![Survey of renewable chemicals produced from lignocellulosic biomass during ionic liquid pretreatment [2013]](https://static.fdokumen.com/doc/165x107/6333905f9d8fc1106803de76/survey-of-renewable-chemicals-produced-from-lignocellulosic-biomass-during-ionic.jpg)
Production of polyhydroxybutyrate in switchgrass, a value-added co-product in an important...
16
Plant Biotechnology Journal (2008) 6, pp. 663–678 doi: 10.1111/j.1467-7652.2008.00350.x © 2008 Metabolix, Inc. Journal compilation © 2008 Blackwell Publishing Ltd 663 Blackwell Publishing Ltd Oxford, UK PBI Plant Biotechnology Journal 1467-7644 1467-7652 © 2008 METABOLIX, INC XXX Original Articles Polyhydroxybutyrate production in switchgrass Maria N. Somleva et al. Production of polyhydroxybutyrate in switchgrass, a value-added co-product in an important lignocellulosic biomass crop Maria N. Somleva, Kristi D. Snell*, Julie J. Beaulieu, Oliver P. Peoples, Bradley R. Garrison† and Nii A. Patterson Metabolix, Inc., 21 Erie St., Cambridge, MA 02139, USA Summary Polyhydroxyalkanoate bio-based plastics made from renewable resources can reduce petroleum consumption and decrease plastic waste disposal issues as they are inherently biodegradable in soil, compost and marine environments. In this paper, the successful engineering of the biomass crop switchgrass (Panicum virgatum L.) for the synthesis of polyhydroxybutyrate (PHB) is reported. Polymer production was monitored in more than 400 primary transformants grown under in vitro and glasshouse conditions. Plants containing up to 3.72% dry weight of PHB in leaf tissues and 1.23% dry weight of PHB in whole tillers were obtained. Results from the analysis of the polymer distribution at the cellular and whole plant levels are presented, and target areas for the improvement of PHB production are highlighted. Polymer accumulation was also analysed in the T 1 generation obtained from controlled crosses of transgenic plants. This study presents the first successful expression of a functional multigene pathway in switchgrass, and demonstrates that this high-yielding biomass crop is amenable to the complex metabolic engineering strategies necessary to produce high-value biomaterials with lignocellulose-derived biofuels. Received 28 July 2007; accepted 1 April 2008. *Correspondence (fax 617-583-1767; e-mail [email protected]) †Present address: John Hopkins University, 1830 E. Monument St., Baltimore, MD 21287, USA Keywords: metabolic engineering, multigene construct, Panicum virgatum, polyhydroxybutyrate, switchgrass, T 1 progeny. Introduction Fuels, plastics and chemicals derived from agricultural feedstocks are receiving considerable attention as the world looks for solu- tions to dwindling non-renewable petroleum resources (Herrera, 2006; Ragauskas et al., 2006; Kamm and Kamm, 2007). In the USA, efforts have primarily focused on biofuels, such as ethanol, produced from the starch in maize kernels. This feedstock will probably be replaced by lignocellulosic biomass as US maize production capacity can only supply a proportion of the feed- stock required for the widespread production of ethanol [Department of Energy (DOE), 2006; Service, 2007]. Current technologies for the conversion of lignocellulosic biomass to biofuels are hindered by high costs, and a significant amount of research effort is underway to create more efficient, less expensive processes (Service, 2007). Engineering bioenergy crops to synthesize industrial materials would provide better economics for both the fuel and co-product components. Polyhydroxyalkanoates (PHAs), a family of naturally renew- able and biodegradable plastics, fit nicely into a biorefinery concept (Ragauskas et al., 2006; Kamm and Kamm, 2007) as a value-added co-product to lignocellulose-derived biofuels. These polymers occur in nature as a storage reserve in some microbes faced with nutrient limitation (Madison and Huisman, 1999), and possess properties enabling their use in a variety of applications currently served by petroleum-based plastics. PHA-biobased plastics can be produced via commercial large-scale fermentations of microbial strains, and the marketing of these plastics in a variety of applications is well underway (Bieles, 2006; Deligio, 2007). Pathways for the production of PHAs have been introduced into a number of crops (for a review, see Suriyamongkol et al., 2007; and references cited therein), including maize (Poirier and Gruys, 2002), sugarcane (Petrasovits et al., 2007; Purnell et al., 2007), flax (Wrobel et al., 2004; Wrobel- Kwiatkowska et al., 2007), cotton (John and Keller, 1996), alfalfa (Saruul et al., 2002), tobacco (Arai et al., 2001; Bohmert et al., 2002; Lössl et al., 2003, 2005), potato (Bohmert et al., 2002) and oilseed rape (Houmiel et al., 1999; Slater et al., 1999; Valentin et al., 1999), resulting in the
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Production of polyhydroxybutyrate in switchgrass, a value-added co-product in an important...

Plant Biotechnology Journal
(2008)
6
, pp. 663–678 doi: 10.1111/j.1467-7652.2008.00350.x
© 2008 Metabolix, Inc.Journal compilation © 2008 Blackwell Publishing Ltd
663
Blackwell Publishing LtdOxford, UKPBIPlant Biotechnology Journal1467-76441467-7652© 2008 METABOLIX, INCXXXOriginal Articles
Polyhydroxybutyrate production in switchgrass
Maria N. Somleva
et al.
Production of polyhydroxybutyrate in switchgrass, a value-added co-product in an important lignocellulosic biomass crop
Maria N. Somleva, Kristi D. Snell*, Julie J. Beaulieu, Oliver P. Peoples, Bradley R. Garrison† and Nii A. Patterson
Metabolix, Inc., 21 Erie St., Cambridge, MA 02139, USA
Summary
Polyhydroxyalkanoate bio-based plastics made from renewable resources can reduce
petroleum consumption and decrease plastic waste disposal issues as they are inherently
biodegradable in soil, compost and marine environments. In this paper, the successful
engineering of the biomass crop switchgrass (
Panicum virgatum
L.) for the synthesis of
polyhydroxybutyrate (PHB) is reported. Polymer production was monitored in more than
400 primary transformants grown under
in vitro
and glasshouse conditions. Plants
containing up to 3.72% dry weight of PHB in leaf tissues and 1.23% dry weight of PHB in
whole tillers were obtained. Results from the analysis of the polymer distribution at the
cellular and whole plant levels are presented, and target areas for the improvement of PHB
production are highlighted. Polymer accumulation was also analysed in the T
1
generation
obtained from controlled crosses of transgenic plants. This study presents the first successful
expression of a functional multigene pathway in switchgrass, and demonstrates that this
high-yielding biomass crop is amenable to the complex metabolic engineering strategies
necessary to produce high-value biomaterials with lignocellulose-derived biofuels.
Received 28 July 2007;
accepted 1 April 2008.
*
Correspondence
(fax 617-583-1767;
e-mail [email protected])
†
Present address
: John Hopkins University,
1830 E. Monument St., Baltimore,
MD 21287, USA
Keywords:
metabolic engineering,
multigene construct,
Panicum
virgatum
, polyhydroxybutyrate,
switchgrass, T
1
progeny.
Introduction
Fuels, plastics and chemicals derived from agricultural feedstocks
are receiving considerable attention as the world looks for solu-
tions to dwindling non-renewable petroleum resources (Herrera,
2006; Ragauskas
et al
., 2006; Kamm and Kamm, 2007). In the
USA, efforts have primarily focused on biofuels, such as ethanol,
produced from the starch in maize kernels. This feedstock will
probably be replaced by lignocellulosic biomass as US maize
production capacity can only supply a proportion of the feed-
stock required for the widespread production of ethanol
[Department of Energy (DOE), 2006; Service, 2007]. Current
technologies for the conversion of lignocellulosic biomass to
biofuels are hindered by high costs, and a significant amount
of research effort is underway to create more efficient, less
expensive processes (Service, 2007). Engineering bioenergy
crops to synthesize industrial materials would provide better
economics for both the fuel and co-product components.
Polyhydroxyalkanoates (PHAs), a family of naturally renew-
able and biodegradable plastics, fit nicely into a biorefinery
concept (Ragauskas
et al
., 2006; Kamm and Kamm, 2007) as
a value-added co-product to lignocellulose-derived biofuels.
These polymers occur in nature as a storage reserve in some
microbes faced with nutrient limitation (Madison and Huisman,
1999), and possess properties enabling their use in a variety
of applications currently served by petroleum-based plastics.
PHA-biobased plastics can be produced via commercial
large-scale fermentations of microbial strains, and the
marketing of these plastics in a variety of applications is well
underway (Bieles, 2006; Deligio, 2007).
Pathways for the production of PHAs have been introduced
into a number of crops (for a review, see Suriyamongkol
et al
., 2007; and references cited therein), including maize
(Poirier and Gruys, 2002), sugarcane (Petrasovits
et al
., 2007;
Purnell
et al
., 2007), flax (Wrobel
et al
., 2004; Wrobel-
Kwiatkowska
et al
., 2007), cotton (John and Keller, 1996),
alfalfa (Saruul
et al
., 2002), tobacco (Arai
et al
., 2001;
Bohmert
et al
., 2002; Lössl
et al
., 2003, 2005), potato
(Bohmert
et al
., 2002) and oilseed rape (Houmiel
et al
., 1999;
Slater
et al
., 1999; Valentin
et al
., 1999), resulting in the

664
Maria N. Somleva
et al.
© 2008 Metabolix, Inc.Journal compilation © 2008 Blackwell Publishing Ltd,
Plant Biotechnology Journal
,
6
, 663–678
production of a range of polymer levels depending on the
crop and mode of transformation. In this study, switchgrass
(
Panicum virgatum
L.), a C4 perennial grass with high biomass
yields, was targeted as the vehicle for PHA production.
Switchgrass is one of the bioenergy crops targeted by the
US DOE (DOE, 2006; Sanderson
et al
., 2006), and recent
studies have suggested that the production of cellulosic
ethanol from this crop nets 540% more renewable energy
than the required non-renewable energy inputs (Schmer
et al
., 2008). Despite the considerable interest in the application
of genomics and transgenic approaches for the improvement
of switchgrass for biofuel production (Bouton, 2007), only
the expression of transgenes encoding visual and selectable
marker genes has been described (Richards
et al
., 2001;
Somleva
et al
., 2002). In this study, the successful engineering
of switchgrass using a multigene pathway for the synthesis
of polyhydroxybutyrate (PHB), the most extensively studied
member of the PHA family (Madison and Huisman, 1999), is
reported. This research is a first step towards the exploration
of the potential of switchgrass as a platform for polymer
production to complement biomass use for bioenergy.
