
Generalized theory for the dynamic analysis of thin shells with
Upload
independentCategory
view
2download
0
Earth and Planetary Science Letters 299 (2010) 310–316
Contents lists available at ScienceDirect
Earth and Planetary Science Letters
j ourna l homepage: www.e lsev ie r.com/ locate /eps l
Rare earth elements in mussel shells of the Mytilidae family as tracers for hidden andfossil high-temperature hydrothermal systems
Michael Bau a,b,⁎, Simona Balan a,1, Katja Schmidt a,2, Andrea Koschinsky a,b
a Earth and Space Science Program, School of Engineering and Science, Jacobs University Bremen, Campus Ring 1, 28759 Bremen, Germanyb Integrated Environmental Studies Program, Jacobs University Bremen, Campus Ring 1, 28759 Bremen, Germany
⁎ Corresponding author. Earth and Space Science ProgScience, Jacobs University Bremen, Campus Ring 1, 2875421 2003564.
E-mail address: [email protected] (M. Bau1 Present address: Department of Environmental Scie
University of California Berkeley,137 Mulford Hall #311States.
2 Present address: Deutsches GeoForschungsZentrumGeothermal Research, Telegraphenberg, 14473 Potsdam
0012-821X/$ – see front matter © 2010 Elsevier B.V. Adoi:10.1016/j.epsl.2010.09.011
a b s t r a c t
a r t i c l e i n f oArticle history:Received 13 March 2010Received in revised form 8 September 2010Accepted 9 September 2010
Editor: M.L. Delaney
Keywords:rare earth elementsMytilusBathymodiolushydrothermal systembiomineralizationproxy
Bathymodiolus mussels of the Mytilidae family live in ecological niches at marine vent sites wherehydrothermal fluids discharge at the seafloor and mix with ambient seawater. We report the firstconcentration data for rare earth elements and yttrium (REY) of Bathymodiolus shells and of low-temperaturediffuse hydrothermal fluids venting in their respective habitat at three hydrothermal vent fields along theMid-Atlantic Ridge (MAR) and for littoral Mytilus edulis shells from the western North Atlantic, Dingle Bay,Ireland, and from the southern North Sea, German Bight, Germany. Similar to high-temperature hydrothermalsolutions expelled in the respective area, the low-temperature diffuse fluids from the hydrothermal vent sitesdisplay positive Eu anomalies. These indicate that the fluids carry a component previously involved in high-temperature water–rock interaction, as decoupling of Eu from neighbouring REY requires temperatures aboveabout 200 °C. While the Bathymodiolus shells from the hydrothermal vent fields reflect these positive Euanomalies, the littoral M. edulis shells studied for comparison, do not show Eu anomalies, consistent with thelack of any high-temperature hydrothermal activity in their coastal environments. Shells of mussels of theMytilidae family, such as Bathymodiolus and M. edulis, therefore, are archives whose REY distribution can beused as a proxy for the Eu anomaly of the waters in which the mussels grew. Although the Ca carbonate of amussel shell is precipitated from the extrapallial fluid of themussel and not directly from seawater, the Eu/Eu*ratio of seawater is not significantly modified under the low-temperature conditions prevailing during vitalprocesses and biomineralization. This allows one to use the positive Eu anomalies in the REY distributionpatterns of Bathymodiolus shells as a tracer for hidden or fossil high-temperature hydrothermal systemswhere other indicators of high-temperature hydrothermalism are not available.
ram, School of Engineering and9 Bremen, Germany. Tel.: +49
).nce, Policy, and Management,4, Berkeley, CA 94720, United
GFZ, International Center of, Germany.
ll rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
The formation of mussel shells is a prime example of biominer-alization and sensitive to changes of the physico-chemical conditionsin a mussel's habitat. Interest in the use of mussel shells as archives ofenvironmental conditions has seen a steady increase since the early1980s, and their chemical composition has been shown to providevaluable information on the physico-chemical conditions in amussel'shabitat (e.g., Cravo et al., 2008; Goldberg, 1986; Koide et al., 1982;Puente et al., 1996; Thébault et al., 2007; Wanamaker et al., 2008, and
references therein). The few studies that investigated the partitioningof the rare earth elements and yttrium (REY) between seawater andthe marine biogenic Ca carbonate of corals (e.g., Akagi et al., 2004;Sholkovitz and Shen, 1995; van de Flierdt et al., 2010), stromatoliticmicrobialites (Webb and Kamber, 2000) or foraminifera (e.g., Haley etal., 2005; Vance and Burton, 1999) suggest that, despite the potentialinfluence of vital effects and biomineralization, neighbouring REYsuch as Nd, Sm, Eu, Gd and Tb are incorporated into the biogeniccarbonate structure without major fractionation. Shells of Bath-ymodiolus mussels and their littoral relative Mytilus edulis, therefore,may serve as archives of the dissolved REY distribution in theirhabitat.
