
Properties of structure and interaction of the receptor for ω-conotoxin, a polypeptide active on...
12
Vol. 150, No. 2, 1988 January 29, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS Pages 1051-l 062 PROPERTIES OF STRUCTURE AND INTERACTION OF THE RECEPTOR FOR w-CONOTOXIN, A POLYPEPTIDE ACTIVE ON Ca2+ CHANNELS Jacques Barhanin, Annie S&mid and Michel Laahmski Centre de Biochimie Centre National de la Recherche Scientifique Part Valrose, 06034 Nice Cedex, France Received December 22, 1987 SUMMARY. Binding properties of w-conotoxin (GVIA) to avian and mammalian neuronal Ca2+ channels were investigated using a radioiodinated toxin derivati- ve. An exceptionally high affinity of 0.6 to 2 pM was found both from equilibrium and kinetics measurements. Only one class of non-interacting binding sites was detected. In chick brain, dissucinimidyl suberate specifically cross-linked the toxin to 170 kDa component that comprises a 140 kDa peptide disulfide linked to a 30 kDa peptide, very similar to the heavily glycosylated component of the L- type DHP-sensitive Ca2+ channel. A large peptide of 210-220 kDa was labelled using the azidonitrobenzoyloxy derivative of w-conotoxin as cross-linking re- agent but not the 170/140+30 kDa component. The results suggest that the neuronal Ca2+ channel could be composed by the association of two distinct high molecular weight peptides of 220 kDa and 170/140+30kDa. B 1988 Academic Press, Inc. INTRODUCTION. Most excitable cells have different types of voltage-dependent Ca2+ channels which differ by their biophysical properties, their physiological function and their pharmacology. The only one of these channels for which there is a detailed structural information is the L-type Ca2+ channel and more specifi- cally the L-type Ca2+ channel from skeletal muscle. This channel is blocked by a variety af molecules such as 1,4-dihydropyridines (DHP), phenylalkylamines, diltiazem, bepridil or fluspirilene. Receptor sites for these different drugs have been identified. They all reside with the same polypeptide chain of I 165-170 kDa (l-4). This important component of the structure of the Ca2+ channel has now been sequenced (5). Other polypeptide chains are present in the purified protein complex that binds Ca2+ channel blockers (6, 7) and it has Ca2+ channel activity (S-10). The abbrwiitions used are : N-(5-azido-2-nitrobenzoyloxy)succinimide : ANB- NOS), disuccinimidyl suberate : DSS, 1 ,Cdihydropyridines : DHP. ooo6-291X/88 $1.50 1051 Copyright 0 1988 by Academic Press, Inc. All rights of reproduction in any form reserved.
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Properties of structure and interaction of the receptor for ω-conotoxin, a polypeptide active on...

Vol. 150, No. 2, 1988 January 29, 1988
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS Pages 1051-l 062
PROPERTIES OF STRUCTURE AND INTERACTION OF THE RECEPTOR FOR w-CONOTOXIN, A POLYPEPTIDE ACTIVE ON Ca2+ CHANNELS
Jacques Barhanin, Annie S&mid and Michel Laahmski
Centre de Biochimie Centre National de la Recherche Scientifique
Part Valrose, 06034 Nice Cedex, France
Received December 22, 1987
SUMMARY. Binding properties of w-conotoxin (GVIA) to avian and mammalian neuronal Ca2+ channels were investigated using a radioiodinated toxin derivati- ve. An exceptionally high affinity of 0.6 to 2 pM was found both from equilibrium and kinetics measurements. Only one class of non-interacting binding sites was detected. In chick brain, dissucinimidyl suberate specifically cross-linked the toxin to 170 kDa component that comprises a 140 kDa peptide disulfide linked to a 30 kDa peptide, very similar to the heavily glycosylated component of the L- type DHP-sensitive Ca2+ channel. A large peptide of 210-220 kDa was labelled using the azidonitrobenzoyloxy derivative of w-conotoxin as cross-linking re- agent but not the 170/140+30 kDa component. The results suggest that the neuronal Ca2+ channel could be composed by the association of two distinct high molecular weight peptides of 220 kDa and 170/140+30 kDa. B 1988 Academic Press,
Inc.
