
Morphological relationships of choleragenoid horseradish peroxidase-labeled spinal primary afferents...
16
THE JOURNAL OF COMPARATIVE NEUROLOGY 347~171-186 (1994) Morphological Relationships of Choleragenoid Horseradish Peroxidase-Labeled Spinal Primary Afferents With Myenteric Ganglia and Mucosal Associated Lymphoid Tissue in the Cat Esophagogastric Junction NADINE CLERC AND CHRISTOPHE MAZZIA Laboratoire de Neurobiologie, Interoception, CNRS, 13402 Marseille Cedex 20, France ABSTRACT The goal of the present study was to gain insight into the environmental factors influencing the activity of primary spinal afferent fibers in the different layers of the esophagogastric junction of the cat and, thus, to analyze the relationships of these afferents with various cellular components. Spinal primary afferent fibers were selectively labeled by anterogradely trans- ported choleragenoid horseradish peroxidase conjugate (B-HRP).B-HRP was injected into the thoracic dorsal root ganglion at the T8-T 13 levels. 6-Hydroxydopamine-induced sympathec- tomy was performed prior to B-HRP injection in order to prevent otherwise unavoidable labeling of sympathetic fibers in the gut wall. Numerous labeled fibers ran between, around, and within the myenteric ganglia. Others crossed the muscle layers directly and entered the mucosa, where some ran near granulocytes and around or through solitary lymphoid follicles. Labeled fibers were observed in the squamous esophageal epithelium but not in the fundic glandular epithelium. The fibers in the myenteric area are probably connected to the muscular tension receptors that have been detected by electrophysiologic techniques. This assumption is based on the observation that only a few fibers appear to terminate in muscle layers and on the fact that the myenteric area is very narrow and subject to powerful forces. Fibers in the myenteric ganglia could be involved in local efferent functions. Fibers in the mucosa could act as nociceptors and might be involved in local immunological responses. o 1994 Wiley-Liss, Inc. Key words: visceroceptors, sensory-efferent function, splanchnic nerves, digestive tract Various studies have demonstrated that dorsal root ganglion (DRG) neurons innervate the lower esophageal sphincter (LES). In the cat, Hazarika et al. (1964) studied elimination of the pupillodilatation reflex evoked by squeez- ing the LES after selective cutting of dorsal roots and found that these afferent neurons enter the spinal cord through dorsal roots from the T3-Tl3 levels. In the same species, retrograde transport studies with horseradish peroxidase (HRP) injected into the LES revealed that the cell bodies of these afferent neurons are distributed in all the thoracic DRG and the first lumbar DRG (Clerc, 1983). Data concerning the distribution of spinal primary affer- ent fibers within the esophageal wall and other parts of the gut are scarce. Most information has been obtained indi- rectly from the gastrointestinal level by electrophysiological recordings. These studies suggested that receptive fields for spinal primary afferents are chiefly in the serosa and the mesentery (see Janig and Morrison, 1986), but mechanore- ceptors were purported to exist within the gut wall (Clerc and Mei, 1983a; Sengupta et al., 1990). In the LES, the functional properties of visceroceptors connected to spinal primary afferents have been character- ized in the cat by using electrophysiological recordings and mechanical stimuli. According to their activation and dis- charge modalities, two types of mechanosensitive viscerocep- tors were characterized, i.e., serosal mechanoreceptors and the so-called muscular tension receptors (Clerc and Mei, Accepted March 15, 1994. Address reprint requests to Nadine Clerc, Laboratoire de Neurobiologie, Interoception, CNRS, 31 Chemin Joseph Aiguier, 13402 Marseille Cedex 20, France. o 1994 WILEY-LISS, INC.
-
Upload
univ-avignon -
Category
Documents
-
view
2 -
download
0
Transcript of Morphological relationships of choleragenoid horseradish peroxidase-labeled spinal primary afferents...

THE JOURNAL OF COMPARATIVE NEUROLOGY 347~171-186 (1994)
Morphological Relationships of Choleragenoid Horseradish
Peroxidase-Labeled Spinal Primary Afferents With Myenteric Ganglia and Mucosal Associated Lymphoid Tissue in the Cat Esophagogastric Junction
NADINE CLERC AND CHRISTOPHE MAZZIA Laboratoire de Neurobiologie, Interoception, CNRS, 13402 Marseille Cedex 20, France
ABSTRACT The goal of the present study was to gain insight into the environmental factors influencing
the activity of primary spinal afferent fibers in the different layers of the esophagogastric junction of the cat and, thus, to analyze the relationships of these afferents with various cellular components. Spinal primary afferent fibers were selectively labeled by anterogradely trans- ported choleragenoid horseradish peroxidase conjugate (B-HRP). B-HRP was injected into the thoracic dorsal root ganglion at the T8-T 13 levels. 6-Hydroxydopamine-induced sympathec- tomy was performed prior to B-HRP injection in order to prevent otherwise unavoidable labeling of sympathetic fibers in the gut wall.
Numerous labeled fibers ran between, around, and within the myenteric ganglia. Others crossed the muscle layers directly and entered the mucosa, where some ran near granulocytes and around or through solitary lymphoid follicles. Labeled fibers were observed in the squamous esophageal epithelium but not in the fundic glandular epithelium.
The fibers in the myenteric area are probably connected to the muscular tension receptors that have been detected by electrophysiologic techniques. This assumption is based on the observation that only a few fibers appear to terminate in muscle layers and on the fact that the myenteric area is very narrow and subject to powerful forces. Fibers in the myenteric ganglia could be involved in local efferent functions. Fibers in the mucosa could act as nociceptors and might be involved in local immunological responses. o 1994 Wiley-Liss, Inc.
Key words: visceroceptors, sensory-efferent function, splanchnic nerves, digestive tract
Various studies have demonstrated that dorsal root ganglion (DRG) neurons innervate the lower esophageal sphincter (LES). In the cat, Hazarika et al. (1964) studied elimination of the pupillodilatation reflex evoked by squeez- ing the LES after selective cutting of dorsal roots and found that these afferent neurons enter the spinal cord through dorsal roots from the T3-Tl3 levels. In the same species, retrograde transport studies with horseradish peroxidase (HRP) injected into the LES revealed that the cell bodies of these afferent neurons are distributed in all the thoracic DRG and the first lumbar DRG (Clerc, 1983).
Data concerning the distribution of spinal primary affer- ent fibers within the esophageal wall and other parts of the gut are scarce. Most information has been obtained indi- rectly from the gastrointestinal level by electrophysiological recordings. These studies suggested that receptive fields for
spinal primary afferents are chiefly in the serosa and the mesentery (see Janig and Morrison, 1986), but mechanore- ceptors were purported to exist within the gut wall (Clerc and Mei, 1983a; Sengupta et al., 1990).
In the LES, the functional properties of visceroceptors connected to spinal primary afferents have been character- ized in the cat by using electrophysiological recordings and mechanical stimuli. According to their activation and dis- charge modalities, two types of mechanosensitive viscerocep- tors were characterized, i.e., serosal mechanoreceptors and the so-called muscular tension receptors (Clerc and Mei,
Accepted March 15, 1994. Address reprint requests to Nadine Clerc, Laboratoire de Neurobiologie,
Interoception, CNRS, 31 Chemin Joseph Aiguier, 13402 Marseille Cedex 20, France.
o 1994 WILEY-LISS, INC.

172 N. CLERC AND C. MAZZIA
1983a). They are functionally identical to their vagal coun- terparts (Clerc and Mei, 198310).
In the LES, as well as in the lower digestive tract, the spinal equivalent of vagal mucosal mechanoreceptors has not been found (Clerc and Mei, 1983a,b). It has thus been assumed that the mucosal visceroceptors of the esophagus, stomach, and small intestine are connected predominantly to vagal afferent neurons (Mei, 1983; Grundy and Scratcherd, 1989). In the dog esophagus, this assumption was first supported by retrograde tracing studies (Hudson and Cummings, 1985), but later experiments using the same technique showed that some mucosal visceroceptors were connected to spinal primary afferent neurons (Khurana and Petras, 1991).
However, the gastrointestinal mucosa is not devoid of spinal afferent innervation because 1) splanchnic glucorecep- tors have been demonstrated in the small intestine of cats by electrophysiological recordings (Perrin et al., 1981) and 2) spinal primary afferents innervating the gastrointestinal mucosa of rats have been shown to mediate a viscerocircula- tory reflex (Rozsa et al., 1988). In addition, in the stomach of the rat, mouse, and guinea pig, immunoreactivity of dense nervous fibers to calcitonin gene-related peptide (CGRP) and substance P was observed to disappear after capsaicin treatment as well as after splanchnic section (Green and Dockray, 1988). However, similar immunohisto- chemical experiments indicate that, in the cat esophagus, substance P-immunoreactive fibers do not originate from DRG neurons (Hayashi et al., 1982a).