Common oxic seawater is characterized by shale- or chondrite-normalised REY distribution patterns (Fig. 1A) that show negative Ceanomalies, large positive Y anomalies, and smaller positive anomaliesfor La and Gd (e.g., Bau et al., 1995; Elderfield, 1988). In markedcontrast, marine hydrothermal systems can produce high-tempera-ture aqueous fluids, with black-smoker fluids at mid-ocean ridges asprominent examples, that lack these anomalies, but display pro-nounced positive Eu anomalies (e.g., Bau and Dulski, 1999; Michard et
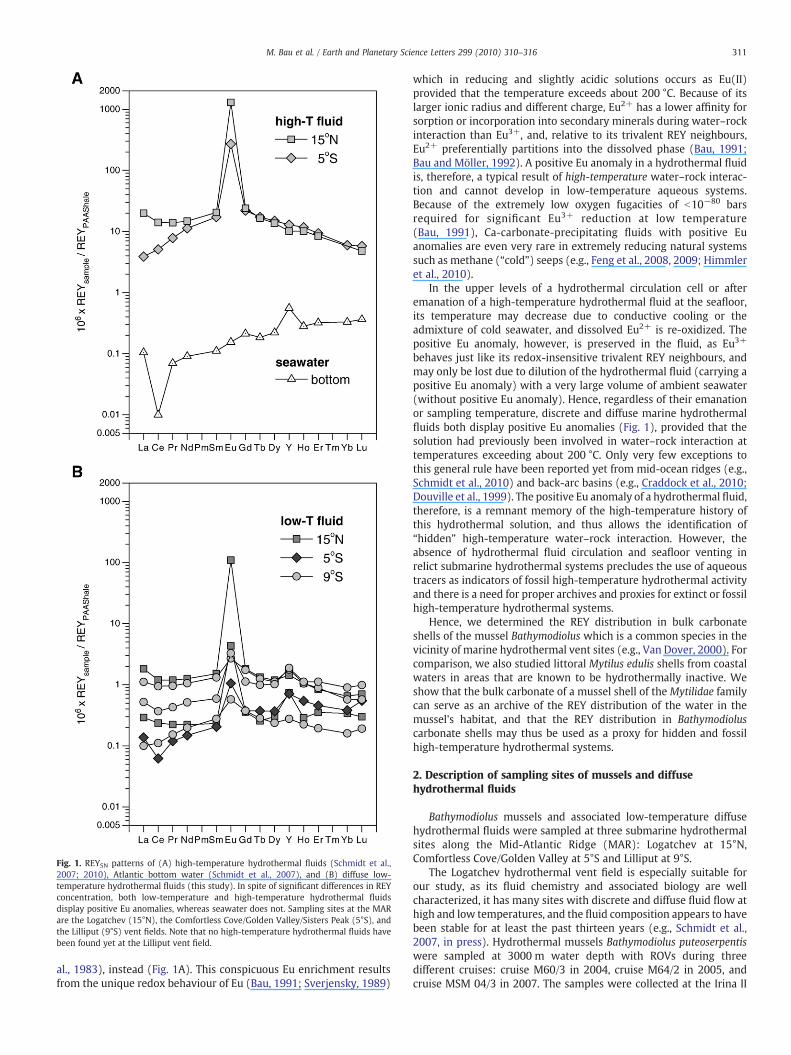
Fig. 1. REYSN patterns of (A) high-temperature hydrothermal fluids (Schmidt et al.,2007; 2010), Atlantic bottom water (Schmidt et al., 2007), and (B) diffuse low-temperature hydrothermal fluids (this study). In spite of significant differences in REYconcentration, both low-temperature and high-temperature hydrothermal fluidsdisplay positive Eu anomalies, whereas seawater does not. Sampling sites at the MARare the Logatchev (15°N), the Comfortless Cove/Golden Valley/Sisters Peak (5°S), andthe Lilliput (9°S) vent fields. Note that no high-temperature hydrothermal fluids havebeen found yet at the Lilliput vent field.
311M. Bau et al. / Earth and Planetary Science Letters 299 (2010) 310–316
al., 1983), instead (Fig. 1A). This conspicuous Eu enrichment resultsfrom the unique redox behaviour of Eu (Bau, 1991; Sverjensky, 1989)
which in reducing and slightly acidic solutions occurs as Eu(II)provided that the temperature exceeds about 200 °C. Because of itslarger ionic radius and different charge, Eu2+ has a lower affinity forsorption or incorporation into secondary minerals during water–rockinteraction than Eu3+, and, relative to its trivalent REY neighbours,Eu2+ preferentially partitions into the dissolved phase (Bau, 1991;Bau and Möller, 1992). A positive Eu anomaly in a hydrothermal fluidis, therefore, a typical result of high-temperature water–rock interac-tion and cannot develop in low-temperature aqueous systems.Because of the extremely low oxygen fugacities of b10−80 barsrequired for significant Eu3+ reduction at low temperature(Bau, 1991), Ca-carbonate-precipitating fluids with positive Euanomalies are even very rare in extremely reducing natural systemssuch as methane (“cold”) seeps (e.g., Feng et al., 2008, 2009; Himmleret al., 2010).
In the upper levels of a hydrothermal circulation cell or afteremanation of a high-temperature hydrothermal fluid at the seafloor,its temperature may decrease due to conductive cooling or theadmixture of cold seawater, and dissolved Eu2+ is re-oxidized. Thepositive Eu anomaly, however, is preserved in the fluid, as Eu3+
behaves just like its redox-insensitive trivalent REY neighbours, andmay only be lost due to dilution of the hydrothermal fluid (carrying apositive Eu anomaly) with a very large volume of ambient seawater(without positive Eu anomaly). Hence, regardless of their emanationor sampling temperature, discrete and diffuse marine hydrothermalfluids both display positive Eu anomalies (Fig. 1), provided that thesolution had previously been involved in water–rock interaction attemperatures exceeding about 200 °C. Only very few exceptions tothis general rule have been reported yet from mid-ocean ridges (e.g.,Schmidt et al., 2010) and back-arc basins (e.g., Craddock et al., 2010;Douville et al., 1999). The positive Eu anomaly of a hydrothermal fluid,therefore, is a remnant memory of the high-temperature history ofthis hydrothermal solution, and thus allows the identification of“hidden” high-temperature water–rock interaction. However, theabsence of hydrothermal fluid circulation and seafloor venting inrelict submarine hydrothermal systems precludes the use of aqueoustracers as indicators of fossil high-temperature hydrothermal activityand there is a need for proper archives and proxies for extinct or fossilhigh-temperature hydrothermal systems.
Hence, we determined the REY distribution in bulk carbonateshells of the mussel Bathymodiolus which is a common species in thevicinity of marine hydrothermal vent sites (e.g., Van Dover, 2000). Forcomparison, we also studied littoral Mytilus edulis shells from coastalwaters in areas that are known to be hydrothermally inactive. Weshow that the bulk carbonate of a mussel shell of the Mytilidae familycan serve as an archive of the REY distribution of the water in themussel's habitat, and that the REY distribution in Bathymodioluscarbonate shells may thus be used as a proxy for hidden and fossilhigh-temperature hydrothermal systems.
2. Description of sampling sites of mussels and diffusehydrothermal fluids
Bathymodiolus mussels and associated low-temperature diffusehydrothermal fluids were sampled at three submarine hydrothermalsites along the Mid-Atlantic Ridge (MAR): Logatchev at 15°N,Comfortless Cove/Golden Valley at 5°S and Lilliput at 9°S.
The Logatchev hydrothermal vent field is especially suitable forour study, as its fluid chemistry and associated biology are wellcharacterized, it has many sites with discrete and diffuse fluid flow athigh and low temperatures, and the fluid composition appears to havebeen stable for at least the past thirteen years (e.g., Schmidt et al.,2007, in press). Hydrothermal mussels Bathymodiolus puteoserpentiswere sampled at 3000 m water depth with ROVs during threedifferent cruises: cruise M60/3 in 2004, cruise M64/2 in 2005, andcruise MSM 04/3 in 2007. The samples were collected at the Irina II

312 M. Bau et al. / Earth and Planetary Science Letters 299 (2010) 310–316
vent site (3000 m depth), which consists of a large sulfide complex(approximately 40 m×60 m) with several active and inactivechimneys. It is surrounded by many sites of diffuse outflow(Tb30 °C) and extensive B. puteoserpentis mussel beds. Most sampleswere taken immediately adjacent to a location of diffuse hydrothermalfluid outflow (244 ROV 9-7 to 244 ROV 9-12), while others were takendirectly at the base of the large sulfide complex away from sites ofpresent-day diffuse fluid outflow (259 ROV 29-8 and 259 ROV 29-15).
