
Proteoglycan interactions with Sonic Hedgehog specify mitogenic responses
22
Proteoglycan interactions with Sonic Hedgehog specify mitogenic responses Jennifer A. Chan 1,2,3,7 , Srividya Balasubramanian 1,7 , Rochelle M. Witt 1,7 , Kellie J. Nazemi 1,4 , Yoojin Choi 1,5 , Maria F. Pazyra-Murphy 1 , Carolyn O. Walsh 1 , Margaret Thompson 6 , and Rosalind A. Segal 1,8 1 Neurobiology Department, Harvard Medical School, 180 Longwood Avenue, Boston, MA 02115; Pediatric Oncology Department, Dana-Farber Cancer Institute, 44 Binney Street, Boston MA 02115 2 Pathology Department, Brigham and Women’s Hospital, 75 Francis Street, Boston, MA 02115 6 Neurology Department, Children’s Hospital, 300 Longwood Avenue, Boston MA 02115 SUMMARY Sonic Hedgehog (Shh) has dual roles in vertebrate development, as it promotes progenitor cell proliferation and induces tissue patterning. Here we show mitogenic and patterning functions of Shh can be uncoupled from one another. Using a genetic approach to selectively inhibit Shh- proteoglycan interactions in a mouse model, we show binding of Shh to proteoglycans is required for proliferation of neural stem/precursor cells but not for tissue patterning. Shh-proteoglycan interactions regulate both spatial and temporal features of Shh signaling. Proteoglycans localize Shh to specialized mitogenic niches and also act at the single cell level to regulate the duration of Shh signaling, thereby promoting a gene expression program important for cell division. As activation of the Shh pathway is a feature of diverse human cancers, selective stimulation of proliferation by Shh-proteoglycan interactions may also figure prominently in neoplastic growth. Development of complex tissues requires concomitant growth and cell fate specification. One mechanism for achieving spatial and temporal coordination of size and form is to utilize the same signaling molecules for both processes. The morphogen Hedgehog (Hh), and its mammalian counterparts Sonic, Desert and Indian Hedgehog (Shh, Dhh and Ihh), are critical in the growth and patterning of developing embryos1. One model for morphogen activity postulates Hedgehog proteins disperse from a localized source and form a gradient that patterns fields of responsive cells 2. Controlled ligand distribution may also localize stem cell proliferation to specialized niches3 , 4. By influencing patterning and proliferation, Hedgehog proteins could coordinate form and size to allow scaling of developing organisms. Here we asked whether proteoglycans differentially regulate Shh-dependent proliferation and patterning. Genetic evidence in Drosophila indicates proteoglycans are required for Hh dispersal and gradient formation5 , 6. Biochemical evidence indicates proteoglycans bind to 8 To whom correspondence should be addressed: Dana-Farber Cancer Institute, 44 Binney Street, Boston MA 02115, 617-632-4737, [email protected]. 3 Current address: Department of Pathology & Laboratory Medicine, University of Calgary, 3330 Hospital Drive NW, Calgary, Alberta, T2N 4N1, Canada 4 Current address: Division of Pediatric Hematology/Oncology, Doernbecher Children’s Hospital, Oregon Health & Science University, 10th floor 3181 SW Sam Jackson Park Road, DC10C, Portland, OR 97239 5 Current address: Science Department, Phillips Exeter Academy, 20 Main Street, Exeter, NH 03833 7 These authors contributed equally. See Supplement for detailed methods. NIH Public Access Author Manuscript Nat Neurosci. Author manuscript; available in PMC 2009 October 1. Published in final edited form as: Nat Neurosci. 2009 April ; 12(4): 409–417. doi:10.1038/nn.2287. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Proteoglycan interactions with Sonic Hedgehog specify mitogenic responses

Proteoglycan interactions with Sonic Hedgehog specifymitogenic responses
Jennifer A. Chan1,2,3,7, Srividya Balasubramanian1,7, Rochelle M. Witt1,7, Kellie J.Nazemi1,4, Yoojin Choi1,5, Maria F. Pazyra-Murphy1, Carolyn O. Walsh1, MargaretThompson6, and Rosalind A. Segal1,8
1 Neurobiology Department, Harvard Medical School, 180 Longwood Avenue, Boston, MA 02115;Pediatric Oncology Department, Dana-Farber Cancer Institute, 44 Binney Street, Boston MA021152 Pathology Department, Brigham and Women’s Hospital, 75 Francis Street, Boston, MA 021156 Neurology Department, Children’s Hospital, 300 Longwood Avenue, Boston MA 02115
SUMMARYSonic Hedgehog (Shh) has dual roles in vertebrate development, as it promotes progenitor cellproliferation and induces tissue patterning. Here we show mitogenic and patterning functions ofShh can be uncoupled from one another. Using a genetic approach to selectively inhibit Shh-proteoglycan interactions in a mouse model, we show binding of Shh to proteoglycans is requiredfor proliferation of neural stem/precursor cells but not for tissue patterning. Shh-proteoglycaninteractions regulate both spatial and temporal features of Shh signaling. Proteoglycans localizeShh to specialized mitogenic niches and also act at the single cell level to regulate the duration ofShh signaling, thereby promoting a gene expression program important for cell division. Asactivation of the Shh pathway is a feature of diverse human cancers, selective stimulation ofproliferation by Shh-proteoglycan interactions may also figure prominently in neoplastic growth.
Development of complex tissues requires concomitant growth and cell fate specification.One mechanism for achieving spatial and temporal coordination of size and form is to utilizethe same signaling molecules for both processes. The morphogen Hedgehog (Hh), and itsmammalian counterparts Sonic, Desert and Indian Hedgehog (Shh, Dhh and Ihh), are criticalin the growth and patterning of developing embryos1. One model for morphogen activitypostulates Hedgehog proteins disperse from a localized source and form a gradient thatpatterns fields of responsive cells 2. Controlled ligand distribution may also localize stemcell proliferation to specialized niches3,4. By influencing patterning and proliferation,Hedgehog proteins could coordinate form and size to allow scaling of developing organisms.
Here we asked whether proteoglycans differentially regulate Shh-dependent proliferationand patterning. Genetic evidence in Drosophila indicates proteoglycans are required for Hhdispersal and gradient formation5,6. Biochemical evidence indicates proteoglycans bind to
8To whom correspondence should be addressed: Dana-Farber Cancer Institute, 44 Binney Street, Boston MA 02115, 617-632-4737,[email protected] address: Department of Pathology & Laboratory Medicine, University of Calgary, 3330 Hospital Drive NW, Calgary,Alberta, T2N 4N1, Canada4Current address: Division of Pediatric Hematology/Oncology, Doernbecher Children’s Hospital, Oregon Health & ScienceUniversity, 10th floor 3181 SW Sam Jackson Park Road, DC10C, Portland, OR 972395Current address: Science Department, Phillips Exeter Academy, 20 Main Street, Exeter, NH 038337These authors contributed equally.
See Supplement for detailed methods.
NIH Public AccessAuthor ManuscriptNat Neurosci. Author manuscript; available in PMC 2009 October 1.
Published in final edited form as:Nat Neurosci. 2009 April ; 12(4): 409–417. doi:10.1038/nn.2287.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Hh and the receptor component Ihog to affect Hh activity7,8. In Drosophila, proteoglycanscritical for Hh functions are heparan sulfate proteoglycans (HSPGs), and the core proteincomponents are glypicans (GPI-linked proteins) 6,9–12. Dally-like glypicans are requiredfor Hedgehog and Wingless dispersal and responses, modulating signaling of bothligands13. However, how HSPGs affect Hedgehog signaling and biological responses arenot yet understood.
Proteoglycans (PGs) have also been implicated in Hedgehog-pathway signaling inmammalian systems11,12,14. Loss of glypican GPC3 causes an overall growth increase,reflecting changes in Shh and/or IGF signaling11, while mutations in HSPG synthesizingenzymes cause dramatic defects that may reflect changes in FGF, Wnt, and/or Shhresponses15,16. To investigate the physiologic role of Shh-proteoglycan interactions, wetook the alternative approach of mutating Shh itself. Shh contains an N-terminal Cardin-Weintraub motif that mediates Shh-proteoglycan interactions (KRRHPKK). As mutations inthis motif (R34A/K38A, designated ShhAla) reduce high-affinity Shh-proteoglycaninteractions without altering Shh’s affinity for its receptor Patched (Ptc)14, we can use thismutation to investigate Shh-HSPG interactions, without confounding effects due to othergrowth factors that bind proteoglycans.
