
Putative Mineral-Specific Proteins Synthesized by the Metal Reducing Bacterium Shewanella oneidensis

Biochimica et Biophysica Acta 1844 (2014) 2229–2240
Contents lists available at ScienceDirect
Biochimica et Biophysica Acta
j ourna l homepage: www.e lsev ie r .com/ locate /bbapap
Proteomic analysis of the adaptation towarming in the Antarctic bacteriaShewanella frigidimarina
Laura García-Descalzo a, Eva García-López a, Alberto Alcázar b, Fernando Baquero a,c, Cristina Cid a,⁎a Centro de Astrobiologia (CSIC-INTA), 28850 Torrejón de Ardoz, Spainb Department of Investigation, Hospital Ramon y Cajal, 28034 Madrid, Spainc Department of Microbiology, Hospital Ramon y Cajal, 28034 Madrid, Spain
Abbreviations: AHR, alkyl hydroperoxide reducsiderophore receptor; AP, alkaline phosphatase; EF-Tu, elotion factor Ts; S1, 30S ribosomal protein S1; Hyp2225, hDHLD, dihydrolipoamide dehydrogenase; OmpK, outer msuccinyl-CoA synthetase subunit beta; rrf, ribosome recyclmutase; FATP, long-chain fatty acid transport proteinnucleotidyltransferase⁎ Corresponding author at: Centro de Astrobiología (
28850 Torrejón de Ardoz, Madrid, Spain. Tel.: +34 91 520E-mail address: [email protected] (C. Cid).
http://dx.doi.org/10.1016/j.bbapap.2014.08.0061570-9639/© 2014 Elsevier B.V. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 27 March 2014Received in revised form 6 August 2014Accepted 11 August 2014Available online 19 August 2014
Keywords:ExtremophilePsychrophileShewanellaAntarcticaThermal adaptationHeat shock protein
Antarctica is subjected to extremely variable conditions, but the importance of the temperature increase in coldadapted bacteria is still unknown. To study themolecular adaptation towarming of Antarctic bacteria, cultures ofShewanella frigidimarinawere incubated at temperatures ranging from0 °C to 30 °C, emulating themost extremeconditions that this strain could tolerate. A proteomic approach was developed to identify the soluble proteinsobtained from cells growing at 4 °C, 20 °C and 28 °C. The most drastic effect when bacteria were grown at28 °C was the accumulation of heat shock proteins as well as other proteins related to stress, redox homeostasisor protein synthesis and degradation, and the decrease of enzymes and components of the cell envelope. Further-more, two main responses in the adaptation to warm temperature were detected: the presence of diverse iso-forms in some differentially expressed proteins, and the composition of chaperone interaction networks at thelimits of growth temperature. The abundance changes of proteins suggest thatwarming induces a stress situationin S. frigidimarina forcing cells to reorganize their molecular networks as an adaptive response to these environ-mental conditions.
© 2014 Elsevier B.V. All rights reserved.
1. Introduction
The discovery of cold-tolerant microorganisms in permanently fro-zen environments has broadened the known range of environmentalconditions which support microbial life. The completion of their ge-nomes has facilitated the monitoring of the global molecular changesand the identification of protein biomarkers for cells undergoing a stressresponse. When compared to other known microorganisms, cold-adapted microorganisms show many unique qualities and molecularmechanisms that allow their adaptation to environment and might beexploited for biotechnology [1,2]. These strategies include polyunsatu-rated fatty acids in their plasmamembranes. Someof themuse enzymesthat continue to function at near freezing ambient temperatures. Addi-tionally, psychrophilic bacteria are able to produce proteins that are
tase; TonB, TonB-dependentngation factor Tu; EF-Ts, elonga-ypothetical protein Sfri_2225;embrane protein OmpK; SCoA,ing factor; SOD, superoxide dis-; PNPase, polyribonucleotide
CSIC-INTA), Ctra. Ajalvir, km 4,5455; fax: +34 91 520 10 74.
stable at cold temperatures such as heat shock proteins and coldshock proteins.
Antarctica is one of the regions in theworld that has experienced thelargest increase in temperature in recent decades: 0.56 °C per decadebetween 1951 and 2000 [3]. Climatic changes do not only result inhigher overall temperature, but also in greater variability in weatherconditions [4]. Environmental conditions, such as freeze–thaw cycles,appear to have also profound effects on microbial microorganisms.
Until now, several transcriptomics and proteomics approaches havebeen achieved on polar microorganisms to investigate gene expressionchanges to temperature. It has been studied howpsychrophiles respondto sudden heat shock or cold shock, and how cold-adaptedmicroorgan-isms can cope with cold [5–8]. Some examples are the studies usingproteomics about the adaptation to cold of the Antarctic bacteriaPseudoalteromonas haloplanktis [5], Pseudomonas syringae [6] andOleispira antarctica [7] or about the cold-adapted archaeon Metha-nococcoides burtonii [8]. Nevertheless, the tolerance to warmtemperatures in cold-adapted microorganisms and their molecularmechanisms of adaptation have not been as extensively investigated.The response at the upper limit of growth temperature has been studiedby proteomics in a transgenic strain of Escherichia coli [9], in the Antarc-tic archaeon M. burtonii [10] and in the Arctic bacterium Sphingopyxisalaskensis [11].
Despite all these studies there are still many interesting questions toanswer: What is the upper limit of temperature for S. frigidimarina?

2230 L. García-Descalzo et al. / Biochimica et Biophysica Acta 1844 (2014) 2229–2240
Several known strategies of adaptation to cold or warm temperaturesare already known, mainly those involving Hsps and Csps, but whichother proteins take part in these mechanisms? S. frigidimarina, a bacte-rial species that was originally isolated from the Antarctic coastal siteshas been considered in this research as it is a good example of anAntarc-tic microorganism that thrives in particularly harsh environmentalconditions. The experimental approach consisted on cultures ofS. frigidimarina incubated at different temperatures, ranging from 0 °Cto 30 °C, emulating the most extreme conditions that this strain cantolerate. In order to gather information regarding the adaptation towarming of this bacterium and its molecular mechanisms of adaptationa comparative proteomic approach using two-dimensional fluorescenceDifference Gel Electrophoresis (2-D DIGE) and matrix-assisted laserdesorption/ionization-time of flight mass spectrometry (MALDI-TOF–TOF MS) analysis was carried out.
2. Materials and methods
2.1. Bacterial strains and growth conditions
Bacteria included in this study were purchased from culturecollections (ATCC®). Broth cultures of S. frigidimarina (ACAM 591,700753™) were prepared as follows: 500-μl aliquots of mid-exponential grown cultures were transferred to 500-ml Erlenmeyerflasks containing 100 ml fresh, temperature-equilibrated medium (Ma-rine broth 2216). Theflaskswere incubated aerobicallywith agitation at150 rpm, at 0 °C, 4 °C, 12 °C, 20 °C, 28 °C or 30 °C for each conditionrespectively, until an absorbance (OD600) of 0.3–0.4 had been attained.
0.000
0.200
0.400
0.600
0.800
1.000
1.200
1.400
1.600
1.800
0 20 40 60
O.D
. 600
nm
4ºC 20ºC 284ºC 20º0ºC
Tim
O.D
. 600
nm
4ºC 20ºC 284ºC 20º0ºC 4ºC 20º0ºC
St 0ºC
97 -66 -45 -
30 -
14 -
4ºC
---
-
-
A
B
Fig. 1. Cell cultures of S. frigidimarina. (A) Growth rates. Cell cultures aerobically incubated at 0 °Cell extracts from cultures incubated at 0 °C, 4 °C, 12 °C, 20 °C and 28 °C were analyzed by SD
Four biological replicates were grown. The cells were then harvested bycentrifugation (10,000 ×g, 15 min), rinsed in PBS and stored at−20 °Cuntil use.
All general chemicals were purchase from Sigma unless statedotherwise in parenthesis. The chemicals used in isoelectric focusing(IEF) and in sodium dodecyl sulfate-polyacrylamide gel electrophoresis(SDS-PAGE) were purchase from GE Healthcare.
2.2. Preparation of cell extracts and protein determination
Bacteria from each biological replicate (four in total) were obtainedfrom the individual cultures. The cells were harvested, washed andlysed in buffer A (20mMTris–HCl, pH 7.6; 140mMpotassium chloride;2 mM benzamidine; 1 mM EDTA; 10 μg/ml pepstatin A, leupeptin andantipain), using a French Press at 4 °C. Cell debris was removed by cen-trifugation at 11,000×g for 10min to obtain a supernatant, and the pro-tein extracts were processed using a 2-D Clean-Up kit (GE Healthcare,Spain). The pellet was frozen and stored at−80 °C. The protein contentwas determined in cell extracts using the BioRad protein assay based onthe Bradford method using different dilutions of BSA as the standard.The samples were adjusted to a protein concentration of 5–10 mg/ml.All steps were carried out at 4 °C.
2.3. Two-dimensional electrophoresis
Horizontal slab gel isoelectric focusing (IEF) was combined withSDS-PAGE for 2-DE by using the Multiphor II apparatus for the firstdimension and standard vertical slab gel electrophoresis for the second
100 120 140
ºC 30ºC 0ºCC 28ºC 30ºC
80
e (h)
ºC 30ºC 0ºCC 28ºC 30ºCC 28ºC 30ºC
12ºC 20ºC 28ºC St
C, 4 °C, 20 °C, 28 °C and 30 °Cweremonitored by optical density at 600 nm. (B) SDS-PAGE.S-PAGE. Protein pattern is very different between 20 °C and 28 °C.

2231L. García-Descalzo et al. / Biochimica et Biophysica Acta 1844 (2014) 2229–2240
dimension, as previously described, with some modifications [12,13].Samples (about 150 μg of protein) were prepared in 7M urea/2 M thio-urea and 5%β-mercaptoethanol, centrifuged and applied to pH gradientstrips for IEF. Carrier ampholyte urea IEF was performed usingimmobilized pH 3–10 and pH 3–11 non-linear gradient strips (18 and24 cm respectively).
After the first dimension, the IEF stripswere equilibrated for the sec-ond dimension in two successive SDS equilibration buffer solutions con-taining Tris buffer 100 mM, urea 6 M and glycerol 30% (w/v). The firstbuffer solution contained DTT 0.5% (w/v) to reduce sulfhydryl groups.Secondly, in order to alkylate sulfhydryl groups, they were incubatedwith iodoacetamide 4.5% (w/v).
The second dimension in SDS-PAGEwas carried out on 12% acrylam-ide (2.6% crosslinking) gels (1.0 mm thick). Gels were stained withCoomassie Blue or with MALDI-MS-compatible silver reagent for pro-tein identification [14].