INTRODUCTION. Most excitable cells have different types of voltage-dependent
Ca2+ channels which differ by their biophysical properties, their physiological
function and their pharmacology. The only one of these channels for which there
is a detailed structural information is the L-type Ca2+ channel and more specifi-
cally the L-type Ca2+ channel from skeletal muscle. This channel is blocked by a
variety af molecules such as 1,4-dihydropyridines (DHP), phenylalkylamines,
diltiazem, bepridil or fluspirilene. Receptor sites for these different drugs have
been identified. They all reside with the same polypeptide chain of I 165-170
kDa (l-4). This important component of the structure of the Ca2+ channel has
now been sequenced (5). Other polypeptide chains are present in the purified
protein complex that binds Ca2+ channel blockers (6, 7) and it has Ca2+ channel
activity (S-10).
The abbrwiitions used are : N-(5-azido-2-nitrobenzoyloxy)succinimide : ANB- NOS), disuccinimidyl suberate : DSS, 1 ,Cdihydropyridines : DHP.
ooo6-291X/88 $1.50
1051 Copyright 0 1988 by Academic Press, Inc.
All rights of reproduction in any form reserved.

Vol. 150, No. 2, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Polypeptide toxins are now available to analyze the properties of CaZi
channels. They have been purified from fish hunting cone snails (11) and called
w-conotoxins and they block presynaptic Ca*+ channels (12) including N-type
Ca*+ channels. Attempts have been made to determine the molecular properties
of the w-conotoxin target using affinity labelling techniques (13, 14). One group
has reported that the MW of the essential peptide in the w-toxin receptor was
135 kDa (131, whereas another one (14) labelled polypeptide of 310 kDa, 240 kDa
and 34 kDa. This paper presents an analysis of the properties of interaction of
w-conotoxin with its receptor and reinvestigates the problem of the subunit
structure of the w -conotoxin receptor.
MATERIALS AND METHODS Materials. Synthetic w-conotoxin GVIA was purchased from Peptide Institute, Inc. (Minoh, Osaka). N-(5-azido-2-nitrobenzoyloxy)succinimide (ANB-NOS) and disuccinimidyl suberate (DSS) were from Sigma. Preparation of lZ51-GVIA. Synthetic GVIA (1 nmol) was iodinated at a high specific activity using 1 nmol (74 MBq) of 1*51-N, (Amersham) in the presence of 15 nmol of chloramine-T in 85 pl of 50 mM sodium phosphate buffer at pH 5.0. The reaction mixture was immediately applied to a 5 ml Sephadex SP-C25 column, equilibrated and eluted with 350 mM NaCl, 20 mM sodium phosphate pH 8.9 and 0.01 mg/ml lysotyme. The number of iodine atoms incorporated per mol of toxin was 0.6. It was measured by applying the iodination and purification procedures described above to a larger amount of GVIA (50 nmol) using 750 nmol of chloramine-T and 49.9 nmol of unlabelled NaI and 0.1 nmol of Na1251. Lysozyme was omitted in the Sephadex SP-C25 eluting buffer and the peptide content of the fractions was determined by a fluorescence method (15). Membrane Preparations. Membrane fractions were prepared as previously descri- bed for chick, rat and rabbit synaptosomes (16), for chick heart microsomes (17), for chick and rabbit skeletal muscle microsomes (6) and for rabbit muscle T- tubules (18). Chicken used were 5 days old, rabbits and rats were about 5 weeks old. Binding Assays. The standard incubation medium for 1251-GVIA binding consisted of 320 mM sucrose, 20 mM Hepes-NaOH pH 7.4, 0.01 mg/ml lysozyme. Bound toxin was separated from free toxin by rapid filtration through GF/C filters (Whatman) kept in 100 mM Tris-HCl pH 7.4, 0.025% polyethyleneimine. Filters were washed twice with 5 ml of 100 mM Tris-HCI pH 7.4, 2 mM CaC12, 0.1% bovine serum albumin. (+)[3H]PN 200-110 (Amersham) binding was performed as described ( 19). Cross-Iinking of 12%CVIA to its receptor. Chick and rat brain synaptosomes were incubated at 20°C with 0.25 nM and 0.5 nM of 1251-GVIA, respectively, in 320 mM sucrose, 20 mM Hepes-NaOH pH 8.5, 0.1 mM phenyl-methylsulfonyl fluoride, without lysozyme. The final concentration of synaptosomes was 0.35 mg of protein per ml, equivalent to a receptor concentration of 0.35 mM for chick membranes and 0.13 nM for rat membranes. Replicate incubations were made with the addition of unlabelled GVIA at a concentration of 10 nM. After a 40 min incubation, DSS in dimethylsulfoxide was added to a final concentration of 30 pM or 300 pM and the mixture was incubated 15 min at 20°C. Tris-HCl pH 8.5 was then added to a final concentration of 100 mM to neutralize excess cross-linking reagent. Synaptosomes were then sedimented by centrifugation and washed twice in the incubation buffer. Photoaffinit mes with the ANB derivative of 115
labelling of chick and rat brain synaptoso- I-GVIA (ANB-125LGVIA) was performed
1052

Vol. 150, No. 2, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
according to Abe and Saisu (14). The toxin used in these experiments was iodinated at high specific activity as described above but Tris was changed for Hepes in the column elution buffer. Finally, labelled synaptosomes were dissol- ved for 3 min at 92°C in a denaturating 75 mM Tris-HCI pH 6.8 buffer, containing 2% SDS, 10% glycerol, 20 mM dithiothreitol (reducing conditions) or 20 mM iodoacetamide (non reducing conditions). 70 pg of protein of each sample were then applied to a 4-14% gradient of polyacrylamide gel and submitted to electrophoresis according to Laemmli (20).