To identify on an organismal level responses that require Shh-proteoglycan interactions wegenerated mice in which wild type Shh is replaced with ShhAla. While Shh is needed forgrowth and patterning of diverse tissues, we find proteoglycan interactions selectively affectShh-induced proliferation rather than Shh-induced patterning. Proteoglycans localize Shhligand to mitogenic niches in developing brain to promote proliferation of neural stem/precursor cells. Proteoglycans also function at the level of individual responding cells; cell-associated proteoglycans bind Shh and alter ligand perdurance. In this way, Shh-proteoglycan interactions stimulate expression of bmi-1, D-type Cyclins and other genesimplicated in proliferation and neoplastic growth17, 18. These studies indicate that HSPGsselectively promote mitogenic responses to Shh.
RESULTSHSPG binding motif mutations affect Shh-PG interactions
We identified a conserved Cardin-Weintraub motif in Shh that is responsible for highaffinity binding of Shh to HSPGs14. To determine whether mutations in this motif can beused to identify functions of Shh-proteoglycan interactions, we quantitatively evaluated Shhbinding to heparin. We assessed binding of equal amounts of wild type Shh or ShhAla toheparin sulfate-coated plates using alkaline phosphatase tagged Shh isoforms. We measuredbinding in the absence, or presence of increasing amounts of soluble heparin sulfate. Overallbinding of mutant Shh was significantly less than wild type. Furthermore, higherconcentrations of soluble heparin sulfate were required to abolish binding of wild type Shhto immobilized heparin than concentrations needed to abolish binding of ShhAla (Fig. 1a).Binding of ShhAla to endogenous proteoglycans was also strikingly impaired. Wild typeShh, but not ShhAla, binds endogenous proteoglycans in CNS tissue sections (14, Fig. 1b).Pre-treatment of sections with heparinases, which remove heparan sulfates, or PI-PLC,which removes GPI-linked molecules, prevented binding of wild type Shh to tissueproteoglycans (Fig. 1b, Supplementary Fig. 1). As glypicans are HSPGs with a GPI-linkage,these results suggest that wild type Shh binds glypican proteoglycans in the developingcerebellum and that the ShhAla mutation interferes with binding to these endogenousglypicans in developing cerebellum. We next introduced the ShhAla mutation into a fulllength Shh expression vector and produced protein in HEK cells. The ShhAla precursorundergoes proteolytic cleavage to generate a mature isoform of the correct size (Fig. 1c)that, like wild type Shh, is palmitoylated (Fig. 1d)19. We previously demonstrated that
Chan et al. Page 2
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

ShhAla binds Ptc with similar affinity as wild type ligand 14. Together, these data showShhAla can be used as a specific reagent to determine functions of Shh-proteoglycaninteractions.
Using homologous recombination, we generated knock-in mice in which endogenous Shhwas replaced with ShhAla (Supplementary Fig. 2). With this targeting strategy, a loxP siteremained within the intron between exons 1 and 2. Because intronic elements can affect Shhexpression, we also generated a line of control animals wherein the only alteration is thisloxP site (ShhCtl); ShhCtl/Ctl animals are indistinguishable from wild type. Furthermore, theShhAla mutation itself does not alter the expression of Shh protein in vivo: Shh levels areindistinguishable in Shh+/+, ShhAla/+ and ShhAla/Ala tissues (Supplementary Fig. 2e, f). Thisnovel genetic approach enables us to determine the role of Shh binding to proteoglycanswithout interference from proteoglycan-dependent modulation of other growth factorpathways.
Shh-PG interactions affect growth but not patterningShh−/− animals are embryonic lethal and display holoprosencephaly and limb patterning -defects20. Hypomorphic alleles of Shh usually alter size and pattern19. In contrast,homozygous ShhAla/Ala mice only exhibited growth defects and showed no patterningdefects. While Shh is critical in generating and patterning diverse organs, mutant animalswere viable and fertile, with all organs present and correctly localized. In contrast,ShhAla/Ala mice displayed multiple growth abnormalities. Overall size was reduced inmutant animals, with particular differences in size of the brain, spinal cord and eyes. Bodyweight of ShhAla/Ala mice was 11% less than wild type or ShhCtl/Ctl animals, and brainweight was reduced by 13% (p=0.0018). There was, however, disproportionate hypoplasiaof cerebellum and olfactory bulb (Fig. 2a). In contrast to these clear differences in tissuegrowth, when we examined areas where Shh is critical for pattern formation1,21,22, wefound ShhAla/Ala mice displayed normal digit number and shape, and the spinal cord laminaewere correctly formed (Fig. 2a,b, Supplementary Fig. 3). Furthermore, whileholoprosencephaly and cyclopia are cardinal features of Shh−/− mice20, ShhAla/Ala animalsdisplayed normal separation of cerebral ventricles and two well-spaced (but small) eyes(Fig. 2a). Together these findings indicate that Shh-proteoglycan interactions mediated bythe Cardin-Weintraub motif are specifically required for growth regulation but not forpatterning activities.
In mice, one copy of Shh is sufficient for normal development as Shh+/− mice exhibitnormal patterning and growth20. One copy of ShhAla is also sufficient for patterning, asShhAla/− mice were viable with normal limbs and digits. However, even more dramaticchanges in growth were evident in ShhAla/− than in ShhAla/Ala animals (Fig. 2c). The overallsize of ShhAla/− mice, and in particular, cerebellar and olfactory bulb size were greatlyreduced compared to Shh+/− animals. However, even when one copy of ShhAla is present, nodefects were seen in tissue patterning (Fig. 2c). The characterization of ShhAla/Ala andShhAla/− mice underscores the selectivity of the phenotype, and indicates Shh-proteoglycaninteractions particularly regulate tissue growth.
Shh-PG interactions promote stem/precursor proliferationTo determine why proteoglycan interactions are critical for tissue growth, we focused on thecerebellum, as cerebella of ShhAla/Ala mice are one third smaller than their wild typelittermates. As Shh regulates cell division and cell death in the nervous system23,24, weasked whether the decrease in cerebellar size in ShhAla/Ala and ShhAla/− mice reflectsabnormal proliferation or apoptosis. In early postnatal life, granule cell precursors (GCPs)undergo extensive proliferation in the external granule cell layer (EGL) then exit the cell
Chan et al. Page 3
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

cycle and migrate inwards past the Purkinje cell layer to reside in the internal granule celllayer (IGL). Previous studies have shown Purkinje cell-synthesized Shh promotes GCPproliferation23,25. In ShhAla/Ala mice, GCP proliferation in the EGL is approximately 30%less than in wild type, as detected by percentage of EGL cells positive for M-phase markerphospho-Histone H3 and by S-phase labeling using bromodeoxyuridine (BrdU) (Fig. 3a–c).This decreased proliferation, which was seen throughout the cerebellum (Fig. 3d), indicatesShh-proteoglycan interactions promote mitogenesis of cerebellar precursors during earlypostnatal life across the rostral-caudal axis of the developing hindbrain.
Ext1 and Ext2 are required for HSPG chain elongation, and so are necessary for generatingglycan chains that interact with Shh. While we previously identified developmental changesin expression of ext1 and ext2 in BALB/c mice during the early postnatal period14, the 129/C57BL6 strain used here exhibited consistent expression of ext1 and ext2 in the postnatalperiod when proliferation is reduced in ShhAla/Ala (Supplementary Fig. 4). Many other genesrequired for synthesizing and modifying glycans are also appropriately expressed at thistime26. Thus, Shh and proteoglycans are expressed appropriately to regulate GCPproliferation, which is reduced when Shh-proteoglycan interactions are impaired.
The subventricular zone (SVZ) adjacent to the lateral ventricles and the subgranular layer(SGL) of the hippocampus represent two additional locations where there is significant Shh-regulated postnatal proliferation of neural stem/precursor cells3,4,27, while the embryonicspinal cord represents a site where Shh regulates prenatal expansion of neural stem/precursors28. Neural precursor proliferation in the spinal cord was reduced in ShhAla/Ala
embryos as compared to Shh+/+ littermates (Fig. 4a,b). Proliferation of neural stem/precursors in the SVZ and SGL of adult mutants was also decreased compared to age- andsex-matched ShhCtl/Ctl (Fig. 4c,d). As stem/precursor cells in the SVZ generate olfactorybulb neurons, decreased SVZ proliferation may contribute to the olfactory bulb hypoplasia(Fig. 2, Supplementary Fig. 5). Together, these findings demonstrate that Shh-proteoglycaninteractions promote neural stem/precursor cell proliferation in multiple locations in both thedeveloping and adult CNS, and thus highlight the widespread importance of proteoglycansin mitogenesis.