Bathymodiolusmussels were also sampled from two hydrothermalfields along the southern MAR (Comfortless Cove/Golden Valley andLilliput) during cruises M64/1 in 2005, M68/1 in 2006, Atalante-Leg2MARSUED IV in 2008 and M78/2 in 2009. At Comfortless Cove,mussels are abundant at, and were sampled from, a fissured area withdiffuse flow (sampled at T=8 °C) known as Golden Valley (3000 mdepth). This area is about 70 m distance from the Sister's Peak area ofhigh-temperature black-smoker activity, where fluids with tempera-tures up to 407 °C exit the seafloor (Haase et al., 2007; Koschinskyet al., 2008).
At the Lilliput hydrothermal field at 9°33′S on the MAR, diffuse low-temperature hydrothermal solutions are present, but no hot vent fluidshave been found yet. Bathymodiolusmusselswere sampled from sites ofdiffuse low-temperature flow (Tb16 °C) at a depth of 1500m (Haase etal., 2009; Koschinsky et al., 2006; Perner et al., 2007). Five relativelysmall diffuse vent sites are known at this field and occur across whatappears to be a large young lava flow that is partly covered by a thinlayer of hydrothermal Fe (hydr)oxide sediment. The fauna at the Lilliputhydrothermal vents largely consists of juvenile Bathymodiolus spp.(b1.5 cm in size) suggesting a recent reactivation of hydrothermal fluidflow. Mussels are mostly concentrated in discrete densely coveredclusters directly at fluid outflow orifices or cracks, surrounded by thickorange-red Fe oxyhydroxide layers.
At all three hydrothermal systems, sampling of diffuse fluids withthe “KIPS” fluid sampling system (Garbe-Schönberg et al., 2006) wascarried out at the same time as sampling of mussel populations.
To allow for comparison between REY distributions of bivalve shellsfrom areas of known hydrothermal activity and from environmentswithout any hydrothermal activity, we also sampled littoral M. edulisshells from shallow coastal waters of Dingle Bay (off Inch, County Kerry,western Republic of Ireland) and German Bight (southern North Sea,Germany; off Cuxhaven and the island of Sylt, resp.).
3. Analytical
3.1. Sample preparation and analysis
The Bathymodiolus andM. edulis shells were treated using identicalprotocols in order to allow for a valid comparison between chemicaldata. After physical cleaning, the shells were exposed to a temper-ature of 200 °C in a muffle oven for 12 h after which the organicperiostracum with its sorbed trace cations was hand separated fromthe carbonate parts of the shells. For both, the Bathymodiolus and theM. edulis shells, we decided against the separation of the outer calciticostracum from the inner aragonitic nacreous layer (hypostracum).Separation of calcite and aragonite shell layers by heating of the shellsto temperatures of N300 °C (the commonly applied method ofBourgoin, 1988; see also Puente et al., 1996) was possible for M.edulis for which it had originally been developed. However, thehydrothermal Bathymodiolus shells disintegrated completely whensubjected to temperatures above 300 °C. To allow proper comparisonof the hydrothermal and littoral shells we, therefore, subjected bothshell types to the same procedure and did not separate the calcite andthe aragonite layers. This approach does not affect the validity of ourresults, because there is no reason to assume that the relative partitioncoefficients of isovalent neighbouring REY, such as Sm, Eu, Gd, and Tb,between aragonite and seawater should be significantly differentfrom those between calcite and seawater.
The bulk carbonate shellwas then crushed in an agatemortar. About1000 mg of carbonate powder was digested for 2 h at 90 °C in 30 ml of5 M suprapure-grade HNO3 in a Teflon beaker. The solution was thenfiltered through a 0.2-μM cellulose acetate filter. The solution wasevaporated to incipient dryness by heating to 110 °C. The residue wastaken up in 10 ml of 0.5 M HNO3, transferred to a polyethylene bottle,and diluted to 25 ml with 0.5 M HNO3. As certified reference materialthe international reference standard JLs-1 (a Japanese limestone) hadbeen chosen, because itmost closelymatches the carbonate shellmatrixand the low REY concentrations of bivalve shells.
Due to the low REY concentrations and to avoid potential matrixproblems, we employed a separation and preconcentration procedure(Bau and Dulski, 1996; Kulaksiz and Bau, 2007; Shabani et al., 1992)that had also been successfully applied in the analysis of carbonateand silicate rock standards (Dulski, 2001): About 20 mL of thedigested sample solution was diluted to 1000 mL with de-ionizedwater, subsequently acidified to pH 2 with suprapure HCl and spikedwith 1 mL of a 250-ppb Tm solution to monitor REY recovery. A smallaliquot was taken for the determination of the Tm referenceconcentration, before the sample was passed over a pre-cleaned ionexchange column (Sep-Pak C18 cartridge) loaded with a mixture ofethyl-hexyl-phosphates. To remove major alkali and alkali earthelements, the cartridge was washed with 0.01 M HCl before the REYwere eluted with 40 ml of 6 M suprapure HCl. The eluate wasevaporated in a Teflon beaker to incipient dryness, and eventuallytaken up in 10 mL of 0.5 M HNO3 including an internal standard forICP-MS analysis, consisting of Ru, Re, and Bi. Procedural blanks and theJLs-1 reference standard were processed along with the samples foranalytical quality control. All subsequentmeasurements of the eluatesand the Tm reference aliquots were performed with a quadrupole-based ICP-MS (PerkinElmer DRC-e). The final dilution of the completeprocedure (acid decomposition plus REY separation/preconcentrationplus analytical measurement) was about 10, which is 20 times higherthan direct measurement without REY preconcentration.
Sample treatment and REY analyses of the diffuse hydrothermalfluids followed the procedure described in detail by Schmidt et al.(2007, in press) and also involved matrix separation and preconcen-tration of REY and analysis by ICP-MS.
3.2. Analytical quality assessment
The analytical precision reflecting the long-term reproducibilitywas determined by applying our analytical procedure of sampledecomposition, REY separation and preconcentration, and measure-ment by ICP-MS tomultiple independent aliquots (n=11) of the JLs-1(Japanese limestone) international reference standard which ischaracterized by very low REY concentrations. The results arereported in Table 1 of the online supplementary material. Precisionis expressed as relative standard deviation (RSD; 2σ) from theaverage, and yields an uncertainty of b5% RSD for Nd, Sm, Eu, Dy andEr, b6% RSD for Ce, Pr, Gd, Tb, Ho and Yb, b7% RSD for La und Lu, andb9% RSD for Y. Similar results were obtained for two aliquots of ahomogenised mussel shell (b5% RSD for La, Ce, Nd, Sm, Eu, Gd, Tb, Dy,Y, Ho, Er, and Yb, and b6% RSD for Pr and Lu).
