
The Effect of Functionally Graded Materials on Temperature ...

593
Mol. Biol. Evol. 18(4):593–605. 2001q 2001 by the Society for Molecular Biology and Evolution. ISSN: 0737-4038
Phylogeny, Function, and Evolution of the Cupins, a StructurallyConserved, Functionally Diverse Superfamily of Proteins
Sawsan Khuri, Freek T. Bakker,1 and Jim M. DunwellSchool of Plant Sciences, University of Reading, Reading, England
The cupin superfamily is a group of functionally diverse proteins that are found in all three kingdoms of life,Archaea, Eubacteria, and Eukaryota. These proteins have a characteristic signature domain comprising two histidine-containing motifs separated by an intermotif region of variable length. This domain consists of six beta strandswithin a conserved beta barrel structure. Most cupins, such as microbial phosphomannose isomerases (PMIs), AraC-type transcriptional regulators, and cereal oxalate oxidases (OXOs), contain only a single domain, whereas others,such as seed storage proteins and oxalate decarboxylases (OXDCs), are bi-cupins with two pairs of motifs. Althoughsome cupins have known functions and have been characterized at the biochemical level, the majority are knownonly from gene cloning or sequencing projects. In this study, phylogenetic analyses were conducted on the conserveddomain to investigate the evolution and structure/function relationships of cupins, with an emphasis on single-domain plant germin-like proteins (GLPs). An unrooted phylogeny of cupins from a wide spectrum of evolutionarylineages identified three main clusters, microbial PMIs, OXDCs, and plant GLPs. The sister group to the plantGLPs in the global analysis was then used to root a phylogeny of all available plant GLPs. The resulting phylogenycontained three main clades, classifying the GLPs into distinct subfamilies. It is suggested that these subfamiliescorrelate with functional categories, one of which contains the bifunctional barley germin that has both OXO andsuperoxide dismutase (SOD) activity. It is proposed that GLPs function primarily as SODs, enzymes that protectplants from the effects of oxidative stress. Closer inspection of the DNA sequence encoding the intermotif regionin plant GLPs showed global conservation of thymine in the second codon position, a character associated withhydrophobic residues. Since many of these proteins are multimeric and enzymatically inactive in their monomericstate, this conservation of hydrophobicity is thought to be associated with the need to maintain the various monomer-monomer interactions. The type of structure-based predictive analysis presented in this paper is an important ap-proach for understanding gene function and evolution in an era when genomes from a wide range of organisms arebeing sequenced at a rapid rate.
Introduction
The recently designated cupin superfamily of pro-teins (Dunwell 1998a) offers a model with which to in-vestigate evolutionary relationships among proteins ofsimilar structures but very different functions (Aravindand Koonin 1999a). The conserved domain, comprisinga six-stranded beta barrel structure (Gane, Dunwell, andWarwicker 1998; Woo et al. 2000), was given the namecupin (from the Latin word cupa, meaning ‘‘small bar-rel’’). Extensive sequence analysis has identified repre-sentatives of this type of protein in all prokaryotic andeukaryotic lineages examined, with the possible excep-tion of the spirochaetes (Dunwell, Khuri, and Gane2000).
The cupin superfamily was discovered when aminoacid sequence analysis, originally based on the nona-peptide ‘‘germin-box’’ (Lane et al. 1991), found a highlevel of similarity between fungal spherulins, producedupon spore formation in the slime mold Physarum po-lycephalum, and wheat germin, a thermostable, glyco-sylated protein produced during germination and shownto have oxalate oxidase (OXO) (EC 1.2.3.4) activity(Lane et al. 1993; Lane 2000). This small family of ger-
1 Present address: Wageningen University, Plant TaxonomyGroup, Wageningen, the Netherlands.
Key words: phylogeny, cupin, germin, oxalate oxidase, phos-phomannose isomerase, protein structure.
Address for correspondence and reprints: Jim M. Dunwell, Schoolof Plant Sciences, University of Reading, Reading RG6 6AS, UnitedKingdom. E-mail: [email protected].
min-related proteins was then found to share a smallnumber of globally conserved residues with 7S (vicilin)and 11S (legumin) seed storage proteins (Baumlein etal. 1995). Subsequent analysis (Dunwell and Gane1998), using an extended version of the germin-box,identified the conserved domain in a large number ofmicrobial proteins. These included type II phosphom-annose isomerases (PMIs) (EC 5.3.1.8), found in a largenumber of prokaryotes, AraC-type (Gallegos et al. 1997)and other (Aravind and Koonin 1999b) transcriptionregulators, gentisate 1,2-dioxygenases (GDOs) (EC1.13.11.4), and oxalate decarboxylases (OXDCs) (EC4.1.1.2).
Although some functional information is available,the majority of bacterial and archaeal cupins, as well asan increasing number from plants (generally known asgermin-like proteins [GLPs]) and animals, have beenidentified from genome sequencing projects and are ofunknown function. Together with the experimental con-firmation of the bifunctional nature of barley germin asan enzyme with both OXO and manganese superoxidedismutase (Mn-SOD) (EC 1.15.1.1) activity (Woo et al.2000), as well as the recent evidence that a GLP frommoss (Yamahara et al. 1999) and one from tobacco(Carter and Thornburg 2000) are Mn-SODs, there is cir-cumstantial evidence relating to the possible functionsof many other germins and GLPs. Some are expressedat critical developmental stages such as embryogenesis(Domon et al. 1995; Neutelings 1998) or floral induction(Heintzen et al. 1994; Staiger, Apel, and Trepp 1999),and many are induced by a range of stresses, either bi-otic, such as infection with powdery mildew (Thordahl-
by guest on August 7, 2016
http://mbe.oxfordjournals.org/
Dow
nloaded from

594 Khuri et al.
FIG. 1.—Alignment of the conserved domain in a sample of typical cupin sequences from archaea (pho1), bacteria (xca, pae3), fungi(fCVEA and B, fPPO1), moss (mBUN), gymnosperms (PCA), and higher plants (SAL, NPL, PSA3, and HVU1). The consensus cupin signatureresidues are highlighted, the metal-binding active site ligands of three histidines and a glutamate are in bold, and the arrows denote the six b-strands.
Christensen et al. 1997; Schweizer, Christoffel, andDudler 1999), or abiotic, such as exposure to salt (Hurk-man and Tanaka 1996), aluminum (Hamel, Breton, andHoude 1998), or high temperatures (Vallelian-Bind-schedler et al. 1998).
Cupins have two histidine-containing motifs (fig.1), corresponding to the C/D and G/H strands of OXO(Woo et al. 2000), separated by an intermotif region(IMR) which includes strands E and F and the interven-ing loop. The IMR of the single domain cupins variesin length from 15 amino acids in most archaeal andbacterial cupins to 26 residues in some of the cerealproteins (Dunwell, Khuri, and Gane 2000). It is pre-sumed that this variation in intermotif length, due prin-cipally to insertions in the interstrand loop, delayed theidentification of the conserved residues in this diversesuperfamily of proteins. It is now known that the twoconserved histidines and a glutamate in the first motif,together with a third conserved histidine in the secondmotif (fig. 1), act as ligands for the active-site metal,which has been shown to be a single manganese atomfor barley germin (Requena and Bornemann 1999; Wooet al. 2000), and for the GLPs from a moss (Barbulaunguiculata; Yamahara et al. 1999) and from tobacco(Carter and Thornburg 2000).
One diagnostic feature that can be used to discrim-inate between the various classes of cupin is whetherthe conserved domain occurs singly, in proteins such astranscription regulators and PMIs (Dunwell 1998a), orin a duplicated form within proteins such as the OXDCs(Dunwell and Gane 1998) and seed storage proteins(Baumlein et al. 1995); these two-domain proteins havebeen termed bi-cupins (Dunwell 1998a). The evolution-ary origin of cupins, and, indeed, that of bi-cupins, hasyet to be resolved; these latter could have been the resultof one or more gene duplication events leading to sep-arate evolution of the different classes (Shutov, Blattner,and Baumlein 1999). This paper presents evidence onthe evolutionary relationships within the cupin super-family, with a focus on single-domain plant GLPs, the
conservation of structure, and the significance of the ac-tive-site ligands in the possible function of theseproteins.