Results
Transformation of switchgrass with multigene
constructs for PHB synthesis
In this study, two different promoters for the expression of
the PHB biosynthetic pathway in switchgrass were compared:
the strong constitutive promoter of the rice ubiquitin 2 gene
(Wang
et al
., 2000) and the
cab-m5
light-inducible promoter
of the chlorophyll
a
/
b
-binding protein in maize (Sullivan
et al
.,
1989; Becker
et al
., 1992), fused to the heat shock protein 70
(
hsp70
) intron (Brown and Santino, 1997). These promoters
were assembled into expression cassettes with PHB pathway
genes that had been previously appended at the 5
′
end with
a DNA sequence encoding a plastid targeting signal (Kourtz
et al
., 2005) to form the
cab-m5
promoter construct
pMBXS155 and the
rubi2
promoter construct pMBXS159.
Highly embryogenic callus cultures were inoculated with
Agrobacterium tumefaciens
strain AGL1 (Lazo
et al
., 1991)
harbouring pMBXS159 or pMBXS155, and transformed
cultures were selected for functional expression of the
bar
gene imparting resistance to bialaphos. Regenerated plantlets
were further screened by treatment with the herbicide Basta,
and the presence of the transgenes was confirmed by
polymerase chain reaction (PCR). Approximately 9% of the
regenerated plants were found to be non-transformed
escapes. Transformation efficiencies, defined as the number
of PCR-positive plants recovered per inoculated explant,
varied from 0% to 93%, depending on the genotype and the
gene construct utilized in the transformation (Table 1).
Southern hybridizations were performed on 48 PCR-
positive plants regenerated from 17 bialaphos-resistant callus
lines transformed with pMBXS155 (Figure 1a). Twenty-seven
independent transformation events were identified by their
unique integration patterns. The T-DNA copy number detected
in these lines ranged from one to nine. About 50% of the
independent transformants analysed had one (six events) or
two (eight events) gene copies. Thirteen transformation
Table 1 Transformation response of mature caryopsis-derived embryogenic callus cultures from various switchgrass genotypes (our designation), cv. Alamo
Alamo
genotype
cab-m5 promoter construct rubi2 promoter construct
Number of calluses
Number of
plantlets‡
Number of calluses
Number of
plantlets‡Inoculated* Transformed† Inoculated§ Transformed†
56 258 83 240 113 28 104
92 128 2 10 49 0 0
119 43 1 3 37 1 3
136 201 4 14 71 12 22
215 113 13 25 124 6 5
247 51 5 13 64 7 21
Total 794 108 305 458 54 155
*Callus cultures inoculated with Agrobacterium tumefaciens AGL1/pMBXS155.
†Bialaphos-resistant calluses producing transgenic plantlets.
‡Transgenic plantlets carrying all four transgenes as confirmed by polymerase chain reaction (PCR).
§Callus cultures inoculated with AGL1/pMBXS159.
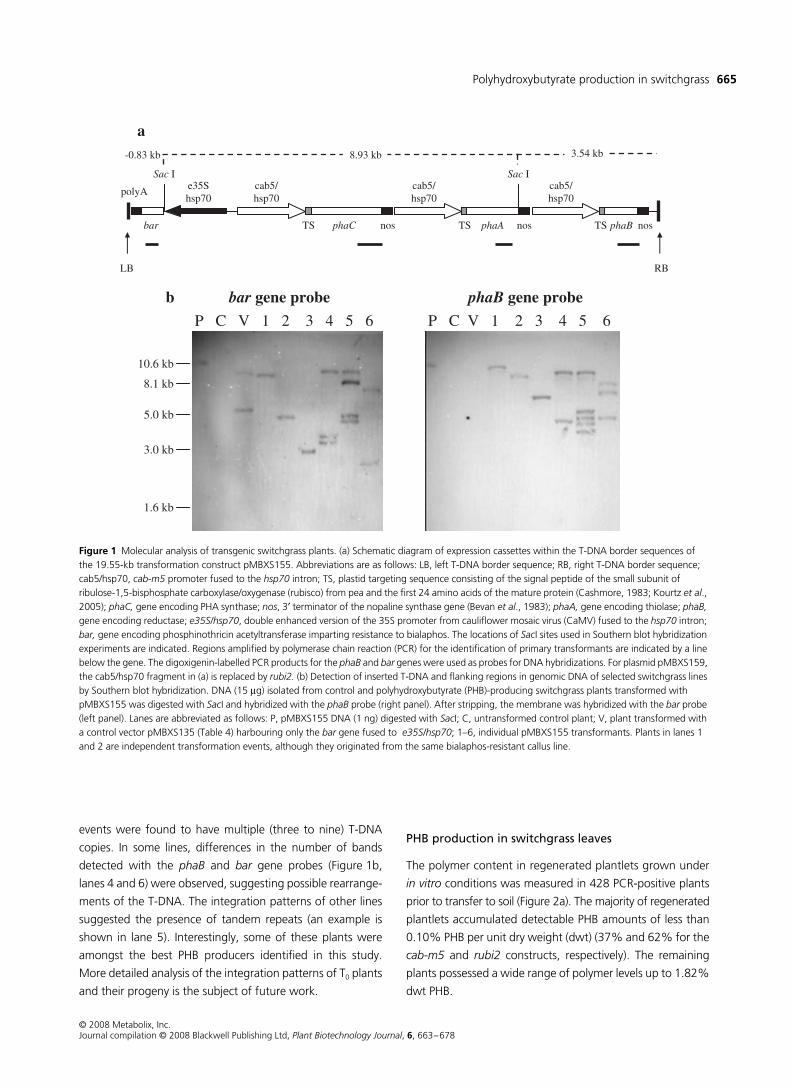
Polyhydroxybutyrate production in switchgrass
665
© 2008 Metabolix, Inc.Journal compilation © 2008 Blackwell Publishing Ltd,
Plant Biotechnology Journal
,
6
, 663–678
events were found to have multiple (three to nine) T-DNA
copies. In some lines, differences in the number of bands
detected with the
phaB
and
bar
gene probes (Figure 1b,
lanes 4 and 6) were observed, suggesting possible rearrange-
ments of the T-DNA. The integration patterns of other lines
suggested the presence of tandem repeats (an example is
shown in lane 5). Interestingly, some of these plants were
amongst the best PHB producers identified in this study.
More detailed analysis of the integration patterns of T
0
plants
and their progeny is the subject of future work.
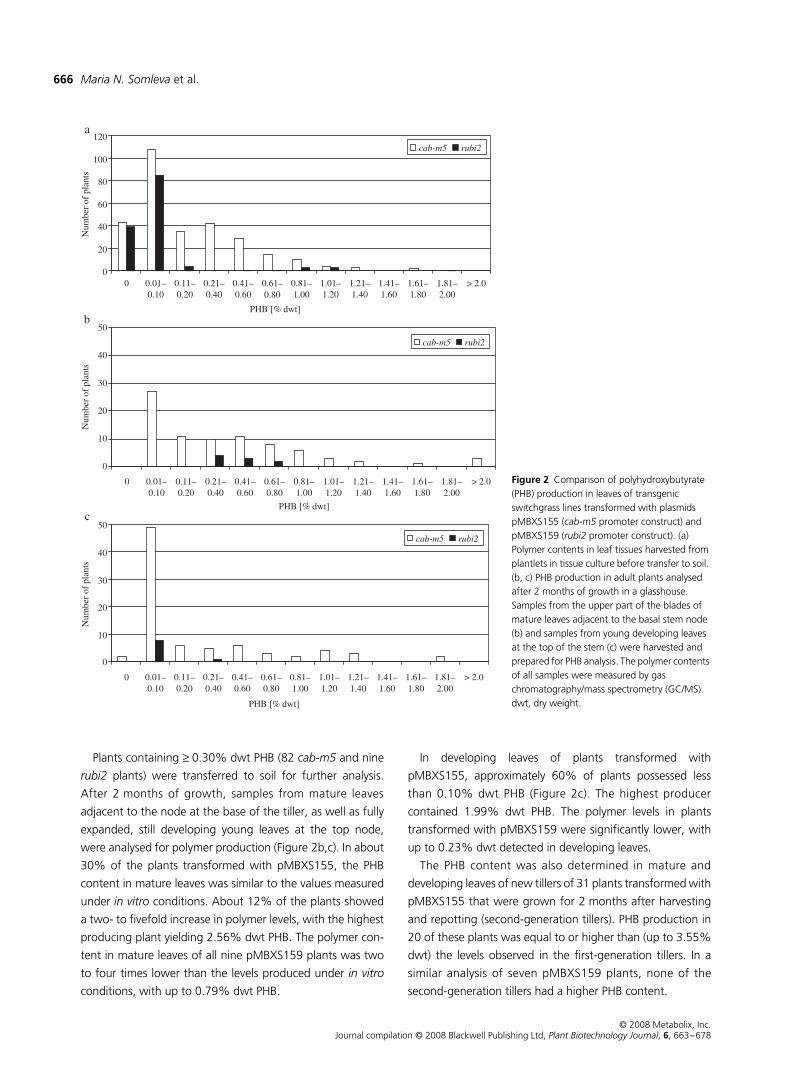
PHB production in switchgrass leaves
The polymer content in regenerated plantlets grown under
in vitro
conditions was measured in 428 PCR-positive plants
prior to transfer to soil (Figure 2a). The majority of regenerated
plantlets accumulated detectable PHB amounts of less than
0.10% PHB per unit dry weight (dwt) (37% and 62% for the
cab-m5
and
rubi2
constructs, respectively). The remaining
plants possessed a wide range of polymer levels up to 1.82%
dwt PHB.