RESULTS. Fig. 1 shows the results of typical binding experiments at low
synaptosome concentrations to ensure that the receptor concentration was in the
range of the Kd value of the toxin-receptor complex. Scatchard plots indicated
that the toxin binds to a single class of receptor sites in both tissues. The Kd
values are 0.82 pM for chick brain synaptosomes and 1.23 pM for rat brain
synaptosomes. The B,,, values for chick and rat membranes are 1.03 pmol and
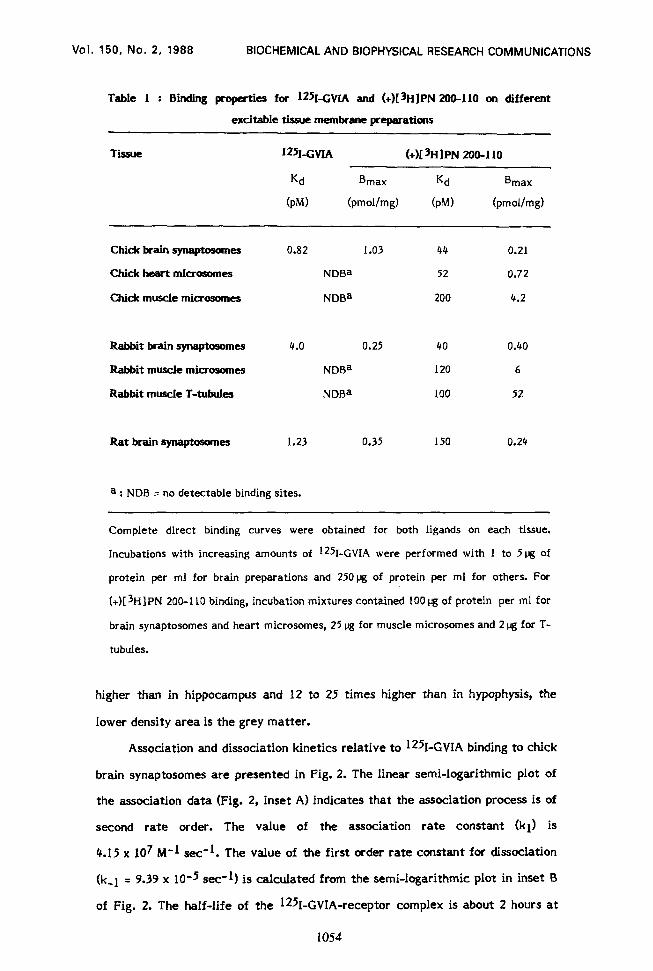
0.35 pmol per mg of protein, respectively. Table 1 indicates B,,, and Kd values
for the association of 1251-GVIA to different tissues and compares them to
corresponding values found for the interaction of the DHP (+)[3H]PN 200-l 10 to
the same membrane preparations.
