
Persistent Sonic Hedgehog Signaling in Adult Brain Determines Neural Stem Cell Positional Identity
13
Neuron Article Persistent Sonic Hedgehog Signaling in Adult Brain Determines Neural Stem Cell Positional Identity Rebecca A. Ihrie, 1,2 Jugal K. Shah, 1,2 Corey C. Harwell, 1,3 Jacob H. Levine, 1,2 Cristina D. Guinto, 1,2 Melissa Lezameta, 4 Arnold R. Kriegstein, 1,3 and Arturo Alvarez-Buylla 1,2, * 1 Eli and Edythe Broad Center of Regeneration Medicine and Stem Cell Research 2 Department of Neurosurgery 3 Department of Neurology University of California, San Francisco, San Francisco, CA 94143, USA 4 Instituto Cavanilles de Biodiversitat i Biologia Evolutiva, 46980 Valencia, Spain *Correspondence: [email protected] DOI 10.1016/j.neuron.2011.05.018 SUMMARY Neural stem cells (NSCs) persist in the subventricular zone (SVZ) of the adult brain. Location within this germinal region determines the type of neuronal progeny NSCs generate, but the mechanism of adult NSC positional specification remains unknown. We show that sonic hedgehog (Shh) signaling, resulting in high gli1 levels, occurs in the ventral SVZ and is associated with the genesis of specific neuronal progeny. Shh is selectively produced by a small group of ventral forebrain neurons. Ablation of Shh decreases production of ventrally derived neuron types, while ectopic activation of this pathway in dorsal NSCs respecifies their progeny to deep granule interneurons and calbindin-positive periglo- merular cells. These results show that Shh is neces- sary and sufficient for the specification of adult ventral NSCs. INTRODUCTION Most of the neurons in the central nervous system are produced in the embryo, and for many years it was thought that this was the only source of neurons in the CNS. We now know that two populations of neurons continue to be produced in the adult: olfactory bulb interneurons and granule neurons in the dentate gyrus of the hippocampus. Neuronal production is limited to the subventricular zone (SVZ), along the walls of the lateral ventricles, and the subgranular zone, within the dentate gyrus. These two regions contain neural stem cells (NSCs) and continue to generate new neurons throughout adult life (Zhao et al., 2008; Ihrie and Alvarez-Buylla, 2011). Within the SVZ, glial fibrillary acidic protein (GFAP)-expressing stem cells (type B cells) give rise to rapidly dividing transit-amplifying progeny (type C cells), which in turn generate immature neuroblasts (type A cells). These neuroblasts migrate to the olfactory bulb (OB) within a network of tangentially oriented chains that coalesce to form the rostral migratory stream (RMS) (Luskin, 1993; Lois and Alvarez- Buylla, 1994; Doetsch et al., 1999a, 1999b). Within the OB, young neurons migrate radially, complete their differentiation, and integrate into the granular and periglomerular layers. In the mouse, the SVZ covers an area greater than six square millimeters along the rostrocaudal and dorsoventral axes (Mirzadeh et al., 2008). Neuroblasts derived from the SVZ traverse a significant distance to reach their final destination in the OB. Why are neurons derived from such an extended prolif- erative zone, and how does site of origin in the SVZ affect cell fate? One clue comes from recent experiments using viral or genetic lineage tracing to label specific subregions of the devel- oping or adult SVZ (Kelsch et al., 2007; Kohwi et al., 2007; Merkle et al., 2007; Ventura and Goldman, 2007; Young et al., 2007). These results suggest that the SVZ is arranged as a mosaic; the position of stem cells within the SVZ determines the types of differentiated progeny generated. In particular, deep granule interneurons and a subpopulation of periglomerular cells arise from the ventral SVZ, while superficial granule interneurons and distinct periglomerular cells are derived from the dorsal SVZ. The molecular mechanisms responsible for this positional spec- ification of primary progenitors in the adult SVZ are not known. The Hedgehog (Hh) signaling pathway is central to the devel- opment and patterning of the nervous system and other organs (McMahon et al., 2003; Fuccillo et al., 2006). In mammals, this signaling pathway is initiated by one of three family members—Indian hedgehog (Ihh), Desert hedgehog (Dhh), or Sonic hedgehog (Shh). Secreted Hedgehog morphogen binds Patched at the cell surface, relieving its inhibition of the trans- membrane protein Smoothened (Rohatgi et al., 2007). Smooth- ened triggers the activation of the Gli transcription factors. In the absence of Hh signal, the Gli3 transcription factor acts as a transcriptional repressor, while Gli2 functions primarily as a transcriptional activator upon Hh stimulation and can initiate transcription of Gli1, a constitutive transcriptional activator that indicates high levels of pathway activity (Ahn and Joyner, 2005; Palma et al., 2005; Clement et al., 2007). The responsiveness of a cell to a given level of Hh ligand is also modulated by intrinsic expression of cell-surface proteins. The transmembrane proteins Cdo, Boc, and Gas1 are thought to positively regulate Hh pathway activation and allow cells at a greater distance from a Hh source to respond to lower levels within a Hh gradient (Tenzen et al., 2006; Allen et al., 2007; McLellan et al., 2008). Conversely, Hedgehog interacting protein 250 Neuron 71, 250–262, July 28, 2011 ª2011 Elsevier Inc.
Transcript of Persistent Sonic Hedgehog Signaling in Adult Brain Determines Neural Stem Cell Positional Identity

Neuron
Article
Persistent Sonic Hedgehog Signaling in Adult BrainDetermines Neural Stem Cell Positional IdentityRebecca A. Ihrie,1,2 Jugal K. Shah,1,2 Corey C. Harwell,1,3 Jacob H. Levine,1,2 Cristina D. Guinto,1,2 Melissa Lezameta,4
Arnold R. Kriegstein,1,3 and Arturo Alvarez-Buylla1,2,*1Eli and Edythe Broad Center of Regeneration Medicine and Stem Cell Research2Department of Neurosurgery3Department of Neurology
University of California, San Francisco, San Francisco, CA 94143, USA4Instituto Cavanilles de Biodiversitat i Biologia Evolutiva, 46980 Valencia, Spain
*Correspondence: [email protected] 10.1016/j.neuron.2011.05.018
SUMMARY
Neural stem cells (NSCs) persist in the subventricularzone (SVZ) of the adult brain. Location within thisgerminal region determines the type of neuronalprogeny NSCs generate, but the mechanism of adultNSC positional specification remains unknown. Weshow that sonic hedgehog (Shh) signaling, resultingin high gli1 levels, occurs in the ventral SVZ and isassociated with the genesis of specific neuronalprogeny. Shh is selectively produced by a smallgroup of ventral forebrain neurons. Ablation of Shhdecreases production of ventrally derived neurontypes, while ectopic activation of this pathway indorsal NSCs respecifies their progeny to deepgranule interneurons and calbindin-positive periglo-merular cells. These results show that Shh is neces-sary and sufficient for the specification of adultventral NSCs.
INTRODUCTION
Most of the neurons in the central nervous system are produced
in the embryo, and formany years it was thought that this was the
only source of neurons in the CNS. We now know that two
populations of neurons continue to be produced in the adult:
olfactory bulb interneurons and granule neurons in the dentate
gyrus of the hippocampus. Neuronal production is limited to
the subventricular zone (SVZ), along the walls of the lateral
ventricles, and the subgranular zone, within the dentate gyrus.
These two regions contain neural stem cells (NSCs) and continue
to generate new neurons throughout adult life (Zhao et al., 2008;
Ihrie and Alvarez-Buylla, 2011). Within the SVZ, glial fibrillary
acidic protein (GFAP)-expressing stem cells (type B cells) give
rise to rapidly dividing transit-amplifying progeny (type C cells),
which in turn generate immature neuroblasts (type A cells).
These neuroblasts migrate to the olfactory bulb (OB) within a
network of tangentially oriented chains that coalesce to form the
rostral migratory stream (RMS) (Luskin, 1993; Lois and Alvarez-
Buylla, 1994; Doetsch et al., 1999a, 1999b). Within the OB,
250 Neuron 71, 250–262, July 28, 2011 ª2011 Elsevier Inc.
young neurons migrate radially, complete their differentiation,
and integrate into the granular and periglomerular layers.
In the mouse, the SVZ covers an area greater than six square
millimeters along the rostrocaudal and dorsoventral axes
(Mirzadeh et al., 2008). Neuroblasts derived from the SVZ
traverse a significant distance to reach their final destination in
the OB. Why are neurons derived from such an extended prolif-
erative zone, and how does site of origin in the SVZ affect cell
fate? One clue comes from recent experiments using viral or
genetic lineage tracing to label specific subregions of the devel-
oping or adult SVZ (Kelsch et al., 2007; Kohwi et al., 2007;Merkle
et al., 2007; Ventura and Goldman, 2007; Young et al., 2007).
These results suggest that the SVZ is arranged as a mosaic; the
position of stem cells within the SVZ determines the types of
differentiated progeny generated. In particular, deep granule
interneurons and a subpopulation of periglomerular cells arise
from the ventral SVZ, while superficial granule interneurons and
distinct periglomerular cells are derived from the dorsal SVZ.
The molecular mechanisms responsible for this positional spec-
ification of primary progenitors in the adult SVZ are not known.
The Hedgehog (Hh) signaling pathway is central to the devel-
opment and patterning of the nervous system and other
organs (McMahon et al., 2003; Fuccillo et al., 2006). Inmammals,
this signaling pathway is initiated by one of three family
members—Indian hedgehog (Ihh), Desert hedgehog (Dhh), or
Sonic hedgehog (Shh). Secreted Hedgehog morphogen binds
Patched at the cell surface, relieving its inhibition of the trans-
membrane protein Smoothened (Rohatgi et al., 2007). Smooth-
ened triggers the activation of the Gli transcription factors. In
the absence of Hh signal, the Gli3 transcription factor acts as
a transcriptional repressor, while Gli2 functions primarily as
a transcriptional activator upon Hh stimulation and can initiate
transcription of Gli1, a constitutive transcriptional activator that
indicates high levels of pathway activity (Ahn and Joyner,
2005; Palma et al., 2005; Clement et al., 2007).