Materials and Methods
In view of the extensive variation in overall lengthof the various cupin proteins, and as a consequence ofpreliminary analyses, the sequences used in the presentstudy were restricted to the conserved cupin domain thatcomprises the two conserved motifs [G(X)5HXH-(X)3,4E(X)6G] and G(X)5PXG(X)2H(X)3N], together withthe intermotif region (IMR) (fig. 1).
The various sequences were identified usingBLAST searches with a variety of cupin sequences fromdifferent species. The nonredundant, nonmouse, andnonhuman EST and microbial GenBank databases at theNational Institute for Biotechnology Information (NIH,Bethesda, Md.) were mainly used via http://www.ncbi.nlm.nih.gov/BLAST; other sequences wereidentified by using the Institute of Genome ResearchTIGR database (Rockville, Md.) at http://www.tigr.org/cgi-bin/BlastSearch/blast.cgi and the Sanger Centre da-tabase (Cambridge, England) at http://sanger.ac.uk. Toavoid errors and misleading results, all microbial se-quences used in these analyses were from completedgenomes, except for the Streptococcus mutans bi-cupinputative OXDC. Similarly, identical cupin sequences(e.g., from Arabidopsis thaliana) were deleted, and onlya small number of plant ESTs were used in the phylo-genetic studies. General alignments were performed us-ing CLUSTAL W at the BCM Search Launcher at http://dot.imgenn.bcm.tmc.edu:9331/ and manually adjustedto produce a largely unambiguous alignment. Subse-quent phylogenetic analysis was performed usingPAUP*, version 4.0b4a (Swofford 1999). A globalalignment was constructed with a total of 120 proteinsequences comprising 25 plant GLPs, 5 fungal sequenc-es, and 90 microbial sequences, of which 13 were fromarchaeal species. A second separate alignment was con-
by guest on August 7, 2016
http://mbe.oxfordjournals.org/
Dow
nloaded from

Phylogeny, Function, and Evolution of Cupins 595
structed, containing 73 GLPs from plants and fungi. Re-sults of the unrooted global analysis indicated that ePBAwas an appropriate root for the GLP analysis (seebelow).
As described above, phylogenetic analyses wereperformed for both alignments with the IMR region in-cluded; excluding this region from the analyses resultedin loss of resolution in the topologies found. Parsimonysearches in both analyses involved heuristic searcheswith the following settings: gaps were treated as ‘‘miss-ing’’; branches were collapsed if maximum branchlength was zero; topological constraints were not en-forced, and STEEPEST DESCENT, ACCTRAN, andMULPARS were in effect. In order to measure cladesupport, jackknife analysis was carried out using PAUPwith settings emulating Parsimony Jackknifer (Farris1969), i.e., percentage of characters deleted in each rep-licate 5 37, ‘‘fast’’ stepwise addition, ‘‘Jac’’ resampling,and only groups with jac frequency .50% kept. Initial(equally weighted) searches were constrained by jack-knife trees thus obtained so as to avoid searchingthrough irrelevant parts of tree space. Given the expec-tation that the sequences in our data sets could be highlydivergent, the ‘‘Protpars’’ stepmatrix as implemented inMacClade (version 3.0.8a; Maddison and Maddison1992) was used in subsequent searches in order to takeinto account differences in probabilities of substitutionsamong amino acids (i.e., stepmatrix values are based onthe minimum number of changes between amino acids,calculated from the genetic code).
The 120-protein sequence alignment contained 70positions, of which 25 were part of the IMR. In total,the alignment contained 65 variable positions, of which62 were phylogenetically informative. A heuristic par-simony search was performed with 100 random se-quence additions (‘‘hold’’ 5 3) without branch swap-ping, with ‘‘MULPARS off,’’ and saving trees evenwhen they were not optimal overall. The resulting treeswere then used as starting trees in a search with ‘‘MUL-PARS on’’ and TBR branch swapping, which yielded.20,300 most-parsimonious trees (MPTs) of length2,039 (consistency index [CI] 5 0.35, retention index[RI] 5 0.57). The search was repeated with the Protparsstepmatrix implemented in order to reflect generally ob-served substitution patterns in amino acid sequences.This yielded 6 MPTs of length 2,688 (CI 5 0.26, RI 50.59), of which the strict consensus, with correspondingjackknife support values, was taken as the final result.This stepmatrix-weighted strict consensus tree waslargely congruent with and better resolved than the un-weighted strict consensus (not shown).
The sequence alignment of 73 plant GLPs con-tained 64 positions, of which 53 were phylogeneticallyinformative. A heuristic parsimony search with 50 rep-licates of random sequence addition (holding three treesper step), with TBR branch swapping, and keeping onlytrees compatible with a jackknife tree calculated fromthe same data set yielded 3,660 MPTs of length 631 (CI5 0.49, RI 5 0.75). Repeating the analysis with theProtpars stepmatrix implemented and without topologi-cal constraints yielded 2,928 MPTs of weighted length
761 (CI 5 0.42, RI 5 0.76), of which the strict consen-sus tree, rooted with the ‘‘ePBA’’ clade, is presentedbelow. This rooting was based on the global topology(see above). As with the global analysis, the effect ofimplementing the Protmars stepmatrix was a further in-crease in resolution in the plant-weighted strict consen-sus topology compared with that from the unweightedanalysis (not shown).
Each sequence used in this study is denoted by athree-letter code made up of the first letter of the genericname and the first two letters of the epithet, with pro-karyotic abbreviations in lowercase and eukaryotic onesin uppercase; e.g., ‘‘pae’’ denotes Pseudomonas aeru-ginosa, and ‘‘HVU’’ denotes Hordeum vulgare. Tables1 and 2 contain a complete listing of the sequences used.
Results and DiscussionGlobal AnalysisPhylogenetic Analysis
The conserved cupin domain section of the proteinsequences used in these analyses contained enough phy-logenetic signal to allow inference of evolutionary andfunctional relationships among them. The phylogenypresented in figure 2 shows that the plant GLP sequenc-es form a separate clade and that cupins from proteinsof similar, or the same, putative functions generally clus-ter together. While it is recognized that branching orderwithin main clades could change upon further taxonsampling, it is expected that the main clades themselveswill remain identifiable.
Most of the hypothesized functions of sequences inthe gene databases are based only on sequence similarityand not on biochemical evidence (see table 1). Thus,any functional relationships that are inferred from thisphylogeny are made in accordance with these assump-tions. The functional groups that resulted from this anal-ysis (fig. 2) were the bacterial and archaeal PMI andPMI-like sequences, the two bacterial GDOs, and thefungal OXDCs which formed a group together with pro-karyotic sequences of putative OXDC function. TheAraC-type and other transcription regulators, however,were dispersed.
The other sequences used were of unknown func-tion. Several changes in IMR length have occurredacross the phylogeny (fig. 2), from the basic, ancestralvalue of 15 residues found in the PMIs and transcrip-tional regulators to a maximum of 26 in a small numberof cereal GLPs (see below).
