
Preictal state identification by synchronization changes in long-term intracranial EEG recordings
10
Preictal state identification by synchronization changes in long-term intracranial EEG recordings Michel Le Van Quyen a, * , Jason Soss b , Vincent Navarro a,c , Richard Robertson d , Mario Chavez a , Michel Baulac c , Jacques Martinerie a a Laboratoire de Neurosciences Cognitives et Imagerie Ce ´re ´brale, LENA,CNRS UPR 640, Ho ˆpital de la Pitie ´-Salpe ˆtrie `re, 75651 Paris, France b Department of Neurology, David Geffen School of Medicine, University of California at Los Angeles, Los Angeles, CA, USA c Epileptology Unit, Ho ˆpital de la Pitie ´-Salpe ˆtrie `re, Paris, France d Mathematics Department, California State Polytechnic University, Pomona, CA, USA Accepted 7 October 2004 Available online 25 December 2004 Abstract Objective: There is accumulated evidence that mesial temporal lobe seizures are preceded by a preictal transition that evolves over minutes to hours. In the present study, we investigated these possible preictal changes in long-term intracranial recordings of five patients by a measure of phase synchronization. In order to clearly distinguish preictal changes from all the other interictal states, we developed an automatic extraction of representative patterns of interictal synchronization activity. This reference library was used to classify the successive synchronization patterns of long-term recordings into groups of similar patterns. Altered states of brain synchronization were identified as deviating from patterns in the reference library and were evaluated relative to the times of seizure onset in terms of sensitivity and specificity. Methods: A phase-locking measure was estimated using a sliding window analysis on 15 frequency bands (2 Hz steps between 0 and 30 Hz), for all pairs of EEG channels in the epileptogenic temporal lobe (14–20 channels), over the entire data sets (total analyzed duration 305 h). The preictal identification encompasses three basic stages: (1) a preprocessing stage involving the determination of a reference library of characteristic interictal synchronization patterns using a K-means algorithm, and the identification of discriminant variables differentiating interictal from preictal states, (2) a classification stage of the synchronization pattern via a minimum Mahalanobis distance to the reference patterns, as well as detection of outliers, (3) an evaluation stage of the sensitivity and specificity of the detection by receiver-operating characteristic curves. Results: In most of the cases (36 of 52 seizures, i.e. 70%), a specific state of brain synchronization can be observed several hours before the actual seizure. The changes involved both increases and decreases of the synchronization levels, occurring mostly within the 4–15 Hz frequency band, and were often localized near the primary epileptogenic zone. Conclusions: The analysis of phase synchronization offers a way to distinguish between a preictal state and normal interictal activity. These findings suggest that brain synchronizations are preictally altered in the epileptogenic temporal lobe, inducing a pathological state of higher susceptibility for seizure activity. Significance: Phase synchronization is capable of extracting information from the EEG that allow the definition of a preictal state. Although the proposed analysis does not constitute genuine seizure anticipation, these changes in neuronal synchronization may provide helpful information for prospective seizure warning. q 2004 International Federation of Clinical Neurophysiology. Published by Elsevier Ireland Ltd. All rights reserved. Keywords: Seizure anticipation; Intracranial EEG; Synchronization; Temporal lobe epilepsy 1. Introduction A fundamental issue in epilepsy research is the identification of patho-physiological changes predictive 1388-2457/$30.00 q 2004 International Federation of Clinical Neurophysiology. Published by Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.clinph.2004.10.014 Clinical Neurophysiology 116 (2005) 559–568 www.elsevier.com/locate/clinph * Corresponding author. Tel.: C33 142161171; fax: C33 145862537. E-mail address: [email protected] (M. Le Van Quyen).
Transcript of Preictal state identification by synchronization changes in long-term intracranial EEG recordings

Preictal state identification by synchronization changes in long-term
intracranial EEG recordings
Michel Le Van Quyena,*, Jason Sossb, Vincent Navarroa,c, Richard Robertsond,Mario Chaveza, Michel Baulacc, Jacques Martineriea
aLaboratoire de Neurosciences Cognitives et Imagerie Cerebrale, LENA,CNRS UPR 640, Hopital de la Pitie-Salpetriere, 75651 Paris, FrancebDepartment of Neurology, David Geffen School of Medicine, University of California at Los Angeles, Los Angeles, CA, USA
cEpileptology Unit, Hopital de la Pitie-Salpetriere, Paris, FrancedMathematics Department, California State Polytechnic University, Pomona, CA, USA
Accepted 7 October 2004
Available online 25 December 2004
Abstract
Objective: There is accumulated evidence that mesial temporal lobe seizures are preceded by a preictal transition that evolves over
minutes to hours. In the present study, we investigated these possible preictal changes in long-term intracranial recordings of five patients by
a measure of phase synchronization. In order to clearly distinguish preictal changes from all the other interictal states, we developed an
automatic extraction of representative patterns of interictal synchronization activity. This reference library was used to classify the
successive synchronization patterns of long-term recordings into groups of similar patterns. Altered states of brain synchronization were
identified as deviating from patterns in the reference library and were evaluated relative to the times of seizure onset in terms of sensitivity
and specificity.
Methods: A phase-locking measure was estimated using a sliding window analysis on 15 frequency bands (2 Hz steps between 0 and
30 Hz), for all pairs of EEG channels in the epileptogenic temporal lobe (14–20 channels), over the entire data sets (total analyzed duration
305 h). The preictal identification encompasses three basic stages: (1) a preprocessing stage involving the determination of a reference library
of characteristic interictal synchronization patterns using a K-means algorithm, and the identification of discriminant variables differentiating
interictal from preictal states, (2) a classification stage of the synchronization pattern via a minimum Mahalanobis distance to the reference
patterns, as well as detection of outliers, (3) an evaluation stage of the sensitivity and specificity of the detection by receiver-operating
characteristic curves.
Results: In most of the cases (36 of 52 seizures, i.e. 70%), a specific state of brain synchronization can be observed several hours before the
actual seizure. The changes involved both increases and decreases of the synchronization levels, occurring mostly within the 4–15 Hz
frequency band, and were often localized near the primary epileptogenic zone.
Conclusions: The analysis of phase synchronization offers a way to distinguish between a preictal state and normal interictal activity.
These findings suggest that brain synchronizations are preictally altered in the epileptogenic temporal lobe, inducing a pathological state of
higher susceptibility for seizure activity.
Significance: Phase synchronization is capable of extracting information from the EEG that allow the definition of a preictal state.
