
Monumentalization and ritual landscapes at Fort Center in the Lake Okeechobee basin of South Florida

ELSEVIER Ecological Engineering 5 (1995) 183-207
ECOLOGICAL ENGINEERING
Phosphorus dynamics in selected wetlands and streams of the lake Okeechobee Basin *
K.R. Reddy *, O.A. Diaz, L.J. Scinto, M. Agami Soil and Water Science Department, 106 Newell Hall, Box 110510, Uniuersity of Florida, Institute of Food
and Agricultural Sciences, Gainesuille, FL 32611, USA
Abstract
Lake Okeechobee is becoming increasingly eutrophic, presumably due to P loading from numerous dairy operations in the Lake's northern drainage basin. Phosphorus released from this basin is transported through canals, streams, and wetlands before its discharge into the lake. This paper summarizes the results of several studies on P dynamics in wetlands and stream sediments in the Lake Okeechobee Basin with primary focus on P interaction with soil/sediment-water column and vegetation.
Stream sediments and wetland soils in the basin were characterized for labile and non-labile pools of P. The labile inorganic P (Pi) pool (KCt-extractable) accounted for 0.1 to 2.3% and 0.1 to 0.7% of the total P in sediments and wetland soils, respectively. The NaOH extractable Pi, representing the P associated with Fe and AI oxyhydroxides, was the dominant Pi in both stream sediments and wetland soils (accounting for up to 71 and 43% total P, respectively). The NaOH-P o (humic and fulvic acid associated organic P) is considered resistant to biological breakdown and accounted for 6 to 56% of total P. Stream sediments showed higher buffer intensity for P sorption than wetlands. Phosphate sorption capacity (Sma x) and buffer intensity (Kd-adsorption coefficient) were highly correlated with oxalate extractable [Fe + AI] and total organic carbon (TOC) suggesting P sorption is associated with amorphous and weakly crystalline forms of Fe and AI, and /o r complexed with organic matter.
Phosphorus assimilation in vegetation was found to be short-term and dependent upon plant species, P loading, and wetland hydrology. Decomposition of detrital tissue resulted in rapid release of P into the water column. Phosphorus release was rapid during decomposi- tion of floating macrophytes, as compared to herbaceous vegetation.
Paper presented at the workshop on Phosphorus Behavior in the Okeechobee Basin, sponsored by the South Florida Water Management Distirct and the University of Florida, Institute of Food and Agricultural Sciences.
" Corresponding author.
0925-8574/95/$09.50 © 1995 Elsevier Science B.V. All rights reserved SSD[ 0925-8574(95)00024-0

184 K.R. Reddy et al. / Ecological Engineering 5 (1995) 183-207
Phosphorus retent ion coefficients were positively correlated with oxalate extractable Fe and AI content of soils and sediments. The average EPC w (threshold P concentrat ion in the water column where P retention = P release) for stream sediments was 0.10 mg P 1-1 and 0.42 mg P 1-1 for wetland soils. The stability of the P retained was regulated by the physico-chemical properties of the soils and sediments.
Keywords: Phosphorus dynamics; Wetlands; Streams; Lake Okeechobee
1. In troduct ion
Lake Okeechobee (surface area 1890 km 2) is the fourth largest lake in the United States of America. This shallow sub-tropical lake may be moving from a naturally eutrophic state to a hypereutrophic state as a result of increased phosphorus (P) loading from surrounding watersheds. For example, the Taylor Creek/Nubbin Slough watershed which provides 5% of the water input supplied approximately 28% of the total P, whereas the Kissimmee River Basin which provides 30% of the water input contributed 20% of the P loading (Federico et al., 1981a). The high P loading from these two basins was attributed to numerous dairies located in this region. Most dairy sites in the Okeechobee Basin are located on Myakka (a sandy, siliceous, hyperthermic Aeric Haplaquod) or Immokalee (a sandy, siliceous, hyperthermic Arenic Haplaquod) fine sand soils (Graetz and Nair, 1995). The poor P retention capacity and high water permeability of these sandy surface soils reduces the amount of particulate-bound P to negligible proportions (Knisel et al., 1985; Allen, 1987). The inefficient P removal by these sandy soils are caused by: low P retention capacities of the A and the highly eluted E horizons, and a lack of deep percolation of soluble P into Al rich lower spodic (Bh) horizons (Yuan, 1965; Allen, 1987). These conditions result in high soluble P concentrations in the drainage discharged from these basins. Therefore, soluble P is transported via overland flow or lateral seepage along eluted horizons above the spodic layer (Campbell et al., 1995). The route of P transport to the lake is by surface and subsurface flow from uplands to adjacent wetlands and streams, which feed into major inflows and finally discharge into the lake.
About 400000 ha of wetlands are located in the Okeeehobee Basin, with > 80% in the Lower Kissimmee River Basin (Table 1). In addition, these wetlands are connected by canals and streams which transport water to Lake Okeechobee.
Table 1 Wetland area in major drainage basin of Lake Okeechobee (Federico et al., 1981a)
Basin Area (ha) % Land use in respective basin
Lower Kissimmee River 320355 18 Taylor Creek/Nubbin Slough 8955 15 Fisheating Creek 48 963 42 Harney Pond/Indian Prairie 32827 36 Lake Istokpoga 5024 4

K.R. Reddy et al. / Ecological Engineering 5 (1995) 183-207 185
Soil/sediment
&,o%.~=~, 7 In/out |
-91 I1,.-
Rainfall ~ Atmosphere n
\ it I ! A :
~ . -! ____.-
V V
l U p l a n d , i , . . . .
Fig. 1. Schematic showing phosphorus exchange processes in upland-wetland-stream system.
Thus, P removal by wetlands in the basins has been suggested as a means to reduce P loading to Lake Okeechobee (Federico et al., 1978; Davis, 1982; Goldstein, 1982). However, wetlands were shown to function as sinks or sources for P depending on water column and sediment physico-chemical properties (Richard- son, 1985). To evaluate the potential of wetlands and stream sediments for P assimilation, it is critical that we determine the roles of various components of these systems (soils/sediments, vegetation, water) in regulating the water quality. Thus, the key cluestions often asked by managers and regulatory agencies.involved in water management are: (1) how much P can be stored in wetland soils and stream sediments? (2) how stable is the stored P and under what conditions will it be released back into the water column? (3) are wetlands and stream sediments sources or sinks for P? and (4) what is the long-term P assimilatory capacity of wetlands and stream sediments? These questions are very complex and involve a comprehensive study of biogeochemical processes regulating P retention/release by soils or sediments, and vegetation of the ecosystem. These processes include: adsorption/desorption reactions, precipitation, mineralization, exchange processes between soil/sediment and the overlying water, and plant uptake and release (Fig. 1).
In this paper, we examined P behavior in soil/sediment-water-plant compo- ,nents of selected wetlands and streams in the Lake Okeechobee drainage basin. In

