
Protective immunogenicity of the paraflagellar rod protein 2 of Leishmania mexicana
12
Vaccine 23 (2005) 984–995 Protective immunogenicity of the paraflagellar rod protein 2 of Leishmania mexicana N.G. Saravia a,∗ , M.H. Hazb ´ on a,1 , Y. Osorio a , L. Valderrama a , J. Walker a , C. Santrich b , T. Cort´ azar a , J.H. LeBowitz b,2 , B.L. Travi a a Centro Internacional de Entrenamiento e Investigaciones M´ edicas (CIDEIM), Apartado A´ ereo 5390, Cali, Colombia b Department of Biochemistry, Purdue University, West Lafayette, IN, USA Received 4 December 2003; received in revised form 19 July 2004; accepted 26 July 2004 Available online 22 September 2004 Abstract Paraflagellar rod proteins (PFR) are a potent immunogen against experimental Trypanosoma cruzi infection. PFR are highly conserved among kinetoplastid parasites. We therefore evaluated the immunogenicity of the Leishmania mexicana pfr-2 gene and protein product in the hamster model of American cutaneous leishmaniasis. Immunization with pfr-2 DNA-induced specific antibody, confirming immunogenicity. Subsequent challenge with 10,000 and 500 stationary phase L. mexicana promastigotes respectively, resulted in delayed appearance of lesions, and significant reduction in lesions post infection in male hamsters, yet exacerbated lesions in female hamsters. Immunization with recombinant PFR-2 protein (rPFR-2) prevented lesion development in female hamsters challenged with L. panamensis, but was ineffective against L. mexicana. Nevertheless, prime boost immunization of female hamsters with rPFR and pfr-2 DNA significantly reduced lesion size following challenge with 500 L. mexicana promastigotes, supporting the relevance of PFR-2 as a potential vaccine constituent. © 2004 Elsevier Ltd. All rights reserved. Keywords: Leishmania; Paraflagellar rod protein; DNA vaccine 1. Introduction Vaccination has been proposed to be the most cost- effective measure to control leishmaniasis. However, there is still no safe and effective vaccine for any form of human leishmaniasis. The current understanding of the immunology of leishma- niasis in experimental murine models indicates that the con- trol of the infection requires an immune response by the Th1 subset of CD4 + T lymphocytes, which produce macrophage activating cytokines [1]. DNA vaccines are able to induce Th1 ∗ Corresponding author. Tel.: +57 2 6682164/6682160; fax: +57 2 6672989. E-mail address: [email protected] (N.G. Saravia). 1 Present address: Center for Emerging Pathogens, University of Medicine and Dentistry of New Jersey, Newark, NJ, USA. 2 Present address: Symbiontics Inc., Saint Louis, MO, USA. responses even in the case of antigens that injected as pro- teins elicit a Th2 response. For example, the -galactosidase or gp63 proteins induce Th2 responses but their correspond- ing genes, LacZ or gp63, respectively, induce Th1 responses [2–4]. Therefore, the Th1 dominant response to genetic vacci- nation is an attractive feature for vaccine development against several parasitic diseases including leishmaniasis [2,5]. Par- tial protection of susceptible BALB/c mice against L. ma- jor infection by immunization with the coding sequence for gp63, the most abundant surface glycoprotein in Leishmania, demonstrated the feasibility of genetic vaccination against leishmaniasis [4]. The rationale for the use of paraflagellar rod proteins (PFR-1 and -2) as immunogens derives from findings on the protective immunogenicity of flagellar proteins of an- other kinetoplastid pathogen, Trypanosoma cruzi. Inocula- tion of mice with flagellar fractions of T. cruzi resulted in 80–90% survival following challenge with lethal doses of 0264-410X/$ – see front matter © 2004 Elsevier Ltd. All rights reserved. doi:10.1016/j.vaccine.2004.07.044
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Protective immunogenicity of the paraflagellar rod protein 2 of Leishmania mexicana

Vaccine 23 (2005) 984–995
Protective immunogenicity of the paraflagellar rod protein 2 ofLeishmania mexicana
N.G. Saraviaa,∗, M.H. Hazbona,1, Y. Osorioa, L. Valderramaa, J. Walkera,C. Santrichb, T. Cortazara, J.H. LeBowitzb,2, B.L. Travia
a Centro Internacional de Entrenamiento e Investigaciones M´edicas (CIDEIM), Apartado A´ereo 5390, Cali, Colombiab Department of Biochemistry, Purdue University, West Lafayette, IN, USA
Received 4 December 2003; received in revised form 19 July 2004; accepted 26 July 2004Available online 22 September 2004
Abstract
Paraflagellar rod proteins (PFR) are a potent immunogen against experimentalTrypanosoma cruziinfection. PFR are highly conserveda heh ity.S ce ofl ization withr ea ef©
K
1
eil
ntsa
f
a
s pro-eond-
sesacci-ainst
for,ainst
einss onan-
of
0d
mong kinetoplastid parasites. We therefore evaluated the immunogenicity of theLeishmania mexicana pfr-2gene and protein product in tamster model of American cutaneous leishmaniasis. Immunization withpfr-2DNA-induced specific antibody, confirming immunogenicubsequent challenge with 10,000 and 500 stationary phaseL. mexicanapromastigotes respectively, resulted in delayed appearan
esions, and significant reduction in lesions post infection in male hamsters, yet exacerbated lesions in female hamsters. Immunecombinant PFR-2 protein (rPFR-2) prevented lesion development in female hamsters challenged withL. panamensis, but was ineffectivgainstL. mexicana. Nevertheless, prime boost immunization of female hamsters with rPFR andpfr-2DNA significantly reduced lesion sizollowing challenge with 500L. mexicanapromastigotes, supporting the relevance of PFR-2 as a potential vaccine constituent.
2004 Elsevier Ltd. All rights reserved.
eywords: Leishmania; Paraflagellar rod protein; DNA vaccine
. Introduction
Vaccination has been proposed to be the most cost-ffective measure to control leishmaniasis. However, there
s still no safe and effective vaccine for any form of humaneishmaniasis.
The current understanding of the immunology of leishma-iasis in experimental murine models indicates that the con-
rol of the infection requires an immune response by the Th1ubset of CD4+ T lymphocytes, which produce macrophagectivating cytokines[1]. DNA vaccines are able to induce Th1
∗ Corresponding author. Tel.: +57 2 6682164/6682160;ax: +57 2 6672989.
E-mail address:[email protected] (N.G. Saravia).1 Present address: Center for Emerging Pathogens, University of Medicinend Dentistry of New Jersey, Newark, NJ, USA.2 Present address: Symbiontics Inc., Saint Louis, MO, USA.
responses even in the case of antigens that injected ateins elicit a Th2 response. For example, the�-galactosidasor gp63 proteins induce Th2 responses but their corresping genes,LacZor gp63, respectively, induce Th1 respon[2–4]. Therefore, the Th1 dominant response to genetic vnation is an attractive feature for vaccine development agseveral parasitic diseases including leishmaniasis[2,5]. Par-tial protection of susceptible BALB/c mice againstL. ma-jor infection by immunization with the coding sequencegp63, the most abundant surface glycoprotein inLeishmaniademonstrated the feasibility of genetic vaccination agleishmaniasis[4].
The rationale for the use of paraflagellar rod prot(PFR-1 and -2) as immunogens derives from findingthe protective immunogenicity of flagellar proteins ofother kinetoplastid pathogen,Trypanosoma cruzi. Inocula-tion of mice with flagellar fractions ofT. cruzi resulted in80–90% survival following challenge with lethal doses
264-410X/$ – see front matter © 2004 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2004.07.044

N.G. Saravia et al. / Vaccine 23 (2005) 984–995 985
infective parasites[5,6]. Decreased parasitemia and tissuelesions were also achieved by immunization with flagellarfractions[7]. Subsequently, the flagellar antigen that elicitedthe protective response was identified as PFR. Immuniza-tion of mice with PFR-1 and PFR-2 purified fromT. cruziepimastigotes, induced an immune response capable of pro-tecting against a lethal challenge ofT. cruzi trypomastig-otes[8], and this response was shown to be a Th1-type re-sponse[9]. The finding that immunized mice depleted ofB cells survived lethal challenge withT. cruzi trypomastig-otes whereas mice deficient in CD8+ and CD4+ cells did notresolve the parasitemia[10], further substantiated the cell-mediated nature of the protective response elicited by PFRproteins.