The analytical accuracy of the applied method was establishedfrom our average value of the JLs-1 standard compared to publishedreference values (Fig. 2) which were determined using two differentsample treatment protocols (“method B without REY preconcentra-tion” and “method C with REY preconcentration” of Dulski, 2001);note that due to its very low REY concentrations, only very limitedREY data have been published for JLs-1. Our data and that obtained byDulski (2001) using the same REY separation and preconcentrationtechnique (“method C” in Fig. 2) are the same within b3% for Nd, Sm,Eu, Gd, Tb, Dy, Ho, Y and Er, b10% for Ce, Pr, Yb and Lu, and 20% for La.The agreement is even better (Fig. 2) between our data and thatobtained by “method B” of Dulski (2001). The reason for the

Fig. 2. Comparison of the REYSN patterns of the international reference standard JLs-1, aPermian limestone from Japan with (ultra)low REY concentrations, determined with(method C) and without (method B) REY separation and preconcentration. Except forLa, the data sets are in good agreement (note that in contrast to Figs. 1 and 3, the y-axisextends only over one order of magnitude). Data for Eu, Yb, and Lu have not beenplotted for method B (Dulski, 2001), due to severe interference problems (data are,however, included in Table 1 of the online supplementary material).
313M. Bau et al. / Earth and Planetary Science Letters 299 (2010) 310–316
difference betweenmeasured La concentrations in reference standardJLs-1 in different studies is unclear, but these data demonstrate theneed for consensus data for (ultra)low-REY marine limestonestandards. Nevertheless, we emphasize that elemental ratios such asGd/Gd*, Y/Ho, and, in particular, the Eu/Eu* ratio (Eu/Eu*=EuSN/(0.5SmSN+0.5GdSN); subscript SN denotes normalisation to Post-Archean Australian Shale of McLennan, 1989) that quantifies the Euanomalywhich is the focus of this contribution, agreewithin b5%withpreviously reported data from Dulski (2001).
4. Results
4.1. Mussel shells
All REY concentration data for mussel shells are reported in Table 2of the online supplementary material. The Bathymodiolus shells fromthe Logatchev field have high Nd concentrations between 30.9 and99.6 μg kg−1. Their REYSN patterns show only minor fractionationbetween light, middle and heavy rare earth elements (LREE, MREEand HREE, respectively), and display small positive Y anomalies andsmall negative Ce anomalies (Fig. 3A); they are, however, character-ized by very large positive Eu anomalies (Eu/Eu*: 26.6–137).
Shells from the Golden Valley site have Nd concentrations between22.4 and 58.7 μg kg−1 and show consistent REYSN patterns that increasefrom the LREE to the MREE and decrease from the MREE to the HREE(Fig. 3B). The negative Ce and positive Y anomalies aremore pronouncedthan those of shells from all other locations. All Golden Valley shells showstrong positive Eu anomalies (Eu/Eu*: 4.0–5.1).
Shells from the Lilliput field may be subdivided into two types(Fig. 3C): type 1 shells have Nd concentrations between 29.0 and44.9 μg kg−1 and show positive La anomalies, no Ce anomalies, andsmall but distinctive positive Eu anomalies (Eu/Eu*: 1.7–2.6); a sub-population referred to as type 2 shells has lower Nd concentrationsbetween 7.47 and 21.9 μg kg−1 and shows negative Ce anomalies andpositive La anomalies, but only very small positive Eu anomalies (Eu/Eu*: 1.3–1.5).
The littoral M. edulis shell from Dingle Bay, Ireland, shows a ratherlow Nd concentration of 13.8 μg kg−1 and a Eu/Eu* ratio of 0.99.Except for the missing Eu anomaly, its REYSN pattern (Fig. 3D) issimilar to that of the Golden Valley shells and displays positive La andY anomalies and a negative Ce anomaly. The Mytilus shells from closeto Cuxhaven and Sylt in the southern North Sea also lack positive Euanomalies (Eu/Eu*: 1.00 and 0.97, resp.) and show low Ndconcentrations (16.5 μg kg−1 and 11.0 μg kg−1, resp.).
Comparison of our REY data for modern marine mussel shells toliterature data is hampered, because published REY data for musselshells are (in contrast to data for other metals; Cravo et al., 2008; andreferences therein) very rare. REY data for a Calyptogena shell fromthe Iheya Ridge hydrothermal vent field in the Okinawa Trough(Hongo and Nozaki, 2001) reveal significant differences to our resultsfor Bathymodiolus shells, as it displays very low REY concentrationsand does not show a distinct positive Eu anomaly (Eu/Eu*=1.3).Hongo and Nozaki (2001) suggested that hydrothermal solutions didnot influence the habitat of this Calyptogena mussel, a conclusionsupported by our results. The only other published REY data formodern bivalve shells, that we are aware of, are for an unspecifiedshell from a cold seep at Alaminos Canyon, Gulf of Mexico, that doesnot show a positive Eu anomaly (Feng et al., 2008), and for two clamsfrom the coast of Western Australia (Webb and Kamber, 2000), thatappear to suffer from analytical uncertainty.
4.2. Low-temperature diffuse hydrothermal fluids
The low-temperature (T: 8.5 to 30 °C) diffuse hydrothermal fluidsfrom all three study sites (Table 3 of the online supplementary material)display significant positive Eu anomalies (Eu/Eu*: 1.8 to 66) and Ndconcentrations that range from 5×10−3 to 42.5×10−3 μg kg−1 (Fig. 1B).For the Golden Valley and Logatchev samples these positive Eu anomaliesreflect those of the respective high-temperature fluids present in the area(illustrated by Eu/Eu* of 14 and 58, respectively, for the high-temperaturefluids in Fig. 1A), although the latter shows significantly higher REYconcentrations (Nd: 384×10−3 to 502×10−3 μg kg−1). This relationshipis similar to thatobservedatotherhydrothermal sites along theMAR, suchas TAG (Bau and Dulski, 1999; James and Elderfield, 1996; Mills andElderfield, 1995).
5. Discussion
5.1. Inter- and intra-population variation of the REY distribution inthe shells
Our sample set of mussel shells from marine hydrothermal environ-ments defines a broad general trend in which the maximum Ndconcentration of hydrothermal shells increases with increasing Eu/Eu*ratio. Only the latter, however, clearly distinguishes the shell populationsfrom the individual vent fields (Fig. 4) and allows one to discriminatebetween mussel shells from hydrothermal and from non-hydrothermallittoral environments. Only mussel shells from the hydrothermalenvironments show positive Eu anomalies. In spite of this inter-population difference, the intra-population variation of the REY patternsis rather small (Fig. 3).However, the littoralM.edulis shell fromDingleBay(LaSN/SmSN=0.52) is depleted in LREE, while those from the southernNorth Sea are not (LaSN/SmSN=0.99 and 0.96); all littoral shells show thesame lack of a Eu anomaly.
