
Preclinical activity profile of α-lactoalbumin, a whey protein rich in tryptophan, in rodent models...
11
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Preclinical activity profile of α-lactoalbumin, a whey protein rich in tryptophan, in rodent models...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Epilepsy Research (2011) 95, 60—69
journa l homepage: www.e lsev ier .com/ locate /ep i lepsyres
Preclinical activity profile of �-lactoalbumin, a wheyprotein rich in tryptophan, in rodent models ofseizures and epilepsy
Rita Citraroa, Francesca Scicchitanoa, Salvatore De Fazioa,Riccardo Raggiob, Paolo Mainardi c, Emilio Peruccad,Giovambattista De Sarroa,∗, Emilio Russoa
a Chair of Pharmacology, Department of Experimental and Clinical Medicine, School of Medicine, University ‘‘Magna Graecia’’ ofCatanzaro, Via T. Campanella, 115, Catanzaro 88100, Italyb Department of Pharmaceutical Science, University of Genova, Italyc Department of Neurosciences, Ophthalmology and Genetics, University of Genova, Italyd Clinical Pharmacology Unit, University of Pavia, and Clinical Trial Center, Institute of Neurology IRCCS C. Mondino Foundation,Pavia, Italy
Received 17 December 2010; received in revised form 18 February 2011; accepted 27 February 2011Available online 1 April 2011
KEYWORDSSerotonin;ALAC;Audiogenic seizures;Pilocarpine;GEPR
SummaryPurpose: To evaluate the potential anticonvulsant activity of �-lactalbumin (ALAC), a wheyprotein rich in tryptophan (TRP) relative to other large neutral aminoacids (LNAAs), in rodentmodels of seizures and epilepsy.Methods: The effects of ALAC administered per os were evaluated by standard protocolsagainst audiogenic seizures in Genetic Epilepsy Prone Rats (GEPR-9 rats), maximal electroshock(MES)-induced seizures in rats, pilocarpine-induced seizures in mice, spontaneous chronicseizures in mice exposed to pilocarpine-induced status epilepticus (SE), and absence seizures inWAG/Rij rats. In some models, carbamazepine (CBZ) was included as an active control. PlasmaTRP/LNAAs ratios were measured by GC—MS.Results: Single doses of ALAC up to 500 or 6000 mg/kg were devoid of anticonvulsant activityin all models tested. Conversely, 5- and 12-day treatment with ALAC (250—1000 mg/kg/day) inGEPR rats reduced dose-dependently seizure scores and prolonged latency to clonus onset,with full persistence of the effect for up to 12 h. ALAC (125—500 mg/kg/day for 15 days)protected against seizures induced by 250 mg/kg pilocarpine, but was less effective against
∗ Corresponding author. Tel.: +39 0961 712323; fax: +39 0961 774424.E-mail address: [email protected] (G. De Sarro).
0920-1211/$ — see front matter © 2011 Elsevier B.V. All rights reserved.doi:10.1016/j.eplepsyres.2011.02.013

Author's personal copy
alpha-Lactoalbumin protects against seizures 61
higher pilocarpine doses. Similarly to CBZ, ALAC (125—500 mg/kg/day for 15 days) was also effec-tive against spontaneous seizures in the post-pilocarpine SE model. ALAC (up to 6000 mg/kg/dayfor 12 days) did not prevent MES-induced seizures, although it reduced the duration of tonicextension at doses between 250 and 1000 mg/kg/day. Absence seizures in WAG/Rij rats were notsignificantly affected by ALAC. Plasma TRP/LNAAS ratios increased 2- to 3-fold after dosing withALAC (250 mg/kg/day) for 7 and 14 days, respectively.Conclusions: ALAC exerts significant protective activity against seizures in animal models, theeffect being especially prominent against audiogenic seizures in GEPR-9 rats, seizures induced bylow-dose pilocarpine in mice, and spontaneous seizures in mice exposed to pilocarpine-inducedSE. This action is likely to be mediated by increased availability of TRP in the brain, with aconsequent increase in 5-HT mediated transmission.© 2011 Elsevier B.V. All rights reserved.
Introduction
Serotonin (5-HT) is involved in several physiological func-tions in the central nervous system (CNS) such as controlof appetite, sleep, memory and learning, temperature reg-ulation, mood, behaviour, maturation of neuronal and glialcells, synaptic connections and neuronal excitability (Barnesand Sharp, 1999; Bagdy et al., 2007). As early as 1957,Bonnycastle et al. (1957) suggested that there may alsobe a link between 5-HT levels and epilepsy. Experimentaldata indicate that 5-HT has anticonvulsant properties bothin experimental models of epilepsy and in humans (Bagdyet al., 2007). Likewise, a reduction in brain 5-HT concen-trations leads to an increase in seizure susceptibility inanimal models of epilepsy (Browning et al., 1997; Statnicket al., 1996) and, possibly, in humans (Maynert et al.,1975; Pallister, 1959). Selective 5-HT reuptake inhibitors(SSRIs), initially believed to be proconvulsant, have beensubsequently reported to possess anticonvulsant proper-ties (Favale et al., 1995, 2003; Albano et al., 2006a). Ingeneral, agents that elevate synaptic 5-HT levels, such as5-hydroxytryptophan (5-HTP) and SSRIs, inhibit both focal(limbic) and generalized seizures (Yan et al., 1994a,b;Albano et al., 2006a). Based on this evidence, SSRIs weretested in the 1990s as potential anticonvulsants as add-onto antiepileptic drugs (AEDs) in patients with drug resis-tant epilepsy with promising results (Favale et al., 1995,2003; Cupello et al., 2005; Albano et al., 2006a). Many AEDsincluding valproic acid, lamotrigine, carbamazepine, pheny-toin and zonisamide elevate brain 5-HT levels as part oftheir actions (Okada et al., 1992; Dailey et al., 1996; Ahmadet al., 2005). Dailey et al. (1997) measured the effects ofAEDs on brain 5-HT levels using microdialysis in geneticallyepilepsy prone rats (GEPRs) and they found that the anticon-vulsant effects of AEDs correlated well with the increase inextracellular 5-HT concentrations.
In agreement with the role of 5-HT in epilepsy, the lev-els of tryptophan (TRP), an essential aminoacid and theonly precursor of 5-HT in the brain, have been found to bereduced in the cerebrospinal fluid and/or plasma of somegroups of patients with seizure disorders (Ko et al., 1993;Marion et al., 1985). Indeed, clinical and neurochemical evi-dence suggests that a decreased brain availability of TRPmay play a pathogenetic role in epilepsy (Russo et al., 2009).A PET study confirmed that a decrease in brain uptake of TRPis paralleled by a decrease in brain synthesis of 5-HT (Diksicet al., 2002), which, in turn, may lead to diminished 5-HT-
mediated anticonvulsant activity. Conversely, an increase inplasma TRP levels results in increased brain 5-HT synthesis.Furthermore, intravenously (i.v.) or intraperitoneally (i.p.)administered 5-hydroxy-TRP have been found to have anti-convulsant effects (Truscott, 1975; Alexander and Kopeloff,1976), while oral TRP has not been found to be consis-tently effective. The lack of anticonvulsant effects of orallyadministered TRP could be related to inadequate intestinalabsorption (Frenhani and Burini, 1999; Heuther et al., 1992).
Because TRP competes with other large neutral aminoacids (LNAAs) for the transport by a specific carrier atthe blood brain barrier (BBB), the cerebral uptake of TRPis enhanced by an increase in TRP/LNAAs ratio in plasma(Diksic et al., 2002). An elevation in plasma TRP/LNAA ratiocan be obtained by administration of whey proteins rich inTRP and poor in other LNAAs content (Feurté et al., 2001;Markus et al., 2000). In particular, �-lactoalbumin (ALAC,Davisco Foods International, Inc., Minneapolis, MN, USA), awhey protein naturally occurring in human milk and com-mercially available as a food supplement, shows the highestTRP/LNAAs ratio among all quantitatively relevant food-derived proteins. Oral doses of ALAC have been found toincrease significantly plasma TRP/LNAAs ratio in rodents andin humans (Yokogoshi and Wurtman, 1986; Markus et al.,2000; Feurté et al., 2001). In addition, ALAC has been foundto increase brain 5-HT concentrations (Orosco et al., 2004)and to reduce sleep disturbances in rats (Minet-Ringuetet al., 2004). In a pilot study, ALAC administration was alsoreported to be associated with improved seizure control inpatients with drug resistant epilepsy (Albano et al., 2006b;Mainardi et al., 2008). Based on these data, we considered itof interest to investigate its activity profile in various animalmodels of seizures and epilepsy.