The pfr-2 gene fromLeishmania mexicana(accessionnumber U45884)[11] encodes a 599-residue of PFR pro-tein, which has 83% amino acid identity with its counterpartfrom T. cruzi.Hence, we reasoned that thepfr-2 gene prod-uct ofL. mexicanacould share common epitopes involved ineliciting a protective immune response as occurred withT.cruziPFR. The aim of this study was to evaluate the potentialof thepfr-2gene product fromL.mexicanaas an immunogenin the immunoprophylaxis of New World dermal leishmaniaswhen administered either as a DNA vaccine or as a recombi-nant protein vaccine. The study was conducted in the goldenh tc cur-r ithL .W x oft , andta
2
2
l-lp n,S
2
edw urer
2
s-f er ions( acts
from the transient transfectants and fromL. mexicana(MNY/BZ/62/M379) subclone M379 promastigotes were re-solved by SDS–PAGE as described[13,14]. Immunoblotswere performed using an ECL labeling kit according to themanufacturer’s instructions (Amersham). Anti-PFR (mono-clonal antibody 2E10, a gift of Diane McMahon–Pratt[15],was used to detect PFR proteins.
2.4. In vitro expression of recombinant PFR (rPFR-2)
An in vitro expression system was produced by insertion ofthe clonedL. mexicana pfr-2gene into the expression vectorpET28a, and the resulting construct was used to transformEs-cherichia coliBL21. Transformed bacterial cultures were se-lected by growth on Luria–Bertani (LB) agar plates (contain-ing 0.1% glucose and 50�g/mL kanamycin). A single colonywas then inoculated into a 25 mL LB/glucose/kanamycin liq-uid preculture, incubated overnight (14 h) at 37◦C in an or-bital shaker. The preculture was then transferred to 1 L ofliquid medium and growth was continued for 2 h at 37◦C.rPFR-2 expression was then induced by addition of isopropyl-�-d-thiogalactopyranoside (IPTG) at a final concentration of1 mM. Following a further 2.5 h of growth at 37◦C, bacte-ria were harvested by centrifugation (10,000× g, 15 min,4◦C).
2
2R-2
w nal,tao in-c tionp da genI singw M)a fferc ndy oge-n lec-t
2
vedu ingE fur-t ore-s . Thee 8%p R-2b licesb 2
amster, which is susceptible to theLeishmaniaspecies thaause cutaneous leishmaniasis in the New World and isently the best available experimental model of infection w. panamensisand other species of theVianniasubspeciese examined the effect of the route of inoculation and se
he experimental host on the immune response inducedhe protection of hamsters against challenge withL.mexicanandL. panamensis.
. Materials and methods
.1. Construction of the DNA vaccine
A 1.8 kbEcoRI–NotI DNA fragment harbouring the fulength L. mexicana pfr-2coding sequence fromplasmidPFR2[12], was cloned into the pCDNA3 vector (Invitrogean Diego, USA), to obtain the plasmid pCDNA3-pfr-2.
.2. Purification of the DNA vaccine
DNA for immunization was overproduced and purifiith the EndoFree Plasmid Giga kit, following the proced
ecommended by the manufacturer (Qiagen, USA).
.3. Transfection and expression of pfr-2 in HeLa cells
The pCDNA3-pfr-2 construct was transiently tranected into human HeLa cells using the FuGenTM
eagent, following the manufacturer’s recommendatBoehringer Mannheim, Germany). Crude protein extr
.5. Purification of rPFR-2
.5.1. Ni2+-affinity chromatographyThe pET28a-pfr-2 expression system generates rPF
ith a tag of six histidine residues at the amino termihus facilitating subsequent purification by nickel (Ni2+)-ffinity chromatography using His-Bind® resin [16]. Lysisf bacterial cells, recovery and extraction of the insolublelusion bodies containing rPFR-2, and general purificarocedures using His-Bind®-affinity resin were performeccording to manufacturer’s recommendations (Nova
nc., USA), except that column washes were conducted uash buffer containing 10 mM imidazole (instead of 60 mnd rPFR-2 was eluted from the resin using elution buontaining 120 mM imidazole (instead of 1 M). Purity aield of rPFR-2 was determined by SDS–PAGE on homous 8% polyacrylamide gels using the Mini-Protean II e
rophoresis system (Bio-Rad).
.6. Preparative SDS–PAGE and electroelution
Although substantial purification of rPFR-2 was achiesing Ni2+-affinity chromatography, several contaminat. coliproteins remained in the final column eluate, and
her purification required the use of preparative electrophis and subsequent electroelution of the rPFR-2 bandluates from the affinity resin were electrophoresed onolyacrylamide preparative SDS–PAGE gels, the rPFand was excised and protein was recovered from gel sy electroelution at 60 mA (3 h, 4◦C) using the Model 42

986 N.G. Saravia et al. / Vaccine 23 (2005) 984–995
Table 1Summary of immunization protocols
No. ofexperiment
Protocol Challenge,specie/inoculuma
No. ofhamsters, total(no./group)
Immunogen Route ofinoculationb
1st immu-nization,dose/site
2nd–3rd im-munizations,dose/site/intervals
Intervals ofobservationpost challenge(days)
1 DNA vaccine; routeof inmunization;male vs. female
L. mexicana;10,000
80 (8) pfr-2 DNA;vector
IM or ID 50 �g;hindpaw andforepaw
50�g;hindpaw andforepaw, 1month
15, 30, 45, 60
2 rPFR-2; site ofinmunization; malevs. female
L. panamensis;10,000
75 (7–16) rPFR-2 SCc/ID 100�g;hindpaw andforepaw
50�g;hindpaw orear, 1 month
15, 30, 45, 75,120
3a DNA vaccine; males L. mexicana;500
48 (16) pfr-2 DNA;vector
ID 100�g;forepaw
50�g;hindpaw andear; 2months
15, 30, 60, 90
3b rPFR-2; male vs.female
L. mexicana;500
42 (11–16) rPFR-2 SCc/ID 100�g;forepaw
50�g;hindpaw andear; 2months
15, 30, 60
4a Prime-boost;DNA-protein vs.protein-DNA;females
L. mexicana;1000
35 (7) DNA-protein;protein-DNA
ID 20�g;forepaw
10�g; ears;1 month
15, 30, 45, 60,75
4b DNA vaccine; malevs. female
L. mexicana;1000
42 (7) pfr-2 DNA;vector
ID 20�g;forepaw
10�g; ears 15, 30, 45, 60,75
a Number of stationary phase promastigotes ofLeishmaniaused as infective inoculum after immunizations.b Route of inoculation: IM = intramuscular; ID = intradermic; SC = subcutaneous.c SC first immunization with rPFR-2 plus Freund’s incomplete adjuvant.
Electro-Eluter (Bio-Rad) and a protein elution buffer (25 mMTrizma base, 192 mM glycine 0.5 M urea and 0.1% SDS) ac-cording to manufacturer’s recommendations. Subsequently,the eluted rPFR-2 was subjected to electrodialysis in orderto remove bound SDS, which interferes with immunologi-cal evaluation of the antigen. The protein elution buffer wasreplaced by a dialysis buffer (as above but without SDS),and four rounds of electrodialysis were performed (each of2 h duration) at 4◦C. The rPFR-2 preparation was passivelydialysed against 1 L of 0.5 M urea for 24 h (with three changesof urea solution), and then concentrated using polyethyleneglycol.