186 K.R. Reddy et al. /Ecological Engineering 5 ~1995) 183-207
Table 2 Flow weighted mean phosphorus concentration for tributary inflows (Federico et al., t981b and SFWMD unpublished results)
Basin 1973-1979 1991 1992
SRP Total P SRP Total P
mg/1- t
Kissimmee River (S-65E) 0.092 0.107 Taylor Creek (S-133) 0.258 0.341 0.327 0.437 Nubbin Slough (S-191) 0.749 0.906 0.613 0.753 Fisheating Creek 0.161 0.235 0.104 0.196 Harney Pond Canal (S-71) 0.185 0.261 0.186 0.238 Indian Prairie Canal (S-72) 0.139 0.217 0.160 0.253 Rainfall 0.041 0.061 0.05
particular, we summarize the results of a comprehensive project on the biogeo- chemistry of P in these systems. Specific topics include: (1) forms of labile and non-labile pools of P in soils and sediments, (2) P sorption capacity of selected stream sediments and adjacent wetlands, (3) mobility of P in sediment, soil and the overlying water column, (4) chemical precipitation of P in the water column, (5) P storage and release capacity of selected aquatic macrophytes in streams and adjacent wetlands and (6) long-term P storage in wetlands and stream sediments. Detailed results of these studies are presented in Reddy et al. (1992a); Reddy et al. (1992b); Reddy et al. (1993); and Diaz et al. (1995).
2. Site description
The Lake Okeechobee drainage basin covers more than 4600 square miles. Inflows into the lake include: rainfall, the Kissimmee River, Fisheating Creek, Taylor Creek/Nubbin Slough, Harney Pond and Indian Prairie Basins, and the Everglades Agricultural Area. The total P concentration of the major inflows has changed little during the past 20 years (Table 2) with a major portion of the inflow water P (64 to 83% of total P) in soluble form. The P concentration of Taylor Creek/Nubbin Slough inflow was higher than other inflows. Approximately 50% of the total P loading to the lake is from Kissimmee River and Taylor Creek/Nub- bin Slough inflows (Federico et al., 1981a).
The sampling sites (Fig. 2; Table 3) used in the study were selected in Taylor Creek/Nubbin Slough Basin, Lower Kissimmee River Basin (S-65D, S-65E, S-154 Basins), Istokpoga Basin (Bates Canal), Fisheating Creek Basin, and Harney Pond/Indian ~ Prairie Basin (C-41 Basin). At each site, stream sediment and adjacent wetland soil and overlying water samples were obtained. For the purpose of this paper, the physico-chemical characteristics of surface soil or sediment to a depth of 15 cm were used. Selected physico-chemical characteristics of the stream
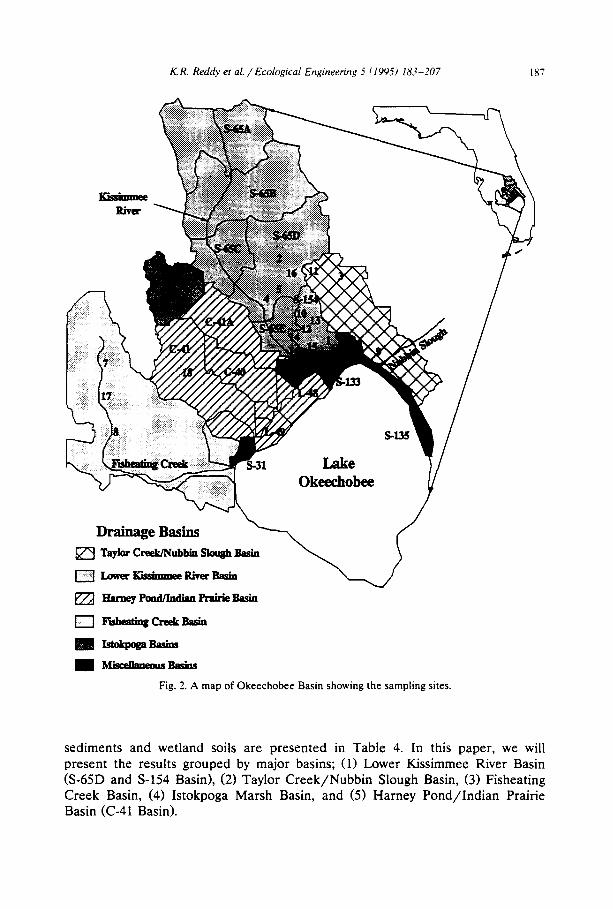
K.R. Reddy et al . / Ecological Engineering 5 (1995) 183-207 187
River
i ? ̧ ¸ i i i ! ! i i : . : . : :
I,~ke Okeechobee
S-135
Tabor Creek/Nubbln Slough ~
Im t~okpo~ e,,s~
Fig. 2. A map of Okeechobee Basin showing the sampling sites.
sediments and wetland soils are presented in Table 4. In this paper, we will present the results grouped by major basins; (1) Lower Kissimmee River Basin (S-65D and S-154 Basin), (2) Taylor Creek/Nubbin Slough Basin, (3) Fisheating Creek Basin, (4) Istokpoga Marsh Basin, and (5) Harney Pond/Indian Prairie Basin (C-41 Basin).

188 K.R. Reddy et al. / Ecological Engineering 5 (1995) 183-207
3. Forms of labile and non-labile pools of phosphorus
To determine the role of wetland soils and stream sediments on water quality in the Lake Okeechobee Basin, it is important to develop a fundamental understand- ing of the form of P in the system. Chemical fractionation was used to characterize selected wetland soils and sediments in the basin with respect to forms of inorganic and organic P (labile and non-labile pools). This method involves sequential extraction of soils or sediments with KC1-NaOH-HCI, with each chemical removing discrete forms of P (Chang and Jackson, 1957; Hieltjes and Lijklema, 1980; Reddy et al., 1992a). Major pools of Pi (inorganic P) identified in this scheme are: (1) loosely adsorbed P, (2) Fe- and Al-bound P, and (3) Ca- and Mg-bound P. The P
Table 3 Selected stream/wetland systems from the Okeechobee Basin used in the study
Sampling sites Drainage basins Number of samples
Lower Kissimmee River Basin Stream Wetland Dry Lake (1) S-154 5 15 Larson Dixie (10) S-154 1 1 Turkey Slough (12) S-154 1 1 Popash Slough 0 S-154 1 1 Pelaez (13) S-154 1 1 Yates Marsh (14) S-154 1 1 W.F. Rucks (2) S-65D 5 14 Chandler Slough (4) S-65D 4 4 Cypress Slough (5) S-65D 4 4 Fish Slough (16) S-65D 1 1
Taylor Creek / Nubbin Slough Basin Otter Creek (3) 8 Nubbin Slough (9) 1 1 McArthur #5 (11) 1 1
Harney Pond/Indian Prairie Basin Archbold Sawgrass (18) C-41 1 1
Bates Canal 1 (6) 4 Bates Canal 2 (6) 4 Bates Canal 3 (6) 4 Organic soil (drainedX6)
Fisheating Creek Basin FEC 1 (7) 4 FEC 2 (8) 4 4 FEC 3 (17) 1 1
Numbers in parenthesis indicate locations on Lake Okeechobee Basin map (Fig. 2). Details of the studies are presented by Reddy et al. (1992a) for Dry Lake and W.F. Rucks; Reddy et al. (1992b) for Otter Creek and Reddy et al. (1993) for the remaining sites.

Tab
le 4
S
elec
ted
phys
ical
and
che
mic
al p
rope
rtie
s o
f st
ream
sed
imen
ts a
nd w
etla
nd s
oils
fro
m t
he O
kee
cho
bee
Bas
in
Sam
plin
g si
tes
pH
TO
C (
mg
g-
i)
Bul
k de
nsit
y (g
cm -
3)
Am
mo
niu
m o
xala
te
Tot
al P
F
e A
I m
g kg
- I
KC
I
Ca
Mg
t~
Stre
am s
edim
ents
L
ower
Kis
sim
mee
Riv
er B
asin
S-
154
Bas
in
6.61
10
5 0.
611
S-65
D B
asin
5.
79
9 1.
110
Tay
lor
Cre
ek/N
ub
bin
Slo
ugh
Bas
in
6.25
39
0.
752
Har
ney
Po
nd
/In
dia
n P
rair
ie B
asin
(C
-41)
5.
54
99
0.53
4 F
ishe
atin
g C
reek
Bas
in
6.49
52
0.
708
lsto
kpog
a B
asin
6.
65
113
0.60
1
Wet
land
soi
ls
Low
er K
issi
mm
ee R
iver
Bas
in
S-15
4 B
asin
6.
21
ll0
0.
460
S-65
D B
asin
5.
37
151
0.52
2 T
aylo
r C
reek
/Nu
bb
in S
loug
h B
asin
6.
61
101
0.56
6 H
arne
y P
on
d/I
nd
ian
Pra
irie
Bas
in (
C-4
11
4.82
10
5 0.
782
Fis
heat
ing
Cre
ek B
asin
5.
77
85
0.89
8 ls
tokp
oga
Bas
in
3.43
52
8 0.
231
502
2544
56
9 33
19
604
72
685
172
477
32
469
776
1187
14
38
133
387
1699
89
3 25
50
326
418
1501
67
6 22
32
380
211
1879
97
9 31
29
4111
615
2616
69
5 38
09
463
702
2394
18
55
4471
36
8 10
79
2096
32
1~
51~0
31
2 33
8 10
73
1003
23
03
378
304
890
342
763
218
367
1297
17
47
5688
50
8
e., I -.4

