
Microbial lipid-based lignocellulosic biorefinery: feasibility and challenges
12
Microbial lipid-based lignocellulosic biorefinery: feasibility and challenges Mingjie Jin 1 , Patricia J. Slininger 2 , Bruce S. Dien 2 , Suresh Waghmode 1 , Bryan R. Moser 2 , Andrea Orjuela 1 , Leonardo da Costa Sousa 1 , and Venkatesh Balan 1 1 Biomass Conversion Research Laboratory (BCRL), Department of Chemical Engineering and Materials Science, Michigan State University, MBI Building, 3815 Technology Boulevard, Lansing, MI 48910, USA 2 National Center for Agricultural Utilization Research, USDA-ARS, 1815 North University Street, Peoria, IL 61604, USA Although single-cell oil (SCO) has been studied for dec- ades, lipid production from lignocellulosic biomass has received substantial attention only in recent years as biofuel research moves toward producing drop-in fuels. This review gives an overview of the feasibility and challenges that exist in realizing microbial lipid produc- tion from lignocellulosic biomass in a biorefinery. The aspects covered here include biorefinery technologies, the microbial oil market, oleaginous microbes, lipid ac- cumulation metabolism, strain development, process configurations, lignocellulosic lipid production, techni- cal hurdles, lipid recovery, and technoeconomics. The lignocellulosic SCO-based biorefinery will be feasible only if a combination of low- and high-value lipids are coproduced, while lignin and protein are upgraded to high-value products. Lignocellulosic SCO biorefinery Second-generation biorefineries Conversion of food materials to fuels in first-generation biorefineries has sparked intensive debate and motivated the development of second-generation biorefineries, which use abundant non-edible lignocellulosic biomass as feedstock [1]. Life-cycle assessments (LCAs) of second-generation biorefineries showed greater greenhouse gas (GHG) emis- sion reduction than in first-generation biorefineries [2]. However, lignocellulosic biomass is highly recalcitrant due to its complex composition of cellulose, hemicellulose, and lignin [3]. Biorefineries that go with the biochemical route apply pretreatment to disrupt the plant cell-wall structure of lignocellulosic biomass, thus facilitating subsequent enzy- matic hydrolysis to obtain fermentable sugars [4]. The fermentable sugars can then be fermented by various micro- organisms into fuels and chemicals [5]. Fuel molecules that are undergoing or have undergone pilot scale trials include ethanol, butanol, isobutanol, biofene, bisbolene, organic acids, long-chain alcohols [6], and lipids from oleaginous microorganisms (SCO) [7]. Although the process cost of second-generation biorefineries is high, it is expected to decrease as the technology matures. SCO production in a second-generation biorefinery is the focus of this review. Potential markets for microbial lipids The lipids produced by oleaginous organisms are mostly in the form of Triacylglycerols (TAGs), with some in the form of free fatty acids (FAs). Lipids are attractive feedstocks for production of renewable fuels due to their high carbon- to-heteroatom ratios. The two most prevalent biofuels produced from lipids are biodiesel (via transesterification of TAGs) and renewable diesel (via hydrotreatment) [8]. The major process for transforming lipids into oleo- chemicals is the hydrolysis of TAGs into glycerol and FAs under the influence of water, temperature, and pressure (Figure 1A) [9–11]. Various products can be formed from FAs, such as oleic acid [10–13] (Figure 1B). Microbial lipids containing essential FAs (EFAs) are especially valuable in food applications [14]. Important EFAs include gamma linoleic acid (GLA), eicosapentaenoic acid (EPA), arachi- donic acid (ARA), and docosahexaenoic acid (DHA). Glyc- erol, produced as a byproduct during lipid processing, is an important feedstock for various C3 commodity chemicals that are currently produced in petrochemical processes from propylene [10–12] and has many possible applications and transformations (Figure 1C) [9,15–20]. SCO microbes and culture conditions Oleaginous microbes An ‘oleaginous’ microorganism is one that is able to accu- mulate greater than 20% of its dry body mass as ‘oil’ in the form of lipids, primarily as TAGs and FAs [7,21]. A literature survey by Subramaniam et al. [22] included: 14 genera of microalgae with the highest reported oil contents ranging from 20% to 77% of dry weight, most notably 50–77% for Schizochytrium sp.; four genera of bacteria with oil accumulations ranging from 24% to 78% of dry weight, the highest achieved by Arthrobacter sp. at >40%, and up to 78% from glucose [23]; four genera of yeast with oil contents ranging from 58% to 72% of cell dry weight, with a Rhodotorula glutinis strain accumulating the highest level; and four genera of molds with oil contents ranging from 57% to 86%, with a strain of Mortierella isabellina accumulating the highest level in the range. The basics of lipid biosynthetic metabolism is well understood (Figure 2 and Box 1) [24–27]. The theoretical yield of lipid production is 0.32 g/g sugar from glucose and Review 0167-7799/ ß 2014 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.tibtech.2014.11.005 Corresponding authors: Jin, M. ([email protected]); Balan, V. ([email protected]). Keywords: lignocellulosic lipids; single-cell oil; biorefinery; microbial lipids. Trends in Biotechnology, January 2015, Vol. 33, No. 1 43
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Microbial lipid-based lignocellulosic biorefinery: feasibility and challenges

Microbial lipid-based lignocellulosicbiorefinery: feasibility and challengesMingjie Jin1, Patricia J. Slininger2, Bruce S. Dien2, Suresh Waghmode1,Bryan R. Moser2, Andrea Orjuela1, Leonardo da Costa Sousa1, and Venkatesh Balan1
1 Biomass Conversion Research Laboratory (BCRL), Department of Chemical Engineering and Materials Science, Michigan State
University, MBI Building, 3815 Technology Boulevard, Lansing, MI 48910, USA2 National Center for Agricultural Utilization Research, USDA-ARS, 1815 North University Street, Peoria, IL 61604, USA
Review
Although single-cell oil (SCO) has been studied for dec-ades, lipid production from lignocellulosic biomass hasreceived substantial attention only in recent years asbiofuel research moves toward producing drop-in fuels.This review gives an overview of the feasibility andchallenges that exist in realizing microbial lipid produc-tion from lignocellulosic biomass in a biorefinery. Theaspects covered here include biorefinery technologies,the microbial oil market, oleaginous microbes, lipid ac-cumulation metabolism, strain development, processconfigurations, lignocellulosic lipid production, techni-cal hurdles, lipid recovery, and technoeconomics. Thelignocellulosic SCO-based biorefinery will be feasibleonly if a combination of low- and high-value lipids arecoproduced, while lignin and protein are upgraded tohigh-value products.
Lignocellulosic SCO biorefinerySecond-generation biorefineries
Conversion of food materials to fuels in first-generationbiorefineries has sparked intensive debate and motivatedthe development of second-generation biorefineries, whichuse abundant non-edible lignocellulosic biomass as feedstock[1]. Life-cycle assessments (LCAs) of second-generationbiorefineries showed greater greenhouse gas (GHG) emis-sion reduction than in first-generation biorefineries [2].However, lignocellulosic biomass is highly recalcitrant dueto its complex composition of cellulose, hemicellulose, andlignin [3]. Biorefineries that go with the biochemical routeapply pretreatment to disrupt the plant cell-wall structure oflignocellulosic biomass, thus facilitating subsequent enzy-matic hydrolysis to obtain fermentable sugars [4]. Thefermentable sugars can then be fermented by various micro-organisms into fuels and chemicals [5]. Fuel molecules thatare undergoing or have undergone pilot scale trials includeethanol, butanol, isobutanol, biofene, bisbolene, organicacids, long-chain alcohols [6], and lipids from oleaginousmicroorganisms (SCO) [7]. Although the process cost ofsecond-generation biorefineries is high, it is expected to
0167-7799/
� 2014 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.tibtech.2014.11.005
Corresponding authors: Jin, M. ([email protected]); Balan, V. ([email protected]).Keywords: lignocellulosic lipids; single-cell oil; biorefinery; microbial lipids.
decrease as the technology matures. SCO production in asecond-generation biorefinery is the focus of this review.