Figure 1 Molecular analysis of transgenic switchgrass plants. (a) Schematic diagram of expression cassettes within the T-DNA border sequences of the 19.55-kb transformation construct pMBXS155. Abbreviations are as follows: LB, left T-DNA border sequence; RB, right T-DNA border sequence; cab5/hsp70, cab-m5 promoter fused to the hsp70 intron; TS, plastid targeting sequence consisting of the signal peptide of the small subunit of ribulose-1,5-bisphosphate carboxylase/oxygenase (rubisco) from pea and the first 24 amino acids of the mature protein (Cashmore, 1983; Kourtz et al., 2005); phaC, gene encoding PHA synthase; nos, 3′ terminator of the nopaline synthase gene (Bevan et al., 1983); phaA, gene encoding thiolase; phaB, gene encoding reductase; e35S/hsp70, double enhanced version of the 35S promoter from cauliflower mosaic virus (CaMV) fused to the hsp70 intron; bar, gene encoding phosphinothricin acetyltransferase imparting resistance to bialaphos. The locations of SacI sites used in Southern blot hybridization experiments are indicated. Regions amplified by polymerase chain reaction (PCR) for the identification of primary transformants are indicated by a line below the gene. The digoxigenin-labelled PCR products for the phaB and bar genes were used as probes for DNA hybridizations. For plasmid pMBXS159, the cab5/hsp70 fragment in (a) is replaced by rubi2. (b) Detection of inserted T-DNA and flanking regions in genomic DNA of selected switchgrass lines by Southern blot hybridization. DNA (15 μg) isolated from control and polyhydroxybutyrate (PHB)-producing switchgrass plants transformed with pMBXS155 was digested with SacI and hybridized with the phaB probe (right panel). After stripping, the membrane was hybridized with the bar probe (left panel). Lanes are abbreviated as follows: P, pMBXS155 DNA (1 ng) digested with SacI; C, untransformed control plant; V, plant transformed with a control vector pMBXS135 (Table 4) harbouring only the bar gene fused to e35S/hsp70; 1–6, individual pMBXS155 transformants. Plants in lanes 1 and 2 are independent transformation events, although they originated from the same bialaphos-resistant callus line.
666
Maria N. Somleva
et al.
© 2008 Metabolix, Inc.Journal compilation © 2008 Blackwell Publishing Ltd,
Plant Biotechnology Journal
,
6
, 663–678
Plants containing
≥
0.30% dwt PHB (82
cab-m5
and nine
rubi2
plants) were transferred to soil for further analysis.
After 2 months of growth, samples from mature leaves
adjacent to the node at the base of the tiller, as well as fully
expanded, still developing young leaves at the top node,
were analysed for polymer production (Figure 2b,c). In about
30% of the plants transformed with pMBXS155, the PHB
content in mature leaves was similar to the values measured
under
in vitro
conditions. About 12% of the plants showed
a two- to fivefold increase in polymer levels, with the highest
producing plant yielding 2.56% dwt PHB. The polymer con-
tent in mature leaves of all nine pMBXS159 plants was two
to four times lower than the levels produced under
in vitro
conditions, with up to 0.79% dwt PHB.
In developing leaves of plants transformed with
pMBXS155, approximately 60% of plants possessed less
than 0.10% dwt PHB (Figure 2c). The highest producer
contained 1.99% dwt PHB. The polymer levels in plants
transformed with pMBXS159 were significantly lower, with
up to 0.23% dwt detected in developing leaves.
The PHB content was also determined in mature and
developing leaves of new tillers of 31 plants transformed with
pMBXS155 that were grown for 2 months after harvesting
and repotting (second-generation tillers). PHB production in
20 of these plants was equal to or higher than (up to 3.55%
dwt) the levels observed in the first-generation tillers. In a
similar analysis of seven pMBXS159 plants, none of the
second-generation tillers had a higher PHB content.
Figure 2 Comparison of polyhydroxybutyrate (PHB) production in leaves of transgenic switchgrass lines transformed with plasmids pMBXS155 (cab-m5 promoter construct) and pMBXS159 (rubi2 promoter construct). (a) Polymer contents in leaf tissues harvested from plantlets in tissue culture before transfer to soil. (b, c) PHB production in adult plants analysed after 2 months of growth in a glasshouse. Samples from the upper part of the blades of mature leaves adjacent to the basal stem node (b) and samples from young developing leaves at the top of the stem (c) were harvested and prepared for PHB analysis. The polymer contents of all samples were measured by gas chromatography/mass spectrometry (GC/MS). dwt, dry weight.

Polyhydroxybutyrate production in switchgrass
667
© 2008 Metabolix, Inc.Journal compilation © 2008 Blackwell Publishing Ltd,
Plant Biotechnology Journal
,
6
, 663–678
In total, 19 plants, representing 6.5% of the population of
cab-m5
plants analysed in this study (291 plants), accumulated
1.00%–3.55% dwt PHB in fully developed mature leaves.
Transgene expression and PHB production
Reverse transcription-polymerase chain reaction (RT-PCR)
and Western blot analysis were conducted with randomly
selected pMBXS155 plants with different PHB contents to
determine whether there was a correlation between
transcript and/or protein levels and product accumulation.
An example is shown in Figure 3. RT-PCR results revealed the
presence of transcripts of the expected size of all the trans-
genes, indicating transcriptional activity of the genes of the
PHB biosynthetic pathway in both mature and developing
leaves (Figure 3a). Similar amounts of transgene mRNAs
were detected in both types of sample in the low and high
producers (Figure 3a, LP and HP), but the transcript levels for
the
phaB
,
phaC
and
bar
genes in the developing leaf of the
medium producer (Figure 3a, MP) were lower than those in
the mature leaf. The highest mRNA levels for all transgenes
were detected in the high producer (Figure 3a, HP).
Western blot analysis of the total soluble proteins isolated
from the same leaves as used for RNA extraction was
performed with anti-thiolase and anti-reductase antibodies
(Figure 3b). The relative amounts of thiolase protein were
found to correlate well with PHB concentration in the corre-
sponding samples, despite the lack of positive correlation
between the
phaA
mRNA levels and thiolase content. In
contrast, the abundance of the reductase protein reflected
the transcript levels of the
phaB
gene, resulting in higher
protein accumulation in both mature and developing leaves
of the low producer than in the medium producer (Figure 3b,
LP and MP). A similar discrepancy in reductase expression and
PHB accumulation has been observed previously in
Arabidopsis
(Kourtz
et al
., 2005). In studies with PHB-producing
sugarcane lines, significant correlations were established
between the relative amounts of transgene-encoded
enzymes and polymer concentration (Purnell
et al
., 2007).
However, the authors reported relatively poor correlations
between each protein and its respective transcript.
PHB formation in switchgrass leaf cells
In Nile blue-stained sections from fully developed mature
leaves of transgenic plants obtained from both pMBXS155
(Figure 4a,b) and pMBXS159 (data not shown) transforma-
tions, fluorescent granular inclusions were located primarily
in the bundle sheath cells surrounding the vascular tissues.
These granules were not present in sections from wild-type
plants (Figure 4c,d). Interestingly, vascular bundles of the
mid-vein of wild-type leaves yielded some staining with Nile
blue that was not present in the transgenic samples. As Nile
blue also stains lipids (Greenspan and Fowler, 1985), this may
indicate a difference in lipid composition of the vascular
bundles of transgenic and non-transgenic plants.
Further analysis by transmission electron microscopy
demonstrated that granules were located in the chloroplasts
of leaves from plants transformed with the pMBXS155 gene
construct. PHB granules were observed primarily in the
chloroplasts of the bundle sheath cells (Figure 4e) and occa-
sionally in the mesophyll cells (Figure 4f). In both cell types,
granule formation was often co-localized with plastoglobules.
In samples from wild-type leaves, no granules of PHB were
observed (Figure 4g,h). Despite incubation of plants in the
dark for 22 h prior to sampling, starch granules were still
Figure 3 Transgene expression in T0 plants producing different levels of polyhydroxybutyrate (PHB). Mature and developing leaves from glasshouse-grown plants were split longitudinally and used for PHB content measurement, RNA extraction and soluble protein isolation. (a) Transcript levels determined by reverse transcription-polymerase chain reaction (RT-PCR) in 0.5 ng of total RNA isolated from mature (left lanes) and developing (right lanes) leaves from a low producer (LP; 0.02% and < 0.01% dry weight (dwt) PHB in mature and developing leaves, respectively), medium producer (MP; 1.11% and 0.42% dwt PHB), high producer (HP; 1.84% and 0.87% dwt PHB) and a control non-transformed plant (WT). Actin expression (bottom panel) was used as a control for uniform loading. (b) Western blots of total soluble proteins (12 μg per lane) incubated with antibodies against thiolase (top panel) and reductase (bottom panel). Lane C contains 150 ng of purified Ralstonia eutropha enzymes.

668
Maria N. Somleva et al.
© 2008 Metabolix, Inc.Journal compilation © 2008 Blackwell Publishing Ltd, Plant Biotechnology Journal, 6, 663–678
visible in the chloroplasts of the bundle sheath cells of wild-type
plants, but were absent in the chloroplasts of PHB producers.
Effect of PHB production on plant growth and
development
All of the plantlets obtained from callus transformed with
pMBXS155 grew normally in tissue culture. A few plants
regenerated from cultures constitutively expressing pha
genes showed some phenotypic changes under in vitro
conditions (data not shown). Some plants accumulating
more than 2% dwt PHB in senescing leaves were observed to
have slight chlorosis and growth reduction (Figure 5a) without
drastic changes in plant development. However, normally
growing plants containing similar amounts of PHB were also
obtained (Figure 5b).
Polymer accumulation in whole plants
The spatial distribution of PHB was examined in leaf blades
and stem tissues (nodes, internodes, leaf sheaths, panicles) of
transgenic plants grown under glasshouse conditions. Tillers
at both vegetative (three to four visible nodes) and reproductive
(four to five nodes with developing panicles) stages were
harvested and analysed. As PHB was not detected in roots
during preliminary experiments, root samples were not
included in this experiment. Four cab-m5 lines, representing
three different groups of plants with regard to previously
measured leaf polymer content in in vitro and soil-grown
plants, were analysed: a low-producing line containing
0.05% (in vitro) and 0.06% (mature leaf) dwt PHB; a medium
producer I with 0.50% and 0.44% dwt PHB, respectively; a
medium producer II containing 1.16% and 0.72% dwt PHB,
Figure 4 Polyhydroxybutyrate (PHB) accumulation in switchgrass leaves. (a–d) Nile blue staining of transverse sections of mature leaves. (a) A leaf section from a T0 plant transformed with the cab-m5 promoter construct under visible light. (b) The specimen shown in (a) under ultraviolet (UV) light. Note the polymer granules primarily in the bundle sheath cells. (c) A leaf section from a wild-type plant under visible light. (d) The leaf segment shown in (c) under UV light. Note the lack of polymer accumulation and the background staining observed in the vascular bundles. The stained samples were examined at × 20 magnification. B, bulliform cells; BS, bundle sheath cells; M, mesophyll cells; P, phloem; X, xylem; XV, xylem vessels. (e) Transmission electron micrograph (TEM) of a BS chloroplast with PHB granules from the cab-m5 plant shown in Figure 5b. (f) TEM of an M chloroplast from the same transgenic plant. The polymer content in the leaf region sampled for TEM was 1.60% dry weight (dwt). (g) TEM of a BS chloroplast from a wild-type plant. Note the large starch granules. (h) An M chloroplast with thylakoid membranes organized in grana from a wild-type plant. Bars, 1 μm. CW, cell wall; P, PHB granules; S, starch granules.