190 K.R. Reddy et al . / Ecological Engineering 5 (1995) 183-207
not extracted was assumed to be highly resistant residual organic P and highly insoluble inorganic P.
Phosphorus extracted with 1M KCI represented the labile pool of P~ (inorganic P). This P pool is important in plant growth and controlling the P concentration of the overlying water column. In stream sediments, labile P~ was in the range of 0.I to 2.3% of total P, with higher ranges observed in Dry Lake and Chandler Slough sediments (Fig. 3a). In wetland soils, labile P~ was in the range of 0.1 to 1.1% of
Lower Kissimmee River Basin (a) $154 Basin
Dry Lakes Stream
2.1%
Total P = 877 mg/kg
Dry Lakes Wetland
~ 0.2%
To ta l P = 320 m g / k g
Dry ! ~tr~ Upland
Total P = 101 mg/kg
W.F. Rucks Stream
~ 1.0%
Total P = 93 mg/kg
$65-D Basin
W.F. Rucks Wetland
0.2%
Total P = 294 mg/kg
W.F. Pucks Upland
0.1%
4.1%
Total P -- 357 mg/kg
Chandler Slough Stream
2.3%
3.9% Total P = 32 mg/kg
Cypress Slough Stream
2.6% ~ I , 4 %
Total P -- 45 mg/kg
Chandler Slough Wetland
1 . 0 ~ 0.5%
Total p = 987 mg/kg
Cypress Slough Wetland
0.7%
0.7% Total P = 277 mg/kg
KCI-Pi
NaOH-Pi
NaOH-Po
HCI-Pi
Residual Po
Fig. 3. (a) Labile and nonlabile pools of soil and sediment phosphorus in Lower Kissimmee River Basin. KCI-Pi = adsorbed P potentially bioavailable; NaOH-Pi = Fe/Al-bound inorganic P; NaOH-Po = organic P associated with humic and fulvic acids; HCI-Pi ~=Ca/Mg-bound inorganic P; Residual Po = resistant organic P.

K.R. Reddy et al. / Ecological Engineering 5 (1995) 183-207
(b) Taylor Creek/
Nubbin Slough Basin Istokpoga Basin
Otter Creek Stream
1.9% 0.3%
Total P = 940 mg/kg
Bates Canal 1
~ 0.4%
Total P = 139 mg/kg
Fisheating Creek Basra
I91
F i s h . r i n g Creek I Stream
6.3% ~0.1%
Total P = 347 mg/kg
Otter Creek Wetland
1 " 3 % ~ 0 I%
Total P = 444 mg/kg
Bates Canal 2
Total P = 113 mg/kg
Fisheating Creek 2 Stream
0.3%
1.7% Total P = 255 mg/kg
NaOH-Pi
~'] NaOH-Po
HCI-Pi
V-q Residual Po
lllllllll~a~8 Canal 3
Total P = 68 mg/kg
Organic Son (drainea3
Fisheating Creek 2 Wetland
@ 1.1~
0.4%
Total P = 190 mg/kg
8.9% 1.5% Total P = 351 mg/kg
Fig. 3. (b) Labile and nonlabile pools of soil and sediment phosphorus in Taylor Creek/Nubbin Slough, Istokpoga and Fisheating Creek Basins. KCI-Pi = adsorbed P potentially bioavailable: NaOH-Pi = Fe/Al-bound inorganic P; NaOH-Po = organic P associated with humic and fulvic acids; HCI-Pi = Ca/Mg-bound inorganic P; Residual Po = resistant organic P.
total P with the exception of a drained wetland in Istokpoga Marsh (Fig. 3b), which had labile Pi pool of 9.5% of total P. The labile Pi readily responds to external P ioadings, as was measured in constructed wetlands (Gale et al., 1994) and natural wetlands (Reddy et al., 1990).
Phosphorus extracted with NaOH represents the P associated with amorphous oxyhydroxide surfaces and crystalline Fe and AI oxides. Phosphorus associated with oxyhydroxides is readily desorbed under most conditions, but the P associated with crystalline Fe and AI oxides is desorbed only under extended waterlogged conditions. The NaOH-P i was the dominant pool of inorganic P in both stream

192 K.R. Reddy et al. /Ecological Engineering 5 (1995) 183-207
sediment and wetland soils, indicating the significance of Fe/Al-bound P. The NaOH-P~ was in the range of 20 to 71% of total P in stream sediments and 17 to 43% of total P in wetland soils. The NaOH-P i was strongly correlated (p < 0.01) with oxalate extractable P and associated cations, Fe and AI. This relationship strongly suggests that the P is bound to Fe/Al-components of the sediments and wetland soils.
The P~ extracted with 0.5 M HCI during sequential extractions of soils and sediments, represents the P associated with Ca and Mg minerals. The HCI-P i pool was low in all soils and sediments, with the exception of stream sediments at Dry Lake site (50% of total P) and 13% of total P in W.F. Rucks wetland. High P and Ca loading to this stream probably resulted in elevated levels of HCI-P i. Under most natural conditions, Ca-bound P is essentially unavailable to biological assimi- lation (Sonzogni et al., 1982). However, in many wetlands and stream sediments of the Okeechobee Basin, the HCI-P~ pool is not significant because of low pH conditions.
The alkaline extraction used in the fractionation scheme also extracted a large proportion of organic P essentially consisting of P associated with humic and fulvic acids. The fulvic acid bound P (NaOH-P o) is biologically reactive and can be hydrolyzed to bioavailable forms. The NaOH-P o represented 6 to 57% of total P (Fig. 3a,b). The residual P not extracted either by KCI, NaOH and HCI is considered to be highly resistant and biologically unavailable. This pool of P was found to be in high proportions (43-51%) of total P in Chandler Slough, Dry Lake upland, and Fisheating Creek sediments.
4. Phosphorus sorption characteristics of sediments and wetland soils
The ability of stream sediments and adjacent wetlands to retain P depends on the physico-chemical characteristics of the sediment or the wetland soil. It has been shown that inorganic P added at concentrations considerably greater than
Water
Soil/S~hn~
Ps-,~-~Pads"
g3
i Smax ~ slope - KD
Phosphorus in Soil/Sediment Pore water
Fig. 4. Schematic of phosphorus adsorption isotherm for soils and sediments. Kd = Phosphorus sorption buffer intensity or adsorption coefficient; Sma x = phosphorus sorption maximum; S o = native P present in sorbed phase; EPC o = equilibrium P concentration in sediments or soils at which point adsorption equals desorption.

Tab
le 5
P
hosp
horu
s so
rpti
on c
hara
cter
isti
cs o
f se
lect
ed s
trea
m s
edim
ents
and
wet
land
soi
l (0
-15
cm d
epth
) fr
om t
he O
kee
cho
bee
Bas
in
....
Sam
plin
g si
tes
Kd
(I k
g-
i)
S o
(rag
kg
- l)
E
PC
o (
rag
I- l
) Sm
a ~ (r
ag k
g-
l)
k (I
rag
- I
p)
P ~
satu
rati
on (
% 3
",,,a
~)
~.
Stre
am s
edim
ents
L
ower
Kis
sim
mee
Riv
er B
asin
~"
S-
154
Bas
in
8.3
32.0
4.
76
162
0.05
3 20
~
. S
-65D
Bas
in
17.4
5.
0 1.
15
95
0.22
2 5.
3 T
aylo
r C
reek
/Nu
bb
in S
loug
h B
asin
34
.7
2.4
0.08
11
7 2.
1 F
ishe
atin
g C
reek
Bas
in
217.
0 8.
3 0.
04
575
0.52
5 1.
4 ~'
1 s
tokp
oga
Bas
in
543.
5 2.
1 0.
03
134
1.1
1.6
Wet
land
soi
ls
Low
er K
issi
mm
ee R
iver
Bas
in
~t"
S-15
4 B
asin
49
.4
10.5
0.
36
297
0.14
3 3.
5 o~
S
-65D
Bas
in
38.0
14
.8
1.28
41
8 0.
111
3.5
Fis
heat
ing
Cre
ek B
asin
25
.8
0.2
0.03
98
0.
233
0.2
Isto
kpog
a B
asin
6.
3 57
. I
9.06
28
0.
473
204
Kd
= P
sorp
tion
coe
ffic
ient
or
buff
er i
nten
sity
; S
O =
P
pres
ent
in s
orbe
d ph
ase
un
der
am
bien
t co
ndit
ion;
Sm
~, =
P s
orpt
ion
max
imum
; k
= a
cons
lant
j
rela
ted
to b
ondi
ng e
nerg
y.
Per
cent
P s
atur
atio
n =
(So
/S,,
~)
100.

