
Genetic Reactivation of Cone Photoreceptors Restores Visual Responses in Retinitis Pigmentosa

Journal o f Neurocytology 8, 1 8 1 - 1 9 5 (1979)
Ultrastructure of photoreceptors in the eye of Hermissenda labelled with intracellular injections of horseradish peroxidase T E R R Y C R O W , E L I A H U H E L D M A N , V A C H I K H A C O P I A N , R O B E R T E N O S and D A N I E L L. A L K O N
Laboratory of Biophysics, Section on Neural Systems, IRP, NINCDS, National Institutes of HealtO and Marine Biological Laboratory, Woods Hole, Massachusetts 02543, U.S.A.
Received 1 June 1978; revised 3 October and 30 November 1978; accepted 5 December 1978
Summary The terminal processes of single and of pairs of identified photoreceptors in the eyes of the nudi- branch mollusc Hermissenda crassicornis were studied by light and transmission electron micro- scopy after their somata were labelled by intracellular iontophoresis of horseradish peroxidase (HRP). The HRP spread from the somata into the axons and fine terminal processes within the neuropil of the cerebropleural ganglia. The photoreceptors ended in extensive secondary branches in the neuropil where previous electrophysiological studies had indicated the probable site of synaptic interactions between photoreceptors. Clear round vesicles (54-126 nm diameter)within labelled processes were similar to the vesicles found in the somata and axon hillocks. The terminal processes of pairs of type B photoreceptors contained different intensities of the HRP label. Uniform intensities of the HRP label were found in the terminal processes of single type B photo- receptors. These differences in intensity suggested that the terminal processes were from different type B photoreceptors. This finding suggests that the connections between type B photoreceptors are probably monosynaptic.
Int roduct ion
The nudibranch mollusc Hermissenda crassicornis has proved useful for the study of synaptic physiology in well defined neural networks (Alkon, 1973a, b; Alkon et al., 1978). A neural network, consisting of two interacting sensory pathways, the visual and vestibular, may mediate previously reported behavioural modifications of Hermissenda (Alkon, 1974, 1975; Crow and Alkon, 1978a, b). Previous work on the ultrastructure of the Hermissenda eye suggested that the synaptic interactions between photoreceptors may take place between or close to the somata, although an analysis of identified photoreceptor processes within the neuropil was not under- taken (Eakin et al., 1967; Stensaas etal., 1969). Recent electrophysiological evi- dence has indicated that the synaptic interaction between photoreceptors is mono-
�9 1979 Chapman and Hall Ltd. Printed in Great Britain

182 CROW, HELDMAN, HACOPIAN, ENOS and ALKON
synaptic and the locus of this interaction is in the neuropil, between the secondary processes of the photoreceptors (Alkon and Fuortes, 1972).
Numerous cell marking techniques have been developed to examine the general morphology of identified neurons of invertebrates (Stretton and Kravitz, 1968; Pit- man et al., 1972; Christensen, 1973; Gillette and Pomeranz, 1973). A few techniques have been used successfully with electron microscopy (Pentreath and Cottrell, 1974; Bailey etal., 1976; Muller and McMahan, 1976; Thompson etal., 1976a, b). The histochemical reaction of the enzyme horseradish peroxidase (HRP) is a suitable marker for both light (Straus, 1964a, b) and electron microscopy (Graham and Karnovsky, 1966).
Individual and pairs of photoreceptors within the same eye of Hermissenda were marked in the present study by the intracellular iontophoresis of HRP in order to identify the primary and secondary processes within the neuropil. We examined the stained processes with light and electron microscopic techniques in order to investi- gate relationships between photoreceptors and to study the ultrastructure of the labelled terminals.
Materials and methods
Specimens of Hermissenda crassicornis were provided by Mr Michael Morris of the Sea Life Supply, Sand City, California. The animals were maintained in individual plastic containers in a flow- through fresh seawater aquarium at 15 ~ C.
Injection of horseradish peroxidase (HRP) The circumoesophageal nervous system was dissected, isolated, and pinned to the Sylgard (Dow Coming Co.) bottom of a recording chamber filled with seawater at room temperature (22~ The nervous system was incubated in a solution of digestive enzyme prior to the electrode penetration of photoreceptors (Protease, Type VII, Sigma). Photoreceptors were identified on the basis of their response to light, electrophysiological characteristics and their position within the eyes (Alkon and Fuortes, 1972). It has been previously demonstrated (Alkon, 1974a) that these charac- teristics are similar from preparation to preparation.