Selective visualization of afferent innervation following sympathetic pathways, i.e., arising from thoracolumbar DRG, has never been performed in the digestive tract. Conversely, visceral afferents that run along the parasym- pathetic pathways have been visualized using antero- gradely transported tracers in the esophagus and the LES (Clerc and Condamin, 1987) and in the gastroduodenal area (Holst and Powley, 1992). The fourfold purpose of this study was 1) to label spinal primary afferents selectively within the different layers of the wall of the cat esophagogas- tric junction, i.e., of the LES and the first millimeters of the fundus; 2) to reconcile these anatomical findings with electrophysiological data; 3) to explain the functional role of spinal primary afferents based on their microenvironment, and 4) to compare the distribution of spinal primary afferents with that of vagal primary afferents. Cholerage- noid horseradish peroxidase conjugate (B-HRP) was used as the tracer, because it had been previously shown to be anterogradely transported (Robertson and Grant, 1985). However, injection of B-HRP into thoracic DRG at the T8-Tl3 levels leads to labeling of numerous cell bodies in paravertebral ganglia. Because neurons of thoracic paraver- tebral ganglia innervate the esophagus (Hudson and Cum- mings, 1985), 6-hydroxydopamine (6-OHDA) treatment was performed prior to B-HRP injections in order to avoid labeling of sympathetic fibers in the wall of the esophagogas- tric junction.
MATERIALS AND METHODS Experiments were performed on 12 adult cats weighing
2.5-5.0 kg. After preanesthesia with ketamine hydrochlo- ride (20 mgikg i.m.), halothane (1-3% with 97-9976 air) was administered throughout the dissection and injection procedures. Upon awakening, animals systematically re- ceived antiinflammatory, analgesic, and antibiotic treat-
ment by i.m. injections of tolfenamic acid (12 mg/kg, Ketoquinol) and flumequine (30 mgikg, Sanofi).
B-HRP anterograde tracing B-HRP conjugate was prepared according to the method
of McIlhinney et al. (1988), with choleragenoid and HRP type VI obtained from Sigma (St. Louis, MO). The final volume of the resulting solution was 800 p1. B-HRP anterograde tracing was performed on nine cats. One to four microliters of the B-HRP solution was injected through a glass micropipette into the left thoracic DRG from T8 to T13.
After postoperative survival times of 72 hours in eight cases and 96 hours in one case, animals were reanesthetized and perfused transcardially . The perfusion was initiated by 1 liter of saline followed by 3 liters of a fixative consisting of 4% paraformaldehyde (PF; see Clerc and Condamin, 1987) in phosphate buffer (PB) 0.1 M, pH 7.4, in the presence or absence of 0.5% glutaraldehyde (GL). Omitting GL pre- serves full immunoreactivity to tyrosine hydroxylase (TH; see below). Fixation was terminated after 30 minutes by flushing the vasculature with 1 liter of 0.1 M PB containing 15% sucrose at room temperature.
The left DRG from the T8-Tl3 levels, the corresponding spinal segments, and the esophagogastric junction were removed and stored overnight in 0.1 M PB containing 15% sucrose. Samples were cut into 40 pm serial sections on a cryostat. Spinal segments were cut transversely, while the esophagogastric junction was cut either transversely or longitudinally. All material was processed according to the tetramethylbenzydine (TMB) method (Mesulam, 19761, modified as previously described (Clerc and Condamin, 1987), and counterstained with neutral red.
6-OHDA sympathectomy Eight of the nine animals that underwent B-HRP injec-
tion were pretreated with 6-OHDA hydrochloride (Sigma). Two 6-OHDA treatment protocols adapted from Tranzer and Thoenen (1968) and Kostrzewa and Jacobowitz (1974) were tested (Table 1). The first protocol consisted of four injections of 17.5 mg/kg 6-OHDA i.p. over a period of 48 hours at 12 hour intervals. B-HRP was injected 17-18 days after the first day of treatment (four animals). The second protocol consisted of two injections of 17.5 mg/kg i.p. at 12 hour intervals on days 1,2,18, and 23. B-HRP was injected 26 days after the first day of treatment (four animals). As a control, each protocol was applied to one animal that was not subjected to B-HRP anterograde tracing.
TH immunohistochemistry TH immunohistochemistry was performed to ascertain
the completeness of 6-OHDA sympathectomy in the eight animals subjected to both 6-OHDA treatment and B-HRP anterograde tracing (group A, Table l ) , in the two animals subjected only to 6-OHDA treatment (group B; Table l), and in one control animal (group C; Table 1). In groups B and C, fixation was achieved with 4% PF, as indicated above, except that the esophagogastric junction was post- fixed for 8-24 hours. After overnight immersion in PB containing 15% sucrose, samples were cut into 20 or 40 pm serial transverse sections on a cryostat and stored at -20°C.
For each TH-immunohistochemistry experiment, sec- tions from the esophagogastric junction of each animal in

SPINAL PRIMARY AFFERENTS IN THE GUT 173
TABLE 1. Experimental Procedures
Transverse fTI or CT-HRP ~~ ~ - , .~
longitudinal anterograde TH Experiment (L) sections tracing Fixative solution 6-OHDA treatment immunohistochemistry
~
1 T + 4% PF + 0.5% GL n 0 Group A
2 3 4 5
9 Group B
10 11
12 Group C
T T T L L T
+ + +
4% PF + 0 5% GL 4% PF 4% PF 4% P F 4 % P F + 0 5 X G L 4% PF + 0 5% GL 4% PF 4% PF
4 4 P F + post-fix 4% PF + post-fix 1
46 PF + post-fix
Protocol 1 . 4 i.p. injections2 over a period of 48 hours a t 12 hour intervals; survival time 20-23 days
Protocol 2: 2 i.p. injections2 at 12 hour intervals on days 1,2,18, and 23 of treatment; survival time. 29 days
Protocol 1 Protocol 2
0
+ +
+ +
+ ‘Post fixation. 2Injection dose = 17 5 mgikg.
group A were processed simultaneously with thawed esopha- gogastric junction sections from animals in groups B and C. Animals in groups Aand B were matched with regard to the 6-OHDA protocol used. This experimental design allowed us to verify that treated animals were immunonegative regardless of the duration of fixation and that only the animal that did not undergo sympathectomy was immu- nopositive. These verifications were necessary because of the short fixation time required for the TMB method and as a safeguard against false immunonegative results.
Sections from group A were treated to inhibit peroxidase activity before TH immunohistochemistry by the method of Straus (1976) before rehydration and rinsing in phosphate- buffered saline (PBS), 0.1 M, pH 7.4. Esophagogastric junction sections from the three groups were immersed for 30 minutes at room temperature in PBS containing lysine 0.1 M and 0.1% sodium azide. After rinsing in PBS, they were submitted to a proteolytic treatment with trypsin in PBS (see Finley and Petrusz, 1982).
After thorough rinsing in PBS, sections were incubated as follows: 1) 6 hours at room temperature in PBS contain- ing 0.1% sodium azide, 1% bovine serum albumin (BSA), 5% normal swine serum (NSS), and 1% Triton X-100; 2) 60 hours at 4°C in the first antiserum raised in rabbits (TH, Institute Jacques Boy), diluted 1:20,000 in the same solu- tion; 3) six times for 10 minutes each in PBS containing 0.1% sodium azide, 5% NSS, and 0.3% TritonX-100 in PBS; 4) 4 hours at room temperature in swine antirabbit IgG (Dako S.A.) diluted 1:25 in the previous solution. After rinsing in PBS, the double-bridge peroxidase-antiperoxi- dase procedure (Vacca et al., 1980) was applied using rabbit peroxidase-antiperoxidase (Dako S.A.) diluted 1:50 in PBS with 0.3% Triton X-100 for 2-3 hours at room temperature. Finally, after a rinse in Tris buffer, 0.05 M, pH 7.6, sections were incubated 10 minutes at room temperature in diamino- benzidine tetrahydrochloride (0.025% in Tris buffer), be- fore adding 0.06% H202.
RESULTS Injection sites
The reaction product displayed a density varying propor- tionally to the amount of B-HRP injected. B-HRP spread throughout the extracellular space but appeared to accumu- late mainly on the cell membrane (Fig. 1). In all but a few
Fig. 1. B-HRP distribution in a section taken from the left dorsal root ganglion (DRG) at the T8 level after injection of B-HRP (4 ~ 1 ) . B-HRP was located either pericellularly (black arrows) or intracellu- larly (black arrowhead). One cell body was totally devoid of labeling (white arrows). Scale bar = 100 pm.
cases, intracellular labeling was weak or absent (Fig. 1). Some cell bodies exhibited neither intra- nor pericellular labeling (Fig. 1). These labeling patterns were observed on both large and small cell bodies. Numerous labeled fibers were seen within ganglia, dorsal roots, and spinal nerves.
Central projections At the spinal cord level (Fig. 2), labeling was homolateral
to the injection site. In the white matter, darkly labeled fibers were observed in Lissauer’s tract and lighter labeling in the dorsal columns. In the grey matter, both the lateral and the medial collateral pathways of Lissauer’s tract were densely labeled. The dorsal horn displayed homogeneous labeling, including all the laminae of Rexed. Dense labeling was also observed on lamina VII mediolateral to the central canal. The ventral horn exhibited a few granules of HRP reaction product.