Potential markets for microbial lipids
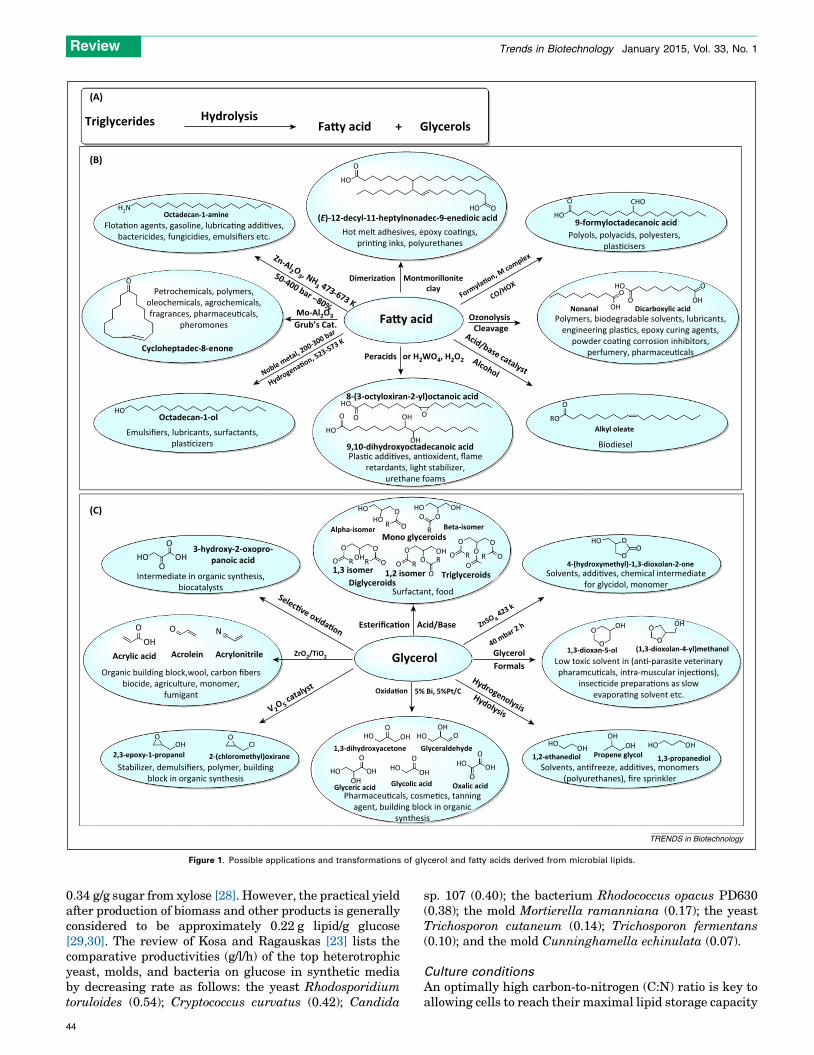
The lipids produced by oleaginous organisms are mostly inthe form of Triacylglycerols (TAGs), with some in the formof free fatty acids (FAs). Lipids are attractive feedstocksfor production of renewable fuels due to their high carbon-to-heteroatom ratios. The two most prevalent biofuelsproduced from lipids are biodiesel (via transesterificationof TAGs) and renewable diesel (via hydrotreatment)[8]. The major process for transforming lipids into oleo-chemicals is the hydrolysis of TAGs into glycerol and FAsunder the influence of water, temperature, and pressure(Figure 1A) [9–11]. Various products can be formed fromFAs, such as oleic acid [10–13] (Figure 1B). Microbial lipidscontaining essential FAs (EFAs) are especially valuable infood applications [14]. Important EFAs include gammalinoleic acid (GLA), eicosapentaenoic acid (EPA), arachi-donic acid (ARA), and docosahexaenoic acid (DHA). Glyc-erol, produced as a byproduct during lipid processing, is animportant feedstock for various C3 commodity chemicalsthat are currently produced in petrochemical processesfrom propylene [10–12] and has many possible applicationsand transformations (Figure 1C) [9,15–20].
SCO microbes and culture conditionsOleaginous microbes
An ‘oleaginous’ microorganism is one that is able to accu-mulate greater than 20% of its dry body mass as ‘oil’ inthe form of lipids, primarily as TAGs and FAs [7,21]. Aliterature survey by Subramaniam et al. [22] included:14 genera of microalgae with the highest reported oilcontents ranging from 20% to 77% of dry weight, mostnotably 50–77% for Schizochytrium sp.; four genera ofbacteria with oil accumulations ranging from 24% to78% of dry weight, the highest achieved by Arthrobactersp. at >40%, and up to 78% from glucose [23]; four genera ofyeast with oil contents ranging from 58% to 72% of cell dryweight, with a Rhodotorula glutinis strain accumulatingthe highest level; and four genera of molds with oil contentsranging from 57% to 86%, with a strain of Mortierellaisabellina accumulating the highest level in the range.
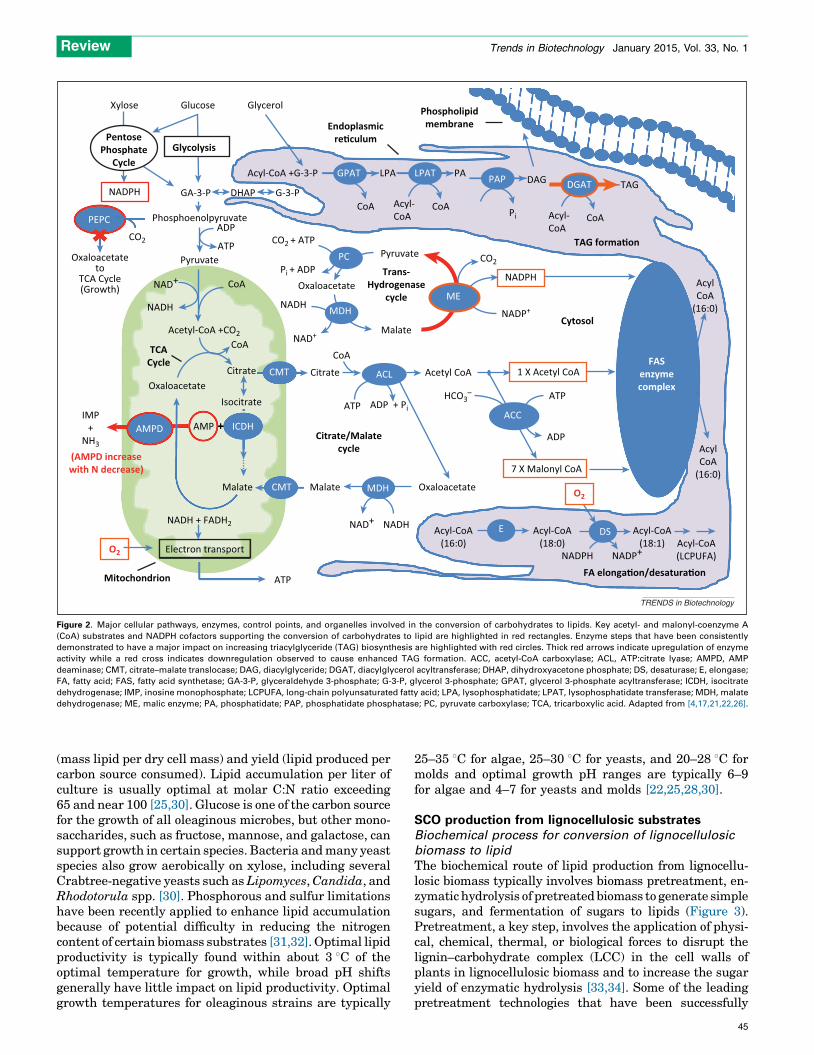
The basics of lipid biosynthetic metabolism is wellunderstood (Figure 2 and Box 1) [24–27]. The theoreticalyield of lipid production is 0.32 g/g sugar from glucose and
Trends in Biotechnology, January 2015, Vol. 33, No. 1 43
Triglycerides HydrolysisFa�y acid + Glycerols
Fa�y acid
Glycerol
(A)
(B)
(C)
H2N
O
Flota�on agents, gasoline, lubrica�ng addi�ves,bactericides, fungicidies, emulsifiers etc.
9-formyloctadecanoic acid(E)-12-decyl-11-heptylnonadec-9-enedioic acid
Nonanal
Alkyl oleate
9,10-dihydroxyoctadecanoic acid
8-(3-octyloxiran-2-yl)octanoic acid
4-(hydroxymethyl)-1,3-dioxolan-2-one
1,3-dioxan-5-ol
1,2-ethanediol1,3-dihydroxyacetone
Oxida�on
Esterifica�on
Alpha-isomer
1,3 isomer 1,2 isomer TriglyceroidsDiglyceroids
Beta-isomerMono glyceroids
Acid/Base
5% Bi, 5%Pt/C
FormalsGlycerol
Glyceric acid Glycolic acid
Glyceraldehyde
Oxalic acid
Propene glycol 1,3-propanediol
(1,3-dioxolan-4-yl)methanol
Dicarboxylic acid
Polyols, polyacids, polyesters,plas�cisers
Hot melt adhesives, epoxy coa�ngs,prin�ng inks, polyurethanes
CHO
HO
RO
HO
Surfactant, foodSelec�ve oxida�on
HydrogenolysisHydolysisV 2O 5 c
atalyst
HO
OHOH
HOHO
Plas�c addi�ves, an�oxident, flameretardants, light stabilizer,
urethane foams
OHO
O O OO O
OOO
OO
O
RR ROOO R R
ZrO2/TiO2
ZnSO 4 423 k
Zn-Al2 O3 , NH
3 473-673 K
50-400 bar ∼80%Formyla�on, M
complex
Noble metal, 200-300 bar
Hydrogena�on, 523-573 K
CO/HOX
40 mbar 2 h
R
OO
OR
R
OH
OH
Solvents, an�freeze, addi�ves, monomers(polyurethanes), fire sprinkler
OHOH OHHOHO
HO HO
Pharmaceu�cals, cosme�cs, tanningagent, building block in organic
synthesis
HO
HOOH
OH OHOH
OH
O
O
O
O O
O
OH
HO
Low toxic solvent in (an�-parasite veterinarypharamcu�cals, intra-muscular injec�ons),
insec�cide prepara�ons as slowevapora�ng solvent etc.