The responsiveness of a cell to a given level of Hh ligand is also
modulated by intrinsic expression of cell-surface proteins. The
transmembrane proteins Cdo, Boc, and Gas1 are thought to
positively regulate Hh pathway activation and allow cells at
a greater distance from a Hh source to respond to lower levels
within a Hh gradient (Tenzen et al., 2006; Allen et al., 2007;
McLellan et al., 2008). Conversely, Hedgehog interacting protein

gli1 mRNA gli2 mRNA gli3 mRNA
dorsal
ventral
sep
str
sep
str
sep
str
sepstr sepstr
sep
str
A
D E F
ptc-lacZ gli1-nlacZ
dorsal
ventral sep
str
str
str
sepstr
I J
HG
CB
-1
-0.5
0
0.5
1
1.5
2
2.5
3
3.5
4
Nkx6.2 Nkx2.1 Gli1 Gli2 Gli3
vSVZ
septum
Relative transcript expression (compared to dorsal SVZ)
fold
cha
nge
(log2
)
K
*
**
*
Figure 1. Gli1 Expression Is Primarily
Ventral
(A–F) Dorsal and ventral areas shown in images
(A)–(F) are indicated in the cartoon at left. In situ
hybridization for the three Gli family members
reveals that gli1 mRNA is present in the ventral
SVZ (D), but not the dorsal SVZ (A). gli2 and gli3
mRNA are present at low levels in both the dorsal
and ventral SVZ (B, C, E, and F). Scale bars: 50
microns.
(G–J) LacZ staining in either ptc-lacZ (G and I) or
gli1-nlacZ (H and J) reporter strains is strongest in
the ventral SVZ (I and J) and is reduced or absent in
the dorsal SVZ (G and H). Red dashed lines indi-
cate the lateral ventricle.
(K) qRT-PCR detection of Gli family members in
microdissected ventral SVZ and septum recapit-
ulates the expression observed in in situ hybrid-
ization. Values shown are relative transcript
expression compared to microdissected dorsal
SVZ. Nkx2.1 and Nkx6.2, which are known to be
upregulated in ventral SVZ, are highly expressed in
both ventral SVZ samples (blue bars) and RNA
derived from the septum (red bars). gli1 is ex-
pressedmore highly in the ventral SVZ and septum
than in the dorsal SVZ. gli2 and gli3 transcript
expression did not differ significantly between the
dorsal and ventral SVZ, but both genes were ex-
pressed to higher levels in the septum. Asterisks =
p < 0.05.
Neuron
Sonic Hedgehog Regulates Neural Stem Cell Identity
(Hhip) binds and sequesters Hh ligand, and acts cooperatively
with Patched as a negative-feedback mechanism to regulate
pathway activation (Chuang and McMahon, 1999; Chuang
et al., 2003). Studies in the developing neural tube have demon-
strated that the varying levels of Hh ligand and interacting
proteins present over time along the dorsoventral axis of this
structure result in graded levels of activity of the Gli transcription
factors, allowing a morphogen gradient to be translated into
transcriptional control of neuronal identity (Jessell, 2000; Ribes
and Briscoe, 2009). Shh signaling also regulates cell proliferation
and fate in the developing forebrain and hindbrain (Machold
et al., 2003; Corrales et al., 2004; Blaess et al., 2006; Balordi
and Fishell, 2007a, 2007b; Xu et al., 2010). Shh is important in
the initial generation and proliferation of postnatal neural stem
cells in the SVZ and DG, but whether it has a role in the specifi-
cation of regional identity in the adult is not known (Lai et al.,
2003; Balordi and Fishell, 2007b; Han et al., 2008).
Here, we demonstrate that Shh signaling has a continuous,
critical role in the dorsoventral specification of adult SVZ neural
stem cells. We find that despite the ubiquitous expression of
Smoothened on stem cells throughout this region, Gli1 expres-
sion primarily occurs in the ventral SVZ and is associated with
particular neuronal fates. We show that Shh signaling is instruc-
tive in cell fate specification, and removal of Shh expression
affects the generation of specific neuronal subtypes. Further-
more, we identify a possible source of secreted Sonic hedgehog
(Shh) ligand close to the ventral SVZ: surprisingly, this source is
neuronal. These results are the first identification of a signaling
pathway that is sufficient to determine neuronal cell fate in adult
SVZ NSCs.
RESULTS
Gli1 Expression Occurs in the Ventral SVZShh pathwaymembers have been implicated in the development
and postnatal maintenance of SVZ neural stem cells (Machold
et al., 2003; Ahn and Joyner, 2005; Balordi and Fishell, 2007a,
2007b). Remarkably, in situ hybridization for gli1, gli2, and gli3
revealed that gli1 expression is higher in the ventral SVZ (Figures
1A and 1D), while gli2 and gli3 are present both ventrally and
dorsally (Figures 1B, 1C, 1E, and 1F). Likewise, staining of brain
sections from mice carrying gli1-nlacZ and ptc-lacZ reporter
alleles (Goodrich et al., 1997; Bai et al., 2002) showed high levels
of reporter expression in the ventral SVZ, in both the lateral and
medial walls (Figures 1I and 1J). We also microdissected these
regions from adult brains and performed qRT-PCR analysis. To
confirm that the correct areas were dissected, we measured
relative expression of the transcription factors Nkx2.1 and
Nkx6.2, which are expressed in the ventral forebrain during
development (Xu et al., 2008, 2010) and are present ventrally in
the adult SVZ (L. Fuentealba and A.A.-B., data not shown). Using
qRT-PCR, we observed elevated gli1 expression in the ventral
SVZ as well as the medial septum when compared to the dorsal
SVZ (Figure 1K).
Neuron 71, 250–262, July 28, 2011 ª2011 Elsevier Inc. 251

Neuron
Sonic Hedgehog Regulates Neural Stem Cell Identity
We next stained adult SVZ for Smoothened (Smo), an obligate
component of the canonical Hh pathway. Smo protein was
present throughout the SVZ in a pattern reminiscent of GFAP,
which is expressed by type B cells in this region (see Figures
S1A–S1F; Doetsch et al., 1999a; Garcia et al., 2004; Tavazoie
et al., 2008). Confocal analysis of both dorsal and ventral SVZ
using two different antibodies demonstrated that Smo is ex-
pressed on a subset (�80%) of GFAP-positive cells in both
subregions. This staining was not observed when the antibody
was incubated with blocking peptide or when primary antibody
was omitted, and was almost entirely absent in the brains of
hGFAP::Cre; Smofl/fl mice, where Smoothened is lost in most
neural stem cells (Figures S1G and S1H; Han et al., 2008).
Smo did not colocalize with Dcx, CD24, or EGFR, which label
other cell types in the SVZ. To confirm that Smoothened is
primarily expressed on stem cells, we infused the antimitotic
cytosine-b-D-arabinofuranoside (Ara-C) into the brains of wild-
type mice for 6 days. This treatment eliminates fast-dividing
transit-amplifying (type C) cells and neuroblasts from the sub-
ventricular zone, while sparing slow-dividing stem cells (Doetsch
et al., 1999b; Long et al., 2001). While a 6 day Ara-C infusion was
sufficient to eliminate doublecortin-positive neuroblasts in wild-
type mice, Smo expression remained after Ara-C treatment
(Figures S1J–S1M).
We next examined pathway activation using Gli1-CreERT2;
R26YFP mice (Ahn and Joyner, 2004), in which cells responding
to high levels of Hedgehog ligand express a tamoxifen-inducible
Cre recombinase. By giving tamoxifen at postnatal day 60 (P60)
in adult mice, we identified SVZ stem cells in which the Hh
pathway was active and followed their YFP-labeled progeny
(Figure 2). We administered tamoxifen for 5 days and examined
YFP expression at 0 days, 5 days, or 30 days after the end of
treatment. In contrast to the widespread expression of Smooth-
ened, activation of Hh signaling was much more restricted.
Intriguingly, pathway activity occurred in a spatially specific
manner—the initial population of Hh-responsive cells was
distributed along the anterior-posterior axis of this region, but
largely confined to the ventral half of the SVZ (Figure 2D), an
area which primarily produces deep granule interneurons within
the OB (Merkle et al., 2007). This labeling was not due to limited
infiltration of tamoxifen in the dorsal SVZ, as equivalent treat-
ment of mice with ubiquitously expressed Cre-ER transgenes
caused recombination throughout the SVZ (Kuo et al., 2006;
data not shown).
The initial population of YFP-labeled ventral cells were largely
GFAP-positive (Figure 2D). YFP-positive migrating neuroblasts
appeared at 5 days after tamoxifen treatment in the dorsal and
ventral SVZ, RMS, and the core of the OB (Figures 2B, 2E, 2H,
2K, and S2). The presence of ventrally derived neuroblasts in
the dorsal wedge is consistent with the pattern of tangential
migration to the OB (Doetsch and Alvarez-Buylla, 1996). These
dorsal YFP-positive cells were GFAP negative and doublecortin
positive, confirming their identity as neuroblasts (Figures 2B and
S2). At 30 days after treatment, YFP-labeled cells were present
throughout the SVZ, in the RMS, and in the granular and periglo-
merular layers of the OB (Figures 2C, 2F, 2I, and 2L). We also
observed many YFP+ cells in the medial septum at 30 days after
tamoxifen treatment (Figure S3). These labeled septal cells were
252 Neuron 71, 250–262, July 28, 2011 ª2011 Elsevier Inc.
primarily S100b+ astrocyte-like cells, similar to those observed
upon EGF infusion into the SVZ (Deloulme et al., 2004; Hachem
et al., 2005; Gonzalez-Perez et al., 2009). We next quantified the
localization of labeled progeny in the OBs of Gli1-CreERT2;
R26YFP animals at one month after tamoxifen. The labeled cells
were primarily interneurons located in the deep granule layer of
the OB, similar to the cells labeled by viral injection in the ventral
SVZ (Figures 2M–2O and S2M). The distribution of labeled cells
differed significantly from distributions generated by combining
dorsal and ventral injection data or from general BrdU labeling
and lineage tracing in the SVZ (shown in Figure 4), confirming
that Gli1 expression delineates a population of ventrally-derived
interneurons.
Sonic Hedgehog Ligand Is Produced in the VentralForebrainWe next investigated possible sources of Hh ligand in the adult
brain. As previously reported (Echelard et al., 1993), we did not
observe expression of indian hedgehog or desert hedgehog in
the adult brain by in situ hybridization (data not shown). We did
not observe expression of shh in the dorsal SVZ. However, shh
mRNA was present in the medial septum, ventral forebrain,
and in infrequent cells close to the ventral SVZ (Figures S4A–
S4C). This was confirmed by qRT-PCR on dissected ventral
SVZ and septum (Figure S4D) and is consistent with previous
reports of in situ hybridization in the rat brain (Traiffort et al.,
1999). We next used a genetic approach to label the cells
producing Shh and visualize cell morphology. We first examined
ShhCre; R26YFP animals (Harfe et al., 2004), in which cells
expressing Shh at any point in development recombine the
R26YFP reporter (Figure S4E). In addition to labeling in the cere-
bellum and cortex, we also observed an accumulation of YFP+
cells in the ventral forebrain.