The purity and concentration of the final rPFR-2 prepa-ration was determined by SDS–PAGE (as above) and amodification of the Bradford protein assay, respectively. Cy-totoxicity of the purified rPFR-2 preparation was evaluatedby co-culture with hamster lymph node cells for 24 h fol-lowed by assessment of viability. Cell viability was evalu-ated by microscopy and/or proliferation assay in response topromastigotes in the presence of rPFR-2.
2.7. Leishmania strains
A subclone (LM16) of the WHO reference strain forL.mexicana(MNY/BZ/62/M379) and a strain ofL. panamen-s nt,w pro-m dium[
2.8. Immunization and evaluation of protection inhamsters
Male and female golden hamsters (Mesocriteus aura-tus), 6–8 weeks of age, were used for the experiments.Animals were maintained under standard caging conditionsprovided with food and water ad libitum according with theGuiding Principles for Biomedical Research Involving An-imals (Council for International Organizations of MedicalSciences) and the law 84 of 1989 “Estatuto Nacional de Pro-teccion de los Animales”, Colombia. Animals were anes-thetized with 50 mg/kg ketamine clorhydrate (Ketamina®,Holliday-Scott S.A.) and 0.02 mg/kg xylazine (Rompun®,Bayer Colombia). At the end of the study, the animals weresedated and euthanized in a CO2 chamber.
Several different immunization strategies were tested: vac-cination with rPFR-2 protein; immunization with the con-structpCDNA3-pfr2 (pfr-2 DNA) or pcDNA3 (vector); andprime-boost immunization consisting of eitherpfr-2 DNAfollowed by rPFR-2 protein (DNA-protein) or vice versa(protein-DNA). The experimental protocols for each strategyare summarized inTable 1. Recombinant PFR-2,pfr-2DNAvaccine and vector were diluted and delivered in PBS. Thesite of intramuscular (IM) inoculation was the tibialis anteri-oris and carpal flexor muscles. Intradermal (ID) inoculationw
af-t goteso s
is(HOM/COL/84/1099) isolated from a Colombian patieere used for challenge experiments. Stationary phaseastigotes were harvested from Senekjie’s diphasic me
17] after culture at 24◦C for 7 days.
as targeted to the ears and dorsal skin of the paws.Challenge inoculations were carried out 15 days
er the last booster using stationary phase promastif L. mexicana(MNYC/BZ/62/M379) or L. panamensi

N.G. Saravia et al. / Vaccine 23 (2005) 984–995 987
(MHOM/COL/84/1099). Parasites were collected from thesupernatant of Senekjie’s culture medium by centrifugation,resuspended in PBS and counted in a hemocytometer. Thechallenge inoculum was adjusted to 500–10,000 parasites(Table 1) in 50�L of PBS and delivered intradermally inthe hindpaw. Naive non-immunized infected controls wereincluded in each experiment. The size of lesions in the pawwas determined with a digital caliper at 15 days intervalsand expressed as the diameter of the infected paw minus thediameter of the contra-lateral uninfected paw, in mm[18].Protection was evaluated as median lesion size of immunizedhamsters compared with the median lesion size of naive ani-mals.
2.9. Delayed-type hypersensitivity response (DTH) torPFR-2
DTH response to rPFR-2 was evaluated over a dose rangeof 5, 25, 50 and 100�g of rPFR-2 protein by ID injectionin the forepaw of hamsters infected withL. panamensis, anduninfected controls. Groups of two to three infected and un-infected hamsters were tested for each dose. Specific DTHresponse was determined by measuring the diameter of in-duration of the forepaw 48 and 72 h after injecting rPFR-2minus the contra-lateral paw injected with PBS containing0 thec
2
phn ationo lfs5 s( (b)r5 ki-n erepL er-m tru-m ragec la-t tainn base1 pos-i devi-a outa
2r
tigenw lenge
infection with Leishmaniaby enzyme-linked immunoab-sorbent assay (ELISA). Microtiter plates (Immulon 2, Dynex,Helsinki, Finland) were coated with rPFR-2 antigen at1�g/mL in 1× coating buffer (Kirkegaard & Perry Labo-ratories Inc., KPL, USA) and were incubated overnight at4◦C. After blocking with 1× bovine serum albumin (BSA)(KPL) for 1 h at 37◦C, 1:25 dilution of hamster serum in1× BSA was added in duplicate and incubated for 1 h at37◦C. Peroxidase-conjugated Protein A (1:1000) (Sigma)was added and incubated at 37◦C, 1 h. After washing with1× PBS–0.1%Tween 20 (Sigma), 100�L of ABTS sub-strate (KPL) was added and the enzymatic reaction allowedto develop for 10 min before evaluation using an ELISAreader (Dynex) at a wavelength of 405 nm. The average op-tical density (OD) was obtained for duplicate serum sam-ples.
2.12. Immunoblotting
rPFR-2 protein, and whole lysates ofL. mexicanasubclone LM16 and the PFR deletion mutant line 13.2were separated by SDS–PAGE followed by electroblot-ting onto nitrocellulose membranes (Amershan PharmaciaBiotech, UK). The membranes were blocked with PBSw ithsc 20.A 000d Ther m)a ityw E10[
2
pro-g ald riedo ga-r dis-t ro-l SA.P i-a odyr wered fromt cul-t re-s uaret andw neyn -p
.05–1 M urea according to the concentration of urea inorresponding dilution of rPFR-2.
.10. Lymphoproliferation
Lymphocytes from retropharingeal and axillary lymodes of hamsters were cultured at a final concentrf 2.5 × 106 cells/mL RPMI 1640 with 5% of foetal caerum. The cells were stimulated with (a) 2.5× 105,× 105 and 1 × 106 promastigotes ofL. panamensi
MHOM/CO/85/1166) lysed by freezing and thawing;PFR-2 at 2.5, 5 and 10�g/mL, during 3 days at 37◦C with% CO2, based on prior experiments to determine theetics of the lymphoproliferative response. Cultures wulsed for 8 h with 1�Ci/well of [3H]thymidine (Amershamife Science, UK) and incorporation into DNA was detined using a beta scintillation counter (Beckman Insents). Each condition was tested in triplicate. The ave
ounts per minute (cpm) of controls (cells without stimuion) was subtracted from cpm of stimulated cells to obet cpm, and the results transformed to logarithm in0 for statistical analysis. A response was considered
tive when the net cpm was greater than 2 standardtions from the mean of control cultures (medium withntigen).
.11. Determination of specific IgG in response toPFR-2 by ELISA
Specific IgG produced in response to the rPFR-2 anas determined in hamster sera obtained before chal
ith 0.2% of goat serum and then incubated for 2 h wera from hamsters inoculated with thepfr-2 DNA vac-ine and vector diluted 1:50 in PBS–0.05% Tweenfter washing, peroxidase-conjugate Protein A in 1:1ilution was added and incubated during one hour.eaction was developed with an ECL kit (Amershafter 10 min of exposure on an X-ray film. Specificas confirmed using monoclonal anti-PFR antibody 2
15].
.13. Statistical analyses
Statistical analyses were performed using the SPSSram 1998 version for Windows. Verification of normistribution of data from quantitative variables was carut using the Kolmogorov–Smirnov statistical test. Loithmic transformation was required to achieve normalribution of the data for the analysis of lymphocyte piferation and antibody response to rPFR-2 using ELIarametric analyses, Student’st-test and analysis of varnce were performed to establish differences in antibesponse and lymphoproliferation. Positive responsesefined as values greater than 2 standard deviations
he mean of healthy controls for antibody and mean ofures without stimulus for lymphocyte transformationponses. This information was analyzed using the chi-sqest. Lesion size showed non-parametric distributionas analyzed using the Kruskal–Wallis or Mann–Whiton-parametric test. SignificantP values (P < 0.05) are reorted.
