
Coordinated expression and immunogenicity of an outer ...
10
Coordinated expression and immunogenicity of an outer membrane protein from Salmonella enterica serovar Typhi under iron limitation, oxidative stress and anaerobic conditions V. Chanana 1 , S. Majumdar 2 , P. Ray 3 , M. Sharma 3 & P. Rishi 1, * 1 Department of Microbiology, Panjab University, Chandigarh, 160014, India; 2 Department of Experimental Medicine and Biotechnology, Post Graduate Institute of Medical Education and Research (PGIMER), Chandigarh, 160012, India; 3 Department of Medical Microbiology, Post Graduate Institute of Medical Education and Research (PGIMER), Chandigarh, India Received 22 July 2005; accepted 24 October 2005 ȑ 2005 National Science Council, Taipei Key words: expression, Salmonella typhi, stress protein Summary Successful pathogens overcome the environmental stresses by the coordinated expression of various genes and eventually proteins. Since, the surface of the microbe is likely to come in contact with the host initially, an attempt was made to identify the outer membrane proteins (OMPs), if any, which may get expressed under more than one environmental conditions simulating the in vivo ones. In the present study, Salmonella enterica serovar Typhi was grown under iron-limited, oxidative stress as well as anaerobic conditions and the OMP profiles were compared. A 69 kDa OMP was found to express with enhanced intensity under the selected stress conditions in comparison to normal conditions. The phenotypic simi- larity among the proteins was assessed on the basis of their molecular weight, cross reactivity and HPLC. The protein expressed under oxidative stress and anaerobic conditions reacted with the antibodies raised against iron-regulated outer membrane protein (IROMP), indicating the sharing of at least some of the epitopes. A single peak observed after subjecting the pooled 69 kDa protein sample and appearance of a single band on SDS-PAGE thereafter, confirmed the purity and phenotypic similarity of the 69 kDa OMP. Reactivity of pooled 69 kDa protein with 85% of sera from typhoid patients revealed the in vivo expression of this protein. The results of this study indicate the coordination of this phenotype under iron stress, oxidative stress and anaerobic conditions. In view of the expression of the 69 kDa protein under the selected stress conditions and their in vivo immunogenicity, these findings may be relevant for the better understanding of the host–microbe interactions and for the further development of diagnostic and pre- ventive strategies. Introduction Typhoid fever is a human disease caused by Salmonella enterica serovar Typhi, a facultative, intracellular pathogen. This disease remains a major public health problem due to significant morbidity and mortality worldwide and therefore, continues to be a priority of the World Health Organization. After S. typhi is ingested through contaminated food and/or water, the organism migrates to distal ileum. Presumably, S. typhi, then adheres to and invades the intestinal epithelium, migrates to the underlying Peyer’s patches and disseminates throughout the reticuloendothelial system [1]. However, little is known about the *To whom correspondence should be addressed. Fax: +91-172- 2541770; E-mail: [email protected] Journal of Biomedical Science (2006) 13:303–312 303 DOI 10.1007/s11373-005-9047-5
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Coordinated expression and immunogenicity of an outer ...

Coordinated expression and immunogenicity of an outer membrane proteinfrom Salmonella enterica serovar Typhi under iron limitation, oxidativestress and anaerobic conditions
V. Chanana1, S. Majumdar2, P. Ray3, M. Sharma3 & P. Rishi1,*1Department of Microbiology, Panjab University, Chandigarh, 160014, India; 2Department of ExperimentalMedicine and Biotechnology, Post Graduate Institute of Medical Education and Research (PGIMER),Chandigarh, 160012, India; 3Department of Medical Microbiology, Post Graduate Institute of MedicalEducation and Research (PGIMER), Chandigarh, India
Received 22 July 2005; accepted 24 October 2005
� 2005 National Science Council, Taipei
Key words: expression, Salmonella typhi, stress protein
Summary
Successful pathogens overcome the environmental stresses by the coordinated expression of various genesand eventually proteins. Since, the surface of the microbe is likely to come in contact with the hostinitially, an attempt was made to identify the outer membrane proteins (OMPs), if any, which may getexpressed under more than one environmental conditions simulating the in vivo ones. In the present study,Salmonella enterica serovar Typhi was grown under iron-limited, oxidative stress as well as anaerobicconditions and the OMP profiles were compared. A 69 kDa OMP was found to express with enhancedintensity under the selected stress conditions in comparison to normal conditions. The phenotypic simi-larity among the proteins was assessed on the basis of their molecular weight, cross reactivity and HPLC.The protein expressed under oxidative stress and anaerobic conditions reacted with the antibodies raisedagainst iron-regulated outer membrane protein (IROMP), indicating the sharing of at least some of theepitopes. A single peak observed after subjecting the pooled 69 kDa protein sample and appearance of asingle band on SDS-PAGE thereafter, confirmed the purity and phenotypic similarity of the 69 kDaOMP. Reactivity of pooled 69 kDa protein with 85% of sera from typhoid patients revealed the in vivoexpression of this protein. The results of this study indicate the coordination of this phenotype under ironstress, oxidative stress and anaerobic conditions. In view of the expression of the 69 kDa protein under theselected stress conditions and their in vivo immunogenicity, these findings may be relevant for the betterunderstanding of the host–microbe interactions and for the further development of diagnostic and pre-ventive strategies.