Although the proposed analysis does not constitute genuine seizure anticipation, these changes in neuronal synchronization may provide
helpful information for prospective seizure warning.
q 2004 International Federation of Clinical Neurophysiology. Published by Elsevier Ireland Ltd. All rights reserved.
Keywords: Seizure anticipation; Intracranial EEG; Synchronization; Temporal lobe epilepsy
1388-2457/$30.00 q 2004 International Federation of Clinical Neurophysiology.
doi:10.1016/j.clinph.2004.10.014
* Corresponding author. Tel.: C33 142161171; fax: C33 145862537.
E-mail address: [email protected] (M. Le Van Quyen).
1. Introduction
A fundamental issue in epilepsy research is the
identification of patho-physiological changes predictive
Clinical Neurophysiology 116 (2005) 559–568
www.elsevier.com/locate/clinph
Published by Elsevier Ireland Ltd. All rights reserved.

M. Le Van Quyen et al. / Clinical Neurophysiology 116 (2005) 559–568560
of an impending seizure. In recent years, new techniques for
seizure anticipation prior to visually detectable intracranial
electrical changes have been intensively investigated using
nonlinear analysis of EEG signals (Iasemedis et al., 1990;
Lehnertz and Elger, 1998; Le Van Quyen et al., 2001a,b;
Martinerie et al., 1998). These studies have shown that
information extracted from the EEG may allow the
definition of a preictal state and its distinction from the
interictal state. More recently, applications of a phase
synchronization measure have been shown to be useful in
gathering spatio-temporal information about the epilepto-
genic process (Le Van Quyen et al., 2001c; Mormann et al.,
2000). The method is based on direct estimation of the
instantaneous phase of the signal and, hence, is ideal for
analyzing nonstationary EEG recordings whose synchroni-
zation properties evolve over time (Lachaux et al., 1999).
Since transient phase-locking between different cortical
areas is ubiquitous during normal physiological conditions
such as sensory-motor processing or sleep (Varela et al.,
2001), assessment of phase synchronization in epilepsy is
also important to obtain a better insight into possible
intermittent dysfunction (Tass et al., 1998).
In this study, we examined the degree of phase
synchronization in long-term intracranial EEG recordings
of five patients suffering from mesial temporal lobe epilepsy
(see summary paper of the Bonn 2002 Workshop for patient
information and data descriptions). We have focused on
looking for quantifiable spatial or temporal shifts in preictal
synchronization far in advance of seizure onset detectable
on the EEG. Because normal EEG is enormously varied,
manifesting qualitative changes depending on behavioral
state, it is important to clearly distinguish preictal changes
from all the other interictal states. Therefore, its final
assessment should be carried out by a statistical approach
involving continuous recordings over long periods of time,
including, in particular, a systematic study of false positives.
In this paper, we propose a general patient-specific approach
for this purpose which involves three stages: (1) an initial
preprocessing stage to select a library of synchronization
patterns representing the normal interictal periods and to
improve the ability to distinguish between interictal and
preictal states, (2) a classification stage where shifts in
synchronization are detected, and (3) an assessment stage
evaluating the sensitivity and specificity of this approach for
preictal identification.
2. Methods
2.1. Synchronization analysis
The analysis of phase synchronization between neuronal
signals was introduced by Lachaux et al. (1999) and Tass
et al. (1998) to overcome some limitations of conventional
methods which not disentangle amplitudes and phases
(Bullock et al., 1995). The term ‘synchronization’ is used in
its strict sense, as a statistical measure of the degree to
which two signals are phase-locked during a short time
period. Recent studies have demonstrated the ability of this
measure to discriminate transient synchronization in
intracranial EEG data (Fell et al., 2001; Mormann et al.,
2000). Our analysis followed several steps: first, signals
from non-overlapping, consecutive 5 s periods were filtered
with a bandpass corresponding to a particular frequency
component. Second, the instantaneous phase of each filtered
window was extracted by means of the Hilbert transform.
Third, the degree of phase-locking between a pair of EEG
channels was quantified by the trial-average of the phase
differences on the unit circle in the complex plane
PLV Z k1
n
Xn
1
ei½f1ðtÞKf2ðtÞ�k
where n is the number of data points in each time window.
This phase-locking value (PLV) varies between 0 (inde-
pendent signals) and 1 (constant phase-lag between the two
signals). The PLVs were computed over consecutive signal
segments for P possible pairs of EEG channels in the
epileptogenic temporal lobe (from 14 to 20 channels, using
both medial and subdural temporal electrodes) and NZ15
frequency bands (2 Hz steps between 0 and 30 Hz). This
computation allows the characterization of the multi-
frequency synchronization patterns of each time window
t as a vector S(t) in a P!N dimensional space. For 20
channels, the dimension of this space is (20!19)/2!15Z2850. The phase-locking values were investigated over the
entire data sets (total analyzed duration 305 h, covering 52
seizures).
2.2. Detecting preictal changes: a general description
of the algorithm
Once the synchronization values are calculated, our
preictal state identification is based on a patient-specific
pattern recognition approach involving three stages:
Stage 1. A preprocessing stage is required in order to
reduce the number of variables used and to enhance the
ability to distinguish between interictal and preictal
features. This first implies extracting and selecting relevant
and appropriate features of the interictal state. For this
purpose, we chose long baseline controls (duration ranging
from 5 to 24 h), separated from any seizure by a few hours.