Materials and methods
Animals
For experiments in mice, we used male C57BL/6 mice weighing22—28 g (8 weeks old, Harlan Italy SRL, Correzzana, Milano, Italy),housed in groups of 8—10 per cage under stable conditions ofhumidity (60 ± 5%) and temperature (21 ± 2 ◦C). The animals weremaintained on a 12 h light and 12 h dark cycle (lights on at 7:00 pm)and were allowed free access to standard laboratory chow (Teklad2018) and tap water until the time of experiments.
For experiments in rats, GEPRs of both sexes, 12—18 weeks ofage, were obtained from our breeding stock (Department of Phar-

Author's personal copy
62 R. Citraro et al.
macology, University of Catanzaro, Italy), which is derived from acolony originally supplied by Prof. Brian S. Meldrum (University ofLondon, UK). Male WAG/Rij and Wistar rats, 4-week old, with abody weight ∼60 g, were purchased from Charles River Laborato-ries Italia s.r.l. (Calco, Lecco, Italy). Rats were housed three/fourper cage under the same conditions mentioned above for mice.
All experimental protocols were approved by the Ethics Com-mittee of the University of Catanzaro. All procedures were incompliance with the National Institutes of Health Guide for Careand Use of Laboratory Animals (publication no. 85-23, revised 1985)and European Communities Council Directive of 24 November 1986(86/609 EEC).
Audiogenic seizures in GEPR rats
GEPRs aged between 6 and 8 weeks were exposed to sound stimu-lation three times at weekly intervals, and only animals showing aseizure (score 9; see below) on all three occasions were used for thepresent experiments (De Sarro et al., 1999). Seizure response wasassessed by two independent observers, according to a standardprocedure which has been previously described (De Sarro et al.,1999; Gareri et al., 2004). Sound-evoked responses were coded byusing the following scale: 0 = no response; 1 = running phase only;2 = two running phases followed by a clonic convulsion (clonus offore and hind limbs, head, pinnae, vibrissae and tail); 3 = one run-ning phase followed by a clonic convulsion; 4 = two running phasesfollowed by tonus of neck, trunk, fore- and hind-limb clonus; 5 = onerunning phase followed by tonus of neck, trunk, fore- and hind-limb clonus; 6 = two running phases followed by tonic extension ofwhole body except hind feet; 7 = one running phase followed bytonic extension of whole body except hind feet; 8 = two runningphases and complete tonic extension; 9 = one running phase andcomplete tonic extension. The sound stimulation was applied for60 s, but it was interrupted earlier if the animal developed a tonicextensor seizure. The maximum response was recorded for eachanimal.
Test compounds or vehicle (controls) were given per os (p.o.)by gastric gavage to groups of at least 10 rats per dose and time oftesting. For the evaluation of the effect of a single dose of ALAC,animals were placed under a hemispheric Perspex dome (58 cm indiameter) and challenged with a 12—16 kHz sinusoidal tone at 109 dBat different times after administration. For evaluation of subchronictreatment with ALAC, rats were tested after treatment p.o. for 5 or12 days. Audiogenic stimulation was delivered as described above,60 min after the last administration. Animals were observed forbehavioural changes during the period between administration ofeach dose and the auditory stimulation. Control animals were testedon the same day as the ALAC-treated animals.
Maximal electroshock seizure (MES) test
Treatments were applied in Wistar rats by using the same proto-col described above for GEPR rats, after single dose administrationand after 12 days treatment. Tonic seizures were induced by elec-troshock via ear electrodes, as described by De Sarro et al. (2003).The stimulus was applied for 0.2 s at a frequency of 60 Hz and 150 mAamplitude. The incidence and duration of tonic (hind limb) seizureswere measured.
Pilocarpine-induced seizures
Adult male C57BL/6 mice were divided into 3 groups, which wereassigned to receive ALAC, carbamazepine (CBZ) or vehicle, respec-tively. Each group was further divided into subgroups of 30 animals,which were assigned to each of the dose levels tested (ALAC 125, 250and 500 mg/kg/day, CBZ 20, 40 and 80 mg/kg) or vehicle. ALAC wasadministered p.o. for 15 consecutive days every morning between
9.00 and 11.00 am, up to 1 h before pilocarpine administration. CBZor vehicle were administered p.o. as a single dose 60 min beforetesting.
Each subgroup of mice received an i.p. injection of pilocarpinehydrochloride at doses of 250, 270 or 300 mg/kg. Methyl scopo-lamine nitrate (Sigma—Aldrich; 1 mg/kg i.p.) was injected 30 minbefore pilocarpine to antagonize the peripheral cholinergic effects.Seizures were scored by using the following scale: 1 = staring withmouth clonus; 2 = automatisms (WDS, scratching); 3 = unilateralforelimb clonus; 4 = bilateral forelimb clonus; 5 = bilateral forelimbclonus with rearing and falling; 6 = tonic—clonic seizure (Velískováand Moshé, 2006). A threshold convulsion was considered a seizureof at least stage 4 with a minimal duration of 10 s.
Chronic seizures after pilocarpine-induced statusepilepticus (SE)
After pilocarpine-induced SE, mice develop chronic spontaneousseizures (Löscher, 2002). A pilocarpine dose of 270 mg/kg was cho-sen to induce SE because it was found to have the best ratio betweennumber of animals undergoing SE and number of animals subject toearly mortality. The injection of pilocarpine was always precededby methyl scopolamine nitrate (1 mg/kg i.p.).
After the pilocarpine injection, mice were observed forthe development of SE, defined behaviourally as continuoustonic/clonic convulsions. The time of SE onset was defined as thetime of a stage 4—5 seizure which did not terminate in the sub-sequent 3 min. The evolution of spontaneous seizures follows thebehavioural and electrographic stages of kindling (Racine, 1972).Once a generalized convulsion similar to stage 5 kindled seizuresoccurs, the vast majority of the seizures that follow are also gen-eralized.
After 60 min from SE onset, animals were administered 10 mg/kgdiazepam i.p. to decrease seizure severity and increase survival.The mice were housed in clear Plexiglas cages for at least 6 weeks.Animals that survived for 6 weeks after SE were divided into groups(at least 15 animals per group), each of which was assigned toreceive one of the treatments to be tested. These included ALAC(125, 250 or 500 mg/kg/day) and CBZ (20 mg/kg/day), each admin-istered for ∼15 consecutive days in the drinking water beforerecordings. Before administration, each ALAC dose was dissolvedin 100 mL of tap water. For CBZ, 10 mg of the drug was dissolvedin a 3—4 mL chloroform, to which 50 mL of a solution containing10 mg Pluronic® PE 6800 (BASF) in water were added; the final solu-tion was then placed on a magnetic stirrer at a temperature of55 ± 1 ◦C to evaporate the chloroform for at least 24 h. The finalvolume was then adjusted to 50 mL by adding water. Doses werecalculated on the basis of the knowledge that mice drink on aver-age 10 mL/100 g/day (van Zutphen et al., 2001). Water bottles werewrapped in silver foil to prevent exposure to light and solutions werefreshly prepared and replaced twice a week.
Animals were anesthetized with chloral hydrate (40 mg/kg i.p.,Carlo Erba, Milan, Italy), and placed in a Kopf stereotaxic instru-ment. The skull was exposed after a midline incision to the skinoverlying the scalp, and five cortical electrodes for EEG recordingwere implanted. Stainless-steel screw electrodes were implantedon the dura mater over the cortex, two in the frontal region and twoin the parietal region according to the atlas coordinates of Paxinosand Franklin (2001). The ground electrode was placed over the cere-bellum. Animals were allowed to recover for at least 1 week prior torecordings. Every recording session lasted for 6 h. Epileptic animalsdevelop typical electrographic seizures with high voltage fast activ-ity involving both the hippocampus and cortex, occurring as burstsof activity preceding seizures and followed by continuous trains ofhigh-voltage spikes and polyspikes. EEG signals were amplified andconditioned by analog filters (filtering: below 1 Hz and above 30 Hzat 6 dB/octave) and subjected to analog-to-digital conversion with

Author's personal copy
alpha-Lactoalbumin protects against seizures 63
a sampling rate of 300 Hz. Quantification of seizures was based onthe number and duration of spikes and polyspikes, summarized forthe 6 h of EEG recording, as previously described (De Sarro et al.,1992; Leite and Cavalheiro, 1995).