Surprisingly, apoptosis was decreased in the EGL of ShhAla/Ala mutants compared with wildtype littermates (Supplementary Fig. 6). As the Shh pathway can stimulate or suppressapoptosis under different conditions24,29, it is possible that pro-apoptotic effects of Shhrequire proteoglycan interactions, while anti-apoptotic effects do not. Alternatively,decreased apoptosis in ShhAla/Ala mice may be an indirect consequence of the mutation. Ineither case, the increased survival did not mitigate the proliferation decrease, and the netresult was a dramatically smaller cerebellum (30% smaller in ShhAla/Ala and 50% smaller inShhAla/−).
PGs delineate the Shh mitogenic niche in the EGLThere are two potential mechanisms to explain the effects of proteoglycans on Shh-inducedproliferation. One is that proteoglycan interactions localize Shh to a mitogenic niche.Previous studies have suggested that proteoglycans regulate Hh dispersal5,9,12,13 andlocalize Ihh in developing bone30. Thus, without proteoglycan binding, Shh might notcollect in the EGL, resulting in decreased proliferation. A second is that binding of Shh toproteoglycans on the surface of GCPs modulates Shh-dependent intracellular signalcascades7,13. As these models are not mutually exclusive, we investigated both.
We previously developed an assay to evaluate the nature and location of mitogenic niches inneural tissues31. In this assay, GFP-labeled precursors are introduced onto organotypicslices and incorporate into the slices. We then incubate the slice culture with BrdU to label
Chan et al. Page 4
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

proliferating cells, and measure the proliferative index of GFP+ precursors in distinctmicroenvironments. Precursors exposed to the cerebellar EGL proliferate extensively due tomitogenic effects of Shh31. To determine whether Shh-proteoglycan interactions areimportant in delineating this niche, we introduced wild type GFP-labeled precursors or DiO-labeled ShhAla/Ala precursors onto slices cultured from wild type or ShhAla/Ala littermates.While wild type or ShhAla/Ala precursor proliferation is enhanced when cells are introducedonto a wild type EGL, the EGL of ShhAla/Ala mice does not provide a mitogenic niche forwild type precursors (Fig. 5a). To determine whether decreased proliferation is due toimpaired proteoglycan interactions, we added heparan sulfates to cultures of wild typeprecursors on wild type slices. Excess glycans can compete with, and so diminish the actionsof endogenous proteoglycans30. Excess heparan sulfates phenocopied the ShhAla mutation,as there was decreased proliferation of cells introduced onto the EGL (Fig. 5b). Togetherthese data indicate that interactions between Shh’s Cardin-Weintraub motif and endogenousproteoglycans promote neural precursor proliferation and establish the EGL mitogenicniche.
In slice overlay cultures, proliferation of introduced precursor cells was decreased in theEGL but increased in the IGL of ShhAla/Ala slices (Fig. 5a). These data suggest thatproteoglycan binding enables Shh to accumulate in the EGL rather than in other layers. Totest this, we carried out immunohistochemistry for Shh. Consistent with previous studies31,
32 in wild type animals, Shh immunostaining was detected in Purkinje cells, and in the IGLand EGL (Fig. 5c, Supplementary Fig. 7). In ShhAla/Ala animals, Shh immunostaining wasdecreased in the outer EGL (EGLa) where proliferation occurs (Fig. 5c). These findingsindicate proteoglycans localize Shh to a mitogenic niche within the outer EGL. Electronmicroscopy studies have identified a proteoglycan matrix in EGLa just under the pia andadjacent to proliferating GCPs33. Thus one mechanism by which Shh-HSPG interactionsstimulate precursor proliferation is by localizing ligand to the EGL mitogenic niche.
Proteoglycan interactions modulate responses to ShhWhile the data above indicate Shh-proteoglycan interactions are important for localizing Shhto proliferative zones and so establishing a mitogenic niche, it is also possible proteoglycanson neural stem/precursors bind Shh, thereby specifying proliferative responses. To addressthis possibility, we asked whether proteoglycans on GCPs affect Shh-induced responses atthe level of an individual responding cell. Consistent with previous studies14,23,25,34,35,wild type Shh induces robust GCP proliferation. In dissociated cell cultures, the proliferativeresponse evoked by wild type Shh was much greater than that evoked by equivalentconcentrations of ShhAla (Fig. 6a). In contrast, no consistent effects on survival were seenwhen dissociated GCP cultures were stimulated with either wild type Shh or ShhAla (Fig.6b). Thus, the decreased proliferation observed in ShhAla/Ala mice is a direct consequence ofthe Cardin-Weintraub mutation, while decreased apoptosis may be an indirect result of themutation. These data demonstrate Shh interacts with proteoglycans on individual GCPs,thereby inducing cells to proliferate, and suggest that the decreased proliferation observed inShhAla/Ala mice results both from altered localization of ligand and from changes inintracellular signaling cascades.
Shh responses are mediated by the transcription factors Gli1, Gli2 and Gli336. Therefore,we analyzed expression of these factors in ShhAla/Ala mice. While Gli transcription factorscan function either as transcriptional activators or repressors37, Gli1 only functions as anactivator, and only one isoform of Gli1 has been reported38. Gli1 protein levels wereequivalent in P3 ShhAla/Ala and Shh+/+ animals (Fig. 7a). Gli2 can function as activator andrepressor, and distinct isoforms have been identified that subserve these differentfunctions38,39. In cerebella of mutant animals, the ratio of Gli2 activator (GliAct) to theshorter Gli2 repressor (Gli2Rep) was reduced by 50.9% (Fig. 7b). These data suggest overall
Chan et al. Page 5
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Gli2-dependent transcription is likely to be altered in mutants. Previous studies haveindicated Gli3 functions primarily as a repressor40. We did not detect any differences inGli3Rep levels, and we did not detect Gli3Act in cerebellar tissue of either wild type ormutant animals (Fig. 7c). Thus Gli2 isoforms represent the Shh-dependent transcriptionfactor(s) that are clearly altered in ShhAla/Ala cerebellum. As Gli2 activity is required tomediate expansion of GCPs41, changes in Gli2 may potentially explain the observedShhAla/Ala phenotype.
Since the Gli2 activator/repressor ratio is altered in vivo in ShhAla/Ala cerebella and Gli2 hasbeen shown to play a critical role in GCP proliferation41, we examined in greater detail Gli2protein levels in GCPs acutely stimulated with wild type or mutant Shh. We found thatGCPs stimulated with either Shh or ShhAla exhibited an increase in Gli2Act protein (9.7%increase and 34.7% increase for Shh- and ShhAla-stimulated GCPs respectively). However,consistent with in vivo results, the Gli2Act/Gli2Rep ratio in GCPs acutely stimulated withShhAla was lower than that seen in GCPs stimulated with Shh (Fig. 7d). Furthermore, thetemporal profiles of Gli2Act and Gli2Rep differed when GCPs were stimulated with ShhAla
rather than wild type Shh (Fig. 7e, Supplementary Fig. 8) (i.e average Gli2Act levelsdeclined by 3.7% and 31.4% between 24 and 32 hours post-stimulation by Shh and ShhAla
respectively). These data indicate Shh interacts with proteoglycans on GCPs to alter thenature and timing of Gli2-dependent transcription.
PGs alter Shh-dependent gene expression and Shh kineticsChanges in the Gli2 activator/repressor ratio are likely to alter expression of Shh-responsivegenes. A number of Shh-responsive genes have been identified, including genes involved inproliferation and tissue patterning17,34,35, 42–44. We analyzed Shh-responsive geneexpression when dissociated wild type GCPs were stimulated acutely with equalconcentrations of wild type Shh or ShhAla. Shh-dependent genes can be separated into threeclusters that are differentially modulated by proteoglycan interactions (Fig. 8a). One cluster,including gli1 and ptc1, are similarly induced by ShhAla and wild type Shh. A second clusterof target genes (gli2 and N-myc) is induced to a greater extent by ShhAla than by wild typeShh. Surprisingly, expression of N-myc and gli2, genes implicated in proliferation, are in thegene set that is better induced by ShhAla. We note that Shh regulates both Gli2 and N-mycvia transcriptional and post-transcriptional mechanisms 39, 45, and so Gli2 and N-mycactivity need not correspond to RNA levels. Induction of a third cluster, including gli3,Cyclin D1, Cyclin D2, and bmi-1, was diminished when cells were stimulated with ShhAla
rather than wild type ligand. Cyclins D1 and D2 are cell cycle regulators at the G1/Stransition that are induced in response to Shh stimulation35 and have been particularlyimplicated in Shh-induced stem/precursor cell proliferation35. The polycomb protein,Bmi-1, is a Shh target that is critical for self-renewal of stem cells and for cancer cellproliferation17,18,44. Together, these data indicate the gene expression program induced byShh is modulated by proteoglycans of responding cells. Moreover, Shh target genes that arealtered when cells are stimulated by ShhAla include many implicated in proliferation andoncogenesis.