Because many seizures in the data sets were often close
together (less than 3 h apart), the baselines were typically
taken at the beginning of each data set. In theory, the values
of synchronization indices are dependent on the frequency
band, the channel pair and the time (Fig. 1A), which implies
a high degree of heterogeneity. Nevertheless, while the
number of possible synchronization patterns can be large,
we found that most of the interictal recordings can be
reasonably fit into 5–10 clusters. This indicates strong
recurrences in the pattern dynamics and may be related to
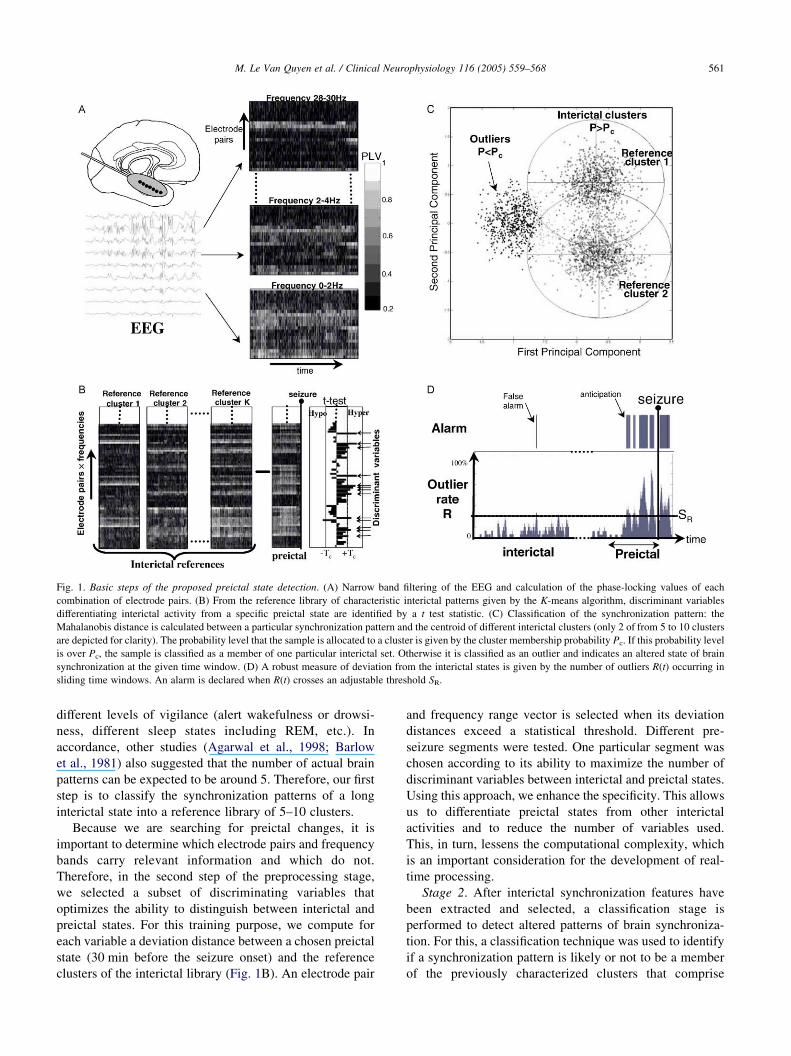
Fig. 1. Basic steps of the proposed preictal state detection. (A) Narrow band filtering of the EEG and calculation of the phase-locking values of each
combination of electrode pairs. (B) From the reference library of characteristic interictal patterns given by the K-means algorithm, discriminant variables
differentiating interictal activity from a specific preictal state are identified by a t test statistic. (C) Classification of the synchronization pattern: the
Mahalanobis distance is calculated between a particular synchronization pattern and the centroid of different interictal clusters (only 2 of from 5 to 10 clusters
are depicted for clarity). The probability level that the sample is allocated to a cluster is given by the cluster membership probability Pc. If this probability level
is over Pc, the sample is classified as a member of one particular interictal set. Otherwise it is classified as an outlier and indicates an altered state of brain
synchronization at the given time window. (D) A robust measure of deviation from the interictal states is given by the number of outliers R(t) occurring in
sliding time windows. An alarm is declared when R(t) crosses an adjustable threshold SR.
M. Le Van Quyen et al. / Clinical Neurophysiology 116 (2005) 559–568 561
different levels of vigilance (alert wakefulness or drowsi-
ness, different sleep states including REM, etc.). In
accordance, other studies (Agarwal et al., 1998; Barlow
et al., 1981) also suggested that the number of actual brain
patterns can be expected to be around 5. Therefore, our first
step is to classify the synchronization patterns of a long
interictal state into a reference library of 5–10 clusters.
Because we are searching for preictal changes, it is
important to determine which electrode pairs and frequency
bands carry relevant information and which do not.
Therefore, in the second step of the preprocessing stage,
we selected a subset of discriminating variables that
optimizes the ability to distinguish between interictal and
preictal states. For this training purpose, we compute for
each variable a deviation distance between a chosen preictal
state (30 min before the seizure onset) and the reference
clusters of the interictal library (Fig. 1B). An electrode pair
and frequency range vector is selected when its deviation
distances exceed a statistical threshold. Different pre-
seizure segments were tested. One particular segment was
chosen according to its ability to maximize the number of
discriminant variables between interictal and preictal states.
Using this approach, we enhance the specificity. This allows
us to differentiate preictal states from other interictal
activities and to reduce the number of variables used.
This, in turn, lessens the computational complexity, which
is an important consideration for the development of real-
time processing.
Stage 2. After interictal synchronization features have
been extracted and selected, a classification stage is
performed to detect altered patterns of brain synchroniza-
tion. For this, a classification technique was used to identify
if a synchronization pattern is likely or not to be a member
of the previously characterized clusters that comprise

M. Le Van Quyen et al. / Clinical Neurophysiology 116 (2005) 559–568562
the interictal library. For each sample, the probability level
of cluster membership Pc is calculated by the distance of the
corresponding synchronization pattern to all the interictal
reference clusters. If this probability level is over Pc, the
sample is classified as a member of one interictal reference
set. Otherwise it is classified as an outlier, indicating an
altered state of brain synchronization, deviating from all
interictal reference patterns. As an example, Fig. 1C
illustrates, in the space of the first two principal com-
ponents, cases where samples lie on the fringes of the
clusters or outside the Pc confidence intervals. In order to
obtain a robust measure of deviation from the interictal
states, we computed the number of outliers R(t) occurring in
sliding windows of 5 min duration, as a function of time t.
For detection purposes, an alarm was declared when R(t)
crossed an adjustable threshold SR (Fig. 1D).
Stage 3. Finally, we evaluated the threshold parameters
and the performance of the corresponding alarm sequences
relative to the times of seizure onset in terms of sensitivity
and specificity. For this validation, the accuracy of the
parameters was assessed by generating an ROC curve (Litt
et al., 2001), and plotting sensitivity versus specificity for a
range of adjustable parameters, here the probability levels
for cluster membership Pc and threshold values SR. On the
basis of the ROC curve, we carried out an in-sample
optimization of these two parameters, maximizing both
sensitivity and specificity for each patient.
2.3. Preprocessing stage
2.3.1. Selection of a library of interictal synchronization
patterns
To identify different synchronization patterns in the
interictal state, a K-means cluster analysis (Anderberg,
1973) was applied to the multi-frequency vectors S(t) of an
interictal state (duration ranging from 5 to 24 h), far
removed from any seizures (separated from them by
3–10 h, depending on the data set). The K-means algorithm
requires the fixing of the number K of clusters that are
expected in the data. Our own experimental evidence
suggests that most of the interictal recordings can be
reasonably fit into 5–10 clusters. After initialization of the
number of clusters, the K-means algorithm assigns each
vector to one of the K clusters, such that the within-cluster
sum of distances between member points and the centroid is
minimized (see mathematical Appendix A.1).