The distribution of 1251,GVIA binding sites in rat brain is heterogeneous
(Table 2). The cortex is the area with the highest density, approximately twice
I A Chick brain
FREE (PM) - -
u Equilibrium binding of 1251-GVIA to chick and rat brain synaptosomes. Chick (A) or rat (8) brain synaptosomes (I and 3 rrg of respectively) were incubated with various concentrations of PBg&,P”f,,“ij hours at 25°C (final volume 4.5 ml). Each value is the mean of duplicate filtrations of 2 ml. Non specific binding (0 , A ) was measured in the presence of IO-9 M unlabelled toxin. Specific binding (0 , A) is the difference between total and non specific binding. C : Scatchard plot of the data. ( l ) chick; (A ) rat. The specific activity of the 1251~CVIA used was 2000 cpm per fmol.
1053
Vol. 150, No. 2, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Table I : Binding properties for wGVIA and (+)[3H]PN ZOO-110 on different
excitable tissue membrane preparations
TiSUe 125~1~ (+)[3HlPN ZOO-110
Kd
(PM)
Bmax
(pmol/mg)
Kd
(PM)
Bmax
(pmol/mg)
chick brain synaptosome!s 0.82 1.03 44 0.21
C&k% heart microsomes NDBa 52 0.72
Chick muscle microsomes NDBa 200 4.2
Rabbit brain qn+osomes
Rabbit muscle microsomes
Rabbit muscle T-tubules
4.0 0.25 40 0.40
NDBa I20 6
NDBa 100 52
Rat brain synaptosomes 1.23 0.35 150 0.24
a : NDB = no detectable binding sites.
Complete direct binding curves were obtained for both ligands on each tissue.
Incubations with increasing amounts of ‘2kGVIA were performed with 1 to 5pg of
protein per ml for brain preparations and 25Opg of protein per ml for others. For
(+)[sH]PN 200-l 10 binding, incubation mixtures contained 100 pg of protein per ml for
brain synaptosomes and heart microsomes, 25 pg for muscle microsomes and 2 pg for T-
tubules.
higher than in hippocampus and 12 to 25 times higher than in hypophysis, the
lower density area is the grey matter.
Association and dissociation kinetics relative to 125I-GVIA binding to chick
brain synaptosomes are presented in Fig. 2. The linear semi-logarithmic plot of
the association data (Fig. 2, inset A) indicates that the association process is of
second rate order. The value of the association rate constant (kl) is
4.15 x 107 M-1 see-1. The value of the first order rate constant for dissociation
(k-1 = 9.39 x 10-5 see-1) is calculated from the semi-logarithmic plot in inset B
of Fig. 2. The half-life of the I251-GVIA-receptor complex is about 2 hours at
1054

Vol. 150, No. 2, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Table 2 : Distribution of 12%-GVIA binding sites ln rat brain
Brain Area I~~I-GVIA
B max
fpmollmg)
Cortex 0.83
Hippocampus 0.47
Hypothalamus 0.11
Neurohypophysis 0.07
Anterior pituitary 0.03
25°C. The equilibrium dissociation constant (Kd) value calculated from kinetic
data is I&j = (k-l/kl) = 2.26 pM in good agreement with the Kd value deduced
from equilibrium experiments.
Fig. 3 (main panel) shows that the concentration of unlabelied toxin that
50% of 1251-GVIA binding (K0.5) is 0.65 pM. Classical L-type Ca2+
l
k-1=9.39 10-5 set- 1
TIME (min)
Association and dissociation kinetics for the binding of 12%GVIA to rain synaptosomes. Synaptosomes (10 lig of protein per ml) were incubated
at 2Y’C in the presence (non specific binding) or the absence (total bindin 10-8 M unlabelled GVIA. Association was started by addition of 3.7 x IO- f
) of 2 M
125I-GVIA. A plateau value was reached after 100 min of association. At that time, indicated by the arrow, 10-8 M unlabelled GVIA was added to provoke the dissociation of the 12%CVIA-receptor complex (0 ). ( l ), concentration of 1251-GVIA specifically bound during the association process. Inset A : Associa- tion kinetic plotted according to the integrated form of the second rate order equation (28). Inset B : First order representation of the time-course of dissocia- tion. X = percentage of 18%-GVIA-receptor complex remaining at t1me.t.