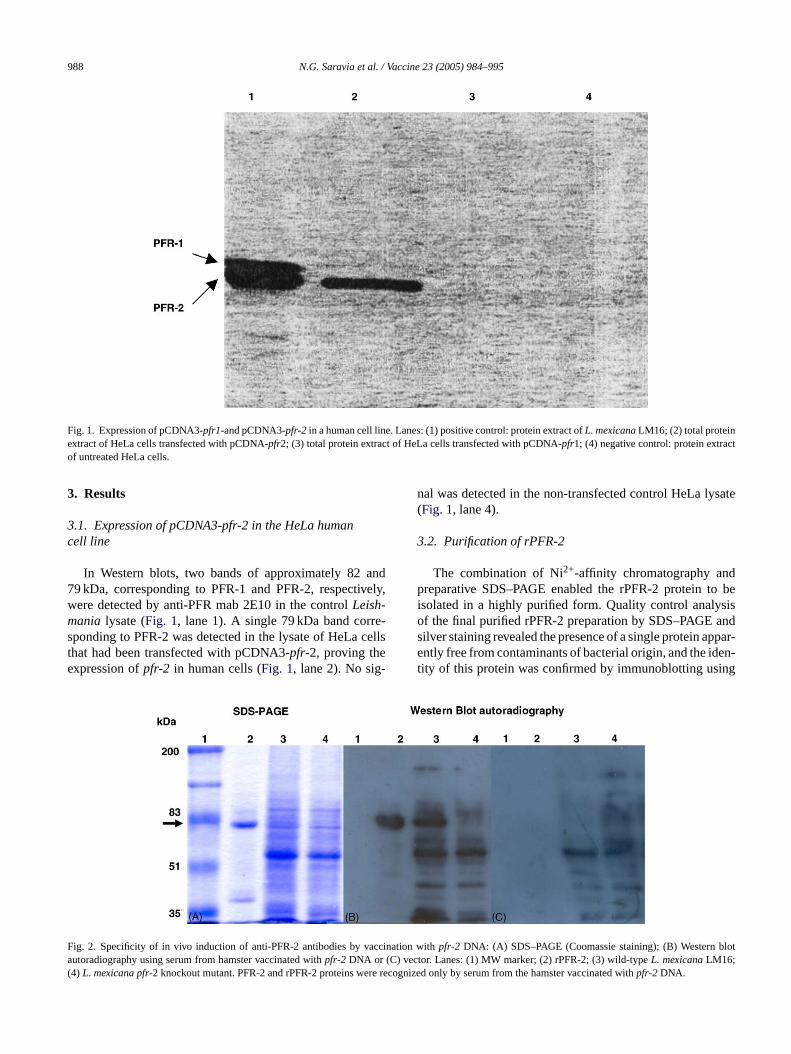
988 N.G. Saravia et al. / Vaccine 23 (2005) 984–995
Fig. 1. Expression of pCDNA3-pfr1-and pCDNA3-pfr-2 in a human cell line. Lanes: (1) positive control: protein extract ofL. mexicanaLM16; (2) total proteinextract of HeLa cells transfected with pCDNA-pfr2; (3) total protein extract of HeLa cells transfected with pCDNA-pfr1; (4) negative control: protein extractof untreated HeLa cells.
3. Results
3.1. Expression of pCDNA3-pfr-2 in the HeLa humancell line
In Western blots, two bands of approximately 82 and79 kDa, corresponding to PFR-1 and PFR-2, respectively,were detected by anti-PFR mab 2E10 in the controlLeish-mania lysate (Fig. 1, lane 1). A single 79 kDa band corre-sponding to PFR-2 was detected in the lysate of HeLa cellsthat had been transfected with pCDNA3-pfr-2, proving theexpression ofpfr-2 in human cells (Fig. 1, lane 2). No sig-
F ccinat blota (C) vec( re reco
nal was detected in the non-transfected control HeLa lysate(Fig. 1, lane 4).
3.2. Purification of rPFR-2
The combination of Ni2+-affinity chromatography andpreparative SDS–PAGE enabled the rPFR-2 protein to beisolated in a highly purified form. Quality control analysisof the final purified rPFR-2 preparation by SDS–PAGE andsilver staining revealed the presence of a single protein appar-ently free from contaminants of bacterial origin, and the iden-tity of this protein was confirmed by immunoblotting using
ig. 2. Specificity of in vivo induction of anti-PFR-2 antibodies by vautoradiography using serum from hamster vaccinated withpfr-2 DNA or4) L. mexicana pfr-2 knockout mutant. PFR-2 and rPFR-2 proteins we
ion withpfr-2 DNA: (A) SDS–PAGE (Coomassie staining); (B) Westerntor. Lanes: (1) MW marker; (2) rPFR-2; (3) wild-typeL. mexicanaLM16;gnized only by serum from the hamster vaccinated withpfr-2DNA.

N.G. Saravia et al. / Vaccine 23 (2005) 984–995 989
Table 2PFR specific antibody production measured by ELISA in response to vaccination withpfr-2 DNA, vector, and rPFR-2 according to sex and route of inoculation
Route of inoculation/immunogen
Antibody (OD) reactive with rPFR-2
Females Males All (females and males)
n % Positivea,b G meanc,d 95% CI n % Positiveb G meand,e 95% CI n % Positiveb G meand,e 95% CI
Intradermicpfr-2 DNA 9 56 0.31 0.15–0.64 41 46 0.29 0.22–0.37 50 48 0.29 0.23–0.37Vector 9 0 0.10 0.09–0.10 39 3 0.10 0.09–0.12 48 2 0.10 0.09–0.11
Intramuscularpfr-2 DNA 7 71 0.68 0.30–1.54 9 33 0.30 0.15–0.59 16 50 0.43 0.25–0.74Vector 10 0 0.11 0.09–0.12 8 0 0.10 0.08–0.11 18 0 0.10 0.09–0.11
rPFR-2 45 100 1.89 1.86–1.92 39 100 1.91 1.87–1.94 84 100 1.90 1.87–1.92Healthy controls 21 5 0.12 0.10–0.15 25 4 0.11 0.10–0.13 46 4 0.12 0.10–0.13
a Positive: optical density greater than 2 standard deviation of the mean of negative controls.b Chi-square test,P < 0.001. Comparison among animals immunized withprf-2 DNA, vector or rPFR-2, and healthy controls.c Geometric mean.d Anova one way,P < 0.001. Comparison among animals immunized withprf-2 DNA, vector or rPFR-2, and healthy controls.e Anova main effects,P < 0.001.pfr-2 DNA vs. vector, controlled by sex and route of inoculation.
mono- and polyclonal antibodies specific for PFR-2. Thequantity of rPFR-2 obtained was diminished by losses duringthe preparative electrophoresis and electro elution steps, yetfinal yields ranging from 3.8 to 9.6 mg of pure rPFR-2 perlitre of E. coli culture were still achieved.
3.3. Antibody response
Immunization withpfr-2 DNA, by different routes andbooster protocols, induced specific antibody response torPFR-2 overall in 48% of 66 vaccinated hamsters while 2%of 66 hamsters inoculated with the vector alone, and 4% of46 non-immunized healthy control hamsters presented an-tibody reactive with rPFR-2. The mean optical density ofsera (diluted 1:25) from hamsters vaccinated withpfr-2DNAwas significantly higher than that of hamsters inoculated withthe vector, independently of the sex or route of inoculation(Table 2) P < 0.001, and that of healthy control sera. No-tably, immunization with rPFR-2 yielded markedly higherantibody responses (mean OD = 1.90) in 100% of 84 ham-sters than thepfr-2 DNA vaccine delivered by either the IDor IM routeP < 0.001. The specificity of the antibody de-
Table 3In vitro proliferative response of hamster lymph node cells elicited byL. panamensispromastigotes and rPFR-2
A
anb
25 81 7rrr 6
ard denet cpm
r differ
tected by ELISA was confirmed by Western blot of rPFR-2protein andL.mexicanacell lysates separated by SDS–PAGE(Fig. 2).