5.2. REY in Bathymodiolus shells reveal high-temperature hydrothermalsystems
The positive Eu anomalies recorded in hydrothermal Bathymodiolusshells straightforwardly suggest that the water in their specific habitatcarries a pronounced positive Eu anomaly. In contrast, the absence of any
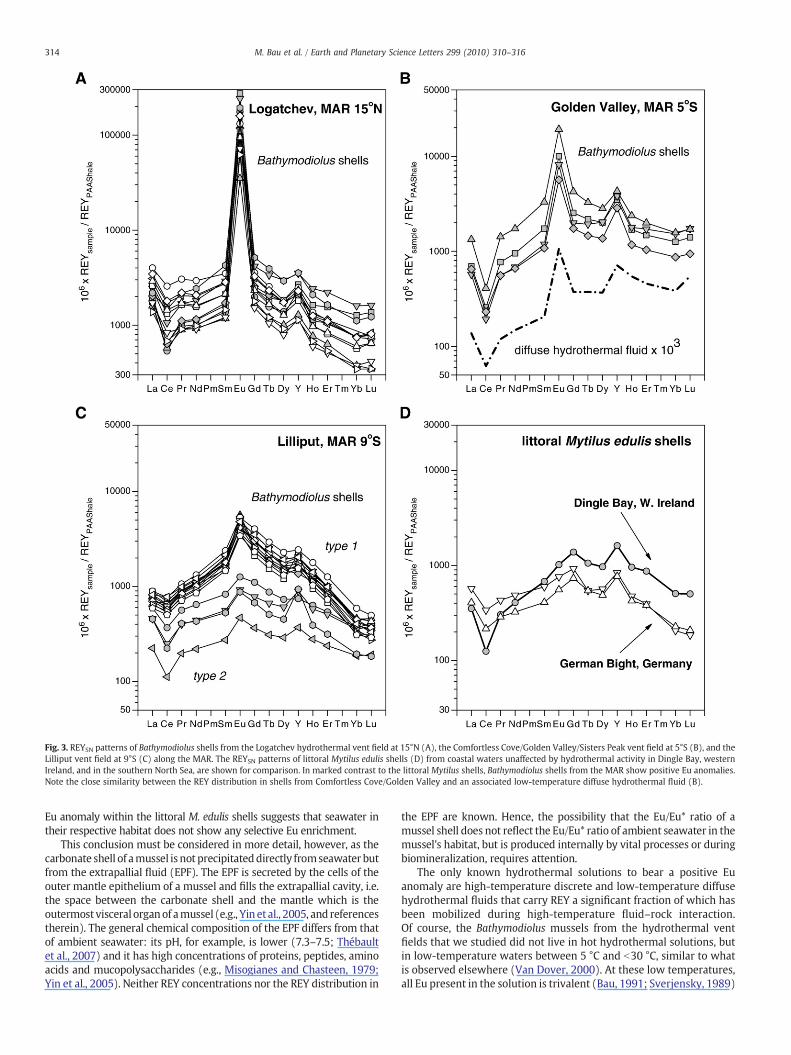
Fig. 3. REYSN patterns of Bathymodiolus shells from the Logatchev hydrothermal vent field at 15°N (A), the Comfortless Cove/Golden Valley/Sisters Peak vent field at 5°S (B), and theLilliput vent field at 9°S (C) along the MAR. The REYSN patterns of littoral Mytilus edulis shells (D) from coastal waters unaffected by hydrothermal activity in Dingle Bay, westernIreland, and in the southern North Sea, are shown for comparison. In marked contrast to the littoral Mytilus shells, Bathymodiolus shells from the MAR show positive Eu anomalies.Note the close similarity between the REY distribution in shells from Comfortless Cove/Golden Valley and an associated low-temperature diffuse hydrothermal fluid (B).
314 M. Bau et al. / Earth and Planetary Science Letters 299 (2010) 310–316
Eu anomaly within the littoral M. edulis shells suggests that seawater intheir respective habitat does not show any selective Eu enrichment.
This conclusion must be considered in more detail, however, as thecarbonate shell of amussel is not precipitateddirectly fromseawater butfrom the extrapallial fluid (EPF). The EPF is secreted by the cells of theouter mantle epithelium of a mussel and fills the extrapallial cavity, i.e.the space between the carbonate shell and the mantle which is theoutermost visceral organof amussel (e.g., Yin et al., 2005, and referencestherein). The general chemical composition of the EPF differs from thatof ambient seawater: its pH, for example, is lower (7.3–7.5; Thébaultet al., 2007) and it has high concentrations of proteins, peptides, aminoacids and mucopolysaccharides (e.g., Misogianes and Chasteen, 1979;Yin et al., 2005). Neither REY concentrations nor the REY distribution in
the EPF are known. Hence, the possibility that the Eu/Eu* ratio of amussel shell does not reflect the Eu/Eu* ratio of ambient seawater in themussel's habitat, but is produced internally by vital processes or duringbiomineralization, requires attention.
