
Mitogenic and Cytotoxic Effects of Pentachlorophenol to AML 12 Mouse Hepatocytes
Upload
independentCategory
view
4download
0
Molecular and Cellular Endocrinology 150 (1999) 1–10
Actions of bFGF on mitogenic activity and lineage expression in ratosteoprogenitor cells: effect of age
Hiroshi Tanaka a,b, Hiroyoshi Ogasa a, Janice Barnes a, C. Tony Liang a,*a Gerontology Research Center, National Institute on Aging, National Institutes of Health, 5600 Nathan Shock Dri6e, Baltimore,
MD 21224-6825, USAb Department of Orthopedic Surgery, Yamaguchi Uni6ersity School of Medicine, 1144 Kogushi Ube, Yamaguchi, 755-8505, Japan
Received 14 August 1998; accepted 18 February 1999
Abstract
Rat osteoprogenitor cells were used to examine the effects of bFGF on DNA synthesis and the expression of osteoblast(OB)-related genes. bFGF, as low as 0.1 ng/ml, stimulated DNA synthesis. bFGF also increased the mRNA level of osteopontin(OP) and decreased that of type I collagen (COL I). When cultures were grown in dexamethasone (DEX) to induce OB lineagecommitment, the expression of COL I, alkaline phosphatase (AP) and OP was greatly enhanced. Subsequent incubation withbFGF partially negated the stimulatory effect of DEX on AP and COL I mRNAs. bFGF also inhibited the expression ofosteocalcin mRNA in cells grown in 1,25(OH)2D3 and DEX. Combined effects of bFGF with IGF-I or PDGF on DNA synthesisand OP expression were examined. bFGF+IGF-I, but not bFGF+PDGF, was more effective than PDGF alone. By comparingcells from adult and old animals, we found that bFGF-induced mitogenic activity was reduced significantly with age. In contrast,the effect of bFGF on the expression of OB genes was not significantly altered by age. These findings suggest that bFGF playsa dual role as a local positive and negative regulator on proliferation and osteogenic lineage expression, respectively, inosteoprogenitor cells, and that the mitogenic activity in response to bFGF was impaired in aging. © 1999 Published by ElsevierScience Ireland Ltd. All rights reserved.
Keywords: bFGF; IGF-I; PDGF; Bone marrow osteoprogenitor cells; Proliferation; Gene expression
1. Introduction
bFGF was first isolated from brain and pituitary(Gospodarowicz, 1990), however, it has now beendemonstrated in a variety of tissues and is known to bemultifunctional (Goustin et al., 1986). FGF family iscomprised of eight multifunctional proteins found innormal and/or malignant tissues (Sporn and Todara,1980; Goustin et al., 1986; Gospodarowicz, 1990). Theprototype members of the FGF family, aFGF andbFGF, have been suggested to play a role in wound
repair, inflammation, transformation, and angiogenesis(Shing et al., 1984; Gimenez-Gallego et al., 1985;Goustin et al., 1986; Gospodarowicz, 1990). The twoforms of FGF have similar biological effects and shareapproximately 55% sequence homology and interactwith the same cell surface receptors.
The mineralized matrix of skeletal tissue is known tocontain various growth factors including aFGF andbFGF (Hauschka et al., 1986). Recent studies showedthat cultured bovine bone cells synthesized bFGF andstored it in extracellular matrix (Globus et al., 1989). Inaddition, bFGF has been reported to enhance DNAand collagen synthesis in rat osteoblast cultures andreduce alkaline phosphatase (AP) activity (Canalis etal., 1988). Therefore, it is likely that bFGF may play aprominent role in the regulation of bone remodeling.
* Corresponding author. Tel.: +1-410-5588468; fax: +1-410-5588317.
E-mail address: [email protected] (C.T. Liang)
0303-7207/99/$ - see front matter © 1999 Published by Elsevier Science Ireland Ltd. All rights reserved.