Immunization withpfr-2 DNA by the IM route tendedto induce higher mean OD than ID immunization in maleand female hamster sera, but the difference did not reachsignificance (Table 2). Female hamsters that receivedpfr-2DNA IM presented both a higher percentage of positive seraand higher mean OD than those receiving the vaccine ID,and male hamsters vaccinated by either route presented alower percentage of positive sera and lower OD than femalehamsters. Nevertheless, there were no significant differencesin antibody response between male and female hamsters(Table 2).
3.4. Recombinant PFR-2 protein is recognized by thecell-mediated immune system of hamsters experimentallyinfected with L. panamensis
Cell-mediated immune response to rPFR-2 protein in-duced by experimental infection was evaluated as lymphocyteproliferation in vitro and cutaneous delayed hypersensitivity
ntigen Healthy controls
n % Positivea Geometric me
.5× 105 promastigotes/mL 10 10c 14d
× 105 promastigotes/mL 11 18c 49d
0× 105 promastigotes/mL 10 20c 125d
PFR-2, 2.5�g/mL 3 0 105PFR-2, 5�g/mL 6 0 102PFR-2, 10�g/mL 11 18 461
a Positive response = net cpm thymidine incorporation > mean + 2 standb [3H]thymidine incorporation in response to antigenic stimulation (c Chi-square test,P < 0.05. Healthy controls vs. infected hamsters.d Anova one way,P < 0.05. Healthy controls vs. infected hamsters fo
Infected withL. panamensis
95% CI n % Positive Geometric mean 95% CI
1–130 12 50 655 61–71026–385 13 62 2038 301–13,77
20–784 16 69 4144 1132–15,165–2160 5 0 132 4–46447–1464 6 17 264 13–5426
99–2153 17 47 1400 441–444
viations of control culture without stimulus.).
ent promastigote concentrations.

990 N.G. Saravia et al. / Vaccine 23 (2005) 984–995
Fig. 3. DTH response to rPFR-2 antigen in hamsters infected withLeishmaniacompared with uninfected controls. Data are presented as box plot of the diameterof induration for all infected or uninfected hamsters independendty of dose of rPFR-2. Induration diameter = diameter of the foot inoculated ID with rPFR-2− diameter of the foot inoculated ID with the vehicle alone at 48 and 72 h. Mann–Whitney test, infected vs. uninfected: (a)P= 0.031 at 48 h; (b)P < 0.001 at72 h.
(DTH). In vitro evidence of specific recognition of rPFR-2include a dose response effect for lymphocytes from infectedhamsters but not for lymphocytes from healthy control ham-sters and lymphoproliferative response to 10 (g/mL rPFR-2protein in 8/17 (47%) hamsters infected withL. panamensis,and 2/11 (18%) healthy control hamsters (Table 3). Althoughstatistical significance was not reached, the geometric meannet cpm elicited by 10�g rPFR-2 was 3× higher for lympho-cytes from infected hamsters than healthy controls (geometricmean net cpm (1400 versus 461 cpm) (Table 3). The positivecontrol response of lymphocytes from infected hamsters tokilled promastigotes ofL. panamensisalso showed a doseresponse.
Specific delayed-type hypersensitivity response waselicited in hamsters with active experimentalL. panamen-sis infections by the intradermal injection of rPFR-2 overa dose range of 5–100�g (Fig. 3). Statistically significantdifferences in the response to dose of rPFR were not ob-served for either infected or uninfected animals. However,comparison of the DTH responses of infected and uninfectedhamsters independently of the dose of rPFR, showed that theinduration in infected hamsters was significantly larger thanin uninfected hamsters at both 48 and 72 h (Mann–Whitney,P= 0.031 andP< 0.001 at 48 and 72 h, respectively) (Fig. 3).Together, the cutaneous DTH and lymphoproliferative re-s tallyi thL ofP
3rmm
stig-o on-i e dif-
ference did not achieve statistical significance in these exper-iments.
Intradermal immunization with thepfr-2 DNA vaccinefollowed by boosters at 1 month intervals prevented the de-velopment of lesions in male hamsters during the first 30days following challenge with 1× 104 stationary phasepromastigotes ofL. mexicana(Kruskal–Wallis,P < 0.05;Mann–Whitney,P= 0.004) (Experiment 1,Table 1) (Fig. 4).However, by 45 days post challenge, lesions comparable tothose in naive hamsters had developed in hamsters inoculatedwith the vaccine or vector. Intramuscular delivery of thepfr-2 DNA also induced a significant reduction in lesion size at30 days post challenge. Notably, the vector alone was alsoimmunostimulatory. Significantly smaller lesions developedin male hamsters inoculated IM with the vector at 30 days(Kruskal–Wallis,P < 0.05; Mann–Whitney,P= 0.017) and45 days (P< 0.05; Mann–Whitney,P= 0.014) post challenge(Fig. 4).
In contrast with the outcome of challenge infection inmales, female hamsters were not protected by ID or IM vacci-nation withpfr-2 DNA. Rather, ID-vaccinated female ham-sters developed significantly larger lesions than vaccinatedmale hamsters (Mann–Whitney,P< 0.001). The vector alonehad a similar exacerbating effect in female hamsters. Hence,the vector was itself immunostimulatory in hamsters of boths pend-it ctoro imin-i f the6
leh hicht wasi ducedta per-i ed
ponse of lymph node cells from hamsters experimennfected withL. panamensisdemonstrate that infection wi. panamensisinduces cell-mediated immune recognitionFR-2.
.5. Immunization with pfr-2 DNA delays onset oreduces lesion size in male hamsters and increasesorbidity in female hamsters challenged with L.exicana
Challenge infection with doses of 500–10,000 promates ofL. mexicanatended to produce larger lesions in n
mmune male hamsters than in female hamsters but th
exes, resulting in decreased or increased lesion size deng upon the sex of the recipient hamsters (Fig. 4). Overall,he immunomodulating effects of both the vaccine and ven lesions in male and female hamsters subsequently d
shed and were not statistically significant at the end o0 days period of observation.
The capacity ofpfr-2 DNA to induce protection in maamsters was improved in a subsequent experiment in w
he interval between intradermal booster immunizationsncreased to 2 months and the challenge dose was reo 500 stationary phase promastigotes ofL. mexicana, whichpproximates the inoculum of an infected sand fly (Ex
ment 3a,Table 1). With this inoculum, lesions develop

N.G. Saravia et al. / Vaccine 23 (2005) 984–995 991
Fig. 4. Clinical response in female (blank symbols) or male (filled symbols)hamsters immunized ID or IM with 100�g pfr-2 DNA (vaccine) or vectorfollowed by two boosters of 50�g of vaccine or vector at 1 month intervalsand challenged 15 days later by ID inoculation of 10,000 stationary phasepromastigotes ofL. mexicana. Data is expressed as the median of the lesionsize of the experimental group (n= 8 hamsters per group). Kruskal–Wallistest,P < 0.05: (a) comparison among animals immunized with vector,pfr-2DNA, and naive at 30 and 45 days. Mann–Whitney test: (b)P= 0.004 males:pfr-2DNA ID vs. naive; (c)P < 0.05 males: vector IM vs. naive.
only after 60 days in controls and hamsters receiving thevector alone. At the end of the period of observation 3months post challenge, lesions were significantly smaller inthe vaccinated hamsters (median 1.9 mm) than in the non-immunized controls (median 3.5 mm) (P= 0.004). Lesionsin hamsters receiving the vector alone (median 2.4 mm) werenot significantly different from controls. Immunization ofmale hamsters with lower doses ofpfr-2DNA or vector alone(Experiment 4b,Table 1) failed to induce significant reduc-tion in lesions after challenge with 1000L. mexicanapro-mastigotes (data not shown).