Introduction
Typhoid fever is a human disease caused bySalmonella enterica serovar Typhi, a facultative,intracellular pathogen. This disease remains amajor public health problem due to significant
morbidity and mortality worldwide and therefore,continues to be a priority of the World HealthOrganization. After S. typhi is ingested throughcontaminated food and/or water, the organismmigrates to distal ileum. Presumably, S. typhi, thenadheres to and invades the intestinal epithelium,migrates to the underlying Peyer’s patches anddisseminates throughout the reticuloendothelialsystem [1]. However, little is known about the
*To whom correspondence should be addressed. Fax: +91-172-2541770; E-mail: [email protected]
Journal of Biomedical Science (2006) 13:303–312 303DOI 10.1007/s11373-005-9047-5

bacterium–host interactions that occur at each ofthese steps in the pathogenesis of S. typhi.
Like other enteric pathogens, Salmonella typhiencounters iron limitation during the diseaseprocess, as the human body has different ironwithholding defense systems, preventing theorganism from acquiring essential iron for surviv-ing inside the host [2]. Besides iron limitation,availability of oxygen, is yet another stressencountered by S. typhi while using molecularoxygen for respiration or oxidation of nutrientsto obtain energy and within the intracellular milieuof macropahge-phagolysosomes as well. Thisresults in continuous generation of reactive oxygenspecies, such as anion radical (O_
2), hydrogenperoxide (H2O2) and highly reactive hydroxylradicals (_OH)) [3]. Anaerobic conditions are alsofrequently encountered by pathogens invading thegastrointestinal tract due to low/limiting oxygenconditions prevalent in the small intestine [4, 5].However, the regular interaction of the host andthe pathogen causes the latter to express certainregulons for sensing the host environments anddevelop survival strategies against the environ-mental stresses [2, 6]. Although, a number of genesincluding fur, sox R/S, mar A, oxy R have beenidentified, for combating these stresses [7–10],there are not many studies indicating the identifi-cation and localization of the proteins involved inpathogenesis under these conditions.
There is some indication from the existingliterature that if stress tolerance occurs in abacterium when exposed to a single stressor, crossprotection can be manifested across multiplestresses and general tolerance of these environ-ments is increased [10, 11]. Iron responsive regu-lator Fur has been found to be directly orindirectly involved in regulation of oxidative stressdefense [12]. Iron regulation has been reported toconfer resistance against oxidative stress too, asthe organisms are also exposed to extracellularoxygen metabolites of stimulated polymorphonu-clear phagocytes in the infected gut. The limitedavailability of iron in mucosal secretions, in tissuesand within the phagocytes, due to the bound ironis well known. The link between iron metabolismand oxidative stress is further demonstrated by theresults that the primary regulators of the oxidativestress response in E. coli i.e. OxyR and SoxRSinduce the synthesis of Fur and that the E. coli furmutant is hypersensitive to oxidative stress [3, 9]. It
is clear from these studies that the pathogensevolve strategies to overcome the stresses bycoordinated expression of genes and proteins.However, very little is known about the coordina-tion of phenotype(s) expression and availability ofiron and oxygen particularly with respect to outermembrane proteins.
The outer membrane proteins of bacteria func-tion as the dynamic interface between the bacte-rium and its surroundings and are involved in themaintenance of cell structure, binding a variety ofsubstances adhesion to other cells, and regulationof transport of both nutrients and bactericidalagents [13]. There is a vast amount of informationabout OMPs and their roles in virulence, inflam-mation, immunogenicity and anti-microbial resis-tance [14–16]. However, the knowledge about theexpression of OMPs of the bacteria, in reference tocoordination of iron and oxygen availability islargely not available. The present study wastherefore; carried out to evaluate the coordinatedexpression of signal sensing phenotype(s) in ser-ovar Typhi under the environmental conditions tomimic the in vivo environmental milieu.
Materials and methods
Animals
Young male New Zealand white rabbits (1–2 kg inweight) used for raising anti-sera, were procuredfrom Central Animal House, Panjab University,Chandigarh. Care and use of laboratory animalswas followed, in accordance with the guidelines ofthe institutional ethical committee.
Microorganisms
The standard strain of S. enterica serovar Typhi(Ty2) procured from CRI Kasauli, India, was usedin the present study. The strain was checked for itspurity and was characterized biochemically as wellas serologically. It was maintained in 10% glycerolbroth and also stored as lyophilized ampules.