Previous studies have demonstrated distinct responses to Shh can be elicited depending onligand concentration22. To determine whether a shift in the dose response curve explainswhy ShhAla induces an altered program of gene expression, we performed dose responseexperiments using Shh or ShhAla protein. C3H10T1/2 differentiation as assessed by alkalinephosphatase induction (Supplementary Fig. 9a) is a standard assay for Shh activity; wefound no shift in the ShhAla dose response curve compared to Shh. Furthermore, maximalefficacy of ShhAla was not reduced compared to Shh. We next measured gli1 and CyclinD2mRNA levels in GCPs stimulated across a wide range of Shh and ShhAla doses(Supplementary Fig. 9b). Whereas gli1 expression did not differ significantly in response to
Chan et al. Page 6
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

equivalent doses of Shh and ShhAla throughout the dose range, CyclinD2 induction byShhAla was diminished regardless of dose. These data indicate the altered biological activityof ShhAla does not reflect a shift in the dose response curve. As recent studies havehighlighted the importance of Shh signaling kinetics in determining Shh responses46, weasked whether proteoglycan interactions alter the ability of Shh to signal over prolongedperiods of time. Equivalent amounts of Shh or ShhAla were added to either C3H10T1/2 orGCP cultures. After twenty-four hours of stimulation, we removed ligand from the mediaand analyzed Shh perdurance in the stimulated cells over the ensuing two days. In bothsystems, ShhAla levels declined more precipitously than wild type (Fig. 8b). Taken together,these data indicate that proteoglycans alter the kinetics of signaling, promoting a geneexpression signature important for Shh-dependent precursor proliferation.
To determine whether changes in Shh-dependent gene expression explain the in vivophenotype of ShhAla/Ala mice, we used quantitative RT-PCR to compare Shh-responsivegene expression in wild type and mutant cerebella. gli1 and ptc1 levels were equivalent inwild type and ShhAla/Ala cerebella. However, expression of a second set of genes includingCyclin D1, Cyclin D2, and bmi-1 was significantly reduced in ShhAla/Ala animals comparedto wild type littermates (Fig. 8c). Decreased expression of CyclinD1, D2 and bmi-1 do notreflect a generalized decrease in cell cycle-associated proteins, as CyclinE expression wasunchanged. A third set of Shh-regulated genes (gli2 and N-myc) was expressed at higherlevels in ShhAla/Ala than in wild type mice. The observed changes in vivo do not reflectalterations in the distribution of cells expressing Shh-responsive genes as indicated by in situhybridization studies for gli1, gli2 and gli3 (Supplementary Fig. 10). A critical feature ofthese data is that proteoglycan interactions do not uniformly increase or decrease Shh-dependent gene expression in vivo. Furthermore, clusters that are unchanged, increased, ordecreased in ShhAla/Ala animals exhibit a striking similarity to gene expression patterns inGCPs acutely stimulated with wild type versus mutant Shh.
Taken together, these studies indicate Shh-proteoglycan interactions selectively promoteneural stem/precursor proliferation by two mechanisms. First proteoglycans localize theligand within developing tissue and so establish the mitogenic niche, and second they alterligand perdurance, preferentially activating intracellular cascades that culminate inmitogenesis and precursor renewal.
DISCUSSIONUsing a genetic approach, we find that Shh-proteoglycan interactions are required forproliferative, rather than patterning responses to Shh. We generated mutant mice thatexpress Shh that cannot bind proteoglycans but can bind to Ptc. Mutants exhibit a selectivedeficit in neural stem/precursor cell proliferation. We identified two distinct activities ofproteoglycans in Shh-dependent proliferation: these molecules localize Shh to mitogenicniches and they also trigger a gene expression program important for cell division and stemcell renewal.
Proteoglycans are tremendously diverse; each proteoglycan represents a complex, yet poorlyunderstood readout of the glycogenes, the genes encoding core proteins, sugar transporters,glycosyltransferases, sulfatases, and acetylating enzymes. As proteoglycans interact withmany growth factors and other molecules, analyses of mutants that interfere with individualsteps of proteoglycan synthesis are difficult to interpret. To identify the functions ofproteoglycans for Shh responses in vivo, we took an alternative approach. Mutations in theCardin-Weintraub motif of Hh proteins interfere with proteoglycan binding, but do not alterPtc affinity, lipid modifications, or expression level. Therefore ShhAla/Ala and ShhAla/−
Chan et al. Page 7
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

mutants provide a unique genetic approach for identifying functions of Shh-proteoglycaninteractions.
We find proteoglycans are specifically needed for proliferative responses to Shh, but aredispensable for most of its patterning activities. One intriguing finding is that proteoglycanslocalize Shh to proliferative zones, and also function at the single cell level to determine thenature of the response. Previous studies using mutations in the HSPG synthesizing enzymesof the ext gene family, mutations in the glypican Dally-like or deletion of the Cardin-Weintraub motif have indicated that HSPGs are critical for appropriate localization of Hhproteins 11–13, 47. Here we show that Shh that cannot bind to HSPGs do not accumulate inthe EGL mitogenic niche. Thus, direct interactions of Shh with proteoglycans are importantfor Shh dispersal from the Purkinje cell layer and/or its sequestration in the EGL. AlteredShh localization in mutant animals impairs the establishment of mitogenic niches indeveloping brain.
In addition to their function in localizing Shh, we find that proteoglycans on receiving cellsmodulate the Shh response. Previous studies have indicated that proteoglycans function inHh-responding cells7,13. Surprisingly, some groups have found that glypicans on receivingcells are needed for full-strength signaling while others have found that responding cellglypicans compete with Ptc1 for Hh interaction, thereby inhibiting Hh activity11,13. Ourstudies suggest proteoglycan interactions differentially affect intracellular signaling cascadesdownstream of Smoothened, and so selectively regulate Shh target gene subsets, therebymodifying the response.
As Shh target gene expression is primarily regulated by the Gli transcription factors36, weexamined levels and isoforms of Gli1, Gli2 and Gli3 proteins in wild type and mutantanimals. Whereas there were no discernable differences between Gli1 and Gli3 proteinlevels of wild type and mutant cerebella, clear differences were seen in Gli2 proteins. Gli2 isthe major transcription factor driving Shh-induced proliferation41,48 and proliferation isreduced in multiple locations pre- and post-natally in ShhAla/Ala animals. Gli2 proteins canfunction as either transcriptional activators or repressors; unprocessed Gli2 functions as aweak transcriptional activator37,39, while Gli2 cleavage and removal of the C-terminalactivator domain generates a transcriptional repressor39. Therefore both Gli2 protein levelsand the ratio between unprocessed and processed forms are likely to be critical for the netGli2-dependent transcriptional response. In vivo, gli2 mRNA is lower in wild type than inShhAla/Ala cerebella, but the Gli2 activator/repressor ratio is two-fold greater in wild type.Similarly, gli2 mRNA levels are lower in GCPs acutely stimulated with Shh than ShhAla,but GCPs acutely stimulated with Shh had a larger Gli2 activator/repressor ratio than thosestimulated with ShhAla.
Gli2 expression and processing are both Shh-regulated39,48. The ability of proteoglycans toaffect the levels and proportions of the Gli2 isoforms in vivo and in vitro would alter Shh-dependent transcriptional activation and repression, and so could account for increases anddecreases in Shh-target gene expression. One set of Shh-regulated genes, including classicaltargets ptc1 and gli1, are similarly induced by Shh that can and cannot bind proteoglycans.A second set, including gli2 and N-myc, are preferentially increased by Shh that cannot bindto proteoglycans. Induction of a third set of Shh-regulated genes, including bmi-1, and D-type Cyclins, genes implicated in stem cell maintenance and in tumor biology, requiresproteoglycan interactions. Differences in Shh target genes are observed in vivo and in vitroand are not due to shifts in dose response curves, but may be explained by changes in ligandsignaling kinetics. Taken together, analysis of the mutant phenotype indicates that Shh-HSPG interactions particularly affect overall Gli2 transcriptional activity, thereby promotingproliferative responses.