2.4. Two-dimensional fluorescence difference gel electrophoresis
Proteins from bacterial cells grown at 4 °C, 20 °C or 28 °C were ana-lyzed by 2-D DIGE experiments, considering 20 °C as control tempera-ture to compare with 4 °C or 28 °C. Proteins were extracted fromdifferent cultures to obtain four biological replicates. Samples (40 μg)from each experimental condition — extreme temperatures (4 °C or28 °C) and control temperature (20 °C) — were made to 7 M urea/2 Mthiourea, centrifuged, and labeled with Cy5 or Cy3 CyDye DIGE Fluorminimal dyes (GE Healthcare); according to the Ettan DIGE protocol. Apooled standard of 4 °C + 20 °C or 28 °C + 20 °C was labeled with
sample preparaShewanella frigidim
Incubate
4º C
cell extr
2-D DIGE
DIA(Differential In-gel Analysis)
Cy2 Cy3Cy5
combine
fluorescence scan *
IEF/SDS-PAGE
* Spots matched accurately
4º C/28ºC 4º C/28ºC + 20ºC 20º C
20º C
pooled standard control temp.extreme temp.
sample preparaShewanella frigidim
Incubate
4º C
cell extr
2-D DIGE
DIA(Differential In-gel Analysis)
Cy2 Cy3Cy5
combine
fluorescence scan *
IEF/SDS-PAGE
* Spots matched accurately
4º C/28ºC 4º C/28ºC + 20ºC 20º C
20º C
pooled standard control temp.extreme temp.
sample preparaShewanella frigidim
Incubate
4º C
cell extrcell extr
2-D DIGE
DIA(Differential In-gel Analysis)
Cy2 Cy3Cy5
combinecombine
fluorescence scan *
IEF/SDS-PAGE
* Spots matched accurately
4º C/28ºC 4º C/28ºC + 20ºC 20º C
20º C
pooled standard control temp.extreme temp.
A
B
Fig. 2. Summary of the overall experimental strategy. Schematic representation of the experShewanella frigidimarina to warming. (A) Sample preparation. Broth cultures of S. frigidimarinaPairs of experimental and control samples were mixed and subjected to 2-DE as described in Sfrom preparative 2-DE gels and stained with Coomassie Blue or with MALDI-MS-compatible si
Cy2 (Fig. 1S). After DTT addition, pairs of experimental and control sam-ples were mixed and subjected to 2-DE as described above.
2.5. Biological variation analysis
Immediately after the 2-DE, CyDye-labeled protein gels were im-agedwithin the glass cassettes using a Typhoon 9400 variablemode im-ager (GE Healthcare) at 100 μm resolution. The resultant gel imageswere processed and analyzed using the DeCyder Differential In-gelAnalysis (DIA) software version 7.0 (GE Healthcare) (Fig. 2S). In DIGEimages, the spots were co-detected, quantified, normalized andmatched between the four replicate experiments.
Protein abundance changes between the experimental groups (4 °C,20 °C or 28 °C) were examined by ANOVA test. Spots present in four offour gels per group, with significant ANOVA test (p b 0.05) and an aver-aged ratio ≥ ±2 with respect to control samples (20 °C), were consid-ered and selected for a further MS analysis in preparative gels.
2.6. Protein identification by mass spectrometry
Protein spots were excised manually from the Coomassie or silverstained 2-DE gels, de-stained and then digested automatically using aProteineer DP protein digestion station (Bruker-Daltonics, Bremen,Germany). The identification protocol used was based on the methodpreviously described in with minor variations [12]. An aliquot of theabove digestion solution was mixed with an aliquot of α-cyano-4-hydroxycinnamic acid (Bruker-Daltonics) in 33% aqueous acetonitrileand 0.1% trifluoroacetic acid. This mixture was deposited onto aMALDI probe and allowed to dry at room temperature. For MALDI-
tion (4 replicates)arina ACAM591d at
28º C
acts
preparative 2-DE
IEF/SDS-PAGE
Coomassie orsilver stain *
in-gel digest
MALDI-TOF-TOF MS
identification(Mascot)
4º C 28º C20º C
tion (4 replicates)arina ACAM591d at
28º C
acts
preparative 2-DE
IEF/SDS-PAGE
Coomassie orsilver stain *
in-gel digest
MALDI-TOF-TOF MS
identification(Mascot)
4º C 28º C20º C
tion (4 replicates)arina ACAM591d at
28º C
actsacts
preparative 2-DE
IEF/SDS-PAGE
Coomassie orsilver stain *
in-gel digest
MALDI-TOF-TOF MS
identification(Mascot)
4º C 28º C20º C
C
imental design followed to identify differentially expressed proteins in the response ofaerobically grown at 4 °C, 20 °C and 28 °C and preparation of cell extracts. (B) 2-D DIGE.ection 2.4. and in Fig. 1S. (C) Preparative 2-DE. Protein identification of the spots resolvedlver reagent.

Table 1Identification of total soluble proteins, isolated from the psychrophilic bacteria S. frigidimarina grown at 4 °C, 20 °C and 28 °C.
Proteins showing increased abundance at 28 °C
N° Protein Th/AppaMw Th/App pI Accession N° in NCBI Score b Fold change
28 °C/20 °C 20 °C/4 °C 28 °C/4 °C N C
Chaperone and stress proteins205 Chaperone protein DnaK 69/69 4.7/4.6 YP_749660 116 2.4 3.7 6.95 10/5 21215 Chaperone protein DnaK 69/69 4.7/4.7 YP_749660 92 2.10 3.12 6.17 65/11 42224 Chaperone protein DnaK 69/69 4.7/4.7 YP_749660 140 2.02 3.16 6.16 65/13 431398 GrpE protein 22/25 4.4/4.4 YP_751470 138 2.70 4.51 8.24 15/7 19377 Chaperonin GroEL 57/57 4.8/5.0 YP_752326 101 2.1 3.04 5.14 40/6 32387 Chaperonin GroEL 57/57 4.8/4.9 YP_752326 282 3.05 6.15 9.15 65/11 511888 Co-chaperonin GroES 11/11 5.8/5.8 YP_752327 120 8.9 17.35 26 46/10 951418 Phage shock protein A, PspA 26/26 5.4/5.4 YP_751257 137 0.98 1.87 2.92 30/13 49
Protein synthesis152 Elongation factor G 77/75 5.0/5.2 YP_749249 148 1.4 2.82 4.24 15/5 6701 Elongation factor Tu 44/44 5.1/5.1 YP_748847 90 1.93 3.81 5.81 65/10 39708 Elongation factor Tu 43/44 5.1/5.2 YP_748835 180 1.38 2.76 4.13 20/6 63711 Elongation factor Tu 44/44 5.1/5.3 YP_748847 208 1.01 2.14 3.18 65/19 721198 Elongation factor Ts 30/30 5.1/5.2 YP_749962 92 2.06 4.11 6.16 36/11 40169 GTPase-translation elongation factor 47/70 5.4/5.2 BAF_64776 295 1.50 3.40 5.03 33/13 23170 Glycyl-tRNA synthetase subunit beta 76/71 5.3/5.2 YP_748707 113 1.50 3.02 4.45 31/13 26459 tRNA modification GTPase TrmE 50/55 4.8/4.8 YP_752726 76 2.50 5.04 7.54 12/9 30
Protein degradation791 Protease Do 47/44 5.8/5.8 YP_752037 92 1.4 2.8 4.06 65/14 411578 ATP-dependent Clp protease 22/22 5.7/5.8 YP_751275 97 1.27 2.61 3.82 22/10 271884 ATP-dp Clp protease adaptor ClpS 12/12 5.1/5.3 YP_750939 87 5.89 ns ns 14/11 64
Ribosomal-related proteins258 30S ribosomal protein S1 61/65 4.8/4.8 YP_750807 90 9.09 ns ns 22/13 30266 30S ribosomal protein S1 61/65 4.9/4.9 YP_750807 81 6.49 ns ns 42/11 24
Redox homeostasis1519 Alkyl hydroperoxide reductase 22/23 5.2/5.2 YP_750145 223 7.17 ns ns 23/11 411522 Alkyl hydroperoxide reductase 22/23 5.2/5.2 YP_750145 222 5.94 ns ns 20/9 342022 Alkyl hydroperoxide reductase 22/22 5.2/5.2 YP_750145 319 3.00 8.34 11.34 35/13 411767 Ferritin, Dps family protein 18/18 5.8/6.1 YP_750527 173 9.21 18.10 27.15 15/4 731773 Ferritin, Dps family protein 18/18 5.8/5.8 YP_750527 196 8.22 16.13 24.2 22/5 231985 Cytochrome c3 11/11 4.7/5.2 2K3V_A 89 6.78 ns ns 12/4 74
Energy pathway1409 5′-nucleotidase surE 26/25 5.0/5.2 YP_749748 98 5.37 ns ns 10/9 341832 Nucleoside-diphosphate kinase 15/15 5.4/5.3 YP_751100 101 1.12 2.23 3.34 19/8 38
Cell motility805 Flagellar hook-associated protein FlgL 42/42 4.4/4.4 YP_749862 141 1.2 1.34 3.49 40/18 32
Other functions1278 GumN family protein 33/28 5.1/4.9 YP_752191 84 6.1 ns ns 15/7 331220 Hypothetical protein Sfri_0844 33/30 4.8/4.8 YP_749535 182 1.8 3.6 5.41 23/10 351598 Hypothetical protein Sfri_0823 24/20 4.6/4.5 YP_749516 88 13.65 ns ns 20/9 301708 Hypothetical protein Sfri_3876 20/19 9.6/10 YP_752540 104 4.79 ns ns 15/7 33
Unknown proteins1688 Unknown protein /19 /4.5 – 5.35 ns ns1691 Unknown protein /19 /4.7 – 10.7 ns ns1727 Unknown protein /19 /6.1 – 30.06 ns ns1730 Unknown protein /19 /5.9 – 16.59 ns ns1830 Unknown protein /15 /5.4 – 2.25 ns ns1974 Unknown protein /10 /5.3 – 5.14 ns ns
Proteins showing decreased abundance at 28 °C
N° Protein Th/App a
MwTh/ApppI
Accession N° in NCBI Score b Fold change
28 °C/20 °C 20 °C/4 °C 28 °C/4 °C
Cold-acclimation proteins420 Trigger factor 48/58 4.7/4.7 YP_751276 72 −1.6 −2.2 −4.8 8/7 24
Cell envelope188 TonB-dependent siderophore receptor 79/75 4.5/4.4 YP_752515 153 −2.00 3.61 −5.61 30/20 26189 TonB-dependent siderophore receptor 79/75 4.5/4.4 YP_752515 172 −2.38 −4.5 −7.15 14/12 74195 TonB-dependent siderophore receptor 79/75 4.5/4.4 YP_752515 112 −3.00 −7.6 −10.63 16/8 32783 Outer membrane channel protein 47/44 4.9/4.9 YP_752007 153 −2.80 −5.71 −8.48 65/20 431020 Porin 38/38 4.6/4.8 YP_749202 138 −0.93 −1.90 −2.78 20/8 331031 Porin 38/38 4.6/4.4 YP_749202 118 −1.27 −2.54 −3.81 53/13 381033 Porin 38/38 4.6/5.1 YP_749202 138 −0.90 −1.87 −2.8 24/12 431051 Porin 34/34 4.5/4.5 YP_751448 151 −5.74 9.1 −15.87 26/13 421052 Porin 34/34 4.5/4.4 YP_751448 124 −30.95 −90.95 −114.95 65/18 47