Polyhydroxybutyrate production in switchgrass 669
© 2008 Metabolix, Inc.Journal compilation © 2008 Blackwell Publishing Ltd, Plant Biotechnology Journal, 6, 663–678
respectively; and a high producer containing 0.46% and
2.28% dwt PHB, respectively. The polymer distribution in a
reproductive tiller from the high producer (Figure 6) revealed
a basipetal gradient of PHB accumulation. In general, the
mature tissues in the base of the tiller contained higher PHB
levels than the tissues at the top of the tiller. In addition, there
was a concentration gradient along each individual leaf. The
highest polymer content was measured in the oldest leaf
regions within 30 cm of the tip. These leaf parts accumulated
significant amounts of polymer (up to 3.72% dwt). Stem
tissues were found to contain much lower polymer levels
than the leaf blades (0–0.17% dwt). Similar PHB distribution
Figure 5 Effect of polyhydroxybutyrate (PHB) production on plant growth and development. (a) Plants from switchgrass genotype 56 after 2 months in the glasshouse: left, a wild-type (untransformed) plant; right, a cab-m5 plant (the high producer; see Table 2 and Figure 6) containing 2.27% dry weight (dwt) PHB in a sample from a mature leaf adjacent to the basal node. (b) Plants from genotype 215 after 2 months in the glasshouse: left, a plant transformed with the cab-m5 gene construct containing 2.52% dwt PHB in a sample from a mature leaf adjacent to the basal node; right, a wild-type plant.
Figure 6 (a) Spatial distribution of polyhydroxybutyrate (PHB) in a reproductive tiller from a high producer (Table 2). Sections from the nodes, internodes, leaf sheaths, leaf blades and panicle were cut, and the length and dry weight of each sample were recorded. Numbers indicate the PHB content (% dry weight) of each sample as measured by gas chromatography/mass spectrometry (GC/MS). (b) Scheme of the basic unit (phytomer) of the grass plant. in, internode; lb, leaf blade; ls, leaf sheath; n, node.

670 Maria N. Somleva et al.
© 2008 Metabolix, Inc.Journal compilation © 2008 Blackwell Publishing Ltd, Plant Biotechnology Journal, 6, 663–678
patterns were observed in the other lines producing smaller
amounts of polymer in soil (Table 2). Reproductive tillers of
each of the lines analysed contained more polymer than
vegetative tillers, suggesting a similar development-dependent
pattern of product accumulation despite significant differences
in the absolute PHB values amongst the lines. Biomass
partitioning data (Table 2) revealed that stem tissues primarily
accounted for the increased dry weight of flowering tillers.
Leaf : stem ratios of 0.9–1.1 in vegetative tillers dropped to
0.5–0.7 in tillers at a reproductive stage in the low and the
two medium producers, whereas flowering tillers from the
high producer maintained a relatively high (1.1) leaf : stem
ratio. The higher polymer levels in reproductive tillers
combined with insignificant changes in leaf biomass at this
developmental stage (0–28%) suggest that PHB synthesis
continues during plant maturation.
PHB production in the progeny of primary
transformants
The segregation ratios for Basta tolerance/susceptibility of T1
plants from five reciprocal crosses between transgenic plants
are shown in Table 3. All of the herbicide-tolerant seedlings
possessed pha genes, as confirmed by PCR. PHB production
in the progeny was analysed in leaf tissues collected from
transgenic seedlings at a growth stage comparable to the
stage previously used for the analysis of the parent plants in
tissue culture. The polymer content in individual T1 plants
varied from 0% to 1.82% dwt. Plants accumulating PHB at
similar or significantly higher levels (up to 155-fold increase)
than the parent lines were identified in the offspring
obtained from each cross (Table 3). These results indicated
that the pha genes were sexually transmitted through both
female and male gametes to the next generation.
The PHB content was also measured in mature and devel-
oping leaves from T1 plants grown in the glasshouse for
2 months. Changes in polymer production were found to
have patterns similar to those observed in the primary
Table 2 Polyhydroxybutyrate (PHB) production and biomass accumulation in leaf and stem tissues of vegetative and reproductive tillers from switchgrass lines transformed with the cab-m5 promoter construct
Transgenic
line
Sample
description
Vegetative stage Reproductive stage
dwt
(mg)
PHB
(% dwt)
dwt
(mg)
PHB
(% dwt)
Low producer Whole tiller 1918 0.01 2667 0.03
Leaf blade 918 0.01 1063 0.08
Leaf sheath 429 0 373 0
Internode 482 0 768 0
Node 89 0 121 0
Panicle – – 343 0
Medium producer I Whole tiller 1848 0.12 2880 0.20
Leaf blade 955 0.23 1225 0.46
Leaf sheath 329 0 553 0.01
Internode 470 0 777 0
Node 94 0 160 0
Panicle – – 202 0
Medium producer II Whole tiller 2287 0.18 3458 0.38
Leaf blade 1165 0.36 1171 1.09
Leaf sheath 318 0 468 0.07
Internode 689 0 1007 0.01
Node 97 0 138 0
Panicle – – 682 0.02
High producer Whole tiller 1250 0.89 1165 1.23
Leaf blade 673 1.62 606 2.18
Leaf sheath 214 0.06 210 0.14
Internode 303 0.01 238 0.07
Node 60 0.05 56 0.08
Panicle – – 56 0.39
dwt, dry weight.
Table 3 Segregation analysis and polyhydroxybutyrate (PHB) production in the T1 progeny obtained from controlled crosses between T0 plants. The χ2 test is based on the expected segregation ratio of 3 : 1. PHB contents were measured by gas chromatography/mass spectrometry (GC/MS) in leaf tissues from seedlings at a growth stage similar to the stage of the parent plants in tissue culture
Cross
Total
number
of seeds
Number
of T1 plants
T1 plants assayed
for Basta tolerance
Ratio
T : S χ2
PHB content (% dwt)
T0 plants T1 plants
Tolerant (T) Sensitive (S) Parent 1 Parent 2 Range Mean Median SD
1 515 489 372 117 3.2 : 1 0.30 0.18 0.26 0–1.82 0.06 0.01 0.17
2 171 159 117 42 2.8 : 1 0.17 1.79 0.01 0–1.55 0.32 0.20 0.39
3 178 154 117 37 3.2 : 1 0.08 0.44 0.39 0–1.60 0.17 0.05 0.28
4 110 105 79 26 3.0 : 1 0.01 0.31 0.39 0–1.24 0.14 0.06 0.22
5 78 74 53 21 2.5 : 1 0.45 0.15 0.11 0–0.65 0.15 0.07 0.16
Dwt, dry weight; SD, standard deviation.

Polyhydroxybutyrate production in switchgrass 671
© 2008 Metabolix, Inc.Journal compilation © 2008 Blackwell Publishing Ltd, Plant Biotechnology Journal, 6, 663–678
transformants after transfer to soil. For example, in the progeny
of a cross between low producers (cross 1 in Table 3), PHB
contents in mature leaves were similar to the contents at the
seedling stage in five of the ten T1 plants, reduced in three
plants, and four to five times higher (up to 4.19% dwt) in two
plants (Figure 7a). Polymer levels in the majority of the T1
plants obtained from cross 2 were lower in the mature leaves
than in the seedlings, but two plants were found to accumu-
late three- to sixfold more PHB during their growth in the
glasshouse (Figure 7b, T1 plants 2 and 6).
Discussion
Lignocellulosic biomass has received considerable attention
as an abundant feedstock for biofuels despite the high costs
associated with conversion processes. The USA has the
agricultural capability to grow vast quantities of this biomass,
with recent estimates exceeding one billion tons without
affecting food or feed (Perlack et al., 2005). These estimates
include 377 million dry tons of biomass from perennial
herbaceous crops that could be dedicated for conversion to
biofuels. Switchgrass, a warm season perennial grass, has
great potential as an industrial crop, as it requires minimal
inputs for growth in many agricultural regions of the USA and
Europe (Lewandowski et al., 2003) and has the ability to
sequester large amounts of carbon in the soil with its exten-
sive root system (Parrish and Fike, 2005). Direct production of
biobased polymers in switchgrass would yield an industrial
plant feedstock that could be converted into plastics and
fuels, providing better economics for both co-products.
In this study, the accumulation of biobased plastics directly
in the green tissues of switchgrass was investigated. The
chloroplast was chosen as the site for PHB synthesis in switch-
grass as this organelle has an endogenous flux of the polymer
Figure 7 Comparison of polyhydroxybutyrate (PHB) production in T0 plants and their progeny obtained from controlled crosses. Polymer contents in all transgenic seedlings were measured by gas chromatography/mass spectrometry (GC/MS) at a growth stage similar to the stage of the parent plants in tissue culture. From each cross, 10–19 randomly selected T1 plants containing different amounts of polymer were grown in the glasshouse for 2 months prior to the analysis of PHB production in their mature and developing leaves. (a) Cross 1: two low producers from genotypes 215 and 56 were used as parent plants in this cross. (b) Cross 2: the parent lines were a low producer and a medium producer from genotypes 92 and 56, respectively. Numbers 1–10 indicate individual T1 plants. dwt, dry weight.