174 N. CLERC AND C. MAZZIA
Fig. 2. Partial transverse section taken from the spinal cord at the T8 level after B-HRP injection into the left DRG from the T8-Tl3 levels. B-HRP-labeled terminals and axons are homolateral to the injection site, in the dorsal columns (dc), in the Lissauer tract (Lt) and
its lateral and medial collateral pathways (Icp and mcp), in the whole dorsal horn (dh), and in the ventral horn (vh), in particular near the central canal (cc). Scale bar = 50 pm.

SPINAL PRIMARY AFFERENTS IN THE GUT
Labeling in sympathetic ganglia In the paravertebral ganglia from the T8-Tl3 levels,
numerous densely labeled cell bodies were observed, usu- ally over the whole surface of the sections. Efferent nervous fibers of the sympathetic trunk and afferent nervous fibers of the bundle stemming from the white rami communi- cantes were frequently labeled.
In the prevertebral celiac ganglia, cell bodies were never labeled. However, numerous labeled nervous fibers were seen homolaterally to the injection sites. Most of these fibers were packed in bundles reaching the nerve rami connected to the visceral effectors.
6-OHDA sympathectomy The efficacy of the two protocols used to destroy sympa-
thetic fibers was assessed using TH immunohistochemistry (see Materials and Methods). After protocol 1, in animals from groups A and B, TH-immunoreactive fibers were still observed crossing the longitudinal and circular muscle and in the myenteric area. The pattern of immunoreactivity was similar to that of the control animal, except that diffuse immunoreactive product concentrated in the medial part of the myenteric ganglia instead of varicose immunoreactive fibers. After protocol 2, no TH immunoreactivity was observed in the wall of the esophagogastric junction.
Labeling in the wall of the esophagogastric junction
B-HRP labeled fibers were observed on serial sections of the esophagogastric junction from the control animal and all sympathectomized animals, regardless of the 6-OHDA injection protocol used. Labeled fibers were seen in the serosa, the longitudinal and circular muscle layers, the myenteric area, and the mucosa. They frequently ran alongside vascular beds (in all layers; Figs. 3B,C, 4B, 6C) or surrounded blood vessels (in the mucosa; Fig. 3A,D). The distribution pattern in each of these layers was as follows.
In the serosa, as well as in the longitudinal muscle layer, labeled fibers were generally grouped in bundles, and there was no evidence of terminal endings. A few B-HRP-labeled fibers penetrated into the longitudinal muscle and ran parallel to muscle fibers, as seen in longitudinal sections (Fig. 4E). Most fibers ran between bundles of longitudinal muscle fibers and made their way toward the myenteric area.
In the myenteric area, two patterns of labeling were observed. In the first pattern, the labeled fibers did not show a close relationship with the myenteric ganglia (Fig. 5A). Observations made on both transverse and longitudi- nal sections indicate that these fibers run parallel to the circular muscle (Fig. 5A) or alongside the longitudinal axis of the esophagus. In the second pattern, the fibers either passed through (Fig. 5B,C) or wrapped around (Fig. 5B) the myenteric ganglia. Within the ganglia, the labeled fibers had a course either sinuous or straight. These intragangli- onic fibers sometimes appeared to ramify and frequently displayed a varicose appearance (Fig. 5C,D). Some fibers may terminate within ganglia. The high density of the B-HRP-labeled fibers in the myenteric area, compared to their low density in the circular muscle, suggests that many end there.
In the circular muscle, the pathway of B-HRP-labeled fibers was usually straight. On transverse sections, fibers ran parallel (Fig. 4A) or, more often, perpendicular to the
175
muscle fibers (Fig. 4B,C). On longitudinal sections, they sometimes crossed the muscle layer obliquely. In a few cases, however, the pathway was extremely winding (Fig. 4D).
Most of the fibers seen in the circular muscle appeared to enter the mucosa. A large number of them, either isolated or grouped in bundles, were found in the submucosa. Their pathway was always sinuous.
On longitudinal sections, a small subset of B-HRP- labeled fibers located in the mucosa was found to run along the muscle fibers of the muscularis mucosae, between or within bundles of muscle fibers (Fig. 6A) or along the borders of the muscularis mucosae. A few fibers seen in that layer were winding (Fig. 6B). Most of the numerous B-HRP- labeled fibers found in the submucosa probably reached the outer layers of the mucosa, i.e., the lamina propria and the epithelium, through bundles of longitudinal muscle fibers of the muscularis mucosae, as shown in Figure 6C.
In the lamina propria, numerous B-HRP-labeled fibers were seen running through clusters of granulocytes (Fig. 7A,B), in close proximity to solitary lymphoid follicles (Fig. 8A,B) and, in one case, reaching the germinal center of a solitary lymphoid follicle (Fig. 8C). Granulocytes and lym- phocytes are components of the gut-associated lymphoid tissue (GALT). Granulocytes were easily visualized under our experimental conditions, because they contain endog- enous HRP. In this regard, the only peroxidase-containing granulocytes in the cat gut are neutrophils (see Arndt et al., 1992). Granulocytes and well developed solitary lymphoid follicles with germinal centers were consistently observed on the gastric side of the esophagogastric junction.
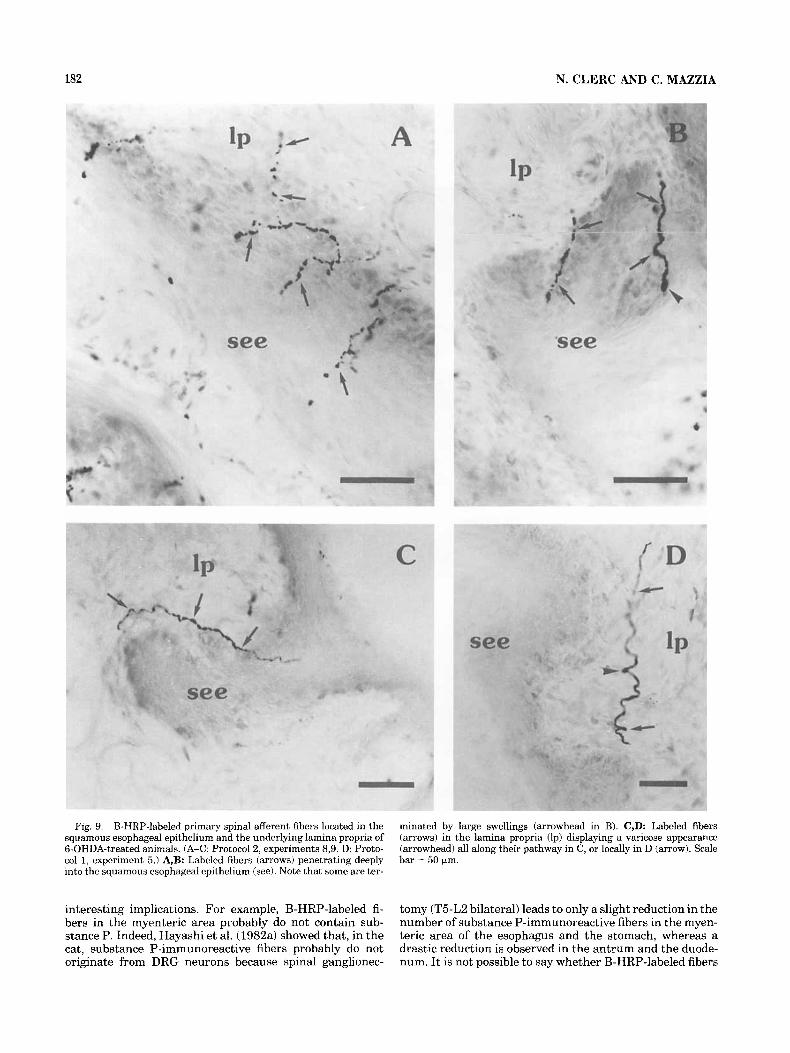
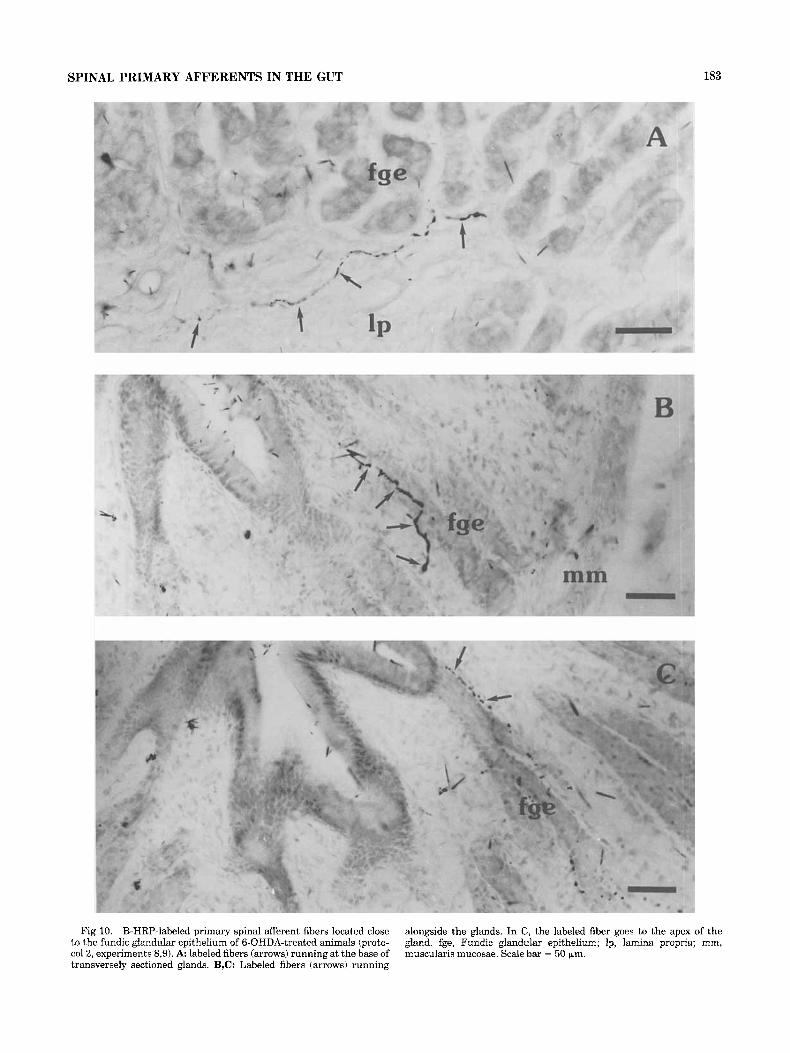
Numerous B-HRP-labeled fibers penetrated into the squamous esophageal epithelium (Fig. 9) and could be observed close to the lumen (Fig. 9A,B). Although no fibers entered the fundic glandular epithelium, a few were seen at the base of (Fig. 10A) or alongside the glands (Fig. 10B,C). In the muscularis mucosae and the squamous epithelium, labeled fibers sometimes presented large swellings that seemed to be endings (Figs. 6A, 8A2,9B).