OH
O
O OO
OO
O
Solvents, addi�ves, chemical intermediatefor glycidol, monomer
O
Biodiesel
Polymers, biodegradable solvents, lubricants,engineering plas�cs, epoxy curing agents,
powder coa�ng corrosion inhibitors,perfumery, pharmaceu�cals
OO
O
OH
Dimeriza�on Montmorilloniteclay
OzonolysisMo-Al2O3Grub’s Cat.
Acid/base catalystAlcohol
Cleavage
Peracids or H2WO 4, H2O2
OH
HO
HO
OH
OH
OOO
HO
O
O
HO
HO O
Petrochemicals, polymers,oleochemicals, agrochemicals,fragrances, pharmaceu�cals,
pheromones
Cycloheptadec-8-enone
Emulsifiers, lubricants, surfactants,plas�cizers
Intermediate in organic synthesis,biocatalysts
Acrylic acid Acrolein
Stabilizer, demulsifiers, polymer, buildingblock in organic synthesis
2,3-epoxy-1-propanol 2-(chloromethyl)oxirane
Organic building block,wool, carbon fibersbiocide, agriculture, monomer,
fumigant
O OOH CI
HO OH
OHO O N
O
O
Acrylonitrile
HOOctadecan-1-ol
3-hydroxy-2-oxopro-panoic acid
Octadecan-1-amine
TRENDS in Biotechnology
Figure 1. Possible applications and transformations of glycerol and fatty acids derived from microbial lipids.
Review Trends in Biotechnology January 2015, Vol. 33, No. 1
0.34 g/g sugar from xylose [28]. However, the practical yieldafter production of biomass and other products is generallyconsidered to be approximately 0.22 g lipid/g glucose[29,30]. The review of Kosa and Ragauskas [23] lists thecomparative productivities (g/l/h) of the top heterotrophicyeast, molds, and bacteria on glucose in synthetic mediaby decreasing rate as follows: the yeast Rhodosporidiumtoruloides (0.54); Cryptococcus curvatus (0.42); Candida
44
sp. 107 (0.40); the bacterium Rhodococcus opacus PD630(0.38); the mold Mortierella ramanniana (0.17); the yeastTrichosporon cutaneum (0.14); Trichosporon fermentans(0.10); and the mold Cunninghamella echinulata (0.07).
Culture conditions
An optimally high carbon-to-nitrogen (C:N) ratio is key toallowing cells to reach their maximal lipid storage capacity
Xylose Glucose Glycerol
Endoplasmicre�culum
Phospholipidmembrane
PentosePhosphate
Cycle
NADPH
Glycolysis
PEPC
Oxaloacetateto
TCA Cycle(Growth)
CO2
CoA
CoA
ATP
ADP
CoA
ATP
NAD+
NADP+
CO2
CoACoACoA
ATP
ADP
Malate
ADP + Pi
NAD+
ATP
NAD+
Cytosol
Acetyl CoA
Malate
HCO3–
Oxaloacetate
NADH
MDH
ACC
ACLCMT
MDHME
PC
CMT
ICDHAMPAMPD
CitrateCitrate
IsocitrateOxaloacetate
Citrate/Malatecycle
Malate
Electron transport
Mitochondrion
NADH + FADH2
O2
O2
E DS
FASenzymecomplex
1 X Acetyl CoA
AcylCoA
(16:0)
AcylCoA
(16:0)
TAG forma�onPyruvate
NADH
7 X Malonyl CoA
Acyl-CoA(16:0)
Acyl-CoA(18:0)
Acyl-CoA(18:1)
Trans-Hydrogenase
cycleOxaloacetate
Pi
Pi + ADP
CO2 + ATP
TCACycle
NADH
Pyruvate
Acetyl-CoA +CO2
IMP+
NH3
(AMPD increasewith N decrease)
NADPH NADP+
FA elonga�on/desatura�on
Acyl-CoA(LCPUFA)
Phosphoenolpyruvate
GA-3-P G-3-P
Acyl-CoA +G-3-P LPA PAGPAT LPAT PAP DAG TAGDGAT
Acyl-CoA
NADPH
Acyl-CoA
DHAP
TRENDS in Biotechnology
Figure 2. Major cellular pathways, enzymes, control points, and organelles involved in the conversion of carbohydrates to lipids. Key acetyl- and malonyl-coenzyme A
(CoA) substrates and NADPH cofactors supporting the conversion of carbohydrates to lipid are highlighted in red rectangles. Enzyme steps that have been consistently
demonstrated to have a major impact on increasing triacylglyceride (TAG) biosynthesis are highlighted with red circles. Thick red arrows indicate upregulation of enzyme
activity while a red cross indicates downregulation observed to cause enhanced TAG formation. ACC, acetyl-CoA carboxylase; ACL, ATP:citrate lyase; AMPD, AMP
deaminase; CMT, citrate–malate translocase; DAG, diacylglyceride; DGAT, diacylglycerol acyltransferase; DHAP, dihydroxyacetone phosphate; DS, desaturase; E, elongase;
FA, fatty acid; FAS, fatty acid synthetase; GA-3-P, glyceraldehyde 3-phosphate; G-3-P, glycerol 3-phosphate; GPAT, glycerol 3-phosphate acyltransferase; ICDH, isocitrate
dehydrogenase; IMP, inosine monophosphate; LCPUFA, long-chain polyunsaturated fatty acid; LPA, lysophosphatidate; LPAT, lysophosphatidate transferase; MDH, malate
dehydrogenase; ME, malic enzyme; PA, phosphatidate; PAP, phosphatidate phosphatase; PC, pyruvate carboxylase; TCA, tricarboxylic acid. Adapted from [4,17,21,22,26].
Review Trends in Biotechnology January 2015, Vol. 33, No. 1
(mass lipid per dry cell mass) and yield (lipid produced percarbon source consumed). Lipid accumulation per liter ofculture is usually optimal at molar C:N ratio exceeding65 and near 100 [25,30]. Glucose is one of the carbon sourcefor the growth of all oleaginous microbes, but other mono-saccharides, such as fructose, mannose, and galactose, cansupport growth in certain species. Bacteria and many yeastspecies also grow aerobically on xylose, including severalCrabtree-negative yeasts such as Lipomyces, Candida, andRhodotorula spp. [30]. Phosphorous and sulfur limitationshave been recently applied to enhance lipid accumulationbecause of potential difficulty in reducing the nitrogencontent of certain biomass substrates [31,32]. Optimal lipidproductivity is typically found within about 3 8C of theoptimal temperature for growth, while broad pH shiftsgenerally have little impact on lipid productivity. Optimalgrowth temperatures for oleaginous strains are typically
25–35 8C for algae, 25–30 8C for yeasts, and 20–28 8C formolds and optimal growth pH ranges are typically 6–9for algae and 4–7 for yeasts and molds [22,25,28,30].
SCO production from lignocellulosic substratesBiochemical process for conversion of lignocellulosic
biomass to lipid
The biochemical route of lipid production from lignocellu-losic biomass typically involves biomass pretreatment, en-zymatic hydrolysis of pretreated biomass to generate simplesugars, and fermentation of sugars to lipids (Figure 3).Pretreatment, a key step, involves the application of physi-cal, chemical, thermal, or biological forces to disrupt thelignin–carbohydrate complex (LCC) in the cell walls ofplants in lignocellulosic biomass and to increase the sugaryield of enzymatic hydrolysis [33,34]. Some of the leadingpretreatment technologies that have been successfully
45

Box 1. Lipid biosynthesis and metabolism
The first event leading to lipid accumulation is believed to be a
nutrient limitation, typically nitrogen. When nitrogen is in short
supply, AMP deaminase activity is upregulated and it breaks down
AMP to release ammonia to nitrogen-starved cells. As a result, the
tricarboxylic acid cycle (TCA) is interrupted at isocitrate as its
required cofactor AMP declines in concentration with rising levels of
AMP deaminase activities. Accumulating citrate is then diverted
from the TCA cycle and transferred from mitochondria to the cytosol
by citrate–malate translocase, where ATP:citrate lyase (ACL) activity
cleaves it into acetyl-CoA and oxaloacetate. Oxaloacetate is
converted by malate dehydrogenase into malate, which can be
translocated to the mitochondria by citrate–malate translocase or
converted by ME into pyruvate with the production of NADPH and
CO2 through the cytosolic transhydrogenase cycle. The net acetyl-
CoA and NADPH produced as a result of nitrogen limitation then
flow into FA and TAG synthesis. It is notable that the presence of ME
activity varies by yeast strain. Particularly, some Lypomyces sp. and
Candida sp. strains do not have this activity and are believed to
produce the NADPH needed to support FA and TAG synthesis via
some other FA-dedicated enzyme system of the cytosol [24] or
perhaps via pentose phosphate pathway cycling [25].