194 K.R. Reddy et al. / Ecological Engineering 5 (1995) 183-207
those present in the porewater of sediments or wetland soils is retained by oxides and hydroxyoxides of iron and aluminum, and calcium carbonate, while at low P loadings, wet land/s t ream sediments were found to release rather than retain P (Khalid et al., 1977; Berkheiser et al., 1980; Bostrom et al., 1982; Logan, 1982; Sonzogni et al., 1982; Froelich, 1988).
A series of batch incubation studies were conducted on selected stream sedi- ments and wetland soils obtained in the Okeechobee Basin to demonstrate their capacity to retain or release P. Details of methodology and results are presented by Reddy et al., 1992a, 1993. These studies provided information on (1) equilib- rium P concentration (EPC), (2) P adsorption coefficients, (3) P adsorption maximum, and (4) the relationship between P sorption parameters and selected soi l /sediment physico-chemical properties. Results are summarized in Table 5.
4.1. Equilibrium phosphorus concentration
Equilibrium P concentration (EPCo) is the P in soil porewater in equilibrium with the P sorbed on the solid surface (Fig. 4). EPC o is referred to as the concentration at which adsorption by the solid phase equals desorption (x-axis intercept of typical adsorption isotherms). At this point, the solids exhibit maxi- mum capacity for buffering porewater P (Froelich, 1988). The EPC o values determined by adsorption isotherms have been used to determine the direction of P release between suspended sediments and the water column (Logan and McLean, 1973).
High EPC 0 values of stream sediments in the Lower Kissimmee River Basin is a reflection of poor buffer intensity as compared to the sediments in other basins (Table 5). It should be noted that the EPC 0 values only indicate whether adsorp- tion or desorption occurs in the sediments or soils, when added P alters the concentration in the porewater (Fig. 4). However, EPC 0 does not indicate the direction of P flux, which is regulated by the concentration gradients across the sediment/soil-f loodwater interface.
4.2. Phosphate sorption maximum (S,,ax)
Phosphate sorption by stream sediments and wetland soils has been described with a Langmuir isotherm (Bache and Williams, 1971):
S = S m a x k C / 1 + k C (1)
Where: Sm~ x = P sorption maximum (mg kg- t ) ; k = a constant related to bonding energy (1 mg-L); C = P in solution (mg I- l) ; S = P sorbed on solid surface. Distinct differences were observed in Sm~ , among different sites (Table 5). It should be noted that the Sma ~ estimate assumes that P in streams and wetlands is in direct contact with the sediments (a completely mixed system). Under field conditions however, diffusion of P from floodwater to underlying sediment or soil regulates the amount of P retained in the system.

K_R. Reddy et al. /Ecological Engtneenng 5 (1995) 183-207 105
In our batch incubation studies, when soils/sediments were equilibrated with high levels of P (1000 mg P kg-t added) it was assumed that all adsorption sites were saturated, thus providing a measure of sorption maximum. Except for one sampling site, all locations showed a highly significant relationship (p <0.01) between Sm~ x and P sorbed at 1000 mg P kg -1 added. S ~ , values were 1.21 times the values obtained at P addition of 1000 mg P kg- ~. This factor was subsequently used to estimate Sm~, for additional sites in the basin using the data based on single point isotherm (sorption measured at 1000 mg P kg-~).
The values of S O represent P in the adsorbed phase under ambient conditions (Table 5). Except for Dry Lake site, the S O represents only 1.4 to 5% of the Sm~,, suggesting that > 95% of the sorption sites are available for additional P sorption. Heavy P loading at the Dry Lake site resulted in saturation of 20% of the sorption sites in the sediments. The S O of wetland sites accounted for 0.2 to 3.5% of the Sm~ x, except for a drained wetland in Istokpoga marsh, which was completely saturated with P (Sma x -- So).
4.3. Buffer intensity of phosphorus sorption (Kd)
At low equilibrium P concentrations (< 10 mg P l-t), P sorption followed a linear relationship, under both aerobic and anaerobic conditions.
S = Kd C - S O (2)
Where: Kd = buffer intensity for P sorption (1 kg-~); C = P in solution (mg l-t); S O = P sorbed under ambient conditions. At EPCo, So = Kd C. The value So represents the amount of P in the labile pool which can be potentially desorbable, and significantly correlated with the KCl-extractable P. For stream sediments S O accounts for 1.6 to 25% of sorption sites (expressed as % of Sma~), while the range was 1 to 12% for wetland soils.
The stream sediments in the Lower Kissimmee River Basin generally have low buffer intensity for P retention, as indicated by low Kd values (Table 5). In the same basin, wetland soils showed higher buffer power with Kd values in the range of 5 to 107 1 kg- ~, as compared to 0.6 to 17 1 kg- t for stream sediments (except for Cypress Slough stream sediments, 86 I kg-~). However, canal sediments in Istok- poga Marsh have the highest buffer intensity, followed by sediments from Fisheat- ing Creek, while the wetlands in these basins showed much lower buffer intensity for P retention. Low Kd values of stream sediments in the lower Kissimmee River Basin suggest poor P binding capacity, and potential of these sediments to readily desorb P into the water column.
4.4. Relationship between soil properties and phosphorus sorption
Significant correlations were observed between P sorption maximum (Sm~ x) and soil properties such as extractable Fe, A1 and Ca, and TOC (p < 0.01). The most striking relationship (r--0.89, n = 115) was observed between Sma x and oxalate extractable [Fe + AI]. Strong correlation suggests that P sorption in these soils and

196 K.R. Reddy et al. /Ecological Engmeering 5 (1995) 183-207
sediments is associated with amorphous and some poorly crystalline forms of Fe and AI. Correlation of Sm~ , with TOC suggests that organic matter can also play a major role in P sorption (p < 0.01). It has been shown that Fe and AI complexed with organic matter may be responsible for P sorption, suggesting an indirect effect of organic matter (Syers et al., 1973; Traina et al., 1986). Several studies have reported the significance of Fe and AI in P sorption by sediments and wetland soils (Harter, 1968; Shukla et al., 1971; Richardson, 1985). Reduction of ferric oxyhy- droxide to more soluble ferrous forms under reduced conditions was shown to increase overall P sorption relative to oxidized conditions (Patrick and Khalid, 1974). These results suggest .that oxidation-reduction changes in wetland soils can significantly alter P solubility.
Phosphate sorption maximum (Sma x) of stream sediments can be adequately predicted by measuring oxalate extractable AI, with about 91% of the variability being accounted for by this parameter. However, for wetland soils, oxalate ex- tractable [Fe + AI] and TOC explained only 53% of the variability. When both stream sediments and wetlands were included in the regression, exchangeable Mg, oxalate extractable AI and TOC explained 71% of the variability in the prediction of Sma x.
The time required to saturate sorption sites depends on P loading, hydraulic retention time, and vegetation. Calculation of maximum retention capacity based on isotherms usually underestimates potential P sorption by soils under field conditions (Beek and de Haan, 1974; Sawhney and Hill, 1975). Under field conditions, fluctuating hydrologic conditions can change the redox potential of the soil, thus affecting P retention and release. Similarly, soil redox potential can also be influenced by organic matter inputs, creating greater oxygen demand, thus affecting P solubility.
The sorption capacities of soils and sediments measured in this study are based on soil continuously mixed with P-enriched water. However, P retention by stream sediments and wetlands under field conditions is influenced by P diffusion from the water column to the sediment, P concentration of the interstitial water and physico-chemical characteristics of the sediment at the sediment-water interface. The sediment or soil sorption data can not be readily used to predict the P assimilatory capacity in a dynamic wetland-stream system, unless consideration is given to P concentration of the overlying water, water depth, biochemical oxygen demand of the water, and associated transport processes.
5. Chemical precipitation of phosphorus in the water column
In the water column of many streams in the basin, it is likely that the increase in photosynthetic activity of algae and other aquatic vegetation can lower dissolved CO 2 in the water, resulting in high pH levels. These high pH values associated with high Ca levels can potentially precipitate P as Ca-phosphate minerals. Batch incubation studies using stirred reactors were conducted to determine chemical precipitation in selected stream waters of the Okeechobee Basin (Diaz et al.,