2.3.2. Identification of discriminant variables best
differentiating interictal from preictal periods
A deviation distance was computed to identify the
variables most capable of distinguishing one chosen preictal
state (30 min before the seizure onset) and the K reference
clusters of the interictal library. For this purpose, we
considered the t index between the PLVs of each electrode
pair and each frequency range
T Z ðmp KmiÞ=
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffis2
p=np Cs2i =ni
q
where mp and mi are the respective means of the preictal and
the interictal PLVs, with corresponding standard deviations
sp and si and sample sizes np and ni. A synchrony change is
considered to be statistically significant at the PZ0.01 level
(via the Student’s t test) when the absolute deviation
distance jTj is above the critical value jTcjZ2.5. An
electrode pair and frequency range is selected when its
deviation distances from the K reference clusters satisfy
either criterion: min(T1,.,TK)OTc (hyper-synchronization)
or max(T1,.,TK)!KTc (hypo-synchronization). Chosen in
this way, the selected electrode pairs and frequency ranges
define a set of discriminant vectors U(t) of S(t).
2.4. Classification of synchronization patterns and detection
of outliers
The goal of this step is to classify each discriminant
vector U(t) as a member of one of the previously
characterized interictal clusters or as ‘outlier’ when it
deviates from all these reference patterns, indicating an
altered state of brain synchronization. The synchronization
vectors U(t), containing m discriminant frequency and
electrode pair combinations, are classified by using a
minimum Mahalanobis distance criterion (Mahalanobis,
1936, see mathematical Appendix A.2). Let mi be the
multidimensional vector containing the average values of
the discriminating variables of cluster i; then the squared
Mahalanobis distance D2 between a vector U(t) and mi is
defined as
D2 Z ðUðtÞKmiÞT CK1
i ðUðtÞKmiÞ
where Ci is the covariance matrix of cluster i. The
distance of a sample from each of the synchronization
clusters can be calculated in this way. The sample is
allocated to the closest cluster of the interictal library or,
otherwise, is defined to be an ‘outlier’ when it lies outside
of all the interictal clusters (Chernoff, 1970). Visual
inspection of feature histograms indicated that the features
used here were approximately Gaussian in distribution. In
this case, probability levels for cluster membership are
determined from the Chi-square distribution, calculated
from the incomplete gamma function with m degrees of
freedom (Johnson and Wichern, 1988). This probability
level Pc can be used to classify samples based on the
simple rule: samples having a probability level over Pc are
classified as members of the training set, whereas samples
having a probability level under Pc are classified as
outliers.
2.5. Evaluation of the sensitivity and specificity
Deviations from baseline controls are coarsely estimated
by the outliers’ rate R(t), representing the number of outliers
M. Le Van Quyen et al. / Clinical Neurophysiology 116 (2005) 559–568 563
within a short time window [tKs,t]. We chose sZ5 min in
this study. Note that the value of each function is attributed
to the end of the window, so we do not depend on
information from the future. The emergence of an abnormal
synchronization pattern is recognized by the condition:
RðtÞRSR
We consider this condition as an ‘alarm’. The evaluation
of the anticipation performance makes sense only if the
success and error scores are insensitive to the variation of
adjustable parameters, here Pc and SR. The receiver-
operating characteristic (ROC) curve shows the relative
scores of successes and errors for different choices of
adjustable parameters (Litt et al., 2001). The vertical axis
of this graph represents the sensitivity of anticipation,
defined as the rate of confirmed anticipation on all the pre-
seizure recordings (duration 30 min). This is plotted
against the specificity of the anticipation, defined as the
rate of false alarm over 50 segments (duration 30 min) of
randomly selected interictal data. Different points corre-
spond to different probability levels Pc used to detect
outliers, with Pc varying from 10K1 at the highest point to
10K7 at the lowest point.
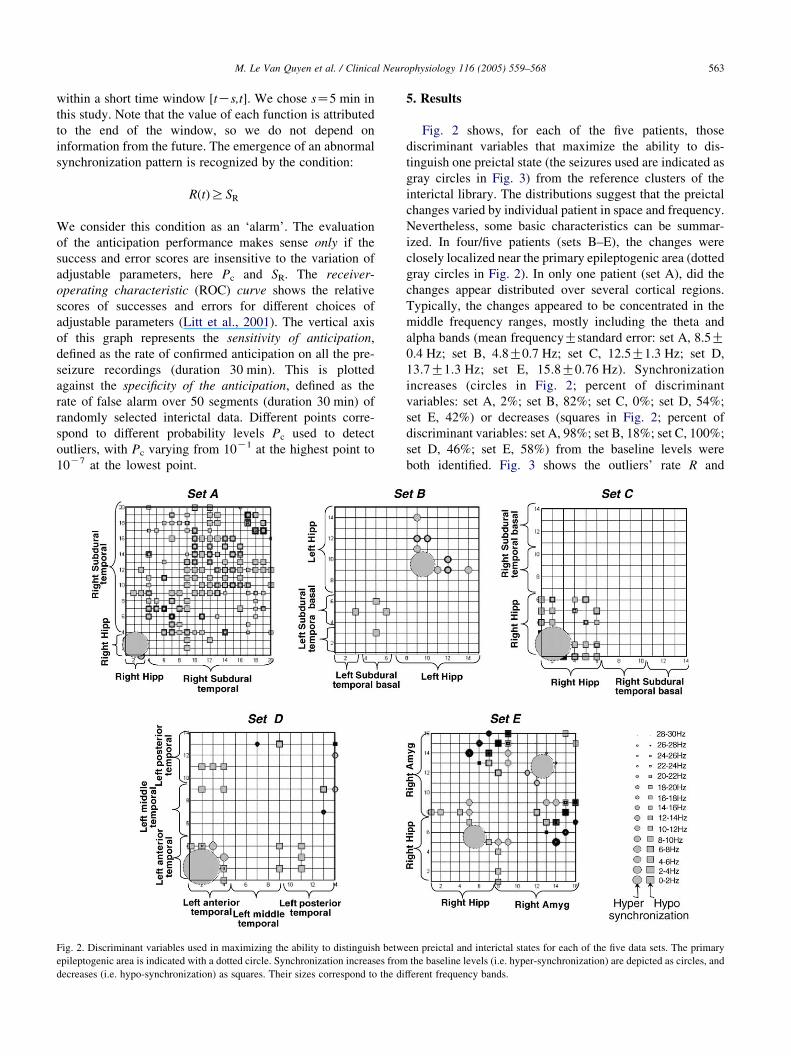
Fig. 2. Discriminant variables used in maximizing the ability to distinguish betw
epileptogenic area is indicated with a dotted circle. Synchronization increases from
decreases (i.e. hypo-synchronization) as squares. Their sizes correspond to the di
5. Results
Fig. 2 shows, for each of the five patients, those
discriminant variables that maximize the ability to dis-
tinguish one preictal state (the seizures used are indicated as
gray circles in Fig. 3) from the reference clusters of the
interictal library. The distributions suggest that the preictal
changes varied by individual patient in space and frequency.