Growth conditions
Bacteria was grown on nutrient agar platesand single colony was used to prepare seedculture. 0.1% inoculum from seed culture was
304

used for growing the bacteria under stressconditions.
Iron stress
Cells were grown in nutrient broth under iron-depleted conditions created by adding differentconcentrations of iron chelator 2,2¢-dipyridyl (100,150, 200 and 250 lM) in accordance with themethod originally described by Fernandez-Beroset al. [17]. Iron content of the media before andafter chelation was measured by the method ofCarter [18]. The concentration of dipyridyl thatcould induce the maximum expression of IR-OMP(s) was chosen for the subsequent experi-ments. Iron-sufficient conditions were created byadding ferrous sulphate in growth medium usingthe concentration comparable to the concentrationof the iron chelator.
Oxidative stress
For the oxidative stress, S. Typhi cells wereexposed to different concentrations of hydrogenperoxide (10, 15 and 20 mM) in accordance withthe method of Ozkanca et al. [19]. The cells weregrown in nutrient broth and hydrogen peroxidewas added to the culture medium in the earlylogarithmic phase. The cells were allowed to growovernight at 37 �C, 150 rpm and were harvested inthe stationary phase. The concentration of H2O2
that could induce the maximum expression ofoxidative stress induced OMP(s) was chosen forthe subsequent experiments.
Anaerobic stress
The bacteria were cultured by growing in nutrientbroth, under anaerobic conditions by the methodof Schimann and Shope [20]. Anaerobic conditionswere maintained by placing the culture in Mack-intosh anaerobic chamber in the presence of 85%nitrogen, 10% H2 and 5% CO2. The chamber waskept at 37 �C overnight and the cells were har-vested.
Combined stress
The bacteria were subjected to the combined stressof iron and oxygen by growing the cells in nutrientbroth under iron depleted conditions created by
the addition of 2,2¢,-dipyridyl as described earlierand S. Typhi cells were then exposed to hydrogenperoxide (15 mM) in the early logarithmic phaseof growth as described by the method of Ozkancaet al. [19]. The cells were allowed to grow over-night at 37 �C, 150 rpm and were harvested in thestationary phase.
Extraction of OMPs
To study the expression of stress-induced proteins,OMPs were prepared according to the methoddescribed by Chander et al. [21]. Briefly, cells weregrown under selected stress conditions as describedearlier. Bacterial growth was harvested and thepellet obtained was suspended in 20 mM Tris–HClbuffer (pH 7.6) containing 5 mM MgCl2. Cellswere harvested twice and suspended in the samebuffer containing 2 mM phenyl methyl sulphonylfluoride (PMSF, Sigma, USA). Cells were thendisrupted by ultrasonication and undisruptedmaterial was removed by low speed centrifugation.Supernatant was then ultracentrifuged (BeckmanCoulter, Optima TM XL-100K Ultracentrifuge,USA) at 100,000� g for 60 min at 4 �C and thepellet was suspended in 1% sodium lauryl sarcosyl(ICN, USA) (in 20 mM Tris–HCl buffer, pH 7.6)for 2 h at 37 �C. Detergent insoluble OMP frac-tion was collected by ultracentrifugation at thesame speed.
Analysis of OMP profile
The protein content in each preparation wasestimated by the method of Markwell and Tolbery[22]. The proteins were separated on SDS-PAGEas described by Laemmli [23]. The molecularweight of the protein(s) expressed with enhancedintensity under iron stress, oxidative stress as wellas anaerobic stress conditions was determined bycalculating the relative front (Rf) values in com-parison to that of standard molecular weightmarkers [phosphorylase B (97 kDa), bovine serumalbumin (66 kDa), ovalbumin (43 kDa) and car-bonic anhydrase (29 kDa)]. Densitometric analysisof stress induced OMPs was determined usingGENE TOOL software on GEL DOCUMENTA-TION (Syngene, USA). For this, one of theprotein bands in the marker lane was taken as 1.Then the protein of interest expressed with
305

enhanced intensity was quantified as how manytimes of the marker band.
Elution of stress induced OMP from polyacrylamidegels
Stress induced OMPs were eluted out of thepreparative gel by the method of Hager andBurgess [24]. Briefly, OMPs extracted from path-ogen grown under different stress conditions asdescribed earlier, were resolved on SDS-PAGEand the region expressing stress induced OMP wasexcised from the gel. Gel section was chopped intosmall pieces and crushed. Proteins were eluted byincubating the crushed mass of gel with elutionbuffer for overnight at 4 �C. The mixture wascentrifuged and the supernatant was then trans-ferred to a fresh vial. Four volumes of ice-coldacetone was added to the gel eluate and the samplewas allowed to precipitate for 30 min at )20 �C.The precipitation was then centrifuged (27,000� g,10 min) and supernatant was drained out. Therecovery of the protein was checked on SDS-PAGE by silver staining.