Bevelled mieroelectrodes containing several strands of fibreglass were filled with a filtered solution of 1% HRP (Type VI, Sigma) in 0.05 M Tris-HC1 buffered (pH 8.5) 0.2 M KC1. Electrode resistance varied between 80 and 150 m~2. The HRP was iontophoresed by 200-400 ms pulses of 10-20 nA constant depolarizing current at 2-3 pulses/s for 30-60 min. Signals were amplified and displayed by conventional techniques. Current passage was monitored by recording the voltage drop across a fixed resistance in the ground path.
Histochemistry Histological procedures similar to those described by Muller and McMahan (1976) were employed. Following the injection, preparations were incubated in seawater (60 min) to permit the spread of the enzyme into the fine processes of the photoreceptors. At the end of this period ganglia were fixed for 30 min with 2.5% glutaraldehyde and 1% paraform- aldehyde in 0.12 M phosphate-buffered (pH 7.4) 4% sucrose. The fixed ganglia were rinsed in 25% sucrose with 0.12 M phosphate buffer (pH 7.4) and incubated in 0.5 mg/ml 3.3'-diamino- benzidine tetrahydrochloride in 25% sucrose with 0.12 M phosphate buffer (pH 7.4). One or two drops of 3% H2 02 were added to the incubation solution and the reaction was followed visually.

Ultrastructure of photoreceptors labelled with horseradish peroxidase 183
Light microscopy Ganglia were fixed in 0.12 M phosphate-buffered (pH 7.4) 2.5% glutaraldehyde, postfixed in 0.12 M phosphate-buffered 2% OsO4 (pH 7.4) and 4% sucrose, dehydrated through graded alcohols, and embedded in Epon-Araldite. Serial 1 - 1 0 #m sections were cut through the ganglion, mounted on glass slides, examined in a microscope and photographed. In some preparations thick sections containing stained secondary processes were glued to trimmed blocks for subsequent thin sectioning. Whole mounts were fixed in glutaraldehyde, dehydrated as described, cleared in xylene and photographed.
Electron microscopy Fixed ganglia were rinsed in 0.12 M phosphate-buffered (pH 7.4) sucrose 25% and postfixed in 2% OsO4 in 0.12 M phosphate buffer (pH 7.4) and 4% sucrose. The ganglia were then rapidly dehydrated in increasing concentrations of cold ethanol and embedded in Epon-Araldite. After polymerization, thin sections were cut for electron microscopy using glass and diamond knives on an LKB ultramicrotome. Thin sections were collected on grids, stained with uranyl acetate and lead citrate and examined with a Jeol 100 C or Phillips 300 electron microscope. In some cases only alternate sections were stained. The alternate stained and unstained sections were examined with the electron microscope and compared to aid in the identification of terminals containing HRP.
Results
The anatomy of the visual system has been previously described (Eakin et al., 1967; Stensaas etal., 1969; Alkon and Fuortes, 1972). The eyes of Hermissenda are situated on the dorsal surface of the circumoesophageal nervous system between the cerebropleural and pedal ganglia (see Fig. 1). Each eye contains five photoreceptors, two type A and three type B which can be distinguished by electrophysiological and morphological criteria (Alkon, 1976). Type A cells are approximately 25/am in diameter and are located in the anteroventral portion of the eye. The type B cells are approximately 40/am in the longest diameter and are located in the posterodorsal portion of each eye. Type B photoreceptors, unlike the type A, are spontaneously active in darkness and are responsive to dim illumination. Two type B photoreceptors are shown in the diagram of the nervous system in Fig. 1. The axons of the photo- receptors emerge from the posterior portion of the eye and form the optic nerve. The optic nerve courses through the optic ganglion and joins with axons from optic ganglion cells to form the optic tract which then enters the cerebropleural ganglion and terminates in the neuropil (Fig. 1).