DISCUSSION Selective labeling is the crucial point in these experi-
ments. To ensure this condition, we performed sympathec- tomy using two distinct protocols because injection of B-HRP into the DRG also results in labeling of sympathetic efferent neurons that may innervate the LES (Hudson and Cummings, 1985). In this regard, labeling of sympathetic cell bodies has been observed in the stellate ganglion of the guinea pig (Quigget al., 1988) and in the thoracic paraverte- bra1 ganglia of the cat (Glerc, 1989) after injection of wheat germ agglutinin conjugated to HRP (WGA-HRP) into DRG. According to Quigg et al. (1988), this labeling would result from anterograde transsynaptic transport of the tracer from spinal primary afferents to the postganglionic sympa- thetic neurons. However, ultrastructural evidence of extra- cellular tracer in the dorsal horn (Weinberg et al., 1990) and in paravertebral ganglia (Clerc and Baude, in preparation) after injection of WGA-HRP into DRG suggest transcellular transport. WGA-HRP and B-HRP labeling of cell bodies in paravertebral ganglia can be assumed to depend on the same underlying phenomenon.
Labeled spinal primary afferent fibers can be conveyed by both the small thoracic sympathetic nerve rami and the splanchnic nerves (Clerc, 1983). No data are available
176 N. CLERC AND C. MAZZIA
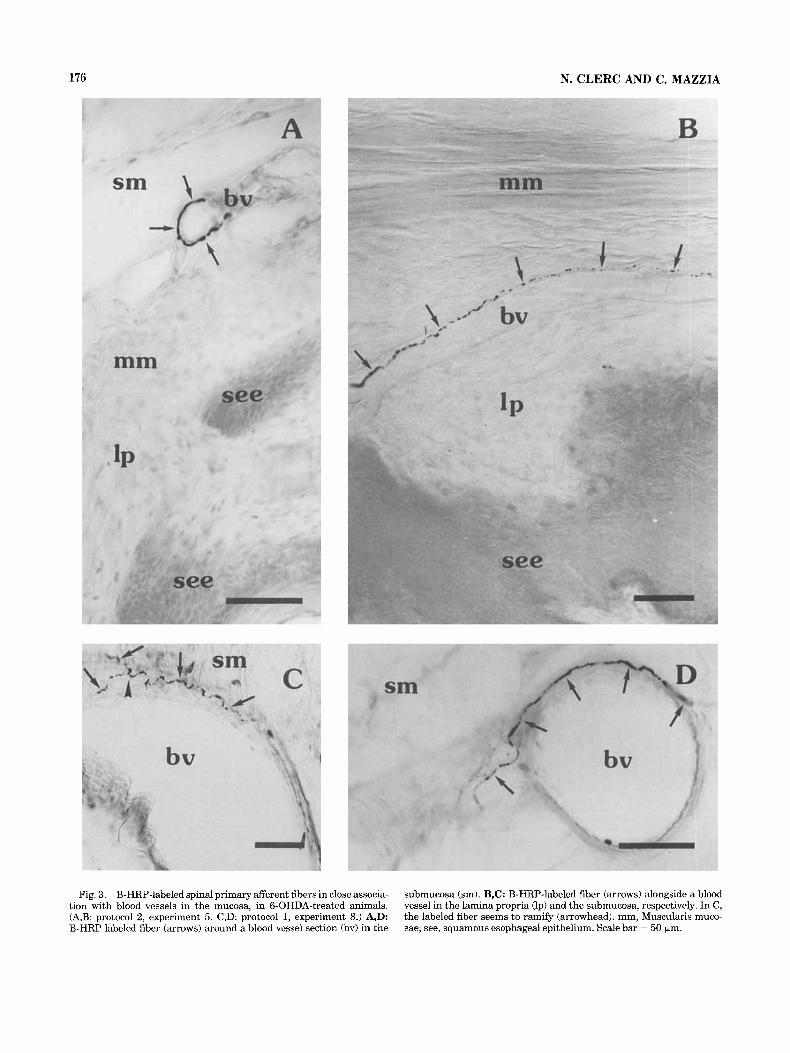
Fig, 3. B-HRP-labeled spinal primary afferent fibers in close associa- tion with blood vessels in the mucosa, in 6-OHDA-treated animals. (A,B: protocol 2, experiment 5. C,D: protocol 1, experiment 8.) A,D: B-HRP labeled fiber (arrows) around a blood vessel section (bv) in the
submucosa (sm). B,C: B-HRP-labeled fiber (arrows) alongside a blood vessel in the lamina propria (Ip) and the submucosa, respectively. In C, the labeled fiber seems to ramify (arrowhead). mm, Muscularis muco- sae; see, squamous esophageai epithelium. Scale bar = 50 bm.

SPINAL PRIMARY AFFERENTS IN THE GUT 177
Fig. 4. Transverse sections of the esophagogastric junction showing B-HRP-labeled spinal primary afferent fibers in the longitudinal and circular muscles of 6-OHDA-treated animals. (Protocol 2. A-D: Experi- ment 8. E: Experiment 9.) A: Labeled fibers parallel to the circular muscle fibers, between the longitudinal muscle (lm) and the circular muscle (cm, fine arrows), or within the circular muscle (thick arrows).
B,C: Labeled fiber crossing the circular muscle perpendicular to the muscle fibers; in B, labeled fibers (arrows) run alongside a blood vessel (bv). D: Labeled fiber (arrows) following a very sinuous pathway in the circular muscle. E: Laheled fiber (arrows) within the longitudinal muscle parallel to the muscle fibers. Scale bar = 50 pm.

178 N. CLERC AND C. MAZZIA
Fig. 5. Longitudinal sections of esophagogastric junction showing B-HRP-labeled spinal primary afferent fibers located in the myenteric area of 6-OHDA-treated animals. (A,C: Protocol 1, experiment 5. B,D: Protocol 2, experiment 9.) A: Labeled fiber showing a varicose appear- ance (arrowhead) in a part of the myenteric area that is devoid of
myenteric ganglia. B-D: Labeled fibers showing a close relationship with myenteric ganglia (mg). In B, one labeled fiber wraps around the ganglion (thin arrows). In B-D, labeled fibers are observed within the ganglion (thick arrows) and may display a varicose aspect (solid arrow- heads) or wrap around ace11 body (open arrowhead). Scale bar = 50 Fm.