K.R. Reddy et al. /Ecological Engineering 5 (1995) 183-207 197
1995). Under ambient conditions, P solubility in Otter Creek water was slightly decreased by changes in dissolved CO 2. The low Ca content of this water and the changes in pH were not high enough to effectively decrease P solubility. Effective P removal from surface waters due to precipitation requires Ca concentrations of > 100 mg l - t and pH > 8.0 (Diaz et al., 1995). Increases in Ca concentrations can be observed in the water column of streams (Dry Lake site) receiving dairy effluents. With Ca concentrations above 100 mg 1-1, more than 60% of the SRP concentrations in the water can be precipitated with a pH 8.0 (Diaz et al., 1995). These reductions in P solubility as pH and Ca levels were increased suggest that some calcium phosphate mineral formation occurred during the study.
Calcium phosphate solid phases formed in the Otter Creek and Dry Lake water samples under high pH conditions, rapidly dissolved when water pH decreased to 7 as a result in increases in dissolved CO 2, about 75% and > 90% of the precipi- tated P was dissolved in Dry Lake and Otter Creek water samples, respectively. Except in highly P loaded areas (such as Dry Lake site), it is unlikely that Ca induced precipitation plays a significant role in decreasing soluble P concentration in streams of the Okeechobee Basin.
6. Phosphorus storage in aquatic vegetation
The P assimilative capacity of aquatic macrophytes is dependent upon their growth rates, composition of water, water depth, sediment characteristics, oxygen transfer capability of the plants into the root-zone, biochemical and physico-chem- ical processes functioning at the root-water-sediment interface, plant density per unit area, plant harvesting and climate (Reddy and DeBusk, 1987). The potential rate of P uptake by a plant is limited by its net productivity (growth rate) and the concentration of nutrients in the plant tissue. Phosphorus storage is similarly dependent on plant tissue concentrations, and also on the ultimate potential for biomass accumulation, that is, the maximum standing crop. In addition to plant assimilation, P removal in streams and wetlands containing plants is also affected by a number of biological, physical and chemical processes functioning in the water, sediment and rhizosphere (Reddy, 1983). Plant assimilation of P accounted for 30 to 80% of the total P removal, suggesting the importance of physico-chem- ical processes in P retention (Reddy and DeBusk, 1987).
Field studies were conducted at Dry Lake and W.F. Rucks sites to determine the potential of selected aquatic vegetation in retention and release of P (Reddy et al., 1992a). Results showed that aquatic macrophytes present in stream and adjacent wetlands of the Okeechobee Basin, function as temporary storage units for P. In general, plant tissue N and P were high in plants as compared to adjacent upland (the N/P ratio was in the range of 4 to 6), suggesting that neither nutrient is limiting the growth of plants in wetlands and streams.
At the Dry Lake site, P storage in standing crop of aquatic macrophytes was in the range of 3 to 43 kg ha-t . The wide range in P storage was due to diversity in the plant community. In the stream, Hydrocotyle showed maximum storage of 43

198 K.R. Reddy et al. / Ecological Engineering 5 (1995) 183-207
kg P ha -t. Polygonum, a common wetland plant at Dry Lake site, showed P assimilatory capacity of 8 to 23 kg P ha-t . Panicum repens, another wetland plant, showed P storage capacity of 9 to 29 kg P ha-t . Generally, aquatic plants at the Rucks site showed lower P storage, reflecting a lower P loading to the system. Pontederia and Juncus exhibited P storage capacity of 5 to 9 kg P ha- t and 1 to 7 kg P ha-~, respectively. Stored P in floating aquatic plants is short-term because of rapid turnover. If the plants are not harvested, the stored nutrients can be rapidly released into the water column during decomposition of detrital tissue. On the other hand, emergent macrophytes such as Typha, Juncus, and Pontedaria have more supportive tissue than floating plants and thus provide greater potential for P storage (Reddy and DeBusk, 1987). Most of the emergent macrophytes were present in the wetland locations of Dry Lake and W.F. Rucks sites, while the majority of water flow was in the streams, thus nutrients discharged may not be in the reach of many emergent macrophytes for effective removal. The forested areas at Rucks wetland site provide long-term storage of nutrients in woody plants, but the uptake rates of these trees are extremely slow.
In streams and wetlands dominated by aquatic vegetation, detrital tissue is generated as a result of such factors as aging of plants, over-crowding, pest damage, and frost damage during winter months. Under all these conditions, detritus is generated in the photic zone of the macrophyte stands and deposited on the soil surface (Fenchel and Jorgensen, 1977). In situ decomposition studies (Reddy et al., 1992a) have indicated that floating macrophytes decompose faster than emergent macrophytes, suggesting that Pistia and Hydrocotyle are poor sinks for P storage (Fig. 5). For the plant species evaluated, the time required to decompose 50% of plant tissues were 17 days for Pistia and Hydrocotyle, 73 days
100,
8 0 D r y Lake - ; ~ a m P u c k s - s t r e a m
60 • Hy(~'0cotyle ,, Hydroco~le ._=
~ :o.~.~_ : ~ ~ _ _ . _ _ _ : _
e ~ _
I00,
20' "N ~onum Rur.,k.s -wetland "~
o o to 20 30 400 tO 20 30 40
Time, weeks
Fig. 5. Phosphorus release during decomposition of detrital plant tissue of selected aquatic macrophytes (Reddy et al., 1992a).

I(R. Reddy et al./ Ecological Engineering 5 (1995) 183-207 199
for Pontederia, 88 days of Polygonum, 143 days for Panicum, 155 days for Juncus and 1940 days for oak leaves. Phosphorus release from decomposing tissues was rapid, with about 80% of P released in 15 days for Pistia and Hydroco~le. However, P release from the wetland plants Pontederia and Polygonum was initially slower but after 24 weeks the TP remaining was 33 and 8%, respectively. Highly significant correlations were observed between TP loss and dry weight loss (p < 0.01). Phosphorus release from aquatic vegetation was in the order of Pistia and Hydrocotyle > Polygonum > Pontederia > Panicum > Juncus > oak leaves. Rate of decomposition was found to decrease exponentially with increasing C / N and C / P ratios of plant tissue. Other researchers have shown C / N ratio as the major factor regulating the decomposition of aquatic plant tissue (Godshalk and Wetzel, 1978; Ogwada et al., 1984). Based on results of our field studies, it can be concluded that aquatic macrophytes present in the Dry Lake and Rucks sites may not be functioning as a net sink for P; any P stored is transitional and eventually released into the system.
7. Phosphorus exchange between soil and the overlying water column
The direction of P flux across the soil/sediment-floodwater interface is regu- lated by the P concentration gradients across this interface, floodwater pH, sorption/precipitation reactions at the soil/sediment-water interface, uptake by algae and aquatic vegetation, physico-chemicai properties of soils and sediments and bioturbation at the interface (Bostrom et al., 1982; Logan, 1982; Froelich, 1988).
In wetland soils and stream sediments, the soil/sediment-floodwater interface is usually oxidized and the thickness of this oxidized layer is dependent on oxygen diffusion and the oxygen demand (Howeler and Bouldin, 1971). This zone can function as a potential sink by immobilizing P into insoluble ferric phosphate, as well as uptake and storage of P into bacterial biomass (Mortimer, 1971; Gachter et al., 1988). Often, it is assumed that oxic conditions completely prevent release of P from sediments, however, mass balance studies have shown that P release can be substantial from sediments to well aerated waters (Ryding and Forsberg, 1977). To demonstrate the role of sediments and soils in regulating the overlying water quality, we have conducted several studies to determine: (1) the P exchange rates across the soil/sediment interface, (2) bulk P assimilation capacity in wetland, stream and canal sediments, and (3) effect of soil physico-chemical properties on P release/retention capacity (Reddy et al., 1992a,1993). Intact soil/sediment cores were obtained from selected sites in Lower Kissimmee River Basin (S-154, S-65D), Taylor Creek/Nubbin Slough, Istokpoga Basin and Fisheating Creek Basin. Phos- phorus retention/release rates were measured at various P loading rates by monitoring the changes in SRP concentration of the water column.
Phosphorus retention capacity of stream sediments and wetland soils was variable and regulated by their physico-chemical properties. Phosphorus reten-