The only known hydrothermal solutions to bear a positive Euanomaly are high-temperature discrete and low-temperature diffusehydrothermal fluids that carry REY a significant fraction of which hasbeen mobilized during high-temperature fluid–rock interaction.Of course, the Bathymodiolus mussels from the hydrothermal ventfields that we studied did not live in hot hydrothermal solutions, butin low-temperature waters between 5 °C and b30 °C, similar to whatis observed elsewhere (Van Dover, 2000). At these low temperatures,all Eu present in the solution is trivalent (Bau, 1991; Sverjensky, 1989)
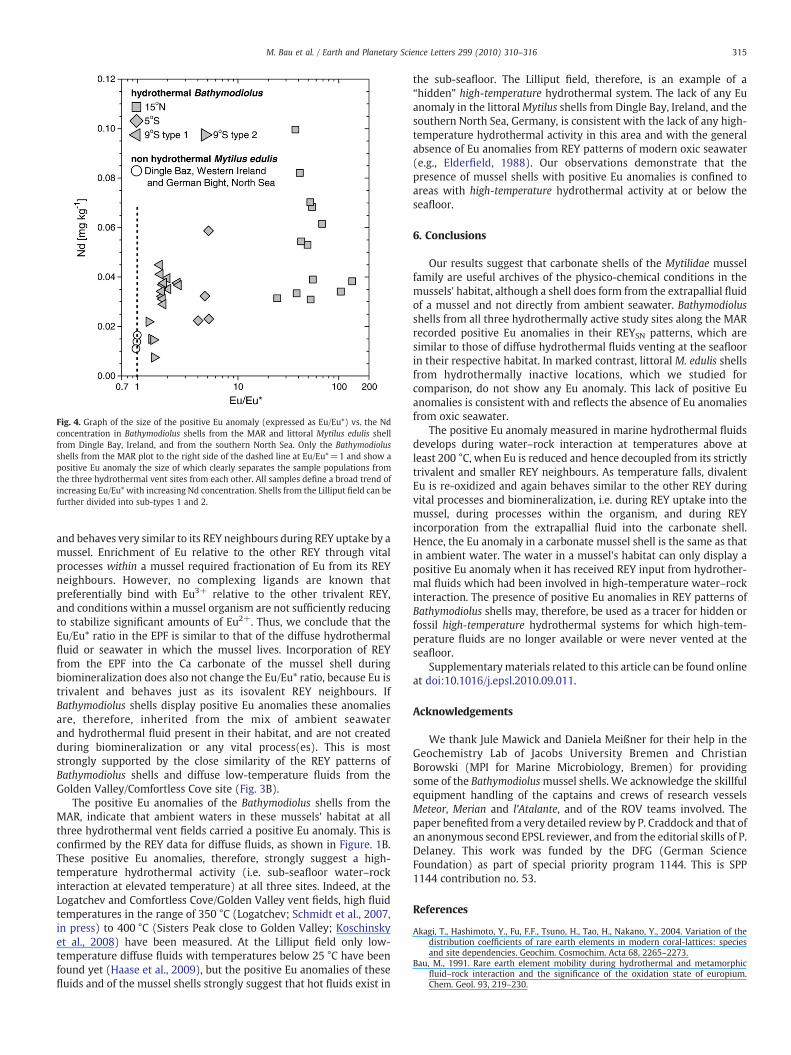
Fig. 4. Graph of the size of the positive Eu anomaly (expressed as Eu/Eu*) vs. the Ndconcentration in Bathymodiolus shells from the MAR and littoral Mytilus edulis shellfrom Dingle Bay, Ireland, and from the southern North Sea. Only the Bathymodiolusshells from the MAR plot to the right side of the dashed line at Eu/Eu*=1 and show apositive Eu anomaly the size of which clearly separates the sample populations fromthe three hydrothermal vent sites from each other. All samples define a broad trend ofincreasing Eu/Eu* with increasing Nd concentration. Shells from the Lilliput field can befurther divided into sub-types 1 and 2.
315M. Bau et al. / Earth and Planetary Science Letters 299 (2010) 310–316
and behaves very similar to its REY neighbours during REY uptake by amussel. Enrichment of Eu relative to the other REY through vitalprocesses within a mussel required fractionation of Eu from its REYneighbours. However, no complexing ligands are known thatpreferentially bind with Eu3+ relative to the other trivalent REY,and conditions within a mussel organism are not sufficiently reducingto stabilize significant amounts of Eu2+. Thus, we conclude that theEu/Eu* ratio in the EPF is similar to that of the diffuse hydrothermalfluid or seawater in which the mussel lives. Incorporation of REYfrom the EPF into the Ca carbonate of the mussel shell duringbiomineralization does also not change the Eu/Eu* ratio, because Eu istrivalent and behaves just as its isovalent REY neighbours. IfBathymodiolus shells display positive Eu anomalies these anomaliesare, therefore, inherited from the mix of ambient seawaterand hydrothermal fluid present in their habitat, and are not createdduring biomineralization or any vital process(es). This is moststrongly supported by the close similarity of the REY patterns ofBathymodiolus shells and diffuse low-temperature fluids from theGolden Valley/Comfortless Cove site (Fig. 3B).
The positive Eu anomalies of the Bathymodiolus shells from theMAR, indicate that ambient waters in these mussels' habitat at allthree hydrothermal vent fields carried a positive Eu anomaly. This isconfirmed by the REY data for diffuse fluids, as shown in Figure. 1B.These positive Eu anomalies, therefore, strongly suggest a high-temperature hydrothermal activity (i.e. sub-seafloor water–rockinteraction at elevated temperature) at all three sites. Indeed, at theLogatchev and Comfortless Cove/Golden Valley vent fields, high fluidtemperatures in the range of 350 °C (Logatchev; Schmidt et al., 2007,in press) to 400 °C (Sisters Peak close to Golden Valley; Koschinskyet al., 2008) have been measured. At the Lilliput field only low-temperature diffuse fluids with temperatures below 25 °C have beenfound yet (Haase et al., 2009), but the positive Eu anomalies of thesefluids and of the mussel shells strongly suggest that hot fluids exist in
the sub-seafloor. The Lilliput field, therefore, is an example of a“hidden” high-temperature hydrothermal system. The lack of any Euanomaly in the littoralMytilus shells from Dingle Bay, Ireland, and thesouthern North Sea, Germany, is consistent with the lack of any high-temperature hydrothermal activity in this area and with the generalabsence of Eu anomalies from REY patterns of modern oxic seawater(e.g., Elderfield, 1988). Our observations demonstrate that thepresence of mussel shells with positive Eu anomalies is confined toareas with high-temperature hydrothermal activity at or below theseafloor.
6. Conclusions
Our results suggest that carbonate shells of the Mytilidae musselfamily are useful archives of the physico-chemical conditions in themussels' habitat, although a shell does form from the extrapallial fluidof a mussel and not directly from ambient seawater. Bathymodiolusshells from all three hydrothermally active study sites along the MARrecorded positive Eu anomalies in their REYSN patterns, which aresimilar to those of diffuse hydrothermal fluids venting at the seafloorin their respective habitat. In marked contrast, littoral M. edulis shellsfrom hydrothermally inactive locations, which we studied forcomparison, do not show any Eu anomaly. This lack of positive Euanomalies is consistent with and reflects the absence of Eu anomaliesfrom oxic seawater.
The positive Eu anomaly measured in marine hydrothermal fluidsdevelops during water–rock interaction at temperatures above atleast 200 °C, when Eu is reduced and hence decoupled from its strictlytrivalent and smaller REY neighbours. As temperature falls, divalentEu is re-oxidized and again behaves similar to the other REY duringvital processes and biomineralization, i.e. during REY uptake into themussel, during processes within the organism, and during REYincorporation from the extrapallial fluid into the carbonate shell.Hence, the Eu anomaly in a carbonate mussel shell is the same as thatin ambient water. The water in a mussel's habitat can only display apositive Eu anomaly when it has received REY input from hydrother-mal fluids which had been involved in high-temperature water–rockinteraction. The presence of positive Eu anomalies in REY patterns ofBathymodiolus shells may, therefore, be used as a tracer for hidden orfossil high-temperature hydrothermal systems for which high-tem-perature fluids are no longer available or were never vented at theseafloor.