Phenotypic similarity amongst the stresses inducedproteins
Phenotypic similarity amonst the stress inducedproteins was determined on the basis of (i)Identical molecular weight: for this, the bandscorresponding to the protein of interest wereexcised and eluted from respective gels [24] asdescribed before. The three proteins were thenpooled and run on the SDS-PAGE again. The gelwas silver stained by the method of Budowle et al.[25] (ii) Cross reactivity: for this, the anti-serumagainst iron regulated outer membrane protein(IROMP) was raised in rabbits according to Claireet al. [26]. The titre of the antibodies was checkedby ELISA [27]. Cross reactivity of oxidative andanaerobic stress induced proteins with IROMPwas assessed using anti-IROMP anti-serum andanti-rabbit IgG HRP conjugate (Dako Inc. USA)by Western blotting [28]; and (iii) HPLC: for this,the selected stress induced outer membrane pro-teins were pooled and subjected to high perfor-mance liquid chromatography (HPLC)(Shimadzu). Analytical details for HPLC included;Column – Shim-pack DIOL-150, 7.9 mmID� 25 cm; Mobile phase – 0.2 M sodium sulfate
in 0.01 M phosphate buffer, pH 7.0; Flow rate –1.0 ml/min; Temperature – 22 �C; Detector –280 nm.
In-vivo expression of the stress-induced phenotype
In-vivo immunogenicity of the selected stress-induced OMP was detected by Western blotting,using sera from 20 typhoid fever patients (Widalpositive sera). The sera from six apparentlyhealthy individuals were used as controls. Forthis, outer membrane proteins extracted fromS. Typhi grown under iron stress, oxidative stressas well as anaerobic stress were resolved on SDS-PAGE. Resolved proteins were transferred ontothe nitrocellulose paper (NCP) as described previ-ously [28] at 200 mA for 3 h. The NCP containingthe transferred antigen (OMPs) was blocked with5% (w/v) skimmed milk in PBS for 2 h at 37 �C.Thereafter, NCP was incubated with 1:1500diluted anti-human IgG HRP conjugate (DakoInc. USA) for 1 h at 37 �C. Strips were thenwashed with washing solution for 3–4 times andthen with PBS. The developing was done with3,3¢diaminobenzidine (DAB) (Sigma Chemical Co.USA) acting as a substrate (5 mg DAB in 10 mlPBS with 5 ll H2O2). The reaction was stoppedwith double distilled water.
Results
Analysis of outer membrane proteins
A number of proteins were found to be expressedby S. Typhi as assessed by SDS-PAGE. Onanalyzing the protein profiles, it was interestingto note that a protein having molecular weight of69 kDa (determined by calculating Rf value) wasgetting expressed with high intensity under all theselected stress conditions. On comparing theexpression of this protein under different concen-trations of dipyridyl, it was found that theexpression was maximum at 200 lM, than at 150or 250 lM concentration of dipyridyl (Figure 1).On exposing the bacteria to different concentra-tions of H2O2 (10, 15 and 20 mM), the expressionof the 69 kDa OMP was found to be highestat 15 mM concentration of H2O2 (Figure 2).Enhanced expression of the 69 kDa phenotypewas also observed when S. Typhi was grown
306
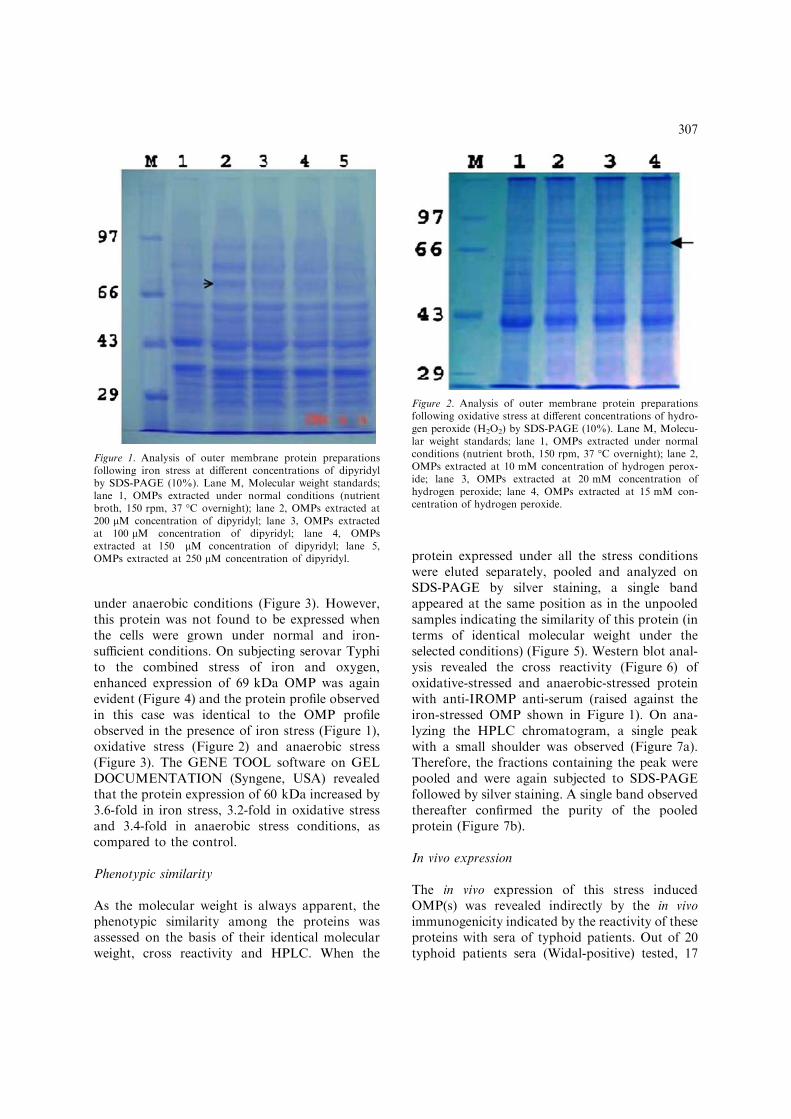