200 K.R. Reddy et al. / Ecological Engineering 5 (1995) 183-207
Table 6 Phosphorus retention/release potential of stream sediments and wetland soils (Reddy et al., 1990; Reddy et al., 1993)
Sampling site Phosphorus % of % of added P retained P
Added Retained Released Retained Released mg P m-" (%) (%)
Stream sediments Lower Kissimmee River Basin S-154 (Dry Lake) 893 679 98 76 14 S-65D W.F. Rucks 848 719 39 85 5 Chandler Slough 2267 849 236 37 28 Cypress Slough 2240 1070 157 48 15
Taylor Creek/ 763 668 11 88 2 Nubbin Slough Otter Creek Fisheating Creek 2200 1514 143 69 9 Basin Istokpoga Basin 2242 1262 81 56 6
Wetland Soils Lower Kissimmee River Basin S-154 856 585 146 68 9-5 S-65D W.F. Rucks 881 670 83 76 1.2 Chandler Slough 2231 1011 207 45 20 Cypress Slough 2199 1550 129 70 8
Taylor Creek/ 749 653 13 87 2 Nubbin Slough Otter Creek Fisheating Creek 2390 561 763 23 136 Basin Istokpoga Basin 2447 51 783 2 1535
t i o n / r e l e a s e ra tes were c o r r e l a t e d with ex t rac tab le F e and AI of soils o r sedi- ments , and w a t e r p a r a m e t e r s such as pH, T O C , and P concen t ra t ions . A l inear inc rease in P r e t e n t i o n was o b s e r v e d up to 6 mg P l - t . Phospho rus r e t en t ion by s t r e a m s e d i m e n t s was in the r ange o f 37 to 85% of a d d e d P (Table 6), when s e d i m e n t s were l o a d e d with low P r a inwa te r abou t 2 to 9% of r e t a i n e d P was r e l e a s e d back into w a t e r co lumn in Rucks , O t t e r Creek , I s tokpoga and F i shea t ing C r e e k Basins, sugges t ing tha t P r e t a i n e d was re la t ive ly s table in the system. In cont ras t , 14 to 28% of the P r e t a i n e d was r e l e a s e d f rom Dry Lake , C h a n d l e r and Cypress S loughs s ed imen t s (Tab le 6). Low P r e t en t i on capac i t i es a re r e f l ec ted in s a t u r a t i on of so rp t ion si tes at the s e d i m e n t - f l o o d w a t e r in te r face as a resul t of heavy load ing and low buf fe r in tens i ty of sed iments .
P h o s p h o r u s r e t en t i on in w e t l a n d soils was in the r ange of 2 to 87%, with low P

KR. Reddy et al. /Ecological Engineering 5 (1995) 183-207
Table 7 Phosphorus retention by selected soil/sediments measured at 24-day hydraulic retention time
201
Sampling sites A (1 m-z) P0 (mg m - 2) EPC~, (rag 1- t)
Stream sediments Lower Kissimmee River Basin
S-154 Basin 124 23.7 0.19 S-65D Basin 105 3.4 0.06
Taylor Creek/Nubbin Slough Basin 140 19.7 0.14 Fisheating Creek Basin 145 17.3 0.17 lstokpoga Basin 128 11.7 0.09 Wetland Soils Lower Kissimmee River Basin S-154 Basin 126 63.0 0.53 S-65D Basin 124 17.0 0.14 Fisheating Creek Basin 118 271.0 2.29 Istokpoga Basin 44 136.4 3.14
A = P assimilation coefficient; P0 = P release potential of sediment or soils. Constants derived from Pr = A C - P0; where Pr = P retained in 24-day HRT and C = SRP concentration in the water column.
retention observed in a drained wetland in Istokpoga Basin (Table 6). During contact with low P rainwater, the drained wetland in Istokpoga Basin readily released a large proportion of native P. Wetland soils in Fisheating Creek Basin were also not effective in P retention. Phosphorus retained in wetland soils of W.F. Rucks, Otter Creek, and Cypress Slough was stable with only 2 to 12% released back into water column (Table 6).
To estimate the P assimilation coefficient (A) for wetland soils and stream sediments, P retained during 24 days was regressed with the initial SRP concentra- tion of the water column using the followingequation:
P r = A C - P ~ (3)
where Pr=Phosphorus assimilated by soils or sediments (mg P m-Z); p l = phosphorus release potential of the soil or sediment under ambient conditions (mg P m-2); A- - phosphorus retention coefficient (1 m-2); C = water column P concentration (mg l-L). The "A '° in Eq. (3) is analogous to the Kd in Eq. (2), except ",4" reflects the effect of P diffusion from floodwater and P sorption by sediments. The A values for sediments and wetlands in S-154 Basin were higher than those observed for other basins (Table 7). The pt for stream sediments was in the range of 1 to 43 mg P m -2, as compared to 10 to 271 mg P m -2 for wetland soils. Similar P assimilation coefficients were observed for all stream sediments (85 to 148 1 m -2) and wetland soils (43 to 142 1 m-2). These results suggest that short-term P assimilation is dependent on P concentration of the water column, but the long-term stability of P retained is regulated by the sediment/soil physico- chemical properties.
Using Eq. (3), EPCw was determined for each site. EPCw is defined as the gquilibrium P concentration in the overlying water column, at which point sedi-

202 K.R. Reddy et al. / Ecological Engineering 5 (1995) 183-207
Water
Sediment
Pw
Ps p,
I~tax
7 .;o7£L ambient eotlditio~
Floodwater P
Fig. 6. Schematic of phosphorus retention isotherm for so i l / sed iment -water column. A = Phosphorus assimilation coefficient; Pmax = phosphorus retention maximum; Po = maximum P release potential of native soils or sediments; EPCw = equilibrium P concentrat ion in the water column at which point P retention equals release.
ments neither function as source nor as sink (Fig. 6). If the ambient SRP concentration of the water column is higher than the EPCw value, the underlying sediments o r soils will function as a sink, and as a source if the ambient SRP concentration is lower than the EPCw. The EPCw for stream sediments and wetland soils is generally low, with the exception of wetland soils from Fisheating Creek and Istokpoga Basins (Table 7). EPCw values ranged from 0.06 to 0.19 mg 1-~, with the lowest values measured in the stream sediments from Lower Kissim- mee River Basin (Table 7). In contrast, EPCw values of the wetland soils from Fisheating Creek and Istokpoga Basins were > 10 fold higher (1.95 and 3.90 mg 1-~, respectively) than the other study sites.
The following linear relationships were observed for all stream sediments and wetlands studied.
Sediments: Pr = 127.7[SRP] - 12.4 (4)
r z = 0.949; n = 175
Wetlands: Pf = l l7 .8 [SRP] - 54.2 (5)
r 2 = 0.709; n = 119
Based on the above equations, the EPCw for sediments and wetlands is calculated as 0.10 and 0.47 mg l - t , respectively. Thus, these values can be used as threshold concentrations to determine whether sediments and wetland soils are sources or sinks.
The EPCw for sediments (Table 7) are generally lower than the ambient P concentration of the inflows, suggesting that sediments will function as net sinks. In sediments with high buffer intensity for P sorption, the P loading will have minimal effect on EPCw. However, continuous P loading can result in decreased buffer intensity and increased EPCw. Under these conditions, P retention may occur only at high P concentrations, and the desorbable P pool will increase in the sediments.