By administering tamoxifen to ShhCreER; R26YFP animals,
we induced YFP expression in cells producing Shh in the adult
(Figure 3; Harfe et al., 2004). YFP expression identified cells in
the ventral forebrain, extending along the rostrocaudal axis
adjacent and ventral to the SVZ. These cells primarily localized
to the medial and ventral septum, the preoptic nuclei near the
hypothalamus, and the bed nuclei of the stria terminalis. We
also observed rare labeled cells in cortex (Garcia et al., 2010).
Within the septum, YFP+ cells localized to both the horizontal
and vertical limbs of the diagonal band, approximately 0.25–
1 mm ventromedial to the SVZ (Figures 3A and 3B). A number
of YFP+ cells in the bed nuclei of the stria terminalis were in close
proximity to the ventral tip of the lateral ventricle (boxed area in
Figures 3A and 3D). We did not observe YFP-labeled cells near
the dorsal SVZ, the RMS, OB, or in the choroid plexus—other
sites which have been suggested to produce this ligand (Balordi
and Fishell, 2007a; Angot et al., 2008). Labeled cells had the
morphology of neurons, and all were colabeled by the neuronal
marker protein NeuN (Figure 3H). Most YFP-labeled cells were
GABAergic (GABA-positive; Figure 3I), with a small number of
cholinergic (ChAT-positive), YFP-positive cells (Figure 3J). We
did not observe YFP-positive dopaminergic (TH-positive) cells
(data not shown). Labeled cells within the bed nucleus of the stria
terminalis were of particular interest because of their close prox-
imity to the ventral SVZ, and we examined these cells in greater

YFP / GFAP
0 d post tamoxifen 5 d post tamoxifen 30 d post tamoxifen
dorsalSVZ
ventralSVZ
RMS
olfactorybulb
YFP / GFAP YFP / GFAP YFP / GFAP
YFP / GFAP YFP / GFAP YFP / GFAP
YFP / Dcx YFP / Dcx YFP / Dcx
YFP / Dcx YFP / Dcx YFP / Dcx
A
D
G
J K L
IH
E F
B C
YFP / DcxN YFP / DcxO
tamoxifen(5 days)
Gli1CreER; R26YFP YFP labeling in SVZ, RMS, OB
M 1
0.8
0.6
0.4
0.2
00 to 0.33
deep0.33 to 0.67intermediate
0.67 to 1superficial
Dorsal viral injection (n = 3)
Ventral viral injection (n = 3)
Gli1CreER; R26YFP (n = 3)
Frac
tion
of to
tal l
abel
ed c
ells
Normalized distance from core
Figure 2. High Hedgehog Pathway Activity Is Limited to the Ventral SVZ
(A–L) Lineage tracing of Gli1-expressing cells in the SVZ. Schematic at top indicates the experimental timeline. Immediately after 5 days of tamoxifen admin-
istration inGli1CreERT2; R26YFP animals, a small number of YFP/GFAP double-positive cells are present in the ventral SVZ (arrowhead and inset, D). YFP-labeled
cells are absent from the dorsal SVZ, RMS, and OB (A, G, and J). At 5 days after tamoxifen administration, YFP-labeled cells are present in the ventral SVZ, and
some YFP labeling coincides with GFAP labeling (arrowheads and inset, E). YFP-labeled cells are also present in the dorsal SVZ (B), medial septum (right side in
E), RMS (H), and the core of the OB, where doublecortin (Dcx)-expressing migrating neuroblasts enter this structure (K). One month after tamoxifen adminis-
tration, there is widespread YFP labeling in the SVZ (C and F), RMS (I), and OB (L). Scale bars: 50 microns.
(M–O) Quantification of the localization of labeled granular layer neurons inGli1CreERT2; R26YFP animals at 1 month after tamoxifen administration (red bars, M).
The neuronal progeny of labeled cells tend to localize in the deep OB granular layer close to the core, a phenotype that is consistent with an origin in the ventral
SVZ. The distributions of cells resulting from dorsal (dark blue) or ventral (light blue) viral injection are shown for comparison. Data shown are average ± SEM
(n in figure is number of animals). This localization can be observed consistently in multiple samples (N and O). The OB core (dashed line) is located at the bottom
of both images.
Neuron
Sonic Hedgehog Regulates Neural Stem Cell Identity
detail. Computerized tracings of YFP-labeled cells highlighted
thin processes that were located close to the basal side of ventral
SVZ cells (Figures 3E and 3F). In order to better characterize
these cells, we used whole-mount preparations of ShhCreER;
R26YFP animals, allowing en face visualization of the lateral
and medial walls of the ventricles. ShhCreER; R26YFP-labeled
cells had thin processes that interdigitated with the basal side
of cell bodies within the SVZ (Figure 3G).
To test whether these cells are actively producing Shh and
might contact the subventricular zone, we pursued two strate-
gies. First, we injected Ad:CreStopLight (Ad:CSL) virus in the
ventral SVZ of Shh-Cre animals. This virus contains a CMV
Neuron 71, 250–262, July 28, 2011 ª2011 Elsevier Inc. 253
αYFP αYFP αYFP
Shh-CreER; R26YFP - 4 days post tamoxifen administration
B C D
YFP / DAPIE
YFP / NeuN /DAPI
YFP / GABA /DAPI
H I YFP / ChAT /DAPI
J
YFP / DAPIGYFP / DAPIF
A
YFP Fluorogold YFP / FG / DAPI
K L M
tamoxifen(2 days)
(2 days)
ShhCreER; R26YFP Inject Fluorogold in ventricle YFP and Fluorogold
Shh/ NeuN /DAPI
Shh / DAPI Shh / RFP /DAPI
Q R
N O P
Shh / DAPIShh / DAPI
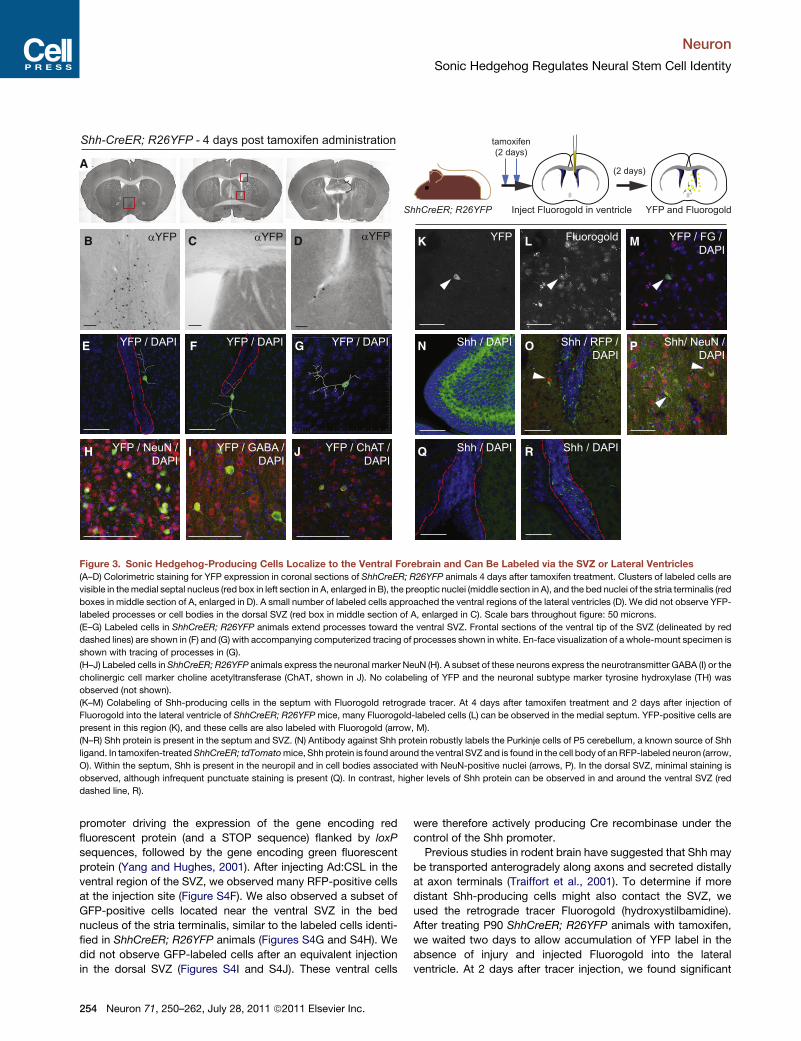
Figure 3. Sonic Hedgehog-Producing Cells Localize to the Ventral Forebrain and Can Be Labeled via the SVZ or Lateral Ventricles
(A–D) Colorimetric staining for YFP expression in coronal sections of ShhCreER; R26YFP animals 4 days after tamoxifen treatment. Clusters of labeled cells are
visible in themedial septal nucleus (red box in left section in A, enlarged in B), the preoptic nuclei (middle section in A), and the bed nuclei of the stria terminalis (red
boxes in middle section of A, enlarged in D). A small number of labeled cells approached the ventral regions of the lateral ventricles (D). We did not observe YFP-
labeled processes or cell bodies in the dorsal SVZ (red box in middle section of A, enlarged in C). Scale bars throughout figure: 50 microns.
(E–G) Labeled cells in ShhCreER; R26YFP animals extend processes toward the ventral SVZ. Frontal sections of the ventral tip of the SVZ (delineated by red
dashed lines) are shown in (F) and (G) with accompanying computerized tracing of processes shown in white. En-face visualization of a whole-mount specimen is
shown with tracing of processes in (G).
(H–J) Labeled cells in ShhCreER; R26YFP animals express the neuronal marker NeuN (H). A subset of these neurons express the neurotransmitter GABA (I) or the
cholinergic cell marker choline acetyltransferase (ChAT, shown in J). No colabeling of YFP and the neuronal subtype marker tyrosine hydroxylase (TH) was
observed (not shown).
(K–M) Colabeling of Shh-producing cells in the septum with Fluorogold retrograde tracer. At 4 days after tamoxifen treatment and 2 days after injection of
Fluorogold into the lateral ventricle of ShhCreER; R26YFPmice, many Fluorogold-labeled cells (L) can be observed in the medial septum. YFP-positive cells are
present in this region (K), and these cells are also labeled with Fluorogold (arrow, M).