1055

Vol. 150, No. 2, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
-15 -14 -13 -12 -11 -10 - ‘9
-log [GVIA] (M)
Fig. 3. Protection of 125LGVIA binding by unlabelled GVIA, classic Ca2+ channel antagonists and ions. Chick brain synaptosomes (6% of protein per ml) were incubated at 25°C with the indicated concentration of native toxin (main paneI) or Ca2+ channel antagonist (inset A) or ions (inset B) for 10 min and then I*%- GVIA (6 x lo-12 M) was added for 2 hours. For the effects of ions, the osmolarity of the incubation medium was maintained constant by varying the sucrose concentration and the CaC12 concentration was buffered with 2 mM EDTA which had no effect on the 1251-GVIA binding.
channel inhibitors, including DHP ((+)PN ZOO-1 IO), phenylalkylamines ((*)verapa-
mil), benzothiazepines (d-cis-diltiazem) and neuroleptics of the diphenylbutylpi-
peridine series (fluspirilene) have no effect on 1wGVIA binding to chick brain
synaptosomes, even at high concentrations (Fig. 3, inset A). The same absence of
effect has been observed with rat and rabbit brain synaptosomes.
125I-GVIA binding is very sensitive to the presence of monovalent and
divalent cations. Fig. 3, inset 6, shows 50% inhibition of 1251-GVIA binding in the
presence of 116 mM Na+, 68 mM Tris or 4 mM Ca2+. The inhibition curves are
cooperative with respect to the salt concentration with Hill numbers close to 2.
The same cations are unable to inhibit 125I-GVIA binding when they are added
after the 1251-GVIA-receptor complex has been formed. This observation indica-
tes that the dissociation of the toxin-receptor complex is not accelerated in the
presence of 140 mM NaCI, 100 mM Tris or 2 mM CaC12.
Two cross-linking reagents were used to covalently link 1251-GVIA to its
receptor components in brain membranes. The first one was DSS at a final
concentration of 30 or 3OOpM. With chick brain synaptosomes and 3O@i DSS,
1056

Vol. 150, No. 2, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
0 4 r 8 13
N-0-C-CHz-CHz-CH2-CH2-CH2-CH2-C- /
- A 12 34 56 78 812 34 -
0 \\
-0-N
3 4
0
56 78
Fig. @. Covalent cross-linking of 1251-CVIA to chick brain receptor components with DSS. Shown are the autoradiograms depicting the peptides labelled by 12% GVIA using 30 @I DSS (lanes I, 3 and 5, 7) or 300 pM DSS (lanes 2, 4 and 6, 8). Lanes 3, 4 and 7, 8 were obtained for experiments performed in the presence of IO-9 M unlabelled GVIA. Gels were run under reducing conditions (lanes 1-4) and non reducing conditions (lanes 5-8). Part A : autoradiograms exposed for 26 hours at -7O’C on Kodak X-OMAT films with a Cronex Hi-plus intensifying screen (DuPont). Part 6 : autoradiograms from the same gels as in A exposed for 140 hours. Positions of molecular weight markers (BioRad) are indicated by horizon- tal bars : myosin (200 0001, b-galactosidase (116 5001, phosphorylase B (96 500), bovine serum albumin (66 200), ovalbumin (45 OOO), carbonic anhydrase (31 000) and soybean trypsin inhibitor (21 500).
autoradiography of the polyacrylamide gels revealed the presence of only one
specifically labelled band of 140 kDa in reducing conditions and 170 kDa in non-
reducing conditions (Fig. 4A lanes I and 5). Superexposition of the film on the gel
(140 hours instead of 26 hours) resulted in the same specific labelling pattern
indicating that no other bands were labelled, even weakly (Fig. 4B lanes 1 and 5).
When DSS was used at a concentration of 300 pM the specific labelling of the
170 kDa protein was more intense in non reducing conditions, as expected for a
better yield of cross-linking of the toxin to its receptor (Fig. 4A and 8, lanes 2
and 6). However under these conditions, and after reduction of disulfide bonds,
the major labelled component had a MW of 170 kDa, and the 140 kDa band was
still present but in a lower amount (Fig. 4A and B, lane 2). The specificity of the
covalent labelling of the 12%GVIA receptor components is demonstrated by the
1057

Vol. 150, No. 2, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
\\ @Lo-;g
4 ‘43 AN&NOS
1 2 34 - -
Fig. 5. Analysis of the photoaffinity labelled peptides using ANB-l*%GVIA on rat and chick brain membranes. Preparations labelled with ANB-1251-GVIA in the absence (lanes I and 4) or the presence (lanes 2 and 3) of 1O-9 M unlabelled GVIA were analyzed under reducing conditions. Lanes 1 and 2 : rat brain membranes; lanes 3 and 4 : chick brain membranes. Molecular weight markers are the same as in Fig. 4.
absence of incorporation of radioactivity in the previously discussed polypeptides
under protected conditions. The same series of cross-linking experiments carried
out with rat brain membranes were unsuccessful. No specific labeiling could be
detected with this material (not shown).