Experiments in WAG/Rij rats
The effects of ALAC (250 and 500 mg/kg p.o.) after a single doseand after 15 consecutive days of treatment were assessed in maleWAG/Rij rats (6 months of age, n = 6 per each dose or vehicle), amodel of generalized absence seizures. For the 15-day treatment,ALAC was administered every morning between 9.00 and 11.00 AM.
Animals were chronically implanted with electrodes for EEGrecordings (Russo et al., 2004), under chloral hydrate anaesthesia(400 mg/kg i.p. Carlo Erba, Milan, Italy). Stainless-steel screw elec-trodes were implanted on the dura mater over the cortex, two inthe frontal region (AP = 2; L = ±2.5) and two in the parietal region(AP = −6; L = ± 2.5) according to the atlas coordinates of Paxinos andWatson (1998). The ground electrode was placed over the cerebel-lum. Control animals were implanted in the same day as treatedanimals. All animals were allowed at least 1 week of recovery andhandled twice a day. To habituate the animals to the recording con-ditions, the rats were connected to the recording cables for at least3 days before the experiments. A multichannel amplifier (StellateHarmonie Electroencephalograph; Montreal, Quebec, Canada) con-nected to a flexible recording cable and an electric swivel, fixedabove the cages, were used, which allowed animals to move freely.
Each recording session lasted for 3 h and started 1 h after the lastadministration of ALAC or vehicle. All EEG signals were amplifiedand conditioned by analog filters (filtering: below 1 Hz and above30 Hz at 6 dB/octave) and subjected to analog-to-digital conver-sion with a sampling rate of 300 Hz. The quantification of absenceseizures was based on the number and duration of EEG spike-wavedischarges (SWDs) (Russo et al., 2010). Briefly, the number andduration of SWDs for each rat were summarized in 30 min inter-vals (epochs) for 3 h of recording, and scored by visual inspectionof the recordings. All recordings were analyzed by two indepen-dent investigators, one of whom was blind to the treatment. A SWDwas considered as an EEG background deflection characterized bya 7—10 Hz frequency and an amplitude at least double that of thebackground, with a minimal duration of 1 s.
Assay of TRP and LNAAs in plasma
For the determination of the TRP/LNAA ratio in plasma, bloodsamples were collected from GEPRs rats treated with ALAC(250 mg/kg/day p.o.) for 7 or 14 consecutive days, respectively.Blood samples were centrifuged for 5 min at 5000 rpm, the plasmawas separated and deproteinization was performed by adding 5 mLof acetone to 1 mL plasma. After centrifugation at 3000 rpm for10 min, the organic phase was transferred to tubes and evaporatedat 80 ◦C.
The quantitative analysis of aminoacids was performedby GC—MS after derivatization with ethyl chloroformate(Sigma—Aldrich, Milan, Italy) as described by Wang et al.(1994). This derivatization procedure is fast and ensures goodsensitivity and resolution. Several stock solutions of the selectedaminoacids (Sigma, St. Louis, MO, USA) in 0.1 M HCl, each at aconcentration of 0.5 mg/mL, were prepared. Derivatization reac-tions were performed by adding 10 �L of the aminoacid mixtureto a hydroalcoholic solution made with 30 �L of ethanol, 60 �Lof distilled water and 10 �L of pyridine as protons scavenger. Anexcess of 10 �L of ethyl chloroformate was added and the reactionmixture was gently vortex-mixed for 5 min and left to cool. Themixture was then extracted with 200 �L of chloroform. Aftercentrifugation, the upper layer was discarded and the extract wasanalyzed by gas chromatography—mass spectrometry (GC—MS).
GC—MS determinations were carried out on a HewlettPackard 6890 GC/5973 MSD system with a capillary column HP-5(30 m × 0.25 mm, 0.25 �m film thickness). A helium carrier gas at1 mL/min constant flow was used with a temperature program setat 100—290 ◦C at a rate of 8 ◦C min−1. The ion source was set at250 ◦C with the ionisation voltage at 70 eV. Injector temperature washeld at 250 ◦C. 1 �L of chloroform extract was injected by meansof direct (splitless) injection. A MS SIM scan on selected ions wasperformed for quantitative analysis. Norvaline was generally addedas internal standard.
Statistical analysis
The maximum behavioural response was recorded for each ani-mal. Seizure severity scores and latencies were compared betweengroups using a Kruskall—Wallis nonparametric analysis of variance(ANOVA) followed by a Mann—Whitney U-test. All tests used weretwo-sided and a P value < 0.05 was considered significant. ED50 val-ues (±95% confidence limits) for each compound were estimated bythe method of Litchfield and Wilcoxon (1949). Statistical compari-son of mice groups with chronic seizures was carried out by one-wayANOVA followed by a Bonferroni’s post hoc test.
For experiments in WAG/Rij rats, EEG recordings were subdi-vided into 30 min epochs, and the duration and number of SWDswere treated separately for every epoch. These values were aver-aged and data obtained were expressed as mean ± SEM for eachdose group. Treated animals were compared by one-way ANOVA withtreatment as the only variable, followed by a Bonferroni’s post hoctest.
Results
Effects of ALAC in GEPR-9 rats
To evaluate the influence of ALAC on audiogenic seizures,subgroups of rats were used with a full seizure responseculminating in a complete tonic extension of fore limbs(GEPR-9), as identified in the three screening tests carriedout previously.
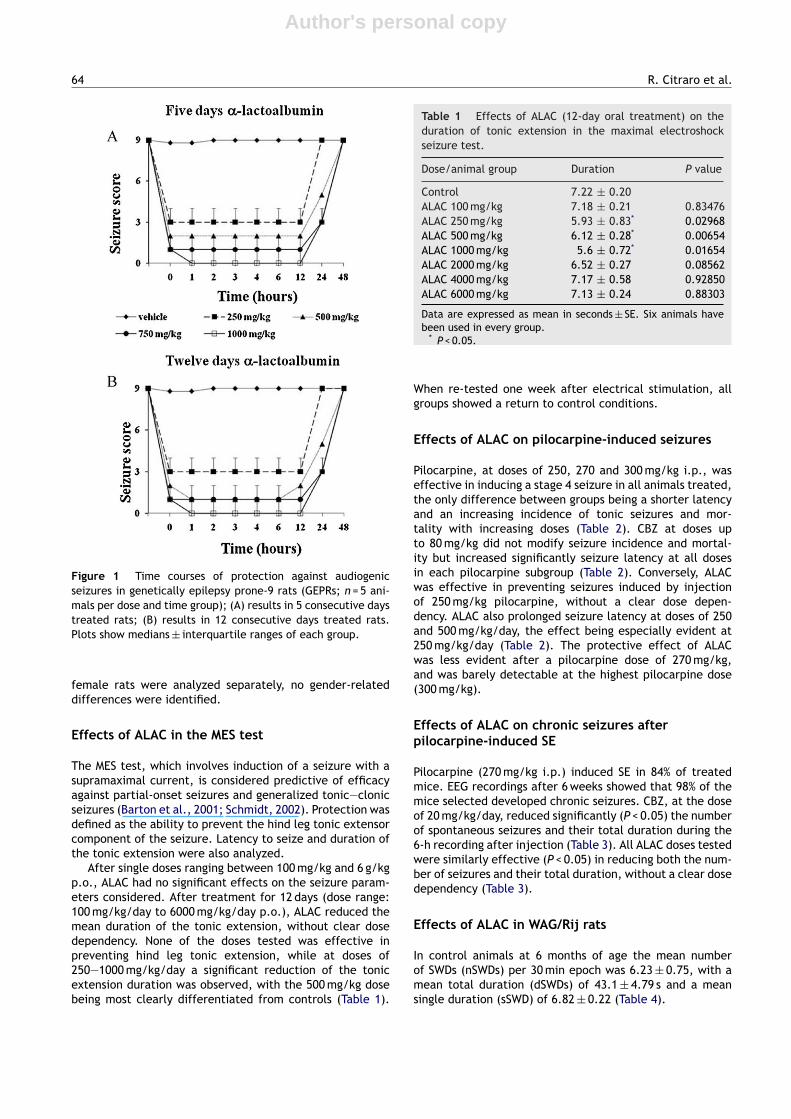
A single dose of ALAC (250, 500, 750 and 1000 mg/kg p.o.)did not induce any significant changes in the seizure scoresassessed at 1, 2, 3, 4, 6 and 12 h after administration (N = 5animals per dose and time group). By contrast, repeatedadministration of ALAC (250, 500, 750 and 1000 mg/kg/dayp.o.) for 5 consecutive days produced a dose-dependentand significant (P < 0.01) reduction in seizure score sever-ity (Fig. 1; N = 5 animals per dose and time group). Theeffect was already evident at the first audiogenic stimula-tion 1 h after the last ALAC dose, persisted for 12 h, andwas clearly reduced after 24 and 48 h. When ALAC treat-ment was continued for 12 days, the reduction in seizurescore severity was slightly more marked, but the differencescompared with the 5-day treatment were not statisti-cally significant (Fig. 1; N = 5 animals per dose and timegroup). A group of animals for each dose was also tested1 week after the last ALAC dose, and complete recoveryof the seizure response was found in all groups (data notshown).