(N–R) Shh protein is present in the septum and SVZ. (N) Antibody against Shh protein robustly labels the Purkinje cells of P5 cerebellum, a known source of Shh
ligand. In tamoxifen-treatedShhCreER; tdTomatomice, Shh protein is found around the ventral SVZ and is found in the cell body of an RFP-labeled neuron (arrow,
O). Within the septum, Shh is present in the neuropil and in cell bodies associated with NeuN-positive nuclei (arrows, P). In the dorsal SVZ, minimal staining is
observed, although infrequent punctuate staining is present (Q). In contrast, higher levels of Shh protein can be observed in and around the ventral SVZ (red
dashed line, R).
Neuron
Sonic Hedgehog Regulates Neural Stem Cell Identity
promoter driving the expression of the gene encoding red
fluorescent protein (and a STOP sequence) flanked by loxP
sequences, followed by the gene encoding green fluorescent
protein (Yang and Hughes, 2001). After injecting Ad:CSL in the
ventral region of the SVZ, we observed many RFP-positive cells
at the injection site (Figure S4F). We also observed a subset of
GFP-positive cells located near the ventral SVZ in the bed
nucleus of the stria terminalis, similar to the labeled cells identi-
fied in ShhCreER; R26YFP animals (Figures S4G and S4H). We
did not observe GFP-labeled cells after an equivalent injection
in the dorsal SVZ (Figures S4I and S4J). These ventral cells
254 Neuron 71, 250–262, July 28, 2011 ª2011 Elsevier Inc.
were therefore actively producing Cre recombinase under the
control of the Shh promoter.
Previous studies in rodent brain have suggested that Shh may
be transported anterogradely along axons and secreted distally
at axon terminals (Traiffort et al., 2001). To determine if more
distant Shh-producing cells might also contact the SVZ, we
used the retrograde tracer Fluorogold (hydroxystilbamidine).
After treating P90 ShhCreER; R26YFP animals with tamoxifen,
we waited two days to allow accumulation of YFP label in the
absence of injury and injected Fluorogold into the lateral
ventricle. At 2 days after tracer injection, we found significant

Shh orfl/+
ShhCreER; Shh fl
tamoxifengli1 mRNAin SVZ
olfactorybulb BrdU distribution
BrdU(1 week)(2 weeks) (3 weeks)
sep
str
sep
str
gli1 mRNA (SVZ)
Shh protein (septum and SVZ)
Shhfl/+ ShhCreER; Shhfl
Shhfl/+ ShhCreER; ShhflBrdU (olfactory bulb)
BrdU / CalB (olfactory bulb)Shhfl/+
ShhCreER; Shhfl
E F
GFr
actio
n of
tota
l Brd
U+Shh / DAPI Shh / DAPIA B
DC Shh / DAPI Shh / DAPI 0
0.1
0.2
0.3
0.4
0.5
BrdU / CalB DAPI
BrdU / CalB DAPI
0 to 0.33deep
0.33 to 0.67intermediate
0.67 to 1superficial
Shhfl/+ (n = 3)ShhCreER; Shh fl
(n = 5)
I
J
**
Normalized distance from core
H
Per
cent
Cal
B-B
rdU
/ to
tal B
rdU
+
genotype
Shh (n = 3)fl/+
ShhCreER; Shh fl(n=6)
0
1
2
3*
Figure 4. Loss of Shh Alters the Production
of Olfactory Interneurons
(A–F) Treatment of ShhCreER/Shhfl animals with
tamoxifen results in the loss of Shh protein and gli1
expression. Schematic at top shows the timeline of
tamoxifen and BrdU administration. At six weeks
after tamoxifen administration, Shh protein is
present in the septum and ventral SVZ of control
animals (A and C). The SVZ is outlined by a red
dashed line. In situ hybridization for gli1 shows the
expected pattern of ventral expression in Shhfl/+
animals (arrowhead, E). Shh protein is largely
absent in ShhCreER/Shhfl animals (B and D).
gli1 expression is also substantially reduced or
absent in ShhCreER/Shhfl animals, indicating that
pathway activity is compromised (arrowhead, F).
White scale bars throughout figure: 50 microns.
Black scale bars: 500 microns.
(G) The distribution of newly born granule neurons
in the OB is altered in the absence of Shh. At
3 weeks after a 1 week pulse of BrdU in Shhfl/+
animals, labeled cells are distributed throughout
the granular layers of the OB, with a greater
percentage of the total present in the deep granule
layers close to the core (blue bars, G). However, in
ShhCreER/Shhfl animals, BrdU-labeled cells were
less frequent in the deep granule layers and more
prevalent in the superficial layers of the OB.
Quantification of the distribution of BrdU-labeled
cells by genotype shows a statistically significant
decrease in the number of deep granule cells and
concomitant increase in the number of superficial
granule cells (asterisks = p = 0.01, unpaired t test).
Data shown are average ± SEM. n indicates
number of biological replicates counted, with at
least 100 OB granular layer cells counted per
animal.
(H–J) The production of calbindin-positive peri-
glomerular cells is decreased in the absence of
Shh. At 3 weeks after a 1 week pulse of BrdU,
a subpopulation of BrdU-labeled cells in the peri-
glomerular layer of Shhfl/+ OBs are calbindin-
positive (colabeling shown in I). This population is
substantially reduced in ShhCreER/Shhfl animals
(p = 0.0033, unpaired t test).
Neuron
Sonic Hedgehog Regulates Neural Stem Cell Identity
Fluorogold labeling around the ventricles and in the bed nuclei
of the stria terminalis, as well as many labeled cells in the
medial septum and preoptic nuclei. A small number of Fluoro-
gold-labeled cells in the septum also expressed YFP (Figures
3K–3M), suggesting that Shh produced in the septum could
also reach the SVZ by transport along axonal extensions. We
did not observe more distant Fluorogold-labeled cells in the
midbrain or hindbrain.
Finally, we performed immunohistochemical staining for Shh
protein (rabbit mAb 95.9, kind gift from Genentech). We found
that Shh protein was present in the septum in cell bodies asso-
ciated with NeuN-positive nuclei and at low levels in the associ-
ated neuropil (Figure 3P). In the ventral SVZ, we also found Shh
protein in the neuropil and in association with the apical and
basal surfaces of SVZ cells (Figures 3O, 3R, and 4C). Infrequent,
punctate staining for Shh protein was also observed in the dorsal
SVZ (Figure 3Q), suggesting that a low level of ligand may be
present in this region despite the absence of Shh-producing cells
or their processes.
Removal of Shh Reduces the Production of VentrallyDerived Olfactory NeuronsAlthough Shh ligand production and pathway activity both
occur in the ventral forebrain, this did not necessarily indicate
a requirement for this signaling in the generation of particular
olfactory interneurons. We examined the requirement for Shh
ligand in the adult brain by conditionally deleting Shh in mice
carrying one copy of a knockin allele of ShhCreER and a floxed
Shh allele in the second locus (ShhCreER/Shhfl). Adult (P60)
ShhCreER/Shhflmice and littermateShhfl/+ controls were treated
with tamoxifen for 5 days to induce deletion of the functional Shh
allele. After a 2 week period to ensure loss of Shh protein expres-
sion and tamoxifen clearance, mice were given a 1 week pulse of
BrdU, followed by a 3 week chase, to label newly produced
Neuron 71, 250–262, July 28, 2011 ª2011 Elsevier Inc. 255

saline cyclopamine αShh (5E1) SAG
dorsalSVZ
ventralSVZ
tamoxifen
Gli1CreER; R26YFP YFP labeling in SVZ
(5 days)
YFP/GFAP YFP/GFAP YFP/GFAP YFP/GFAP
YFP/GFAPYFP/GFAPYFP/GFAPYFP/GFAP
A B C D
E F G H
Figure 5. Dorsal SVZ Cells Are Resistant to Pathway Activation
(A–H) Infusion of Hedgehog pathway agonists in the SVZ. Schematic at top illustrates pump cannula placement, allowing infusion of solution into the SVZ and
CSF. Infusion of vehicle (sterile saline) does not alter the predominantly ventral distribution of cells labeled by theGli1CreERT2; R26YFP reporter (A and E). Infusion
of either the small-molecule antagonist cyclopamine (B and F) or an anti-Shh antibody (C and G) results in a reduction in YFP-positive cells. Administration of the
small-molecule agonist SAG causes a robust increase in YFP labeling in the ventral SVZ (H), but does not cause a similar upregulation of YFP expression in the
dorsal SVZ (D). Scale bars: 50 microns.
Neuron
Sonic Hedgehog Regulates Neural Stem Cell Identity
interneurons in the OB. At the end of this time course, we
observed a decrease in Shh protein in both the septum and
ventral SVZ of ShhCreER/Shhfl mice (Figures 4A–4D). Impor-
tantly, we also observed a loss of gli1 mRNA expression in
tamoxifen-treated ShhCreER/Shhfl mice but not treated Shhfl/+
controls, indicating that Shh pathway activity was significantly
decreased in ventral SVZ (Figures 4E and 4F). As expected,
the OBs of control animals had BrdU–labeled cells distributed
throughout the granular layer, with 36% of cells observed in
the deep granule layer (Figures 4G and S5). Similar numbers of
BrdU-labeled interneurons were present in all genotypes, and
label-retaining cells as well as proliferating cells were present
in the SVZ for both genotypes, suggesting that stem cell self-
renewal and progenitor proliferation were not grossly affected
(data not shown). However, in ShhCreER/Shhfl animals, there
was a shift in the distribution of labeled cells, with 15% fewer
deep granule cells present and a 25% increase in superficial
granule cells in the labeled population when compared to
controls (Figure 4G; p = 0.02, unpaired t test). This suggests
that a subset of deep granule interneurons is lost when Shh
ligand is removed from the adult brain.
We also examined the effects of Shh loss on the population
of calbindin (CalB)-positive periglomerular cells normally pro-
duced by the ventral SVZ (Merkle et al., 2007). In ShhCreER/
Shhfl animals, production of new CalB+ periglomerular cells
(BrdU/CalB double-positive cells) was decreased by almost
90% compared to controls (Figures 4H–4J; p = 0.0033, unpaired
t test). This reduction in CalB-positive cells was more pro-
256 Neuron 71, 250–262, July 28, 2011 ª2011 Elsevier Inc.
nounced than the reduction in deep granule cells. Shh signaling
may be required for the generation of specific subgroups of
deep granule cells, but not others, resulting in a smaller decrease
in the total population of deep granule cells. However, in both
populations of cells, we observed a meaningful change in the
cell types generated, indicating that Shh production plays
a role in the production of different types of neurons destined
for the OB by ventral NSCs.
Dorsal SVZ Cells Are Intrinsically Resistantto Smo AgonistTo test whether dorsal and ventral SVZ cells are equivalently
responsive to Shh pathway activation, we administered Smooth-
ened agonist (SAG) in the cerebrospinal fluid via an intracranial
osmotic pump into the lateral ventricles. SAG is a small molecule
that efficiently activates the Hh pathway (Chen et al., 2002).