672 Maria N. Somleva et al.
© 2008 Metabolix, Inc.Journal compilation © 2008 Blackwell Publishing Ltd, Plant Biotechnology Journal, 6, 663–678
precursor acetyl-CoA for fatty acid biosynthesis, and has
yielded the highest levels of polymer in plants to date (Bohmert
et al., 2000, 2004). Co-ordinated expression of the three
transgenes necessary for conversion of acetyl-CoA to PHB
was controlled by either the constitutive rice ubiquitin 2
promoter or the maize light-inducible cab-m5 promoter in
multigene expression constructs. It has been shown previously
that plants transformed with multigene constructs produce
higher levels of polymer than plants obtained from crossing
single transgene lines (Valentin et al., 1999; Bohmert et al.,
2000). Agrobacterium-mediated transfer of the transgenes
into highly embryogenic switchgrass callus cultures resulted
in the generation of 428 lines producing PHB at levels ranging
from less than 0.01% to 3.55% dwt. Interestingly, the trans-
formation efficiencies obtained with these multigene vectors
were similar to those previously reported with a basic selectable
and visual marker construct (Somleva et al., 2002; Somleva,
2006), demonstrating the robustness of the procedure.
During plant growth and development, differences in
polymer accumulation patterns were observed in plants
transformed with the two multigene constructs. Approximately
30% of the plants transformed with the cab-m5 promoter
construct contained similar or higher polymer values after
growth in soil than in tissue cultures, suggesting that polymer
production in these lines is not dependent on external carbon
sources as reported previously in tobacco (Bohmert et al.,
2002; Lössl et al., 2003). In contrast, values in plants engineered
with the rubi2 construct decreased in soil. Data for the
average PHB contents in switchgrass suggest that the maize
cab-m5 promoter exerts better control of transgene expression
than the constitutive rice ubiquitin 2 promoter for the synthesis
of polymer during plant maturation.
Examination of the spatial distribution of polymer in
switchgrass plants demonstrated that PHB accumulation was
consistently higher in fully developed mature tissues of the
plant. In a single leaf, polymer contents increased from the
base to the tip, and mature leaves yielded more polymer than
young developing leaves. PHB levels were also found to be
much higher in switchgrass leaves than stems. These findings
are in accordance with the recently published data for the
spatio-temporal pattern of polymer accumulation in sugar-
cane (Purnell et al., 2007), despite the different promoters
and transformation methods utilized. However, the maximum
amount of polymer in leaf tissues and whole tillers of switch-
grass was higher in our best characterized lines (leaf, 3.72%
dwt; whole tiller, 1.23% dwt) than the values reported for
sugarcane (leaf, 1.77% dwt; whole plant, 0.79% dwt).
Further developmental differences in PHB formation in
switchgrass were found in flowering and non-flowering
tillers. Tillers at a reproductive stage consistently accumulated
more polymer than vegetative tillers, suggesting that polymer
production still occurs at this later developmental stage.
At the cellular level, PHB granules in switchgrass leaves
were predominantly located in the chloroplasts of the bundle
sheath cells, although some inclusions were also visible in the
chloroplasts of the mesophyll cells. Similar analyses with
sugarcane (Petrasovits et al., 2007) and maize (Poirier and
Gruys, 2002) have also revealed abundant accumulation of
granules in the chloroplasts of the bundle sheath cells,
although with no apparent polymer formation in the mesophyll
chloroplasts. The factors responsible for the differences in
PHB production between the chloroplasts of the bundle
sheath and mesophyll cells are currently not known, but it has
previously been suggested that the availability of precursors
or the efficiency of plastid targeting to mesophyll chloroplasts
may be too low for sufficient polymer production (Poirier and
Gruys, 2002; Petrasovits et al., 2007). Transgene expression
from the maize cab-m5 promoter used in pMBXS155 and in
constructs for PHB production in maize (Poirier and Gruys,
2002) is unlikely to be a factor, as the cab-m5 gene has been
shown to be expressed in both mesophyll and bundle sheath
cells, with more abundant transcripts in mesophyll cells
(Sheen and Bogorad, 1986). The reasons for the differences
observed in polymer accumulation in the mesophyll cells of
switchgrass compared with sugarcane and maize are
currently unknown and require further study.
Consistent with studies in other plants, PHB production at
higher levels induced variable changes in the growth and fitness
of switchgrass plants. The highest producers containing up to
3.72% dwt PHB in leaf regions developed normally and
accumulated biomass. Only one plant with a leaf polymer
content of 3.55% dwt was stunted and exhibited strong
chlorophyll deficiency. Further studies are necessary to
establish whether the site of transgene insertion yielded
undesirable effects in this plant. The ability to generate fertile
PHB-producing plants phenotypically similar to wild-type
plants suggests that product accumulation at these levels
is well tolerated by switchgrass plants. An analysis of the
progeny of primary transformants indicates that stable,
high-level polymer production can be maintained in the next
generation.
Although the polymer levels in our transgenic lines were
lower than the 7.5% dwt threshold estimated to be necessary
for the commercialization of PHB-producing switchgrass
(Metabolix commercial target, biomass crops), this study is a
significant first step towards achieving this goal. Our work
also demonstrates the technology for efficient, large-scale
engineering of switchgrass with complex multigene pathways

Polyhydroxybutyrate production in switchgrass 673
© 2008 Metabolix, Inc.Journal compilation © 2008 Blackwell Publishing Ltd, Plant Biotechnology Journal, 6, 663–678
that will be necessary for the formation of many value-added
co-products in this important bioenergy crop.
Experimental procedures
Multigene constructs
pMBXS159
This binary vector contains expression cassettes for the three-genePHB biosynthetic pathway under the control of the rice polyubiquitin2 (rubi2) promoter (Wang et al., 2000). The PHB genes chosen forthis construct include a hybrid Pseudomonas oleovorans/Zoogloearamigera PHA synthase (Huisman et al., 2001) and the thiolase andreductase from Ralstonia eutropha (Peoples and Sinskey, 1989).Each PHA gene is fused to a plastid targeting sequence encoding thesignal peptide of the small subunit of ribulose-1,5-bisphosphatecarboxylase/oxygenase (rubisco) from pea and the first 24 aminoacids of the mature protein (Cashmore, 1983), as described previously(Kourtz et al., 2005). Plasmid pMBXS159 was constructed using thefollowing multistep procedure.
(i) pMBXS124, pMBXS125 and pMBXS126
These plasmids contain rubi2 and the 3′ termination sequence ofnopaline synthase (nos), and differ with respect to the restrictionsites available for subsequent cloning purposes (Table 4). The rubi2and nos fragments were isolated from template plasmids pRGL112-1 (Wang et al. 2000) and pNEB(A) (Kourtz et al., 2007), respectively,using standard PCR techniques. The rubi2 and nos fragments weresubcloned back into pRGL112-1, forming plasmids pMBXS124–126.
(ii) pMBXS135
Plasmid pMBXS135 is a pCAMBIA3300-derived vector in which themaize hsp70 intron (Brown and Santino, 1997) has been placedbetween the e35S promoter and bar to increase expression of theselectable marker in monocots. The maize hsp70 intron was isolatedby PCR from PCR-Ready maize genomic DNA (BioChain, Hayward,CA, USA) using primers BGNP108 and BGNP109, and insertedbetween the e35S promoter and bar gene using conventionalcloning techniques.
(iii) pMBXS154
Cassettes from pMBXS124, pMBXS125 and pMBXS126, containingrubi2 and nos, were sequentially transferred to pMBXS135 to formpMBXS154. Plasmid pMBXS124 was digested with SmaI and HindIII,and the rubi2 nos fragment was ligated into pMBXS135, which hadbeen digested previously with EcoRI, treated with the Klenowfragment of DNA polymerase 1 and digested with HindIII, to formintermediary plasmid pMBXS138. The rubi2 nos cassette of pMBXS125was isolated on digestion with BspEI and HindIII, and the resultingfragment was inserted into the equivalent sites of pMBXS138 toform pMBXS140. The rubi2 nos cassette of pMBXS126 was isolatedon digestion with BsrGI and HindIII, and inserted into the equivalentsites of pMBXS140 to form pMBXS154.
(iv) pMBXS159
The PHB biosynthetic genes, each modified with the plastid targetingsignal from pea (TS), were sequentially inserted into plasmid
pMBXS154 using the following multistep procedure to formpMBXS159. A PCR fragment containing TS-phaC was isolated frompNEB(C) (Table 4) using conventional PCR procedures, and insertedinto the AvrII and BamHI sites of pMBXS154 to form plasmidpMBXS156. A fragment containing TS-phaA-nos was isolated byPCR from plasmid pNEB(A), and inserted into plasmid pMBXS156,which had been digested previously with BsrGI, blunt-ended withKlenow and digested with BstBI, to generate pMBXS157. A frag-ment containing TS-phaB-nos was isolated by PCR from plasmidpNEB(B) (Table 4), and inserted into plasmid pMBXS157, which hadbeen digested previously with AscI, blunt-ended with Klenow anddigested with HindIII, to form plasmid pMBXS159.
pMBXS155
Plasmid pMBXS155 is a binary vector in which the PHB genes describedabove are expressed under the control of a maize chlorophylla/b-binding protein promoter (Sullivan et al., 1989). This promoter isequivalent to the cab-m5 promoter described in later work (Beckeret al., 1992). In pMBXS155, the cab-m5 promoter is fused to thehsp70 intron (Brown and Santino, 1997) for enhanced expression inmonocots. pMBXS155 was constructed using the following multistepprocedure.
(i) pMBXS137, pMBXS143 and pMBXS144
These plasmids contain cab-m5, the hsp70 intron and nos, and differwith respect to the restriction sites available for subsequent cloningpurposes (Table 4). The fragment containing cab-m5 was amplifiedfrom PCR-Ready maize genomic DNA (BioChain) with primersBGNP110 and BGNP111.
(ii) pMBXS148
Expression cassettes from plasmids pMBXS137, pMBSX143 andpMBXS144 were sequentially inserted into plant transformationvector pMBXS135 as follows. Plasmid pMBXS137 was digestedwith SmaI and HindIII, and the cab-m5/hsp70 nos fragment wasligated to plasmid pMBXS135, which had been digested previouslywith EcoRI, treated with Klenow and digested with HindIII, to createpMBXS145. This plasmid was modified by introducing an EcoRI siteat the 3′ end of nos to generate pMBXS146. The cab-m5/hsp70nos fragment of pMBXS143 was isolated with an EcoRI and HindIIIdigest, and cloned into the EcoRI and HindIII sites of pMBXS146to generate pMBXS147. The third cab-m5/hsp70 nos cassette wasisolated from pMBXS144 by digestion with BsrGI and HindIII and wascloned into the BsrGI and HindIII sites of pMBXS147 to createpMBXS148.