Nevertheless, some basic characteristics can be summar-
ized. In four/five patients (sets B–E), the changes were
closely localized near the primary epileptogenic area (dotted
gray circles in Fig. 2). In only one patient (set A), did the
changes appear distributed over several cortical regions.
Typically, the changes appeared to be concentrated in the
middle frequency ranges, mostly including the theta and
alpha bands (mean frequencyGstandard error: set A, 8.5G0.4 Hz; set B, 4.8G0.7 Hz; set C, 12.5G1.3 Hz; set D,
13.7G1.3 Hz; set E, 15.8G0.76 Hz). Synchronization
increases (circles in Fig. 2; percent of discriminant
variables: set A, 2%; set B, 82%; set C, 0%; set D, 54%;
set E, 42%) or decreases (squares in Fig. 2; percent of
discriminant variables: set A, 98%; set B, 18%; set C, 100%;
set D, 46%; set E, 58%) from the baseline levels were
both identified. Fig. 3 shows the outliers’ rate R and
een preictal and interictal states for each of the five data sets. The primary
the baseline levels (i.e. hyper-synchronization) are depicted as circles, and
fferent frequency bands.

Fig. 3. Outliers’ rate for each data set and the corresponding alarm sequences for each of the five data sets. The seizures used in the determination of the
discriminant variables (see Fig. 2) are indicated as gray circles. The threshold value used, SR, is indicated as a doted line. Note that the R curve crossed the alarm
threshold several hours before the actual seizures and that, during the reference interictal state, there were only a few spurious detections.
M. Le Van Quyen et al. / Clinical Neurophysiology 116 (2005) 559–568564
the corresponding alarm sequences for each patient. In the
majority of the analyzed seizures, distinct differences
between interictal and preictal states were revealed (36/52
of all seizures, i.e. 70%). For example, in data set B, where
the longest interictal state was chosen for clustering
purposes (24 h), we see that R crossed the alarm threshold
several hours before the actual seizures and stayed at high
levels until the seizure onset. After the seizure, R often
dropped to low levels and increased again later. Note that,
during the reference interictal state, there were only a few
spurious detections, suggesting that the qualitative changes
in brain dynamics (like the wakefulness-sleep transitions)
did not induce false alarms. To better demonstrate this,
Fig. 4A shows the ROC curve for data set B, plotting
sensitivity versus specificity for different probability levels
Pc and threshold values SR. Choosing the parameters at the
‘corner’ of the ROC curve (PcZ2!10K3, SRZ10%
defining the parameters in Fig. 3), data epochs were
correctly classified as either preictal or interictal with a
sensitivity of 84% and specificity of 95%. It can be seen that
the performance is relatively insensitive to the variation of
adjustable parameters. For the other patients, using the
optimal parameters to obtain near 100% specificity, the
sensitivity of the analysis is: set A, 67%; set C, 66%; set D,
67%; set E, 86%. In order to estimate the time duration of
the preictal trends, we defined the preictal duration as the
first time at which the R curve crossed the SR level before a
particular seizure and after the preceding one. In this
estimation, we excluded the 10 min of post-ictal activity
from the preceding seizure. Mean time from declaration of
impending seizure to seizure EEG onset was 187G56 min
(set A, 23G12 min; set B, 274G66 min; set C, 117G45 min; set D, 253G148 min; set E, 164G46 min; Fig. 4B).
It is important to stress that these times are strongly
dependent on the criteria we used and should be regarded
as a weak estimate that allows both intra- and inter-
comparisons. Further improvements have to involve a
constraint on the length of time that R must remain at or
above the threshold (Le Van Quyen et al., 2001). The effects
of these stronger criteria on sensitivity and specificity for

Fig. 4. (A) Receiver-operating characteristic (ROC) curve for data set B, plotting sensitivity versus specificity for different probability levels Pc and threshold
values SR. The specificity of the anticipation is defined as the mean rate of (false) alarms occurring during randomly selected interictal segments. Choosing the
parameters PcZ2!10K3, SRZ10% at the ‘corner’ of the ROC curves, data epochs were classified as either preictal or interictal with a sensitivity of 84% and
specificity of 95%. (B) Preictal duration in hours, defined as the first time at which the R curve crossed the SR level before a particular seizure, excluding 10 min
of post-ictal activity from the preceding seizure. The black (resp. gray) bars indicate the preictal duration determined using no a priori (resp. some a priori)
knowledge of future behavior.
M. Le Van Quyen et al. / Clinical Neurophysiology 116 (2005) 559–568 565
individual or group optimization of the algorithm are
currently being investigated, but are beyond the scope of
this paper. Furthermore, for a rigorous definition of seizure
anticipation, the system’s output at time t must be solely a
function of the information available to that system at or
before time t (i.e. no use of future information from any time
later than t is allowed). In order to assess the anticipation
performance of our analysis, Fig. 4B shows using black bars
the preictal duration determined with no a priori knowledge
of future behavior. These results suggest that our approach
can be used for warning purposes.