under anaerobic conditions (Figure 3). However,this protein was not found to be expressed whenthe cells were grown under normal and iron-sufficient conditions. On subjecting serovar Typhito the combined stress of iron and oxygen,enhanced expression of 69 kDa OMP was againevident (Figure 4) and the protein profile observedin this case was identical to the OMP profileobserved in the presence of iron stress (Figure 1),oxidative stress (Figure 2) and anaerobic stress(Figure 3). The GENE TOOL software on GELDOCUMENTATION (Syngene, USA) revealedthat the protein expression of 60 kDa increased by3.6-fold in iron stress, 3.2-fold in oxidative stressand 3.4-fold in anaerobic stress conditions, ascompared to the control.
Phenotypic similarity
As the molecular weight is always apparent, thephenotypic similarity among the proteins wasassessed on the basis of their identical molecularweight, cross reactivity and HPLC. When the
protein expressed under all the stress conditionswere eluted separately, pooled and analyzed onSDS-PAGE by silver staining, a single bandappeared at the same position as in the unpooledsamples indicating the similarity of this protein (interms of identical molecular weight under theselected conditions) (Figure 5). Western blot anal-ysis revealed the cross reactivity (Figure 6) ofoxidative-stressed and anaerobic-stressed proteinwith anti-IROMP anti-serum (raised against theiron-stressed OMP shown in Figure 1). On ana-lyzing the HPLC chromatogram, a single peakwith a small shoulder was observed (Figure 7a).Therefore, the fractions containing the peak werepooled and were again subjected to SDS-PAGEfollowed by silver staining. A single band observedthereafter confirmed the purity of the pooledprotein (Figure 7b).
In vivo expression
The in vivo expression of this stress inducedOMP(s) was revealed indirectly by the in vivoimmunogenicity indicated by the reactivity of theseproteins with sera of typhoid patients. Out of 20typhoid patients sera (Widal-positive) tested, 17
Figure 2. Analysis of outer membrane protein preparationsfollowing oxidative stress at different concentrations of hydro-gen peroxide (H2O2) by SDS-PAGE (10%). Lane M, Molecu-lar weight standards; lane 1, OMPs extracted under normalconditions (nutrient broth, 150 rpm, 37 �C overnight); lane 2,OMPs extracted at 10 mM concentration of hydrogen perox-ide; lane 3, OMPs extracted at 20 mM concentration ofhydrogen peroxide; lane 4, OMPs extracted at 15 mM con-centration of hydrogen peroxide.
Figure 1. Analysis of outer membrane protein preparationsfollowing iron stress at different concentrations of dipyridylby SDS-PAGE (10%). Lane M, Molecular weight standards;lane 1, OMPs extracted under normal conditions (nutrientbroth, 150 rpm, 37 �C overnight); lane 2, OMPs extracted at200 lM concentration of dipyridyl; lane 3, OMPs extractedat 100 lM concentration of dipyridyl; lane 4, OMPsextracted at 150 lM concentration of dipyridyl; lane 5,OMPs extracted at 250 lM concentration of dipyridyl.
307

sera were recognized by all the three stress-inducedOMP(s) (Figure 8). In contrast, none of the sixnormal sera from apparently healthy individualsreacted with these proteins.