2232 L. García-Descalzo et al. / Biochimica et Biophysica Acta 1844 (2014) 2229–2240

Table 1 (continued)
Proteins showing increased abundance at 28 °C
N° Protein Th/AppaMw Th/App pI Accession N° in NCBI Score b Fold change
28 °C/20 °C 20 °C/4 °C 28 °C/4 °C N C
Redox homeostasis634 Glutathione reductase 48/48 5.6/5.8 YP_752541 171 −1.20 −2.49 −3.73 22/10 211602 Alkyl hydroperoxide reductase 22/20 5.2/5.3 YP_750145 99 −12.69 −25.49 −38.07 65/12 682023 Alkyl hydroperoxide reductase 22/22 5.2/5.4 YP_750145 137 −4.75 −8.01 −12.75 22/7 34
Energy metabolism328 Alkaline phosphatase 56/60 4.5/4.5 YP_751222 176 −15.98 −32.54 −45.00 34/11 18331 Alkaline phosphatase 56/60 4.5/4.5 YP_751222 131 −11.87 −21.33 −32.23 21/8 15385 Alkaline phosphatase 56/55 4.5/4.5 YP_751222 103 −2.33 −4.77 −6.92 17/15 26411 Alkaline phosphatase 67/55 4.32/ YP_751223 266 −7.72 −16.11 −23.24 65/22 492019 Alkaline phosphatase 56/61 4.5/4.5 YP_751222 197 47.88 −95.66 −142.81 65/22 532020 Alkaline phosphatase 67/59 4.3/4.3 YP_751223 172 20.14 −46.99 −65.17 65/22 49358 Peptidase M61 domain-cont protein 68/65 5.3/5.3 YP_751451 155 ns −9.57 ns 34/11 18951 Putative agmatine deiminase 41/41 4.6/4.6 YP_750304 89 −3.4 −6.8 −10.2 14/11 39993 Fructose-1, 6-bisphosphatase class 1 36/35 4.5/4.6 YP_749230 74 −6.03 −12.1 −18.01 12/10 32
Transcription821 Transcriptional regulator, LysR fam. 34/40 6.5/5.3 YP_749519 78 ns −4.24 ns 15/5 25
Other functions278 Hypothetical protein Sfri_0573 80/70 5.5/5.3 YP_749272 158 −5.91 −11.01 −16.91 32/21 31288 Hypothetical protein Sfri_2225 75/68 4.4/4.3 YP_750909 124 −1.7 −4.12 −5.7 45/20293 Hypothetical protein Sfri_2225 75/68 4.4/4.3 YP_750909 141 −2.50 −5.21 −7.71 65/22 361689 Hypothetical protein Sfri_2875 20/20 5.9/5.3 YP_751553 139 −8.22 −16.51 −24.66 45/13 52
Unknown proteins1117 Unknown protein /32 /4.4 – ns −8.41 ns
a Abbreviations used: Theoreticalmolecularweight (ThMw), apparentmolecularweight (AppMw), theoretical isoelectric point (ThpI), apparent isoelectric point (ApppI), non-detectablespot at 4 °C or 28 °C (ns), number of mass values searched/number of mass values matched (N), % sequence coverage (%C).
b Protein scores higher than 72 were significant (p b 0.05) in the Mascot database search algorithm (Matrix science, UK).
2233L. García-Descalzo et al. / Biochimica et Biophysica Acta 1844 (2014) 2229–2240
TOF/TOF analysis, samples were automatically acquired in an ABi 4800MALDI TOF/TOF mass spectrometer (Applied Biosystems) in positiveion reflector mode (the ion acceleration voltage was 25 kV toMS acqui-sition and 1 kV to MS/MS). PMF and MS/MS fragment ion spectra weresmoothed and corrected to zero baseline using ABi 4000 Series ExplorerSoftware. Two of the strongest peaks of the TOF spectra per samplewerechosen for MS/MS analysis.
2.7. Data search
Spectral data were analyzed to search the NCBInr database using theMascot database search algorithm (Matrix Science, London, UK). Com-bined peptide mass fingerprint and MS/MS ion search modes wereperformed against the S. frigidimarina NCIMB 400 genome data bankavailable at NCBI (release date September 14, 2006; including25510175 sequences; 8788594840 residues). Only peptides and pro-teins with confidence scores of at least 95% (p b 0.05) were consideredpositive identifications.Mascot searchparameterswere tryptic peptideswith one missed cleavage, using 80 ppm and 0.3 Da for precursor andfragment tolerances, respectively. Theoretical molecular weight andisoelectric point were calculated for each protein with the ComputepI/MW tool (ExPASy, Swiss Institute of Bioinformatics, Switzerland).Apparent molecular weight and isoelectric point were obtained fromeach spot position in gels.
2.8. Immunoprecipitation of DnaK and GroEL
Cell extracts were immunoprecipitated with monoclonal anti-DnaKand anti-GroEL antibodies (Enzo Life Sciences, USA) in order to studythe DnaK- and the GroEL-associated proteins, as previously described[13]. Anti-DnaK and anti-GroEL antibodies were raised against E. coli.These antibodies also recognize these proteins in other bacterial species.In S. frigidimarina, we had previously checked the specificity of these an-tibodies with recombinant DnaK and GroEL from S. frigidimarina, pur-chased from Bionova (Spain). The detection of bands was carried out
usingHRP-linked anti-mouse IgG, followed by ECLWestern blotting de-tection reagents (GE Healthcare).
Then, DnaK or GroEL was immunoprecipitated in each sample(100 μg) with 1 μg of antibody for 4 h at 4 °C in buffer A. Protein G-Sepharose (25 μl of 50% slurry) was then added for 1 h at 4 °C andcentrifuged. The immunoprecipitation was developed in stringent con-ditions to minimize unspecific complexes. The immunoprecipitateswere washed in 1 ml of cold buffer with 0.5 M lithium chloride, andthen washed two times in 1 ml of cold buffer without lithium chloride,leaving the solid phase in thewash buffers for 10min at eachwash, andsubjected to 2-DE followed by protein identificationbyMALDI-TOF–TOFMS. Control experiments of nonspecific binding to protein G-Sepharosebeadswere run in parallel. Other control experimentswith immunopre-cipitateswere performed to check thedifferences between immunopre-cipitation at 4 °C and at room temperature. Resultswere similar in termsof specificity — the interaction partners did not change — but spots ingels from experiments at room temperature were weaker. Maybe pro-teins had been degraded somehow, although cells had been lysed inbuffer with protease inhibitors. So, all experiments were carried at 4 °C.
3. Results and discussion
3.1. Cell growth at extreme temperatures
According to previous studies, S. frigidimarina grows at temperaturesbetween 0 °C and 27 °C–28 °C, with a maximum growth rate around20–22 °C [15]. In our experiments, S. frigidimarina could be cultured attemperatures from 0 °C to 28 °C and reached absorbance values of1.4–1.6 (OD 600) (Fig. 1A), but very scarce growth occurred at 30 °C,and a change to reddish color could be observed at warm temperatures,mainly in cultures at 28 °C. Although S. frigidimarinawas able to grow attemperatures ranging from 0 °C to 28 °C we only compared 4 °C, 20 °Cand 28 °C as the solubility of culture media at 0 °C decreased and a lim-ited bioavailability of essential nutrients could alter the results aboutadaptation to temperature. Cell extracts from cultures incubated at
2234 L. García-Descalzo et al. / Biochimica et Biophysica Acta 1844 (2014) 2229–2240
0 °C, 4 °C, 12 °C, 20 °C and 28 °C were analyzed by SDS-PAGE. Proteinpattern was very different between 20 °C and 28 °C (Fig. 1B). So, 20 °Cwas considered as an intermediate temperature for comparison in 2-DDIGE gels.
A change to reddish color could be observed in cultures ofS. frigidimarina at warm temperatures. Several bacterial pigments havebeen identified in other polar bacteria such as the Antarctic bacteriumSphingobacterium antarcticus [16] or some strains of Janthinobacteriumlividum isolated from Alaskan soils [17]. In most reports, the amountsof pigments increased in cells grown at cold temperatures meanwhilein S. frigidimarina the change to reddish color is observed at warmtemperature.
3.2. Identification of proteins expressed at increasingly warmertemperatures
2-DDIGE gels fromcultures growing in temperature-equilibratedme-dium (Marine broth 2216) at 4 °C, 20 °C (maximum growth rate) and28 °C (upper temperature limit) were compared. Schematic representa-tions of the experimental design are shown in Figs. 2 and 1S. Proteinmaps produced from four independent protein extractions showed ahigh level of reproducibility. Fig. 2S represents 2D maps of soluble pro-teins extracted from the three culture conditions, in which the intensityof 75 spots was significantly altered by an averaged fold change of ±2with respect to control samples (20 °C). Among these 75 spots, the abun-dance of 41 proteins increased in cells growing under thermal stress at28 °C meanwhile the abundance of 28 proteins decreased (Table 1). Allof themwere expressed at 20 °C but someprotein spotswere only detect-able at 4 °C or 28 °C and were marked as “ns” (non-detectable spot). Atotal of 75 differentially expressed protein spots were excised frompreparative gels (Fig. 3S) and analyzed by MALDI-TOF–TOF MS. Ofthese, 69 spots were identified (Table 1 and Supplementary dataTables S2–S114).