Chan et al. Page 8
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Changes in ligand intracellular localization or ligand perdurance provide two possiblemechanisms whereby Shh-proteoglycan interactions could selectively modulate signaling.Shh signaling kinetics are important in determining morphogen responses46. Here we findthat Shh-proteoglycan interactions affect Shh perdurance in responding cells and alter thetemporal profile of Shh signaling. It has been proposed that signal duration is proportional toShh concentration46, our data indicate that proteoglycan interactions alter that relationship.In this way, proteoglycan interactions can modulate the pattern of gene induction and thebiological response to Shh. Proliferation without appropriate patterning is a cardinal featureof tumor biology. The Hedgehog pathway may promote oncogenesis when it stimulatesproliferation without patterning. It is intriguing that glypican overexpression can augmentgrowth in cancers that depend on Shh activity, including rhabdomyosarcoma, prostatecancer, and pancreatic carcinoma49,50. Similarly, D-type Cyclins and bmi-1, gene targetsthat require Shh-HSPG interactions, play central roles in tumor stem cell biology17,18.
In summary, we have demonstrated that Shh-proteoglycan interactions are selectivelycritical for Shh-dependent mitogenesis, functioning in two ways. First, proteoglycanslocalize Shh to establish mitogenic niches. Second, proteoglycans on responsive cellsselectively promote intracellular pathways that lead to precursor proliferation. Bothmechanisms contribute to the phenotype of the ShhAla/Ala mice. Thus, Shh-proteoglycaninteractions promote proliferation within mitogenic niches by localizing Shh to theseproliferative zones and by modulating intracellular signaling cascades and transcriptionalprograms.
METHODSSection binding assay
To evaluate proteoglycan binding in situ14, cryosections were treated with PBS, vehiclecontrol, 500 mU/ml of heparinases (Sigma), or 500 mU/ml of PI-PLC for 1 h (37°C), thenO/N (4°C). Shh:AP or ShhAla:AP was added (1 h, RT), sections were washed andimmunostained with anti-alkaline phosphatase.
Size measurements6 month old animals were weighed, then sacrificed. Total brain and cerebella were weighed.Olfactory bulb dimensions (OB) were measured from 10 μm thick coronal cryosections andoverall volume was calculated. At least 3 pairs of ShhAla/Ala and age-matched ShhCtl/Ctl
were assessed.
Shh staining quantificationQuantification was done on mid-sagittal cerebellar sections from three wild type-ShhAla/Ala
littermate pairs, stained in parallel for Shh. Staining was analyzed in primary and secondaryfissures using NIH Image J software. In each fissure, three lines from the pia through theIGL were drawn perpendicular to the pia, and an intensity plot was determined for each line(plot profile function).
Proliferation and apoptosis quantification in vivoAnalyses of p-Histone H3 staining, BrdU labeling, and TUNEL were performed oncerebella from 3–5 matched wild-type and mutant littermate pairs. Proliferation or apoptoticindex was calculated by dividing the number of positive cells by the number of DAPI-stained cells in the EGL. Comparable locations (midpoint of primary, secondary and tertiaryfissures within vermis) were assessed. In coronal sections of adults, every 9th section of theSGL or SVZ was stained with p-Histone H3, BrdU, and activated caspase 3. At least 3 pairs
Chan et al. Page 9
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

of adult ShhAla/Ala animals and age-matched controls were analyzed. SGL was the inner halfof the thickness of the dentate gyrus, extending two cell widths into the hilus.
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
AcknowledgmentsSupported by NIH (JAC, RAS, SB), DFCI Mahoney Center for Neuro-Oncology (JAC), Musella Foundation(KJN), Quan Fellowship (RMW), Children’s Hospital MRDDRC and Harvard NeuroDiscovery Center. We thankJennifer Despinoy for excellent assistance; C Stiles, D Rowitch, M Greenberg and the Segal lab for helpfuldiscussions; D Rowitch, Q Ma, P Chuang, D Paul, S O’Gorman, P Silver and A McMahon for reagents.
References1. Capdevila J, Izpisua Belmonte JC. Patterning mechanisms controlling vertebrate limb development.
Annu Rev Cell Dev Biol. 2001; 17:87–132. [PubMed: 11687485]
2. Hooper JE, Scott MP. Communicating with Hedgehogs. Nature reviews. 2005; 6:306–317.
3. Machold R, et al. Sonic hedgehog is required for progenitor cell maintenance in telencephalic stemcell niches. Neuron. 2003; 39:937–950. [PubMed: 12971894]
4. Palma V, et al. Sonic hedgehog controls stem cell behavior in the postnatal and adult brain.Development. 2005; 132:335–344. [PubMed: 15604099]
5. Bellaiche Y, The I, Perrimon N. Tout-velu is a Drosophila homologue of the putative tumoursuppressor EXT-1 and is needed for Hh diffusion. Nature. 1998; 394:85–88. [PubMed: 9665133]
6. Desbordes SC, Sanson B. The glypican Dally-like is required for Hedgehog signalling in theembryonic epidermis of Drosophila. Development. 2003; 130:6245–6255. [PubMed: 14602684]
7. Lum L, et al. Identification of Hedgehog pathway components by RNAi in Drosophila culturedcells. Science. 2003; 299:2039–2045. [PubMed: 12663920]
8. Yao S, Lum L, Beachy P. The ihog cell-surface proteins bind Hedgehog and mediate pathwayactivation. Cell. 2006; 125:343–357. [PubMed: 16630821]
9. Han C, Belenkaya TY, Khodoun M, Tauchi M, Lin X. Distinct and collaborative roles of DrosophilaEXT family proteins in morphogen signalling and gradient formation. Development. 2004;131:1563–1575. [PubMed: 14998928]
10. Bornemann DJ, Duncan JE, Staatz W, Selleck S, Warrior R. Abrogation of heparan sulfatesynthesis in Drosophila disrupts the Wingless, Hedgehog and Decapentaplegic signaling pathways.Development. 2004; 131:1927–1938. [PubMed: 15056609]
11. Capurro MI, et al. Glypican-3 inhibits Hedgehog signaling during development by competing withpatched for Hedgehog binding. Dev Cell. 2008; 14:700–711. [PubMed: 18477453]
12. Hacker U, Nybakken K, Perrimon N. Heparan sulphate proteoglycans: the sweet side ofdevelopment. Nature reviews. 2005; 6:530–541.