Review Trends in Biotechnology January 2015, Vol. 33, No. 1
scaled up to industrially relevant conditions include Ammo-nia Fiber Expansion (AFEX), Dilute Acid (DA), SteamExplosion, and Organosolve. Enzymatic hydrolysis appliesenzyme cocktails comprising cellulases and hemicellulasesto hydrolyze the major polysaccharides (cellulose and hemi-cellulose) into simple sugars [35]. Since the sugars derivedfrom lignocellulosic biomass are mostly glucose and xylose,oleaginous strains that consume both glucose and xylose arepreferred, and the ability to consume minor sugars such asarabinose, mannose, or galactose is also desirable [36]. Twooptions for enzymatic hydrolysis are separate hydrolysisand fermentation (SHF) (Figure 3) and simultaneous sac-charification and fermentation (SSF). SSF converts simplesugars immediately as they are generated by enzymatichydrolysis, preventing the inhibition of enzymes by sugarand improving the yield of enzymatic hydrolysis [37]. SSFalso reduces the contamination risk and capital cost oflipid production. Enzyme production can be integrated intothe SSF system and the process configuration then becomes
Enzyme produc(Fermenta�o
either on-site or Microbial
strain seed(e.g. Trichoderma reesei )
Enzymes(e.g. cellulases
& hemicellulases)
Water& nutrients
Water Water(if applicable)
PretreatmentEnzyma�chydrolysis
Solid/liquidsepara�on
(if applicable)
Solid/liquidsepara�onPretreated
biomass Slurry
Unhs
Combus�onHeat or electr
Chemicals(e.g. sulfuric acid,ammonia, sodium
hydroxide)
Lignocellulosicbiomass
Glucose orPretreated
biomass
Figure 3. Biochemical route for lipid prod
46
Consolidated Bioprocessing (CBP), which is deemed theultimate low-cost process for biofuel production [38]. Sub-merged culture is the dominant culture method used in theliterature for lipid production from lignocellulosic biomass,but solid-state fermentation is also a viable option [39]. How-ever, solid-state fermentation has drawbacks in areas suchas heat and mass transfer, scale-up and cell and lipidharvest.
Examples of lignocellulose conversion to SCO
In cellulosic lipid research efforts, oleaginous yeast andfungal strains were the main organisms investigated(Table 1). Heterophilic cultivation of microalgal strainsfor the conversion of organic substances to lipids has alsoattracted attention [40]. However, only a few studies haveused real lignocellulose-derived sugars [40–43]. Most ofthese microalgal studies focused on the inhibitory effectof degradation products, since this effect limits the utiliza-tion of microalgae for lipid production from lignocellulosicbiomass [43].
The lipid content of cell biomass generated duringfermentation of lignocellulosic hydrolysates was loweroverall than that seen in synthetic media. For instance,R. toruloides Y4 accumulated 51.8% of its cell biomass aslipid in a synthetic medium, but only 36.4% in the ionicliquid corn stover hydrolysate with similar cell biomassproduction [44]. The maximal lipid content seen in theliterature is 58.5% (Table 1). Lipid yield is an essentialparameter that affects the cost of lignocellulosic lipid pro-duction. Lipid yields were often below 0.25 g/g consumedsugar (<74% of the theoretical maximum) (Table 1), withone exception, which reached as high as 0.33 g/g consumedsugar (almost the theoretical maximum) in a detoxifiedhydrochloric acid hydrolysate of sugarcane bagasse usingYarrowia lipolytica Po1 g [45]. The same strain, however,reached a lipid yield of only 0.10 g/g in a detoxified sulfuricacid hydrolysate of rice bran [46].
Lipid fermentation is generally a slower process comparedwith ethanol fermentation. However, aerobic fermentation
�onn):off-site
Oleaginous microbe seed& nutrients (if applicable)
Oleaginousmicrobe cells
Microbial meal(∼50% protein)
Lipids
Air
Hydrolysate
Liquid broth
Fermenta�onbroth
ydrolyzedolids Lipid
Fermenta�on foricity
Lipidextrac�on
Solid/liquidsepara�on
Waste watertreatment
TRENDS in Biotechnology
uction from lignocellulosic biomass.

Table 1. Examples of lignocellulose conversion to SCO
Feedstock Pretreatment Microbial strain Fermentation medium/mode Lipid
(g/l)
Fermentation
product (g/l/d)
Lipid
content
Lipid yield
(g/g consumed
carbon source)
Lipid yield
(g/g
feedstock)
Note (shake-flask scale
unless otherwise
stated)
Refs
Corn stover DA Mortierella
isabellina
ATCC 42613
Enzymatic hydrolysate
of whole slurry
4.8 1.20 34.5% 0.15 N/A Liquid was separated
from solids after
pretreatment,
neutralized, and then
combined again with
solids before enzymatic
hydrolysis
[76]
Corn stover DA Trichosporon
cutaneum CX1
SSF 3.2 0.81 N/A N/A N/A Pretreated biomass was
detoxified using
Amorphotheca resinae
ZN1 at 25 8C for 3 days;
10% (w/w) solid loading
during SSF was applied;
50-l scale
[51]
Corn stover DA T. cutaneum
AS 2.571
Enzymatic hydrolysate 7.6 1.89 39.2% 0.15 N/A Glucose and xylose were
consumed
simultaneously; 3-l scale
[77]
Corn stover DA T. cutaneum
CX1
Enzymatic hydrolysate
of detoxified biomass
3.1 1.24 23.5% 0.06 N/A Pretreated biomass was
detoxified using A.
resinae ZN1; 3-l scale
[78]
Corn stover DA Rhodotorula
graminis
Enzymatic hydrolysate 14.4 5.04 34% 0.08 N/A Strain consumes
glucose, xylose,
galactose, mannose,
cellobiose, and glycerol;
20-l scale
[79]
Corn stover DA M. isabellina
ATCC 42613
Enzymatic hydrolysate
of whole slurry
3.2 0.87 24.8% 0.12 0.063 Initial hydrolysate acetic
acid, HMF, and furfural
concentrations were 2.4,
0.02, and 0.19 g/l,
respectively
[80]
Switchgrass DA As above As above 4.4 0.90 35.6% 0.15 0.072 Initial hydrolysate acetic
acid, HMF, and furfural
concentrations were
3.25, 0.07, and 0.97 g/l,
respectively
[80]
Miscanthus DA As above As above 3.7 0.76 32.2% 0.13 0.050 Initial hydrolysate acetic
acid, HMF, and furfural
concentrations were
5.01, 0.07, and 0.92 g/l,
respectively
[80]
Giant reed DA As above As above 3.0 0.53 21.2% 0.11 0.042 Initial hydrolysate acetic
acid, HMF, and furfural
concentrations were 6.2,
0.19, and 0.51 g/l,
respectively
[80]
Wheat straw DA Yarrowia
lipolytica
Non-detoxified liquid
stream of DA
0.4 0.06 4.6% 0.01 0.032 Detoxification did not
improve fermentation
performance
[81]
Revie
wT
rends
in B
iote
chnolo
gy
January
2015,
Vol.
33,
No.