(iii) pMBXS155
A fragment containing TS-phaC was isolated from pNEB(C) usingconventional PCR procedures, and inserted into the AvrII and BamHIsites of pMBXS148 to form plasmid pMBXS151. Similarly, a fragmentcontaining TS-phaA-nos was isolated by PCR from plasmid pNEB(A),and inserted into plasmid pMBXS151, which had been digestedpreviously with BsrGI, blunt-ended with Klenow and digested withBstBI, to generate pMBXS153. Insertion of the PHB genes was com-pleted by isolating a fragment containing TS-phaB from plasmidpNEB(B) and inserting it at the PacI and AscI sites of pMBXS153 tocreate pMBXS155.

674 Maria N. Somleva et al.
© 2008 Metabolix, Inc.Journal compilation © 2008 Blackwell Publishing Ltd, Plant Biotechnology Journal, 6, 663–678
Table 4 Plasmids and primers used in this study
Relevant characteristics Reference
Plasmids
pRGL112-1 pUC19-based vector with rubi2 fused to GUS; source of rubi2 for subsequent cloning efforts Wang et al. (2000)
pNEB(A) pNEB193 derivative (New England Biolabs, Ipswich, MA, USA), contains 35S-C4PPDK promoter, TS, phaA, nos Kourtz et al. (2007)
pMBX124 pUC19-based vector; contains SmaI-rubi2 and AvrII-BamHI-nos-BspEI-HindIII cassettes This study
pMBX125 pUC19-based vector; contains BspEI-rubi2 and BstBI-nos-BsrGI-HindIII cassettes This study
pMBX126 pUC19-based vector; contains BsrGI-rubi2 and AscI-nos-HindIII cassettes This study
pCAM3300 pCAMBIA3300 binary vector; contains e35S bar expression cassette conferring resistance to bialaphos CAMBIA
pMBXS135 modified pCAMBIA3300 binary vector; contains e35S hsp70 bar cassette to increase bar expression This study
pMBXS138 pMBXS135 with rubi2-AvrII-BamHI-nos-BspEI-HindIII This study
pMBXS140 pMBXS135 with rubi2-AvrII-BamHI-nos-BspEI-rubi2-BstBI-nos-BsrGI-HindIII This study
pMBXS154 pMBXS135 with rubi2-AvrII-BamHI-nos-BspEI-rubi2-BstBI-nos-BsrGI-rubi2-AscI-nos-HindIII This study
pNEB(C) Similar to pNEB(A); contains 35S-C4PPDK promoter, TS, phaC, nos This study
pMBXS156 pMBXS135 with rubi2-AvrII-TS-phaC-BamHI-nos-BspEI-rubi2-BstBI-nos-BsrGI-rubi2-AscI-nos-HindIII This study
pMBXS157 pMBXS135 with rubi2-AvrII-TS-phaC-BamHI-nos-BspEI-rubi2-BstBI-TS-phaA-nos-rubi2-AscI-nos-HindIII This study
pNEB(B) Similar to pNEB(A); contains 35S-C4PPDK promoter, TS, phaB, nos This study
pMBXS159 pMBXS135 with rubi2-AvrII-TS-phaC-BamHI-nos-BspEI-rubi2-BstBI-TS-phaA-nos-rubi2-TS-phaB-nos-HindIII This study
pMBXS137 pNEB193-based vector with SmaI-cab-m5/hsp70 and AvrII-BamHI-nos-HindIII cassettes This study
pMBXS143 pNEB193-based vector with EcoRI-cab-m5/hsp70 and BstBI-nos-BsrGI-HindIII cassettes This study
pMBXS144 pNEB193-based vector; contains BsrGI-cab-m5/hsp70 and PacI, AscI-nos-HindIII cassettes This study
pMBXS145 pMBX135 with cab-m5/hsp70-AvrII-BamHI-nos-HindIII This study
pMBXS146 pMBX135 with cab-m5/hsp70-AvrII-BamHI-nos-EcoRI-HindIII This study
pMBXS147 pMBX135 with cab-m5/hsp70-AvrII-BamHI-nos-EcoRI-cab-m5/hsp70-BstBI-nos-BsrGI-HindIII This study
pMBXS148 pMBX135 with cab-m5/hsp70-AvrII-BamHI-nos-EcoRI-cab-m5/hsp70-BstBI-nos-BsrGI-cab-m5/
hsp70-PacI-AscI-nos-HindIII
This study
pMBXS151 pMBX135 with cab-m5/hsp70-AvrII-TS-phaC-BamHI-nos-EcoRI-cab-m5/hsp70-BstBI-nos-BsrGI-cab-m5/
hsp70-PacI-AscI-nos-HindIII
This study
pMBXS153 pMBX135 with cab-m5/hsp70-AvrII-TS-phaC-BamHI-nos-EcoRI-cab-m5/hsp70-BstBI-TS-phaA-nos-cab-m5/
hsp70-PacI-AscI-nos-HindIII
This study
pMBXS155 pMBX135 with cab-m5/hsp70-AvrII-TS-phaC-BamHI-nos-EcoRI-cab-m5/hsp70-BstBI-TS-phaA-nos-cab-m5/
hsp70-PacI-TS-phaB-AscI-nos-HindIII
This study
Primers
BGNP108 5′-AGATCTACCGTCTTCGGTACGCGCTC-3′ This study
BGNP109 5′-CCATGGCCGCTTGGTATCTGCATTAC-3′ This study
BGNP110 5′-GAA TTCACGGAAGATCCAGGT-3′ This study
BGNP111 5′-AGATCTTGCTGAAGCTGAGCGTGAAAG-3′ This study
Bar_fwd 5′-GCACCATCGTCAACCACTACATCG-3′ This study
Bar_rev 5′-TCATGCCAGTTCCCGTGCTTG-3′ This study
phaA_fwd 5′-ATCATCGCAAGACCGGCAACAG-3′ This study
phaA_rev 5′-TACAAGAGCTATGCCAACGC-3′ This study
phaB_fwd 5′-GCAAGACCGGCAACAGGATTCA-3′ This study
phaB_rev 5′-TCGGCGAGGTTGATGTGCTGAT-3′ This study
phaC_fwd 5′-GTAACATAGATGACACCGCG-3′ This study
phaC_rev 5′-GAAACAGCCTGAAAGTGCC-3′ This study
phaC_fwd1 5′-GCTGGGCGATATCAACAA-3′ This study
phaC_rev1 5′-GCACATAGTTCCATACCAGGTC-3′ This study
ActA 5′-CACTGGAATGGTCAAGGATG-3′ Okubara et al. (2002)
ActB 5′-CTCCATGTCATCCCAGTTG-3′ Okubara et al. (2002)
cab-m5/hsp70, cab-m5 promoter from maize fused to the hsp70 intron from maize; e35S promoter, double enhanced version of the 35S promoter
from cauliflower mosaic virus; GUS, β-glucuronidase gene; phaA, gene encoding thiolase; phaB, gene encoding reductase; phaC, gene encoding synthase;
nos, 3′ terminator from nopaline synthase gene; TS, plastid targeting sequence. Relevant restriction sites mentioned in the text are indicated in bold.

Polyhydroxybutyrate production in switchgrass 675
© 2008 Metabolix, Inc.Journal compilation © 2008 Blackwell Publishing Ltd, Plant Biotechnology Journal, 6, 663–678
Plant material, transformation and selection
Highly embryogenic callus cultures were initiated from maturecaryopses of cv. Alamo according to established procedures (Denchevand Conger, 1994). Cultures were grown at 28 °C in the dark andmaintained by monthly subcultures. After 3 months, callus regenerationability was tested. Cultures capable of producing more than 350plantlets per gram of callus were used for Agrobacterium-mediatedtransformation. Embryogenic cultures were infected and co-cultivatedwith Agrobacterium tumefaciens strain AGL1 carrying the binaryvector pMBXS155 or pMBXS159 in the presence of 100 μM ofacetosyringone, as described previously (Somleva et al., 2002;Somleva, 2006). Cultures were selected with 10 mg/L bialaphos for2–4 months. Resistant calluses were transferred to a regenerationmedium containing 10 mg/L bialaphos (Somleva et al., 2002;Somleva, 2006). Cultures were incubated at 28 °C with a 16-hphotoperiod (cool white fluorescent bulbs, 80 μmol/m2/s) for 4–6 weeks with biweekly subcultures. The resultant plantlets weretreated with the herbicide Basta, as described previously (Somlevaet al., 2002; Somleva, 2006). Transgenic and control plants weregrown in a glasshouse at 28 °C with supplemental lighting (16-hphotoperiod, sodium halide lamps).
PHB analysis
Leaf tissues (20–80 mg) from primary transformants in tissue culturewere collected, lyophilized and prepared for analysis by gas chroma-tography/mass spectrometry (GC/MS) using a previously describedsimultaneous extraction and butanolysis procedure (Kourtz et al.,2007). For screening of PHB-producing lines grown under glasshouseconditions, samples from mature leaves adjacent to the node at thebase of the stem and younger, still developing leaves at the top ofthe stem of plants grown in soil for 2 months were analysed. For spatialdistribution analysis, whole tillers (the basic units of harvestedswitchgrass biomass; Moser and Jennings, 2007) at vegetative andreproductive growth stages were used. The following tissues werecollected: nodes, internodes, leaf sheaths, leaf blades and panicles(from flowering tillers). Samples were prepared as described above,and analysed in selected ion monitoring mode using an Agilent 5973GC/MS instrument (Agilent, Santa Clara, CA, USA) equipped with aDB-225MS column and guard (Kourtz et al., 2007). The polymercontent is expressed as the percentage of PHB per unit dry weight ofplant tissue.