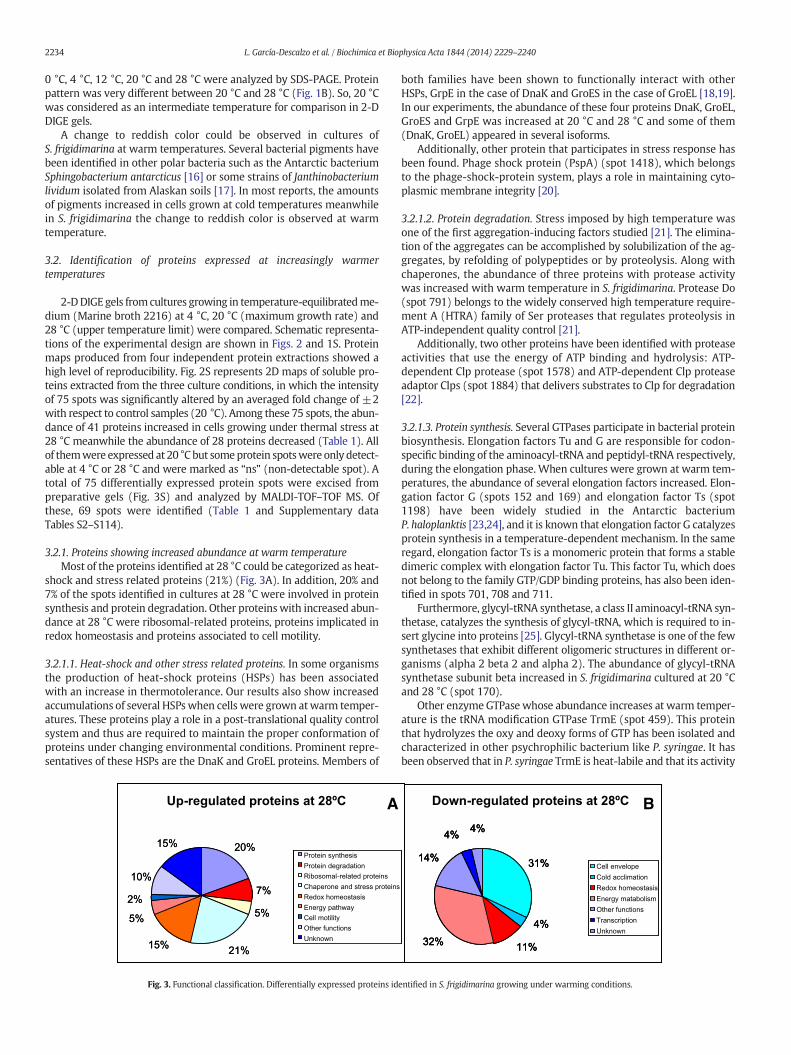
3.2.1. Proteins showing increased abundance at warm temperatureMost of the proteins identified at 28 °C could be categorized as heat-
shock and stress related proteins (21%) (Fig. 3A). In addition, 20% and7% of the spots identified in cultures at 28 °C were involved in proteinsynthesis and protein degradation. Other proteinswith increased abun-dance at 28 °C were ribosomal-related proteins, proteins implicated inredox homeostasis and proteins associated to cell motility.
3.2.1.1. Heat-shock and other stress related proteins. In some organismsthe production of heat-shock proteins (HSPs) has been associatedwith an increase in thermotolerance. Our results also show increasedaccumulations of several HSPswhen cells were grown atwarm temper-atures. These proteins play a role in a post-translational quality controlsystem and thus are required to maintain the proper conformation ofproteins under changing environmental conditions. Prominent repre-sentatives of these HSPs are the DnaK and GroEL proteins. Members of
20%
7%
5%
21%15%
5%
2%
10%
15%Protein synthesisProtein degradationRibosomal-related proteinsChaperone and stress proteinsRedox homeostasisEnergy pathwayCell motilityOther functionsUnknown
Up-regulated proteins at 28ºC
20%
7%
5%
21%15%
5%
2%
10%
15%Protein synthesisProtein degradationRibosomal-related proteinsChaperone and stress proteinsRedox homeostasisEnergy pathwayCell motilityOther functionsUnknown
Protein synthesisProtein degradationRibosomal-related proteinsChaperone and stress proteinsRedox homeostasisEnergy pathwayCell motilityOther functionsUnknown
A
Fig. 3. Functional classification. Differentially expressed proteins id
both families have been shown to functionally interact with otherHSPs, GrpE in the case of DnaK and GroES in the case of GroEL [18,19].In our experiments, the abundance of these four proteins DnaK, GroEL,GroES and GrpE was increased at 20 °C and 28 °C and some of them(DnaK, GroEL) appeared in several isoforms.
Additionally, other protein that participates in stress response hasbeen found. Phage shock protein (PspA) (spot 1418), which belongsto the phage-shock-protein system, plays a role in maintaining cyto-plasmic membrane integrity [20].
3.2.1.2. Protein degradation. Stress imposed by high temperature wasone of the first aggregation-inducing factors studied [21]. The elimina-tion of the aggregates can be accomplished by solubilization of the ag-gregates, by refolding of polypeptides or by proteolysis. Along withchaperones, the abundance of three proteins with protease activitywas increased with warm temperature in S. frigidimarina. Protease Do(spot 791) belongs to the widely conserved high temperature require-ment A (HTRA) family of Ser proteases that regulates proteolysis inATP-independent quality control [21].
Additionally, two other proteins have been identified with proteaseactivities that use the energy of ATP binding and hydrolysis: ATP-dependent Clp protease (spot 1578) and ATP-dependent Clp proteaseadaptor Clps (spot 1884) that delivers substrates to Clp for degradation[22].
3.2.1.3. Protein synthesis. Several GTPases participate in bacterial proteinbiosynthesis. Elongation factors Tu and G are responsible for codon-specific binding of the aminoacyl-tRNA and peptidyl-tRNA respectively,during the elongation phase. When cultures were grown at warm tem-peratures, the abundance of several elongation factors increased. Elon-gation factor G (spots 152 and 169) and elongation factor Ts (spot1198) have been widely studied in the Antarctic bacteriumP. haloplanktis [23,24], and it is known that elongation factor G catalyzesprotein synthesis in a temperature-dependent mechanism. In the sameregard, elongation factor Ts is a monomeric protein that forms a stabledimeric complex with elongation factor Tu. This factor Tu, which doesnot belong to the family GTP/GDP binding proteins, has also been iden-tified in spots 701, 708 and 711.
Furthermore, glycyl-tRNA synthetase, a class II aminoacyl-tRNA syn-thetase, catalyzes the synthesis of glycyl-tRNA, which is required to in-sert glycine into proteins [25]. Glycyl-tRNA synthetase is one of the fewsynthetases that exhibit different oligomeric structures in different or-ganisms (alpha 2 beta 2 and alpha 2). The abundance of glycyl-tRNAsynthetase subunit beta increased in S. frigidimarina cultured at 20 °Cand 28 °C (spot 170).
Other enzyme GTPase whose abundance increases at warm temper-ature is the tRNA modification GTPase TrmE (spot 459). This proteinthat hydrolyzes the oxy and deoxy forms of GTP has been isolated andcharacterized in other psychrophilic bacterium like P. syringae. It hasbeen observed that in P. syringae TrmE is heat-labile and that its activity
31%
4%
11%32%
14%
4% 4%
Cell envelopeCold acclimationRedox homeostasisEnergy matabolismOther functionsTranscriptionUnknown
Down-regulated proteins at 28ºC
31%
4%
11%32%
14%
4% 4%
31%
4%
11%32%
14%
4% 4%
Cell envelopeCold acclimationRedox homeostasisEnergy matabolismOther functionsTranscriptionUnknown
Cell envelopeCold acclimationRedox homeostasisEnergy matabolismOther functionsTranscriptionUnknown
B
entified in S. frigidimarina growing under warming conditions.

2235L. García-Descalzo et al. / Biochimica et Biophysica Acta 1844 (2014) 2229–2240
decreases at temperatures higher than 30 °C [26]. The high concentra-tion of elongation factors and GTPases in S. frigidimarina might repre-sent an adaptive tool to ensure an efficient nucleotide exchange evenin stress situations [27].
3.2.1.4. Ribosomal-related proteins. Surprisingly, although accumulationsof several elongation factors have been observed, the abundance of initi-ation factors did not increase. It is generally believed that prokaryotictranslation is initiated by the interaction between the Shine–Dalgarno(SD) sequence at the 5′UTR of mRNA and the anti-SD sequence at the3′end of the 16S rRNA. However, there are some alternativemechanismswhich do not require the SD sequence for translation initiation such astranslation of leaderlessmRNA that lacks its 5′UTR [28]. Other alternativemechanism involves the ribosomal protein S1 [29]. The S1 protein medi-ates the recognition and binding betweenmRNA and 30S ribosomal par-ticles, through its C-terminal domain with high affinity for the mRNApyrimidine-rich sequences and the association with 30S subunit via itsN-terminal globular domain by protein–protein interactions [29].
2000000000000
293288
205 224 215
374387
377 701 708 711
258 266
1519 15222022
20
1602
188 189195
10311052
1051 10201033
4112020385
2019328
331
DnaK
GroEL
S1TonB
Hyp2225
AP
Porin
EF-Tu
AHR
293293288288
205205 224224 215215
374374387387
377377 701701 708708 711711
258258 266266
15191519 1522152220222022
2020
16021602
188188 189189195195
1031103110521052
10511051 1020102010331033
41141120202020385385
20192019328328
331331
DnaK
GroEL
S1TonB
Hyp2225
AP
Porin
EF-Tu
AHR
A
293288
205 224 215
374387
377 701 708 711
258 266
1519 15222022
20
1602
188 189195
10311052
1051 10201033
4112020385
2019328
331
DnaK
GroEL
S1TonB
Hyp2225
AP
Porin
EF-Tu
AHR
293293288288
205205 224224 215215
374374387387
377377 701701 708708 711711
258258 266266
15191519 1522152220222022
2020
16021602
188188 189189195195
1031103110521052
10511051 1020102010331033
41141120202020385385
20192019328328
331331
DnaK
GroEL
S1TonB
Hyp2225
AP
Porin
EF-Tu
AHR
A
B
202215221519
2023
1602
1020
1052
103310311051 195
189188
387 377 2374
-160-140-120-100-80-60-40-20
0
4060
Rel
ativ
e ab
un
dan
ce o
f is
ofo
rms
AHR
Porin
TonBGroEL
pH 3pH 3
Fig. 4. Close-up of possible isoforms detected by 2-DE. (A) Soluble proteins of S. frigidimarina resare shown. The numbered spots correspond to proteins listed in Table 1. Thefigure is a represenprotein isoforms at 28 °C with respect to 4 °C. Proteins are colored according to cellular procereceptor; AP, alkaline phosphatase; EF-Tu, elongation factor Tu; S1, 30S ribosomal protein S1;
In our experiments, two isoforms (spots 258 and 266) of the ribo-somal protein S1 had increased abundance whereas other members oftranslation initiation machinery did not show changes. Although trans-lation initiation is indispensable for all protein-coding genes, its mecha-nisms can dramatically change in stress situations. So, in S. frigidimarinathe ribosomal protein S1 may play an essential role for an alternativetranslation initiation in response to high temperature stress.