The second cross-linking technique used to identify the i251-GVIA receptor
consisted in reacting first ANB-NO5 with the toxin to produce ANB-i251-GVIA, a
photoaffinity derivative of the toxin. In our hands, this photoaffinity labelling
resulted in a considerably Iower specific incorporation of the label to the
receptor than in using the DSS technique with chick brain membranes. Moreover,
the 170-140 kDa components were not labelled with ANB-i251-GVIA. The
photoactivated toxin derivative incorporated into a component of about 220 kDa
in rat and chick brain membranes, and also into a peptide of 33 kDa (36 kDa
minus 3 kDa, the MW of the bound toxin) in rat brain membranes (Fig. 5). Under
1058

Vol. 150, No. 2, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
non reducing conditions, these bands were not identified, possibly because they
aggregated and did not enter the separating polyacrylamide gel.
DIXXJSSION. The first important property of w-conotoxin is its exceptionally
high affinity for its target. Equiiibrium binding and kinetic studies have indicated
that Kd values for the w-conotoxin-receptor complex, both in chick and in rat
brain, are in the pM range (0.8 - 2 pM for chick brain and 1.2 pM for rat brain).
These Kd values are even lower than found previously for the Tityus toxin y-Na’
channel interaction (21), for the apamin-Ca2+ -activated K+ channel interaction
(221, and for the dendrotoxin I-K+ channel interaction (23). This very high
affinity of w-toxin for its binding site is accompanied by a slow rate of
dissociation of the complex (t I/2 = 2 hr). Kd values found in this work are
different from those reported by other authors elsewhere. Results by Cruz and
Olivera suggested a Kd value at about 500 pM for chick brain membranes (24).
This high value was probably due to the fact that the concentration of receptors
used in the assay (1.9 nM) was too high as compared to the true dissociation
constant Kd. Under such conditions, one cannot determine a dissociation
constant. In a first paper Abe et al. (25) identified two types of binding sites for --
W -conotoxin in rat brain membranes, one with a Kd value of 10 pM, nearly one
order of magnitude higher than the one found in the present work, and the other
one with a Kd value of 0.5 nM. In a more recent publication (14), the same group
found a half-maximum displacement of 1251-GVIA by the unlabelled toxin at a
concentration near 1 nM. The affinity found in the present work using both
equilibrium and kinetic data is clearly higher than that found previously. It is
nearly the same in avian and mammalian brain membranes and we have found no
evidence for a second type of binding site in either chick or rat brain membranes
even when higher membrane concentrations (100 pg protein/ml) were used in the
binding assays.
Our data are in agreement with others (13) in showing that binding sites for
125LGVIA are absent in other excitable membranes such as muscle or cardiac
membranes which are good sources of material for DHP binding sites. It should
1059

Vol. 150, No. 2, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
not be concluded however from these negative results that there are no channels
in cardiac and skeletal muscles with the biophysical properties of Ca2+ channels
which are the target of w-conotoxin in brain cells. The toxin may be tissue
specific. An example of tissue specificity has previously been observed with
voltage-dependent Na+ channels. Another Conus toxin, p-conotoxin, discrimina-
tes between the different voltage-dependent Na+ channels in different tissues
and only blocks the muscle Na+ channel (26).
At first it is intriguing that the different techniques used to cross-link
w-toxin to its binding site in different brain membranes lead to results
suggesting different polypeptide constitutions for the receptor. The results
obtained here show that previously observed (13, 14) differences in labelling are
probably due to differences in length of the two cross-linking reagents. Photo-
affinity labelling with ANB-12%GVIA identifies two components of 220 kDa and
33 kDa in rat brain membranes and a 210 kDa polypeptide in chick brain
membranes. Cross-linking of 12%GVIA to its receptor in rat brain membranes
with DSS did not lead to a specific incorporation of the toxin to a membrane
polypeptide. However it identified in chick membranes a peptide of 170 kDa
under non reducing conditions that converted to 140 kDa under reducing condi-
tions. This result would suggest that upon reduction the 170 kDa peptide gives
rise to two peptides of about 140 kDa and 30 kDa respectively. The 30 kDa
peptide could be the same as the one that has been found to migrate with a MW
of 33 kDa in affinity labelling experiments using ANB-1251-GVIA and rat
membranes.