The time of onset of clonus was increased significantly byALAC treatment for 5 or 12 days (P < 0.003 vs. vehicle treatedanimals, ANOVA). Multiple-dose treatment with ALAC did notaffect the latency time from stimulus onset to the initiationof wild running (data not shown). When data from male and
Author's personal copy
64 R. Citraro et al.
Figure 1 Time courses of protection against audiogenicseizures in genetically epilepsy prone-9 rats (GEPRs; n = 5 ani-mals per dose and time group); (A) results in 5 consecutive daystreated rats; (B) results in 12 consecutive days treated rats.Plots show medians ± interquartile ranges of each group.
female rats were analyzed separately, no gender-relateddifferences were identified.
Effects of ALAC in the MES test
The MES test, which involves induction of a seizure with asupramaximal current, is considered predictive of efficacyagainst partial-onset seizures and generalized tonic—clonicseizures (Barton et al., 2001; Schmidt, 2002). Protection wasdefined as the ability to prevent the hind leg tonic extensorcomponent of the seizure. Latency to seize and duration ofthe tonic extension were also analyzed.
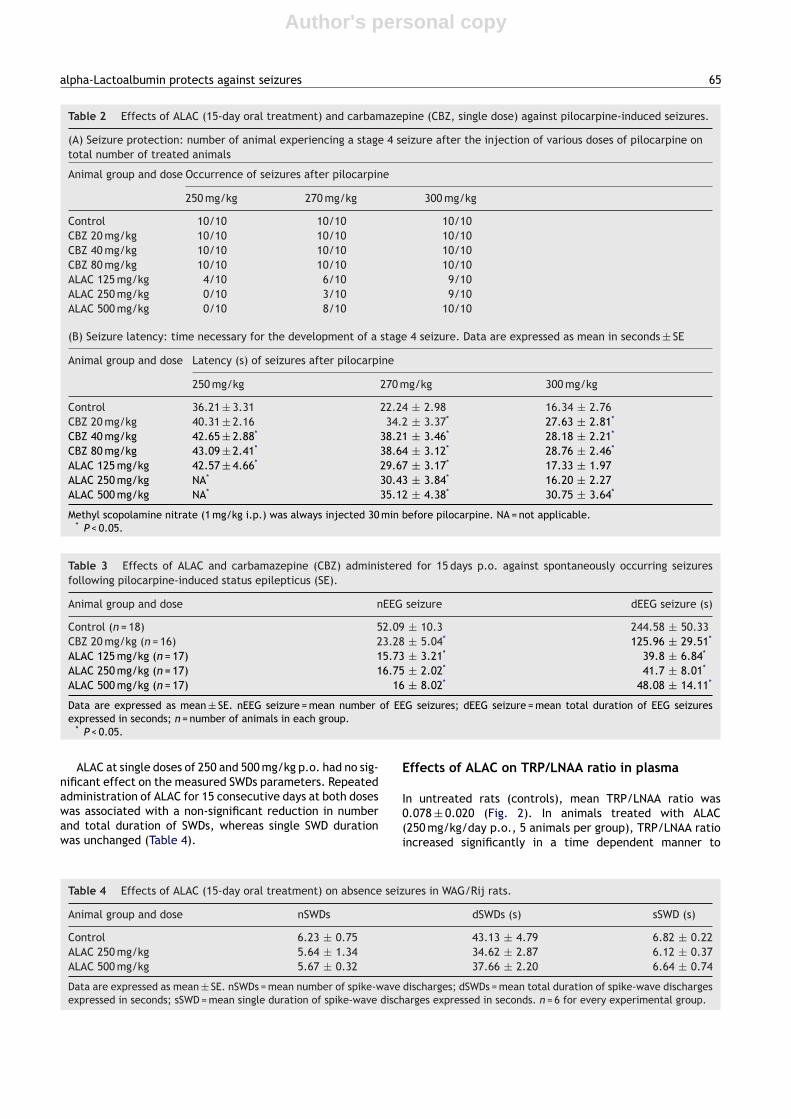
After single doses ranging between 100 mg/kg and 6 g/kgp.o., ALAC had no significant effects on the seizure param-eters considered. After treatment for 12 days (dose range:100 mg/kg/day to 6000 mg/kg/day p.o.), ALAC reduced themean duration of the tonic extension, without clear dosedependency. None of the doses tested was effective inpreventing hind leg tonic extension, while at doses of250—1000 mg/kg/day a significant reduction of the tonicextension duration was observed, with the 500 mg/kg dosebeing most clearly differentiated from controls (Table 1).
Table 1 Effects of ALAC (12-day oral treatment) on theduration of tonic extension in the maximal electroshockseizure test.
Dose/animal group Duration P value
Control 7.22 ± 0.20ALAC 100 mg/kg 7.18 ± 0.21 0.83476ALAC 250 mg/kg 5.93 ± 0.83* 0.02968ALAC 500 mg/kg 6.12 ± 0.28* 0.00654ALAC 1000 mg/kg 5.6 ± 0.72* 0.01654ALAC 2000 mg/kg 6.52 ± 0.27 0.08562ALAC 4000 mg/kg 7.17 ± 0.58 0.92850ALAC 6000 mg/kg 7.13 ± 0.24 0.88303
Data are expressed as mean in seconds ± SE. Six animals havebeen used in every group.
* P < 0.05.
When re-tested one week after electrical stimulation, allgroups showed a return to control conditions.
Effects of ALAC on pilocarpine-induced seizures
Pilocarpine, at doses of 250, 270 and 300 mg/kg i.p., waseffective in inducing a stage 4 seizure in all animals treated,the only difference between groups being a shorter latencyand an increasing incidence of tonic seizures and mor-tality with increasing doses (Table 2). CBZ at doses upto 80 mg/kg did not modify seizure incidence and mortal-ity but increased significantly seizure latency at all dosesin each pilocarpine subgroup (Table 2). Conversely, ALACwas effective in preventing seizures induced by injectionof 250 mg/kg pilocarpine, without a clear dose depen-dency. ALAC also prolonged seizure latency at doses of 250and 500 mg/kg/day, the effect being especially evident at250 mg/kg/day (Table 2). The protective effect of ALACwas less evident after a pilocarpine dose of 270 mg/kg,and was barely detectable at the highest pilocarpine dose(300 mg/kg).
Effects of ALAC on chronic seizures afterpilocarpine-induced SE
Pilocarpine (270 mg/kg i.p.) induced SE in 84% of treatedmice. EEG recordings after 6 weeks showed that 98% of themice selected developed chronic seizures. CBZ, at the doseof 20 mg/kg/day, reduced significantly (P < 0.05) the numberof spontaneous seizures and their total duration during the6-h recording after injection (Table 3). All ALAC doses testedwere similarly effective (P < 0.05) in reducing both the num-ber of seizures and their total duration, without a clear dosedependency (Table 3).
Effects of ALAC in WAG/Rij rats
In control animals at 6 months of age the mean numberof SWDs (nSWDs) per 30 min epoch was 6.23 ± 0.75, with amean total duration (dSWDs) of 43.1 ± 4.79 s and a meansingle duration (sSWD) of 6.82 ± 0.22 (Table 4).