Molecular analyses
PCR
The presence of transgenes in plantlets regenerated from bialaphos-resistant calluses was detected in total nucleic acid extracts obtainedfrom leaf tissues of in vitro-grown plantlets with three to four leavesusing the REDExtract Plant PCR Kit (Sigma-Aldrich, St. Louis,MO, USA). Sequences of primers specific for the coding regions oftransgenes are listed in Table 4. The primer pairs were as follows:phaA, primers phaA_fwd and phaA_rev; phaB, primers phaB_fwdand phaB_rev; phaC, primers phaC_fwd and phaC_rev; bar, primersbar_fwd and bar_rev. PCR conditions for all amplifications were asfollows: 96 °C for 30 s (one cycle); 95 °C for 1 min, 57 °C for 1 min,72 °C for 1 min (35 cycles); 72 °C extension for 15 min.
Southern blot hybridizations
Switchgrass genomic DNA was isolated from young leaf tissues and,after complete digestion with SacI, was separated and blotted asdescribed previously (Somleva et al., 2002). Hybridizations wereperformed at 62 °C using fragments of the coding regions of the bar(394 bp) and phaB (578 bp) genes as probes. The digoxigenin-labelledprobes were generated by PCR amplification (PCR DIG Probe SynthesisKit, Roche, Indianapolis, IN, USA) using the primers in Table 4 and thePCR conditions described above. Digoxigenin-hybridized fragmentswere detected with the DIG Luminescent Detection Kit (Roche) usingDisodium 3-(4-methoxyspiro {1,2-dioxetane-3,2′-(5′-chloro) tricyclo[3.3.1.13,7] decan}-4-yl) phenyl phosphate (CSPD) as a chemilumines-cent substrate, according to the manufacturer’s protocol.
Expression analyses
For the analysis of transgene expression at mRNA and protein levels,mature and developing leaves from transgenic plants with differentPHB contents and control plants at the stage used for routine GC/MSanalysis (see above) were divided longitudinally into three parts forRNA extraction, protein isolation and PHB content determination.
RT-PCR
Leaf tissues (200–300 mg fresh weight) were used for total RNAisolation by phenol–chloroform extraction, followed by precipitationwith 8 M LiCl (Kay et al., 1987). RNA extracts were incubated withRNase-free RQ1 DNase (Promega, Madison, WI, USA) at 37 °C for10 min. Preliminary experiments showed no genomic DNA amplifi-cation products in the treated samples. After column purificationwith an RNeasy Mini Kit (Qiagen, Valencia, CA, USA), total RNA(0.5 or 1.0 ng) was subjected to reverse transcription and DNAamplification in a one-step RT-PCR assay (Qiagen) using the follow-ing gene-specific primers: phaA, primers phaA_fwd and phaA_rev;phaB, primers phaB_fwd and phaB_rev; phaC, primers phaC_fwd1and phaC_rev1; bar, primers bar_fwd and bar_rev; actin, primersActA and ActB (Table 4). The RT-PCR conditions for all amplificationswere as follows: 50 °C for 30 min; 95 °C for 15 min; 94 °C for1 min, 57 °C for 1 min, 72 °C for 1 min (35 cycles); 72 °C extensionfor 10 min. Preliminary experiments showed that the amplificationwas linear under these conditions. Actin amplification productswere used as a control for uniform loading (Okubara et al., 2002).Non-saturated signals were detected using ChemiDoc XRS (Biorad,Hercules, CA, USA).
Western blot analysis
Total soluble proteins were isolated from 200–300 mg of leaf tissuein 0.25 mL of a cell lysis buffer, as described previously (Kourtz et al.,2005). Protein concentrations were determined by the Bradfordassay (Bradford, 1976) using bovine serum albumin as a standard.Proteins (12 μg) were separated on 4%–12% polyacrylamidegradient gels (Invitrogen, Carlsbad, CA, USA). Samples of Ralstoniaeutropha thiolase and reductase (150 ng each) (Kourtz et al., 2005)were used as positive controls. Protein transfer on to polyvinylidenedifluoride membrane (GE Healthcare, Piscataway, NJ, USA) andmembrane blotting were performed as described previously(Kourtz et al., 2005) with slight modifications: the blocking andwashing buffers contained 0.5% (w/v) and 0.2% (w/v) gelatin,respectively. Polyclonal antibodies specific to the enzymes encoded

676 Maria N. Somleva et al.
© 2008 Metabolix, Inc.Journal compilation © 2008 Blackwell Publishing Ltd, Plant Biotechnology Journal, 6, 663–678
by the phaA and phaB genes (Kourtz et al., 2005) were used at adilution of 1 : 2000. Detection was performed using a goat anti-rabbit antiserum conjugated to alkaline phosphatase (dilution1 : 10 000) and 5-bromo-4-chloroindol-3-yl phosphate (BCIP) andnitroblue tetrazolium (NBT) (Roche Diagnostics, Indianapolis, IN,USA) as substrates. Non-saturated signals were visualized byChemiDoc XRS (Biorad).
Nile blue staining
Staining of leaf samples with Nile blue was performed as describedpreviously (Kourtz et al., 2005). Samples were viewed by fluorescencemicroscopy on a Zeiss Axiolab light microscope equipped with a ZeissHBO 100 fluorescence attachment (Zeiss, Thornwood, NY, USA)using the following filter set: exciter, HQ545/30; beam splitter,Q570lp; emitter D590/20 (Chroma Technology, Brattleboro, VT,USA). Images were recorded with a Zeiss MC 80 DX MicroscopeCamera using Kodak Elite Chrome 100 film (Kodak, Rochester,NY, USA).
Transmission electron microscopy
Plants were placed in a dark room for 22 h prior to sampling fortransmission electron microscopy to allow the degradation of starch.Segments from the basal leaf of tillers with four nodes were harvested6 cm below the tip of both wild-type and PHB-producing plants.Polymer contents in leaf segments were measured by GC/MS. Smallpieces of leaf were fixed in 2% paraformaldehyde and 2% glutaral-dehyde in sodium cacodylate buffer (50 mM, pH 7.0) for 6 h at22 °C. Samples were post-fixed in 1% osmium tetroxide in the samebuffer for 2 h, dehydrated in a graded series of acetone and embeddedin Spurr’s epoxy resin. Transverse sections (thickness, 60 nm) werestained with uranyl acetate and lead citrate and observed in a JEOLJSM-100s transmission electron microscope (JEOL USA, Inc., Peabody,MA, USA) operated at 80 kV.
Progeny analyses
As a result of the high self-incompatibility of switchgrass, T1 seedswere obtained from cross-pollination of transgenic plants fromdifferent Alamo genotypes used as both male and female parents, asdescribed previously (Somleva et al., 2002). Dry seeds from individualcrosses were germinated and the resultant seedlings were transferredto soil. Their response to the herbicide Basta was tested (Somlevaet al., 2002), and the presence of the transgenes in the tolerantplants was confirmed by PCR following the procedure for theidentification of primary transformants (see above). The segregationratios were analysed by the χ2 test at P = 0.05. Transgenic T1 plants(10–19 per cross) were grown in the glasshouse for further analyses.PHB contents were determined by GC/MS (Kourtz et al., 2007) inleaf samples collected from T1 plants at the developmental stagesused for analysis of the parent T0 plants in tissue culture and soil.
Acknowledgements
The authors thank Aminat Ali, Sean Daughtry, Mirel Sharxhi,
Steve Goldfless and Chris Johnson for technical contributions,
and James Oard of Louisiana State University for supplying
the plasmid pRGL112-1. Transmission electron microscopy
was performed by Dale Callaham at the Central Microscopy
Facility of the University of Massachusetts, Amherst, MA,
USA. This work was supported in part by a Department of
Energy Industry of the Future Award (DE-FC07–011D14214)
and a grant from the United States Department of Agriculture
(USDA-68-3A75-3-142).
References
Arai, Y., Nakashita, H., Doi, Y. and Yamaguchi, I. (2001) Plastidtargeting of polyhydroxybutyrate biosynthetic pathway intobacco. Plant Biotechnol. 18, 289–293.
Becker, T., Templeman, T., Viret, J. and Bogorad, L. (1992) Thecab-m7 gene: a light-inducible, mesophyll-specific gene of maize.Plant Mol. Biol. 20, 49–60.
Bevan, M., Barnes, W.M. and Chilton, M.-D. (1983) Structure andtranscription of the nopaline synthase gene region of T-DNA.Nucleic Acids Res. 11, 369–385.
Bieles, J. (2006) US facility brews up crop-based polymers. Plasticsand Rubber Weekly February 17, 1.
Bohmert, K., Balbo, I., Kopka, J., Mittendorf, V., Nawrath, C.,Poirier, Y., Tischendorf, G., Trethewey, R.N. and Willmitzer, L. (2000)Transgenic Arabidopsis plants can accumulate polyhydroxybutyrateto up to 4% of their fresh weight. Planta, 211, 841–845.
Bohmert, K., Balbo, I., Steinbuchel, A., Tischendorf, G. andWillmitzer, L. (2002) Constitutive expression of the β-ketothiolasegene in transgenic plants. A major obstacle for obtainingpolyhydroxybutyrate-producing plants. Plant Physiol. 128, 1282–1290.
Bohmert, K., Peoples, O.P. and Snell, K.D. (2004) Metabolicengineering: plastids as bioreactors. In: Molecular Biology andBiotechnology of Plant Organelles (Daniell, H. and Chase, C.D.,eds), pp. 559–585. Dordrecht: Kluwer Academic Publishers.
Bouton, J.H. (2007) Molecular breeding of switchgrass for use as abiofuel crop. Curr. Opin. Genet. Dev. 17, 553–558.
Bradford, M.M. (1976) A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein–dye binding. Anal. Biochem. 72, 248–254.
Brown, S.M. and Santino, C.G. (1997) Enhanced expression inplants. US Patent 05593874.
Cashmore, A.R. (1983) Nuclear genes encoding the small subunit ofribulose-1,5-bisphosphate carboxylase. In: Genetic Engineering ofPlants (Kosuge, T., Meredith, C.P. and Hollaender, A., eds),pp. 29–38. New York: Plenum.
Deligio, T. (2007) The greening of an industry: a growing market.Modern Plastics June 1. URL http://www.modplas.com/inc/mparticle.php?section=cvrStory&thefilename=cvrStory06012007_01.[accessed on 09 May 2008].
Denchev, P.D. and Conger, B.V. (1994) Plant regeneration fromcallus cultures of switchgrass. Crop Sci. 34, 1623–1627.