K.R. Reddy et aL /Ecological Engineering 5 (1995) 183-207 203
The EPCw = EPC o, when dissolved P in soil/sediment porewater is in equilib- rium with the P concentration in the overlying water column. Lack of agreement between these two values suggest that the system is not at equilibrium ( Tables 5 and 7). This is not surprising considering the dynamic interactions of P with other ions in the water column and in the oxidized sediment/soil-floodwater interface, associated with various processes such as diffusion, precipitation, adsorption and microbial assimilation.
8. Discussion and conclusions
Wetlands and streams are sensitive to N and P inputs. A significant portion of N added to wetlands and streams is lost from the system, while P inputs are usually accumulated within the system. As P enriched water flows through these systems, it interacts with vegetation, aquatic biota and soils/sediments. Assimilation of P in vegetation is usually short-term and decomposition of detrital plant tissue is usually rapid resulting in release of P into the water column. However, the undecomposed organic P accumulates in the system and becomes an integral part of the soil/sediment P pool. Depending on physico-chemical conditions within the system, some wetlands and stream sediments function as net sinks for P, while others function as net sources of P, thus raising a doubt on the P retention capacities of these systems. The prerequisite to understanding the role of wetlands and streams in P retention is an assessment of the interaction of P between the soil/sediment and the overlying water column, and the factors regulating P retention or release.
Phosphate sorption data presented have several practical implications with respect to P release into the water column. Phosphate sorption in wetland soils and stream sediments studied is apparently governed by amorphous and poorly crystalline forms of Fe and Al and organic matter. Aluminum is not involved in oxidation-reduction reactions, and thus is not affected by aerobic or anaerobic conditions. The P sorption by Al compounds is regulated by pH and organic matter, as observed for many sites. Phosphate interactions with Fe can be signifi- cant at the soil/sediment-floodwater interface, where Fe 3÷ in the oxidized zone can function as a sink for: (i) P diffusing from the water column into this layer, and (ii) P diffusing from the underlying anaerobic soil layer. The thickness of this boundary depends on oxygen demanding species at the interface. During rapid stream flow, suspended sediment particles can also sequester P from the water column. These particles with their large surface area and high concentrations of Fe and Al oxides will have high buffer intensity for P sorption. The role of suspended sediments in P retention/release has not been quantified in the Okeechobee Basin.
Phosphorus content of stream sediments and wetland soils is generally higher than the unimpacted upland areas in the basin. Even though the total P content is high, Sm~ x estimates indicate that stream sediments and wetlands are capable of retaining additional P. According to these estimates, stream sediments can poten-

204 K.R. Reddy et al. /Ecological Engineering 5 (1995) 183-207
tially retain an additional 30 to 130% of total P, with an average value of about 50%. Similarly, wetland soils can also retain an additional 54% of total P (32 to 60%). It should be noted that these estimates do not take into account additional P retention sites created in the system as a result of mineral particle deposition during stream flow and incorporation of P into the organic pool.
Only a small portion of the total P (KCl-ex'tractable) is bioavailable (0.1 to 2.3% for stream sediments and 0.2 to 9.5% for wetlands). It is assumed that the P extracted with KCI is potentially bioavailable. Phosphorus loaded to these systems is rapidly retained by Fe/AI solid phases, as evidenced by a large proportion of P in this pool (20 to 70% for stream sediments and 17 to 62% for wetland soils). Phosphorus present in this pool is relatively stable and depending on mineral equilibria a steady slow release can occur. Similarly, Ca/Mg bound P and organic P may also contribute to the bioavailable pool at a relatively slow rate.
Phosphorus assimilation by sediments and wetland soils (as measured by loss of P from floodwater) is controlled by (1) diffusion of P from water to the underlying sediment or soil, and (2) adsorption by soil or sediment. This combined effect of both diffusion and adsorption is shown as a lumped assimilation coefficient (A). The A value was found to be independent of sediment or soil type, over the short-term, and was dependent on P concentration of the water column. However, over the long-term P retained is rapidly complexed with Fe/AI compounds and for most of the soils and sediments only 2 to 28% of the retained P was released into water column. The EPCw values for stream sediments (Table 7) are lower than the SRP concentrations of the respective inflows, suggesting that the sediments are potential sinks for P. However, wetland soils were not as effective sinks, especially in Istokpoga and Fisheating Creek Basins. The variability in P assimilation coeffi- cients was found to be minimal.
Many conclusions made in this study are based on batch column experiments conducted on soil and sediment samples collected from the field. Phosphorus sorption under field conditions is controlled by alterations in hydrologic conditions which will effect soil redox potential and pH, concentration of P in the water column, seasonal changes in vegetation (storage and release) and hydraulic reten- tion time. Under current management practices, flow is primarily confined to streams, thus it is likely that only stream sediments are functioning in retaining P. The wetland soils adjacent to streams have significantly larger capacities to retain P than the stream sediments. Thus, if the flow is directed throughout the wetland, it likely will improve the overall P retention by the wetland stream system. The wetland/stream systems will function as a sink as long as P concentrations of the effluent discharged into the wetland are higher than the EPCw. However, during periods of high rainfall, significant dilution in the effluent can occur, resulting in the sediments functioning as a source of P to the water column.
Acknowledgements
This research was funded in part by a grant from South Florida Water Management District (SFWMD), West Palm Beach, Florida. The authors acknowl-