Vehicle (saline), SAG, or the pathway antagonists cyclopamine
or anti-Shh antibody (5E1) were infused for 5 days inGli1CreERT2;
R26YFPmice. During infusion, mice were treated with tamoxifen
to acutely label cells in which the Hh pathway was active. YFP
labeling in vehicle-infused mice was predominantly ventral,
demonstrating that pump installation alone did not alter the
pattern of Hh signaling (Figures 5A and 5E). We observed
increased GFAP labeling after all pump implantations, likely
due to increased numbers of reactive astrocytes. Administration
of either cyclopamine or 5E1 antibody reduced the number of
YFP-positive cells in the ventral SVZ, confirming that YFP
labeling was dependent on pathway activation (Figures 5B, 5C,

Neuron
Sonic Hedgehog Regulates Neural Stem Cell Identity
5F, and 5G). SAG infusion resulted in a dramatic increase in YFP-
positive cells, both GFAP-positive and –negative, in the ventral
SVZ, but did not significantly alter the pattern of YFP labeling
in the dorsal SVZ (Figures 5D and 5H).
Infusion of cyclopamine, 5E1, or SAG may also affect SVZ
cell survival or proliferation, as suggested by previous experi-
ments in which Smoothened was ablated in the SVZ (Balordi
and Fishell, 2007b). Staining for the proliferation marker Ki67
indicated that large changes in proliferation did not occur during
the time frame of this experiment. In both controls and SAG-
infused animals, we observed small populations of YFP-positive
cells in the dorsal SVZ. Most of these cells were GFAP negative
and Dcx positive, suggesting that they correspond to young
migrating neurons (Figure 5 and data not shown). We cannot
exclude that a small subpopulation of Gli1-expressing type B
or C cells are present in dorsal regions. The regional difference
in Gli1 distribution remains after agonist infusion, suggesting
that additional cell-intrinsic factorsmay affect the ability of dorsal
cells to activate the Hh pathway.
Ectopic Pathway Activation ReprogramsNeuronal ProgenyWhile high Hedgehog pathway activity was required for the
production of particular types of neuronal progeny, this observa-
tion did not necessarily indicate that Hedgehog signaling had an
instructive role in cell fate. To investigate this possibility, we per-
formed targeted injections of Ad:GFAPpCre virus in SmoM2-
YFP; R26R mice (Mao et al., 2006). The Ad:GFAPp-Cre virus,
in which Cre recombinase expression is driven by the murine
GFAP promoter, results in recombination in primary progenitors
(type B cells) within this region (Merkle et al., 2007). In these
animals, Cre-mediated recombination causes the expression of
SmoM2, a constitutively active mutant of Smo, and activation
of the Hh pathway. By injecting Ad:GFAPpCre in the dorsal
SVZ of SmoM2-YFP; R26R animals, we activated the Hh
pathway to high levels in a ligand-independent, cell-intrinsic
fashion while simultaneously labeling these cells with b-galacto-
sidase. This allowed us to follow the labeled progeny of these in-
fected stem cells. Remarkably, while expression of the SmoM2
protein is sufficient to drive rapid tumorigenesis in other contexts
(Schuller et al., 2008), we did not observe abnormalities within
the SVZ of injected SmoM2-YFP; R26R animals. One month
after injection, lacZ-labeled cells were found at the site of injec-
tion in control and SmoM2-YFP; R26R animals (Figure 6). gli1
expression was absent in the dorsal SVZ of injected R26R
mice. SmoM2-YFP; R26R animals showed a marked upregula-
tion of gli1, but not Shh, mRNA at the site of virus injection
(Figures S6A–S6I), confirming that Hh signaling was active in
infected cells.
At 1 month after dorsal injection of Ad:GFAPpCre in control
R26R animals, lacZ labeling marked a population of cells
located in the superficial granular layer of the OB, consistent
with previous work (Figure 6B). Remarkably, dorsal injections
in SmoM2-YFP; R26R animals generated a population of labeled
cells that localized to the deep granule layer of the OB (Figures
6E, 6M, and S6J), similar to the progeny resulting from injections
in the ventral SVZ of R26R or SmoM2-YFP/R26R animals (Fig-
ures 6H and 6K). These labeled cells expressed NeuN (Figures
6C, 6F, 6I, and 6L), suggesting that SmoM2 expression in
infected neural stem cells did not block maturation but did alter
the type of progeny generated.
We next injected Ad:GFAPpCre in SmoM2-YFP; CAG animals
andCAG littermates to generate progeny expressingGFP, which
fills the cell and allows visualization of cell morphology (Figure 7).
Ad:GFAPpCre infection of dorsal SVZ cells in SmoM2-YFP; CAG
animals caused a shift in the localization of GFP-expressing
progeny in the OB like that observed with the R26R reporter
(Figures 7A and 7D). Within the SmoM2-YFP; CAG SVZ, we
observed an almost 4-fold increase (p = 0.0025) in the dorsal
expression of the transcription factor Pbx3a, which is normally
limited to the ventral SVZ (Figures 7J, 7L, and 7R). We also
observed a decrease in expression of Pax6, which is normally
present in the dorsal SVZ, in YFP-positive cells in SmoM2/CAG
animals (Figures 7N and 7P). Within the OB, SmoM2/GFP-
expressing cells were positive for NeuN and the neurotransmitter
GABA (Figures 7B, 7C, 7E, and 7F), confirming that the relocali-
zation of progeny does not block their maturation into inter-
neurons. The projection patterns of deep and superficial inter-
neurons differ (Merkle et al., 2007; Whitman and Greer, 2009),
so in addition to soma location, we traced the arborizations of
GFP-labeled cells. The progeny of dorsally injectedCAG animals
were primarily superficial interneurons with dendrites that
reached past the midline of the external plexiform layer of the
OB (Figure 7G). After dorsal injections in SmoM2/CAG animals,
labeled olfactory interneurons tended to have dendrites that
contacted the inner half of the external plexiform layer, a feature
that is typical of deep granule interneurons (Figure 7H). In addi-
tion to the deep granule cells that arise from the ventral SVZ,
calbindin-expressing periglomerular cells are also derived from
this region. Within the periglomerular cell population derived
from dorsal injections, we saw a three-fold enrichment of CalB/
GFP-double positive cells in SmoM2-YFP; CAG animals when
compared to controls (Figures 7O, 7Q, and 7S). We conclude
that ectopic activation of the Hh pathway is sufficient to respecify
dorsal SVZ neuronal progeny.
DISCUSSION
We demonstrate that the Hedgehog pathway has a critical func-
tion in the regional specification of stem cells of the adult brain.
We found that Smo is expressed by stem cells throughout the
SVZ. However, high Hh pathway activation is more prevalent in
the ventral SVZ and is sufficient to respecify dorsal SVZ stem
cells when ectopically induced. In the forebrain, we found that
Shh is the only Hh isoform expressed. Interestingly, Shh produc-
tion occurs in neurons that are located very close to the ventral
subventricular zone. These results indicate that specification of
subpopulations of neural stem cells and their progeny in the adult
brain is actively regulated by Hh signaling.
It has previously been suggested that the Hh pathway func-
tions in multiple cell types within the SVZ. Genetic and pharma-
cologic studies have argued that this pathway is important in
stem cell self-renewal, the generation of transit-amplifying
progeny, and neuroblast migration to the OB (Machold et al.,
2003; Palma et al., 2005; Balordi and Fishell, 2007a, 2007b).
Our results indicate that astrocytes of the SVZ are the major
Neuron 71, 250–262, July 28, 2011 ª2011 Elsevier Inc. 257

SVZ (injection site) olfactory bulbβ-galactosidase
R26Ronly -dorsal
injection
SmoM2/R26R -dorsal
injection
R26Ronly -
ventralinjection
SmoM2/R26R -ventral
injection
β-gal / NeuN / DAPI
A
D
B C
M
FE
G
J
H I
LK
0
0.2
0.4
0.6
0.8
1 R26R dorsal (n = 3)
SmoM2 ventral (n = 3)
R26R ventral (n = 3)
SmoM2 dorsal (n = 5)
0 to 0.33deep
0.33 to 0.67intermediate
0.67 to 1superficial
Frac
tion
of to
tal l
acZ+
gra
nule
cel
ls
Normalized distance from core
*
* *
Figure 6. High Pathway Activity Causes Relocali-
zation of Neuronal Progeny
(A–L) Ad:GFAPpCre was injected into the dorsal or ventral
SVZ of R26R and SmoM2-YFP/R26R animals and
brains were collected one month later to analyze labeled
progeny. LacZ labeling within the SVZ (A, D, G, and J)
confirmed the site of virus injection. Dorsal injections in
control animals primarily generated superficial labeled
cells (B) that were distant from the core of the OB (indi-
cated by a dashed line). Injections in the dorsal SVZ of
SmoM2 animals gave rise to deep cells located close to
the core of the OB (E), similar to those derived from ventral
viral injections in either genotype (H and K). b-galactosi-
dase-expressing progeny in both control and SmoM2
animals coexpressed the NeuN protein, a marker of
mature neurons (arrows in C, F, I, and L). Black scale bars:
50 microns. White scale bars: 14 microns.
(M) Quantification of the spatial distribution of labeled
progeny in R26R and SmoM2-YFP; R26R animals after
dorsal or ventral Ad:GFAPpCre injection. Ventral virus
injection in either genotype (pale red and pale blue bars)
results in cells that are mostly located close to the core of
the OB. Dorsal virus injection in R26R animals (bright blue
bars) results in many cells that are distant from the core.
However, dorsal virus injection in SmoM2; R26R animals
results in cells that are located close to the core (bright red
bars), with a distribution similar to that from ventral injec-
tions. Data shown are average ± SEM, with total number of
animals indicated on graph. Asterisks = p < 0.01.
Neuron
Sonic Hedgehog Regulates Neural Stem Cell Identity
Hh-responsive population within the SVZ: expression of Smo
protein, which is required for a cell to respond to Hh ligand, is
restricted to the GFAP-positive population (including type B1
cells) in this region. Intriguingly, Smo expression labels a subset
of GFAP-expressing SVZ cells, suggesting either that Smo is
258 Neuron 71, 250–262, July 28, 2011 ª2011 Elsevier Inc.
only expressed in a subset of stem cells, or
that it is a more specific marker of stem cells
than GFAP, which is also expressed by some
astrocytes in non-germinal regions (Garcia
et al., 2010).