13. Gallet A, Staccini-Lavenant L, Therond PP. Cellular trafficking of the glypican Dally-like isrequired for full-strength Hedgehog signaling and wingless transcytosis. Dev Cell. 2008; 14:712–725. [PubMed: 18477454]
14. Rubin JB, Choi Y, Segal RA. Cerebellar proteoglycans regulate sonic hedgehog responses duringdevelopment. Development. 2002; 129:2223–2232. [PubMed: 11959830]
15. Pallerla SR, Pan Y, Zhang X, Esko JD, Grobe K. Heparan sulfate Ndst1 gene function variablyregulates multiple signaling pathways during mouse development. Dev Dyn. 2007; 236:556–563.[PubMed: 17183530]
16. Inatani M, Irie F, Plump AS, Tessier-Lavigne M, Yamaguchi Y. Mammalian brain morphogenesisand midline axon guidance require heparan sulfate. Science. 2003; 302:1044–1046. [PubMed:14605369]
17. Molofsky AV, et al. Bmi-1 dependence distinguishes neural stem cell self-renewal from progenitorproliferation. Nature. 2003; 425:962–967. [PubMed: 14574365]
Chan et al. Page 10
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

18. Pardal R, Molofsky AV, He S, Morrison SJ. Stem cell self-renewal and cancer cell proliferationare regulated by common networks that balance the activation of proto-oncogenes and tumorsuppressors. Cold Spring Harb Symp Quant Biol. 2005; 70:177–185. [PubMed: 16869752]
19. Chen MH, Li YJ, Kawakami T, Xu SM, Chuang PT. Palmitoylation is required for the productionof a soluble multimeric Hedgehog protein complex and long-range signaling in vertebrates. GenesDev. 2004; 18:641–659. [PubMed: 15075292]
20. Chiang C, et al. Cyclopia and defective axial patterning in mice lacking Sonic hedgehog genefunction. Nature. 1996; 383:407–413. [PubMed: 8837770]
21. Harfe BD, et al. Evidence for an expansion-based temporal Shh gradient in specifying vertebratedigit identities. Cell. 2004; 118:517–528. [PubMed: 15315763]
22. Briscoe J, Ericson J. The specification of neuronal identity by graded Sonic Hedgehog signalling.Semin Cell Dev Biol. 1999; 10:353–362. [PubMed: 10441550]
23. Wechsler-Reya RJ, Scott MP. Control of neuronal precursor proliferation in the cerebellum bySonic Hedgehog. Neuron. 1999; 22:103–114. [PubMed: 10027293]
24. Charrier JB, Lapointe F, Le Douarin NM, Teillet MA. Anti-apoptotic role of Sonic hedgehogprotein at the early stages of nervous system organogenesis. Development. 2001; 128:4011–4020.[PubMed: 11641224]
25. Wallace VA. Purkinje-cell-derived Sonic hedgehog regulates granule neuron precursor cellproliferation in the developing mouse cerebellum. Curr Biol. 1999; 9:445–448. [PubMed:10226030]
26. Yabe T, Hata T, He J, Maeda N. Developmental and regional expression of heparan sulfatesulfotransferase genes in the mouse brain. Glycobiology. 2005; 15:982–993. [PubMed: 15944372]
27. Ahn S, Joyner AL. In vivo analysis of quiescent adult neural sterm cells responding to Sonichedgehog. Nature. 2005; 437:894–897. [PubMed: 16208373]
28. Jeong J, McMahon AP. Growth and pattern of the mammalian neural tube are governed bypartially overlapping feedback activities of the hedgehog antagonists patched 1 and Hhip1.Development. 2005; 132:143–154. [PubMed: 15576403]
29. Yamamoto Y, Stock DW, Jeffery WR. Hedgehog signalling controls eye degeneration in blindcavefish. Nature. 2004; 431:844–847. [PubMed: 15483612]
30. Hilton MJ, Tu X, Cook J, Hu H, Long F. Ihh controls cartilage development by antagonizing Gli3,but requires additional effectors to regulate osteoblast and vascular development. Development.2005; 132:4339–4351. [PubMed: 16141219]
31. Choi Y, Borghesani PR, Chan JA, Segal RA. Migration from a mitogenic niche promotes cell-cycle exit. J Neurosci. 2005; 25:10437–10445. [PubMed: 16280582]
32. Gritli-Linde A, Lewis P, McMahon AP, Linde A. The whereabouts of a morphogen: directevidence for short- and graded long-range activity of hedgehog signaling peptides. DevelopmentalBiology. 2001; 236:364–386. [PubMed: 11476578]
33. Hausmann B, Sievers J. Cerebellar external granule cells are attached to the basal lamina from theonset of migration up to the end of their proliferative activity. J Comp Neurol. 1985; 241:50–62.[PubMed: 4056112]
34. Kenney AM, Cole MD, Rowitch DH. Nmyc upregulation by sonic hedgehog signaling promotesproliferation in developing cerebellar granule neuron precursors. Development. 2003; 130:15–28.[PubMed: 12441288]
35. Kenney AM, Rowitch DH. Sonic hedgehog promotes G(1) cyclin expression and sustained cellcycle progression in mammalian neuronal precursors. Mol Cell Biol. 2000; 20:9055–9067.[PubMed: 11074003]
36. Ulloa F, Briscoe J. Morphogens and the control of cell proliferation and patterning in the spinalcord. Cell Cycle. 2007; 6:2640–2649. [PubMed: 17912034]
37. Sasaki H, Nishizaki Y, Hui C, Nakafuku M, Kondoh H. Regulation of Gli2 and Gli3 activities byan amino-terminal repression domain: implication of Gli2 and Gli3 as primary mediators of Shhsignaling. Development. 1999; 126:3915–3924. [PubMed: 10433919]
38. Aza-Blanc P, Lin HY, Ruiz i Altaba A, Kornberg TB. Expression of the vertebrate Gli proteins inDrosophila reveals a distribution of activator and repressor activities. Development. 2000;127:4293–4301. [PubMed: 10976059]
Chan et al. Page 11
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

39. Pan Y, Bai CB, Joyner AL, Wang B. Sonic hedgehog signaling regulates Gli2 transcriptionalactivity by suppressing its processing and degradation. Mol Cell Biol. 2006; 26:3365–3377.[PubMed: 16611981]
40. Blaess S, Corrales JD, Joyner AL. Sonic hedgehog regulates Gli activator and repressor functionswith spatial and temporal precision in the mid/hindbrain region. Development. 2006; 133:1799–1809. [PubMed: 16571630]
41. Corrales JD, Rocco GL, Blaess S, Guo Q, Joyner AL. Spatial pattern of sonic hedgehog signalingthrough Gli genes during cerebellum development. Development. 2004; 131:5581–5590.[PubMed: 15496441]
42. Yoon JW, et al. Gene expression profiling leads to identification of GLI1-binding elements intarget genes and a role for multiple downstream pathways in GLI1-induced cell transformation. JBiol Chem. 2002; 277:5548–5555. [PubMed: 11719506]
43. Duman-Scheel M, Weng L, Xin S, Du W. Hedgehog regulates cell growth and proliferation byinducing Cyclin D and Cyclin E. Nature. 2002; 417:299–304. [PubMed: 12015606]
44. Leung C, et al. Bmi1 is essential for cerebellar development and is overexpressed in humanmedulloblastomas. Nature. 2004; 428:337–341. [PubMed: 15029199]
45. Kenney AM, Widlund HR, Rowitch DH. Hedgehog and PI-3 kinase signaling converge on Nmyc1to promote cell cycle progression in cerebellar neuronal precursors. Development. 2004; 131:217–228. [PubMed: 14660435]
46. Dessaud E, et al. Interpretation of the sonic hedgehog morphogen gradient by a temporaladaptation mechanism. Nature. 2007; 450:717–720. [PubMed: 18046410]
47. Vyas N, et al. Nanoscale organization of hedgehog is essential for long-range signaling. Cell. 2008;133:1214–1227. [PubMed: 18585355]
48. Galvin KE, Ye H, Wetmore C. Differential gene induction by genetic and ligand-mediatedactivation of the Sonic hedgehog pathway in neural stem cells. Dev Biol. 2007; 308:331–342.[PubMed: 17599824]
49. Kleeff J, et al. The cell-surface heparan sulfate proteoglycan glypican-1 regulates growth factoraction in pancreatic carcinoma cells and is overexpressed in human pancreatic cancer. J ClinInvest. 1998; 102:1662–1673. [PubMed: 9802880]
50. Williamson D, et al. Role for amplification and expression of glypican-5 in rhabdomyosarcoma.Cancer Res. 2007; 67:57–65. [PubMed: 17210683]
Chan et al. Page 12
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. ShhAla specifically alters proteoglycan binding(a) ShhAla shows reduced binding to heparin-coated plates. Shh-AP (circles), ShhAla-AP(squares), or AP (triangles) were incubated with heparin-coated plates in the presence ofincreasing concentrations of soluble heparin.(b) Shh binds GPI-linked proteoglycans in cerebellar sections; ShhAla does not. P6 sectionstreated with vehicle control (−), Heparinases or PI-PLC, incubated with Shh-AP, ShhAla-APor vehicle controls, and processed for binding of AP-tagged ligand. Scale bar, 100 μm.(c) ShhAla is processed to mature isoform. Lysates of HEK293 transfected with Shh orShhAla were analyzed by immunoblot with anti-Shh. Immature 45kD (arrowhead) andmature 20kD isoform (arrow) are seen.
Chan et al. Page 13
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

(d) ShhAla is palmitoylated. HEK293 cells expressing Shh, ShhAla or ShhC24S were labeledwith 3H-palmitate, analyzed for palmitoylation, and probed with anti-Shh. Shh and ShhAla
are palmitoylated. ShhC24S is not.