Author's personal copy
alpha-Lactoalbumin protects against seizures 65
Table 2 Effects of ALAC (15-day oral treatment) and carbamazepine (CBZ, single dose) against pilocarpine-induced seizures.
(A) Seizure protection: number of animal experiencing a stage 4 seizure after the injection of various doses of pilocarpine ontotal number of treated animals
Animal group and dose Occurrence of seizures after pilocarpine
250 mg/kg 270 mg/kg 300 mg/kg
Control 10/10 10/10 10/10CBZ 20 mg/kg 10/10 10/10 10/10CBZ 40 mg/kg 10/10 10/10 10/10CBZ 80 mg/kg 10/10 10/10 10/10ALAC 125 mg/kg 4/10 6/10 9/10ALAC 250 mg/kg 0/10 3/10 9/10ALAC 500 mg/kg 0/10 8/10 10/10
(B) Seizure latency: time necessary for the development of a stage 4 seizure. Data are expressed as mean in seconds ± SE
Animal group and dose Latency (s) of seizures after pilocarpine
250 mg/kg 270 mg/kg 300 mg/kg
Control 36.21 ± 3.31 22.24 ± 2.98 16.34 ± 2.76CBZ 20 mg/kg 40.31 ± 2.16 34.2 ± 3.37* 27.63 ± 2.81*
CBZ 40 mg/kg 42.65 ± 2.88* 38.21 ± 3.46* 28.18 ± 2.21*
CBZ 80 mg/kg 43.09 ± 2.41* 38.64 ± 3.12* 28.76 ± 2.46*
ALAC 125 mg/kg 42.57 ± 4.66* 29.67 ± 3.17* 17.33 ± 1.97ALAC 250 mg/kg NA* 30.43 ± 3.84* 16.20 ± 2.27ALAC 500 mg/kg NA* 35.12 ± 4.38* 30.75 ± 3.64*
Methyl scopolamine nitrate (1 mg/kg i.p.) was always injected 30 min before pilocarpine. NA = not applicable.* P < 0.05.
Table 3 Effects of ALAC and carbamazepine (CBZ) administered for 15 days p.o. against spontaneously occurring seizuresfollowing pilocarpine-induced status epilepticus (SE).
Animal group and dose nEEG seizure dEEG seizure (s)
Control (n = 18) 52.09 ± 10.3 244.58 ± 50.33CBZ 20 mg/kg (n = 16) 23.28 ± 5.04* 125.96 ± 29.51*
ALAC 125 mg/kg (n = 17) 15.73 ± 3.21* 39.8 ± 6.84*
ALAC 250 mg/kg (n = 17) 16.75 ± 2.02* 41.7 ± 8.01*
ALAC 500 mg/kg (n = 17) 16 ± 8.02* 48.08 ± 14.11*
Data are expressed as mean ± SE. nEEG seizure = mean number of EEG seizures; dEEG seizure = mean total duration of EEG seizuresexpressed in seconds; n = number of animals in each group.
* P < 0.05.
ALAC at single doses of 250 and 500 mg/kg p.o. had no sig-nificant effect on the measured SWDs parameters. Repeatedadministration of ALAC for 15 consecutive days at both doseswas associated with a non-significant reduction in numberand total duration of SWDs, whereas single SWD durationwas unchanged (Table 4).
Effects of ALAC on TRP/LNAA ratio in plasma
In untreated rats (controls), mean TRP/LNAA ratio was0.078 ± 0.020 (Fig. 2). In animals treated with ALAC(250 mg/kg/day p.o., 5 animals per group), TRP/LNAA ratioincreased significantly in a time dependent manner to
Table 4 Effects of ALAC (15-day oral treatment) on absence seizures in WAG/Rij rats.
Animal group and dose nSWDs dSWDs (s) sSWD (s)
Control 6.23 ± 0.75 43.13 ± 4.79 6.82 ± 0.22ALAC 250 mg/kg 5.64 ± 1.34 34.62 ± 2.87 6.12 ± 0.37ALAC 500 mg/kg 5.67 ± 0.32 37.66 ± 2.20 6.64 ± 0.74
Data are expressed as mean ± SE. nSWDs = mean number of spike-wave discharges; dSWDs = mean total duration of spike-wave dischargesexpressed in seconds; sSWD = mean single duration of spike-wave discharges expressed in seconds. n = 6 for every experimental group.

Author's personal copy
66 R. Citraro et al.
Figure 2 TRP/LNAA ratio in plasma. n = 7 in every group. Dataare expressed as means ± SEM. TRP = tryptophan; LNAA = largeneutral aminoacids.
0.183 ± 0.032 after 7 days and 0.210 ± 0.040 after 14 days(Fig. 2).
Discussion
Epilepsies represent a heterogeneous group of disorders withdiverse etiologies, electrographical and behavioural seizurepatterns, and pharmacological sensitivities. 5-HT receptorsare expressed in almost all networks involved in ictoge-nesis. In fact, 5-HT appears to play an important role inthe processes involved in seizure development, propagationand maintenance (Buterbaugh, 1978; Racine and Coscina,1979; Browning et al., 1997; Prendiville and Gale, 1993;Wada et al., 1993; Jobe et al., 1973; Dailey et al., 1989,1992a,b; Pranzatelli, 1994). Depletion of brain 5-HT is knownto increase the severity of convulsions induced by audio-genic, chemical and electrical stimuli, usually by loweringseizure threshold (Jobe et al., 1973; Statnick et al., 1996).Conversely, compounds that increase levels of extracellular5-HT, such as 5-HTP and SSRIs, inhibit both limbic and gen-eralized seizures (Dailey et al., 1992a,b; Dailey et al., 1996;Yan et al., 1992, 1994a,b; Browning et al., 1997).
Since 5-HT can cause a significant shift of excitabil-ity in networks involved in epilepsy, compounds that alterthe concentration of 5-HT may impact on the pathogenesisand, possibly, the treatment of some epileptic conditions.In this study, we demonstrated that ALAC exhibits anticon-vulsant properties in some models and modulates seizureresponses in other models. These actions are associated withan increase in plasma TRP/LNAA ratio, which most likelytranslates into an increase in cerebral 5-HT synthesis. Acause effect relationship between ALAC treatment and anincrease in cerebral 5-HT availability, which requires timeto develop, is strongly supported by the ineffectiveness of asingle dose of ALAC in all seizure models tested. The resultsobtained in each of the models are discussed in more detailin the sections below.
Effects of ALAC against audiogenic seizures in theGEPR-9 model
GEPR-9 rats are a model of epilepsy and affective disorderin which the innate deficit of both the serotoninergic andnoradrenergic systems results in lowered seizure threshold
and development of severe seizures after sound stimulation(Jobe et al., 1994; Jobe, 2003; Jobe and Dailey, 2000). Theseverity of audiogenic seizures in these rats is known to be atleast in part dependent on deficiency of brain 5-HT. Previousexperiments have shown that drugs increasing synaptic 5-HT levels, such as fluoxetine, exert anticonvulsant effects inthis model (Dailey et al., 1992a,b, 1996; Yan et al., 1994a,b;Browning et al., 1997; Prendiville and Gale, 1993). Audio-genic seizures seem to originate from the brainstem, whichis a potential target for the anticonvulsant action of drugsacting through 5-HT. The substantia nigra is also known tomodulate the expression of epileptic seizures (Velísková andMoshé, 2006) and is another possible site of action, espe-cially in view of the observation that changes in the levels ofnigral 5-HT impact on seizures severity (Pasini et al., 1996).
In our experiments, ALAC was able to protect against thetonic phase, but time to onset of clonus was also signifi-cantly increased after multiple dose treatment with ALAC incomparison to vehicle treated animals. The effect of ALACshowed a significant dose-dependent relationship, and wasmaintained for up to 24 h after the last administration, witha clear attenuation after 48 h and complete recovery after1 week. This time course is consistent with the assumptionthat the anticonvulsant effects are due to an increase inbrain 5-HT, and with the observation that plasma TRP/LNAAsratio increased 2- to 3-fold after treatment with ALAC for5—14 days, respectively.