We searched for components of the Hh
signaling pathway that might be differentially
expressed in the subregions of the SVZ. We
did not observe significant differences in the
expression of Smo, Gli2, or Gli3 between the
ventral and dorsal SVZ, but found that Gli1
expression is much higher in the ventral SVZ.
Lineage tracing of these cells using Gli1-
CreERT2 mice indicates that progenitors re-
sponding to high levels of Shh are located in
the ventral SVZ and primarily generate deep
granule interneurons. Moreover, Hh signaling
appears to be instructive in cell fate deci-
sions: ablation of Shh expression reduced the
production of ventrally-derived progeny, and
ectopic activation of the pathway resulted in
the respecification of dorsally-derived progeny
to a ventral fate. These results are surprising,
as previous studies using viral targeting and
cell culture indicated that cell identitywas largely
encoded using a cell-intrinsic program: transplantation of dorsal
cells to a ventral location, even after intervening time in culture,
was not sufficient to respecify their progeny (Merkle et al., 2007).
The differences in cell fate between previous transplantation
studies and the results shown here may be due to several

olfactory bulb
CAGonly
SmoM2/CAG
dorsal SVZ(injection site) ventral SVZ
sep
str
sep
str
str
str
GFP / Pbx3a / DAPI GFP / CalB / DAPIGFP / Pbx3a / DAPI
dorsal SVZ(injection site)
GFP / Pax6 / DAPI
I
L M Q
R S
OJ
str
str
P
N
0
4
8
12
16SmoM2/
CAG
CAG only
genotype
Pbx
3a-G
FP /
tota
l GFP
+
0
2
4
6
8
10SmoM2/
CAG
CAG only
genotype
Cal
B-G
FP /
tota
l GFP
+
olfactory bulb
CAGonly
SmoM2/CAG
αGFP / hematoxylin GFP / NeuN / DAPI GFP / GABA / DAPI
A
D E F
CB
CAG dorsal SmoM2 / CAG dorsalG H
exte
rnal
ple
xifo
rmla
yer
Figure 7. SmoM2 Expression Results in
Relocalization, Altered Dendritic Projec-
tions, and Ventral Marker Expression
(A–F) Dorsal injection of Ad:GFAPpCre in SmoM2-
YFP; CAG animals (D–F) also resulted in relocali-
zation of labeled progeny close to the core of
the OB (dashed line) when compared to CAG
controls (A–C). GFP-expressing progeny of both
genotypes expressed NeuN (arrows, B and E) and
the neurotransmitter GABA (arrows, C and F),
consistent with their status as differentiated
inhibitory interneurons. Scale bars: 50 microns.
(G and H). Camera lucida tracings of individual
GFP-labeled cells in CAG and SmoM2; CAG
animals show the projections of these cells into the
external plexiform layer of the OB (delineated by
gray shading). Dashed lines indicate the mitral cell
layer (lower line) and periglomerular layer (upper
line) of the OB. While dorsal SVZ-derived cells in
CAG animals typically project to the outer half of
the EPL (G), dorsal SVZ-derived cells from
SmoM2; CAG animals project to the inner half of
the EPL, a feature associated with a deep granule
interneuron identity (H).
(I, J, L–N, P, andR) At 1month after dorsal injection
of Ad:GFAPpCre in CAG animals, many GFP-
labeled cells are present at the site of injection in
the dorsal SVZ (I). However, the transcription
factor Pbx3a is more prevalent in the ventral SVZ
(J). After a similar injection in SmoM2-YFP; CAG
animals, Pbx3a is present at similar frequency in
the uninjected cells of the ventral SVZ (M), but is
also enriched in dorsal GFP-positive cells (L). str =
striatum, sep = septum. This enrichment is quan-
tified in (R)—graph shows average ± SEM for three
independent injections per genotype (p = 0.0025,
unpaired t test). In concert with the increased
expression of the ventral marker Pbx3a, we also
observed a loss of the dorsal marker Pax6—while
many dorsal GFP-positive cells normally express
Pax6 (arrows, N), this expression is absent in
SmoM2/CAG animals (P).
(O, Q, and S) Within the OBs of CAG animals at
one month after dorsal Ad:GFAPpCre injection,
few GFP-positive periglomerular cells (O and S)
expressed the marker calbindin (shown in red).
However, we did observe an increase in the
number of CalB/GFP double-positive cells in
periglomerular cells derived from injections in
SmoM2-YFP; CAG animals (Q and S). Graph
shows average ± SEM for three independent
injections per genotype.
Neuron
Sonic Hedgehog Regulates Neural Stem Cell Identity
Neuron 71, 250–262, July 28, 2011 ª2011 Elsevier Inc. 259

Neuron
Sonic Hedgehog Regulates Neural Stem Cell Identity
possiblemechanisms: dorsal SVZ cellsmay be intrinsically resis-
tant to the Hh signal, resulting in lower pathway activation even
upon exposure to equivalent levels of ligand, or a second devel-
opmental pathway may act to oppose Hh action in the dorsal
region, as is the case in the developing neural tube (Liem et al.,
2000). It has also recently been recognized that SVZ stem cells
have a specialized apical-basal orientation within the SVZ niche
(Mirzadeh et al., 2008; Shen et al., 2008; Tavazoie et al., 2008).
Transplantation experiments may not allow the grafted popula-
tion to integrate and adopt proper apical-basal positioning within
the niche. Hh ligand may be delivered to ventral SVZ cells via
specialized local contacts which are not recapitulated after
transplantation. Importantly, the present results indicate that
strong activation of the Shh pathway can override the intrinsic
programming of dorsal neural progenitors, suggesting that
reprogramming of neural stem cells for therapeutic purposes
may depend on the identification of the relevant molecular signal
for a desired cell type. This study also provides the first in vivo
respecification of adult neural cell fate by modulation of the Hh
pathway.
We identify clusters of Shh-producing neurons in the ventral
forebrain, in locations that are consistent with previous studies
at the RNA level. A subset of these cells, in the bed nucleus of
the stria terminalis, have processes that are immediately adja-
cent to the ventral SVZ. In addition, some Shh-producing cells
in the ventral and medial septum are able to take up retrograde
tracer molecules that are injected into the lateral ventricle, sug-
gesting that Shh ligand may also reach the ventral SVZ by
anterograde transport from the septum (Traiffort et al., 2001).
The localized activation of ventral SVZ stem cells, and expres-
sion of Gli1 in these cells, might be part of an adult brain regula-
tory mechanism to locally modulate production of specific
neuronal subtypes destined for different OB circuits.
Shh is produced by Purkinje neurons in the developing
cerebellum and by cells of the floor plate in the neural tube
(Ho and Scott, 2002; Fuccillo et al., 2006). In these instances,
Shh signaling directs significant large-scale remodeling and
patterning of developing tissue. Our results demonstrate that in
the adult brain, this pathway remains active and directs the
production of specific subtypes of neurons. The finding that
mature neurons in the adult brain are a likely source of Shh ligand
suggests that neural network activity may regulate generation of
certain types of neurons within the SVZ. It remains unclear if Shh
reaches ventral stem cells via diffusion or whether more special-
ized contacts exist between Shh-producing neurons and stem
cells. It will be of interest to study the mechanism of Hh ligand
delivery to the SVZ, and to identify what circuits in the brainmight
activate the release of Shh ligand.
The SVZ is the largest germinal zone in the adult brain, and
is capable of generating thousands of immature neurons each
day. Importantly, it is organized as a mosaic, with stem cells in
different locations producing different types of neurons (Hack
et al., 2005; Kohwi et al., 2005; Kelsch et al., 2007; Merkle et al.,
2007; Ventura and Goldman, 2007; Young et al., 2007; Alvarez-
Buylla et al., 2008). These groups of olfactory interneurons
differ in location and are also thought to differ functionally
(Gheusi et al., 2000; Cecchi et al., 2001; Kosaka and Kosaka,
2007). Here, we elucidate a molecular mechanism for the spec-
260 Neuron 71, 250–262, July 28, 2011 ª2011 Elsevier Inc.
ification of a subpopulation of neural stem cells within this exten-
sive adult germinal layer. We show that manipulation of this
pathway allows stem cells to be redirected to a different fate.
These studies demonstrate that adult neural stem cells, which
normally produce a restricted repertoire of progeny, may be
reprogrammed if the relevant specification signals are identified.
EXPERIMENTAL PROCEDURES
Tamoxifen Administration
All animal procedures were carried out in accordancewith institutional (IACUC)
and NIH guidelines. Tamoxifen was prepared at 20 mg/ml in corn oil and
administered via oral gavage. Adult mice (P60–P90) were treated with 1 mg
(Gli1-CreER) or 5 mg (Shh-CreER)/day tamoxifen for 5 consecutive days and
sacrificed at the specified times.
Antimitotic Treatment
Adult mice were treated with a solution of 2% cytosine-b-arabinofuranoside
(Sigma) or sterile saline alone as a control. Solutions were infused for 6 days
at the pial surface using a miniosmotic pump (Alzet 1007D).
Injections and Adenovirus Preparation
Ad:GFAPpCre into was injected into dorsal and ventral adult SVZ as described
(Merkle et al., 2007) using 50 nl of virus. Injections of Ad:CSL were carried out
using 100 nl of virus, and Fluorogold injections were carried out using 250 nl of
tracer. Injections used the following coordinates: dorsal SVZ —0.5 anterior,
3.2 lateral, 1.8 deep, needle at 45� angle; ventral SVZ—0.5 anterior, 4.6 lateral,
3.3 deep, needle at 45� angle; lateral ventricle—0.38 posterior, 1.0 lateral,
2.25 deep, with needle vertical. x and y coordinates were zeroed at bregma,
and the z coordinate was zeroed at the pial surface.
Miniosmotic Pump Implantation
Miniosmotic pumps (Alzet 1007D) were assembled under sterile conditions,
filled with vehicle (0.9% saline), 0.5 mMSmoothened agonist (EMDChemicals),
5 mM cyclopamine (Sigma), or 5 mg/ml 5E1 antibody (DSHB), and allowed to
equilibrate overnight at 37�C. Pump installation was performed on a stereo-
taxic rig as previously described.