3.2.1.5. Redox homeostasis. One of the proteins exceptionally induced byhigh temperature was the AHR. This protein corresponded to severalspots: 1519, 1522, 1602, 2022 and 2023. It has been described thatthis enzyme is required for protection against peroxides in E. coli andin osmotic stress in Staphylococcus aureus. In Bacillus subtilis it was iden-tified as induced in response to heat, oxidative stress or metal ion levels[30], while in Shewanella putrefaciens its activity takes also part in coldacclimation [31]. In our experiments we have found five spots corre-sponding to this protein; the abundance of three spots (1519, 1522and 2022) increased (induced by heat) while the abundance of twospots (1602 and 2023) decreased (induced by cold).
23
1773 1767
Ferritin
2323
17731773 17671767
Ferritin
23
1773 1767
Ferritin
2323
17731773 17671767
Ferritin
24215205 1773
1767
2020
411
2019
331328
385
711708701 266258
293288DnaK
FerritinAP
EF-Tu S1
Hyp2225
1111 10
150
kDa
SD
S-P
AG
E
10
150
kDa
SD
S-P
AG
E
olved by 2-DE. All 34 differentially expressed protein spotsmatched to 10 unique proteinstative of four 2-DE experiments. (B) Expression bar plot representing relative abundance ofsses as in Fig. 3. AHR, Alkyl hydroperoxide reductase; TonB, TonB-dependent siderophoreHyp2225, hypothetical protein Sfri_2225.

2236 L. García-Descalzo et al. / Biochimica et Biophysica Acta 1844 (2014) 2229–2240
Besides these proteins related to cellular stress, the activity of otherenzyme capable of providing protection to cells during exposure to se-vere environmental assaults was also increased in cultures at warmtemperature: Ferritin/DNA-binding protein from starved cells (Dps)(spots 1767 and 1773, fold scores−27 and−24 respectively). The abil-ity of Dps to provide multifaceted protection is based on three intrinsicproperties of the protein: DNA binding, iron sequestration and its fer-roxidase activity which oxidize bound ferrous ions to the ferric state[32].
Further, when the spot 1985was identified, it corresponded to cyto-chrome c3. This flavocytochrome c3, a tetrahaem periplasmic proteinwhich possesses a catalytic fumarate reductase activity, has beenwidelystudied in S. frigidimarina [33]. Its abundance increased in cultures at20 °C and 28 °C, but was not detected at 4 °C.
3.2.1.6. Energy pathway. Nucleoside-diphosphate kinase (Ndk) is an im-portant enzyme that generates nucleoside triphosphates by terminalphospho transfer from ATP or GTP to any nucleoside diphosphate andplays an important role in bacterial growth and signal transduction[34]. This protein (spot 1832) was induced in S. frigidimarina at hightemperature.
Besides this kinase, the stationary phase survival protein SurE,whichis a metal ion-dependent phosphatase with activities as nucleotidaseand exopolyphosphatase is involved in stress response [35], showed in-creased abundance (spot 1409).
EF -Tu
TonB
Porin
AP
DnaK
HtpG
AHR
GroELDHLD
EF -Tu
TonB
Porin
AP
DnaK
HtpG
AHR
GroELDHLD
B
EF -TuDnaK
HtpG
TonB
Porin
AP
Omp
EF -TuDnaK
HtpG
TonB
Porin
AP
Omp
EF -TuDnaK
HtpG
TonB
Porin
AP
Omp
F
pH 3pH 3
pH 3pH 3
115
112 107
106
113
A116
108
109
114
101
102
103
104
105110
111
pH 3 1111
150kDa
SD
S-P
AG
E
301302
303
E304
305
306
311307
308309
pH 3 111110
150kDa
SD
S-P
AG
E
Fig. 5. Chaperone networks at growth temperature limits. (A, C, E, G) 2-DE gels of GroEL- and Dtained from S. frigidimarina grown at 4 °C (A, E) or 28 °C (C, G) and subjected to immunoprecipmunoprecipitation are given in Materials and methods section. Immunoprecipitates werecorresponded to proteins identified by MALDI-TOF–TOF and described in Tables 2, 3, 4 andGroEL and DnaK interaction data. Network overview showing the proteins interacting with Growith the STRING v 9.1 database have been represented in blue color. Interactions represented iTonB, TonB-dependent siderophore receptor; AP, alkaline phosphatase; EF-Tu, elongation factmembrane protein OmpK; SCoA, succinyl-CoA synthetase subunit beta; rrf, ribosome recyclingprotein; PNPase, polyribonucleotide nucleotidyltransferase.
3.2.1.7. Cell motility. Flagellar hook-associated protein (FlgL) was identi-fied in spot 805. This protein is a chaperone substrate of type III secre-tion chaperones that act as cytoplasmic bodyguards, protecting theirsecretion substrates from degradation and aggregation and in a sensingmechanism to determine the stage of flagellar assembly [36].
3.2.2. Proteins showing decreased abundance at warm temperatureProteins whose abundance declined in bacteria growing at warm
temperatures were involved in metabolic processes (32%), in redox ho-meostasis, formed part of the cell envelope (31%), and took part in coldacclimation (Fig. 3B).
3.2.2.1. Cold-acclimation proteins. Trigger factor which cooperates withDnaK in folding of newly synthesized proteins [5,37], corresponds tospot 420. This factor has been identified in P. haloplanktis as the maincold-acclimation protein up-regulated at low temperature. InS. frigidimarina, its abundance is also increased at cold temperaturebut it does not reach a high fold change at 28 °C with respect to 4 °C(average fold change −4.8).
3.2.2.2. Cell envelope. At low temperatures iron solubility is decreasedand its acquisition bymicroorganisms is a considerable challenge. An ef-ficient mechanism evolved by many microorganisms to obtain iron inlimited environments is the production and secretion of specific lowmolecularweight iron chelators termed siderophores. High-affinity sur-face receptor proteins bind siderophores, and subsequently facilitate
EF -Tu
Porin
DnaK
rrf
SOD
AHR
GroEL
EF -Tu
Porin
DnaK
rrf
SOD
AHR
GroEL
EF -Tu
Porin
DnaK
rrf
SOD
AHR
GroEL
H
DDnaK
EF-Tu
TonB
Porin
EF-Ts
OmpK
AHR
SCoA
GroEL
GroES
OmpPNPase
FATP
DnaK
EF-Tu
TonB
Porin
EF-Ts
OmpK
AHR
SCoA
GroEL
GroES
OmpPNPase
FATP
DnaK
EF-Tu
TonB
Porin
EF-Ts
OmpK
AHR
SCoA
GroEL
GroES
OmpPNPase
FATP
401
G
406
412 413
422
423424 427
428
411
1111
201 C
202
203204
205
206
207 208 209
210211
212
213
215
214
216
111110
150kDa
SD
S-P
AG
E
10
150kDa
SD
S-P
AG
E
425
DnaJ
naK-interacting proteins in S. frigidimarina grown at 4 °C and 28 °C. Cell extracts were ob-itation with monoclonal anti-GroEL (A, C) or anti-DnaK antibody (E, G). Conditions for im-resolved by 2-DE gels following image analysis. Numbered spots marked with circles5. The figure is a representative of four 2-DE experiments. (B, D, F, G) Overview of theEL (B, D) and DnaK (F, H) in S.frigidimarina. GroEL and DnaK interactions already predictedn red color have been identified in the present study. AHR, alkyl hydroperoxide reductase;or Tu; EF-Ts, elongation factor Ts; DHLD, dihydrolipoamide dehydrogenase; OmpK, outerfactor; SOD, superoxide dismutase; FATP, aromatic hydrocarbon degradation membrane
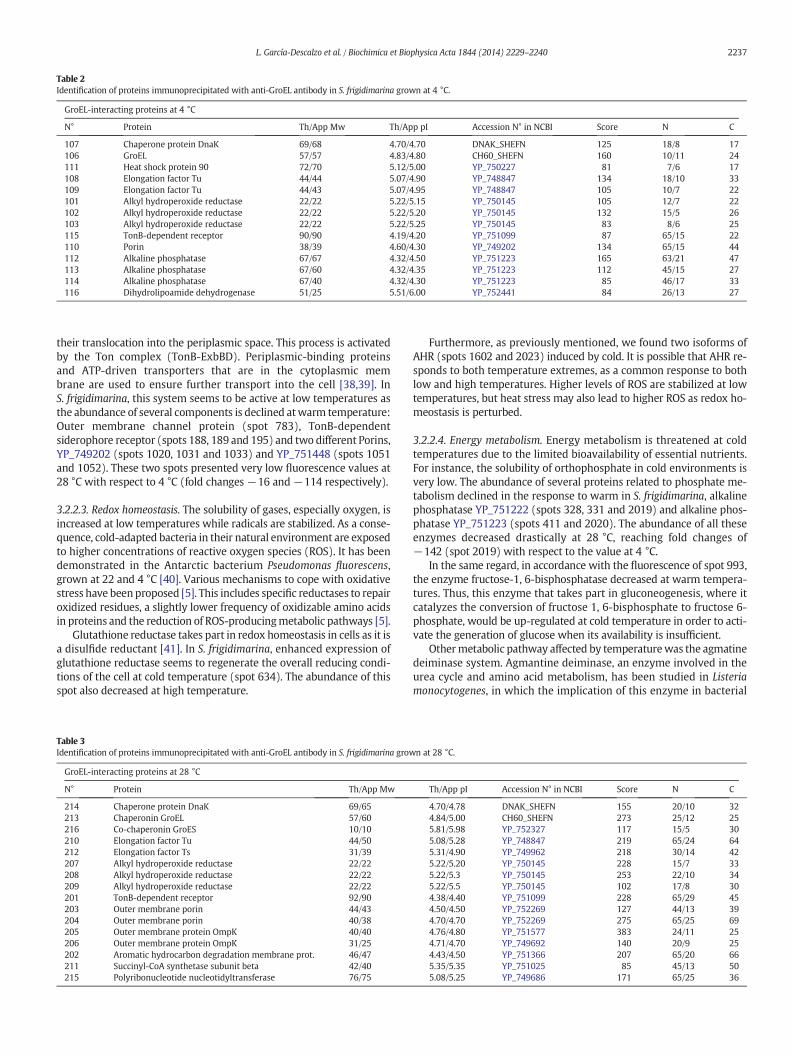
Table 2Identification of proteins immunoprecipitated with anti-GroEL antibody in S. frigidimarina grown at 4 °C.