Effects of ALAC against MES-induced seizures
ALAC was not effective against MES-induced seizures,although it modified the mean duration of tonic extensionat doses ranging between 250 mg/kg/day and 1000 mg/kgwith no clear dose dependency and no evidence for thiseffect persisting at higher doses. Our results are only par-tially in agreement with evidence indicating that fluoxetineexerts anticonvulsant activity against MES-induced seizuresin rats (Borowicz et al., 2006) by modifying the brain lev-els of 5-HT (Browning et al., 1997). It has have suggestedthat enhancement of 5-HT transmission may selectivelyreduce the sensitivity of brainstem circuits to seizure initia-tion (Prendiville and Gale, 1993). Whether this effect couldexplain the ALAC-induced shortening of the duration of tonicextension in this model remains open to speculation.
Effects of ALAC against pilocarpine-inducedseizures
Pilocarpine-induced seizures have been considered as amodel of refractory epilepsy (Turski et al., 1983). Injec-tion of pilocarpine (270 mg/kg) produces a sequence ofbehavioural alterations which progress to SE (Cavalheiroet al., 1991; Turski et al., 1989). Previous investigatorsreported that CBZ is ineffective in this model and that fluox-etine at doses of 10 and 20 mg/kg has proconvulsant actions(Freitas et al., 2006; Turski et al., 1989). In our experi-ments, we confirmed that CBZ does not protect animals fromdeveloping SE and seizures, although it did increase seizurelatency in a dose-dependent manner. Administration of ALACfor 15 consecutive days was effective in preventing seizuresin most animals exposed to a 250 mg dose of pilocarpine. The

Author's personal copy
alpha-Lactoalbumin protects against seizures 67
response did not show a clear dose dependent relationship,with the exception of the latency to clonus which increaseddose-dependently. In conclusion, ALAC showed some effec-tiveness in this model, at least in protecting against seizuresinduced by the lowest dose of pilocarpine. Our results areapparently in contrast with those reported for fluoxetine.However, it should be pointed out that in the fluoxetineexperiments conducted by Freitas et al. (2006), the doseof pilocarpine was much higher (400 mg/kg) than that foundto be antagonized by ALAC in our study. Moreover, in thestudy by Freitas et al. (2006) fluoxetine was administered30 min before seizure induction, while in our experimentstests were conducted after 15 days of ALAC treatment,which may have resulted in greater increase in cerebral 5-HT availability and, possibly, other adaptive changes in thebrain.
Effects of ALAC against spontaneous seizures afterpilocarpine-induced SE
The pilocarpine post-SE model is widely used to repro-duce spontaneously occurring seizures accompanied byhippocampal neurodegeneration, with some similarities tohuman temporal lobe epilepsy. The serotoninergic sys-tem may play a role in modulating AED responsiveness inpatients with temporal lobe epilepsy (Kauffman et al., 2009;Hecimovic et al., 2010). Animals exposed to pilocarpine-induced SE develop over time EEG alterations which areoften followed by behavioural seizures. We confirmed thatCBZ is an effective anticonvulsant in this model, as previ-ously reported (Leite and Cavalheiro, 1995). Fluoxetine hasalso been found to be effective in this model (Hernandezet al., 2002), presumably by increasing synaptic 5-HT con-centrations. In our experiments, ALAC reduced the EEGalterations at all doses tested, but the effect was notdose-dependent. This could be speculatively explainedby inability of increased brain 5-HT content to com-pletely prevent seizure generation and propagation in thismodel.
Effects of ALAC against absence seizures inWAG/Rij rats
ALAC was devoid of significant effects in the WAG/Rij rats,which are extensively used as a model of absence epilepsy(Crunelli and Leresche, 2002; Coenen and Van Luijtelaar,2003). Several data indicate that, in addition to alterationsin GABAergic and glutamatergic systems, serotoninergicmechanisms play a role in the triggering and maintenanceof epileptiform activity in absence epilepsy (Filakovszkyet al., 1999, 2001; Bercovici et al., 2007; Graf et al.,2004). However, studies investigating the effect of 5-HTon absence seizures have yielded inconsistent results. Forexample, in the genetic absence epilepsy rat from Stras-bourg (GAERS), no effects were seen after decreasing 5-HTlevels with para-cholorophenylalanine, or after adminis-tering 5-hydroxy-TRP or methysergide (Marescaux et al.,1992). In the WAG/Rij absence seizure model, Coenen andVan Luijtelaar (2003) reported that 5-HT may indirectlymodulate absence seizures by modifying glutamatergictransmission. However, SSRIs have been reported to exac-
erbate typical absence seizures, presumably by increasing5-HT activity (Jakus et al., 2003).
Conclusions
We evaluated the efficacy of ALAC in various animal modelsof seizures and epilepsy. Our results clearly show that acuteadministration of this compound is ineffective, whereas pro-longed treatment for 5 days or longer is able to preventseizures or attenuate epileptic activity in a number mod-els, an effect which is associated with an increased plasmaTRP/LNAA ratio. The activity profile of ALAC in these modelsshowed some similarities with that reported for fluoxe-tine, an observation which supports the hypothesis that theunderlying mechanisms involve an increase in brain 5-HT. Inmost models, the effects of ALAC were not dose-dependent,suggesting some saturation or self limiting mechanism.
Based on these findings, ALAC may be considered as apromising compound for potential protection against epilep-tic seizures. Further experiments in animals are needed todefine its mode of action and activity in other models. Givenits good safety profile in man, ALAC may also be consideredfor further pilot investigations to determine potential inter-actions with existing AEDs and to assess potential beneficialeffects against drug resistant seizures in the clinical setting.
Acknowledgments
This work was supported by the Italian Ministry of Educa-tion, University and Research (MIUR, Cofin 2007, Rome) andthe National Research Council (CNR, Rome) are gratefullyacknowledged. Dr. Donato Cosco is kindly acknowledged forthe technical preparation of carbamazepine solution.
References
Ahmad, S., Fowler, L.J., Whitton, P.S., 2005. Lamotrigine, carba-mazepine and phenytoin differentially alter extracellular levelsof 5-hydroxytryptamine, dopamine and amino acids. EpilepsyRes. 63, 141—149.
Albano, C., Cupello, A., Mainardi, P., Scarrone, S., Favale, E.,2006a. Successful treatment of epilepsy with serotonin reup-take inhibitors: proposed mechanism. Neurochem. Res. 31,509—514.
Albano, C., Mainardi, P., Leonardi, A., 2006b. An increasing in brainserotonin improves clinical outcome in drug resistant epilepticpatients. In: 7th European Congress on Epileptology , Helsinki2—8 July (abstract).
Alexander, G.J., Kopeloff, L.M., 1976. Additive effect of 5-hydroxytryptophan and p-chloro-phenyalanine in preventingaudiogenic seizures in inbred mice. Psychopharmacol. Commun.2, 379—390.
Bagdy, G., Kecskemeti, V., Riba, P., Jakus, R., 2007. Serotonin andepilepsy. J. Neurochem. 100, 857—873.
Barnes, N.M., Sharp, T., 1999. A review of central 5-HT receptorsand their function. Neuropharmacology 38, 1083—1152.
Barton, M.E., Klein, B.D., Wolf, H.H., White, H.S., 2001. Pharmaco-logical characterization of the 6 Hz psychomotor seizure modelof partial epilepsy. Epilepsy Res. 47, 217—227.
Bercovici, E., Cortez, M.A., Snead 3rd, O.C., 2007. 5-HT2 modu-lation of AY-9944 induced atypical absence seizures. Neurosci.Lett. 418, 13—17.

Author's personal copy
68 R. Citraro et al.
Bonnycastle, D.D., Giarman, N.J., Paasonen, M.K., 1957. Anticon-vulsant compounds and 5-hydroxytryptamine in rat brain. Br. J.Pharmacol. 12, 228—231.
Borowicz, K.K., Stepien, K., Czuczwar, S.J., 2006. Fluoxetineenhances the anticonvulsant effects of conventional antiepilep-tic drugs in maximal electroshock seizures in mice. Pharmacol.Rep. 58, 83—90.
Browning, R.A., Wood, A.V., Merrill, M.A., Dailey, J.W., Jobe, P.C.,1997. Enhancement of the anticonvulsant effect of fluoxetinefollowing blockade of 5-HT 1A receptors. Eur. J. Pharmacol. 336,1—6.
Buterbaugh, C.G., 1978. Effects of drugs modifying central seroton-ergic function on the response of extensor and non-extensor ratsto maximal electroshock. Life Sci. 23, 2393—2904.