Immunostaining
Immunostainings were carried out on methanol-fixed 10 micron frozen
sections (Figure S1) or paraformaldehyde-fixed 50 micron free floating Vibra-
tome sections (all other stains) according to standard procedures. Primary
antibodies used were goat anti-Smo (Santa Cruz, C-17), rabbit anti-Smo
(MBL 2668, kind gift of J. Reiter), mouse anti-GFAP (Chemicon), rabbit anti-
doublecortin (Cell Signaling), mouse anti-PSA-NCAM (Chemicon), chicken
anti-GFP (Aves Labs), rabbit anti-RFP (MBL), mouse anti-b-galactosidase
(Promega), anti-NG2, rabbit anti-Olig2 (kind gift of D. Rowitch), rabbit anti-
S100b, mouse anti-GABA (Sigma), goat anti-ChAT (Chemicon), anti-NeuN
(Chemicon), rabbit anti-tyrosine hydroxylase (Pel-Freez Biologicals), rabbit
95.9 anti-Shh (kind gift from S. Scales, Genentech), mouse anti-Pbx3a (Santa
Cruz), and rabbi anti-calbindin D28k (Chemicon). The secondary antibodies
used were conjugated to AlexaFluor dyes (Invitrogen/Molecular Probes). For
preblocking of Smoothened antibody (Santa Cruz, C-17), the antibody was
incubated with the accompanying blocking peptide (sc-6367P, Santa Cruz).
LacZ (X-gal) staining was carried out after brief fixation in paraformaldehyde
according to standard procedures.
Microscopic Analysis, Tracing, and Quantification
Fluorescent staining was visualized using a Leica SP5 confocal microscope
and analyzed using NIH ImageJ. Tracings of neuronal processes from fluores-
cent staining were completed using the Filament module in Imaris analysis
software (Bitplane). Colorimetric staining was visualized using an Olympus
AX70 microscope, Retiga 2000R camera and LabVelocity software. Measure-
ments of olfactory interneuron localization were carried out using the Measure
and Label plugin in NIH ImageJ, with normalization to granular layer width
carried out as described (Merkle et al., 2007). Data were quantified and

Neuron
Sonic Hedgehog Regulates Neural Stem Cell Identity
analyzed using GraphPad Prism 5. Tracing of colorimetric anti-GFP staining
was completed using a Nikon microscope with camera lucida adaptor. Hand
tracings were scanned and imported into Adobe Illustrator using the LiveTrace
and LivePaint modules.
qRT-PCR
For qRT-PCR, dorsal SVZ, ventral SVZ, striatum, and septum were microdis-
sected from 2 mm slices of unfixed brain. Dissected tissue was immediately
placed in RNAlater solution (Ambion) and stored at �20�C until all brains
(14 total) were dissected. RNA isolation was done with the RNEasy Mini kit
(QIAGEN). cDNA was synthesized using SuperScript III RT (Invitrogen), and
qRT-PCR was completed using SYBR Green PCR Master Mix (Applied
BioSystems) on an ABI7900HT. Data analysis and statistical tests were carried
out using the REST algorithm (Pfaffl et al., 2002).
Whole-Mount Dissection
Whole-mount dissection of Shh-CreER; R26YFP brains was performed as
described (Mirzadeh et al., 2008). Full methods including video are available
online (Mirzadeh et al., 2010).
In Situ Hybridization
In situ hybridization was performed using standard protocols (Han et al., 2008).
Antisense and sense RNA probes were labeled with digoxigenin and visualized
with alkaline phosphatase-NBT/BCIP reaction (Roche).
SUPPLEMENTAL INFORMATION
Supplemental Information includes six figures and can be found with this
article online at doi:10.1016/j.neuron.2011.05.018.
ACKNOWLEDGMENTS
The authors thank Alexandra Joyner for sharing the Gli1CreERT2 mouse line
prior to publication, David Rowitch and the members of the Rowitch and
Alvarez-Buylla labs for many useful conversations, and T. Nguyen for
assistance with qRT-PCR preparations. R.A.I. was supported by postdoctoral
fellowships from the Damon Runyon Cancer Research Foundation (DRG1935-
07) and the AACR/NBTS. C.C.H. was supported by a Ford Foundation Post-
doctoral Fellowship and a UNCF Merck Postdoctoral Science Research
Fellowship. This work was supported by grants from the US National Institutes
of Health (NS28478 and HD32116), the John G. Bowes Research Fund, and
a grant from the Goldhirsh Foundation to A.A.-B. A.A.-B. is the Heather and
Melanie Muss Endowed Chair of Neurological Surgery at UCSF.
Accepted: May 16, 2011
Published: July 27, 2011
REFERENCES
Ahn, S., and Joyner, A.L. (2004). Dynamic changes in the response of cells to
positive hedgehog signaling during mouse limb patterning. Cell 118, 505–516.
Ahn, S., and Joyner, A.L. (2005). In vivo analysis of quiescent adult neural stem
cells responding to Sonic hedgehog. Nature 437, 894–897.
Allen, B.L., Tenzen, T., and McMahon, A.P. (2007). The Hedgehog-binding
proteins Gas1 and Cdo cooperate to positively regulate Shh signaling during
mouse development. Genes Dev. 21, 1244–1257.
Alvarez-Buylla, A., Kohwi, M., Nguyen, T.M., and Merkle, F.T. (2008). The
heterogeneity of adult neural stem cells and the emerging complexity of their
niche. Cold Spring Harb. Symp. Quant. Biol. 73, 357–365.
Angot, E., Loulier, K., Nguyen-Ba-Charvet, K.T., Gadeau, A.P., Ruat, M., and
Traiffort, E. (2008). Chemoattractive activity of sonic hedgehog in the adult
subventricular zone modulates the number of neural precursors reaching the
olfactory bulb. Stem Cells 26, 2311–2320.
Bai, C.B., Auerbach, W., Lee, J.S., Stephen, D., and Joyner, A.L. (2002). Gli2,
but not Gli1, is required for initial Shh signaling and ectopic activation of the
Shh pathway. Development 129, 4753–4761.
Balordi, F., and Fishell, G. (2007a). Hedgehog signaling in the subventricular
zone is required for both the maintenance of stem cells and the migration of
newborn neurons. J. Neurosci. 27, 5936–5947.
Balordi, F., and Fishell, G. (2007b). Mosaic removal of hedgehog signaling in
the adult SVZ reveals that the residual wild-type stem cells have a limited
capacity for self-renewal. J. Neurosci. 27, 14248–14259.
Blaess, S., Corrales, J.D., and Joyner, A.L. (2006). Sonic hedgehog regulates
Gli activator and repressor functions with spatial and temporal precision in the
mid/hindbrain region. Development 133, 1799–1809.
Cecchi, G.A., Petreanu, L.T., Alvarez-Buylla, A., and Magnasco, M.O. (2001).
Unsupervised learning and adaptation in a model of adult neurogenesis.
J. Comput. Neurosci. 11, 175–182.
Chen, J.K., Taipale, J., Young, K.E., Maiti, T., and Beachy, P.A. (2002). Small
molecule modulation of Smoothened activity. Proc. Natl. Acad. Sci. USA 99,
14071–14076.
Chuang, P.T., and McMahon, A.P. (1999). Vertebrate Hedgehog signalling
modulated by induction of a Hedgehog-binding protein. Nature 397, 617–621.
Chuang, P.T., Kawcak, T., and McMahon, A.P. (2003). Feedback control of
mammalian Hedgehog signaling by the Hedgehog-binding protein, Hip1,
modulates Fgf signaling during branching morphogenesis of the lung. Genes
Dev. 17, 342–347.
Clement, V., Sanchez, P., de Tribolet, N., Radovanovic, I., and Ruiz i Altaba, A.
(2007). HEDGEHOG-GLI1 signaling regulates human glioma growth, cancer
stem cell self-renewal, and tumorigenicity. Curr. Biol. 17, 165–172.
Corrales, J.D., Rocco, G.L., Blaess, S., Guo, Q., and Joyner, A.L. (2004).
Spatial pattern of sonic hedgehog signaling through Gli genes during cere-
bellum development. Development 131, 5581–5590.
Deloulme, J.C., Raponi, E., Gentil, B.J., Bertacchi, N., Marks, A., Labourdette,
G., and Baudier, J. (2004). Nuclear expression of S100B in oligodendrocyte
progenitor cells correlates with differentiation toward the oligodendroglial
lineage and modulates oligodendrocytes maturation. Mol. Cell. Neurosci. 27,
453–465.
Doetsch, F., and Alvarez-Buylla, A. (1996). Network of tangential pathways for
neuronal migration in adult mammalian brain. Proc. Natl. Acad. Sci. USA 93,
14895–14900.
Doetsch, F., Caille, I., Lim, D.A., Garcıa-Verdugo, J.M., and Alvarez-Buylla, A.
(1999a). Subventricular zone astrocytes are neural stem cells in the adult
mammalian brain. Cell 97, 703–716.
Doetsch, F., Garcıa-Verdugo, J.M., and Alvarez-Buylla, A. (1999b).
Regeneration of a germinal layer in the adult mammalian brain. Proc. Natl.
Acad. Sci. USA 96, 11619–11624.
Echelard, Y., Epstein, D.J., St-Jacques, B., Shen, L., Mohler, J., McMahon,
J.A., and McMahon, A.P. (1993). Sonic hedgehog, a member of a family of
putative signaling molecules, is implicated in the regulation of CNS polarity.
Cell 75, 1417–1430.
Fuccillo, M., Joyner, A.L., and Fishell, G. (2006). Morphogen to mitogen: the
multiple roles of hedgehog signalling in vertebrate neural development. Nat.
Rev. Neurosci. 7, 772–783.
Garcia, A.D., Doan, N.B., Imura, T., Bush, T.G., and Sofroniew, M.V. (2004).
GFAP-expressing progenitors are the principal source of constitutive neuro-
genesis in adult mouse forebrain. Nat. Neurosci. 7, 1233–1241.
Garcia, A.D., Petrova, R., Eng, L., and Joyner, A.L. (2010). Sonic hedgehog
regulates discrete populations of astrocytes in the adult mouse forebrain.
J. Neurosci. 30, 13597–13608.
Gheusi, G., Cremer, H., McLean, H., Chazal, G., Vincent, J.D., and Lledo, P.M.
(2000). Importance of newly generated neurons in the adult olfactory bulb for
odor discrimination. Proc. Natl. Acad. Sci. USA 97, 1823–1828.
Gonzalez-Perez, O., Romero-Rodriguez, R., Soriano-Navarro, M., Garcia-
Verdugo, J.M., and Alvarez-Buylla, A. (2009). Epidermal growth factor induces
the progeny of subventricular zone type B cells tomigrate and differentiate into
oligodendrocytes. Stem Cells 27, 2032–2043.
Neuron 71, 250–262, July 28, 2011 ª2011 Elsevier Inc. 261

Neuron
Sonic Hedgehog Regulates Neural Stem Cell Identity
Goodrich, L.V., Milenkovi�c, L., Higgins, K.M., and Scott, M.P. (1997). Altered
neural cell fates and medulloblastoma in mouse patched mutants. Science
277, 1109–1113.