GroEL-interacting proteins at 4 °C
N° Protein Th/App Mw Th/App pI Accession N° in NCBI Score N C
107 Chaperone protein DnaK 69/68 4.70/4.70 DNAK_SHEFN 125 18/8 17106 GroEL 57/57 4.83/4.80 CH60_SHEFN 160 10/11 24111 Heat shock protein 90 72/70 5.12/5.00 YP_750227 81 7/6 17108 Elongation factor Tu 44/44 5.07/4.90 YP_748847 134 18/10 33109 Elongation factor Tu 44/43 5.07/4.95 YP_748847 105 10/7 22101 Alkyl hydroperoxide reductase 22/22 5.22/5.15 YP_750145 105 12/7 22102 Alkyl hydroperoxide reductase 22/22 5.22/5.20 YP_750145 132 15/5 26103 Alkyl hydroperoxide reductase 22/22 5.22/5.25 YP_750145 83 8/6 25115 TonB-dependent receptor 90/90 4.19/4.20 YP_751099 87 65/15 22110 Porin 38/39 4.60/4.30 YP_749202 134 65/15 44112 Alkaline phosphatase 67/67 4.32/4.50 YP_751223 165 63/21 47113 Alkaline phosphatase 67/60 4.32/4.35 YP_751223 112 45/15 27114 Alkaline phosphatase 67/40 4.32/4.30 YP_751223 85 46/17 33116 Dihydrolipoamide dehydrogenase 51/25 5.51/6.00 YP_752441 84 26/13 27
2237L. García-Descalzo et al. / Biochimica et Biophysica Acta 1844 (2014) 2229–2240
their translocation into the periplasmic space. This process is activatedby the Ton complex (TonB-ExbBD). Periplasmic-binding proteinsand ATP-driven transporters that are in the cytoplasmic membrane are used to ensure further transport into the cell [38,39]. InS. frigidimarina, this system seems to be active at low temperatures asthe abundance of several components is declined atwarm temperature:Outer membrane channel protein (spot 783), TonB-dependentsiderophore receptor (spots 188, 189 and 195) and two different Porins,YP_749202 (spots 1020, 1031 and 1033) and YP_751448 (spots 1051and 1052). These two spots presented very low fluorescence values at28 °C with respect to 4 °C (fold changes −16 and−114 respectively).
3.2.2.3. Redox homeostasis. The solubility of gases, especially oxygen, isincreased at low temperatures while radicals are stabilized. As a conse-quence, cold-adapted bacteria in their natural environment are exposedto higher concentrations of reactive oxygen species (ROS). It has beendemonstrated in the Antarctic bacterium Pseudomonas fluorescens,grown at 22 and 4 °C [40]. Various mechanisms to cope with oxidativestress have beenproposed [5]. This includes specific reductases to repairoxidized residues, a slightly lower frequency of oxidizable amino acidsin proteins and the reduction of ROS-producingmetabolic pathways [5].
Glutathione reductase takes part in redox homeostasis in cells as it isa disulfide reductant [41]. In S. frigidimarina, enhanced expression ofglutathione reductase seems to regenerate the overall reducing condi-tions of the cell at cold temperature (spot 634). The abundance of thisspot also decreased at high temperature.
Table 3Identification of proteins immunoprecipitated with anti-GroEL antibody in S. frigidimarina grow
GroEL-interacting proteins at 28 °C
N° Protein Th/App Mw
214 Chaperone protein DnaK 69/65213 Chaperonin GroEL 57/60216 Co-chaperonin GroES 10/10210 Elongation factor Tu 44/50212 Elongation factor Ts 31/39207 Alkyl hydroperoxide reductase 22/22208 Alkyl hydroperoxide reductase 22/22209 Alkyl hydroperoxide reductase 22/22201 TonB-dependent receptor 92/90203 Outer membrane porin 44/43204 Outer membrane porin 40/38205 Outer membrane protein OmpK 40/40206 Outer membrane protein OmpK 31/25202 Aromatic hydrocarbon degradation membrane prot. 46/47211 Succinyl-CoA synthetase subunit beta 42/40215 Polyribonucleotide nucleotidyltransferase 76/75
Furthermore, as previously mentioned, we found two isoforms ofAHR (spots 1602 and 2023) induced by cold. It is possible that AHR re-sponds to both temperature extremes, as a common response to bothlow and high temperatures. Higher levels of ROS are stabilized at lowtemperatures, but heat stress may also lead to higher ROS as redox ho-meostasis is perturbed.
3.2.2.4. Energy metabolism. Energy metabolism is threatened at coldtemperatures due to the limited bioavailability of essential nutrients.For instance, the solubility of orthophosphate in cold environments isvery low. The abundance of several proteins related to phosphate me-tabolism declined in the response to warm in S. frigidimarina, alkalinephosphatase YP_751222 (spots 328, 331 and 2019) and alkaline phos-phatase YP_751223 (spots 411 and 2020). The abundance of all theseenzymes decreased drastically at 28 °C, reaching fold changes of−142 (spot 2019) with respect to the value at 4 °C.
In the same regard, in accordance with the fluorescence of spot 993,the enzyme fructose-1, 6-bisphosphatase decreased at warm tempera-tures. Thus, this enzyme that takes part in gluconeogenesis, where itcatalyzes the conversion of fructose 1, 6-bisphosphate to fructose 6-phosphate, would be up-regulated at cold temperature in order to acti-vate the generation of glucose when its availability is insufficient.
Othermetabolic pathway affected by temperaturewas the agmatinedeiminase system. Agmantine deiminase, an enzyme involved in theurea cycle and amino acid metabolism, has been studied in Listeriamonocytogenes, in which the implication of this enzyme in bacterial
n at 28 °C.
Th/App pI Accession N° in NCBI Score N C
4.70/4.78 DNAK_SHEFN 155 20/10 324.84/5.00 CH60_SHEFN 273 25/12 255.81/5.98 YP_752327 117 15/5 305.08/5.28 YP_748847 219 65/24 645.31/4.90 YP_749962 218 30/14 425.22/5.20 YP_750145 228 15/7 335.22/5.3 YP_750145 253 22/10 345.22/5.5 YP_750145 102 17/8 304.38/4.40 YP_751099 228 65/29 454.50/4.50 YP_752269 127 44/13 394.70/4.70 YP_752269 275 65/25 694.76/4.80 YP_751577 383 24/11 254.71/4.70 YP_749692 140 20/9 254.43/4.50 YP_751366 207 65/20 665.35/5.35 YP_751025 85 45/13 505.08/5.25 YP_749686 171 65/25 36

Table 4Identification of proteins immunoprecipitated with anti-DnaK antibody in S. frigidimarina grown at 4 °C.
DnaK -interacting proteins at 4 °C
N° Protein Th/AppMw Th/App pI Accession N° in NCBI Score N C
304 Chaperone protein DnaK 69/70 4.70/4.70 DNAK_SHEFN 80 15/6 20303 Heat shock protein 90 72/70 5.12/4.50 YP_750227 88 15/7 25307 Elongation factor Tu 44/41 5.07/5.00 YP_748847 92 41/10 31301 TonB-dependent receptor 90/90 4.19/4.2 YP_751099 122 65/19 30302 TonB-dependent receptor 105/100 4.35/4.40 YP_751154 191 65/24 36306 Porin 38/35 4.60/4.60 YP_749202 168 65/18 43311 Porin 38/35 4.60/4.20 YP_749202 122 65/14 39309 OmpA/MotB domain-containing protein 42/42 5.44/5.50 YP_751577 177 15/6 17308 Hypothetical protein Sfri_1465 41/41 5.13/5.15 YP_750155 310 27/13 34305 Alkaline phosphatase 67/65 4.32/4.60 YP_751223 142 64/22 39
2238 L. García-Descalzo et al. / Biochimica et Biophysica Acta 1844 (2014) 2229–2240
tolerance to stress and its contribution to the acid tolerance has beenshown [42]. Our results demonstrate also a role in adaptation to coldin S. frigidimarina (spot 951), showing a decreased abundance at 28 °Cwith respect to 4 °C.
At last, PeptidaseM61 belongs to a family of aminopeptidases foundin Proteobacteria with the ability to selectively cleave N-terminal acidicresidues of peptide substrates [43]. Its abundance also decreased at20 °C (spot 358) with respect to 28 °C but was not detectable at 4 °C.
Additionally, our results showed that 10 proteins appeared as 34 dif-ferent spots as marked in Fig. 4A. Interestingly all sets of isoforms hadthe same increasing or decreasing change in abundance in response towarm temperature, whereas one protein, AHR appeared as 5 isoformsthat exhibited opposite expression pattern (Fig. 4B and Table 1). Similarphenomena were demonstrated in other previously reported proteo-mics studies. For instance, GroEL isoforms were also found in the bacte-ria Gluconacetobacter, but those were non-differentially expressedproteins [44]. In our study, the abundance of all the detected isoformschanges depending on the growth temperature. In addition, it isknown that the phosphorylation state of several translation initiationfactors and ribosomal proteins are subjected to alteration during differ-ent types of stress [45]. This finding allows us to reasonably speculatethat some of the isoforms of EF-Tu and 30S ribosomal protein S1 arelikely to be posttranslational modifications by phosphorylation and de-phosphorylation. The activity of other proteins such as AHR has beenshown to be regulated by phosphorylation [46]. This may suggest thatseveral isoforms of AHRmay play different roles in modulating defenceresponses to extreme temperatures in S. frigidimarina.