3.6. rPFR-2 protein induces protection of female but notmale hamsters against challenge with L. panamensis
Subcutaneous immunization with rPFR-2 protein plusFreund’s incomplete adjuvant followed by ID boosters ad-
Fig. 5. Lesion evolution in female (blank symbols) and male hamsters (filledsymbols) immunized subcutaneously in the hindpaw with 100�g rPFR-2plus Freund’s incomplete adjuvant, followed 2 months later by two boostersat 60 days intervals with 50�g rPFR-2 in PBS, then challenged 15 dayslater with 10,000 stationary phase promastigotes ofL. panamensisID intothe forepaw. Data are presented as median of lesion size of the respectiveexperimental group (n= 16 hamsters per group). Kruskal–Wallis test,P <0.01: (a) comparison among female hamsters receiving booster with rPFR-2 ID in forepaw or ear, and naive at 45 and 75 days. Mann–Whitney test:comparison male vs. female hamsters: (b)P= 0.001 naive; (c)P < 0.001rPFR-2 forepaw; (d)P < 0.001 rPFR-2 ear.
ministered into the skin of the ear or paw markedly reducedlesions in female hamsters during 4 months observation fol-lowing challenge with 10,000 stationary phase promastig-otes of L. panamensis(Experiment 2,Table 1) (Fig. 5).Median lesion size was significantly smaller at 45 and 75days post challenge in female hamsters immunized ID andboosted in either the ear (Kruskal–Wallis,P < 0.01). Malehamsters were not protected (Fig. 5). Both the course ofdisease in naive controls, and the response to immunizationof male and female hamsters upon challenge withL. pana-mensiswere widely disparate (Mann–Whitney,P < 0.001)(Fig. 5).
3.7. rPFR-2 protein induces transitory reduction inlesion size in male and female hamsters challenged withL. mexicana
Thirty days post challenge with 500 stationary phase pro-mastigotes ofL. mexicana, male hamsters immunized withrPFR-2 presented smaller lesions than non-immunized con-trols (median lesion size immunized versus naive: males,0.1 mm versus 0.5 mm; females, 0 mm versus 0 mm) butthis difference did not reach significance (Experiment 3b,Table 1). By 2 months post challenge, lesions had progressedand there was no difference between immunized and non-i im-m ales,2
mmunized hamsters of both sexes (median lesion sizeunized versus naive: males, 4 mm versus 3.6 mm; femmm versus 1.8 mm).

992 N.G. Saravia et al. / Vaccine 23 (2005) 984–995
Fig. 6. Lesion size in female hamsters 30–75 days after infection with 1000promastigotes ofL. mexicanafollowing immunization with “prime-boost”protocols,pfr-2 DNA vaccine or vector alone vs. naive hamsters. DNA-protein: ID immunization withpfr-2 DNA (20�g in forepaw, two boostersof 10�g in ears) followed by a final booster with 10�g rPFR-2; protein-DNA: ID immunization with rPFR-2 (20�g in forepaw, two boosters of10�g in ears) followed by a final booster with 10�g pfr-2DNA; DNA: IDimmunization withpfr-2DNA vaccine ID in forepaw and ears as for primeboost groups; vector: ID immunization with vector plasmid alone in forepawand ears as for prime boost groups. Median lesion size for each group (n= 7hamsters per group). Kruskal–Wallis test,P= 0.018: (a) comparison amongfemales immunized with protein-DNA, DNA-protein,pfr-2 DNA, vectorand naive at 75 days. Mann–Whitney test,P= 0.017: (b)pfr-2 DNA vs.DNA-protein or protein-DNA; (c) vector vs. protein-DNA at 75 days.
3.8. Prime-boost combination of pfr-2 DNA and rPFR-2protein induces protection against challenge with L.mexicana in female hamsters
In contrast to the exacerbation of lesions in female ham-sters challenged withL. mexicanaafter immunization withpfr-2DNA, prime-boost immunization with combined DNAand protein vaccine presentations resulted in reduced lesionsize (Experiment 4a,Table 1). This occurred regardless ofthe sequence of exposure topfr-2DNA and rPFR-2. Signifi-cance was reached at day 75 post infection (Kruskal–Wallis,P= 0.018) (Fig. 6). Administration ofpfr-2DNA vaccine orvector alone, even in reduced quantity, induced exacerbationof lesions in female hamsters, as observed in previous exper-iments (Mann–Whitney,P= 0.017 DNA-protein versus pfr-2DNA and protein-DNA versus vector).
4. Discussion
This investigation provides evidence that the gene and cor-responding recombinant protein of the PFR-2 component ofthe paraflagellar rod ofL. mexicanaare immunogenic andthe elicited response can ameliorate or exacerbate lesionsin golden hamsters challenged with infective promastigoteso nv spe-
cific response to rPFR-2 was induced by genetic vaccina-tion as shown by the antibody response. The long-term re-duction of lesions in male and female hamsters by differentimmunization schemes following challenge infection witha biologically relevant inoculum of infective promastigotesalso supports the induction of specific adaptive immunity byPFR-2.
The presentation of PFR as DNA and/or protein, sex ofthe experimental host and species ofLeishmaniaused forchallenge, independently influenced the outcome of immu-nization. Factors underlying the divergent clinical outcomesof challenge infection include differences in the pathogenic-ity of the two species ofLeishmaniain the hamster[19,20]and in the susceptibility of male and female hamsters[18].The very high sequence of homology ofpfr genes in kineto-plastids predicted that the response induced withL.mexicanaPFR-2 would be to conserved epitopes, and therefore not bespecies specific. The cell-mediated immune recognition ofL. mexicanarPFR-2 by hamsters infected withL. panamen-sis and the protection of female hamsters immunized withrPFR-2 againstL. panamensisinfection support this notion.
Genetic vaccination of male hamsters delayed lesion de-velopment post challenge with the relatively high (10,000)number of stationary phaseL. mexicanapromastigotes, andsignificantly reduced lesion size over a longer period of ob-sp ectedb ions,s aryp n-i i-n chesh ic vac-c d-i useoi im-m aryt rs byvmi
f e andd self-r ions[a os heno-t alsob[ vantf aticu f thec r ex-a male
f L. mexicanaandL. panamensis. Although the expressioector clearly exercised an immunostimulatory effect, a
ervation against a challenge inoculum of 500L. mexicanaromastigotes. Although female hamsters were not proty the DNA vaccine alone and even presented larger lesignificant protection from a challenge of 1000 stationhase promastigotes ofL. mexicanawas achieved by admi
stration ofpfr-2 DNA in a prime boost protocol in combation with rPFR-2 protein. Various prime-boost approaave been employed to enhance the response to genetines[21]. Use of DNA in combination with the corresponng recombinant protein[22] has been superseded by thef non-replicating, non-integrating viral vectors[21], which
nduce high levels of both CD4 and CD8 cell-mediatedunity. Our results provide proof of principle that contr
o the deleterious response induced in female hamsteaccination withpfr-2DNA, a protective response toLeish-ania mexicanacan be achieved by combiningpfr-2 DNA
mmunization with protein in prime boost strategies.The pathogenicity ofL. mexicanaandL. panamensisdif-
er in the golden hamster. The former causes progressivisseminated lesions, while the latter produces smalleresolving primary lesions and eventually metastatic les19]. Male hamsters are more susceptible to bothL.mexicanandL. panamensisthan female hamsters[18], hence the twexes represent models of susceptible and resistant pypes, respectively. Sex differences in pathogenicity haveeen described forL.majorandL.mexicanainfection in mice
23–25]. Both susceptible and resistant models are releor preclinical evaluations of vaccines since the pragmse of only male animals because of easier readout olinical outcome, may obscure the potential beneficial ocerbating effect of the same vaccine and regimen in fe

N.G. Saravia et al. / Vaccine 23 (2005) 984–995 993
animals. The distinct clinical responses of male and femalehamsters to different vaccine presentations and challenge in-fection with different species ofLeishmaniademonstratedthe influence of host sex on the outcome of immunizationas well as the pathogenesis of leishmaniasis. These disparateoutcomes underscore the need to consider sex in the designand analysis of preclinical and clinical evaluation of vaccinesfor leishmaniasis. Indeed, the results of this study raise theissue of whether the evidence for protection of boys but notgirls, in the trial of the killedL. majorplus BCG vaccine inIran may have a biological basis linked to gender[26].