Chan et al. Page 14
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2. ShhAla/Ala and ShhAla/− mice exhibit defects in growth, with normal patterning(a) Skeletal morphology, body and brain patterning are normal in ShhAla/Ala mice, but sizeof adult ShhAla/Ala mice is 11% less than wild type, and sizes of olfactory bulbs andcerebella are 30 and 31% reduced (arrows) respectively. Sagittal view of cerebellum showsnormal patterning with reduced size in ShhAla/Ala. Scale bar, 1 mm. ShhAla/Ala animals alsodisplay well-spaced eyes.(b) Spinal cord patterning is normal in ShhAla/Ala mice. In situ hybridization for Shh(expressed in notochord and floor plate), Nkx2.2 (motor neuron precursors), Nkx6.1 (ventralspinal cord), Isl1 (dorsal root ganglia and motor neurons), and Dbx1 (V0 interneurons) inE10.5 littermates. Scale bar, 100 μm.(c) Growth defects are seen in ShhAla/−, without defects in patterning. Olfactory bulb andcerebellum are reduced in size (red arrows). Scale bar, 1 mm. Eyes of ShhAla/− mice are wellspaced.
Chan et al. Page 15
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
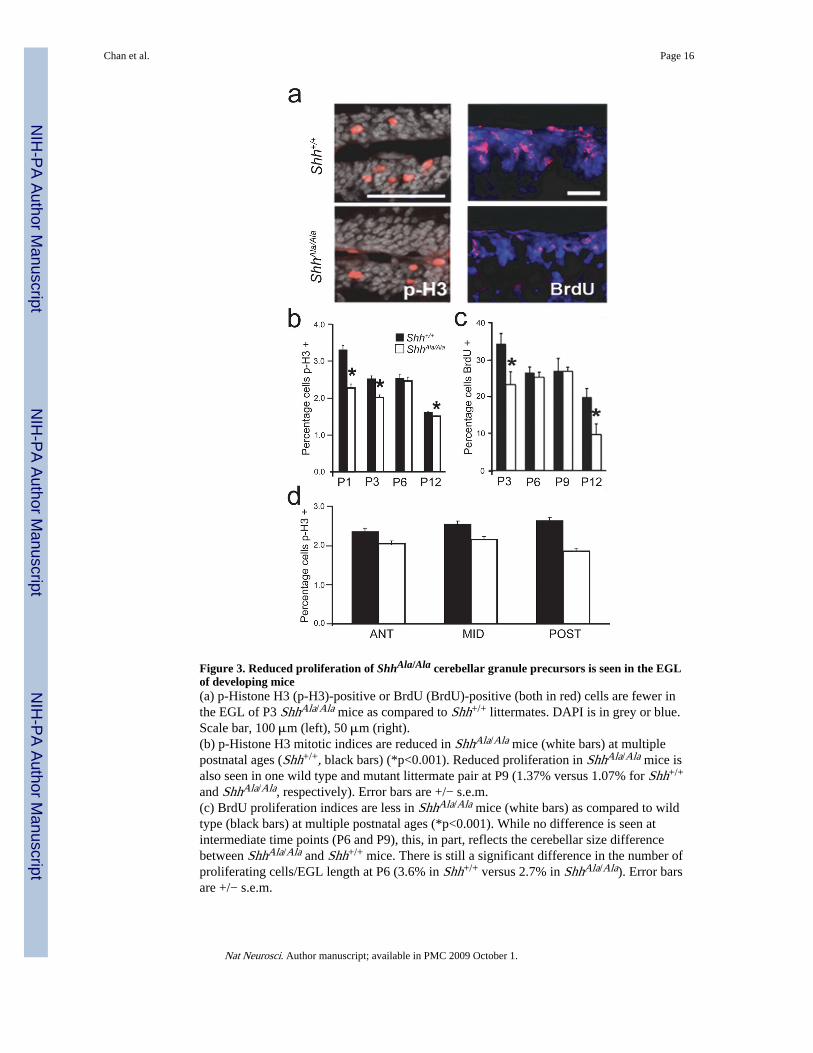
Figure 3. Reduced proliferation of ShhAla/Ala cerebellar granule precursors is seen in the EGLof developing mice(a) p-Histone H3 (p-H3)-positive or BrdU (BrdU)-positive (both in red) cells are fewer inthe EGL of P3 ShhAla/Ala mice as compared to Shh+/+ littermates. DAPI is in grey or blue.Scale bar, 100 μm (left), 50 μm (right).(b) p-Histone H3 mitotic indices are reduced in ShhAla/Ala mice (white bars) at multiplepostnatal ages (Shh+/+, black bars) (*p<0.001). Reduced proliferation in ShhAla/Ala mice isalso seen in one wild type and mutant littermate pair at P9 (1.37% versus 1.07% for Shh+/+
and ShhAla/Ala, respectively). Error bars are +/− s.e.m.(c) BrdU proliferation indices are less in ShhAla/Ala mice (white bars) as compared to wildtype (black bars) at multiple postnatal ages (*p<0.001). While no difference is seen atintermediate time points (P6 and P9), this, in part, reflects the cerebellar size differencebetween ShhAla/Ala and Shh+/+ mice. There is still a significant difference in the number ofproliferating cells/EGL length at P6 (3.6% in Shh+/+ versus 2.7% in ShhAla/Ala). Error barsare +/− s.e.m.
Chan et al. Page 16
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

(d) p-Histone H3 mitotic indices in P3 wild type (black bars) and ShhAla/Ala mice (whitebars) in the anterior, middle and posterior cerebellum are not statistically different from theirrespective totals in (b), demonstrating that lobes throughout the cerebellum are affected inthe mutant. Error bars are +/− s.e.m.
Chan et al. Page 17
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
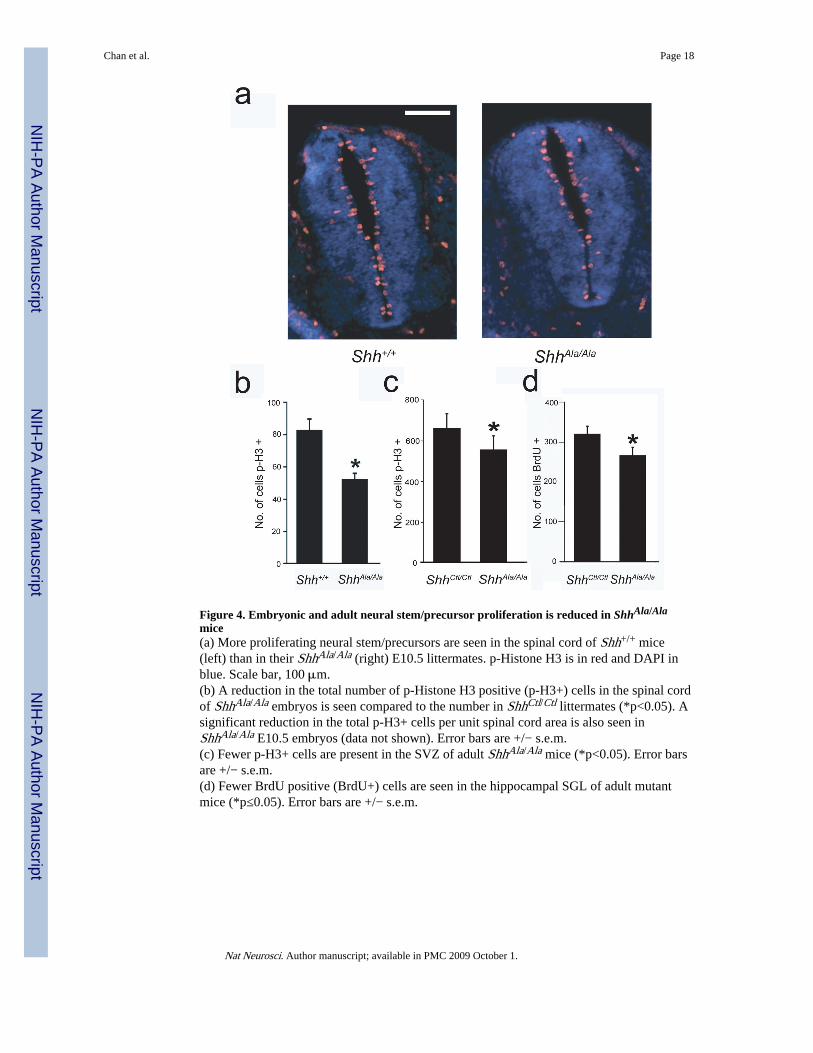
Figure 4. Embryonic and adult neural stem/precursor proliferation is reduced in ShhAla/Ala
mice(a) More proliferating neural stem/precursors are seen in the spinal cord of Shh+/+ mice(left) than in their ShhAla/Ala (right) E10.5 littermates. p-Histone H3 is in red and DAPI inblue. Scale bar, 100 μm.(b) A reduction in the total number of p-Histone H3 positive (p-H3+) cells in the spinal cordof ShhAla/Ala embryos is seen compared to the number in ShhCtl/Ctl littermates (*p<0.05). Asignificant reduction in the total p-H3+ cells per unit spinal cord area is also seen inShhAla/Ala E10.5 embryos (data not shown). Error bars are +/− s.e.m.(c) Fewer p-H3+ cells are present in the SVZ of adult ShhAla/Ala mice (*p<0.05). Error barsare +/− s.e.m.(d) Fewer BrdU positive (BrdU+) cells are seen in the hippocampal SGL of adult mutantmice (*p≤0.05). Error bars are +/− s.e.m.