Light microscopic observations Photoreceptor axons terminated in a series of fine secondary processes or branches in the neuropil approximately 20/am after entering the cerebropleural ganglion. This can be visualized in the photographs of a whole mount preparation shown in Fig. 2. In this preparation, the type A photoreceptor was injected with HRP. The HRP- labelled axon (Fig. 2B) entered the cerebropleural ganglion and ended in a cluster of secondary branches approximately 75-100/am from the eye (Fig. 2A). The depolar- izing current pulses used to iontophorese the HRP resulted in an increase in the spon-

184 CROW, H E L D M A N , H A C O P I A N , ENOS and A L K O N
NP ~ NP
E
/
NP ~ t , 1 ~ ~ ~)Q--~.~..~.~-] /--4 NP
ST CPG PG
A
L/• 12mV
- -9 o. s
Fig. 1. Diagram of the dorsal surface of the circumoesophageal nervous system of Hermissenda crassicornis. Two type B photoreceptor somata, black areas in each eye, their axons and terminal processes have been drawn schematically. Photoreceptors receive synaptic input from other photoreceptors at their terminal endings in the neuropil. E, eye; OG, optic ganglion; NP, neuropil; ST, statocyst; CPG, cerebropleural ganglia; PG, pedal ganglia. Intracellular recordings from two type B photoreceptors within the same eye are shown in A to illustrate the direct inhibitory interactions between type B photoreceptors. A current pulse delivered to one type B photo- receptor evokes a spike (top trace) which is followed by an inhibitory post-synaptic potential in the second type B photoreceptor (bottom trace). This result is invariant from preparation to preparation. The top of the spike in A has been retouched.

Ult ras t ruc ture o f p h o t o r e c e p t o r s labelled with horseradish peroxidase 185
Figs. 2A and B. Photomicrographs at two depths of focus of a type A photoreceptor from a whole mount of the cerebropleural ganglion. The photoreceptor was injected with HRP and reacted with diaminobenzidine and H202. The primary process (axon) enters the cerebropleural ganglion (2B) originating from the eye. A cluster of secondary processes terminates in the neuropil (2A). Scale bar: 100 gm. Fig. 3. Bright field photomicrographs of a thick section showing two type B photoreceptors within the same eye injected with HRP. The two arrows mark the two labelled axons. (Inset) phase contrast micrograph showing point where the two axons (arrows) enter the cerebropleural ganglion.
t aneous act ivi ty o f the p h o t o r e c e p t o r spikes, m e m b r a n e depo la r iza t ion , and eventu- ally a r educ t ion in the ampl i tude of the depolar iz ing genera to r po ten t ia l s to light flashes. However , these e lec t rophys io logica l changes were reversible. Int racel lular recordings f rom the p h o t o r e c e p t o r s af ter the inject ion per iod revealed tha t the

186 CROW, HELDMAN, HACOPIAN, ENOS and ALKON
spontaneous activity gradually returned to the normal level for dark adapted photo- receptors and the generator potentials elicited by light flashes were of normal amplitude and duration.
An example of two photoreceptors within the same eye of Hermissenda injected with HRP is shown in Fig. 3. The two HRP-labelled axons of the two type B photo- receptors in the optic nerve enter the cerebropleural ganglion approximately 60 #m from the posterior part of the eye. The area where the optic nerve enters the gang- lion can be clearly visualized under phase contrast (inset, Fig. 3). Serial sections through the cerebropleural ganglion of the double HRP-labelled preparation shown in Fig. 3 revealed two overlapping clusters of stained secondary processes in the neuropil (Figs. 4 and 5). These serial sections starting in the area where the optic nerve entered the ganglion (see inset, Fig. 3), allowed easy identification of the two labelled axons in the optic tract and their respective terminations in the neuropil. Serial sections from preparations in which one type B photoreceptor was injected with HRP were collected and revealed a much smaller area of labelled secondary processes in the neuropil (Fig. 6). Small differences in the number of observed endings between preparations may occur because of the cutting angle through the block. In all cases the HRP-labelled processes from one injected type B photoreceptor could be clearly distinguished from background staining. Sections from control preparations treated with the same histochemical procedures as the HRP-labelled preparations did reveal some cases of nonspecific background staining. The degree of nonspecific background staining was usually related to the incubation time in diaminobenzidine. Nonspecific background staining was widely dispersed through- out the ganglia and was never concentrated in areas of intense staining as shown in Figs. 4--6. Endogenous peroxidase was observed in the somata of cells in the cere- bropleural ganglion and not in the neuropil.