Department of Energy (DOE) (2006) Breaking the Biological Barriersto Cellulosic Ethanol: a Joint Research Agenda, DOE/SC-0095.Rockville, Maryland: US Department of Energy Office of Scienceand Office of Energy Efficiency and Renewable Energy. URLhttp://www.doegenomestolife.org/biofuels/ [accessed on 03 May2008].

Polyhydroxybutyrate production in switchgrass 677
© 2008 Metabolix, Inc.Journal compilation © 2008 Blackwell Publishing Ltd, Plant Biotechnology Journal, 6, 663–678
Greenspan, P. and Fowler, S. (1985) Spectrofluorometric studies ofthe lipid probe, Nile red. J. Lipid Res. 26, 781–789.
Herrera, S. (2006) Bonkers about biofuels. Nat. Biotechnol. 24, 755–760.
Houmiel, K.L., Slater, S., Broyles, D., Casagrande, L., Colburn, S.,Gonzalez, K., Mitsky, T.A., Reiser, S.E., Shah, D., Taylor, N.B., Tran, M.,Valentin, H.E. and Gruys, K.J. (1999) Poly(beta-hydroxybutyrate)production in oilseed leukoplasts of Brassica napus. Planta, 209,547–550.
Huisman, G.W., Skraly, F., Martin, D.P. and Peoples, O.P. (2001)Biological systems for manufacture of polyhydroxyalkanoatepolymers containing 4-hydroxyacids. US Patent 6,316,262.
John, M.E. and Keller, G. (1996) Metabolic pathway engineering incotton: biosynthesis of polyhydroxybutyrate in fiber cells. Proc.Natl. Acad. Sci. USA, 93, 12 768–12 773.
Kamm, B. and Kamm, M. (2007) Biorefineries – multiproduct processes.Adv. Biochem. Eng. Biotechnol. 105, 175–204.
Kay, R., Chan, A., Daly, M. and Mcpherson, J. (1987) Duplication ofCaMV 35S promoter sequences creates a strong enhancer forplant genes. Science, 236, 1299–1302.
Kourtz, L., Dillon, K., Daughtry, S., Madison, L.L., Peoples, O. andSnell, K.D. (2005) A novel thiolase–reductase gene fusion promotesthe production of polyhydroxybutyrate in Arabidopsis. PlantBiotechnol. J. 3, 435–447.
Kourtz, L., Dillon, K., Daughtry, S., Peoples, O.P. and Snell, K.D.(2007) Chemically inducible expression of the PHB biosyntheticpathway in Arabidopsis. Transgenic Res. 16, 759–769.
Lazo, G.R., Stein, P.A. and Ludwig, R.A. (1991) A DNA transformation-competent Arabidopsis genomic library in Agrobacterium. Bio/Technol. 9, 963–967.
Lewandowski, I., Scurlock, J.M.O., Lindvall, E. and Christou, M.(2003) The development and current status of perennial rhizomatousgrasses as energy crops in the US and Europe. Biomass Bioenerg.25, 335–361.
Lössl, A., Eibl, C., Harloff, H.J., Jung, C. and Koop, H.-U. (2003)Polyester synthesis in transplastomic tobacco (Nicotiana tabacumL.): significant contents of polyhydroxybutyrate are associatedwith growth reduction. Plant Cell Rep. 21, 891–899.
Lössl, A., Bohmert, K., Harloff, H., Eibl, C., Muhlbauer, S. and Koop,H.-U. (2005) Inducible trans-activation of plastid transgenes:expression of the R. eutropha phb operon in transplastomictobacco. Plant Cell Physiol. 46, 1462–1471.
Madison, L.L. and Huisman, G.W. (1999) Metabolic engineering ofpoly(3-hydroxyalkanoates): from DNA to plastic. Microbiol. Mol.Biol. Rev. 63, 21–53.
Moser, L.E. and Jennings, J.A. (2007) Grass and legume structureand morphology. In: Forages the Science of Grassland Agriculture(Barnes, R.F., Nelson, C.J., Moore, K.J. and Collins, M., eds),pp. 15–35. Ames, IA: Blackwell Publishing.
Okubara, P., Blechl, A., McCormick, S., Alexander, N., Dill-Macky, R.and Hohn, T. (2002) Engineering deoxynivalenol metabolism inwheat through the expression of a fungal trichotheceneacetyltransferase gene. Theor. Appl. Genet. 106, 74–83.
Parrish, D.J. and Fike, J.H. (2005) The biology and agronomy ofswitchgrass for biofuels. Crit. Rev. Plant Sci. 24, 423–459.
Peoples, O.P. and Sinskey, A.J. (1989) Poly-β-hydroxybutyratebiosynthesis in Alcaligenes eutrophus H16. Characterization ofthe genes encoding β-ketothiolase and acetoacetyl-CoA reductase.J. Biol. Chem. 264, 15 293–15 297.
Perlack, R.D., Wright, L.L., Turhollow, A.F., Graham, R.L., Stokes, B.J.
and Erbach, D.C. (2005) Biomass as Feedstock for a Bioenergyand Bioproducts Industry: the Technical Feasibility of a Billion-TonAnnual Supply. Oak Ridge, TN, USA: Oak Ridge Natl. Lab. ORNLPubl. No. TM-2005/66.
Petrasovits, L.A., Purnell, M.P., Nielsen, L.K. and Brumbley, S.M.(2007) Production of polyhydroxybutyrate in sugarcane. PlantBiotechnol. J. 5, 162–172.
Poirier, Y. and Gruys, K.J. (2002) Production of polyhydroxyal-kanoates in transgenic plants. In: Biopolymers: Polyesters I –Biological Systems and Biotechnological Production (Doi, Y. andSteinbüchel, A., eds), pp. 401–435. Weinheim: Wiley-VCH.
Purnell, M.P., Petrasovits, L.A., Nielsen, L.K. and Brumbley, S.M.(2007) Spatio-temporal characterisation of polyhydroxybutyrateaccumulation in sugarcane. Plant Biotechnol. J. 5, 173–184.
Ragauskas, A.J., Williams, C.K., Davison, B.H., Britovsek, G.,Cairney, J., Eckert, C.A., Frederick, W.J. Jr, Hallett, J.P., Leak, D.J.,Liotta, C.L., Mielenz, J.R., Murphy, R., Templer, R. and Tschaplinski,T. (2006) The path forward for biofuels and biomaterials. Science,311, 484–489.
Richards, H.A., Rudas, V.A., Sun, H., McDaniel, J.K., Tomaszewski, Z.and Conger, B.V. (2001) Genetic transformation and hybridization:construction of a GFP-BAR plasmid and its use for switchgrasstransformation. Plant Cell Rep. 20, 48–54.
Sanderson, M.A., Adler, P.R., Boateng, A.A., Casler, M.D. andSarath, G. (2006) Switchgrass as a biofuels feedstock in the USA.Can. J. Plant Sci. 86, 1315–1325.
Saruul, P., Srienc, F., Somers, D.A. and Samac, D.A. (2002) Pro-duction of a biodegradable plastic polymer, poly-β-hydroxybutyrate,in transgenic alfalfa. Crop Sci. 42, 919–927.
Schmer, M.R., Vogel, K.P., Mitchell, R.B. and Perrin, R.K. (2008) Netenergy of cellulosic ethanol from switchgrass. Proc. Natl. Acad.Sci. 105, 464–469.
Service, R.F. (2007) Cellulosic ethanol: biofuel researchers prepare toreap a new harvest. Science, 315, 1488–1491.
Sheen, J.-Y. and Bogorad, L. (1986) Differential expression of sixlight-harvesting chlorophyll a/b binding protein genes in maizeleaf cell types. Proc. Natl. Acad. Sci. USA, 83, 7811–7815.
Slater, S., Mitsky, T.A., Houmiel, K.L., Hao, M., Reiser, S.E., Taylor, N.B.,Tran, M., Valentin, H.E., Rodriguez, D.J., Stone, D.A., Padgette, S.R.,Kishore, G. and Gruys, K.J. (1999) Metabolic engineering ofArabidopsis and Brassica for poly(3-hydroxybutyrate-co-3-hydroxyvalerate) copolymer production. Nat. Biotechnol. 17,1011–1016.
Somleva, M.N. (2006) Switchgrass. In: Agrobacterium Protocols(Wang, K., ed.), pp. 65–74. Totowa, NJ, USA: Humana Press.
Somleva, M.N., Tomaszewski, Z. and Conger, B.V. (2002)Agrobacterium-mediated genetic transformation of switchgrass.Crop Sci. 42, 2080–2087.
Sullivan, T., Christensen, A. and Quail, P. (1989) Isolation andcharacterization of a maize chlorophyll a/b binding protein genethat produces high levels of mRNA in the dark. Mol. Gen. Genet.215, 431–440.
Suriyamongkol, P., Weselake, R., Narine, S., Moloney, M. and Shah, S.(2007) Biotechnological approaches for the production ofpolyhydroxyalkanoates in microorganisms and plants – a review.Biotechnol. Adv. 25, 148–175.
Valentin, H.E., Broyles, D.L., Casagrande, L.A., Colburn, S.M.,Creely, W.L., DeLaquil, P.A., Felton, H.M., Gonzalez, K.A.,Houmiel, K.L., Lutke, K., Mahadeo, D.A., Mitsky, T.A., Padgette,S.R., Reiser, S.E., Slater, S., Stark, D.M., Stock, R.T., Stone, D.A.,

678 Maria N. Somleva et al.
© 2008 Metabolix, Inc.Journal compilation © 2008 Blackwell Publishing Ltd, Plant Biotechnology Journal, 6, 663–678
Taylor, N.B., Thorne, G.M., Tran, M. and Gruys, K. (1999) PHAproduction, from bacteria to plants. Int. J. Biol. Macromol. 25,303–306.
Wang, J., Jiang, J. and Oard, J. (2000) Structure, expression andpromoter activity of two polyubiquitin genes from rice (Oryzasativa L.). Plant Sci. 156, 201–211.
Wrobel, M., Zebrowski, J. and Szopa, J. (2004) Polyhydroxybutyratesynthesis in transgenic flax. J. Biotechnol. 107, 41–54.
Wrobel-Kwiatkowska, M., Zebrowski, J., Starzycki, M., Oszmianski,J. and Szopa, J. (2007) Engineering of PHB synthesis causesimproved elastic properties of flax fibers. Biotechnol. Prog. 23,269–277.




