SPINAL PRIMARY AFFERENTS IN THE GUT 179
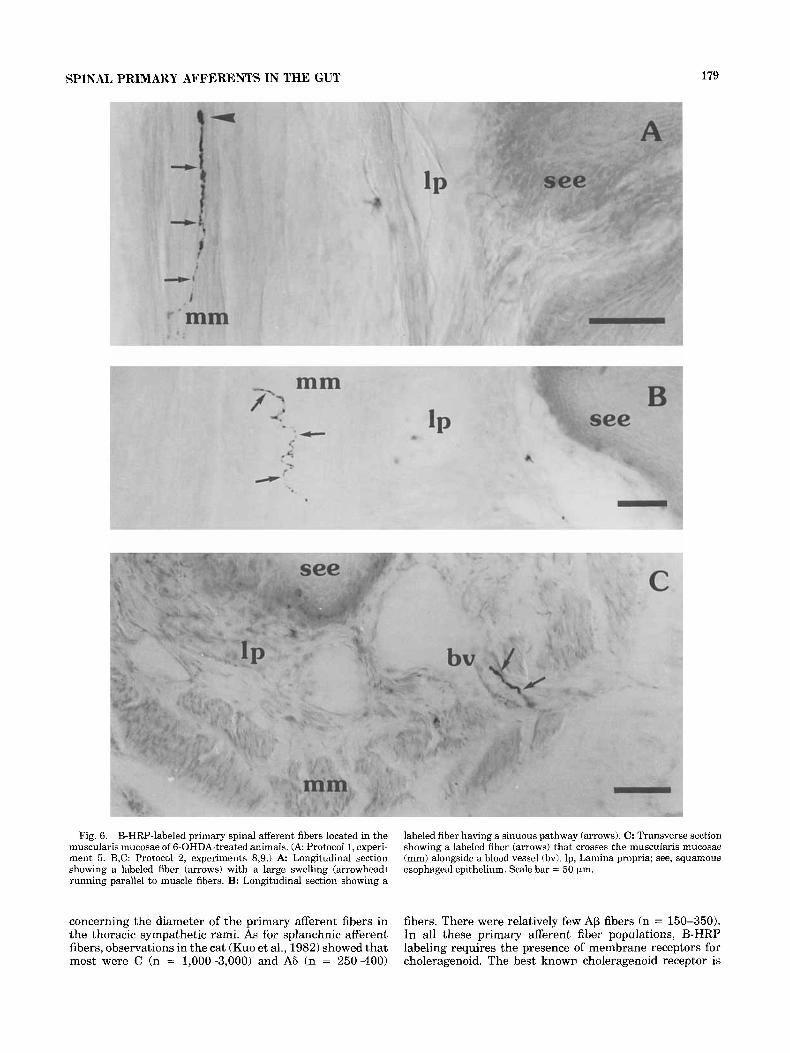
Fig. 6. B-HRP-labeled primary spinal afferent fibers located in the muscularis mucosae of 6-OHDA-treated animals. ( A Protocol 1, experi- ment 5. B,C: Protocol 2, experiments 8,9.) A Longitudinal section showing a labeled fiber (arrows) with a large swelling (arrowhead) running parallel to muscle fibers. B: Longitudinal section showing a
labeled fiber having a sinuous pathway (arrows). C: Transverse section showing a labeled fiber (arrows) that crosses the muscularis mucosae (mm) alongside a blood vessel (bv). lp, Lamina propria; see, squamous esophageal epithelium. Scale bar = 50 pin.
concerning the diameter of the primary afferent fibers in the thoracic sympathetic rami. As for splanchnic afferent fibers, observations in the cat (Kuo et al., 1982) showed that most were C (n = 1,000-3,000) and AS (n = 250-400)
fibers. There were relatively few AP fibers (n = 150-350). In all these primary afferent fiber populations, B-HRP labeling requires the presence of membrane receptors for choleragenoid. The best known choleragenoid receptor is
180 N. CLERC AND C. MAZZIA
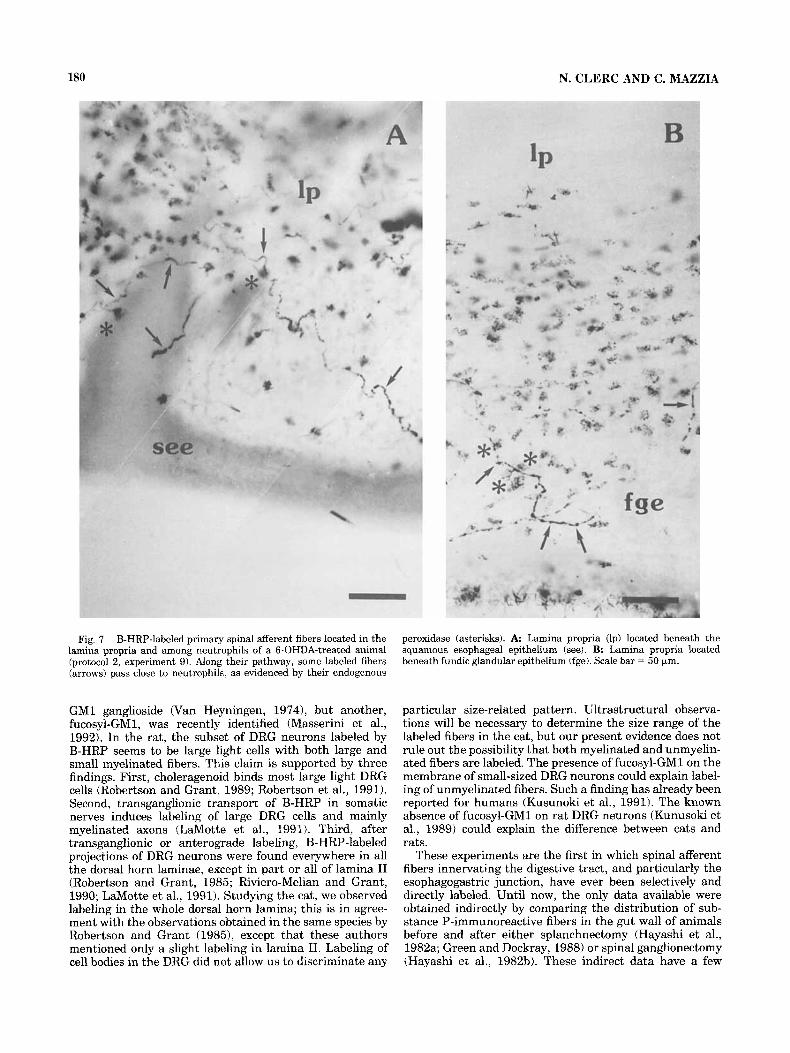
Fig. 7. B-HRP-labeled primary spinal afferent fibers located in the lamina propria and among neutrophils of a 6-OHDA-treated animal (protocol 2, experiment 9). Along their pathway, some labeled fibers (arrows) pass close to neutrophils, as evidenced by their endogenous
peroxidase (asterisks). A Lamina propria (Ip) located beneath the squamous esophageal epithelium (see). B. Lamina propria located beneath fundic glandular epithelium (fge). Scale bar = 50 (*m.
GM1 ganglioside (Van Heyningen, 19741, but another, fucosyl-GM1, was recently identified (Masserini et al., 1992). In the rat, the subset of DRG neurons labeled by B-HRP seems to be large light cells with both large and small myelinated fibers. This claim is supported by three findings. First, choleragenoid binds most large light DRG cells (Robertson and Grant, 1989; Robertson et al., 1991). Second, transganglionic transport of B-HRP in somatic nerves induces labeling of large DRG cells and mainly myelinated axons (LaMotte et al., 1991). Third, after transganglionic or anterograde labeling, B-HRP-labeled projections of DRG neurons were found everywhere in all the dorsal horn laminae, except in part or all of lamina I1 (Robertson and Grant, 1985; Riviero-Melian and Grant, 1990; LaMotte et al., 1991). Studying the cat, we observed labeling in the whole dorsal horn lamina; this is in agree- ment with the observations obtained in the same species by Robertson and Grant (1989, except that these authors mentioned only a slight labeling in lamina 11. Labeling of cell bodies in the DRG did not allow us to discriminate any
particular size-related pattern. Ultrastructural observa- tions will be necessary to determine the size range of the labeled fibers in the cat, but our present evidence does not rule out the possibility that both myelinated and unmyelin- ated fibers are labeled. The presence of fucosyl-GM1 on the membrane of small-sized DRG neurons could explain label- ing of unmyelinated fibers. Such a finding has already been reported for humans (Kusunoki et al., 1991). The known absence of fucosyl-GM1 on rat DRG neurons (Kunusoki et al., 1989) could explain the difference between cats and rats.