The PMI Cluster
Within the monophyletic PMI clade (fig. 2), thereare two key observations to note. First, the A. aeolicus(giz2983213, aae) sequence is on a relatively long branchderived from within the PMI group, a pattern consistentwith hypotheses about the ancient origin of this bacte-rium (Deckert et al. 1998). This cupin has only two con-served histidines; the glutamate and third histidine thatcomplete the putative active site are substituted by va-line and tyrosine, respectively. There is only a singlenucleotide difference between glutamate (GAA/G) and
by guest on August 7, 2016
http://mbe.oxfordjournals.org/
Dow
nloaded from
596 Khuri et al.
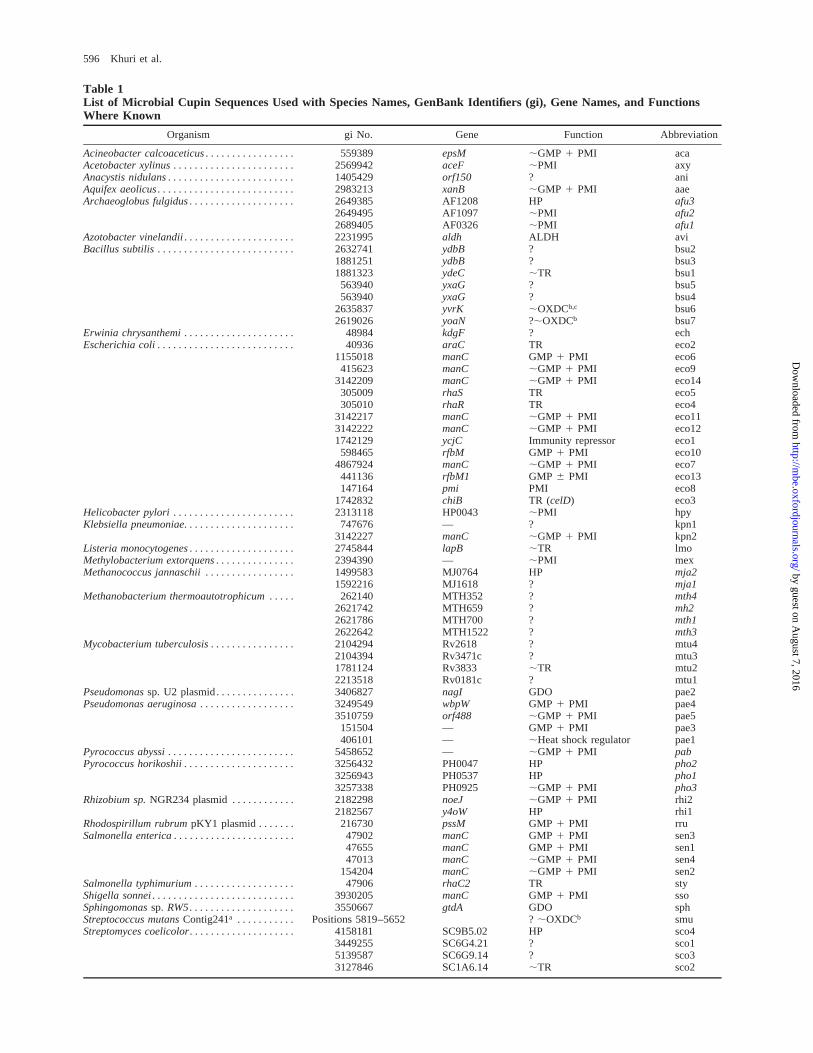
Table 1List of Microbial Cupin Sequences Used with Species Names, GenBank Identifiers (gi), Gene Names, and FunctionsWhere Known
Organism gi No. Gene Function Abbreviation
Acineobacter calcoaceticus . . . . . . . . . . . . . . . . .Acetobacter xylinus . . . . . . . . . . . . . . . . . . . . . . .Anacystis nidulans . . . . . . . . . . . . . . . . . . . . . . . .Aquifex aeolicus . . . . . . . . . . . . . . . . . . . . . . . . . .Archaeoglobus fulgidus . . . . . . . . . . . . . . . . . . . .
559389256994214054292983213264938526494952689405
epsMaceForf150xanBAF1208AF1097AF0326
;GMP 1 PMI;PMI?;GMP 1 PMIHP;PMI;PMI
acaaxyaniaaeafu3afu2afu1
Azotobacter vinelandii . . . . . . . . . . . . . . . . . . . . .Bacillus subtilis . . . . . . . . . . . . . . . . . . . . . . . . . .
2231995263274118812511881323
563940563940
26358372619026
aldhydbBydbBydeCyxaGyxaGyvrKyoaN
ALDH??;TR??;OXDCb,c
?;OXDCb
avibsu2bsu3bsu1bsu5bsu4bsu6bsu7
Erwinia chrysanthemi . . . . . . . . . . . . . . . . . . . . .Escherichia coli . . . . . . . . . . . . . . . . . . . . . . . . . .
4898440936
1155018415623
kdgFaraCmanCmanC
?TRGMP 1 PMI;GMP 1 PMI
echeco2eco6eco9
3142209305009305010
31422173142222
manCrhaSrhaRmanCmanC
;GMP 1 PMITRTR;GMP 1 PMI;GMP 1 PMI
eco14eco5eco4eco11eco12
1742129598465
4867924441136147164
1742832
ycjCrfbMmanCrfbM1pmichiB
Immunity repressorGMP 1 PMI;GMP 1 PMIGMP 6 PMIPMITR (celD)
eco1eco10eco7eco13eco8eco3
Helicobacter pylori . . . . . . . . . . . . . . . . . . . . . . .Klebsiella pneumoniae. . . . . . . . . . . . . . . . . . . . .
Listeria monocytogenes . . . . . . . . . . . . . . . . . . . .Methylobacterium extorquens . . . . . . . . . . . . . . .Methanococcus jannaschii . . . . . . . . . . . . . . . . .
2313118747676
31422272745844239439014995831592216
HP0043—manClapB—MJ0764MJ1618
;PMI?;GMP 1 PMI;TR;PMIHP?
hpykpn1kpn2lmomexmja2mja1
Methanobacterium thermoautotrophicum . . . . .
Mycobacterium tuberculosis . . . . . . . . . . . . . . . .
2621402621742262178626226422104294
MTH352MTH659MTH700MTH1522Rv2618
?????
mth4mh2mth1mth3mtu4
210439417811242213518
Rv3471cRv3833Rv0181c
?;TR?
mtu3mtu2mtu1
Pseudomonas sp. U2 plasmid . . . . . . . . . . . . . . .Pseudomonas aeruginosa . . . . . . . . . . . . . . . . . .
340682732495493510759
151504406101
nagIwbpWorf488——
GDOGMP 1 PMI;GMP 1 PMIGMP 1 PMI;Heat shock regulator
pae2pae4pae5pae3pae1
Pyrococcus abyssi . . . . . . . . . . . . . . . . . . . . . . . .Pyrococcus horikoshii . . . . . . . . . . . . . . . . . . . . .
5458652325643232569433257338
—PH0047PH0537PH0925
;GMP 1 PMIHPHP;GMP 1 PMI
pabpho2pho1pho3
Rhizobium sp. NGR234 plasmid . . . . . . . . . . . .
Rhodospirillum rubrum pKY1 plasmid . . . . . . .Salmonella enterica . . . . . . . . . . . . . . . . . . . . . . .
21822982182567
216730479024765547013
154204
noeJy4oWpssMmanCmanCmanCmanC
;GMP 1 PMIHPGMP 1 PMIGMP 1 PMIGMP 1 PMI;GMP 1 PMI;GMP 1 PMI
rhi2rhi1rrusen3sen1sen4sen2
Salmonella typhimurium . . . . . . . . . . . . . . . . . . .Shigella sonnei . . . . . . . . . . . . . . . . . . . . . . . . . . .Sphingomonas sp. RW5. . . . . . . . . . . . . . . . . . . .
4790639302053550667
rhaC2manCgtdA
TRGMP 1 PMIGDO
styssosph
Streptococcus mutans Contig241a . . . . . . . . . . .Streptomyces coelicolor. . . . . . . . . . . . . . . . . . . .