147

Table 1 (Continued )
Feedstock Pretreatment Microbial strain Fermentation medium/mode Lipid
(g/l)
Fermentation
product (g/l/d)
Lipid
content
Lipid yield
(g/g consumedcarbon source)
Lipid yield
(g/gfeedstock)
Note (shake-flask scale
unless otherwisestated)
Refs
Wheat straw DA Cryptococcus
curvatus
Non-detoxified liquid
stream of DA
5.8 0.96 33.5% 0.20 0.047 Detoxification did not
improve fermentation
performance
[81]
Wheat straw DA Rhodotorula
glutinis
As above 3.5 0.58 25.0% 0.12 0.028 Detoxification did not
improve fermentation
performance
[81]
Wheat straw DA Lipomyces
starkeyi
As above 4.6 0.76 31.2% 0.16 0.037 Detoxification did not
improve fermentation
performance
[81]
Wheat straw DA Rhodosporidium
toruloides
Detoxified liquid
stream of DA
2.4 0.41 24.6% 0.08 0.019 Strain did not grow in
non-detoxified medium
[81]
Wheat straw DA M. isabellina
NRRL 1757
Liquid stream of DA 4.3 0.72 34% 0.17 0.044 Beside glucose, xylose,
arabinose, and
galactose, acetic acid
and furan were also
metabolized
[82]
Corncob DA Trichosporon
coremiiforme
Detoxified liquid
stream of DA
7.7 0.96 37.8% 0.17 N/A Strain fermented
glucose, xylose, and
arabinose
[83]
Rice straw DA Trichosporon
fermentans
CICC 1368
Detoxified liquid
stream of DA
11.5 1.44 40.1% 0.1 N/A Strain grew poorly in
undetoxified
hydrolysate; consumed
glucose, xylose,
mannose, galactose, and
cellobiose
[84]
Sugar cane bagasse DA T. fermentans Detoxified liquid
stream of DA
15.8 1.76 N/A 0.14 N/A Fermentation conditions
were optimized using
Box–Behnken design
[85]
Miscanthus D Neurospora
crassa
Whole pretreatment
slurry
1.3 0.32 N/A N/A 0.038 Genetically engineered
strain with glycogen
synthase deleted and
acetyl-CoA synthase
enhanced
[60]
Corn stover Ionic liquid Cryptococcus
curvatus
SSF 6.0 3.00 N/A N/A 0.081 High inoculation size
(initial fermentation cell
density 5.4 g/l) was
applied
[52]
Corn stover Ionic liquid As above SHF 7.2 3.60 43.4% 0.23 0.100 High inoculation size was
used, as above; more
cellulases were used
than above
[52]
Corn stover Ionic liquid R. toruloides Y4 Enzymatic hydrolysate 5.5 0.83 36.4% 0.10 N/A Lipid fermentation
conditions were not
optimized
[44]
Corn stover Dilute alkali
(NaOH)
M. isabellina
ATCC 42613
Enzymatic hydrolysate
of whole slurry
2.5 0.65 29.5% 0.09 N/A Liquid was separated
from solids after
pretreatment,
neutralized, and then
[76]
Revie
wT
rends
in B
iote
chnolo
gy
January
2015,
Vol.
33,
No.
1
48

combined again with
solids before enzymatic
hydrolysis
Sugarcane bagasse Hydrochloric
acid hydrolysis
Y. lipolytica Po1g Detoxified acid
hydrolysate
6.7 1.67 58.5% 0.33 N/A Strain fermented both
glucose and xylose
[45]
Defatted rice bran Sulfuric acid
hydrolysis
Y. lipolytica Po1g Detoxified acid
hydrolysate
5.2 N/A 48.0% 0.10 N/A Detoxification was
achieved by neutralizing
the hydrolysate using
Ca(OH)2
[46]
Soybean hull None M. isabellina Solid-state fermentation N/A N/A N/A N/A 0.048 Results were based on
8 weeks of cultivation
[39]
Pelletized wood Pyrolysis C. curvatus Fermentation of
carboxylic
acids in pyrolytic
aqueous phase
2.2 0.55 31.9% 0.11 N/A Bio-oil and pyrolytic
aqueous phase were
separated after pyrolysis;
aqueous phase was
detoxified and fermented
[54]
Corncob Organic solvents Trichosporon
dermatis CH007
Enzymatic hydrolysate 9.8 1.40 40.1% 0.17 N/A Enzymatic hydrolysis
was performed using in-
house-produced
Trichoderma reesei
enzymes
[86]
Kraft hardwood pulp None Rhodococcus
opacus PD630 Xsp8
Enzymatic hydrolysate 11.0 1.57 45.8% 0.18 N/A Strain was genetically
engineered for xylose
consumption
[87]
Avicel None Mucor circinelloides CBP with supply of
cellobiohydrolases
0.7 N/A 3.32% 0.06 0.025 Strain contains genes
encoding
13 endoglucanases,
three beta-D-
glucosidases, and
243 other glycoside
hydrolase proteins, but
not genes encoding
exoglucanases
[53]
Rice straw Trifluoroa-cetate Chlorella
pyrenoidosa
Enzymatic hydrolysate 1.59 0.80 56.3% 0.12 N/A One of the few
microalgal studies in real
lignocellulosic
hydrolysate
[41]
Corn stover DA, alkali M. isabellina Enzymatic hydrolysate
of mixture of DA- and
alkali-pretreated biomass
6.9 0.90 37% 0.15 N/A Glucose, xylose, and
acetate were all
consumed; 7.5-l scale
[88]
Revie
wT
rends
in B
iote
chnolo
gy
January
2015,
Vol.
33,
No.
149

Review Trends in Biotechnology January 2015, Vol. 33, No. 1
facilitates xylose consumption during lipid fermentation,which is a major issue in most cases during lignocellulosicethanol fermentation [47]. As a result, lipid fermentationtime is comparable with ethanol fermentation time. Never-theless, since lipid yield (typically <0.25) is much lower thanethanol yield (typically 0.40–0.51), lipid productivities arerelatively low, with a maximum of 5.04 g/l/day (Table 1). Toobtain high lipid productivities, high lipid titers are desir-able. High lipid titers also benefit downstream processing.Although lipid titers above 70 g/l were achieved in syntheticmedia through sugar fed-batch strategies [48,49], lignocellu-losic lipid production was below 20 g/l (Table 1). To achievehigher lipid titers, higher sugar concentrations are a prereq-uisite, which in turn require greater lignocellulosic biomassloading during enzymatic hydrolysis. Unless extensive de-toxification is applied, greater lignocellulosic biomass load-ing inevitably leads to higher levels of degradation productsor inhibitors generated during pretreatment, which thenflow into the lipid fermentation system and reduce lipidyield. Moreover, greater lignocellulosic biomass loadingleads to lower sugar yield during hydrolysis, partly due toend-product feedback inhibition [50], and eventually resultsin lower lipid yield per unit lignocellulosic biomass processed.Ideally, the SSF process prevents end-product inhibition andimproves the sugar yield of enzymatic hydrolysis. Lignocel-lulosic biomass can also be fed in several batches, to reducethe initial inhibitor concentration and viscosity burdens ofhigh solid loading. However, SSF has two major drawbacks.First, SSF is typically conducted at optimal conditions forfermentation (�30 8C, pH �5.0) whereas enzymatic hydro-lysis has different optimal operating conditions (�50 8C, pH�4.8) that lead to slow sugar release. Sugar release ratesmost likely decrease as SSF proceeds because the substratebecomes more recalcitrant. Once the sugar release rate isbelow what is required for cell maintenance, the microbestend to metabolize accumulated lipid as an energy source, atwhich point SSF has to be terminated. Additionally, whenunhydrolyzed solids from lignocellulose are in the fermenta-tion broth, as in the SSF operation, the cost of lipid recovery isincreased and the separation of yeast protein coproducts iscomplicated. Thus, lignocellulose-to-lipid conversion via SSFmay not yet be a satisfactory approach.
Currently, only a few studies have applied SSF tolipid production (Table 1). Liu et al. compared SSF withSHF on DA-pretreated corn stover and found that SSFyielded more lipids [51]. Gong et al. also compared SSFand SHF on ionic liquid-pretreated corn stover butapplied a different fermentation strategy [52]. They grewthe C. curvatus strain in rich medium for cell propaga-tion and then inoculated these cells at a high inoculationsize into either SSF or SHF. No nutrients were addedfor SSF or SHF so that cell propagation was limited andconsumed sugars were channeled into lipid production.Relatively high lipid yields and fermentation productivi-ty were achieved by this approach (Table 1). CBP wasalso attempted for lipid production, resulting in lowyields [53]. Researchers have tried to couple lipid fer-mentation with pyrolysis of lignocellulosic biomass. Forinstance, Lian et al. converted carboxylic acids generatedfrom pyrolysis into lipids through C. curvatus fermenta-tion [54].
50
Technical hurdles
Low lipid content, low lipid yield, low lipid productivity,and low lipid titer are mainly caused by several technicalchallenges in the culture of oleaginous strains on lignocel-lulosic hydrolysates. These challenges include: (i) limitedability to control C:N ratio because of the need to addnitrogen-rich commercial cellulases and hemicellulases;(ii) limited sugar concentrations (e.g., �100 g/l); and (iii)the presence of sugar mixtures (e.g., diauxic growtheffects). However, inhibitor tolerance is the most immedi-ate technical roadblock. Inhibitors are side-productsformed during pretreatment and can include furfural,hydroxymethylfurfural (HMF), acetic acid, neutral andacidic phenolics, and other chemical species [55]. Managinginhibitors and developing yeasts capable of tolerating themhas been the subject of considerable research in cellulosicethanol production. R. toruloides was screened against sixcommon inhibitors and as little as 1 mM furfural inhibitedcell growth by 45% [56]. Some microbes can reduce furfuralto furfuryl alcohol or oxidize it to furoic acid. R. toruloideswas equally sensitive to each of these intermediates.By contrast, the maximal HMF concentration studied(14 mM) decreased cell growth by only approximately10%. The lignin-associated inhibitors p-hydroxybenzalde-hyde and vanillin were toxic at 10 mM. A more extensiveset of studies [57–59] using T. fermentans observed asimilar pattern where lipid production was reduced by50% at a much lower concentration of furfural (4.7 mM)than of HMF (37.7 mM) and the yeast was equally sensi-tive to furfural alcohol (3.8 mM). The yeast was also sensi-tive to ferulic acid (7.7 mM), which may be problematic forherbaceous plant biomass. For both yeasts, FA compositionwas unaltered by the presence of the inhibitors.