Cavalheiro, E.A., Leite, J.P., Bortolotto, Z.A., Turski, W.A., Ikonomi-dou, C., Turki, L., 1991. Long-term effects of pilocarpine in rats:structural damage of the brain triggers kindling and spontaneousrecurrent seizures. Epilepsia 32, 778—782.
Coenen, A.M., Van Luijtelaar, E.L., 2003. Genetic animal models forabsence epilepsy: a review of the WAG/Rij strain of rats. Behav.Genet. 33, 635—655.
Crunelli, V., Leresche, N., 2002. Childhood absence epilepsy: genes,channels, neurons and networks. Nat. Rev. Neurosci. 3, 371—382.
Cupello, A., Favale, E., Audenino, D., Scarrone, S., Gastaldi, S.,Albano, C., 2005. Decrease of serotonin transporters in bloodplatelets after epileptic seizures. Neurochem. Res. 30, 425—428.
Dailey, J.W., Reigel, C.E., Mishra, P.K., Jobe, P.C., 1989. Neurobi-ology of seizure predisposition in the genetically epilepsy-pronerat. Epilepsy Res. 3, 3—17.
Dailey, J.W., Yan, Q.S., Mishra, P.K., Burger, R.L., Jobe, P.C.,1992a. Effects of fluoxetine on convulsions and brain serotoninas detected by microdialysis in genetically epilepsy-prone rats.J. Pharmacol. 260, 533—540.
Dailey, J.W., Mishra, P.K., Ko, K.H., Penny, J.E., Jobe, P.C.,1992b. Serotonergic abnormalities in the central nervous sys-tem of seizure-naive genetically epilepsy-prone rats. Life Sci.50, 319—326.
Dailey, J.W., Yan, Q.S., Adams-Curtis, L.E., Ryu, J.R., Ko, K.H.,Mishra, P.K., Jobe, P.C., 1996. Neurochemical correlates of anti-epileptic drugs in the genetically epilepsy-prone rat (GEPR). LifeSci. 58, 259—266.
Dailey, J.W., Reith, M.E., Yan, Q.S., Li, M.Y., Jobe, P.C., 1997.Anticonvulsant doses of carbamazepine increase hippocam-pal extracellular serotonin in genetically epilepsy-prone rats:dose—response relationships. Neurosci. Lett. 227, 13—16.
De Sarro, G., Ferreri, G., Gareri, P., Russo, E., De Sarro, A., Gitto,R., Chimirri, A., 2003. Comparative anticonvulsant activity ofsome 2,3-benzodiazepine derivatives in rodents. Pharmacol.Biochem. Behav. 74, 595—602.
De Sarro, G., Rizzo, M., Spagnolo, C., Gitto, R., De Sarro, A.,Scotto, G., Zappala, M., Chimirri, A., 1999. Anticonvulsantactivity and plasma level of 2,3-benzodiazepin-4-ones (CFMs) ingenetically epilepsy-prone rats. Pharmacol. Biochem. Behav. 63,621—627.
De Sarro, G.B., Meldrum, B.S., De Sarro, A., Patel, S., 1992.Excitatory neurotransmitters in the lateral habenula and pedun-colopontine nucleus of rat modulate limbic seizures induced bypilocarpine. Brain Res. 591, 209—222.
Diksic, M.S., Okazawa, S., Nishizawa, M.L., Young, S.N., 2002. Adecrease in blood tryptophan concentration produces region-ally specific changes in brain serotonin synthesis in men andwomen: A PET study with alpha-[11C]methyl-L-tryptophan. Eur.Neuropsychopharmacol. 12, 403—404.
Favale, E., Audenino, D., Cocito, L., Albano, C., 2003. The anti-convulsant effect of citalopram as an indirect evidence ofserotonergic impairment in human epileptogenesis. Seizure 12,316—318.
Favale, E., Rubino, V., Mainardi, P., Lunardi, G., Albano, C., 1995.Anticonvulsant effect of fluoxetine in humans. Neurology 45,1926—1927.
Feurté, S., Gerozissis, K., Regnault, A., Paul, F.M., 2001. PlasmaTrp/LNAA ratio increases during chronic ingestion of an alpha-lactalbumin diet in rats. Nutr. Neurosci. 4, 413—418.
Filakovszky, J., Gerber, K., Bagdy, G., 1999. A serotonin-1A receptoragonist and N-methyl-D-aspartate receptor antagonist opposeeach others effects in a genetic rat epilepsy model. Neurosci.Lett. 261, 89—92.
Filakovszky, J., Kantor, S., Halasz, P., Bagdy, G., 2001. 8-OH-DPATand MK-801 affect epileptic activity independently of vigilance.Neurochem. Int. 38, 551—556.
Freitas, R.M., Sousa, F.C., Viana, G.S., Fonteles, M.M., 2006. Effectof gabaergic, glutamatergic, antipsychotic and antidepressantdrugs on pilocarpine-induced seizures and status epilepticus.Neurosci. Lett. 408, 79—83.
Frenhani, P.B., Burini, R.C., 1999. Mechanism of action and con-trol in the digestion of proteins and peptides in humans. Arq.Gastroenterol. 36, 139—147.
Gareri, P., Condorelli, D., Belluardo, N., Russo, E., Loiacono, A.,Barresi, V., Trovato-Salinaro, A., Mirone, M.B., Ferreri Ibbadu,G., De Sarro, G., 2004. Anticonvulsant effects of carbenoxolonein genetically epilepsy prone rats (GEPRs). Neuropharmacology47, 1205—1216.
Graf, M., Jakus, R., Kantor, S., Levay, G., Bagdy, G., 2004. Selec-tive 5-HT1A and 5-HT7 antagonists decrease epileptic activity inthe WAG/Rij rat model of absence epilepsy. Neurosci. Lett. 359,45—48.
Hecimovic, H., Jasminka, S., Lipa, C.S., Vida, D., Branimir, J., 2010.Association of serotonin transporter promoter (5-HTTLPR) andintron 2 (VNTR-2) polymorphisms with treatment response intemporal lobe epilepsy. Epilepsy Res. 91, 35—38.
Hernandez, E.J., Williams, P.A., Dudek, F.E., 2002. Effects offluoxetine and TFMPP on spontaneous seizures in rats withpilocarpine-induced epilepsy. Epilepsia 43, 1337—1345.
Heuther, G., Hajak, G., Reimer, A., Poeggeler, B., Blömer, M.,Rodenbeck, A., Rüther, E., 1992. The metabolic fate of infusedL-tryptophan in men: possible clinical implications of the accu-mulation of circulating tryptophan and tryptophan metabolites.Psychopharmacology (Berlin) 109, 422—432.
Jakus, R., Graf, M., Juhasz, G., Gerber, K., Levay, G., Halasz, P.,Bagdy, G., 2003. 5-HT2C receptors inhibit and 5-HT1A receptorsactivate the generation of spike-wave discharges in a genetic ratmodel of absence epilepsy. Exp. Neurol. 184, 964—972.
Jobe, P.C., Picchioni, A.L., Chin, L., 1973. Role of brain nore-pinephrine in audiogenic seizure in the rat. J. Pharmacol. Exp.Ther. 184, 1—10.
Jobe, P.C., Mishra, P.K., Browning, R.A., Wang, C., Adams-Curtis,L.E., Ko, K.H., Dailey, J.W., 1994. Noradrenergic abnormali-ties in the genetically epilepsy-prone rat. Brain Res. Bull. 35,493—504.
Jobe, P.C., Dailey, J.W., 2000. Genetically epilepsy-prone rats(GEPRs) in drug research. CNS Drug Rev. 6, 241—260.
Jobe, P.C., 2003. Common pathogenic mechanisms between depres-sion and epilepsy: an experimental perspective. Epilepsy Behav.3, 14—24.
Kauffman, M.A., Consalvo, D., Gonzalez-Moron, D., Aguirre, F.,D’Alessio, L., Kochen, S., 2009. Serotonin transporter gene vari-ation and refractory mesial temporal epilepsy with hippocampalsclerosis. Epilepsy Res. 85, 231—234.
Ko, F.J., Chiang, C.H., Liu, W.J., Chiang, W., 1993. Alteration ofamino acid in plasma and cerebrospinal fluid of children withseizure disorders. Gaoxiong Yi Xue Ke Xue Za Zhi 9, 131—142.
Leite, J.P., Cavalheiro, E.A., 1995. Effects of conventionalantiepileptic drugs in a model of spontaneous recurrent seizuresin rats. Epilepsy Res. 20, 93—104.