3.3. Chaperone networks in S. frigidimarina at growth temperature limits
Chaperone complexes were investigated in S. frigidimarina to deter-mine changes in their interactions with client proteins in response tothe adaptation to growth temperature. Proteins of S. frigidimarina cul-tured at 4 °C and 28 °Cwere immunoprecipitatedwith specific antibod-ies anti-GroEL and anti-DnaK and analyzed by 2-DE (Fig. 5A, C, E, G). The
Table 5Identification of proteins immunoprecipitated with anti-DnaK antibody in S. frigidimarina grow
DnaK-interacting proteins at 28 °C
N° Protein Th/App Mw Th/App
425 Chaperone protein DnaK 69/70 4.70/4.7427 Chaperone protein DnaJ 41/41 8.56/8.5422 Chaperonin GroEL 57/60 4.84/5.8423 Elongation factor Tu 44/42 5.07/4.8424 Elongation factor Tu 44/44 5.07/5.1406 Ribosome recycling factor 21/21 5.27/5.3411 Alkyl hydroperoxide reductase 22/22 5.22/4.9412 Alkyl hydroperoxide reductase 22/22 5.22/5.0413 Alkyl hydroperoxide reductase 22/22 5.22/5.2428 Porin 38/40 4.60/4.6401 Superoxide dismutase 22/22 5.16/5.0
most abundant spots were then isolated from gels and identified byMALDI-TOF–TOF (Tables 2, 3, 4 and 5). At both 4 °C and 28 °C, GroELand DnaK proteins formed complexes, but their interaction partnerswere different depending on temperature. GroEL at 4 °C interacts withAHR, DnaK, EF-Tu, Porin, HtpG, AP, TonB and DHLD. GroEL also interactsat 28 °C with AHR, DnaK, EF-Tu, EF-Ts, SCoA, OmPK, OmP, TonB, FATP,PNPase and GroES. Additionally, the observed DnaK-interacting pro-teins were TonB, Hsp90, Porin, AP, EF-Tu, Hyp1465, PmpA, at 4 °C,meanwhile SOD, rrf, AHR, GroEL, EF-Tu, DnaJ and Porin were observedat 28 °C.
Direct interactions between proteins in cells represent an importantstep towards a comprehensive description of cellular mechanisms andfunctions. A network overview of GroEL and DnaK in S. frigidimarinawas performed using the STRING v 9.1 database (http://string-db.org/)[47]. As a result of these comparisons some new chaperone interactionshave been found (Figs. 5B, D, F, H and 6).
Most of the proteins involved in the complexes were also differen-tially expressed at 4 °C and 28 °C (Table 1), and many of them showedprotein isoforms (Fig. 4). Among chaperones, DnaK was alwaysobserved in 2-DE gels from samples immunoprecipitated with anti-GroEL antibody, but GroEL only precipitated with DnaK at high temper-ature, forming a complex with DnaJ (Figs. 5 and 6). Further, GroELinteracted with GroES at 28 °C, confirming the results seen in 2-DDIGE experiments (Fig. 3S and Table 1). Another stress protein thatimmunoprecipitated with both GroEL and DnaK at 4 °C was theHsp90. Furthermore, alkaline phosphatase was observed at 4 °Cinteracting with both GroEL and DnaK. Taking into account that theabundance of all isoforms of this enzyme decreased drastically at28 °C, reaching fold changes of −142 with respect to the value at4 °C, the formation of these complexes could contribute to the stress re-sponse at cold temperature.With respect to AHR, three spots of this pro-tein were observed in 2-DE gels A and C (Fig. 5), what revealed theirinteraction with GroEL at 4 °C and 28 °C. Other proteins such as DHLD,FATP, PNPase, rff and SOD only participated in one of the complexes.The unique factor that appeared in all the networks was EF-Tu (Fig. 6),
n at 28 °C.
pI Accession N° in NCBI Score N C
0 DNAK_SHEFN 223 18/14 370 YP_749661 109 8/7 265 CH60_SHEFN 605 12/14 230 YP_748847 147 54/16 520 YP_748847 164 28/14 420 YP_749964 95 24/13 275 YP_750145 105 10/7 420 YP_750145 92 17/7 302 YP_750145 95 16/6 350 YP_749202 92 65/12 450 YP_750435 124 44/11 41

A B
C D
Adaptation to warming
GroEL
DHLD
Hyp1465PmpA
Hsp90AP
EF-TsSCoAOmPKOmPFATP PNPaseGroES
SODrrfDnaJ
EF-TuDnaK
TonB GroELAHR
Porin
BEF-TsSCoAOmPKOmPFATP PNPaseGroES
C
Hyp1465PmpA
ADHLD
Hsp90AP
TonB
D
SODrrfDnaJ
GroELAHR
Porin
EF-TuDnaK
A B
C D
Adaptation to warming
DnaK
DHLD
Hyp1465PmpA
Hsp90AP
EF-TsSCoAOmPKOmPFATP PNPaseGroES
SODrrfDnaJ
EF-TuDnaK
TonB GroELAHR
Porin
Fig. 6. Venn diagram analysis of the GroEL- and DnaK-interacting proteins inS. frigidimarina grown at 4 °C and 28 °C. Proteins found in complexes immunoprecipitatedwith specific anti-GroEL (A, B) and anti-DnaK (C, D) antibodies in cell extracts ofS. frigidimarina cultured at 4 °C (A, C) or 28 °C (B, D) are shown in the different segments.
2239L. García-Descalzo et al. / Biochimica et Biophysica Acta 1844 (2014) 2229–2240
in contrast to EF-Ts that was only involved in the complexwith GroEL at28 °C. It has been assessed that one of the major effects of cold temper-atures onmicroorganisms is the inhibition of translation elongation [13,48]. Finally, different cell envelope proteins appeared implicated in allcomplexes. The association of chaperones with translation factors orwith cell envelope proteins may represent a method of coping withthe stress situation at both extreme temperatures.
In total, our results suggest that warming may have a profound im-pact on Antarctic bacteria. Although the detection of these effects withassays of total bacterial community structure is difficult, they dobecomemanifest in the distribution of key functional proteins (Fig. 3). In awarming environment, a tight regulation is necessary to maintain thesurvival of these microorganisms [49,50]. An intense stress response istriggered from 20 °C (maximum growth rate) to 28 °C (upper tempera-ture limit). And a very little increase of temperature from 28 °C to 30 °Ccompletely threatens S. frigidimarina survival (Fig. 1). Taken together,findings presented here suggest that two different types of physiologicchanges are involved in the adaptation of S. frigidimarina to extremetemperatures. At high temperature, cells induced stress proteins (heatshock proteins, proteases and antioxidants), protein biosynthesis andmotility in order to avoid cellular injury (Fig. 3A). In contrast, in theresponse of cells growing at 4 °C induced proteins related to cold-acclimation, cell envelope, energy metabolism and redox homeostasiswere induced (Fig. 3B). Redox homeostasis is also controlled throughdifferent mechanisms, at high temperature with the activity of Dps,meanwhile at low temperatures glutathione reductase plays the mainrole as antioxidant. Additionally, diverse isoforms of the AHRparticipatein redox homeostasis depending on the environmental temperature.This is the only identified protein whose abundance increased at bothextreme temperatures. Furthermore, at low temperature the mainstress is caused by low availability of nutrients and iron that inducesthe adjustment of the metabolic network and antioxidative system.
4. Conclusions
Until now, scarce data are available to characterize the molecularmechanism by which cold-adapted bacteria respond to warming,including the metabolic pathways and fundamental physiological pro-cesses that are specifically affected. Using 2D-DIGE and MALDI-TOF–TOF MS new insights into how the Antarctic bacterium S. frigidimarinacope with a warming environment was explored. Cells reorganize
their chaperone networks as an adaptive response to these environ-mental conditions but a continuous increase of temperature willthreaten their survival, what is likely relevant as they are essential com-ponents in their ecosystems.
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.bbapap.2014.08.006.
Acknowledgments
This research was supported by grants CTM/2008-00304/ANT,CTM2010-12134-E/ANT and CTM2011-16003-E from the SpanishMinisterio de Economía y Competitividad (MINECO). E.G.L. is recipientof a MINECO Fellowship (Programa Nacional de Contratación eIncorporación de RRHH, Spain).
References
[1] R. Cavicchioli, K.S. Siddiqui, D. Andrews, K.R. Sowers, Low-temperatureextremophiles and their applications, Curr. Opin. Biotechnol. 3 (2002) 253–261.
[2] L. García-Descalzo, A. Alcazar, F. Baquero, C. Cid, Biotechnological applications ofcold-adapted bacteria, Extremophiles: Sustainable Resources and BiotechnologicalImplications, Wiley-Blackwell, Hoboken, New Jersey, 2012, pp. 159–174.
[3] J. Turner, J.C. King, T.A. Lachlan-Cope, P.D. Jones, Recent temperature trends in theAntarctic, Nature 418 (2002) 291–292.
[4] E. Yergeau, G.A. Kowalchuk, Responses of Antarctic soil microbial communities andassociated functions to temperature and freeze–thaw cycle frequency, Environ.Microbiol. 10 (2008) 2223–2235.
[5] F. Piette, S. D'Amico, C. Struvay, G. Mazzucchelli, J. Renaut, M.L. Tutino, A. Danchin, P.Leprince, G. Feller, Proteomics of life at low temperatures: trigger factor is the pri-mary chaperone in the Antarctic bacterium Pseudoalteromonas haloplanktisTAC125, Mol. Microbiol. 76 (2010) 120–132.
[6] M.V. Jagannadham, C. Chowdhury, Differential expression of membrane proteinshelps Antarctic Pseudomonas syringae to acclimatize upon temperature variations,J. Proteomics 75 (2012) 2488–2499.
[7] M. Kube, T.N. Chernikova, Y. Al-Ramahi, A. Beloqui, N. Lopez-Cortez, M.E.Guazzaroni, H.J. Heipieper, S. Klages, O.R. Kotsyurbenko, I. Langer, T.Y. Nechitaylo,H. Lünsdorf, M. Fernández, S. Juárez, S. Ciordia, A. Singer, O. Kagan, O. Egorova, P.A. Petit, P. Stogios, Y. Kim, A. Tchigvintsev, R. Flick, R. Denaro, M. Genovese, J.P.Albar, O.N. Reva, M. Martínez-Gomariz, H. Tran, M. Ferrer, A. Savchenko, A.F.Yakunin, M.M. Yakimov, O.V. Golyshina, R. Reinhardt, P.N. Golyshin, Genome se-quence and functional genomic analysis of the oil-degrading bacterium OleispiraAntarctica, Nat. Commun. 4 (2013) 2156.
[8] A. Goodchild, M. Raftery, N.F. Saunders, M. Guilhaus, R. Cavicchioli, Biology of thecold adapted archaeon, Methanococcoides burtonii determined by proteomicsusing liquid chromatography–tandem mass spectrometry, J. Proteome Res. 3(2004) 1164–1176.
[9] M. Strocchi, M. Ferrer, K.N. Timmis, P.N. Golyshin, Low temperature-induced sys-tems failure in Escherichia coli: insights from rescue by cold-adapted chaperones,Proteomics 6 (2006) 193–206.
[10] T.J. Williams, F.M. Lauro, H. Ertan, D.W. Burg, A. Poljak, M.J. Raftery, R. Cavicchioli,Defining the response of a microorganism to temperatures that span its completegrowth temperature range (−2 °C to 28 °C) using multiplex quantitative proteo-mics, Environ. Microbiol. 13 (2011) 2186–2203.