These experiments are the first in which spinal afferent fibers innervating the digestive tract, and particularly the esophagogastric junction, have ever been selectively and directly labeled. Until now, the only data available were obtained indirectly by comparing the distribution of sub- stance P-immunoreactive fibers in the gut wall of animals before and after either splanchnectomy (Hayashi et al., 1982a; Green and Dockray, 1988) or spinal ganglionectomy (Hayashi et al., 1982b). These indirect data have a few
SPINAL PRIMARY AFFERENTS IN THE GUT 181
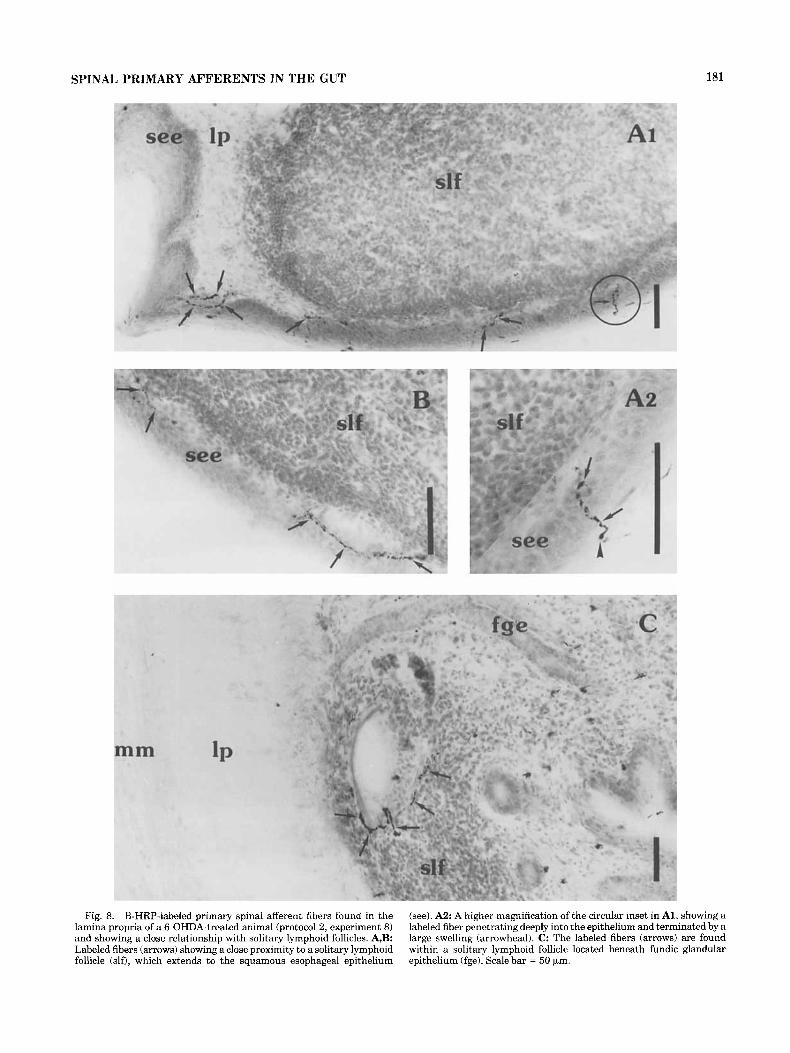
Fig. 8. B-HRP-labeled primary spinal afferent fibers found in the lamina propria of a 6-OHDA-treated animal (protocol 2, experiment 8) and showing a close relationship with solitary lymphoid follicles. A,B: Labeled fibers (arrows) showing a close proximity to a solitary lymphoid follicle (do, which extends to the squamous esophageal epithelium
(see). A 2 A higher magnification of the circular inset in Al, showing a labeled fiber penetrating deeply into the epithelium and terminated by a large swelling (arrowhead). C: The labeled fibers (arrows) are found within a solitary lymphoid follicle located beneath fundic glandular epithelium (fge). Scale bar = 50 pm.
182 N. CLERC AND C. MAZZIA
Fig. 9. B-HRP-labeled primary spinal afferent fibers located in the squamous esophageal epithelium and the underlying lamina propria of 6-OHDA-treated animals. (A-C: Protocol 2, experiments 8,9. D: Proto- col 1, experiment 5.) A,B: Labeled fibers (arrows) penetrating deeply into the squamous esophageal epithelium (see). Note that some are ter-
minated by large swellings (arrowhead in B). C,D: Labeled fibers (arrows) in the lamina propria (lp) displaying a varicose appearance (arrowhead) all along their pathway in C, or locally in D (arrow). Scale bar = 50 bm.
interesting implications. For example, B-HRP-labeled fi- bers in the myenteric area probably do not contain sub- stance P. Indeed, Hayashi et al. (1982a) showed that, in the cat, substance P-immunoreactive fibers probably do not originate from DRG neurons because spinal ganglionec-
tomy (T5-LZ bilateral) leads to only a slight reduction in the number of substance P-immunoreactive fibers in the myen- teric area of the esophagus and the stomach, whereas a drastic reduction is observed in the antrum and the duode- num. It is not possible to say whether B-HRP-labeled fibers
SPINAL PRIMARY AFFERENTS IN THE GUT 183
Fig 10. B-HRP-labeled primary spinal afferent fibers located close to the fundic glandular epithelium of 6-OHDA-treated animals (proto- col2, experiments 8,9). A: labeled fibers (arrows) running at the base of transversely sectioned glands. B,C: Labeled fibers (arrows) running
alongside the glands. In C, the labeled fiber goes to the apex of the gland. fge, Fundic glandular epithelium; Ip, lamina propria; mm, muscularis mucosae. Scale bar = 50 pm.

184 N. CLERC AND C. MAZZIA
innervating the mucosa are related to the subset of the substance P-immunoreactive fibers described in the submu- cosa of the cat esophagus and stomach (Hayashi et al., 1982a; Leander et al., 1982) because no information is available concerning the effect of spinal ganglionectomy on this subset. However, it is noteworthy that Leander et al. (1982) did not find substance P-immunoreactive fibers in the cat esophageal epithelium, where B-HRP-labeled fibers are numerous. Evidence reported by Rodrigo et al. (1985) that CGRP-containing extrinsic nerve fibers of the cat esophagus correspond mainly to vagal afferents make it seem unlikely that B-HRP-labeled fibers contain CGRP.
B-HRP-labeled fibers were observed in every layer of the esophagogastric junction. The densest concentration of B-HRP fibers was observed in the myenteric area and mucosa, including the squamous esophageal epithelium. Most fibers seemed only to cross the longitudinal muscle because they were found running between, rather than within, muscle bundles. In the circular muscle, most fibers ran perpendicular to the muscle fibers, but a few displayed a sinuous pathway. Perpendicular fibers probably go to the mucosa. To reach this layer, these fibers do not necessarily cross the myenteric area directly; some pass alongside the myenteric area either circularly or longitudinally. Except for a few displaying sinuous pathways in the circular muscle, B-HRP-labeled spinal primary afferent fibers pre- sented practically the same aspect and distribution as vagal counterparts selectively labeled with WGA-HRP (Clerc and Condamin, 1987). Both seemed to innervate chiefly the myenteric area and the mucosa.
The demonstration of spinal afferent innervation in the mucosa of the esophagogastric junction contradicts previ- ous retrograde tracing studies (Hudson and Cummings, 1985), immunohistochemical studies (Hayashi et al., 1982b), and electrophysiological studies (Clerc and Mei, 1983a). All these methods failed to detect mucosal receptors connected to spinal primary afferents and suggested that the mucosa does not receive any innervation from DRG neurons. Several explanations can be offered for these previously negative results. In retrograde tracing studies carried out on dogs (Hudson and Cummings, 19851, it is likely that methodological problems occurred because even injections in the muscle layers of the esophagus underestimated the number of labeled DRG cell bodies in comparison with data obtained in the same species by Khurana and Petras (1991). In immunohistochemical experiments, negative results sim- ply indicated that the esophageal mucosa is not innervated by DRG neurons expressing the substance P phenotype but do not rule out the possible involvement of subpopulations with other phenotypes. In electrophysiological studies, negative findings could be due to a lack of response from these visceroceptors to current search stimuli. Evidence has been found recently for unresponsive afferent fibers in the skin (Handwerker et al., 1991).
Some of the spinal primary afferent fibers labeled by B-HRP in the mucosa exhibit morphological relationships with epithelial cells of the squamous esophageal epithe- lium, into which they enter deeply, or other histological structures such as gastric glands, blood vessels, or the muscularis mucosae. Our findings also indicated that some spinal primary afferent fibers were closely associated with solitary lymphoid follicles and granulocytes located in the lamina propria. Until now, morphological relationships between primary afferent fibers and GALT cells had only been speculated to exist for mucosal mast cells, based on
immunohistochemical findings (see Stead et al., 1990). Data were not available concerning relationships with solitary lymphoid follicles located in the gut mucosa or Peyer’s patches. Innervation of secondary lymphoid tissue (ie., lymph nodes and spleen) by primary afferent fibers was previously described from a study involving the guinea pig spleen (Elfvin et al., 1992). However, in that report, sympathectomy was not performed, and the possibility of labeling of sympathetic fibers cannot be ruled out. Compari- son of the 6-OHDA-treated animals with the untreated animal suggests that chemical sympathectomy does not affect either the number of B-HRP-labeled fibers in the mucosa, especially in the lamina propria, or the size and number of solitary lymphoid follicles.
In assessing the physiological role of these spinal primary afferent fibers innervating the mucosa, most of which are electrophysiologically silent, it is necessary to take into account their close relationship with cellular or histological components of the mucosa as well as their location close to the border of the lumen or deeper in the mucosa. In this framework, the best overall hypothesis is the noceffector concept proposed by Kruger (1988), which involves both nociceptor and local effector functions. Concerning the local effector function, a large body of evidence now suggests that, when the peripheral endings of primary afferent neurons are adequately stimulated, they release their neu- ropeptidic content. Up until now, all such evidence has been obtained from capsaicin-sensitive primary afferent neurons innervating the viscera and the skin (see Holzer, 1988; Maggi and Meli, 1988).