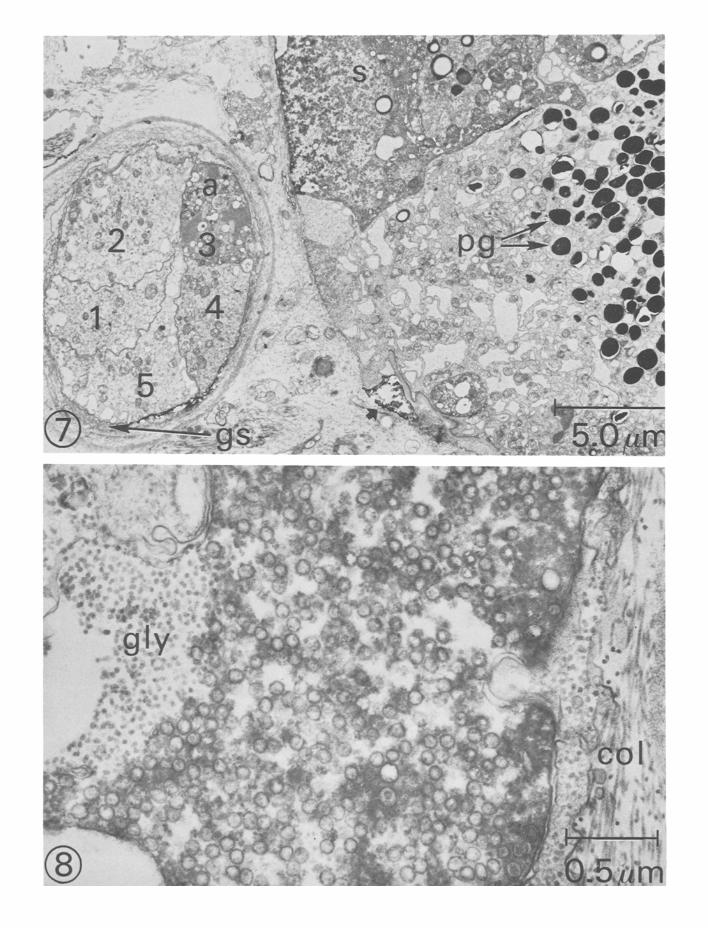
Electron microscopic observations The structure of photoreceptor axons and terminal processes in the cerebropleural ganglion was investigated by examining darkly stained areas containing HRP. A section through part of the eye and a cross-section through the optic nerve is shown in Fig. 7. The soma of one type B photoreceptor is densely filled with the HRP (Fig. 7). The labelled area is surrounded by adjacent pigmented cells containing numerous pigment granules. The cross-section of the optic nerve revealed a darkly labelled axon and four unlabelled photoreceptor axons. The axon hillock region of
Fig. 4. Photomicrograph of a thick section through the cerebropleural ganglion of the preparation shown in Fig. 3 showing HRP-labelled secondary processes (arrows) in the neuropil. Fig. 5. A section of the same preparation as in Figs. 3 and 4 at higher magnification showing two overlapping areas of terminal branches (arrows). Fig. 6. A section through the cerebropleural ganglion showing terminal branches (arrows) follow- ing injection of a single type B photoreceptor with HRP.


188 C R O W , H E L D M A N , H A C O P I A N , E N O S and A L K O N
the stained photoreceptor is shown in Fig. 8. The area filled with electron-dense material contains numerous small (60 -70 nm) clear round vesicles and granular material thought to be glycogen.
Electron microscopic analysis of the terminal processes from four preparations where single type B photoreceptors were injected with HRP revealed stained pro- cesses containing numerous vesicles and mitochondria. These will be referred to as identified processes. An examination of 12 sections containing identified processes showed that the processes contained clear round (spherical) vesicles with a range in diameter of 5 4 - 1 2 0 n m (Fig. 9). Frequently these labelled areas contained mitochondria and infrequently vesicles containing a granular material were found. A single identified process surrounded by numerous unlabelled processes was often observed. An example of a terminal process from a single identified photoreceptor is shown in Fig. 9. The labelled terminal contains aggregations of vesicles and abuts against several processes that do not contain vesicles. Small clusters of two to five identified processes were often found following injection of a single photoreceptor. In some cases these involved three labelled areas containing vesicles surrounding a clear centre process. Infrequently, as many as five labelled processes could be found in these clusters. In some of these examples two stained processes were found partially to surround a central area without vesicles. Seven examples of the latter were found from an examination of the 12 sections.