Discussion
The pathogen is exposed to a variety of hostenvironmental conditions that may influence thefunctional capacity. Such environmental cuesinclude acidic pH, elevated temperature, oxidativestress, different osmotic strengths and the presenceof cationic peptides as well as the availability ofions and nutrients. As the infection proceeds, theexpression of the virulence determinants varieswith time and anatomical sites [6]. Therefore, our
hypothesis is to identify the set of proteins, whichmay get expressed under different environmentalconditions with an ultimate aim to develop apreventive intervention.
Several forms of stresses increase the rate ofsynthesis of stress genes and subsequently proteinsexpressed under different stress conditions. Inresponse to the generation of resistance mecha-nisms in bacteria when exposed to a particularstress, cross protection has also been shown tomanifest across multiple stresses [10, 11]. Baillonet al. [29] have shown that iron regulated proteinsof Campylobacter jejuni are involved in its defense
Figure 4. Analysis of outer membrane protein preparations ofS. Typhi subjected to the combined stress of iron (200 lMdipyridyl) and oxidative (15 mM H2O2) by SDS-PAGE(10%). Lane M, Molecular weight standards; lane 1, OMPsextracted under normal conditions (nutrient broth, 150 rpm,37 �C overnight); lane 2 & 3, OMPs extracted from S. Typhisubjected to the combined iron and oxidative stress.
Figure 3. Analysis of outer membrane protein preparations ofS. Typhi, grown under anaerobic conditions, by SDS-PAGE(10%). Lane M, Molecular weight standards; lane 1, OMPsextracted under normal conditions (nutrient broth, 150 rpm,37 �C overnight); lane 2 & 3, OMPs extracted under anaero-bic conditions.
308

against oxidative stress. Rocha et al. [30] haveshown that response to oxidative stress in anaer-obically grown B. fragilis induces the synthesis ofat least 28 proteins in the presence of H2O2 thatconfers protection to the organism against thehydrogen peroxide induced oxidative damage.Pathogens have been reported to combat thesestress conditions through the induction of variousiron-regulated proteins [31, 16], oxidative stressproteins [19, 32] and anaerobic stressed proteins[33]. However, the coordination of phenotype(s)and their immunodominance under iron andoxygen availability have not been studied in detail.The present study revealed the expression, pheno-typic similarity and immunodominance of the69 kDa signal sensing phenotype under thesestress conditions. This protein may be one of thefactors, which may help the organism to combatthe stresses offered by the host.
Enhanced expression of the 69 kDa OMP in allthe selected stresses is suggestive of the hypothesisthat stress may be responsible for a signal, whichinduces the synthesis of this protein, which thencrosses the inner membrane to become associatedwith outer membrane. Fernandez-Beros et al. [17]and Baumler et al. [34] have also reported thatiron regulated proteins observed in their studywere also associated with the outer membrane.Cross reactivity of anti-IROMP anti-sera withboth the oxidative and anaerobic stressed proteins
indicates the sharing of atleast some of theepitopes expressed on these proteins under thestressed conditions. A single peak after subjectingthe pooled 69 kDa protein further confirmed thephenotypic similarity among the 69 kDa OMPexpressed under all the selected stress conditionsand its purity. The ability of the organism toswitch on a common regulatory network suggestthat there is a class of ‘stress regulated’ genes thatare regulated by common sensing mechanism inresponse to different environmental signals [6, 35].The results of the present study indicating theoverlapping expression of proteins under differentstress conditions are in agreement with earlierstudies of Chander et al. [21] and Ramamoorthyand Schoel-Meeker [36]. The results suggest thatexpression of proteins does not depend on single
Figure 6. Western blot analysis showing the cross-reactivityof IROMP with oxidative, anaerobic and combined (iron andoxidative) stress induced protein using anti-IROMP antibod-ies. Lane 1, Iron stress protein; lane 2, oxidative stress pro-tein; lane 3, anaerobic stress protein; lane 4, combined (ironand oxidative) stress protein.
Figure 5. Analysis of stress induced 69 kDa OMP under dif-ferent conditions by SDS-PAGE (10%) after chemical elu-tion. Lane M, Molecular weight standards; lane 1, IROMP;lane 2, oxidative stress outer membrane protein; lane 3,anaerobic stress outer membrane protein; lane 4, OMP ex-tracted under iron and oxidative combined stress; lane 5, poolof the above stress proteins.
309

environmental stimulus in host but there may bean interaction among stimuli. Expression of69 kDa phenotype under iron, oxidative andanaerobic stress conditions and their cross reac-tivity raises the question to confirm their identityin terms of commonality. Hence, our future goal isto compare their N-terminal sequences and toevaluate their genetic expression under the selectedconditions.
The in-vivo expression of this stress protein isindicated indirectly by its reactivity with the seraof typhoid fever patients. These results are inagreement with the earlier studies where IROMPsin the serovar Typhi were found to be immuno-genic in-vivo [17]. Non-reactivity of two seraobserved in our study can be due to the undefinedstage of illness at the time of sample collection.Although the sera collected from patients sufferingwith pyrexia (but widal test negative) did not reactwith this protein, but sera from non-typhi Salmo-nella bacteremia patients along with a largernumber of typhoid patient sera need to beexplored to determine the specificity and thediagnostic utility of this protein.