Finally, we evaluated possible correlations between
changes in synchronization and visual inspection of EEG
recordings. First, visual inspection of the EEG recordings
sometimes showed the presence of undetected sub-clinical
electrographic seizures. For the data set D, in addition to the
six seizures with both EEG and clinical manifestations, we
observed 12 electrographic seizures between the first and
the last electro-clinical seizures. This can be related to the
fact that R crossed the alarm threshold in a sustained way
after the third electro-clinical seizure (Fig. 3). Second,
changes of the interictal epileptic activities were also
observed. For the data sets B and D, the seizures were
systematically preceded by changes of the spiking
discharges in the epileptogenic focus, consisting of spikes
and waves of long duration and high amplitude, becoming
rhythmic around 0.5–1.0 Hz (Fig. 5E). These changes
occurred 10–15 s before the seizures, but were also
observed temporally far from the seizures. For the data set
B, this activity did not show any clear correlation with the
time course of R (Fig. 5B). The mesial temporal focus also
presented another pattern of spiking discharges, more
frequent and more irregular, consisting of spikes, poly-
spikes, or spikes and waves (Fig. 5D). For the data set B, we
observed that this latter activity neither preceded the seizure
occurrence, nor was correlated with the time course of R
(Fig. 5B). Finally, we observed a third pattern of spiking
discharges (Fig. 5F) that consisted of spikes diffusing
locally (to the whole hippocampus) and regionally (to the
adjacent temporal neocortex). This activity did not precede
the seizure occurrence, but seemed to occur during periods
where R was below the alarm threshold (Fig. 5B).
6. Discussion
This study of five MTLE patients demonstrates that the
analysis of phase synchronization might offer a way to
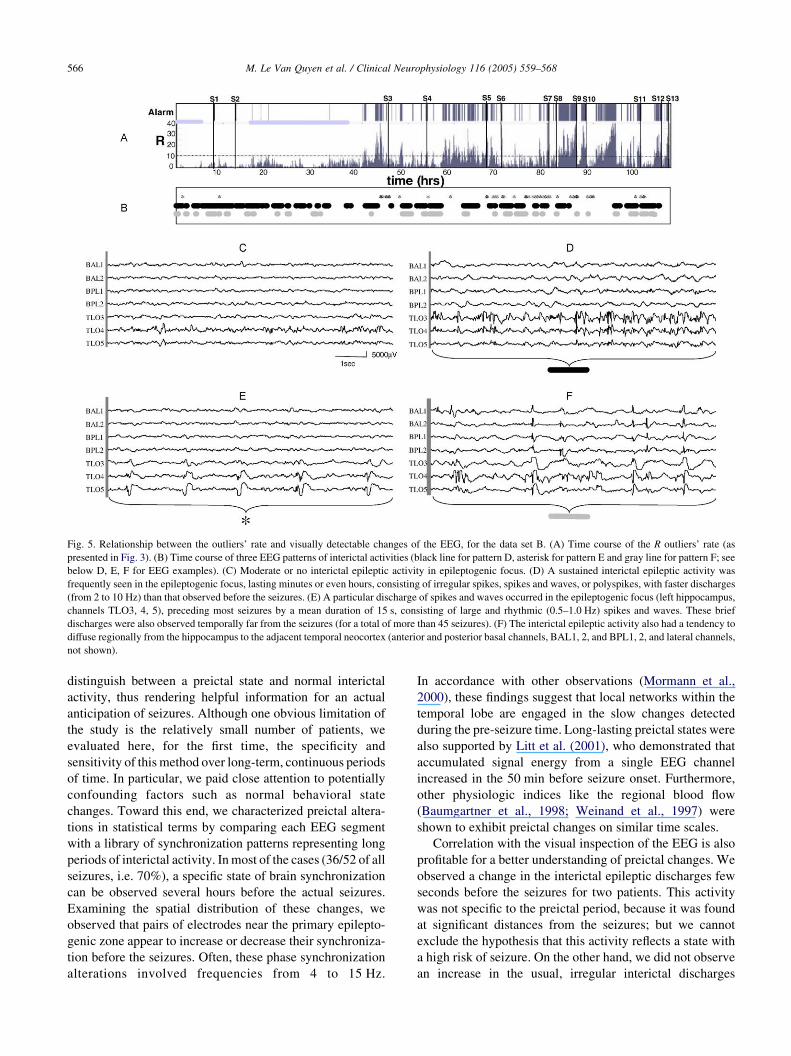
Fig. 5. Relationship between the outliers’ rate and visually detectable changes of the EEG, for the data set B. (A) Time course of the R outliers’ rate (as
presented in Fig. 3). (B) Time course of three EEG patterns of interictal activities (black line for pattern D, asterisk for pattern E and gray line for pattern F; see
below D, E, F for EEG examples). (C) Moderate or no interictal epileptic activity in epileptogenic focus. (D) A sustained interictal epileptic activity was
frequently seen in the epileptogenic focus, lasting minutes or even hours, consisting of irregular spikes, spikes and waves, or polyspikes, with faster discharges
(from 2 to 10 Hz) than that observed before the seizures. (E) A particular discharge of spikes and waves occurred in the epileptogenic focus (left hippocampus,
channels TLO3, 4, 5), preceding most seizures by a mean duration of 15 s, consisting of large and rhythmic (0.5–1.0 Hz) spikes and waves. These brief
discharges were also observed temporally far from the seizures (for a total of more than 45 seizures). (F) The interictal epileptic activity also had a tendency to
diffuse regionally from the hippocampus to the adjacent temporal neocortex (anterior and posterior basal channels, BAL1, 2, and BPL1, 2, and lateral channels,
not shown).
M. Le Van Quyen et al. / Clinical Neurophysiology 116 (2005) 559–568566
distinguish between a preictal state and normal interictal
activity, thus rendering helpful information for an actual
anticipation of seizures. Although one obvious limitation of
the study is the relatively small number of patients, we
evaluated here, for the first time, the specificity and
sensitivity of this method over long-term, continuous periods
of time. In particular, we paid close attention to potentially
confounding factors such as normal behavioral state
changes. Toward this end, we characterized preictal altera-
tions in statistical terms by comparing each EEG segment
with a library of synchronization patterns representing long
periods of interictal activity. In most of the cases (36/52 of all
seizures, i.e. 70%), a specific state of brain synchronization
can be observed several hours before the actual seizures.
Examining the spatial distribution of these changes, we
observed that pairs of electrodes near the primary epilepto-
genic zone appear to increase or decrease their synchroniza-
tion before the seizures. Often, these phase synchronization
alterations involved frequencies from 4 to 15 Hz.
In accordance with other observations (Mormann et al.,
2000), these findings suggest that local networks within the
temporal lobe are engaged in the slow changes detected
during the pre-seizure time. Long-lasting preictal states were
also supported by Litt et al. (2001), who demonstrated that
accumulated signal energy from a single EEG channel
increased in the 50 min before seizure onset. Furthermore,
other physiologic indices like the regional blood flow
(Baumgartner et al., 1998; Weinand et al., 1997) were
shown to exhibit preictal changes on similar time scales.