[11] L. Ting, T.J. Williams, M.J. Cowley, F.M. Lauro, M. Guilhaus, M.J. Raftery, R. Cavicchioli,Cold adaptation in the marine bacterium, Sphingopyxis alaskensis, assessed usingquantitative proteomics, Environ. Microbiol. 12 (2010) 2658–2676.
[12] C. Cid, L. Garcia-Descalzo, V. Casado-Lafuente, R. Amils, A. Aguilera, Proteomic anal-ysis of the response of an acidophilic strain of Chlamydomonas sp. (Chlorophyta) tonatural metal-rich water, Proteomics 10 (2010) 1–11.
[13] L. García-Descalzo, A. Alcazar, F. Baquero, C. Cid, Identification of in vivo HSP90-interacting proteins reveals modularity of HSP90 complexes is dependent on theenvironment in psychrophilic bacteria, Cell Stress Chaperones 16 (2011) 203–218.
[14] I. Miller, J. Crawford, E. Gianazza, Protein stains for proteomic applications: which,when, why? Proteomics 6 (2006) 5385–5408.
[15] J.P. Bowman, S.A. McCammon, D.S. Nichols, J.H. Skerratt, S.M. Rea, P.D. Nichols, T.A.McMeekin, Shewanella gelidimarina sp. nov. and Shewanella frigidimarina sp. nov.,novel Antarctic species with the ability to produce eicosapentaenoic acid (20:5omega 3) and grow anaerobically by dissimilatory Fe(III) reduction, Int. J. Syst.Bacteriol. 47 (1997) 1040–1047.
[16] M.V. Jagannadham, M.K. Chattopadhyay, C. Subbalakshmi, M. Vairamani, K.Narayanan, C.M. Rao, S. Shivaji, Carotenoids of an Antarctic psychrotolerant bacteri-um, Sphingobacterium antarcticus, and a mesophilic bacterium, Sphingobacteriummultivorum, Arch. Microbiol. 173 (2000) 418–424.
[17] P.D. Schloss, H.K. Allen, A.K. Klimowicz, C. Mlot, J.A. Gross, S. Savengsuksa, J. McEllin,J. Clardy, R.W. Ruess, J. Handelsman, Psychrotrophic strain of Janthinobacteriumlividum from a cold Alaskan soil produces prodigiosin, DNA Cell Biol. 29 (2010)533–541.
[18] J.S. McCarty, A. Buchberger, J. Reinstein, B. Bukau, The role of ATP in the functionalcycle of the DnaK chaperone system, J. Mol. Biol. 249 (1995) 126–137.
[19] M.K. Hayer-Hartl, J. Martin, F.U. Hartl, Asymmetrical interaction of GroEL and GroESin the ATPase cycle of assisted protein folding, Science 269 (1995) 836–841.

2240 L. García-Descalzo et al. / Biochimica et Biophysica Acta 1844 (2014) 2229–2240
[20] A.J. Darwin, The phage-shock-protein response, Mol. Microbiol. 57 (2005) 621–628.[21] J.R. Glover, S. Lindquist, Hsp104, Hsp70, and Hsp40: a novel chaperone system that
rescues previously aggregated proteins, Cell 94 (1998) 73–82.[22] G. Román-Hernández, J.Y. Hou, R.A. Grant, R.T. Sauer, T.A. Baker, The ClpS adaptor
mediates staged delivery of N-end rule substrates to the AAA + ClpAP protease,Mol. Cell 43 (2011) 217–228.
[23] I. Ruggiero, G. Raimo, M. Palma, P. Arcari, M. Masullo, Molecular and functionalproperties of the psychrophilic elongation factor G from the Antarctic EubacteriumPseudoalteromonas haloplanktis TAC 125, Extremophiles 11 (2007) 699–709.
[24] G. Raimo, B. Lombardo, M. Masullo, A. Lamberti, O. Longo, P. Arcari, Elongation factorTs from the Antarctic eubacterium Pseudoalteromonas haloplanktis TAC 125: bio-chemical characterization and cloning of the encoding gene, Biochemistry 43(2004) 14759–14766.
[25] W. Freist, D.T. Logan, D.H. Gauss, Glycyl-tRNA synthetase, Biol. Chem. Hoppe Seyler377 (1996) 343–356.
[26] A.K. Singh, S. Shivaji, A cold-active heat-labile t-RNA modification GTPase from a psy-chrophilic bacterium Pseudomonas syringae (Lz4W), Res. Microbiol. 161 (2010) 46–50.
[27] M. Sprinzl, S. Brock, Y. Huang, P. Milovnik, M. Nanninga, M. Nesper-Brock, H.Rütthard, K. Szkaradkiewicz, Regulation of GTPases in the bacterial translation ma-chinery, Biol. Chem. 381 (2000) 367–375.
[28] S. Nakagawa, Y. Niimura, K. Miura, T. Gojobori, Dynamic evolution of translation ini-tiation mechanisms in prokaryotes, Proc. Natl. Acad. Sci. U. S. A. 107 (2010)6382–6387.
[29] I.V. Boni, D.M. Isaeva, M.L. Musychenko, N.V. Tzareva, Ribosome-messenger recogni-tion:mRNA target sites for ribosomal protein S1,Nucleic AcidsRes. 19 (1991)155–162.
[30] L. Chen, L. Keramati, J.D. Helmann, Coordinate regulation of Bacillus subtilis peroxidestress genes by hydrogen peroxide and metal ions, Proc. Natl. Acad. Sci. U. S. A. 92(1995) 8190–8194.
[31] L. Leblanc, C. Leboeuf, F. Leroi, A. Hartke, Y. Auffray, Comparison between NaCl tol-erance response and acclimation to cold temperature in Shewanella putrefaciens,Curr. Microbiol. 46 (2003) 157–162.
[32] L.N. Calhoun, Y.M. Kwon, Structure, function and regulation of the DNA-binding pro-tein Dps and its role in acid and oxidative stress resistance in Escherichia coli: a re-view, J. Appl. Microbiol. 110 (2011) 375–386.
[33] K.L. Turner, M.K. Doherty, H.A. Heering, F.A. Armstrong, G.A. Reid, S.K. Chapman,Redox properties of flavocytochrome c3 from Shewanella frigidimarina NCIMB400,Biochemistry 38 (1999) 3302–3309.
[34] A.M. Chakrabarty, Nucleoside diphosphate kinase: role in bacterial growth, viru-lence, cell signaling and polysaccharide synthesis, Mol. Microbiol. 28 (1998)875–882.
[35] W. Iwasaki, K. Miki, Crystal structure of the stationary phase survival protein SurEwith metal ion and AMP, J. Mol. Biol. 371 (2007) 123–136.
[36] P. Aldridge, J. Karlinsey, K.T. Hughes, The type III secretion chaperone FlgN regulatesflagellar assembly via a negative feedback loop containing its chaperone substratesFlgK and FlgL, Mol. Microbiol. 49 (2003) 1333–1345.
[37] E. Deuerling, A. Schulze-Specking, T. Tomoyasu, A. Mogk, B. Bukau, Trigger factorand DnaK cooperate in folding of newly synthesized proteins, Nature 400 (1999)693–696.
[38] K.D. Krewulak, H.J. Vogel, TonB or not TonB: is that the question? Biochem. Cell Biol.89 (2011) 87–97.
[39] J.D. Faraldo-Gómez, M.S. Sansom, Acquisition of siderophores in gram-negative bac-teria, Nat. Rev. Mol. Cell Biol. 4 (2003) 105–116.
[40] M.K. Chattopadhyay, G. Raghu, Y.V. Sharma, A.R. Biju, M.V. Rajasekharan, S. Shivaji,Increase in oxidative stress at low temperature in an Antarctic bacterium, Curr.Microbiol. 62 (2011) 544–546.
[41] F. Aslund, J. Beckwith, Bridge over troubled waters: sensing stress by disulfide bondformation, Cell 96 (1999) 751–753.
[42] J. Chen, C. Cheng, Y. Xia, H. Zhao, C. Fang, Y. Shan, B. Wu, W. Fang, Lmo0036, an or-nithine and putrescine carbamoyltransferase in Listeria monocytogenes, participatesin arginine deiminase and agmatine deiminase pathways and mediates acid toler-ance, Microbiology 157 (2011) 3150–3161.
[43] S.N. Jamdar, A novel aminopeptidase from Burkholderia cepacia specific for acidicamino acids, FEMS Microbiol. Lett. 295 (2009) 230–237.
[44] M.F. dos Santos, V.L. Muniz de Pádua, E. de Matos Nogueira, A.S. Hemerly, G.B.Domont, Proteome of Gluconacetobacter diazotrophicus co-cultivated with sugar-cane plantlets, J. Proteomics 73 (2010) 917–931.
[45] B. El-Fahmawi, G.W. Owttrim, Cold-stress-altered phosphorylation of EF-Tu in thecyanobacterium Anabaena sp. strain PCC 7120, Can. J. Microbiol. 53 (2007) 551–558.
[46] T.S. Chang, W. Jeong, S.Y. Choi, S. Yu, S.W. Kang, S.G. Rhee, Regulation ofperoxiredoxin I activity by Cdc2-mediated phosphorylation, J. Biol. Chem. 277(2002) 25370–25376.
[47] A. Franceschini, D. Szklarczyk, S. Frankild, M. Kuhn, M. Simonovic, A. Roth, J. Lin, P.Minguez, P. Bork, C. von Mering, J.L. Jensen, STRING v9.1: protein–protein interac-tion networks, with increased coverage and integration, Nucleic Acids Res. 41(2013) 808–815.
[48] D. Georlette, V. Blaise, T. Collins, S. D'Amico, E. Gratia, A. Hoyoux, J.C. Marx, G. Sonan,G. Feller, C. Gerday, Some like it cold: biocatalysis at low temperatures, FEMSMicrobiol. Rev. 28 (2004) 25–42.
[49] J. Han, J. Jung, M. Park, S. Hyun, W. Park, Short-term effect of elevated temperatureon the abundance and diversity of bacterial and archaeal amoA genes in Antarcticsoils, J. Microbiol. Biotechnol. 6 (2013) 692–702.
[50] E. Yergeau, S. Bokhorst, S. Kang, J. Zhou, C. Greer, R.W. Aerts, G.A. Kowalchuk, Shiftsin soil microorganisms in response to warming are consistent across a range of Ant-arctic environments, ISME J. 6 (2012) 692–702.
Copyright © 2022 FDOKUMEN




