In view of the expression of 69 kDa OMPunder iron limitation, oxidative stress as well asanaerobic conditions and its reactivity with thetyphoid patient sera, it may be an importantfactor, induced during the evolution of salmonel-losis. This finding may be relevant for the betterunderstanding of the disease pathophysiology andfor the future developments of diagnostic andpreventive strategies.
Acknowledgements
The authors wish to thank Indian Council ofMedical Research (ICMR), New Delhi, India,for providing financial assistance to carry outthis project.
Figure 8. Immuno detection of S. Typhi stress induced outermembrane proteins with serum from patients with typhoid fe-ver. lane 1, Normal outer membrane proteins; lane 2, ironstress proteins (IROMPs); lane 3, oxidative stress proteins;lane 4, combined (iron and oxidative) stress proteins. lane 5,anaerobic stress proteins; fi , indicates 69 kDa protein; ����>,indicates porins and other proteins.
Figure 7. (a) Auto-scaled HPLC chromatogram of 69 kDa stress protein. (b) Silver staining of purified 69 kDa protein after HPLC.
310

References
1. Hornick R.B., Greisman S.E., Woodward T.E., DupontH.L., Dawkins A.T. and Synder M.J., Typhoid fever:pathogenesis and immunological control. N. Engl. J. Med.283: 739–746, 1970.
2. Ratledge C. and Dover L.G., Iron metabolism in patho-genic bacteria. Annu. Rev. Microbiol. 54: 881–941, 2000.
3. Farr S.B. and Kogoma T., Oxidative stress responses inEscherichia coli and Salmonella typhimurium. Microbiol.Rev. 55: 561–585, 1991.
4. Khullar M., Singh R.D., Smriti M. and Ganguly N.K.,Anaerobiosis induced virulence of Salmonella entericasubsp. enterica serovar Typhimurium: role of phospholi-pase C signaling cascade. J. Med. Microbiol. 52: 741–745,2003.
5. Krishnan H.H., Ghosh A., Paul K. and Chowdhury R.,Effect of anaerobiosis on expression of virulence factors inVibrio cholerae. Infect. Immun. 72: 3961–3967, 2004.
6. Smith H., What happens to bacterial pathogens in vivo.Trends Microbiol. 6: 239–243, 1998.
7. Harvie D.R., Vilchez S., Steggles J.R. and Ellar D.J.,Bacillus cereus Fur regulates iron metabolism and isrequired for full virulence. Microbiology 151: 569–577,2005.
8. Kozyrev D.P. and Vasinova N.A., The role of ironregulated genes in microbial pathogenesis. Tsitologiia 46:465–473, 2004.
9. Straaten T., Zulianello L., Diepen A., Granger D.L.,Janssen R. and Dissel J.T., Salmonella enterica serovarTyphimurium RamA, intracellular oxidative stress re-sponses, and bacterial virulence. Infect. Immun. 72: 996–1003, 2004.
10. Foster J.W. and Spector M.P., How Salmonella surviveagainst the odds. Annu. Rev. Microbiol. 49: 145–174, 1995.
11. Archer D.L., Preservation microbiology and safety: evi-dence that stress enhances virulence and triggers adaptivemutations. Trends Food. Sci. Technol. 7: 91–95, 1996.
12. Ernst F.D., Homuth G., Stoof J., Mader U., Waidner B.,Kuipers E.J., Kist M., Kusters J.G., Bereswill S. and VanVliet A.H., Iron-responsive regulation of the Helicobacterpylori iron-cofactored superoxide dismutase SodB is med-iated by Fur. J. Bacteriol. 187: 3687–3692, 2005.
13. Wexler H.M., Outer-membrane pore-forming proteins ingram-negative anaerobic bacteria. Clin. Infect. Dis. 35:65–71, 2002.
14. Sood S., Rishi P., Dhawan V., Sharma S. and GangulyN.K., Protection mediated by antibodies to iron-regulatedouter membrane proteins of S. typhi in a mouse peritonitismodel. Mol. Cell. Biochem. 273: 421–423, 2005.
15. Sood S., Rishi P., Vohra H., Sharma S. and Ganguly N.K.,Cellular immune response induced by Salmonella entericaserotype Typhi iron-regulated outer-membrane proteins atperipheral and mucosal levels. J. Med. Microbiol. 54:815–821, 2005.
16. Chanana V., Seghal R. and Rishi P., Salmonella typhi iron-regulated outer-membrane proteins cause oedema andhyperalgesia during inflammation induced in a rat model.J. Med. Microbiol. 54: 421–423, 2005.