Correlation with the visual inspection of the EEG is also
profitable for a better understanding of preictal changes. We
observed a change in the interictal epileptic discharges few
seconds before the seizures for two patients. This activity
was not specific to the preictal period, because it was found
at significant distances from the seizures; but we cannot
exclude the hypothesis that this activity reflects a state with
a high risk of seizure. On the other hand, we did not observe
an increase in the usual, irregular interictal discharges

M. Le Van Quyen et al. / Clinical Neurophysiology 116 (2005) 559–568 567
before seizures. Such changes have been previously
reported (Litt et al., 2001), but also excluded by others
(Gotman and Marciani, 1985). Finally, we observed another
pattern of diffusing spikes that preferentially appeared when
the R index was low, suggesting that this activity may use a
network of synchronies that is not favorable to seizure
emergence.
It is important to point out that our proposed synchro-
nization analysis does not constitute genuine seizure
anticipation, because this would require a prospective
analysis of data when time to seizure is unknown. Never-
theless, in the present state, our analysis could, after more
extensive validation, be used for warning purposes. In
particular, the architecture of our algorithm is such that it
may be readily adapted to further improve its performance
in real-time conditions. Further improvements can be
obtained by selecting (on- or offline) multiple pre-seizure
segments for training purposes or by adding continuously
new synchronization clusters in the interictal library. For
example, if the method produces undesired detections of a
certain type of activity, a representative sample of this
activity can be included in the interictal library to prevent
similar undesired detections in the future. Furthermore, the
interictal library can be adapted to the changing clinical
status of the patient by including changes in medication or
post-ictal abnormalities. Also, expert-selected signal infor-
mation can also be included, depending on the type of signal
characteristics or changes the physician/user wants to detect
(or not detect).
There are numerous indications that the spatial and
temporal structures of synchronization patterns are dis-
tributed in space and transient in time (Bullock et al., 1995;
Fell et al., 2001). It was hypothesized that this transient
phase-locking between different parts of the brain could be a
basic mechanism for the functional large-scale integration
observed during cognition (Varela et al., 2001). Thus, given
this central role of synchrony in normal states, our results
suggest that the participation of particular networks in
normal physiological synchronizations appears to be
preictally altered, inducing a state of higher susceptibility
for seizure activity. This is also in accordance with
experimental models of epilepsy showing that specific
networks around the epileptic focus seem to be of crucial
importance in determining whether or not a seizure is likely
to occur and spread (Wyler et al., 1973). Our present
observations suggest that two related processes seem to be
involved here: (1) A state of increased synchronization. This
state may reflect recruitment phenomena within the primary
epileptogenic area and its surroundings regions. (2) A state
of decreased synchronization. This state may isolate
pathologically discharging neuronal populations of the
epileptic focus from the influence of activity in wider
brain areas, thus facilitating the development of local
pathological recruitments. On the other hand, a loss of
synchrony might also provide an ‘idle’ population of
neurons which may be more easily recruited into the
epileptic process (Mormann et al., 2000). Finally, the
preictal loss of synchrony could reflect a depression of
synaptic inhibition in areas surrounding the epileptogenic
focus, as in certain experimental models of epilepsy
(Matsumo and Ajmone Marsan, 1964).
Further investigations of the spatial characteristics of
phase synchronization or studies at a cellular level (Staba
et al., 2002) are necessary to validate these hypotheses.
Whatever the pathophysiologic basis of preictal changes,
the observed duration is sufficient to offer new possibilities
for therapeutic intervention. Such measures might include
preventive strategies to modify the synchronization around
the critical regions by direct electrical stimulation of brain
structures (Fanselow et al., 2000) or by cognitive therapies
(Birbaumer et al., 1990).
Appendix A. Mathematical appendix
A.1. K-means clustering
1.0 The goal of clustering is to reduce the amount of data
by categorizing or grouping similar data items together
Partitional clustering attempts to directly decompose the set
of objects, perhaps p-dimensional ‘feature’ vectors, into a
set of disjoint subsets called ‘clusters’. A commonly used
partitional clustering method is called K-means clustering,
which produces K different clusters that are intended to be
as distinct as possible (Anderberg, 1973).
1.1 A computer program for automating the K-means
algorithm starts with K random clusters and then moves
objects between those clusters with the dual goals of:
(1) minimizing variability within clusters, and (2)
maximizing variability between clusters. Each point
from the feature space is added to the closest cluster.
The ‘closeness’ to the cluster is determined by calculating
the distance between a point and the centroid of that
cluster. Then each point is visited again to recalculate the
distance to the updated cluster. If the closest cluster is not
the one to which it currently belongs, the point will
switch to the new cluster. When switching occurs,
centroids of both modified clusters have to be recalcu-
lated. This procedure is repeated until no more switching
takes place.
1.2 The measure of this variability within and between
clusters is based on a metric or distance function in the space
of p-dimensional feature vectors. One common choice for
such a distance function is the so-called Mahalanobis
metric.
A.2. The Mahalanobis distance
2.0 The Mahalanobis distance D is based on the notion of
‘standardized distance’ d. Let x(i) be the value of feature i of
vector x, with m(i,j) the mean of feature i for cluster j, and
s(i,j) the corresponding standard deviation.

M. Le Van Quyen et al. / Clinical Neurophysiology 116 (2005) 559–568568
Then
d2ðx;mjÞ Zxð1ÞKmð1; jÞ
sð1; jÞ
� �2
Cxð2ÞKmð2; jÞ
sð2; jÞ
� �2
C/
CxðpÞKmðp; jÞ
sðp; jÞ
� �2
;
where mj is the vector of feature means for cluster j (or, its
‘centroid’).
2.1 This metric has the important property that it is ‘scale
invariant’: if distances are measured in this way, the units
used for various features have no effect on the resulting
distances, and thus no effect on the final classification. The
Mahalanobis distance is a generalization of this standar-
dized distance.
2.2 The covariance of two features measures their
tendency to vary together, defined to be the average of the
products of the deviations of the feature values from their
means. More precisely, if {x(1,r),x(2,r),.,x(n,r)} and
{x(1,s),x(2,s),., x(n,s)} are the sets of features r and s for
the n points in a cluster, respectively (i.e. x(j,r) and x(j,s) are
the corresponding features of the jth point), then cðr; sÞZð1=nK1Þ
PnjZ1 ½xðj; rÞKmðrÞ�½xðj; sÞKmðsÞ�f g, with m(r) the
cluster mean of feature r and m(s) the mean of feature s.
2.3 The matrix of all these covariances CZ(c(r,s)), the
covariance matrix, provides a way to measure distance that
is invariant under linear transformations of the space of
feature vectors.