Positions 5819–56524158181344925551395873127846
SC9B5.02SC6G4.21SC6G9.14SC1A6.14
? ;OXDCb
HP??;TR
smusco4sco1sco3sco2
by guest on August 7, 2016
http://mbe.oxfordjournals.org/
Dow
nloaded from
Phylogeny, Function, and Evolution of Cupins 597
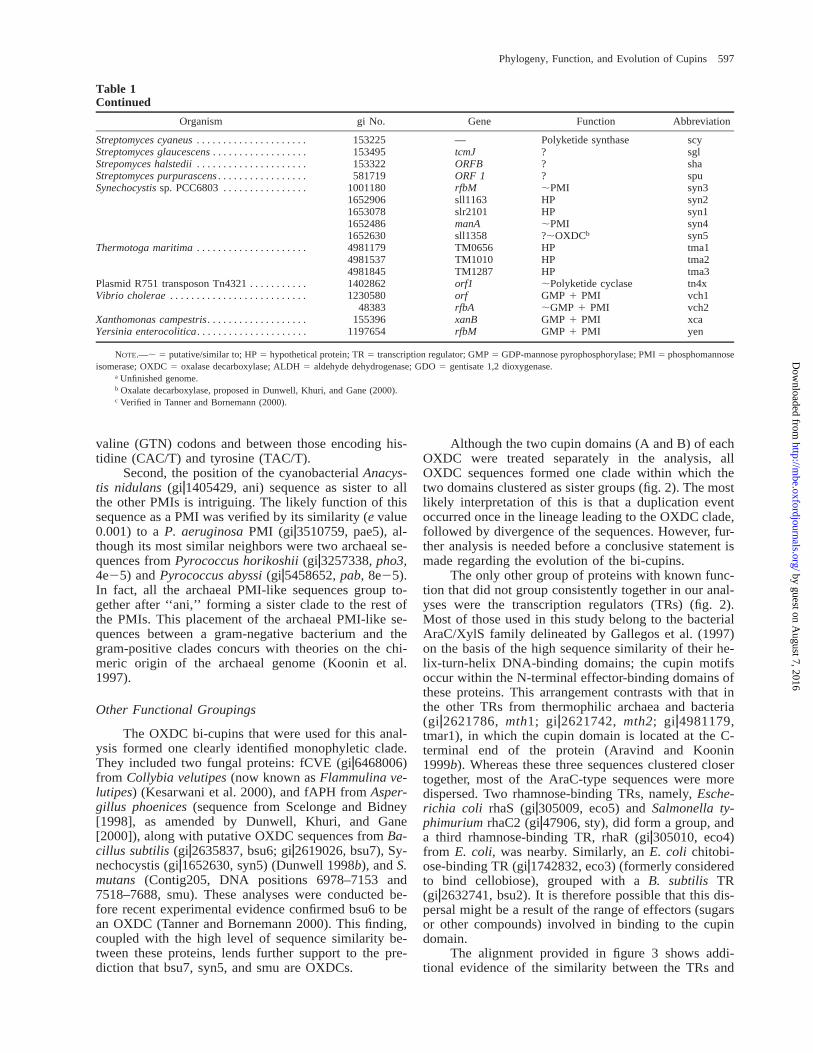
Table 1Continued
Organism gi No. Gene Function Abbreviation
Streptomyces cyaneus . . . . . . . . . . . . . . . . . . . . .Streptomyces glaucescens . . . . . . . . . . . . . . . . . .Strepomyces halstedii . . . . . . . . . . . . . . . . . . . . .Streptomyces purpurascens . . . . . . . . . . . . . . . . .Synechocystis sp. PCC6803 . . . . . . . . . . . . . . . .
153225153495153322581719
1001180
—tcmJORFBORF 1rfbM
Polyketide synthase???;PMI
scysglshaspusyn3
1652906165307816524861652630
sll1163slr2101manAsll1358
HPHP;PMI?;OXDCb
syn2syn1syn4syn5
Thermotoga maritima . . . . . . . . . . . . . . . . . . . . . 498117949815374981845
TM0656TM1010TM1287
HPHPHP
tma1tma2tma3
Plasmid R751 transposon Tn4321 . . . . . . . . . . .Vibrio cholerae . . . . . . . . . . . . . . . . . . . . . . . . . .
Xanthomonas campestris . . . . . . . . . . . . . . . . . . .Yersinia enterocolitica. . . . . . . . . . . . . . . . . . . . .
14028621230580
48383155396
1197654
orf1orfrfbAxanBrfbM
;Polyketide cyclaseGMP 1 PMI;GMP 1 PMIGMP 1 PMIGMP 1 PMI
tn4xvch1vch2xcayen
NOTE.—; 5 putative/similar to; HP 5 hypothetical protein; TR 5 transcription regulator; GMP 5 GDP-mannose pyrophosphorylase; PMI 5 phosphomannoseisomerase; OXDC 5 oxalase decarboxylase; ALDH 5 aldehyde dehydrogenase; GDO 5 gentisate 1,2 dioxygenase.
a Unfinished genome.b Oxalate decarboxylase, proposed in Dunwell, Khuri, and Gane (2000).c Verified in Tanner and Bornemann (2000).
valine (GTN) codons and between those encoding his-tidine (CAC/T) and tyrosine (TAC/T).
Second, the position of the cyanobacterial Anacys-tis nidulans (giz1405429, ani) sequence as sister to allthe other PMIs is intriguing. The likely function of thissequence as a PMI was verified by its similarity (e value0.001) to a P. aeruginosa PMI (giz3510759, pae5), al-though its most similar neighbors were two archaeal se-quences from Pyrococcus horikoshii (giz3257338, pho3,4e25) and Pyrococcus abyssi (giz5458652, pab, 8e25).In fact, all the archaeal PMI-like sequences group to-gether after ‘‘ani,’’ forming a sister clade to the rest ofthe PMIs. This placement of the archaeal PMI-like se-quences between a gram-negative bacterium and thegram-positive clades concurs with theories on the chi-meric origin of the archaeal genome (Koonin et al.1997).
Other Functional Groupings
The OXDC bi-cupins that were used for this anal-ysis formed one clearly identified monophyletic clade.They included two fungal proteins: fCVE (giz6468006)from Collybia velutipes (now known as Flammulina ve-lutipes) (Kesarwani et al. 2000), and fAPH from Asper-gillus phoenices (sequence from Scelonge and Bidney[1998], as amended by Dunwell, Khuri, and Gane[2000]), along with putative OXDC sequences from Ba-cillus subtilis (giz2635837, bsu6; giz2619026, bsu7), Sy-nechocystis (giz1652630, syn5) (Dunwell 1998b), and S.mutans (Contig205, DNA positions 6978–7153 and7518–7688, smu). These analyses were conducted be-fore recent experimental evidence confirmed bsu6 to bean OXDC (Tanner and Bornemann 2000). This finding,coupled with the high level of sequence similarity be-tween these proteins, lends further support to the pre-diction that bsu7, syn5, and smu are OXDCs.
Although the two cupin domains (A and B) of eachOXDC were treated separately in the analysis, allOXDC sequences formed one clade within which thetwo domains clustered as sister groups (fig. 2). The mostlikely interpretation of this is that a duplication eventoccurred once in the lineage leading to the OXDC clade,followed by divergence of the sequences. However, fur-ther analysis is needed before a conclusive statement ismade regarding the evolution of the bi-cupins.
The only other group of proteins with known func-tion that did not group consistently together in our anal-yses were the transcription regulators (TRs) (fig. 2).Most of those used in this study belong to the bacterialAraC/XylS family delineated by Gallegos et al. (1997)on the basis of the high sequence similarity of their he-lix-turn-helix DNA-binding domains; the cupin motifsoccur within the N-terminal effector-binding domains ofthese proteins. This arrangement contrasts with that inthe other TRs from thermophilic archaea and bacteria(giz2621786, mth1; giz2621742, mth2; giz4981179,tmar1), in which the cupin domain is located at the C-terminal end of the protein (Aravind and Koonin1999b). Whereas these three sequences clustered closertogether, most of the AraC-type sequences were moredispersed. Two rhamnose-binding TRs, namely, Esche-richia coli rhaS (giz305009, eco5) and Salmonella ty-phimurium rhaC2 (giz47906, sty), did form a group, anda third rhamnose-binding TR, rhaR (giz305010, eco4)from E. coli, was nearby. Similarly, an E. coli chitobi-ose-binding TR (giz1742832, eco3) (formerly consideredto bind cellobiose), grouped with a B. subtilis TR(giz2632741, bsu2). It is therefore possible that this dis-persal might be a result of the range of effectors (sugarsor other compounds) involved in binding to the cupindomain.