Strain development
At minimum, oleaginous strains need to have high growthrates, productivities, and yields. More specialized traits arealso needed to allow the efficient use of hydrolysates forlipid production, including the ability to metabolize a broadrange of sugars and tolerance to inhibitors present in hydro-lysates under diverse nutritional environments. The oleag-inous strains reported so far are not as efficient or robustas would be preferred for industrial applications (Table 1).It is anticipated that the combination of screening microbialdiversity, directed evolution, and pathway-engineeringapproaches can lead to commercially useful strains for lipidproduction from lignocellulosic biomass substrates.
Numerous studies have been conducted using metabol-ic-engineering techniques to enhance lipid accumulation invarious species, including: (i) overexpression of FA biosyn-thetic enzymes; (ii) overexpression of TAG biosynthesis-enhancing enzymes; (iii) regulation of TAG bypass routes;(iv) blocking competing pathways; and (v) multigene trans-genics [27]. Of these approaches, the second, third, andfourth have achieved the most success due to overexpres-sion of acyl-coenzyme A (CoA):diacylglycerol acyltransfer-ase (DGAT), overexpression of malic enzyme (ME), and therepression of phosphoenolpyruvate (PEP) carboxylase(PEPC), respectively. The conversion of diacylglycerides(DAGs) to TAGs catalyzed by DGAT is rate limiting[27]. The activity of ME is important for the flux of carbon

Review Trends in Biotechnology January 2015, Vol. 33, No. 1
to lipid biosynthesis because it supplies NADPH in thecytosol to lipogenic enzymes such as FA synthase anddesaturase [24]. A third primary control point is believedto be PEPC, which catalyzes the conversion of PEP tooxaloacetate for direct participation in the anabolism ofprotein. The literature suggests that there is a crucial rolefor PEPC in the regulation of lipid biosynthesis in plantsand microalgae, but this has not been found in fungi[27]. Roche et al. genetically modified a filamentous fungus,Neurospora crassa, by deleting glycogen synthase andenhancing acetyl-CoA synthase and obtained a strain ac-cumulating 2.3 times more FAs on pretreated lignocellu-losic biomass [60].
Lipid recovery
Lipid recovery from fermentation broth involves harvestingmicrobial cells from the broth, either by drying cell biomassor by forcing cell disruption, and lipid extraction. Cell har-vest is expensive when cell density is low in the fermentationbroth, which renders high-cell-density fermentation desir-able. Commonly used cell-harvesting methods include cen-trifugation, filtration, and coagulation or flocculation[61]. Drying cell biomass after harvesting typically leadsto higher lipid yield during extraction compared with dis-rupted wet cell biomass [62]. However, drying is an energy-intensive process. Therefore, numerous methods have beeninvented, such as high-pressure homogenization, bead beat-ing, ultrasonication, microwave treatment, enzymatic hy-drolysis of cell walls, and acid hydrolysis, to efficientlydisrupt wet microbial cells so that high lipid yield can alsobe obtained during extraction [62–64]. The Bligh and Dyermethod and the Soxhlet method are traditionally used forlipid extraction using organic solvents such as chloroform,methanol, or hexane [64]. While solvent extraction is anefficient and mature technology, the solvent-recovery pro-cess is energy intensive. Instead of organic solvents, super-critical liquids (e.g., CO2, N2) have also been applied to lipidextraction so that energy-intensive solvent recovery isavoided [62,64]. In the case of using lipids for biodieselapplications, transesterification is sometimes applied di-rectly on either dry or wet cell biomass so that extractionand transesterification occursimultaneously [64,65]. Hydro-thermal liquefaction technology, which has been used foralgal biomass conversion to bio-oil and nutrient recycling forcultivation of algae [66], is potentially a promising technol-ogy for lignocellulosic lipid biorefineries. Coproducts such aspolysaccharides can also be generated through sequentialhydrothermal liquefaction [67]. Other energy-extractionmethods that can be applied include pyrolysis, gasification,and direct combustion [68].
Development of a SCO biorefineryFeedstocks and microbes for SCO biorefineries
In early explorations, expensive carbon sources such asstarch were the substrates of interest, but the microbialproduction of TAGs from starch, which was aimed atmarketing alternatives for low-value plant oils and fats,was not profitable. Although the raw materials for algaecan be sunlight and CO2 harvested from, for example,exhaust gases, the ultimate economic feasibility of algaland other microbial TAG production for biodiesel will
ultimately depend on biomass productivity, cellular lipidcontent, overall lipid productivity, and yield. For profit-ability, the relatively low-value biodiesel end-product willrequire a highly efficient conversion system using low-costsubstrates and with the potential to yield separable, val-ued coproducts. For the most efficient heterotrophic growthand lipid production, yeast- and bacterium-based fermen-tations of low-cost lignocellulosic sources of carbohydratesmay best fit these requirements. They are relatively highyield and rapid and are usually dedicated to the productionof lipids in the form of TAGs, unlike algal fermentations,which are relatively slow and difficult to scale up. Althoughmolds tend to have lower productivities and are sometimesdifficult to establish in submerged culture, they can pro-duce high levels of lipids that are typically mixtures ofTAGs with significant quantities of very high-value poly-unsaturated FAs such as GLA, ARA (an omega-6 FA), andDHA (an omega-3 FA) [24]. Whereas polyunsaturatedFAs produced by molds can fit a high-value, low-volumeproduction model, the TAGs derived from yeast, or ascoproducts from molds, could be pursued for biodiesel fuelto fit a low-value, high-volume production model. Thefeasibility of a biorefinery may require an optimal combi-nation of these high- and low-value products to suit marketdemand and the economics of the system.
Coproducts
Coproduct generation in a lignocellulosic SCO biorefineryis important to offset the processing cost of biofuels. Ligninand microbe cake after lipid processing are the prominentmaterials for coproduct generation (Figure 4). It is widelybelieved that lignin would be burned to supply energy forthe SCO biorefinery. However, there is more scope forlignin to be valorized into fuels and chemicals that cangenerate additional revenue for the biorefinery [69]. In thefirst-generation corn ethanol biorefinery, dried distillersgrains with solubles (DDGS) is a key coproduct that is usedas animal feed and provides a significant revenue stream tothe biorefinery [70]. A portion of the microbes is recycledafter fermentation, while the remaining microbes are soldas a coproduct for animal feed that could easily displace thetraditional soy meal or other oil cakes used for feedinganimals. In the case of oleaginous organisms, after proces-sing the lipids, the microbe cake could be sold as animalfeed or be further processed to produce amino acids orpeptides, which have wide applications (e.g., biomaterials,bioplastic, biofoam) [71]. These processing steps help addvalue to proteins that are extracted from microbes andgenerate more revenue for the biorefinery.
Technoeconomics
Economics is the key to a successful lignocellulosic SCObiorefinery. Currently, only a few technoeconomic studieshave been conducted on the use of heterotrophic oleaginousmicroorganisms for the production of SCO. Ratledge andCohen reported an expected price for the microbial oil ofUS$3000/ton (excluding the cost of feedstock), with calcu-lations based on previous work in New Zealand on theproduction of Cocoa Butter Equivalent from lactose usingCandida curvata [72]. Recently, Koutinas et al. studied theuse of R. toruloides for the production of microbial oil and
51

Lignocellulosic biomassSugars Oleaginous
microbe
Lignin
–Electricity/heat–Chemicals/fuels
–Low value lipids–High value lipids:Omega-6 fa�y acids: ALA, ARAOmega-3 fa�y acids: DHA, EPA
–Fuels–Chemicals–Materials,–Nutraceu�cals
Biomaterials(bioplas�c,
biofoam, etc.)
Animal feed
Amino acids,pep�des, etc.Microbe cake
$120,000
$/m
etric
ton $100,000
$80,000
$60,000
$40,000
$20,000
$0Vegetable
oilMicrobiallipids
GLA DHA ARA
TRENDS in Biotechnology
Figure 4. Products that could be generated in a lignocellulosic lipid biorefinery.