2.4 The matrix generalization of the scalar expression for
squared standardized distance, d2, from feature vector x to
cluster j’s mean vector (or centroid), mj, turns out to be just
D2ðx;mjÞZ ðxKmjÞtrCK1
j ðxKmjÞ, where Cj is the cluster’s
covariance matrix; D is called the Mahalanobis distance.
2.5 Let m1,m2,.,mK be the mean vectors for the K
clusters, with C1,C2,.,CK the corresponding covariance
matrices. A feature vector x is classified by calculating the
Mahalanobis distance D from x to each of the mj, and then
assigning x to the class for which D(x,mj) is a minimum.
2.6 The use of the Mahalanobis distance removes several
of the limitations of the standard Euclidean metric, such as:
(1) it automatically accounts for the scaling of the
coordinate axes, and (2) it corrects for correlations between
different features.
References
Agarwal R, Gotman J, Flanagan D, Rosenblatt B. Automatic EEG analysis
during long-term monitoring in the ICU. Electroencephalogr Clin
Neurophysiol 1998;107:44–58.
Anderberg MR. Cluster analysis for applications. New York: Academic
Press; 1973.
Barlow JS, Creutzfeldt OD, Michael D, Houchin J, Epelbaum H. Automatic
adaptive segmentation of EEGs. Electroencephalpogr Clin Neurophy-
siol 1981;51:512–25.
Baumgartner C, Serles W, Leutmezer F, Pataraia E, Aull S, Czech T,
Pietrzyk U, Relic A, Podreka I. Pre-ictal SPECT in temporal lobe
epilepsy: regional cerebral blood flow is increased prior to electro-
encephalography-seizure onset. J Nucl Med 1998;39:978–82.
Birbaumer N, Elbert T, Canavan A, Rockstroh B. Slow potentials of the
cerebral cortex and behavior. Physiol Rev 1990;70:1–41.
Bullock TH, McClune MC, Achimowicz JZ, Iragui-Madoz VJ, Duckrow RB,
Spencer SS. EEG coherence has structure in the millimeter domain:
subdural and hippocampal recordings from epileptic patients. Electro-
encephalogr Clin Neurophysiol 1995;95:161–77.
Chernoff H. Metric considerations in cluster analysis. Technical report 67.
Society of Statistics, Standford University; 1970.
Fanselow E, Reid A, Nicolelis M. Reduction of pentylenetrazole-induced
seizure activity in awake rats by seizure-trigeminal nerve stimulation.
J Neurosci 2000;20:8160–8.
Fell J, Klaver P, Lehnertz K, Grunwald T, Schaller C, Elger CE,
Fernandez G. Human memory formation is accompanied by rhinal–
hippocampal coupling and decoupling. Nat Neurosci 2001;4:1259–64.
Gotman J, Marciani MG. Electroencephalographic spiking activity, drug
levels, and seizure occurrence in epileptic patients. Ann Neurol 1985;
17:597–603.
Iasemedis LD, Sackellares JC, Zaveri HP, Williams WJ. Phase space
topography and the Lyapunov exponent of electrocorticograms in
partial seizures. Brain Topogr 1990;2:187–201.
Johnson RA, Wichern DW. Applied multivariate statistical analysis.
Englewood Cliffs, NJ: Prentice Hall; 1988. p. 134–5.
Lachaux JP, Rodriguez E, Martinerie J, Varela FJ. Measuring phase-
synchrony in brain signal. Hum Brain Mapp 1999;8:194–208.
Lehnertz K, Elger CE. Can epileptic seizures be predicted? Evidence from
nonlinear time series analysis of brain electrical activity Phys Rev Lett
1998;80:5019–22.
Le Van Quyen M, Martinerie J, Navarro V, Boon P, D’Have M, Adam C,
Renault B, Varela FJ, Baulac M. Anticipation of epileptic seizures from
standard EEG recordings. Lancet 2001a;357:183–8.
Le Van Quyen M, Martinerie J, Navarro V, Baulac M, Varela FJ.
Characterizing neurodynamic changes before seizures. J Clin Neuro-
physiol 2001b;18:191–208.
Le Van Quyen M, Fouchez J, Lachaux JP, Rodriguez E, Lutz A,
Martinerie J, Varela F. Comparison of Hilbert transform and wavelet
methods for the analysis of neuronal synchrony. J Neurosci Methods
2001c;111:83–98.
Litt B, Esteller R, Echauz J, D’Alessandro M, Shor R, Henry T, Pennel P,
Epstein C, Bakay R, Dichter M, Vachtsevanos G. Epileptic seizures
may begin hours in advance of clinical onset: a report of five patients.
Neuron 2001;30:51–64.
Mahalanobis PC. Proc Natl Inst Sci India 1936;2:49.
Martinerie J, Adam C, Le Van Quyen M, Baulac M, Clemenceau S,
Renault B, Varela FJ. Epileptic seizures can be anticipated by non-
linear analysis. Nat Med 1998;4:1173–6.
Matsumo H, Ajmone Marsan C. Cortical cellular phenomena in experimental
epilepsy: ictal manifestations. Exp Neurol 1964;9:305–26.
Mormann F, Lehnertz K, David P, Elger CE. Mean phase coherence as a
measure for phase synchronization and its application to the EEG of
epilepsy patients. Physica D 2000;144:358–69.
Staba RJ, Wilson Cl, Bragin A, Fried I, Engel J. Sleep states differentiate
single neuron activity recorded from human epileptic hippocampus,
entorhinal cortec and subiculum. J Neurosci 2002;22:5694–704.
Tass P, Rosenblum MG, Weule J, Kurths J, Pikovsky A, Volkmann J,
Schnitzler A, Freund HJ. Detection of n:m phase locking from noisy data:
application to magnetoencephalography. Phys Rev Lett 1998;81:3291–4.
Varela FJ, Lachaux JP, Rodriguez E, Martinerie J. The brainweb: phase
synchronization and large-scale integration. Nat Rev Neurosci 2001;2:
229–39.
Weinand ME, Carter LP, El-Saadany WF, Sioutos PJ, Labiner DM,
Oommen KJ. Cerebral blood flow and temporal lobe epileptogenicity.
J Neurosurg 1997;86:226–32.
Wyler AR, Fetz EE, Ward AA. Spontaneous firing patterns of epileptic
neurons in the monkey motor cortex. Exp Neurol 1973;40:567–85.




