17. Fernandez-Beros M.E., Gonzalez C., McIntosh M.A. andCabello F.C., Immuneresponse to the iron deprivationinduced proteins of S. typhi in typhoid fever. Infect.Immun. 57: 1271–1275, 1989.
18. Carter P., Spectrophotometric determination of serum ironat the submicrogram level with a new reagent (Ferrozine).Anal. Biochem. 40: 450–458, 1971.
19. Ozkanca R., Sahin N., Isik K., Kariptas E. and Flint K.P.,The effect of toluidine blue on the survival, dormancy andouter membrane porin proteins (OmpC and OmpF) andSalmonella typhimurium LT2 in seawater. J. Appl. Micro-biol. 92: 1097–1104, 2002.
20. Schimann D.A. and Shope S.R., Anaerobic growth ofSalmonella typhimurium results in increased uptake byHenle 407 epithelial and mouse peritoneal cells in vitro andrepression of a major outermembrane protein. Infect.Immun. 59: 437–440, 1991.
21. Chander H., Majumdar S., Sapru S. and Rishi P.,Reactivity of typhoid patients sera with stress induced55 kDa phenotype in Salmonella enterica serovar Typhi.Mol. Cell. Biochem. 267: 75–82, 2004.
22. Markwell M.A.K. and Tolberi N.E., A modification of theLowry procedure to simplify protein determination inmembrane and lipoprotein samples. Anal. Biochem. 87:206–210, 1978.
23. Laemmli U.K., Cleavage of structural proteins duringthe assembly of bacteriophage T4. Nature 227: 680–685,1970.
24. Hager D.A. and Burgess R.R., Elution of proteins fromSDS-PAGE gels. Removal of SDS and renaturation ofenzyme activity. Anal. Biochem. 109: 76–86, 1980.
25. Budowle B., Chakraborty R., Giusti A.M., Eisenberg A.J.and Allen R.C., Analysis of VNTR locus DIS 80 by thePCR followed by high resolution PAGE. Am. J. Hum.Genet. 48: 137–144, 1991.
26. Claire H., Fabrice P., Waligora-Dupriet Anne-Jubith,Anne C., Marie-Claude B. and Pierre B., GroEL (HSP60) of Clostridium difficile is involved in cell adherence.Microbiology 147: 87–96, 2001.
27. Panezzuti H., James O., Hanson E.J., Choi Y., HarkmessR.E., Klein M.H. and Chang P., Identification of surfaceexposed B cell epitopes recognized by Haemophilus influ-enzae type b, P, specific monoclonal antibodies. Infect.Immun. 61: 1867–1872, 1993.
28. Towbin H., Staachelin T. and Gordon J., Electrophoretictransfer of proteins from acrylamide gels onto nitrocellu-lose sheet: procedure and some applications. Proc. Natl.Acad. Sci. USA 76: 4350–4354, 1979.
29. BaillonM.L.,VanVlietA.H.,KetleyJ.M.,ConstantinidouC.and Penn C.W., An iron-regulated alkyl hydroperoxidereductase (AhpC) confers aerotolerance and oxidative stressresistance to the microaerophilic pathogen Campylobacterjejuni. J. Bacteriol. 181: 4798–4804, 1999.
30. Rocha E.R., Selby T., Coleman J.P. and Smith C.J., Theoxidative stress response in an anaerobe, Bacteroidesfragilis: a role for catalase in protection against hydrogenperoxide. J. Bacteriol. 178: 6895–6903, 1996.
31. RabschW.,MethnerU.,VoigtW.,TschapeH.,ReissbrodtR.andWilliamsP.H.,RoleofreceptorproteinsforEnterobactinand 2,3-dihydroxybenzoylserine in virulence of Salmonellaenterica. Infect. Immun. 71: 6953–6961, 2003.
32. Rocha E.R. and Smith C.J., Characterization of a peroxideresistant mutant of the anaerobic bacterium Bacteroidesfragilis. J. Bacteriol. 180: 5906–5912, 1998.
33. Kapoor S., Singh R.D., Sharma P.C. and Khullar M.,Anaerobiosis induced virulence of Salmonella typhi. Ind. J.Med. Res. 115: 184–188, 2002.
311

34. Baumler A.J., Tsolis R.M., Fitch T.A. and Adams L.D.,Evolution of host adaptation in Salmonella enterica. Infect.Immun. 66: 4579–4587, 1998.
35. Bhriain N.N., Dorman C.J. and Higgins C.F., An overlapbetween osmotic and anaerobic stress responses: a potential
role for DNA supercoiling in the coordinate regulation ofgene expression. Mol. Microbiol. 3: 933–942, 1989.
36. Ramamoorthy R. and Schoel-Meeker D., Borrelia Burgdor-feri proteins whose expression is similarly affected by culturetemperature and pH. Infect. Immun. 69: 2739–2742, 2001.
312




