Increasing evidence indicates that the profile of immuneresponse that is protective or disease promoting is distinctfor different species ofLeishmaniain mouse models[27].The contrasting clinical evolution of infection withL. mexi-canaandL. panamensisfollowing vaccination with rPFR-2protein substantiates the distinct immunobiology of infec-tion by these species ofLeishmania.Studies by others usinginbred mice and different species ofLeishmaniahave alsofound that immunization with an antigen from one speciescan be more effective in ameliorating disease caused by otherspecies[28,29]. Together these findings show that the type ofresponse induced by a particular vaccine and regimen, even ingenetically identical hosts, can distinctly impact subsequentinfection by differentLeishmaniaspecies.
la-t theD suchm thatD ry inmp ts,w sterscIT nseoA ea heiv
i sions n ofD hicha -l cebit n toii in thep pectso
r rodp in
the promastigote stage[11], has potential as a vaccine con-stituent. The evidence of immune recognition of rPFR-2 pro-tein by lymph node cells and the elicitation of a specific DTHresponse to this protein in hamsters experimentally infectedwith L. panamensis, indicate that the immune system is ex-posed to PFR-2 protein during infection withLeishmania.Whether this occurs during inoculation of promastigotes oris the result of low level of expression of this gene product byamastigotes is uncertain, yet the transitory exposure to pro-mastigotes during inoculation seems unlikely to explain therecall response to PFR-2.
The protective response elicited by PFR may be optimizedby modification of the vaccine construct to include othergene and adjuvant sequences, and further development of theprime boost combination of rPFR-2 protein and DNA. In ad-dition, the use of other types of adjuvant, including locally orsystemically administered immunopotentiating drugs such astucaresol that acts through T cell receptor independent signal-ing, or imidazoquinolones that induce Toll receptor signalingcould be used to steer the initial response in a more favor-able direction[42–44]. Optimization of protective immuno-genicity (i.e. long lasting and complete prevention of disease)remains a challenge common toLeishmaniavaccine candi-dates. Nevertheless, DNA vaccines, particularly polygenicpresentations have yielded promising results in the suscepti-b -p le orr minga eredi
A
269U orR OL-C Col-c Thet vitroa efullya ini-t thec blea
R
andience
re-mid
gly-onse
Notably, the pCDNA3 vector was itself immunostimuory and this is likely to have resulted from CpG motifs inNA sequence of the construct. In fact, there were 21otifs in the vector and 31 in the vaccine. It is knownNA sequences of bacterial origin are immunostimulatoammals, and that CpG motifs (5′-Pur-Pur-C-G-Pyr-Pyr-3′)lay an important role[30,31]. In independent experimene observed 17–20% reduction of lesion size in hamhallenged with 10,000 promastigotes ofL. panamensisafterD inoculations with the ODN-CpG (5′-TCG TTT TGT CGTTT GTC GTT-3′) proven to stimulate the immune respof human dendritic cells[32], or the ODN-CpG (5′-TCC ATGCG TTC CTG ATG CT-3′) that induces protection in micgainstL. major[31]. This adjuvant effect is mediated by t
nduction of innate immune mediators, especially IL-12[9]ia Toll receptor 9[33,34].
Intradermal inoculation ofpfr-2DNA proved to be highlymmunostimulatory in terms of reduced or increased leize compared with naive controls. The administratioNA by the intradermal route engages dendritic cells, wre potent antigen presenting cells[35,36]. The site of inocu
ation ofLeishmaniahas previously been shown to influenoth the outcome ofLeishmaniainfection [37,38] and the
mmune response elicited in hamsters[39,40]. Likewise, theissue site of inoculation of DNA vaccines has been shownfluence the outcome of immunization[41]. The differencen protective response induced by intradermal boostersaws or ears illustrates the crucial role that qualitative asf immunization can have on the recall response.
The results of this study demonstrate that paraflagellarotein ofLeishmania, despite being expressed principally
le BALB/c-L. majormodel[45,46]. Besides antigen comosition and presentation of the vaccine, the susceptibesistant phenotype of the experimental host, the route, tind magnitude of challenge infection must also be consid
n vaccine design and evaluation[47].
cknowledgments
This study was supported in part by grant ID 980NDP/WORLD BANK/WHO Special Programme fesearch and Training in Tropical Diseases (TDR), and CIENCIAS grant 2229-05-224-99, 2229-05-228-99 andiencias young investigator program CT #021-2000.echnical assistance of Diego Goyeneche in the innalysis of the immune response in hamsters are gratcknowledged. We thank Andres Hurtado, Ph.D. for his
iative and role in the design of the DNA vaccine foronduct of this study, and Mauricio Perez for his invaluassistance in the statistical analysis of the data.
eferences
[1] Reiner SL, Wang ZE, Hatam F, Scott P, Locksley RM. Th1Th2 cell antigen receptors in experimental leishmaniasis. Sc1993;259:1457–60.
[2] Raz E, Tighe H, Sato Y. Preferential induction of a TH1 inmunesponse and inhibition of specific IgE antibody formation by plasDNA immunization. Proc Natl Acad Sci USA 1996;93:5141–5.
[3] Walker PS, Shaorton-Kersten, et al. Genetic immunization withcoprotein 63 cDNA results in a helper T Cell type immune resp

994 N.G. Saravia et al. / Vaccine 23 (2005) 984–995
and protection in a murine model of leishmaniasis. Hum Gene Ther1998;9(13):1849–50.
[4] Damo X, Liew FY. Genetic vaccination against leishmaniasis. Vac-cine 1994;12:1534–6.
[5] Segura EL, Paulone J, Cerisola JA. Experimental Chagas’ diseaseprotective activity in relation with subcellular fractions of the para-site. J Parasitol 1976;62:131–3.
[6] Segura EL, Vasquez C, Bronzina A, Campos JM, Cerisola JA, CappaSM. Antigens of the subcellular fractions ofTrypanosoma cruziII:flagella and membrane fractions. J Protozol 1977;24:540–3.
[7] Ruiz AM, Esteva M, Cabeza Merkert P, Laguens RP, Segura EL.Protective immunity and pathology induced by inoculation of micewith different subcellular fractions ofTrypanosoma cruzi. Acta Trop1985;42:299–309.
[8] Wrightsman RA, Miller MJ, Saborio JL, Manning JE. Pure paraflag-ellar rod protein protects mice againstTrypanosoma cruziinfection.Infect Immun 1995;63:122–5.
[9] Miller MJ, Wrightsman RA, Manning JE.Trypanosoma cruzi: pro-tective immunity in mice immunized with paraflagellar rod pro-teins in associated with a T helper type Y response. Exp Parasitol1996;84:156–67.
[10] Miller MJ, Wrightsman RA, Striker GA, Manning JE. Protection ofM-mice againstTrypanosoma cruziby immunization with paraflag-ellar rod proteins requires T cell but not B cell, function. J Immunol1997;158:5330–7.
[11] Moore LL, Santrich C, Lebowitz JH. Stage specific expression of theLeishmania mexicanaparaflagellar rod protein PFR-2. Mol BiochemParasitol 1996;80:125–35.