The labeled spinal afferent fibers located near the lumen, i.e., within the squamous esophageal epithelium or along- side the gastric gland, can be activated by applying harmful stimuli. Indeed, spinal reflexes have been described in rats after application of fluid at 45°C on the gastrointestinal mucosa (Rozsa et al., 1988) or after application of a mild irritant consisting of 1 M NaCl on the gastric mucosa (Delbro, 1989). However, no such information is available concerning the mucosa at the level of the esophagogastric junction.
The terminal portions of autonomic nerve fibers being varicose and naked, neuropeptides could be released en passage (see Burnstock, 1986; Stead, 1992) from the spinal primary afferent fibers, which show close relationships with GALT cells. These neuropeptides might either modify immune functions by acting specifically on the membrane receptors of immunocompetent cells or modulate immuno- logical responses in the microenvironment of the mucosa (see Donnerer et al., 1990). Extensive evidence points to the presence of neuropeptide receptors on lymphoid cells in the mucosa (Mantyh et al., 1988; Gates et al., 1989; Reubi et al., 1992) and to functional modulations of immune responses by neuropeptides (Stanisz et al., 1986). The influence of primary afferent fibers on immune competent cells has been demonstrated for connective mast cells of the rat dura mater and tongue after antidromic stimulation of trigemi- nal afferent fibers (Dimitriadou et al., 1991). Interestingly, the electrical stimulation parameters used suggest that at least some of these afferent trigeminal fibers are large- caliber fibers.
The B-HRP-labeled fibers in the circular muscle or myenteric area could correspond to the muscular tension receptors previously characterized in electrophysiological studies (Clerc and Mei, 1983a). In the circular muscle, only the few fibers with sinuous pathways could end there and

SPINAL PRIMARY AFFERENTS IN THE GUT 185
thus would qualify as tension receptors. Conversely, in the myenteric area, the numerous B-HRP-labeled fibers that we observed could fulfill this function. This is logical because the myenteric area is subjected to strong deforma- tions during contraction and relaxation of both the circular and the longitudinal muscles. Intraganglionic fibers may also be exposed to movements able to stretch ganglionic cells. Such stretching is well known at the intestinal level, where myenteric ganglia are anchored to the muscle layers by interstitial cells (see Thuneberg, 1989). For the smooth- muscle part of the cat esophagus, including the LES, electrophysiological studies show that muscular tension receptors connected to vagal and spinal primary afferents have the same function (Clerc and Mei, 1983a,b) in a given location, i.e., detection of distension in the esophagus and detection of the degree of contraction in the LES (Clerc, unpublished results). Sensitivity to distension in the smooth- muscle part of the esophagus was further investigated in recent electrophysiological studies of the stimulus-response function to graded intraluminal distension. Three types of tensioreceptors were identified, namely 1) low-threshold mechanoreceptors connected to vagal afferent neurons (Sengupta et al., 1989), 2) encoding intensity mechanonoci- ceptors connected to DRG neurons (Sengupta et al., 19901, and 3) high-threshold mechanonociceptors connected to DRG neurons (Sengupta et al., 1990). These muscular tension receptors show differential sensitivities to bradyki- nin, suggesting that the vagal afferents are activated by longitudinal muscle contraction, whereas the spinal affer- ents are not (Sengupta et al., 1992). Similarly, in the LES, muscular tension receptors connected to vagal and spinal primary afferents may show differences in activation thresh- old as well as sensitivity to contraction. Light microscopy after anterograde tracing in the myenteric area of the cat esophagogastric junction did not allow clear differentiation of spinal and vagal primary afferent fibers: The extra- or intraganglionic location of vagal primary afferent fibers labeled by WGA-HRP (Clerc and Condamin, 1987) was similar to that of the B-HRP-labeled fibers described in the present study.
An ultrastructural study performed on the esophagus and LES of the rat (Neuhuber, 1987) indicated that vagal primary afferent fibers ended presynaptically to cells of the myenteric ganglia. This observation suggests that they might be involved in local effector functions. This is also a possible hypothesis concerning the spinal primary afferent fibers, but further ultrastructural study is needed.
ACKNOWLEDGMENTS The authors thank Dr. Jean Gonella for reading the
manuscript, Mrs. Madeleine Condamin for excellent assis- tance with histochemistry, Mr. Andre Boyer for assistance with surgery, Mr. Alain Yvinec for developing the photo- graphs, and Mrs. FranGoise Farnarier and Mr. Andy Corsini for reviewing the English.
LITERATURE CITED Arndt, H., P. Kubes, M.B. Grisham, E. Gonzalez, and D.N. Granger (1992)
Granulocyte turnover in the feline intestine. Inflammation 16.549-559. Burnstock, G. (1986) The changing face of autonomic transmission. Acta
Physiol. Scand. 126:67-91. Clerc, N. (1983) Afferent innervation of the lower oesophageal sphincter of
the cat. An HRP study. J. Autonomic Nerv. Syst. 9:623-636. Clerc, N. (1989) Labeling of cell bodies in sympathetic ganglia after
WGA-HRP injection in dorsal root ganglia of the cat Dig. Dis. Sci. 34:972.
Clerc, N., and M. Condamin (1987) Selective labeling of vagal sensory nerve fibers in the lower esophageal sphincter with anterogradely transported WGA-HRP. Brain Res. 424:216-224.
Clerc, N., and N. Mei (1983a) Thoracic esophageal mechanoreceptors connected with fibers following sympathetic pathways. Brain Res. Bull. tD:l-7.
Clerc, N., and N. Mei (1983b) Vagal mechanoreceptors located in the lower oesophageal sphincter of the cat. J. Physiol. 336:487-498.
Delbro, D. (1989) Spinovagal reflex modulation of gastric motility in response to mucosal nociceptive stimulation in the anaesthetized rat. Scand. J. Gastroenterol. 24:933-938.
Dimittiadou, V., M.G. Buzzi, M.A. Moskowitz, and T.C. Theoharides (1991) Trigeminal sensory fiber stimulation induces morphological changes reflecting secretion in rat dura mater mast cells. Neuroscience 44:97- 112.
Donnerer, J., A. Eglezos, and R.D. Helme (1990) Neuroendocrine and immune function in the capsaicin-treated rat: Evidence for afferent neural modulation in vivo. In S. Frier (ed): The Neuroendocrine Immune Network. Boca Raton, FL: CRC Press, Inc., pp. 69-83.
Elfvin, L.G., H. Aldskogius, and J. Johanson (1992) Splenic primary sensory afferents in the guinea pig demonstrated with anterogradely transported wheat-germ agglutinin conjugated to horseradish peroxidase. Cell Tissue Res. 269229-234.
Finley, J.C.W., and P. Petrusz (1982) The use of proteolytic enzymes for improved localization of tissue antigens with immunocytochemistry. In G.R. Bullock and P. Petrusz (eds): Techniques in Immunocytochemistry. Vol. I. New York: Academic Press, pp. 239-249.
Gates, T.S., R.P. Zimmerman, C.R. Mantyh, S.R. Vigna, and P.W. Mantyh (1989) Calcitonin gene-related peptide-a receptor binding sites in the gastrointestinal tract. Neuroscience 31 :757-770.
Green, T., and G.J. Dockray (1988) Characterization of the peptidergic afferent innervation of the stomach in the rat, mouse and guinea-pig. Neuroscience 25: 181-193.
Grundy, D., and T. Scratcherd (1989) Sensory afferents from the gastrointes- tinal tract. In J.D. Scbultz, S.G. Wood, and B.B. Rauner (eds): Handbook of Physiology: The Gastrointestinal System. Vol. 1. Bethesda, MD: Am. Physiol. Soc., pp. 593-620.
Handwerker, H.O., S. Kilo, and P.W. Reeh (1991) Unresponsive afferent nerve fibers in the sural nerve of the rat. J. Physiol. 435:229-242.
Hayashi, H., K. Ohsumi, N. Ueda, M. Fujiwara, and N. Mizumo (1982a) Effect of spinal ganglionectomy on substance P-like immunoreactivity in the gastroduodenal tract of cats. Brain Res. 232227-230.
Hayashi, H., K. Ohsumi, M. Fujiwara, N. Mizumo, I. Kanazawa, and H. Yajima (198213) Immunohistochemical studies on enteric substance P of extrinsic origin in the cat. J. Autonomic New. Syst. 5207-217.
Hazarika, N.H., J. Coote, and C.B.B. Downman (1964) Gastrointestinal viscerotomes in the cat. J. Neurophysiol. 27:107-116.
Holst, M.C., and T.L. Powley (1992) Vagal afferent innervation of the proximal duodenum. Dig. Dis. Sci. 37:A12.974.
Holzer, P. (1988) Local effector functions of capsaicin-sensitive sensory nerve endings: Involvement of tachykinins, calcitonin gene-related pep- tide and other neuropeptides. Neuroscience 24.739-768.
Hudson, L.C., and J.F. Cummings (1985) The origins of innervation of the esophagus of the dog. Brain Res. 326:125-136.