Review Trends in Biotechnology January 2015, Vol. 33, No. 1
biodiesel from glucose, assuming a lipid yield of 0.23 g/g,and obtained an estimated production cost of US$5500/tonoil and US$5900/ton biodiesel [73], which is not viable inthe current market, where vegetable oil prices are aroundUS$800–900/ton and biodiesel is at US$4.0/gallon(US$1220/ton) [73].
Coproduct microbe meal, which has a price of aroundUS$400–800/ton, was not considered in these technoeco-nomic studies. If considered, the economics of a SCObiorefinery would be slightly more favorable (�US$5000/ton oil), but would still not be viable. Improvement of lipidyield, productivity, and titer on lignocellulosic biomass iscritical in making a SCO biorefinery economically viable.Production of high-value FAs (e.g., GLA, DHA, ARA) andupgrading lignin and microbe meal to high-value productsare also essential in leveraging biorefinery economics(Figure 4). Furthermore, using simpler cultivation meth-ods, such as open ponds [74], could also significantly reducethe production cost. Converting oligosaccharides togetherwith monosaccharides derived from lignocellulosic bio-mass into lipids using oligosaccharide-consuming microbes[75] is also beneficial in improving lipid yield and reducingthe production cost.
Concluding remarks and future perspectivesAfter decades of development, second-generation biorefin-ery technologies are maturing. It is believed that variousproducts will be produced in a biorefinery and SCO will beone of the important platform products. A successful SCObiorefinery depends on making the right choices of feed-stock, pretreatment, oleaginous microorganism, processconfiguration, lipid-recovery method, and coproduct gen-eration. The current bottlenecks impeding economic pro-duction of SCO include low yield, low titer and lowproductivity of lipid, low tolerance to pretreatment degra-dation products, and cogeneration of valuable products.
52
Genetic modification and directed evolution of oleaginousmicroorganisms, further improvement of pretreatmenttechnologies, process design, and the development of tech-nologies for valuable coproduct generation are essential inaddressing the major bottlenecks.
References1 Sims, R.E. et al. (2010) An overview of second generation biofuel
technologies. Bioresour. Technol. 101, 1570–15802 Cherubini, F. and Ulgiati, S. (2010) Crop residues as raw materials for
biorefinery systems – a LCA case study. Appl. Energy 87, 47–573 Chundawat, S.P. et al. (2011) Deconstruction of lignocellulosic
biomass to fuels and chemicals. Annu. Rev. Chem. Biomol. Eng. 2,121–145
4 da Costa Sousa, L. et al. (2009) ‘Cradle-to-grave’assessment of existinglignocellulose pretreatment technologies. Curr. Opin. Biotechnol. 20,339–347
5 Zhang, Z. et al. (2012) Advancements and future directions in enzymetechnology for biomass conversion. Biotechnol. Adv. 30, 913–919
6 Balan, V. et al. (2013) Review of US and EU initiatives towarddevelopment, demonstration, and commercialization of lignocellulosicbiofuels. Biofuels Bioprod. Biorefin. 7, 732–759
7 Ratledge, C. and Wynn, J.P. (2002) The biochemistry and molecularbiology of lipid accumulation in oleaginous microorganisms. Adv.Appl. Microbiol. 51, 1–51
8 Knothe, G. (2010) Biodiesel and renewable diesel: a comparison. Prog.Energy Combust. Sci. 36, 364–373
9 Santacesaria, E. et al. (2012) Main technologies in biodiesel production:state of the art and future challenges. Catal. Today 195, 2–13
10 Tan, H. et al. (2013) Glycerol production and its applications as a rawmaterial: a review. Renew Sust. Energy Rev. 27, 118–127
11 Behr, A. et al. (2008) Improved utilisation of renewable resources: newimportant derivatives of glycerol. Green Chem. 10, 13–30
12 Corma, A. et al. (2007) Chemical routes for the transformation ofbiomass into chemicals. Chem. Rev. 107, 2411–2502
13 Biermann, U. et al. (2000) New syntheses with oils and fats asrenewable raw materials for the chemical industry. Angew. Chem.Int. Ed. 39, 2206–2224
14 Das, U.N. (2006) Essential fatty acids: biochemistry, physiology andpathology. Biotechnol. J. 1, 420–439
15 Leung, D.Y. et al. (2010) A review on biodiesel production usingcatalyzed transesterification. Appl. Energy 87, 1083–1095

Review Trends in Biotechnology January 2015, Vol. 33, No. 1
16 Maris, E.P. and Davis, R.J. (2007) Hydrogenolysis of glycerol overcarbon-supported Ru and Pt catalysts. J. Catal. 249, 328–337
17 Guerrero-Perez, M.O. and Banares, M.A. (2008) New reaction:conversion of glycerol into acrylonitrile. ChemSusChem 1, 511–513
18 Beltran-Prieto, J.C. et al. (2013) A review of catalytic systems forglycerol oxidation: alternatives for waste valorization. Aust. J.Chem. 66, 511–521
19 Carrettin, S. et al. (2004) Oxidation of glycerol using supported goldcatalysts. Top. Catal. 27, 131–136
20 Witsuthammakul, A. and Sooknoi, T. (2012) Direct conversion ofglycerol to acrylic acid via integrated dehydration–oxidation bedsystem. Appl. Catal. A Gen. 413, 109–116
21 Ratledge, C. (2011) A portrait in microbial oils. Int. News Fats OilsRelat. Mater. 22, 440–442
22 Subramaniam, R. et al. (2010) Microbial lipids from renewableresources: production and characterization. J. Ind. Microbiol.Biotechnol. 37, 1271–1287
23 Kosa, M. and Ragauskas, A.J. (2011) Lipids from heterotrophicmicrobes: advances in metabolism research. Trends Biotechnol. 29,53–61
24 Ratledge, C. (2004) Fatty acid biosynthesis in microorganisms beingused for single cell oil production. Biochimie 86, 807–815
25 Ageitos, J.M. et al. (2011) Oily yeasts as oleaginous cell factories. Appl.Microbiol. Biotechnol. 90, 1219–1227
26 Beopoulos, A. et al. (2011) An overview of lipid metabolism in yeastsand its impact on biotechnological processes. Appl. Microbiol.Biotechnol. 90, 1193–1206
27 Liang, M-H. and Jiang, J-G. (2013) Advancing oleaginousmicroorganisms to produce lipid via metabolic engineering technology.Prog. Lipid Res. 52, 395–408
28 Papanikolaou, S. and Aggelis, G. (2011) Lipids of oleaginous yeasts.Part I: biochemistry of single cell oil production. Eur. J. Lipid Sci.Technol. 113, 1031–1051
29 Li, Q. et al. (2008) Perspectives of microbial oils for biodieselproduction. Appl. Microbiol. Biotechnol. 80, 749–756
30 Weete, J.D. and Weber, D.J. (1980) Lipid Biochemistry of Fungi andOther Organisms, Plenum Press
31 Wu, S.G. et al. (2010) Phosphate-limitation mediated lipidproduction by Rhodosporidium toruloides. Bioresour. Technol. 101,6124–6129
32 Wu, S.G. et al. (2011) Microbial lipid production by Rhodosporidiumtoruloides under sulfate-limited conditions. Bioresour. Technol. 102,1803–1807
33 Yang, B. and Wyman, C.E. (2008) Pretreatment: the key to unlockinglow-cost cellulosic ethanol. Biofuels Bioprod. Biorefin. 2, 26–40
34 Balan, V. (2014) Current Challenges in Commercially ProducingBiofuels from Lignocellulosic Biomass. ISRN Biotechnol. 2014,1–31
35 Van Dyk, J.S. and Pletschke, B.I. (2012) A review of lignocellulosebioconversion using enzymatic hydrolysis and synergistic cooperationbetween enzymes – factors affecting enzymes, conversion and synergy.Biotechnol. Adv. 30, 1458–1480
36 Huang, C. et al. (2013) Single cell oil production from low-costsubstrates: the possibility and potential of its industrialization.Biotechnol. Adv. 31, 129–139
37 Jin, M. et al. (2012) Simultaneous saccharification and co-fermentation(SSCF) of AFEXTM pretreated corn stover for ethanol production usingcommercial enzymes and Saccharomyces cerevisiae 424A(LNH-ST).Bioresour. Technol. 110, 587–594
38 Lynd, L.R. (1996) Overview and evaluation of fuel ethanol fromcellulosic biomass: technology, economics, the environment, andpolicy. Annu. Rev. Energy Environ. 21, 403–465
39 Zhang, J. and Hu, B. (2012) Solid-state fermentation of Mortierellaisabellina for lipid production from soybean hull. Appl. Biochem.Biotechnol. 166, 1034–1046