[12] Maga JA, Sherwin T, Francis S, Gull K, LeBowitz JH. Geneticdissection of theLeishmaniaparaflagellar rod, a unique flagellar
[ mbly
[ tory9.
[ on--zool
[[
[ aviaand
[ pa-)
[ ec-
[ iting–65.
[ c-en-SA
[ f cu-.
[ tancein-male
[ iffer-
[26] Sharifi I, FeKri AR, Aflatonian MR, et al. Randomised vaccinetrial of single dose of killedLeishmania majorplus BCG againstanthroponotic cutaneous leishmaniasis in Bam. Lancet 1998;351:1540–3.
[27] Colmenares M, Kar S, Goldsmith-Pestana K, McMahon-PrattD. Mechanisms of pathogenesis: differences amongstLeishmaniaspecies. Trans R Soc Trop Med Hyg 2002;96(1):S3–7.
[28] Rachamim N, Jaffe CL. Pure protein fromLeishmania donovaniprotects mice against both cutaneous and visceral leishmaniasis. JImmunol 1993;150:2322–31.
[29] McSorley SJ, Xu D, Liew FY. Vaccine efficacy of Salmonella strainsexpressing glycoprotein 63 with different promoters. Infect Immun1997;65:171–8.
[30] Zimmerman S, Egeter O, Hausman S, et al. Cutting edge:CpG oligodeoxynucleotides trigger protective and curative Th1 re-sponses in lethal murine leishmaniasis. J Immunol 1998;160:3627–30.
[31] Pisetsky DS. Immune activation by bacterial DNA: a new geneticcode. Immunity 1996;5:303–10.
[32] Hartmann G, Weiner GJ, et al. CpG DNA: a potent signal for growth,activation, and maturation of human dendritic cell. Proc Natl AcadSci USA 1999;96(16):9305–10.
[33] Hemmi H, Takeuchi O, Kawai T, et al. Toll-like receptor recognizesbacterial DNA. Nature 2000;408:659–60.
[34] Bauer S, Kirschning CJ, Hacker H, et al. TLR9 confers responsive-ness to bacterial DNA via species specific CpG motif recognition.Proc Natl Acad Sci USA 2001;98:9237–42.
[35] Hengge UR, Chan EF, Foster RA, Walker PS, Vogel JC. Cytokinegene expression in epidermis with biological effects following injec-tion of naked DNA. Nat Genet 1995;10:161–6.
[ Anti-NA
viral
[cula-
[uno-is
[ex-mun
[ flu-t of
9–
[ de-DNA
[ ent ofNA-un
[ s of71–
[ dsV-1-
[ tec-with
cytoskeleton structure. J Cell Sci 1999;112:2753–63.13] LaemmLi UL. Cleavage of structural proteins during the asse
of the head of bacteriophage T4. Nature 1970;227:680–5.14] Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a labora
manual. New York: Cold Spring Harbor Laboratory Press; 19815] Ismach R, Cianci CM, Caulfield JP, Langer PJ, Hein A, McMah
Pratt D. Flagellar membrane and paraxial rod proteins ofLeishmania: characterization employing monoclonal antibodies. J Proto1989;36:617–24.
16] “His-Bind® Kits” Technical Bulletin 054 11/99B; Novagen Inc.17] Senekjie HA. Studies on the culture ofLeishmania tropica. Trans R
Soc Trop Med Hyg 1939;33:267–9.18] Travi BL, Osorio Y, Melby PC, Chandrasekar B, Arteaga L, Sar
NG. Gender is a major determinant of the clinical evolutionimmune response of hamsters infected withLeishmania vianniaspp.Infect Immun 2002;70:2288–96.
19] Martinez JE, Travi BL, Valencia AZ, Saravia NG. Metastatic cability of Leishmania (Viannia) panamensisandLeishmania (Vianniaguyanensisin golden hamsters. J Parasitol 1991;77(5):762–8.
20] Zeledon R, Soto R, Gonzalez G. Experimental superimposed inftion of the hamster withLeishmania mexicanaand L. braziliensis.Acta Trop 1982;39(4):367–72.
21] Ramshaw IA, Ramsay AJ. The prime-boost strategy: excprospects for improved vaccination. Immunol Today 2000;21:16
22] Letvin NL, Montefiori DC, Yasutomi Y, et al. Potent, protetive anti-HIV immune responses generated by bimodal HIVvelope DNA plus protein vaccination. Proc Natl Acad Sci U1997;19(94):9378–83.
23] Giannini MSH. Sex-influenced response in the pathogenesis otaneous leishmaniasis in mice. Parasite Immunol 1986;8:31–7
24] Satoskar A, Al-Quassi HH, Alexander J. Sex-determined resisagaintsLeishmania mexicanais associated with the preferentialduction of a Th1-like response and IFN-gamma production by febut no male DBA/2 mice. Immunol Cell Biol 1998;76:159–66.
25] Satoskar A, Alexander J. Sex-determined susceptibility and dential IFN-gamma and TNF-alpha mRNA expresion in DBA/2 miceinfected withLeishmania mexicana. Inmunology 1995;84:1–4.
36] Casares S, Inaba K, Brumeanu TD, Steinman RM, Bona CA.gen presentation by dendritic cells after immunization with Dencoding a major histocompatibility complex class II restrictedepitope. J Exp Med 1997;186:1481–6.
37] Kirkpatrick CE, Nolan TJ, Farrell JP. Rate ofLeishmaniainducedskin-lesion development in rodents depends on the site of inotion. Parasitology 1987;94:451–65.
38] Osorio Y, Melby P, Pirmez C, Chandrasekar B, Guarın N, Travi BL.The site of cutaneous infection influences the clinical and immlogical response of hamsters infected withLeishmania panamens.Parasite Immunol 2003;25:139–48.
39] Nabors GS, Farrell JP. Site-specific immunity toLeishmania inSWR mice: the site of infection influences susceptibility andpression of the antileishmanial immune response. Infect Im1994;62:3655–62.
40] Nabors GS, Nolan T, Croop W, Li J, Farrell JP. The inence of the site of parasite inoculation on the developmenTh1 and Th2 type immune responses in (BALB/c× C57BL/6)F1 mice infected withL. major. Parasite Immunol 1995;17:5679.
41] Torres CA, Iwasaki A, Barber BH, Robinson HI. Differentialpendence on target site tissue for gene gun and intramuscularimmunization. J Immunol 1997;158:529–32.
42] Charo J, Sundback M, Wasserman K, et al. Marked enhancemthe antigen specific immune response by combining plasmid Dbased immunization with a Schiff base-forming drug. Infect Imm2002;70:6652–7.
43] Stanley MA. Imiquimod and the imidazoquinolones: mechanismaction and therapeutic potential. Clin Exp Dermatol 2002;27:57.
44] Lore K, Betts MR, Brenchley JM, et al. Toll-like receptor liganmodulate dendritic cells to augment cytomegalovirus and HIspecific T cell responses. J Immunol 2003;171:4320–8.
45] Rafati S, Salmanian AH, Taheri T, Vafra M, Fasel N. A protive cocktail vaccine against murine cutaneous leishmaniasisDNA encoding cysteine proteinases ofLeishmania major. Vaccine2001;19:369–75.

N.G. Saravia et al. / Vaccine 23 (2005) 984–995 995
[46] Campos-Neto A, Webb JR, Greeson K, Coler RN, Skeiky YA, ReedSG. Vaccination with plasmid DNA encoding TSA/LmSTI1 leishma-nial fusion proteins confers protection againstLeishmania majorin-fection in susceptible BALB/c mice. Infect Immun 2002;70:2828–36.
[47] Mendez S, Gurunathan S, Kamhawi S, et al. The potency anddurability of DNA- and protein-based vaccines againstLeishmaniamajor evaluated using low-dose, intradermal challenge. J Immunol2001;166:5122–8.




