Chan et al. Page 18
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5. ShhAla cannot specify a mitogenic niche(a) Granule cell precursors (GCPs) from GFP+ mice were cultured on P6 Shh+/+ orShhAla/Ala cerebellar slices, or DiO-labeled ShhAla/Ala GCPs were cultured on wild typeslices. Proliferation of introduced GCPs was analyzed by BrdU incorporation. Theproliferative index is the percent (%) of GFP- or DiO-positive cells in each layer that areBrdU+ (*p<0.05 versus EGL of WT slice; **p<0.05 vs IGL of WT slice; no significantdifference between EGL and IGL proliferation on ShhAla/Ala slices is seen). Error bars are +/− s.e.m.(b) EGL mitogenic niche in ShhAla/Ala slices (light gray bar) is phenocopied by addedexogenous heparan sulfates (HS) (dark gray bar) (*p<0.05). Error bars are +/− s.e.m.(c) Shh immunostaining of wild type and ShhAla/Ala P6 cerebella shows a reduction in Shhstaining in the EGLa of mutant mice (arrowhead) compared to wild type cerebella (arrows)(52%±11% of wild type (p=0.01)). EGLa: external granule cell layer, outer proliferativezone, EGLb: external granule cell layer, inner post-mitotic zone; ML: molecular layer; PCL:Purkinje cell layer; IGL: internal granule cell layer. Scale bar, 50 μm
Chan et al. Page 19
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 6. Shh-proteoglycan interactions promote proliferation in dissociated cell cultures ofGCPs, but are not needed for survival(a) The proliferative response to Shh (gray) (as assessed by BrdU incorporation) is greaterthan that of GCPs to ShhAla (black) (*p≤0.05, **p<0.01). Error bars are +/− s.e.m.(b) No consistent effects on survival (as assessed by activated caspase 3 immunostaining)are seen in GCPs stimulated with Shh (gray) or ShhAla (black). Error bars are +/− s.e.m.
Chan et al. Page 20
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
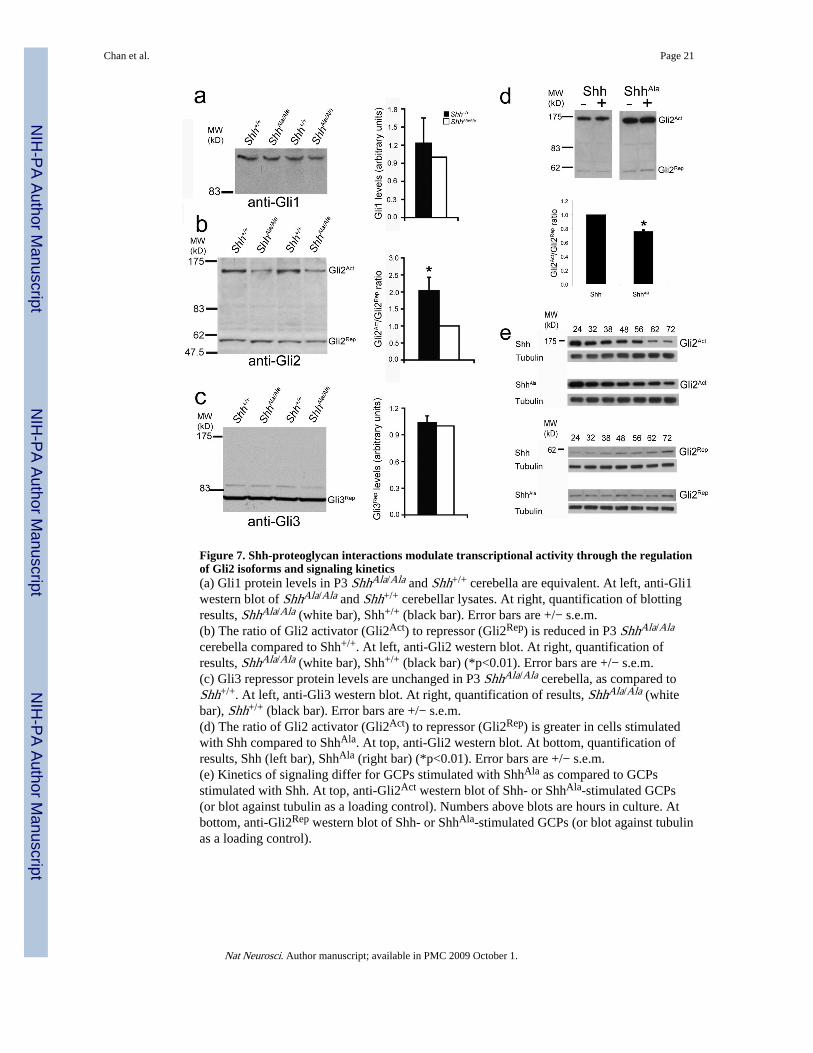
Figure 7. Shh-proteoglycan interactions modulate transcriptional activity through the regulationof Gli2 isoforms and signaling kinetics(a) Gli1 protein levels in P3 ShhAla/Ala and Shh+/+ cerebella are equivalent. At left, anti-Gli1western blot of ShhAla/Ala and Shh+/+ cerebellar lysates. At right, quantification of blottingresults, ShhAla/Ala (white bar), Shh+/+ (black bar). Error bars are +/− s.e.m.(b) The ratio of Gli2 activator (Gli2Act) to repressor (Gli2Rep) is reduced in P3 ShhAla/Ala
cerebella compared to Shh+/+. At left, anti-Gli2 western blot. At right, quantification ofresults, ShhAla/Ala (white bar), Shh+/+ (black bar) (*p<0.01). Error bars are +/− s.e.m.(c) Gli3 repressor protein levels are unchanged in P3 ShhAla/Ala cerebella, as compared toShh+/+. At left, anti-Gli3 western blot. At right, quantification of results, ShhAla/Ala (whitebar), Shh+/+ (black bar). Error bars are +/− s.e.m.(d) The ratio of Gli2 activator (Gli2Act) to repressor (Gli2Rep) is greater in cells stimulatedwith Shh compared to ShhAla. At top, anti-Gli2 western blot. At bottom, quantification ofresults, Shh (left bar), ShhAla (right bar) (*p<0.01). Error bars are +/− s.e.m.(e) Kinetics of signaling differ for GCPs stimulated with ShhAla as compared to GCPsstimulated with Shh. At top, anti-Gli2Act western blot of Shh- or ShhAla-stimulated GCPs(or blot against tubulin as a loading control). Numbers above blots are hours in culture. Atbottom, anti-Gli2Rep western blot of Shh- or ShhAla-stimulated GCPs (or blot against tubulinas a loading control).
Chan et al. Page 21
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 8. Proteoglycan interactions modulate Shh perdurance and differentially affect Shh-dependent gene expression(a) The gene pattern induced by ShhAla stimulation of wild type P6 GCPs is different thanthat of GCPs stimulated by equivalent amounts of Shh (*p<0.05). Error bars are +/− s.e.m.(b) GCP proteoglycans modulate Shh ligand perdurance. At left, (top) anti-Shh western blotof Shh- or ShhAla-stimulated C3H10T1/2s (or blot against actin as loading control). At right,(top) anti-Shh western blot of Shh- or ShhAla-stimulated GCPs (or blot against tubulin asloading control). Numbers above blots are hours in culture. At bottom, quantification ofresults from one representative experiment. Shh (solid), ShhAla (dashed).(c) Expression of Shh target genes in P1-2 ShhAla/Ala cerebella (P1-3 ShhAla/Ala cerebella forgli2) relative to Shh+/+ littermates demonstrates that Shh-proteoglycan interactionsdifferentially affect gene subsets (*p<0.01, **p<0.05). Error bars are +/− s.e.m.
Chan et al. Page 22
Nat Neurosci. Author manuscript; available in PMC 2009 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript




