Ten sections containing identified processes were examined from two prepara- tions in which two type B photoreceptors, within the same eye, had been injected with HRP. These processes contained clear round vesicles ( 5 6 - 1 0 6 n m ) and occasionally mitochondria and were often grouped in large clusters of up to 10 processes.
In further contrast to the situation after injection of a single photoreceptor, some of the identified processes surrounded another centrally situated identified process. An example of this is shown in Fig. 10. In this example the lightly stained process (asterisk) may be from a different photoreceptor than the three darkly stained processes that surround it. However, some examples of labelled processes surround- ing an unlabelled process were found after injecting two photoreceptors. A profile from a double stained section is shown in Fig. 11. In this example from an experi- ment in which two type B photoreceptors were injected, terminals from a lightly
Fig. 7. Electron micrograph showing HRP-labelled photoreceptor soma (s) and cross-section of optic nerve. The pigment cells contain numerous pigment granules (pg). The optic nerve consists of five axons ( 1 - 5 ) surrounded by a glial sheath (gs). A darkly labelled axon (a) corresponds to the labelled soma. A partially filled area indicated by arrow (bot tom centre) may be the result of some leakage from a previous unstable electrode penetration. Fig. 8. Electron micrograph of an axon hillock of a HRP-labelled photoreceptor . The area contains numerous vesicles 6 0 - 8 0 nm in diameter, and granular material thought to be glycogen (gly). Outside the stained axon are collagen fibrils (col). A small process on the right of the micrograph invaginates the axon.

190 c r o w , H E L D M A N , H A C O P I A N , E N O S and A L K O N
Fig. 9. Electron micrograph of a terminal process following injection of a single type B photo- receptor. The terminal process (asterisk) contains numerous clear round vesicles and abuts against several clear processes. Scale bar: 0 .5/ lm. Fig. 10. A moderately labelled terminal process (asterisk) surrounded by three darkly labelled processes following injection of HRP into two type B photoreceptors. Scale bar: 0.5/~m. Fig. 11. Two terminal processes, probably from two different photoreceptors after injection of two type B photoreceptors. The lightly labelled process (asterisk) contains numerous clear round vesicles and the darkly labelled process contains vesicles and mitochondria. Stain contamination may have obscured some detail in unlabelled areas. Scale bar: 0.5 ~tm.

Ultrastructure of photoreceptors labelled with horseradish peroxidase 191
stained process containing vesicles (asterisk) abuts a darkly stained process contain- ing vesicles and mitochondria. Several examples of such differences in the intensity of labelling in adjacent profiles were found after injection of two photoreceptors. The differences are not due to differential staining with lead citrate since they were also apparent in the unstained sections. This suggests that the processes may have been from two different photoreceptors. We found numerous examples of two HRP-labelled processes, both containing vesicles and mitochondria, contacting each other following injection of a single photoreceptor. In these cases both processes had approximately equal concentrations of vesicles and comparable intensity of the label. Synaptic membrane specializations were not found in the terminal regions of the identified photoreceptor processes.
Discussion
A number of electrophysiological tests have been used to demonstrate direct (mono- synaptic) connections (for review see Berry and Pentreath, 1976). One test is usually not sufficient for demonstrating direct connections. Anatomical evidence in con- junction with several electrophysiological tests would provide strong support for direct connections.
These results are consistent with electrophysiological evidence indicating that the connections between type B photoreceptors in the eye of Hermissenda are mono- synapic. Results from previous electrophysiological studies as well as the data presented in Fig. 1A have fulfilled some of the criteria proposed by Berry and Pentreath (1976) for testing monosynaptic connections. Simultaneous recordings from pairs of photoreceptors within the same eye have shown that the ceils are mutually inhibitory (Alkon and Fuortes, 1972). Electrical stimulation of one photoreceptor evoked unitary IPSPs in the other with a constant latency (see Fig. 1A). Spontaneous spikes in one cell were synchronized with unitary IPSPs in the other photoreceptor and the IPSPs followed impulses one-for-one with a constant delay during high frequency stimulation. This mutual inhibition was shown to be characteristic of the interactions between the type B photoreceptors and between the type A and type B photoreceptors (Alkon and Fuortes, 1972).