Supplementarymaterials related to this article can be found onlineat doi:10.1016/j.epsl.2010.09.011.
Acknowledgements
We thank Jule Mawick and Daniela Meißner for their help in theGeochemistry Lab of Jacobs University Bremen and ChristianBorowski (MPI for Marine Microbiology, Bremen) for providingsome of the Bathymodiolusmussel shells. We acknowledge the skillfulequipment handling of the captains and crews of research vesselsMeteor, Merian and l'Atalante, and of the ROV teams involved. Thepaper benefited from a very detailed review by P. Craddock and that ofan anonymous second EPSL reviewer, and from the editorial skills of P.Delaney. This work was funded by the DFG (German ScienceFoundation) as part of special priority program 1144. This is SPP1144 contribution no. 53.
References
Akagi, T., Hashimoto, Y., Fu, F.F., Tsuno, H., Tao, H., Nakano, Y., 2004. Variation of thedistribution coefficients of rare earth elements in modern coral-lattices: speciesand site dependencies. Geochim. Cosmochim. Acta 68, 2265–2273.
Bau, M., 1991. Rare earth element mobility during hydrothermal and metamorphicfluid–rock interaction and the significance of the oxidation state of europium.Chem. Geol. 93, 219–230.

316 M. Bau et al. / Earth and Planetary Science Letters 299 (2010) 310–316
Bau, M., Dulski, P., 1996. Anthropogenic origin of positive gadolinium anomalies in riverwaters. Earth Planet. Sci. Lett. 143, 245–255.
Bau, M., Dulski, P., 1999. Comparing yttrium and rare earths in hydrothermal fluidsfrom the Mid-Atlantic Ridge: implications for Y and REE behaviour during near-vent mixing and for the Y/Ho ratio of Proterozoic seawater. Chem. Geol. 155, 77–90.
Bau, M., Möller, P., 1992. Rare-earth element fractionation in metamorphogenichydrothermal calcite, magnesite and siderite. Mineral. Petrol. 45, 231–246.
Bau, M., Dulski, P., Möller, P., 1995. Yttrium and holmium in South Pacific seawater:vertical distribution and possible fractionation mechanisms. Chem. Erde -Geochemistry 55, 1–15.
Bourgoin, B.P., 1988. A rapid and inexpensive technique to separate the calcite andnacreous layers in Mytilus edulis shells. Mar. Environ. Res. 25, 125–129.
Craddock, P.R., Bach, W., Seewald, J.S., Rouxel, O.J., Reevesa, E., Tivey, M.K., 2010. Rareearth element abundances in hydrothermal fluids from the Manus Basin. PapuaNew Guinea: indicators of sub-seafloor hydrothermal processes in back-arc basins.Geochim. Cosmochim. Acta 74, 5494–5513.
Cravo, A., Foster, P., Almeida, C., Bebianno, M.J., Company, R., 2008. Metal concentra-tions in the shell of Bathymodiolus azoricus from contrasting hydrothermal ventfields on the Mid-Atlantic Ridge. Mar. Env. Res. 65, 338–348.
Douville, E., Bienvenu, P., Charlou, J.L., Donval, J.P., Fouquet, Y., Appriou, P., Gamo, T.,1999. Yttrium and rare earth elements in fluids from various deep-seahydrothermal systems. Geochim. Cosmochim. Acta 63, 627–643.
Dulski, P., 2001. Reference materials for geochemical studies: new analytical data byICP-MS and critical discussion of reference values. Geostand. Newslett. 25, 87–125.
Elderfield, H., 1988. The oceanic chemistry of the rare-earth elements. R. Soc. Lond., Ser.A 325, 105–126.
Feng, D., Chen, D., Liang, Q., Roberts, H.H., 2008. Petrographic and geochemicalcharacterization of seep carbonate from Alaminos Canyon, Gulf of Mexico. Chin. Sci.Bull. 53, 1716–1724.
Feng, D., Chen, D., Peckmann, J., 2009. Rare earth elements in seep carbonates as tracersof variable redox conditions at ancient hydrocarbon seeps. Terra Nova 21, 49–56.
Garbe-Schönberg, D., Koschinsky, A., Ratmeyer, V., Jähmlich, H., Westernströer, U.,2006. KIPS—a newmultiport valve-based all-teflon fluid sampling system for ROVs.Geophys. Res. Abs. 8 EGU2006-A-07032.833.
Goldberg, E.D., 1986. The Mussel Watch concept. Environ. Monit. Assess. 7, 91–103.Haase, K.M., Petersen, S., Koschinsky, A., Seifert, R., Devey, C.W., Keir, R., Lackschewitz,
K.S., Melchert, B., Perner, M., Schmale, O., Süling, J., Dubilier, N., Zielinski, F.,Fretzdorff, S., Garbe-Schönberg, C.D., Westernströer, U., German, C.R., Shank, T.M.,Yoerger, D., Giere, O., Kuever, J., Marbler, H., Mawick, J., Mertens, C., Stöber, U.,Walter, M., Ostertag-Henning, C., Paulick, H., Peters, M., Strauss, H., Sander, S.,Stecher, J., Warmuth, M., Weber, S., 2007. Young volcanism and relatedhydrothermal activity at 5°S on the slow-spreading southern Mid-Atlantic Ridge.Geochem. Geophys. Geosyst. 8, Q11002. doi:10.1029/2006GC001509.
Haase, K.M., Koschinsky, A., Petersen, S., Devey, C.W., German, C., Lackschewitz, K.S.,Melchert, B., Seifert, R., Stecher, J., Giere, O., Paulick, H., and the M64/1, M68/1 andM78/2 Scientific Parties, 2009. Diking, young volcanism and diffuse hydrothermalactivity on the southern Mid-Atlantic Ridge: the Lilliput field at 9°33′S. Mar. Geol.266, 52–64.
Haley, B.A., Klinkhammer, G.P., Mix, A.C., 2005. Revisiting the rare earth elements inforaminiferal tests. Earth Planet. Sci. Lett. 239, 79–97.
Himmler, T., Bach, W., Bohrmann, G., Peckmann, J., 2010. Rare earth elements inauthigenic methane-seep carbonates as tracers for fluid composition during earlydiagenesis. Chem. Geol. 277, 126–136.
Hongo, Y., Nozaki, Y., 2001. Rare earth element geochemistry of hydrothermal depositsand Calyptogena shell from the Iheya Ridge vent field, Okinawa Trough. Geochem. J.35, 347–354.