The alignment provided in figure 3 shows addi-tional evidence of the similarity between the TRs and
by guest on August 7, 2016
http://mbe.oxfordjournals.org/
Dow
nloaded from

598 Khuri et al.
Table 2List of Eukaryotic Cupin Sequences Used with Species Names, GenBank Identifiers (gi), and Protein Names
Species gi No. Protein Abbreviation
Arabidopsis thaliana . . . . . . . . . . . . . . . . . . . . . . 17551526714406664816766467826143869
GLP4GlpGlpGlpGlp
ATH1ATH10ATH13ATH14ATH15
56793282244819175517717551846899900
GLP7GlpGLP5GLP3aGLP10
ATH16ATH128ATH2ATH25ATH3
17551626721170409896821295941934730
GLP2aGlpGLP9glp type 2GLP10
ATH33ATH4ATH41ATH43ATH44
17551821755166672116830470781592672
GLP7GLP6GlpGlpGermin1
ATH46ATH47ATH5ATH51ATH52
67211676721166
GlpGlp
ATH6ATH7
Aspergillus phoenices . . . . . . . . . . . . . . . . . . . . .Atriplex lentiformis . . . . . . . . . . . . . . . . . . . . . . .Barbula unguiculata . . . . . . . . . . . . . . . . . . . . . .
67211656714408
NA49966226429233
GlpGlp—GlpPartiala
ATH8ATH9fAPHALEmBUN
Brassica napus . . . . . . . . . . . . . . . . . . . . . . . . . . .Ceratodon purpureus. . . . . . . . . . . . . . . . . . . . . .Collybia velutipes. . . . . . . . . . . . . . . . . . . . . . . . .Fragaria x ananassa . . . . . . . . . . . . . . . . . . . . . .Glycine max . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
91491060686946468006
NA4289984
GlpESTOXDCb
GlpEST
BNAemCPUfCVEFANeGMA
Gossypium hirsutum . . . . . . . . . . . . . . . . . . . . . .
Hordeum vulgare . . . . . . . . . . . . . . . . . . . . . . . . .
50478066650732289357
117193728152921070358
ESTglp1OXOa,c (1993)OXOa,c (1994)GlpGlp
eGHIGHIHVU1HVU2HVU3HVU4
Lycopersicon esculentum. . . . . . . . . . . . . . . . . . .Medicago truncatula . . . . . . . . . . . . . . . . . . . . . .Mesembryanthemum crystallinum. . . . . . . . . . . .Nicotiana plumbaginifolia. . . . . . . . . . . . . . . . . .
29794946101843
1672586090828
?ESTGlpNEC1a
LESeMTRMCRNPL
Oryza sativa . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2655291265528926552872655285
GER4GER3GER2GER1
OSA1OSA11OSA12OSA13
2801803549973050424615852087
Glp16RGLP1GlpGlp
OSA14OSA2OSA3OSA4
59973242398213293559
RGLP2Osglp1GER7
OSA5OSA6OSA7
Pharbitis nil . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Physarum polycephalum . . . . . . . . . . . . . . . . . . .
Pinus caribaea . . . . . . . . . . . . . . . . . . . . . . . . . . .
662292161262134860
2745849
GlpSpherulin 1ASpherulin 1BPcGER1
PNIfPPO1fPPO2PCA
Pinus radiata . . . . . . . . . . . . . . . . . . . . . . . . . . . . 293552127392606689036668903
PRGER1PartialGer2aGer1
PRAPSA1PSA3PSA4
Populus balsamifera . . . . . . . . . . . . . . . . . . . . . .Prunus persica . . . . . . . . . . . . . . . . . . . . . . . . . . .
3857819409851719168091916807
ESTABP19d
ABP19d
ABP20d
ePBAPPE1PPE2PPE3
Sinapis alba . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Solanum tuberosum . . . . . . . . . . . . . . . . . . . . . . .Triticum aestivum. . . . . . . . . . . . . . . . . . . . . . . . .
6834883171251
170698
GlplOXAOXAgf-2.8c
SALSTUTAE1
by guest on August 7, 2016
http://mbe.oxfordjournals.org/
Dow
nloaded from

Phylogeny, Function, and Evolution of Cupins 599
Table 2Continued
Species gi No. Protein Abbreviation
1772597121131
5869975
PSBGer1gf-3.8glp2b
TAE10TAE2TAE3
Zea mays. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
5869973177260117725994152154
glp2apSBGer3pSBGer2EST
TAES4TAE8TAE9eZMA
NOTE.—NA 5 not available.a Mn-superoxide dismutase.b Oxalate decarboxylase.c Oxalate oxidase.d Auxin-binding protein.
highlights three points. First, the AraC-type group canbe distinguished clearly from the thermophilic group inhaving only one of the conserved histidine residues inmotif 1. Second, the archetypal arabinose-binding E.coli AraC (giz40936, eco2), although similar to othercupins in terms of its beta-barrel structure (Soisson etal. 1997; Dunwell, Khuri, and Gane 2000), is not typicalof its group in terms of sequence. Third, the two se-quences from B. subtilis (giz1881323, bsu1) and Myco-bacterium tuberculosis (giz2213518, mtu2) both have aglutamine residue in place of the glutamate in the pu-tative binding site of these proteins. A more compre-hensive alignment of 112 TRs and putative TR sequenc-es showed that a total of 54 proteins with a glutamineat that position compared with 33 that had a glutamateresidue (data not shown). This contrasts with the higherconservation frequencies of the three histidines, whichwere 94, 95, and 104, respectively.
As expected for the majority of the eukaryotic cu-pins used, the plant sequences formed a monophyleticgroup which included the fungal sequence used, P. po-lycephalum spherulin (giz161261, fPPO1) (fig. 2). Thebest characterized of the plant GLPs are the germinsfrom barley (giz289356, HVU1) and wheat (giz170697,TAE1). The detailed analysis of the plant and fungalGLPs is provided below.
Plant and Fungal Germin-like ProteinsPhylogenetic Analysis
The plant GLPs used in this study included ex-amples from most main plant lineages, including cere-als, legumes, gymnosperms, and two moss GLPs (B.unguiculata, giz6429233, mBUN; Ceratodon purpureusEST giz6068694, emCPU). In addition, two fungalspherulins from P. polycephalum (giz161262 andgiz134860; fPPO1 and 2) were included. The resultingphylogeny contained three main groups (fig. 4), a resultwhich differs from that recently proposed by Carter andThornburg (1999, 2000), who identified five separateclades. However, Carter and Thornburg produced theirphylogeny using the whole protein sequence, rather thanonly the cupin domain. Furthermore, although the totalnumber of sequences used for the phylogenies was co-incidentally the same, Carter and Thornburg used 30sequences from A. thaliana, 10 from Oryza sativa and
33 GLPs from other plant species, including one moss,whereas this analysis utilized 24 from A. thaliana, 11from O. sativa, and 36 from other plant species, includ-ing (see above) two moss species and two from a fungus(table 2). The phylogeny presented in figure 4, therefore,gives a more complete picture of the evolutionary, aswell as the functional, relationships among GLPs.
The first group (subfamily 1 [s/f 1]) can be clas-sified as the ‘‘true germin’’ (Carter and Thornburg 1999)subclade and consists of the wheat and barley germins(giz170698, TAE1; giz289357, HVU1) along with someother GLPs of those species. Evidence is now accumu-lating that the true germins may be bifunctional enzymeswith both SOD and OXO activity (Woo et al. 2000).
Subfamily 2 (s/f 2) includes GLPs from a widerange of taxonomic groups, including other cereals,gymnosperms, and halophytic species, namely Atriplexlentiformis (giz4996622, ALE) and Mesembryanthe-mum crystallinum (giz167258, MCR). Furthermore,these cupins were isolated from diverse and seeminglyunconnected sources. The solanaceous group, for ex-ample, consisted of GLPs from Mn-deficient roots oftomato (giz2979494, LES), nectar of tobacco(giz6090828, NPL), and cell suspension cultures of po-tato (giz3171251, STU). The two gymnosperm GLPswere both isolated from somatic embryos (Neutelings1998).