The data of the present study are also in agreement with the previous electro- physiological evidence indicating that the locus of synaptic interaction between secondary processes is in the neuropil. Simultaneous soma-axon recordings from the same photoreceptor revealed that the IPSPs were larger and had shorter time to peak at the axon recording site. Sectioning the optic tract between the optic ganglion and the cerebropleural ganglion eliminated synaptic activity as recorded from the photoreceptor somata (Alkon and Fuortes, 1972; Alkon and Grossman, 1977). The electrophysiological evidence and tile differences between labelled terminals after injecting two as opposed to one photoreceptor strongly suggest, although does not unequivocally demonstrate, that the connections between type B photoreceptors are monosynaptic.

192 CROW, HELDMAN, HACOPIAN, ENOS and ALKON
It has been suggested that the synaptic interaction between photoreceptors occurs in the somata, axon hillock, between axons in the optic nerve, or near the optic ganglion (Eakin et al., 1967; Stensaas et al., 1969). This suggestion was based on the observation that the somata and axon hillock contained numerous aggregations of vesicles. These authors also found areas of infolding between membranes and small projections that invaginated axons. We also have noted this (see Fig. 8) and, in addition, have found areas in the neuropil which contain aggregations of vesicles in identified photoreceptor processes. Contacts between several identified terminal processes observed in the present study would provide a means for the direct inhibitory interactions between photoreceptors. A photoreceptor is both pre- and post-synaptic to other photoreceptors. This interaction then could be in distinct regions of the same terminal or in different terminal branches of the same photo- receptor. The neuropil we studied contains processes from cells other than photo- receptors. Unlabelled processes, such as those contacting a labelled central process (as in Fig. 10) may be from another photoreceptor since at most only two of five photoreceptors were injected with HRP in one eye, or from another unidentified neural element.
Uitrastructural studies of identified nerve endings in molluscs 'are difficult to interpret because of an absence of specific criteria for defining synapses. The synap- tic morphology of molluscs may be similar to vertebrates, although a comprehensive analysis of the neuropil of molluscs comparable to vertebrate work (Gray, 1971; Palay and Chan-Palay, 1975; Peters et al., 1976) has not been reported. Specialized synaptic.regions resembling vertebrate synapses were rarely observed in early work with Aplys ia , w~ th the most frequent examples being vesicle-filled terminal processes abutting clear neuronal processes (Coggeshall, 1967). An examination of the neuropil in central ganglia of the mollusc Planorbis corneus did reveal several types of simple specializations (Berry et al., 1975; Pentreath et al., 1975b). These included membrane thickening and apposition with vesicle aggregations on one side. Some of the specializations of apposed axonal processes resembled desmosomes. In many cases processes were varicose with vesicle aggregations. However, these specializa- tions were not found in an identified neuron in the central ganglia of Planorbis (Pentreath and Berry, 1975a). We did not find any examples of membrane specializations in the identified photoreceptor processes of Hermissenda, and even after staining with ethanolic phosphotungstic acid (Bloom and Aghajanian, 1966) synapfic densities were not identified (Heldman and Vincent, unpublished observa- tions). Many other efforts to find specialized synaptic structures in molluscs have also been unsuccessful (Gerschenfeld, 1963; Rosenbluth, 1963; Cobb and Mullins, 1973; Pentreath and Berry, 1975a, Pentreath, 1976). In fact, examples purporting to show some membrane specialization were actually the result of overlap of vesicles and axon membrane that could be resolved into unspecialized membrane and vesicles by tilting the section (Cobb and Pentreath, 1977). As previously noted (Pentreath and Berry, 1975a) the probability of finding specialized synaptic struc- tures would be low without consecutive serial sections and the fixations used in

Ultrastructure of photoreceptors labelled with horseradish peroxidase 193
many of these studies may not have preserved specialized structures. This last point is critical considering the difficulties of adequately fixing many marine invertebrate tissues. Complete reconstruction of these terminal processes may be necessary in order to describe the morphology of these synapses and the nature of the synaptic interactions between photoreceptors.
Acknowledgements
We thank Drs William J. Adelman and Alan Hodge for their review of the manuscript. This work was supported in part by National Research Service Award 1 F32 N S 0 5 0 2 1 - 1 2 to Dr Terry J. Crow.