James, R.H., Elderfield, H., 1996. The chemistry of ore-forming fluids and mineralprecipitation rates in an active hydrothermal sulphide deposit on the Mid-AtlanticRidge. Geology 24, 1147–1150.
Koide, M., Lee, D.S., Goldberg, E.D., 1982. Metal and transuranic records inmussel shells,byssal threads and tissues. Estuar. Coastal Shelf Sci. 15, 679–695.
Koschinsky, A., Billings, A., Devey, C.W.,Dubilier,N., Duester, A., Edge,D., Garbe-Schönberg,D., German, C., Giere, O., Keir, R., Lackschewitz, K., Mai, H.A., Marbler, H., Mawick, J.,Melchert, B., Mertens, C., Peters, M., Sander, S., Schmale, O., Schmidt, W., Seifert, R.,Seiter, C., Stöber, U., Suck, I.,Walter, M.,Weber, S., Yoerger, D., Zarrouk,M., Zielinski, F.,
2006. Discovery of newhydrothermal vents on the southernMid-Atlantic Ridge (4°S–10°S) during cruise M68/1. InterRidge News 15, 9–15.
Koschinsky, A., Garbe-Schönberg, D., Sander, S., Schmidt, K., Gennerich, H.H., Strauss, H.,2008. Hydrothermal venting at pressure-temperature conditions above the criticalpoint of seawater, 5°S on the Mid-Atlantic Ridge. Geology 36, 615–618.
Kulaksiz, S., Bau, M., 2007. Contrasting behaviour of anthropogenic gadolinium andnatural rare earth elements in estuaries and the gadolinium input into the NorthSea. Earth Planet. Sci. Lett. 260, 361–371.
McLennan, S.M., 1989. Rare earth elements in sedimentary rocks; influence ofprovenance and sedimentary processes. Rev. Mineral. Geochem. 21, 169–200.
Michard, A., Albarede, F., Michard, G., Minster, J.F., Charlou, J.L., 1983. Rare-earthelements and uranium in high-temperature solutions from East Pacific Risehydrothermal vent field (13°N). Nature 303, 795–797.
Mills, R.A., Elderfield, H., 1995. Rare earth element geochemistry of hydrothermaldeposits from the active TAG mound, 26°N. Mid-Atlantic Ridge. Geochim.Cosmochim. Acta 59, 3511–3524.
Misogianes, M.J., Chasteen, N.D., 1979. A chemical and spectral characterization of theextrapallial fluid of Mytilus edulis. Anal. Biochem. 100, 324–334.
Perner, M., Seifert, R., Weber, S., Koschinsky, A., Schmidt, K., Strauss, H., Peters, M.,Haase, K., Imhoff, J.F., 2007. Microbial CO2 fixation and sulfur cycling associatedwith low-temperature emissions at the Lilliput hydrothermal field, southern Mid-Atlantic Ridge (9°S). Environ. Microbiol. 9, 1186–1201.
Puente, X., Villares, R., Carral, E., Carballeira, A., 1996. Nacreous shell of Mytilusgalloprovincialis as a biomonitor of heavy metal pollution in Galiza (NW Spain).Science of The Total Environment 183, 205–211.
Schmidt, K., Koschinsky, A., Garbe-Schönberg, D., de Carvalho, L., Seifert, R., 2007.Geochemistry of hydrothermal fluids from the ultramafic-hosted Logatchevhydrothermal field, 15°N on the Mid-Atlantic Ridge: temporal and spatialinvestigation. Chem. Geol. 242, 1–21.
Schmidt, K., Garbe-Schönberg, D., Bau, M., Koschinsky, A., 2010. Rare earth elementdistribution in N400 °C hot hydrothermal fluids from 5°S. MAR: controls on anomalousand highly variable distribution patterns. Geochim. Cosmochim. Acta 74, 4058–4077.
Schmidt, K., Garbe-Schönberg, D., Koschinsky, A., Strauss, H., Jost, C., Klevenz, V.,Königer, P., in press. Fluid elemental and stable isotope composition of theNibelungen hydrothermal field (8°18′S, Mid-Atlantic Ridge): constraints on fluid–rock interaction in heterogeneous lithosphere. Chem. Geol. doi:10.1016/j.chemgeo.2010.07.008.
Shabani, M.B., Akagi, T., Masuda, A., 1992. Preconcentration of trace rare-earth elementsin seawater by complexation with bis(2-ethylhexyl) hydrogen phosphate and 2-ethylhexyl dihydrogen phosphate adsorbed on a C18 cartridge and determinationby inductively coupled plasma mass spectrometry. Anal. Chem. 64, 737–743.
Sholkovitz, E., Shen, G.T., 1995. The incorporation of rare earth elements in moderncoral. Geochim. Cosmochim. Acta 59, 2749–2756.
Sverjensky, D.A., 1989. Europium redox equilibria in aqueous solution. Earth Planet. Sci.Lett. 67, 70–78.
Thébault, J., Chauvaud, L., Clavier, J., Guarini, J., Dunbar, R.B., Fichez, R., Mucciarone, D.A.,Morize, E., 2007. Reconstruction of seasonal temperature variability in the tropicalPacific Ocean from the shell of the scallop Comptopallium radula. Geochim.Cosmochim. Acta 71, 918–928.
van de Flierdt, T., Robinson, L.F., Adkins, J.F., 2010. Deep-sea coral aragonite as arecorder for the neodymium isotopic composition of seawater. Geochim.Cosmochim. Acta 74, 6014–6032.
Van Dover, C.L., 2000. The Ecology of Deep-sea Hydrothermal Vents. PrincetonUniversity Press, Princeton, NJ. 424 pp.
Vance, D., Burton, K.W., 1999. Neodymium isotopes in planktonic foraminifera: a recordof the response of continental weathering and ocean circulation rates to climatechange. Earth Planet. Sci. Lett. 173, 365–379.
Wanamaker, A.D., Kreutz, K.J., Wilson, T., Borns jr., H.W., Introne, D.S., Feindel, S., 2008.Experimentally determinedMg/Ca and Sr/Ca ratios in juvenile bivalve calcite forMytilusedulis: implications for paleotemperature reconstructions. Geo-Mar. Lett. 28, 359–368.
Webb, G.E., Kamber, B.S., 2000. Rare earth elements in Holocene reefal microbialites: anew shallow seawater proxy. Geochim. Cosmochim. Acta 64, 1557–1565.
Yin, Y., Huang, J., Paine, M.L., Reinhold, V.N., Chasteen, N.D., 2005. Structuralcharacterization of the major extrapallial fluid protein of the mollusc Mytilusedulis: implications for function. Biochemistry 44, 10720–10731.
Copyright © 2022 FDOKUMEN




