Janig, W., and J.F.B. Morrison (1986) Functional properties of spinal visceral afferent supplying abdominal and pelvic organs, with special emphasis on visceral nociception. In Cervero and J.F.B. Morrison (eds): Visceral Sensation (Progr. Brain Res.). pp. 87-114.
Khurana, R.K., and J.M. Petras (1991) Sensory innervation of the canine esophagus, stomach and duodenum. Am. J. Anat. 192293-306.
Kostrzewa, R.M., and D.M. Jacobowitz (1974) Pharmacological actions of 6-hydroxydopamine. Pbarmacol. Rev. 26; 199-288.
Kruger, L. (1988) Morphological features of thin afferent fibers: A new interpretation of nociceptor function. Progr. Brain Res. 74:253-257.
Kunusoki, S., K. Inoue, M. Iwamori, Y. Nagai, and T. Mannen (1989) Discrimination of human dorsal root ganglion cells by antifucosyl GM1 antibody. Brain Res. 494:391-395.
Kunusoki, S., K. Inoue, M. Iwamori, Y. Nagai, and T. Mannen (1991) Fucosylated glycoconjugates in human dorsal root ganglion cells with unmyelinated axons. Neurosci. Lett. 126:159-162.
Kuo, D.C., G.C.H. Yang, D.S. Yamasaki, and G.M. Krauthamer (1982) A wide field electron microscopic analysis of the fiher constituents of the major splanchnic nerve in cat. J. Comp. Neurol. 210:49-58.

186 N. CLERC AND C. MAZZIA
LaMotte, C.C., S.E. Kapadia, and C.M. Shapiro (1991) Central projections of the sciatic, saphenous, median, and ulnar nerves of the rat demonstrated by transganglionic transport of choleragenoid-HRP (B-HRP) and wheat- germ agglutinin-HRP (WGA-HRP). J. Comp. Neurol. 311.546-562.
Leander, S., E. Brodin, R. Hakanson, F. Sundler, and R. Uddman (1982) Neuronal substance P in the esophagus. Distribution and effects on motor activity. Acta Physiol. Scand. 115427435.
Maggi, C.A., and A. Meli (1988) The sensory-efferent function of capsacein- sensitive sensory neurons. Gen. Pharmacol. 19: 1-43.
Mantyh, P.W., C.R, Mantyh, T. Gates, S.R. Vigna, and J.E. Maggio (1988) Receptors binding sites for substance P and substance K in the canine gastrointestinal tract and their possible role in inflammatory bowel disease. Neuroscience 25:817-837.
Masserini, M., E. Freire, P. Palestini, E. Calappi, and G. Tettamanti (1992) Fuc-GM1 ganglioside mimics the receptor function of GM1 for cholera toxin. Biochemistry 3112422-2426.
McIlhinney, R.A.J., S.J. Bacon, and A.D. Smith (1988) A simple and rapid method for the production of cholera B-chain coupled to horseradish peroxidase for neuronal tracing. J. Neurosci. Methods 22: 189-194.
Mei, N. (1983) Recent studies on intestinal vagal afferent innervation. Functional implications. J. Autonomic Nerv. Syst. 9r199-206.
Mesulam, M.M. (1976) Principles of horseradish peroxidase neurohistochem- istry and their applications for tracing neural pathways. Axonal trans- port, enzyme histochemistry and light microscopic analysis. In M.M. Mesulam (ed): Tracing Neural Connections With Horseradish Peroxi- dase. Chichester: Wiley, pp. 1-153.
Neuhuber, W. (1987) Sensory vagal innervation of the rat esophagus and cardia: A light and electron microscopic anterograde tracing study. J. Autonomic Nerv. Syst. 20r243-255.
Perrin, J., J. Crousillat, and N. Mei (1981) Assessment of true splanchnic glucoreceptors in the jejuno-ileum of the cat. Brain Res. Bull. 7:625-628.
Quigg, M., L.G. Elfvin, and H. Aldskogius (1988) Distribution of cardiac sympathetic afferent fibers in the guinea pig heart labeled by antero- grade transport of wheat germ agglutinin-horseradish peroxidase. J. Autonomic Nerv. Syst. 25107-118.
Reubi, J.C., U. Horisberger, B. Waser, J.O. Gebbers, and J. Laissue (1992) Preferential location of Somatostatin receptors in germinal centers of human gut lymphoid tissue. Gastroenterology 103:1207-1214.
Riviero-Melian, C., and G. Grant (1990) Distribution of lumbar dorsal root fibers in the lower thoracic and lumbosacral spinal cord of the rat studied with choleragenoid horseradish peroxidase conjugate. J. Comp. Neurol. 299:470481.
Robertson, B., and G. Grant (1985) A comparison between wheat germ agglutinin- and choleragenoid-horseradish peroxidase as anterogradely transported markers in central branches of primary sensory neurones in the rat with some observations in the cat. Neuroscience 14r895-905.
Robertson, B., and G. Grant (1989) Immunocytochemical evidence for the localization of the GM1 ganglioside in carbonic anhydrase-containing and RT 97-immunoreactive rat primary sensory neurons. J. Neurocytol. 18:77-86.
Robertson, B., M.J. Perry, and S.N. Lawson (1991) Populations of rat spinal primary afferent neurons with choleragenoid binding compared with those labelled by markers for neurofilament and carbohydrate groups: A quantitative immunocytochemical study. J. Neurocytol. 20r387-395.
Rodrigo, J., J.M. Polak, L. Fernandez, M.A. Ghatei, P. Mulderry, and S.R. Bloom (1985) Calcitonin gene-related peptide immunoreactive sensory and motor nerves of the rat, cat, and monkey esophagus. Gastroenterol- ogy88:444451.
Rozsa, Z., J. Mattila, and E.D. Jacobson (1988) substance P mediates a gastrointestinal thermoreflex in rats. Gastroenterology 95r265-276.
Sengupta, J.N., D. Kauvar, and R.K. Goyal (1989) Characteristics of vagal esophageal tension-sensitive afferent fibers in the opossum. J. Neuro- physiol. 61 : 1001-1010.
Sengupta, J.N., J.K. Saha, and R.K. Goyal (1990) Stimulus-response func- tion studies of esophageal mechanosensitive nociceptors in sympathetic afferents of opossum. J. Neurophysiol. 64:796-812.
Sengupta, J.N., J.K. Saha, and R.K. Goyal (1992) Differential sensitivity to Bradykinin of esophageal distension-sensitive mechanoreceptors in va- gal and sympathetic afferents of the opossum. J. Neurophysiol. 68r1053- 1067.
Stanisz, A.M., Befus D., and J. Bienenstock (1986) Differential effects of Vasoactive Intestinal Peptide, substance P and Somatostatin on immuno- globulin synthesis and proliferations by lymphocytes from Peyer’s patches, mesenteric lymph nodes and spleen. J. Immunol. 136:152-156.
Stead, R.H. (1992) Nerve remodelling during intestinal inflammation. Ann. N.Y. Acad. Sci., Ann. N.Y. Acad. Sci., Vol. 664, Edited by Stead, R.H., N.H. Perdue, H. Cooke, D.W. Powell, and K.E. Barret, pp. 443-455.
Stead, R.H., M.H. Perdue, M.G. Blennerhasset, Y. Kakuta, P. Sestini, and J. Bienenstock (1990) In S. Freier (ed): The Innervation of Mast Cells. Neuroendocrine-Immune Network. Boca Raton, FL: CRC Press, Inc., pp. 19-37.
Straw, W. (1976) Use of peroxidase inhibitors for immunoperoxidase procedures. In G. Feldman, P. Druet, J. Bignon, and S. Avraemas (eds): Immunoenzymatic Techniques (Proc. of the first international sympo- sium on immunoenzymatic techniques INSERM, Symposium No. 2). Amsterdam: North-Holland Publishing Company, pp. 117-124.
Thuneberg L. (1989) Interstitial cells of Cajd. In J.D. Schultz, S.G. Wood, and B.B. Rauner (eds): Handbook of Physiology: The Gastrointestinal System. Vol I. Bethesda, MD: Am. Physiol. Soc., pp. 349-386.
Tranzer, J.P., and H. Thoenen (1968) An electron microscopic study of selective, acute degeneration of sympathetic nerve terminals after administration of 6-hydroxydopamine. Experientia 24: 155-156.
Vacca, L.L., S.J. Abrahams, and N.E. Naftchi (1980) A modified peroxidase- antiperoxidase procedure improved localization of tissue antigens: Local- ization of substance P in rat spinal cord. J. Histochem. Cytochem. 28t297-307.
Van Heyningen, W.E. (1974) Gangliosides as membrane receptors for tetanus toxin, cholera toxin and serotonin. Nature 249:415417.
Weinberg, R.J., D.J. Tracey, and A. Rustioni (1990) Extracellular labeling of unmyelinated dorsal root terminals after WGA-HRP injections in spinal ganglia. Brain Res. 523:351-355.




