References
ALKON, D.L. (1973a) Neural organization of a molluscan visual system. Journal of General Physiology 61, 444--61.
ALKON, D. L. (1973b) Intersensory interactions in Hermissenda. Journal of General Physiology 62, 185-202 .
ALKON, D. L. (1974) Associative training of Hermissenda. Journal of General Physiology 64, 7 0 - 8 4 .
ALKON, D. L. (1975) Neural correlates of associative training in Hermissenda. Journal of General Physiology 65, 4 6 - 5 6 .
ALKON, D. L. (1976) The economy of photoreceptor function in a primitive nervous system. In Neural Principles in Vision (edited by ZETTLER, F. and WEILER, R.) pp. 410--26. Berlin, Heidelberg, New York: Springer-Verlag.
ALKON, D.L. and FUORTES, M. G. F. (1972) Responses of photoreceptors in Hermissenda. Journal of General Physiology 60, 6 3 1 - 4 9 .
ALKON, D. L. and GROSSMAN, Y. (1977) Prolonged inhibition and excitation in the Herrnis- senda eye. Neuroscience Abstracts 3,171.
ALKON, D. L., AKAIKE, T. and HARRIGAN, J. (1978) Interaction of chemosensory, visual, and statocyst pathways in Hermissenda crassicornis. Journal of General Pbysiology 91, 177 -94 .
BAILEY, C.H., THOMPSON, E. B., CASTELLUCCI, V. F. and KANDEL, E. R. (1976) Fine structure of synapses of identified sensory cells which mediate the gill-withdrawal reflex in Aplysia cali]brnica. Neuroscience Abstracts 3, 314.
BERRY, M. S., COTTRELL, G. A. and PENTREATH, V. W. (1975) Nerve-terminal specializations in the central nervous system of Planorbis corneus. Journal of Physiology (London) 251, 3-4P.
BERRY, M. S. and P E N T R E A T H , V. W. (1976) Criteria for distinguishing between monosynaptic and polysynaptic transmission. Brain Research 105, 1 - 2 0 .
BLOOM, F. E. and A G H A J A N I A N , G. H. (1966)Cytochemis t ry of synapses: A selective staining method for eleetronmicroscopy. Science 154, 1575-7 .
CHRISTENSEN, B.N. (1973) Procion brown: an intracellular dye for light and electronmicro- scopy. Science 182, 1255-6 .
COBB, J. L. S and MULLINS, P .A. (1973) Synaptic structure in the visceral ganglion of the lamellibranch mollusc, Spisula solida. Zeitscbrift fiir Zellforscbung und mikroskopiscbe Anatomie 138, 75--83.
COBB, J. L. S. and PENTREATH, V.W. (1977) Anatomical studies of simple invertebrate synapses utilizing stage rotation electron microscopy and densitometry. Tissue and Cell 9, 125--35.

194 C R O W , H E L D M A N , H A C O P I A N , E N O S and A L K O N
COGGESHALL, R. E. (1967) A light and electron microscope study of the abdominal ganglion of Aplysia californica. Journal of Neurophysiology 30, 1263-87 .
CROW, T .J . and ALKON, D.L . (1978a) Retention of an associative behavioral change in Hermissenda. Science 201, 1239-41 .
CROW, T. J. and ALKON, D. L. (1978b) Neural correlates of an associative behavioral change in Hermissenda crassicornis. Society for Neuroscience Abstracts 4, 191.
EAKIN, R.M., WESTFALL, J. A and DENNIS, M.J . (1967) Fine structure of the eye of a nudibranch mollusc Hermissenda crassieornis. Journal of Cell Science 2, 349 -58 .
GERSCHENFELD, H.M. (1963) Observations on the ultrasturcture of synapses in some pulmo- nate molluscs. Zeitschrift fiir Zellforschung und mikroskopische Anatomic 6 0 , 2 5 8 - 7 5 .
GILLETTE, R. and POMERANZ, B. (1973) Neuron geometry and circuitry via the electron microscope: intracellular staining with osmiophilic polymer. Science 182, 1256-8 .
GRAHAM, R.C. and KARNOVSKY, M.J. (1966) The early stages of absorption of injected horseradish peroxidase in the proximal tubules of mouse kidney: ul trastmctural cytochemistry by a new technique. Journal of Histochemistry and Cytochemistry 14, 291-302 .