Despite this diversity of origin, nested within thissubfamily is a group of proteins linked directly to stresstolerance: MCR was isolated from salt-stressed tissue,and OSA2 (giz5499730) and OSA5 (giz5499732) werelabeled stress-responsive root GLPs. In addition, twocentrally positioned proteins within s/f 2 are now knownto have Mn-SOD activity, namely NPL (giz6090828)from Nicotiana plumbaginifolia (Carter and Thornburg2000) and mBUN (giz6429233) from the moss B. un-guiculata (Yamahara et al. 1999). It is therefore possibleto predict that the GLPs in s/f 2 all have a role in tol-erance to oxidative stress and are most likely to be Mn-SODs. This is in accordance with the predictions ofCarter and Thornburg (2000), and it is expected thatmore experimental evidence will be forthcoming whichwill confirm these suggestions.
The third grouping, subfamily 3 (s/f 3), includeslow-affinity auxin-binding proteins from peach
by guest on August 7, 2016
http://mbe.oxfordjournals.org/
Dow
nloaded from

600 Khuri et al.
FIG. 2.—Global phylogeny (unrooted analysis): strict consensus of six most-parsimonious trees after Protpars stepmatrix character stateweighting (see text). Numbers at nodes indicate jackknife values (10,000 replicates). GLP 5 germin-like protein; PMI 5 phosphomannoseisomerase; OXDC 5 oxalate decarboxylase; GDO 5 gentisate 1,2 dioxygenase. Plant and fungal sequences are in uppercase letters; archaealones are italicized and boxed. Sequences have intermotif regions of 15 amino acids, with the following exceptions: dashed line 5 16 (sco2 has17); dotted line 5 20; dot-and-dash line 5 21 or more. TR 5 transcription regulator; prefix f 5 fungal; m 5 moss; e 5 EST; suffix A and B5 first (N-terminal) and second (C-terminal) domains of OXDC, respectively. Full species names and accession numbers of the sequences usedare presented in tables 1 and 2.
(giz4098517, PPE1; giz1916809, PPE2; giz1916807,PPE3), as well as GLPs known to have a role linked tocircadian rhythms and floral induction in A. thaliana(giz1755184, ATH25; Staiger, Apel, and Trepp 1999),Sinapis alba (giz683488, SAL; Heintzen et al. 1994),and Pharbitis nil (giz662292, PNI; Ono et al. 1996). Thissubfamily may therefore consist mainly of regulatory
proteins involved with auxin metabolism, either directlyor indirectly.
The Metal-Binding Active Site
As seen above, with few exceptions, the enzymaticfunction(s) of GLPs remain largely unknown. It is clear
by guest on August 7, 2016
http://mbe.oxfordjournals.org/
Dow
nloaded from

Phylogeny, Function, and Evolution of Cupins 601
FIG. 3.—Alignment of a sample of transcription regulators, with their effector sugars, where known. Residues are highlighted as in figure1. The upper group includes the bacterial AraC-type sequences with sequence sco4 included for comparison (see fig. 1). The lower groupcomprises the three sequences from thermophilic archaea and bacteria (Aravind and Koonin 1999b).
that most of them contain a single metal-binding activesite with three conserved histidine ligands and one glu-tamate ligand (fig. 1). The position of the active site inthe protein is protected within the beta-barrel structure(Woo et al. 2000), and it has recently been shown thatthe barley germin with OXO and SOD activity (Requenaand Bornemann 1999; Woo et al. 2000), and the moss(Yamahara et al. 1999) and tobacco (Carter and Thorn-bug 2000) GLPs with SOD activity are manganese-con-taining enzymes. Both of these enzymes generate hy-drogen peroxide, OXO by the oxidation of the oxalateion, and SOD by the dismutation of superoxide radicals.It is likely that the types of catalysis are similar in allmetal-binding GLPs or that there is a group, possibly s/f 3 as delineated above, that contains a different metaland therefore has slightly different properties.
Interestingly, a small number of GLPs do not con-tain all four of the highly conserved active-site residues.Instead of the third histidine (coded by CAY) residue inthe second motif, PNI (giz662292) has an aspartate(GAY), while ATH4 (giz6721170), ATH5 (giz6721168),and ATH7 (giz6721166) have a glutamine (CAR). Thelack of the third histidine infers that these proteins maynot bind a manganese atom, or that they bind it veryloosely and only under certain pH conditions. The con-served glutamate (GAR) in the first cupin motif can besubstituted, again, by either aspartate (GAY), as inTAE8 (giz1772601), or glutamine (CAR), as in mCPU(giz6068694), ATH6 (giz6721167), and ATH4(giz6721170). The absence of the negatively chargedglutamate makes it unlikely that these proteins wouldbind manganese, and it is not known whether the equallynegatively charged aspartate would provide the samefunction. In some Glycine max ESTs (giz5759981,giz6725711, and giz6566187), this position is occupiedby a positively charged lysine (AAR). Since all of thesechanges are due to a single nucleotide difference, theymay simply be mutations that maintain the structure ofthese proteins but alter the binding characteristics of theactive site and thus change its function.
The Intermotif Region
In the plant and fungal single-domain GLPs ex-amined to date, the intermotif region varies in lengthfrom the usual minimum of 20 (exceptionally, 19 insome legume sequences) to 26 amino acids in some bar-ley and wheat sequences (fig. 4). There is a high levelof conservation within the IMR across all of the cupinsubfamilies identified above, at both the protein and theDNA levels. An alignment of the DNA sequences (fig.5) encoding the GLPs used in the global analysis allowsthe identification of certain patterns within the regionencoding the IMR. First, two GLPs (PPE1 giz4098517and PSA1 giz2739260) show evidence of two compen-satory frameshifts. Second, where the IMR is longerthan 20 amino acids, the position of the additional res-idues is consistently within the loop between structuralstrands E and F, to either side of a conserved asparagine.Third, the alignment shows a 100% conservation of thy-mine in the second base positions of three codons withinstrand E and two codons in strand F. Apart from methi-onine, codons with thymine in the second position codefor leucine (L), isoleucine (I), valine (V), and phenyl-alanine (F), all of which are hydrophobic residues (Trin-quier and Sanejouand 1998) and may help to maintainthe intermonomer interactions necessary for the assem-bly of the homohexamers required for enzyme activityin germins (see references in Dunwell, Khuri, and Gane2000). These observations are also linked to conserva-tion of the core region of the barrel structure of cupins,in that the length and composition of the b-strands re-mains largely conserved, with most variability occurringin the structurally less important central loop region be-tween strands E and F.
Cupin ESTs
There are a rapidly increasing number of cupinEST sequences in the databases, from a wide range ofspecies; only a sample of them was used in the phylog-enies described above. An alignment of all 61 cupin
by guest on August 7, 2016
http://mbe.oxfordjournals.org/
Dow
nloaded from

602 Khuri et al.
FIG. 4.—Plant and fungal GLPs: strict consensus of 2,928 most-parsimonious trees after Protpars stepmatrix character state weighting (see text),rooted on basal branch as found in the global analysis (fig. 1). Numbers at nodes indicate jackknife values (10,000 replicates). Intermotif regions are20 amino acids, with the following exceptions: double dotted line 5 21, double line 5 22, bold line 5 23, dotted line 5 25; dot-and-dash line 5 26.Prefix f 5 fungal; m 5 moss; e 5 EST. All sequences used and their accession numbers are presented in table 2.
by guest on August 7, 2016
http://mbe.oxfordjournals.org/
Dow
nloaded from

Phylogeny, Function, and Evolution of Cupins 603
FIG. 5.—Alignment of DNA sequences encoding the intermotif regions (IMRs) of representative germin-like proteins, grouped accordingto the length of the IMR. Alternate codons are shaded, bases showing $50% conservation are in bold, and an asterisk denotes a 100% conservedthymine in the second position.
ESTs available at the time of this analysis (data notshown) confirmed the high level of conservation withinthe cupin motifs and the IMRs of these proteins. Thecupin ESTs appeared from cDNA libraries constructedfrom a variety of plant tissues, such as leaf, shoot androot material, as well as callus tissue (giz5602575) andfloral tissues (giz4152154). Furthermore, a number ofthese ESTs originated from tissue under biotic or abioticstress, such as maize silks infected with Fusarium(giz6681825) or Medicago truncatula roots infected withPhytophthora (giz6654979). Mesembryanthemum crys-tallinum tissue subjected to high salt levels yields anumber of cupin ESTs (giz2911925, giz5917863,giz2734069, giz5443071, giz5442661, and giz4464834),as does M. truncatula tissue grown under phosphate de-ficiency (giz7675310, giz7675658, and giz7675990).These results are reminiscent of the expression of wheatGLPs during similar biotic (Schweizer, Christoffel, andDudler 1999) and abiotic stress responses (Berna andBernier 1999).