40 Liang, Y.N. (2013) Producing liquid transportation fuels fromheterotrophic microalgae. Appl. Energy 104, 860–868
41 Li, P. et al. (2011) In situ biodiesel production from fast-growing andhigh oil content Chlorella pyrenoidosa in rice straw hydrolysate.Biomed. Res. Int. 2011, 141207
42 EL-Sheekh, M.M. et al. (2012) Mixotrophic and heterotrophic growth ofsome microalgae using extract of fungal-treated wheat bran. Int. J.Recycling Org. Waste Agric. 1, 1–9
43 Miazek, K. et al. (2014) Effect of lignocellulose related compounds onmicroalgae growth and product biosynthesis: a review. Energies 7,4446–4481
44 Xie, H.B. et al. (2012) Enzymatic hydrolysates of corn stover pretreatedby a N-methylpyrrolidone–ionic liquid solution for microbial lipidproduction. Green Chem. 14, 1202–1210
45 Tsigie, Y.A. et al. (2011) Lipid production from Yarrowia lipolyticaPo1 g grown in sugarcane bagasse hydrolysate. Bioresour. Technol.102, 9216–9222
46 Tsigie, Y.A. et al. (2012) Oil production from Yarrowia lipolytica Po1 gusing rice bran hydrolysate. J. Biomed. Biotechnol. 2012, 378384
47 Geddes, C.C. et al. (2011) Advances in ethanol production. Curr. Opin.Biotechnol. 22, 312–319
48 Zhao, X. et al. (2011) Lipid production by Rhodosporidium toruloidesY4 using different substrate feeding strategies. J. Ind. Microbiol.Biotechnol. 38, 627–632
49 Li, Y.H. et al. (2007) High-density cultivation of oleaginous yeastRhodosporidium toruloides Y4 in fed-batch culture. Enzyme Microb.Technol. 41, 312–317
50 Kristensen, J.B. et al. (2009) Yield-determining factors in high-solidsenzymatic hydrolysis of lignocellulose. Biotechnol. Biofuels 2, 11
51 Liu, W. et al. (2012) Simultaneous saccharification and microbial lipidfermentation of corn stover by oleaginous yeast Trichosporoncutaneum. Bioresour. Technol. 118, 13–18
52 Gong, Z.W. et al. (2013) Efficient conversion of biomass into lipids byusing the simultaneous saccharification and enhanced lipid productionprocess. Biotechnol. Biofuels 6, 36
53 Wei, H. et al. (2013) Genomic, proteomic, and biochemical analyses ofoleaginous Mucor circinelloides: evaluating its capability in utilizingcellulolytic substrates for lipid production. PLoS ONE 8, e71068
54 Lian, J.N. et al. (2012) Yeast fermentation of carboxylic acids obtainedfrom pyrolytic aqueous phases for lipid production. Bioresour. Technol.118, 177–186
55 Palmqvist, E. and Hahn-Hagerdal, B. (2000) Fermentation oflignocellulosic hydrolysates. II: inhibitors and mechanisms ofinhibition. Bioresour. Technol. 74, 25–33
56 Hu, C. et al. (2009) Effects of biomass hydrolysis by-products onoleaginous yeast Rhodosporidium toruloides. Bioresour. Technol.100, 4843–4847
57 Huang, C. et al. (2012) Effects of alcohol compounds on the growth andlipid accumulation of oleaginous yeast Trichosporon fermentans. PLoSONE 7, e46975
58 Huang, C. et al. (2011) Effects of aldehydes on the growth and lipidaccumulation of oleaginous yeast Trichosporon fermentans. J. Agric.Food Chem. 59, 4606–4613
59 Huang, C. et al. (2012) Effect of organic acids on the growth and lipidaccumulation of oleaginous yeast Trichosporon fermentans. Biotechnol.Biofuels 5, 4
60 Roche, C.M. et al. (2014) Engineering the filamentous fungusNeurospora crassa for lipid production from lignocellulosic biomass.Biotechnol. Bioeng. 111, 1097–1107
61 Ahmad, A.L. et al. (2014) Comparison of harvesting methods formicroalgae Chlorella sp. and its potential use as a biodieselfeedstock. Environ. Technol. 35, 2244–2253
62 Kim, J. et al. (2013) Methods of downstream processing for theproduction of biodiesel from microalgae. Biotechnol. Adv. 31, 862–876
63 Jin, G.J. et al. (2012) Enzyme-assisted extraction of lipids directly fromthe culture of the oleaginous yeast Rhodosporidium toruloides.Bioresour. Technol. 111, 378–382
64 de Boer, K. et al. (2012) Extraction and conversion pathways formicroalgae to biodiesel: a review focused on energy consumption. J.Appl. Phycol. 24, 1681–1698
65 Zhang, X. et al. (2014) Ultrasonication aided in situ transesterificationof microbial lipids to biodiesel. Bioresour. Technol. 169, 175–180
66 Alba, L.G. et al. (2013) Recycling nutrients in algae biorefinery.ChemSusChem 6, 1330–1333
67 Chakraborty, M. et al. (2012) Concomitant extraction of bio-oil andvalue added polysaccharides from Chlorella sorokiniana using aunique sequential hydrothermal extraction technology. Fuel 95, 63–70
68 Milledge, J.J. and Heaven, S. (2014) Methods of energy extraction frommicroalgal biomass: a review. Rev. Environ. Sci. Biotechnol. 13, 301–320
69 Zakzeski, J. et al. (2010) The catalytic valorization of lignin for theproduction of renewable chemicals. Chem. Rev. 110, 3552–3599
53

Review Trends in Biotechnology January 2015, Vol. 33, No. 1
70 Bothast, R. and Schlicher, M. (2005) Biotechnological processes forconversion of corn into ethanol. Appl. Microbiol. Biotechnol. 67, 19–25
71 Konst, P.M. et al. (2011) Stabilization and immobilization ofTrypanosoma brucei ornithine decarboxylase for the biobasedproduction of 1,4-diaminobutane. Green Chem. 13, 1167–1174
72 Ratledge, C. and Cohen, Z. (2008) Microbial and algal oils: do they havea future for biodiesel or as commodity oils? Lipid Technol. 20, 155–160
73 Koutinas, A.A. et al. (2014) Design and technoeconomic evaluation ofmicrobial oil production as a renewable resource for biodiesel andoleochemical production. Fuel 116, 566–577
74 Santamauro, F. et al. (2014) Low-cost lipid production by an oleaginousyeast cultured in non-sterile conditions using model waste resources.Biotechnol. Biofuels 7, 34
75 Gong, Z. et al. (2014) Conversion of biomass-derived oligosaccharidesinto lipids. Biotechnol. Biofuels 7, 1–11
76 Ruan, Z.H. et al. (2012) Evaluation of lipid accumulation fromlignocellulosic sugars by Mortierella isabellina for biodiesel production.Bioresour. Technol. 110, 198–205
77 Hu, C.M. et al. (2011) Simultaneous utilization of glucose and xylose forlipid production by Trichosporon cutaneum. Biotechnol. Biofuels 4, 25
78 Huang, X. et al. (2011) Biological removal of inhibitors leads to theimproved lipid production in the lipid fermentation of corn stoverhydrolysate by Trichosporon cutaneum. Bioresour. Technol. 102,9705–9709
79 Galafassi, S. et al. (2012) Lipid production for second generationbiodiesel by the oleaginous yeast Rhodotorula graminis. Bioresour.Technol. 111, 398–403
54
80 Ruan, Z.H. et al. (2013) Co-hydrolysis of lignocellulosic biomassfor microbial lipid accumulation. Biotechnol. Bioeng. 110, 1039–1049
81 Yu, X.C. et al. (2011) Oil production by oleaginous yeasts using thehydrolysate from pretreatment of wheat straw with dilute sulfuric acid.Bioresour. Technol. 102, 6134–6140
82 Zeng, J.J. et al. (2013) Lignocellulosic biomass as a carbohydrate sourcefor lipid production by Mortierella isabellina. Bioresour. Technol. 128,385–391
83 Huang, C. et al. (2013) Microbial oil production from corncob acidhydrolysate by oleaginous yeast Trichosporon coremiiforme. BiomassBioenerg. 49, 273–278
84 Huang, C. et al. (2009) Microbial oil production from rice strawhydrolysate by Trichosporon fermentans. Bioresour. Technol. 100,4535–4538
85 Huang, C. et al. (2012) Improving lipid production from bagassehydrolysate with Trichosporon fermentans by response surfacemethodology. N. Biotechnol. 29, 372–378
86 Huang, C. et al. (2012) Oil production by the yeast Trichosporondermatis cultured in enzymatic hydrolysates of corncobs. Bioresour.Technol. 110, 711–714
87 Kurosawa, K. et al. (2013) Engineering xylose metabolism intriacylglycerol-producing Rhodococcus opacus for lignocellulosic fuelproduction. Biotechnol. Biofuels 6, 134
88 Ruan, Z.H. et al. (2014) Oleaginous fungal lipid fermentation oncombined acid- and alkali-pretreated corn stover hydrolysate foradvanced biofuel production. Bioresour. Technol. 163, 12–17




