Assuming that w -conotoxin binds to the same receptor corresponding to
the same Ca2+ channel in rat and chick brain membranes, the simplest structural
composition of this receptor, taking all results together, would then include one
large polypeptide of 220 kDa and a polypeptide of 170 kDa made by the assembly
of a 140 kDa and of a 30-33 kDa polypeptide.
The DHP-sensitive Ca2+ channel is known to be made of several polypepti-
des. The main one has a MW of 165-170 kDa, its mobility in SDS gel is not
changed upon reduction and it contains receptor sites for all classical Ca2+
1060

Vol. 150, No. 2, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
channel blockers. In addition, the purified channel protein having the binding
properties for Ca*+ channel blockers is known to contain a heavily glycosylated
polypeptide of 170 kDa (3, 4, 6, 7) which is constituted by the covalent
association through disulfide bridges of a 140 kDa and a -30 kDa polypeptide
(27). Cloning of the cDNA corresponding to the 165-170 kDa subunit has been
recently achieved (5) and surprisingly enough, the sequence deduced has a MW of
210 kDa (without glycosylation) suggesting that a processing occurs during
integration into the membrane or in purification studies. There is obviously a
close analogy between the MW of 210 kDa obtained from cDNA sequencing of
the DHP receptor and the MW of 220 kDa obtained from photoaffinity labelling
of the w -conotoxin receptor. There is also a close analogy between the MW of
the 170/140 kDa+32 kDa protein identified in the purification of the DHP
receptor and the 170/140 kDa+30 kDa protein identified by cross-linking experi-
ments with DSS. It should be kept in mind that w-conotoxin blocks both L-type
and N-type Ca*+ channels (12). Therefore one cannot completely eliminate an
interpretation of the results which would imply that the 170/140 kDa+30 kDa
protein belongs to the L-type channel (since it has previously been identified as
part of the DHP receptor) while the 220 kDa polypeptide labelled with ANB-
1*51-GVIA belongs to the N-type Ca *+ channel (since it has not been yet
biochemically identified as part of the DHP receptor). A final answer to this
problem will have to come from the sequence determination through cDNA
cloning of the different components of the w-conotoxin receptor.
ACKNOWLEDGEMENTS. We wish to thank Dr. Hof (Sandot, Basel, Switzerland) for the gift of unlabelled (+)PN 200-100, Dr. Traut, Dr. Hollman, and Dr. Lietz (Knoll AC, Ludwigshafen, FRG) for the gift of (*)verapamil, Dr. Laduron (Janssen Pharmaceutics, Beerse, Belgium) for the gift of fluspirilene and Synthelabo (Paris, France) for the gift of d-cis-diltiazem. We also thank Dr. A. Drouva (INSERM U.159, Paris, France) for the preparation of membranes from different brain areas. The excellent technical assistance of Marie-Madeleine Larroque and expert secretarial assistance of C. Roulinat-Bettelheim are greatly appreciated and acknowledged. This work was supported by the ‘Association des Myopathes’, the ‘Centre National de la Recherche Scientifique’, and the ‘Minis&e de la Defense Nationale’ (grant DRET No 85/138).
REFERENCES
1. Galizzi, J.-P., Borsotto, M., Barhanin, J., Fosset, M., and Lazdunski, (1986) J. Biol. Chem. 261, 1393-1397.
M.
1061

Vol. 150, No. 2, 1988 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
2.
3.
4.
5.
6.
7.
8. 9.
10.
11.
12.
13.
14. 15.
16.
17.
18.
19.
20. 21.
22.
23.
24. 25.
26.
27.
28.