Conclusions
Cupins have been found in organisms from ther-mophilic bacteria to plants and animals that inhabit awhole spectrum of environments (Dunwell 1998a). Thewidespread occurrence of the conserved domain in pro-and eukaryotic proteins that include a variety of en-zymes and binding proteins (Dunwell, Khuri, and Gane2000) led to the conclusion that the compact beta-barrel
structure that makes up the cupin core (Gane, Dunwell,and Warwicker 1998; Woo et al. 2000) provides a stablescaffold that allows these proteins to survive and func-tion under a great variety of extreme conditions(Thompson and Eisenberg 1999) and can be functionallymodified by relatively minor changes to the active-siteregion.
An unrooted phylogeny of all the known cupin mo-tifs at the time of analysis (tables 1 and 2) gave rise toa function-based clustering (fig. 2), proving that evenwithin the six-strand cupin domain there was sufficientinformation to permit discrimination between differentclades. If this tree were to be rooted using the cupin(aae) from the thermophilic A. aeolicus (considered themost deeply branched bacterium; Deckert et al. 1998)as the outgroup, it is expected that bacterial PMIs wouldbe the most primitive cupins, with the eukaryotic GLPs,and particularly the true germins (from cereals), beingthe most advanced.
In terms of functional evolution, this means that thefirst ‘‘protocupin’’ arose as a small b-barrel protein(about 100 residues), perhaps containing iron or man-ganese (Kirschvink et al. 2000) and initially able to binda variety of simple sugars. As the environment changed,the protocupin gene was modified and diversified as theprokaryote evolved. The proteins underwent amino acidadditions and/or substitutions to bind more complexsugars and fused with other ‘‘protoproteins’’ to catalyzemore complex reactions. Throughout this evolutionary
by guest on August 7, 2016
http://mbe.oxfordjournals.org/
Dow
nloaded from

604 Khuri et al.
process, the key structural residues were conserved andthe distinctive beta-barrel structure was maintained. Atsome point, one such cupin fused to another, or wasduplicated, with the emergence of bi-cupins such as theOXDCs. It is hoped that the data set from this super-family of proteins will be of value in addressing theobjections raised against phylogenetic analyses by thosewho argue that most sequences are heavily saturatedwith respect to amino acid substitutions and that it isusually not possible to define a stable character in agiven domain (a specific conserved residue) becausethese positions have evolved too rapidly (Philippe andForterre 1999). In the cupin sequences, in contrast, thereare definite fixed residues known to have importantstructural and functional significance.
Relationship of GLPs to other Mn-SODs
Recently, two independent sources have providedevidence for a functional similarity between GLPs andthe well-characterized group of iron/manganese SODsfound in prokaryotes and in the mitochondria of eu-karyotes. First, resolution of the three-dimensionalstructure of barley germin at 1.6 A (Woo et al. 2000)revealed an active site comprising a single Mn atomliganded by the three histidines and the single glutamatewithin the two cupin motifs (fig. 1). This active site hasclose geometric similarity to that of the Mn/Fe class ofSOD, which consists of three histidines and an aspartateresidue (although the overall protein fold of these SODsis different from that of the cupins). Direct experimentalevidence for the connection comes from the finding ofSOD activity in a manganese-containing GLP from themoss B. unguiculata (Yamahara et al. 1999), from thedetection of Mn-SOD activity from a tobacco GLP(Carter and Thornburg 2000), and from the detection ofboth OXO and SOD activities in a barley germin (Wooet al. 2000).
It now seems logical to suggest that the major func-tional role for all GLPs is as a SOD enzyme that isactive at neutral pH and protects plants from oxidativestress induced by the range of biotic and abiotic stressesreferred to above. Compared with the majority of plantGLPs, the more specialized cereal germins (figs. 2 and5) are assumed to have evolved an additional OXOfunction at low pH. This activity provides the additionalbenefit of enabling detoxification of the oxalic acid pro-duced by several plant pathogens (Dunwell, Khuri, andGane 2000).
It is now thought that the ancestral form of thepreviously characterized type of microbial SOD wascambialistic (i.e., it was able to bind both Fe and Mn)and that the two isoforms (Mn form and Fe form) di-verged at a later stage of evolution, after the divergenceof the Archaea and Eubacteria. This change in metalbinding specificity has recently been related to changesin the redox potential of the surface oceans during theperiods around the intense global glaciations known as‘‘snowball earth’’ (Kirschvink et al. 2000). The enzy-matic cupin precursors may have undergone a similarevolutionary path.
As this study has shown, in this genome-sequenc-ing era, it is possible to continue to define new struc-turally related families and superfamilies of functionallydiverse proteins. It is imperative, however, that predic-tions of the enzymatic functions of proteins be supportedby biochemical evidence, although this is not always asimple issue. The identification of barley germin as anOXO (Lane et al. 1993), for example, came 80 yearsafter evidence for the enzyme was first provided (Zales-ki and Reinhard 1912), and it was nearly 10 years laterthat its second function as a SOD was discovered (Wooet al. 2000).
Acknowledgments
The authors are grateful to the Biotechnology andBiological Sciences Research Council, United Kingdom(S.K., J.M.D.), the Natural Environment Research Coun-cil, United Kingdom (F.T.B.), and Syngenta Ltd (J.M.D.)for financial support. We also thank Dr. Alastair Culhamfor valuable advice, and the two reviewers for positiveand constructive comments.
LITERATURE CITED
ARAVIND, L., and E. V. KOONIN. 1999a. Gleaning non-trivialstructural, functional and evolutionary information aboutproteins by iterative database searches. J. Mol. Biol. 287:1023–1040.
———. 1999b. DNA-binding proteins and evolution of tran-scription regulation in the archaea. Nucleic Acids Res. 27:4658–4670.
BAUMLEIN, H., H. BRAUN, I. A. KAKHOVSKAYA, and A. D.SHUTOV. 1995. Seed storage proteins of spermatophytesshare a common ancestor with desiccation proteins of fungi.J. Mol. Evol. 41:1070–1075.
BERNA, A., and F. BERNIER. 1999. Regulation by biotic andabiotic stress of a wheat germin gene encoding oxalate ox-idase, a H2O2-producing enzyme. Plant Mol. Biol. 39:539–549.
CARTER, C., and R. W. THORNBURG. 1999. Germin-like pro-teins: structure, phylogeny and function. J. Plant Biol. 42:97–108.
———. 2000. Tobacco Nectarin 1: purification and character-isation as a germin-like, manganese superoxide dismutaseimplicated in the defense of floral reproductive tissues. J.Biol. Chem. 275:36726–36733.
DECKERT, G., P. V. WARREN, T. GAASTERLAND et al. (15 co-authors). 1998. The complete genome of the hyperthermo-philic bacterium Aquifex aeolicus. Nature 392:353–358.
DOMON, J.-M., B. DUMAS, E. LAINE, Y. MEYER, A. DAVID, andH. DAVID. 1995. Three glycosylated polypeptides secretedby several embryogenic cell lines of pine show highly spe-cific serological affinity to antibodies directed against thewheat germin apoprotein monomer. Plant Physiol. 108:141–148.
DUNWELL, J. M. 1998a. Cupins: a new superfamily of func-tionally diverse proteins that include germins and plant stor-age proteins. Biotechnol. Genet. Eng. Rev. 15:1–32.
———. 1998b. Sequence analysis of the cupin gene family inSynechocystis PCC6803. Microb. Comp. Genomics 3:141–148.
