andpromote secretion of TNF-alpha by macrophages. J Immunol2000;165:5345–51.
16. Tran HB, Ohlsson M, Beroukas D, Hiscock J, Bradley J, Buyon JP,et al. Subcellular redistribution of La/SS-B autoantigen duringphysiologic apoptosis in the fetal mouse heart and conductionsystem: a clue to the pathogenesis of congenital heart block.Arthritis Rheum 2002;45:202–8.
17. Vitali C, Bombardieri S, Moutsopoulos HM, Balestrieri G, Ben-civelli W, Bernstein RM, et al. Preliminary criteria for theclassification of Sjogren’s syndrome: results of a prospectiveconcerted action supported by the European Community. Arthri-tis Rheum 1993;36:340–7.
18. Schoenlebe J, Buyon JP, Zitelli BJ, Friedman D, Greco MA,Knisely AS. Neonatal hemochromatosis associated with maternalautoantibodies against Ro/SS-A and La/SS-B ribonucleoproteins.Am J Dis Child 1993;147:1072–5.
19. McNeilage LJ, Umapathysivam K, Macmillan E, Guidolin A,Whittingham S, Gordon T. Definition of a discontinuous immu-nodominant epitope at the NH2 terminus of the La/SS-B ribonu-cleoprotein autoantigen. J Clin Invest 1992;89:1652–6.
20. Steinbuch M, Audran R. The isolation of IgG from mammaliansera with the aid of caprylic acid. Arch Biochem Biophys 1969;134:279–84.
21. Jacobson L, Polizzi A, Morris-Kay G, Vincent A. Plasma fromhuman mothers of fetuses with severe arthrogryposis multiplexcongenita causes deformities in mice. J Clin Invest 1999;103:1031–8.
22. Keech CL, Gordon TP, McCluskey J. The immune response to52-kilodalton Ro and 60-kilodalton Ro is linked in experimentalautoimmunity. J Immunol 1996;157:3694–9.
23. Keech CL, Gordon TP, McCluskey J. Structural differences be-tween the human and mouse 52-kDa Ro autoantigens associatedwith poorly conserved autoantibody activity across species. ClinExp Immunol 1996;104:255–63.
24. Guyton AC, Hall JE. Textbook of medical physiology. 10th ed.Philadelphia: WB Saunders; 2000. p. 164.
25. Lee LA, Weston WL, Krueger GG, Emam M, Reichlin M, StevensJO, et al. An animal model of antibody binding in cutaneous lupus.Arthritis Rheum 1986;29:782–8.
26. Lee LA, Gaither KK, Coulter SN, Norris DA, Harley JB. Patternof cutaneous immunoglobulin G deposition in subacute cutaneouslupus erythematosus is reproduced by infusing purified anti-Ro(SSA) autoantibodies into human skin-grafted mice. J Clin Invest1989;83:1556–62.
27. Lee LA, Farris AD. Photosensitivity diseases: cutaneous lupuserythematosus. J Investig Dermatol Symp Proc 1999;4:73–8.
ORGAN-SPECIFIC BINDING OF ANTI-LA/SSB IGG TO FETAL APOPTOTIC CELLS 1579




