ICR. Reddy et al. / Ecological Engineering 5 (1995) 183-207 205
edge the assistance of Dr. Eric Flaig, SFWMD in selection of sites and in sampling of soils and sediments. (Florida Agricultural Experiment Station Journal Series No, R-03629.)
References
Allen, L.H., Jr., 1987. Dairy-siting criteria and other options for wastewater management on high water-table soils. Soil Crop Sci. SOc. Fla. Proc., 47: 108-127.
Bache, B.W. and E.G. Williams, 1971. A phosphorus sorption index for soils. J. Soil Sci., 22: 289-300. Barrow, N.J., 1983. A mechanistic model for describing the sorption and desorption of phosphate by
soil. J. Soil Sci., 34: 733-750. Beck, J. and F.A.M. de Haan, 1974. Phosphate removal by soil in relation to waste removal by soil in
relation to waste disposal, Proc. Inter. Conf. on Land for Waste Management, Ottawa, Canada, pp. 77-86.
Berkheiser, V.E., J.J. Street, P.S.C. Rao and T.L Yuan, 1980. Partitioning of inorganic orthophosphate in soil-water systems. In: CRC Critical Reviews in Environmental Control, October 1980, pp. 179-224.
Bostrom, B., M. Jansson and C. Forsberg, 1982. Phosphorus release from lake sediments. Arch. Hydrobiol. Beih., 18: 5-59.
Campbell, K.L., J.C. Capece and T.K. Tremwell, 1995. Surface/subsurface hydrology and phosphorus transport in the Kissimmee River Basin, Florida. Ecol. Eng. (this issue).
Chang, S.C. and M.L Jackson, 1957. Fractionation of soil phosphorus. Soil Sci., 84: 133-144. Davis, F.E., 1982. Strazzulla grove study. In: L.B. Baldwin and A.B. Bottcher (Eds.), Nonpoint
Pollution Control Technology in Florida. Proceedings of the IFAS Conference, Gainesville, Florida, March 9-10, pp. 190-200.
Diaz, O.A., K.R. Reddy and P.A. Moore, 1995. Solubility of inorganic P in stream water as influenced by pH and Ca concentration. Water Res. (in press).
Federico, A.C., J.F. Milleson, P.S. Millar and M. Rosen, 1978. Environmental studies in the chandler slough watershed. South Florida Water Management District, West Palm Beach, Florida.
Federico, A.C., ICG. Dickson, C.R. Kratzer and F.E. Davis, 1981a. Lake Okeechobee water quality studies and eutrophication assessment. Tech. Pub. 81-2. South Florida Water Management District, West Palm Beach, Florida, 270 pp.
Federico, A.C., F.E. Davis and J. Milleson, 1981b. Nutrient retention in an unmodified flow-through marsh. In: P.M. MeCaffrey, T. Beemer and S.E. Gatewood (Eds.), Proceedings of a Symposium on Progress in Wetlands Utilization and Management. Orlando, Florida, June 9-12, p. 432.
Fencbel, T.M. and B.B. Jorgensen, 1977. Detritus food chains of aquatic ecosystems: The role of bacteria. Adv. Microbiol. Ecol., 1: 1-58.
Froelich, P.N., 1988. Kinetic control of dissolved phosphate in natural rivers and estuaries: A primer on the phosphate buffer mechanism. Limnol. Oceanogr., 33: 649-668.
Gachter, R., J.S. Meyer and A. Mares, 1988. Contribution of bacteria to release and fixation of phosphorus in lake sediments. Limnol. Oceanogr., 33: 1542-1558.
Gale, P.M., K.R. Reddy and D.A. Graetz, 1994. Phosphorus retention by wetland soil used for treated wastewater disposal. J. Environ. Qual., 23: 875-877.
Godshalk, G.L and R.G. Wetzel, 1978. Decomposition of aquatic angiosperms. I. Dissolved com- pounds. Aquat. Bot., 5: 281-300.
Goldstein, A.L., 1982. Efficiency of SCS-designated waste stabilization lagoons for reducing nitrogen and phosphorus in milking barn effluent. In: L.B. Baldwin and A.B. Bottcber (Eds.), Proceedings of the IFAS Conference on Nonpoint Pollution Control Technology in Florida, Gainesville, Florida. March 9-10, pp. 78-105.

206 K.R. Reddy et al. / Ecological Engineering 5 (1995) 183-207
Graetz, D.A. and V.D. Nair, 1995. Fate of phosphorus in Florida Spodosols contaminated with cattle manure. Ecol. Eng. (this issue).
Harter, R.D., 1968. Phosphorus adsorption sites in soils. Soil Sci. Soc..~u-n. Proc., 33: 630-631. Hieltjes, H.M. and L. Lijklema, 1980. Fractionation of inorganic phosphates in calcareous sediments. J.
Environ. Qual., 9: 405-407. Howeler, R.H. and D.R. Bouldin, 1971. The diffusion and consumption of oxygen in submerged soils.
Soil Sci. Soc. Am. Proc., 35: 202-208. Khalid, R.A., W.H. Patrick, Jr. and R.D. DeLaune, 1977. Phosphorus sorption characteristics of
flooded soils. Soil Sci. Soc. Am., J. 41: 305-310. Knisel, W.G., Jr., P. Yates, J.M. Sheridan, T.K. Woody, III, L.H. Allen, Jr. and L.E. Asmussen, 1985.
Hydrology and hydrogeology of upper Taylor Creek watershed, Okeechobee county, Florida: Data and analysis. USDA-ARS-25.
Logan, T.J. and E.O. McLean, 1973. Nature of phosphorus and adsorption with depth in soil columns. Soil Sci. Soc. Am. Proc., 37: 351-355.
Logan, T.J., 1982. Mechanism for release of sediment-bound phosphate to water and the effects of agricultural land management on fluvial transport of particulate and dissolved phosphate. Hydrobi- ologia, 92: 519-530.
Mortimer, C.H., 1971. Chemical exchange between sediments and water in the Great Lakes - Speculations on probable regulatory mechanisms. Limnol. Oceanogr., 16: 387-404.
Ogwada, R.A., K.R. Reddy and D.A. Graetz, 1984. Effects of aeration and temperature on nutrient regeneration from selected aquatic macrophytes. J. Environ. Qual., 13: 239-243.
Olila, O.G. and K.R. Reddy, 1995. Comparative evaluation of inorganic phosphorus fractionation methods. (Unpubl. results).
Patrick, W.H. Jr. and R.A. Khalid, 1974. Phosphate release and sorption by soils and sediments: Effect of aerobic and anaerobic conditions. Science, 186: 53-55.
Psenner, R., B. Bostron, M. Dinka, K. Pettersson and M. Sager, 1988. Fractionation of phosphorus in suspended matter and sediment. Arch. Hydrobiol. Beih. Ergebn. Limnol., 30: 43-59.
Reddy, K.R., 1983. Fate of nitrogen and phosphorus in wastewater retention reservoir containing aquatic macrophytes. J. Environ. Qual., 12: 137-141.
Reddy, K.R. and W.F. DeBusk, 1987. Nutrient storage capabilities of aquatic and wetland plants. In: K.R. Reddy and W.H. Smith (Eds.), Aquatic Plants for Water Treatment and Resource Recovery. Magnolia Publishing Inc., Orlando, Florida, pp. 337-357.
Reddy, K.R., K.S. Clough and T.A. DeBusk, 1990. Nutrient removal by wetland soils used for wastewater treatment. Annual report submitted to Reedy Creek Utilities Co., Lake Buena Vista, Florida.
Reddy, K.R., O.A. Diaz, M. Agami, L.J. Scinto and L. LaClaire, 1992a. Phosphorus dynamics in selected wetland/stream systems of South Florida. Final Report submitted to the South Florida Water Management District, West Palm Beach, Flordia.
Reddy, K.R., L.J. Scinto, T.A. DeBusk and J.E. Peterson, 1992b. Phosphorus assimilation by stream sediments II. Final Report submitted to the South Florida Water Management District, West Palm Beach, Florida.
Reddy, K.R., W.G. Harris, O.A. Diaz and H. Wang, 1993. Phosphorus retention in Okeechobee Basin. Final Report submitted to the South Florida Water Management District, West Palm Beach, Florida.
Richardson, C.J., 1985. Mechanisms controlling phosphorus capacity in freshwater wetlands. Science, 228: 1424-1427.
Ryding, S.O. and C. Forsberg, 1977. Sediments as a nutrient source in shallow polluted lakes. In: H. Golterman (Ed.), Interactions Between Sediments and Fresh Water. Dr. W. Junk, The Hague, pp. 227-235.
Sawhney, B.L. and D.E. Hill, 1975. Phosphate sorption characteristics of soils treated with domestic water. J. Environ. Qual., 4: 342-346.
Shukla, S.S., J.K. Syers, J.D.H. Williams, D.E. Armstrong and R.F. Harris, 1971. Sorption of inorganic phosphate by lake sediments. Soil Sci. Soc. Am. Proc., 35: 244-249.

K.R. Reddy et al. / Ecological Engineering 5 (1995) 183-207 20 ~
Sonzogni, W.C., S.C. Chapra, D.E. Armstrong and TJ. Logan, 1982. Bioavailability of phosphorus inputs to lakes. J. Environ. Qual., I h 555-563.
Syers, J.K., R.F. Harris and D.E. Armstrong, 1973. Phosphate chemistry in lake sediments. J. Environ. Qual., 2: 1-14.
Traina, S.J., G. Sposito, D. Hesterberg and U. Kafkafi, 1986. Effects of pH and organic acids on orthophosphate solubility in an acidic, montmorillonitic soil. Soil Sci. Soc. Am. J., 50: 45-52.
Williams, J.D.H., J.K. Syers, R.F. Harris and D.E. Armstrong, 1971. Fractionation of inorganic phosphate in calcareous lake sediments. Soil Sci. SOc. Am. Proc., 35: 250-255.
Yuan, T.L., 1965. A survey of aluminum status in Florida soils. Soil and Crop Sci. SOc. Fla.. 25: 143-152.
Copyright © 2022 FDOKUMEN




