DUNWELL, J. M., and P. J. GANE. 1998. Microbial relatives ofseed storage proteins; conservation of motifs in a function-
by guest on August 7, 2016
http://mbe.oxfordjournals.org/
Dow
nloaded from

Phylogeny, Function, and Evolution of Cupins 605
ally diverse superfamily of enzymes. J. Mol. Evol. 46:147–154.
DUNWELL, J. M., S. KHURI, and P. J. GANE. 2000. Microbialrelatives of the seed storage proteins of higher plants: con-servation of structure and diversification of function duringthe evolution of the cupin superfamily. Microbiol. Mol.Biol. Rev. 64:153–179.
FARRIS, J. S. 1969. A successive approximations approach tocharacter weighting. Syst. Zool. 18:374–385.
GALLEGOS, M.-T., R. SCHLEIF, A. BAIROCH, K. HOFMAN, andJ. L. RAMOS. 1997. AraC/XylS family of transcription reg-ulators. Microbiol. Mol. Biol. Rev. 61:393–410.
GANE, P. J., J. M. DUNWELL, and J. WARWICKER. 1998. Mod-elling based on the structure of vicilins predicts a histidinecluster in the active site of oxalate oxidase. J. Mol. Evol.46:488–493.
HAMEL, F., C. BRETON, and M. HOUDE. 1998. Isolation andcharacterization of wheat aluminum-regulated genes: pos-sible involvement of aluminum as a pathogenesis responseelicitor. Planta 205:531–538.
HEINTZEN, C., R. FISCHER, S. MELZER, K. KAPPELER, K. APEL,and D. STAIGER. 1994. Circadian oscillations of a transcriptencoding a germin-like protein that is associated with cellwalls in young leaves of the long-day plant Sinapis alba L.Plant Physiol. 106:905–915.
HURKMAN, W. J., and C. K. TANAKA. 1996. Effect of salt stresson germin gene expression in barley roots. Plant Physiol.110:971–977.
KESARWANI, M., M. AZAM, K. NATARAJAN, A. MEHTA, and A.DATTA. 2000. Oxalate decarboxylase from Collybia veluti-pes. Molecular cloning and its overexpression to confer re-sistance to fungal infection in transgenic tobacco and to-mato. J. Biol. Chem. 275:7230–7238.
KIRSCHVINK, J. L., E. J. GAIDOS, L. E. BERTANI, N. J. BEUKES,J. GUTZMER, L. N. MAEPA, and R. E. STEINBERGER. 2000.Paleoproterozoic snowball earth: extreme climatic and geo-chemical global change and its biological consequences.Proc. Natl. Acad. Sci. USA 97:1400–1405.
KOONIN, E. V., A. R. MUSHEGIAN, M. Y. GALPERIN, and D. R.WALKER. 1997. Comparison of archaeal and bacterial ge-nomes: computer analysis of protein sequences predictsnovel functions and suggests a chimeric origin for the Ar-chaea. Mol. Microbiol. 25:619–637.
LANE, B. G. 2000. Oxalate oxidases and differentiating surfacestructure in wheat: germins. Biochem. J. 349:309–321.
LANE, B. G., F. BERNIER, E. DRATEWKA-KOS, R. SHAFAI, T. D.KENNEDY, C. PYNE, J. R. MUNRO, T. VAUGHAN, D. WAL-TERS, and F. ALTOMARE. 1991. Homologies between mem-bers of the germin gene family in hexaploid wheat and sim-ilarities between these wheat germins and certain Physarumspherulins. J. Biol. Chem. 266:10461–10469.
LANE, B. G., J. M. DUNWELL, J. RAY, M. R. SCHMITT, and A.C. CUMING. 1993. Germin, a marker of early plant devel-opment, is an oxalate oxidase. J. Biol. Chem. 268:12239–12242.
MADDISON, W. P., and D. R. MADDISON. 1992. MacClade. Ver-sion 3.04. Analysis of phylogeny and character evolution.Sinauer, Sunderland, Mass.
NEUTELINGS, G., J. M. DOMON, N. MEMBRE, F. BERNIER, Y.MEYER, A. DAVID, and H. DAVID. 1998. Characterizationof a germin-like protein gene expressed in somatic and zy-gotic embryos of pine. Plant Mol. Biol. 38:1179–1190.
ONO, M., K. SAGE-ONO, M. INOUE, H. KAMADA, and H. HAR-ADA. 1996. Transient increase in the level of mRNA for agermin-like protein in leaves of the short-day plant Phar-bitis nil during the photoperiodic induction of flowering.Plant Cell Physiol. 37:855–861.
PHILIPPE, H., and P. FORTERRE. 1999. The rooting of the uni-versal tree of life is not reliable. J. Mol. Evol. 49:509–523.
REQUENA, L., and S. BORNEMANN. 1999. Barley (Hordeum vul-gare) oxalate oxidase is a manganese-containing enzyme.Biochem. J. 343:185–190.
SCELONGE, C. J., and D. L. BIDNEY. 1998. Gene encoding ox-alate decarboxylase from Aspergillus phoenices. Patent Ap-plication WO 98/42827.
SCHWEIZER, P., A. CHRISTOFFEL, and R. DUDLER. 1999. Tran-sient expression of members of the germin-like gene familyin epidermal cells of wheat confers disease resistance. PlantJ. 20:541–552.
SHUTOV, A. D., F. R. BLATTNER, and H. BAUMLEIN. 1999. Evo-lution of a conserved protein module from Archaea toplants. Trends Genet. 15:348–349.
SOISSON, S. M., B. MACDOUGALL-SHACKLETON, R. SCHLEIF,and C. WOLBERGER. 1997. Structural basis for ligand-reg-ulated oligomerization of AraC. Science 276:421–425.
STAIGER, D., K. APEL, and G. TREPP. 1999. The Atger3 pro-moter confers circadian clock-regulated transcription withpeak expression at the beginning of the night. Plant Mol.Biol. 40:873–882.
SWOFFORD, D. L. 1999. PAUP*: phylogenetic analysis usingparsimony (*and other methods). Version 4.0b4a. Sinauer,Sunderland, Mass.
TANNER, A., and S. BORNEMANN. 2000. Bacillus subtilis YvrKis an acid-induced oxalate decarboxylase. J. Bacteriol. 18:5271–5273.
THOMPSON, M. J., and D. EISENBERG. 1999. Transproteomicevidence of a loop-deletion mechanism for enhancing pro-tein thermostability. J. Mol. Biol. 290:595–604.
THORDAHL-CHRISTENSEN, H., Z. ZHANG, Y. WEI, and D. B.COLLINGE. 1997. Subcellular localization of H2O2 in plants:H2O2 accumulation in papillae and hypersensitive responseduring powdery-mildew interaction. Plant J. 11:1187–1194.
TRINQUIER, G., and Y. H. SANEJOUAND. 1998. Which effectiveproperty of amino acids is best preserved by the geneticcode? Protein Eng. 11:153–169.
VALLELIAN-BINDSCHEDLER, L., E. MOSINGER, J. P. METRAUX,and P. SCHWEIZER. 1998. Structure, expression and locali-zation of a germin-like protein in barley (Hordeum vulgareL.) that is insolubilized in stressed leaves. Plant Mol. Biol.37:297–308.
WOO, E.-J., J. M. DUNWELL, P. W. GOODENOUGH, A. C. MAR-VIER, and R. W. PICKERSGILL. 2000. Barley germin is amanganese containing homohexamer with oxalate oxidaseand superoxide dismutase activities. Nat. Struct. Biol. 7:1036–1040.
YAMAHARA, T., T. SHIONO, T. SUZUKI, K. TANAKA, S. TAKIO,K. SATO, S. YAMAZAKI, and T. SATOH. 1999. Isolation of agermin-like protein with manganese superoxide dismutaseactivity from cells of a moss, Barbula unguiculata. J. Biol.Chem. 274:33274–33278.
ZALESKI, W., and A. REINHARD. 1912. Uber die fermentativeOxydation der Oxalsaure. Biochem. Zeitung 33:449–455.
ELIZABETH KELLOGG, reviewing editor
Accepted December 13, 2000
by guest on August 7, 2016
http://mbe.oxfordjournals.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