Hachem, S., Aguirre, A., Vives, V., Marks, A., Gallo, V., and Legraverend, C.
(2005). Spatial and temporal expression of S100B in cells of oligodendrocyte
lineage. Glia 51, 81–97.
Hack, M.A., Saghatelyan, A., de Chevigny, A., Pfeifer, A., Ashery-Padan, R.,
Lledo, P.M., and Gotz, M. (2005). Neuronal fate determinants of adult olfactory
bulb neurogenesis. Nat. Neurosci. 8, 865–872.
Han, Y.G., Spassky, N., Romaguera-Ros, M., Garcia-Verdugo, J.M., Aguilar,
A., Schneider-Maunoury, S., and Alvarez-Buylla, A. (2008). Hedgehog
signaling and primary cilia are required for the formation of adult neural stem
cells. Nat. Neurosci. 11, 277–284.
Harfe, B.D., Scherz, P.J., Nissim, S., Tian, H., McMahon, A.P., and Tabin, C.J.
(2004). Evidence for an expansion-based temporal Shh gradient in specifying
vertebrate digit identities. Cell 118, 517–528.
Ho, K.S., and Scott, M.P. (2002). Sonic hedgehog in the nervous system: func-
tions, modifications and mechanisms. Curr. Opin. Neurobiol. 12, 57–63.
Ihrie, R.A., and Alvarez-Buylla, A. (2011). Lake-front property: a unique
germinal niche by the lateral ventricles of the adult brain. Neuron 70, 674–686.
Jessell, T.M. (2000). Neuronal specification in the spinal cord: inductive signals
and transcriptional codes. Nat. Rev. Genet. 1, 20–29.
Kelsch, W., Mosley, C.P., Lin, C.W., and Lois, C. (2007). Distinct mammalian
precursors are committed to generate neurons with defined dendritic projec-
tion patterns. PLoS Biol. 5, e300. 10.1371/journal.pbio.0050300.
Kohwi, M., Osumi, N., Rubenstein, J.L., and Alvarez-Buylla, A. (2005). Pax6 is
required for making specific subpopulations of granule and periglomerular
neurons in the olfactory bulb. J. Neurosci. 25, 6997–7003.
Kohwi, M., Petryniak, M.A., Long, J.E., Ekker, M., Obata, K., Yanagawa, Y.,
Rubenstein, J.L., and Alvarez-Buylla, A. (2007). A subpopulation of olfactory
bulb GABAergic interneurons is derived from Emx1- and Dlx5/6-expressing
progenitors. J. Neurosci. 27, 6878–6891.
Kosaka, K., and Kosaka, T. (2007). Chemical properties of type 1 and type 2
periglomerular cells in the mouse olfactory bulb are different from those in
the rat olfactory bulb. Brain Res. 1167, 42–55.
Kuo, C.T., Mirzadeh, Z., Soriano-Navarro, M., Rasin, M., Wang, D., Shen, J.,
Sestan, N., Garcia-Verdugo, J., Alvarez-Buylla, A., Jan, L.Y., and Jan, Y.N.
(2006). Postnatal deletion of Numb/Numblike reveals repair and remodeling
capacity in the subventricular neurogenic niche. Cell 127, 1253–1264.
Lai, K., Kaspar, B.K., Gage, F.H., and Schaffer, D.V. (2003). Sonic hedgehog
regulates adult neural progenitor proliferation in vitro and in vivo. Nat.
Neurosci. 6, 21–27.
Liem, K.F., Jr., Jessell, T.M., and Briscoe, J. (2000). Regulation of the neural
patterning activity of sonic hedgehog by secreted BMP inhibitors expressed
by notochord and somites. Development 127, 4855–4866.
Lois, C., and Alvarez-Buylla, A. (1994). Long-distance neuronal migration in the
adult mammalian brain. Science 264, 1145–1148.
Long, F., Zhang, X.M., Karp, S., Yang, Y., and McMahon, A.P. (2001). Genetic
manipulationofhedgehogsignaling in theendochondralskeletonrevealsadirect
role in the regulationof chondrocyteproliferation.Development128, 5099–5108.
Luskin, M.B. (1993). Restricted proliferation and migration of postnatally gener-
atedneuronsderived fromthe forebrainsubventricularzone.Neuron11, 173–189.
Machold, R., Hayashi, S., Rutlin, M., Muzumdar, M.D., Nery, S., Corbin, J.G.,
Gritli-Linde, A., Dellovade, T., Porter, J.A., Rubin, L.L., et al. (2003). Sonic
hedgehog is required for progenitor cell maintenance in telencephalic stem
cell niches. Neuron 39, 937–950.
Mao, J., Ligon,K.L., Rakhlin, E.Y., Thayer, S.P., Bronson,R.T., Rowitch, D., and
McMahon, A.P. (2006). A novel somatic mouse model to survey tumorigenic
potential applied to the Hedgehog pathway. Cancer Res. 66, 10171–10178.
McLellan, J.S., Zheng, X., Hauk, G., Ghirlando, R., Beachy, P.A., and Leahy,
D.J. (2008). The mode of Hedgehog binding to Ihog homologues is not
conserved across different phyla. Nature 455, 979–983.
262 Neuron 71, 250–262, July 28, 2011 ª2011 Elsevier Inc.
McMahon, A.P., Ingham, P.W., and Tabin, C.J. (2003). Developmental roles
and clinical significance of hedgehog signaling. Curr. Top. Dev. Biol. 53, 1–114.
Merkle, F.T., Mirzadeh, Z., and Alvarez-Buylla, A. (2007). Mosaic organization
of neural stem cells in the adult brain. Science 317, 381–384.
Mirzadeh, Z., Merkle, F.T., Soriano-Navarro, M., Garcia-Verdugo, J.M., and
Alvarez-Buylla, A. (2008). Neural stem cells confer unique pinwheel architec-
ture to the ventricular surface in neurogenic regions of the adult brain. Cell
Stem Cell 3, 265–278.
Mirzadeh, Z., Doetsch, F., Sawamoto, K., Wichterle, H., and Alvarez-Buylla, A.
(2010). The subventricular zone en-face: wholemount staining and ependymal
flow. J. Vis. Exp. 39, 1938.
Palma, V., Lim, D.A., Dahmane, N., Sanchez, P., Brionne, T.C., Herzberg, C.D.,
Gitton, Y., Carleton, A., Alvarez-Buylla, A., and Ruiz i Altaba, A. (2005). Sonic
hedgehog controls stem cell behavior in the postnatal and adult brain.
Development 132, 335–344.
Pfaffl, M.W., Horgan, G.W., and Dempfle, L. (2002). Relative expression soft-
ware tool (REST) for group-wise comparison and statistical analysis of relative
expression results in real-time PCR. Nucleic Acids Res. 30, e36.
Ribes, V., and Briscoe, J. (2009). Establishing and interpreting graded Sonic
Hedgehog signaling during vertebrate neural tube patterning: the role of nega-
tive feedback. Cold Spring Harb Perspect Biol 1, a002014.
Rohatgi, R., Milenkovic, L., and Scott, M.P. (2007). Patched1 regulates
hedgehog signaling at the primary cilium. Science 317, 372–376.
Schuller, U., Heine, V.M., Mao, J., Kho, A.T., Dillon, A.K., Han, Y.G., Huillard,
E., Sun, T., Ligon, A.H., Qian, Y., et al. (2008). Acquisition of granule neuron
precursor identity is a critical determinant of progenitor cell competence to
form Shh-induced medulloblastoma. Cancer Cell 14, 123–134.
Shen, Q., Wang, Y., Kokovay, E., Lin, G., Chuang, S.M., Goderie, S.K.,
Roysam, B., and Temple, S. (2008). Adult SVZ stem cells lie in a vascular niche:
a quantitative analysis of niche cell-cell interactions. Cell StemCell 3, 289–300.
Tavazoie, M., Van der Veken, L., Silva-Vargas, V., Louissaint, M., Colonna, L.,
Zaidi, B., Garcia-Verdugo, J.M., and Doetsch, F. (2008). A specialized vascular
niche for adult neural stem cells. Cell Stem Cell 3, 279–288.
Tenzen, T., Allen, B.L., Cole, F., Kang, J.S., Krauss, R.S., and McMahon, A.P.
(2006). The cell surface membrane proteins Cdo and Boc are components and
targets of the Hedgehog signaling pathway and feedback network in mice.
Dev. Cell 10, 647–656.
Traiffort, E., Charytoniuk, D., Watroba, L., Faure, H., Sales, N., and Ruat, M.
(1999). Discrete localizations of hedgehog signalling components in the devel-
oping and adult rat nervous system. Eur. J. Neurosci. 11, 3199–3214.
Traiffort, E., Moya, K.L., Faure, H., Hassig, R., and Ruat, M. (2001). High
expression and anterograde axonal transport of aminoterminal sonic
hedgehog in the adult hamster brain. Eur. J. Neurosci. 14, 839–850.
Ventura, R.E., and Goldman, J.E. (2007). Dorsal radial glia generate olfactory
bulb interneurons in the postnatal murine brain. J. Neurosci. 27, 4297–4302.
Whitman, M.C., and Greer, C.A. (2009). Adult neurogenesis and the olfactory
system. Prog. Neurobiol. 89, 162–175.
Xu, Q., Tam,M., and Anderson, S.A. (2008). Fatemapping Nkx2.1-lineage cells
in the mouse telencephalon. J. Comp. Neurol. 506, 16–29.
Xu, Q., Guo, L., Moore, H., Waclaw, R.R., Campbell, K., and Anderson, S.A.
(2010). Sonic hedgehog signaling confers ventral telencephalic progenitors
with distinct cortical interneuron fates. Neuron 65, 328–340.
Yang, Y.S., and Hughes, T.E. (2001). Cre stoplight: a red/green fluorescent
reporter of Cre recombinase expression in living cells. Biotechniques 31,
1036, 1038, 1040–1041.
Young, K.M., Fogarty, M., Kessaris, N., and Richardson, W.D. (2007).
Subventricular zone stem cells are heterogeneous with respect to their embry-
onic origins and neurogenic fates in the adult olfactory bulb. J. Neurosci. 27,
8286–8296.
Zhao, C., Deng, W., and Gage, F.H. (2008). Mechanisms and functional impli-
cations of adult neurogenesis. Cell 132, 645–660.




![u[sonic] Modbus - Lambrecht meteo](https://static.fdokumen.com/doc/165x107/6334bd04a6138719eb0b33dc/usonic-modbus-lambrecht-meteo.jpg)