Sharp, A.H., Imagawa, T., Leung, A.T., and Campbell, K.P. (1987) 3. Biol. Chem. 262, 12309-12315. Vaghy, P.L., Striessnig, J., Miwa, K., Knaus, H.G., Itagaki, K., McKenna, E., Glossmann, H., and Schwartz, A. (1987) J. Biol. Chem. 262, 14337- 14342. Hosey, M.M., Barhanin, J., Schmid, A., Vandaele, S., Ptasienski, J., O’CalIahan, C., Cooper, C., and Lazdunski, M. (1987) Biochem. Biophys. Res. Commun. 147, 1137-1145. Tanabe, T., Takeshima, H., Mikami, A., Flockerzi, V., Takahashi, H., Kangawa, K., Kojima, M., Matsuo, H., Hirose, T., and Numa, S. (1987) Nature 328, 313-318. Borsotto, M., Barhanin, J., Fosset, M., and Lazdunski, M. (1985) 3. Biol. Chem. 260, 14255-14263. Takahashi, M., Seagar, M.J., Jones, J.F., Reber, B.F.X., and Catterall, W.A. (1987) Proc. Natl. Acad. Sci. USA 84, 5478-5482. Curtis, B.M., and Catterall, W.A. (1986) Biochemistry 25, 3077-3083. Flockerzi, V., Oeken, H.J., Hofmann, F., Pelzer, D., Cavalii, A., Trautwein, W. (1986) Nature 323, 66-68. Smith, J.S., McKenna, E.J., Ma, J., Vilven, J., Vaghy, P.L., Schwartz, A., and Coronado, R. (1987) Biochemistry 26, 7182-7188. Olivera, B.M., McIntosh, M., Cruz, L.J., Luque, F.A., and Gray, W.R. (1984) Biochemistry 23, 5087-5090. McCleskey, E.W. Fox, A.P., Feldman, D.H., Cruz, L.J., Olivera, B.M., Tsien, R.W., and Yoshikami, D. (1987) Proc. Natl. Acad. Sci, USA 84, 4327- 4331. Cruz, L.J., Johnson, D.S., and Olivera, B.M. (1987) Biochemistry 26, 820- 824. Abe, T., and Saisu, H. (1987) 3. Biol. Chem. 262, 9877-9882. Bohlen, P., Stein, S., Dairman, W., and Udenfriend, S. (1987) Arch. Biochem. Biophys. 155, 213-219. Abita, J.-P., Chicheportiche, R., Schweitz, H., and Lazdunski, M. (1977) Biochemistry 16, 1838-1844. Schmid, A., Barhanin, J., Coppola, T., Borsotto, M., and Lazdunski, M. (1986) Biochemistry 25, 3492-3495. Fosset, M., Jaimovich, E., Delpont, E., and Lazdunski, M. (1983) J. Biol. Chem. 258, 6086-6092. Cognard, C., Romey, G., Galizzi, J.P., Fosset, M., and Lazdunski, M. (1986) Proc. Natl. Acad. Sci. USA 83, 1518-1522. Laemmli, U.K. (1970) Nature (London) 227, 680-685. Barhanin, J., Pauron, D., Lombet, A., Norman, R.I., Vijverberg, H.P.M., Giglio, J.R., and Lazdunski, M. (1983) EMBO 3. 2, 915-920. Hugues, M., Romey, G., Duval, D., Vincent, J.-P., and Lazdunski, M. (1982) Prof. Natl. Acad. Sci. USA 79, 1308-1312. Bidard, J.-N., Mourre, C., and Lazdunski, M. (1987) Biochem. Biophys. Res. Commun. 143, 383-389. Cruz, L.J., and Olivera, B. (1986) J. Biol. Chem. 261, 6230-6233. Abe, T., Koyano, K., Salsu, H., Nishiuchi, Y., and Sakakibara, S. (1986) Neurosci. Lett. 71, 203-208. Cruz, L.J., Gray, W.R., Olivera, B.M., Zeikus, R.D., Kerr, L., Yoshikami, D., and Moczydlowski, E. (1985) J. Biol. Chem. 260, 9280-9288. Barhanin, J., Coppola, T., Schmid, A., Borsotto, M., and Lazdunski, M. (1987) Eur. J. Biochem. 164, 525-531. Weiland, G.A., and Molinoff, P.B. (1981) Life Sciences 29, 313-330.
1062








![Near-Membrane [Ca2+] Transients Resolved Using the Ca2+ Indicator FFP18](https://static.fdokumen.com/doc/165x107/631286873ed465f0570a4533/near-membrane-ca2-transients-resolved-using-the-ca2-indicator-ffp18.jpg)