G RAY, E. G. (1971) The fine structural characterization of different types of synapse. Progress in Brain Research 34, 149 -60 .
MULLER, K.J . and McMAHAN, U. J. (1976) The shapes of sensory and motor neurons and the distribution of their synapses in ganglia of the leech: a study using intracellular injection of horseradish peroxidase. Proceedings of the Royal Society of London B194, 4 8 1 - 9 9 .
PALAY, S. L. and CHAN-PALAY, V. (1975) A guide to the synaptic analysis of the neuropil. Cold Spring Harbor Symposia on Quantitative Biology 40, 1 - 1 6 .
PENTREATH, V.W. (1976) Ultrastructure of the terminals of an identified 5-hydroxy- tryptamine-containing neurone marked by intracellular injection of radioactive 5-hydroxy- tryptamine. Journal of Neurocytology 5, 4 3 - 5 1 .
PENTREATH, V. W. and BERRY, M. S. (1975a) Ultrastructure of the terminals of an identified dopamine-containing neurone marked by intracellular injection of radioactive dopamine. Journal of Neurocytology 4, 249 -60 .
PENTREATH, V. W., BERRY, M. S. and COBB, J. L. S. (1975b) Nerve ending specializations in the central ganglia of Planorbis corneus. Cell and Tissue Research 163, 99-110.
PENTREATH, V. W. and COTTRELL, G. A. (1974) Anatomy of an identified serotonin neurone studied by means of injection of tri t iated ' transmitter ' . Nature 250, 655-8.
PETERS, A., PALAY, S. L. and WEBSTER, H. de F. (1976~ The Fine Structure of the Nervous System. Philadelphia: W. B. Saunders.
PITMAN, R.M., TWEEDLE, C.D. and COHEN, M.J. (1972). Branching of central neurons: intracellular cobalt injection for light and electron microscopy. Science 1 7 6 , 4 1 2 - 4 .
ROSENBLUTH, J. (1963) The visceral ganglion of Aplysia californica. Zeitschrift fiir Zell- forschung und mikroskopische Anatomic 60, 213 -36 .
STENSAAS, L.-J., STENSAAS, S.S. and TRUJILLO-CENOZ, O. ( 1 9 6 9 ) S o m e morphological aspects of the visual system of Hermissenda crassicornis (mollusca: Nudibranchia). Journal of Ultrastructure Research 27, 510- 32.
STRAUS, W. (1964a). Factors affecting the cytochemical reaction of peroxidase with benzidine and the stability of the blue reaction product. Journal of Histochemistry and Cytochemistry 1 2 , 4 6 2 - 9 .
STRAUS, W. (1964b) Factors affecting the state of injected horseradish peroxidase in animal tissues and procedures for the study of phagosomes and phago-lysosomes. Journal of Histo- chemistry and Cytocbemistry 12, 4 7 0 - 8 0 .
STRETTON, A. O. W. and KRAVITZ, E .A. (1968) Neuronal geometry: determination with a technique of intracellular dye injection. Science 162, 132 -4 .
THOMPSON, E.B., BAILEY, C. H., CASTELLUCCI, V. F. and KANDEL, E. R. (1976a) Two different and compatible intracellular labels: A preliminary structural study of identified

Ul t r a s t ruc tu re o f p h o t o r e c e p t o r s label led wi th horserad ish pe rox idase 195
sensory and motor neurons which mediate the gill-withdrawal reflex in Aplysia californica. Neuroscien ce Abstracts 2, 337.
THOMPSON, E. B., SCHWARTZ, J. H. and KANDEL, E. R. (1976b) A radioautographic analysis in the light and electron microscope of identified Aplysia neurons and their processes after intrasomatic injection of L-{ all} fucose. Brain Research 112, 251--81.
Copyright © 2022 FDOKUMEN










![Synthesis of labelled [15N3]-6-bromopurine, a useful precursor of 15N-labelled O6-alkylguanines, to be used as internal standards for quantitative GC-MS analyses](https://static.fdokumen.com/doc/165x107/63327b5cf008040551047e85/synthesis-of-labelled-15n3-6-bromopurine-a-useful-precursor-of-15n-labelled-o6-alkylguanines.jpg)