Author's personal copy
alpha-Lactoalbumin protects against seizures 69
Litchfield Jr., J.T., Wilcoxon, F.A., 1949. A simplified method of eval-uating dose-effect experiments. J. Pharmacol. Exp. Ther. 96,99—113.
Löscher, W., 2002. Animal models of epilepsy for the developmentof antiepileptogenic and disease-modifying drugs. A comparisonof pharmacology of kindling and post-status epilepticus modelsof temporal lobe epilepsy. Epilepsy Res. 50, 105—123.
Mainardi, P., Leonardi, A., Albano, C., 2008. Potentiation of brainserotonin activity may inhibit seizures, especially in drug-resistant epilepsy. Med. Hypotheses 70, 876—879.
Marescaux, C., Vergnes, M., Depaulis, A., 1992. Neurotransmis-sion in rats’ spontaneous generalized nonconvulsive epilepsy.Epilepsy Res. Suppl. 8, 335—343.
Marion, J.L., Bigot, J.C., Goas, J.Y., 1985. Alcoholic epilepsy:decrease of tryptophan levels in the blood and cerebrospinalfluid. Presse Med. 14, 681—683.
Markus, C.R., Olivier, B., Panhuysen, G.E., Van Der Gugten, J., Alles,M.S., Tuiten, A., Westenberg, H.G., Fekkes, D., Koppeschaar,H.F., de Haan, E.E., 2000. The bovine protein alpha-lactalbuminincreases the plasma ratio of tryptophan to the other largeneutral amino acids, and in vulnerable subjects raises brainserotonin activity, reduces cortisol concentration, and improvesmood under stress. Am. J. Clin. Nutr. 71, 1536—1544.
Maynert, E.W., Marczynski, T.J., Browning, R.A., 1975. The roleof the neurotransmitters in the epilepsies. Adv. Neurol. 13,79—147.
Minet-Ringuet, J., Le Ruyet, P.M., Tomé, D., Even, P.C., 2004. Atryptophan-rich protein diet efficiently restores sleep after fooddeprivation in the rat. Behav. Brain Res. 152, 335—340.
Okada, M., Kaneko, S., Hirano, T., Ishida, M., Kondo, T., Otani, K.,Fukushima, Y., 1992. Effects of zonisamide on extracellular lev-els of monoamine and its metabolite, and on Ca2+-dependentdopamine release. Epilepsy Res. 13, 113—119.
Orosco, M., Rouch, C., Beslot, F., Feurte, S., Regnault, A., Dauge,V., 2004. Alpha-lactalbumin-enriched diets enhance serotoninrelease and induce anxiolytic and rewarding effects in the rat.Behav. Brain Res. 148, 1—10.
Pallister, P.D., 1959. Aggravation of epilepsy by reserpine. Rocky Mt.Med. J. 56, 45—50.
Pasini, A., Tortorella, A., Gale, K., 1996. Anticonvulsant effect offluoxetine in substantia nigra is dependent upon endogenousserotonin. Brain Res. 724, 84—88.
Paxinos, G., Franklin, K.B.J., 2001. The Mouse Brain in StereotaxicCoordinates, 2nd ed. Academic Press, San Diego.
Paxinos, G., Watson, C., 1998. The Rat Brain in Stereotaxic Coordi-nates, 4th ed. Academic Press, San Diego, CA.
Pranzatelli, M.R., 1994. Serotonin and human myoclonus. Rationalefor the use of serotonin receptor agonists and antagonists. Arch.Neurol. 51, 605—617.
Prendiville, S., Gale, K., 1993. Anticonvulsant effect of fluoxetineon focally evoked limbic motor seizures in rats. Epilepsia 34,381—384.
Racine, R., Coscina, D.V., 1979. Effects of midbrain raphe lesions orsystemic p-chlorophenylalanine on the development of kindledseizures in rats. Brain Res. Bull. 4, 1—7.
Racine, R.J., 1972. Modification of seizure activity by electricalstimulation. I. After-discharge threshold. Electroencephalogr.Clin. Neurophysiol. 32, 269—279.
Russo, S., Kema, I.P., Bosker, F., Haavik, J., Korf, J., 2009. Tryp-tophan as an evolutionarily conserved signal to brain serotonin:molecular evidence and psychiatric implications. World J. Biol.Psychiatry 10, 258—268, Review.
Russo, E., Citraro, R., Scicchitano, F., De Fazio, S., Di Paola,E.D., Constanti, A., De Sarro, G., 2010. Comparison of theantiepileptogenic effects of an early long-term treatment withethosuximide or levetiracetam in a genetic animal model ofabsence epilepsy. Epilepsia 51, 1560—1569.
Russo, E., Constanti, A., Ferreri, G., Citraro, R., De Sarro, G.,2004. Nifedipine affects the anticonvulsant activity of topira-mate in various animal models of epilepsy. Neuropharmacology46, 865—878.
Schmidt, D., 2002. Strategies to prevent overtreatment withantiepileptic drugs in patients with epilepsy. Epilepsy Res. 52,61—69 (review).
Statnick, M.A., Maring-Smith, M.L., Clough, R.W., Wang, C.,Dailey, J.W., Jobe, P.C., Browning, R.A., 1996. Effect of5,7-dihydroxytryptamine on audiogenic seizures in geneticallyepilepsy-prone rats. Life Sci. 59, 1763—1771.
Truscott, T.C., 1975. Effects of phenyalanine and 5-hydroxytryptophan on seizures severity in mice. Pharmacol.Biochem. Behav. 3, 939—941.
Turski, W.A., Cavalheiro, E.A., Schwarz, M., Czuczwar, S.J.,Kleinronk, Z., Turski, L., 1983. Limbic seizures produced bypilocarpine in rats: behavioural, eletroencephalographic andneuropathological study, Behav. Brain Res. 9, 315—336.
Turski, L., Ikonomidou, C., Turski, W.A., Bortolotto, Z.A., Cav-alheiro, E.A., 1989. Review: cholinergic mechanisms andepileptogenesis. The seizures induced by pilocarpine: anovel experimental model of intractable epilepsy. Synapse 3,154—171.
van Zutphen, L.F., Baumans, V., Beynen, A.C., 2001. Principles ofLaboratory Animal Science, Rev ed. Elsevier, Amsterdam.
Velísková, J., Moshé, S.L., 2006. Update on the role of substantianigra pars reticulata in the regulation of seizures. Epilepsy Curr.6, 83—87.
Wada, Y., Nakamura, M., Hasegawa, H., Yamaguchi, N.,1993. Intra-hippocampal injection of 8-hydroxy-2-(di-n-propylamino)tetralin (8-OH-DPAT) inhibits partial andgeneralized seizures induced by kindling stimulation in cats.Neurosci. Lett. 159, 179—182.
Wang, J., Huang, Z.H., Gage, D.A., Watson, J.T., 1994. Anal-ysis of amino acids by gas chromatography—flame ionizationdetection and gas chromatography—mass spectrometry: simul-taneous derivatization of functional groups by an aqueous-phasechloroformate-mediated reaction. J. Chromatogr. A 663,71—78.
Yan, Q.S., Jobe, P.C., Dailey, J.W., 1994a. Evidence that a sero-tonergic mechanism is involved in the anticonvulsant effect offluoxetine in genetically epilepsy-prone rats. Eur. J. Pharmacol.252, 105—112.
Yan, Q.S., Jobe, P.C., Cheong, J.H., Ko, K.H., Dailey, J.W., 1994b.Role of serotonin in the anticonvulsant effect of fluoxetine ingenetically epilepsy-prone rats. Naunyn Schmiedebergs Arch.Pharmacol. 350, 149—152.
Yan, Q.S., Mishra, P.K., Burger, R.L., Bettendorf, A.F., Jobe,P.C., Dailey, J.W., 1992. Evidence that carbamazepine andantiepilepsirine may produce a component of their anti-convulsant effects by activating serotonergic neurons ingenetically epilepsy-prone rats. J. Pharmacol. Exp. Ther. 261,652—659.
Yokogoshi, H., Wurtman, R.J., 1986. Meal composition and plasmaamino acid ratios: effect of various proteins or carbohy-drates, and of various protein concentrations. Metabolism 35,837—842.




