PII: S 0 3 0 3 -7207 (99 )00046 -5

H. Tanaka et al. / Molecular and Cellular Endocrinology 150 (1999) 1–102
The marrow stromal system, which is associated withthe soft connective tissues of marrow and bone surface,contains stem cells capable of giving rise to several celltypes including fibroblasts, adipocytes and bone cells(Owen and Friedenstein, 1988). Current concepts pro-pose that these adherent stromal cells are enriched inosteoprogenitor cells that are involved in bone forma-tion. It has been shown that fibroblast-like cells derivedfrom bone marrow have the ability to produce a bone-like tissue in diffusion chambers (Ashton et al., 1980;Bab et al., 1986) and in collagen-hydroxyapatite im-plants (Goshima et al., 1991; Quarto et al., 1995) invivo. Recently it has been demonstrated that bonemarrow stromal cells, when cultured in the presence ofb-glycerophosphate, ascorbic acid, and dexamethasone(DEX), can induce the formation of mineralized bone-like nodules in vitro as characterized by the appearanceof collagen, osteocalcin (OC) and osteonectin and thehydroxyapatite nature of the deposited mineral (Mania-topoulos et al., 1988).
In aging, bone formation activity was significantlyreduced (Liang et al., 1992). It has been postulated thata deficiency in osteoprogenitor cells plays a major rolein the decline of bone induction activity in senesense(Quarto et al., 1995). Proliferation and osteogenic lin-eage expression of bone progenitor cells are regulatedby local and systemic growth factors. It is quite possiblethat changes in the availability of growth factors in oldbones or the response of osteoprogenitor cells from oldbones to growth factors may reduce the number ofprogenitor cells and osteoblasts in old bones and con-tribute to the decline in bone formation activity. In-deed, we have shown that reduced expression of IGF-I(Tanaka et al., 1996), impaired response of osteopro-genitor cells from old animals to IGF-I and PDGF(Slootweg et al., 1990; Tanaka et al., 1994; Tanaka andLiang, 1995) are the possible causes for impaired boneformation activity in old animals. In the present study,we extended our study to examine the response ofosteoprogenitor cells to bFGF, an important regulatorygrowth factor for bone remodeling. Results obtainedfrom this study can strengthen the general concept thatthe impaired response of osteoprogenitor cells togrowth factors may contribute to the insufficient boneformation activity commonly observed in elderlypeople.
2. Materials and methods
2.1. Primary cultures of rat bone marrowosteoprogenitor cells
A suspension of marrow stromal cell was prepared asdescribed previously (Tanaka et al., 1994). Marrowcells were flushed from the midshafts of femurs of 6-
(adult) and 24-month (old) male Wistar rats (the Ani-mal Facility, Gerontology Research Center, NationalInstitute on Aging), with Coon’s modified Ham’s F-12medium (NIH Media Unit, Bethesda, MD) supple-mented with 10% fetal calf serum (Biofluids, Rockville,MD). Single cell suspension was obtained by passagethrough 18- and 23- gauge needles. Cell suspension wasdiluted with saline for cell counting. After a briefincubation in 1% Zapoglobin (Curtin Matheson Scien-tific, Houston, TX) to lyse red blood cells, the numberof nucleated cells were counted with a Coulter Counter(Coulter Corporate Communications, Hialeah, FL).Cells were plated out into 60-mm plates (Falcon BectonDickinson, Oxnard, CA) at a density of 1×106 nucle-ated cells/cm2 and cultured in Coon’s modified Ham’sF-12 medium with 10% fetal calf serum, 50 mg/mlL-ascorbic acid, 100 i.u./ml penicillin and 100 mg/mlstreptomycin (NIH Media Unit, Bethesda, MD) at37°C in 5% CO2 in a humidified incubator. Media werechanged after 4–5 days and the non-adherent cells werediscarded. Thereafter, media were changed twice weeklyuntil the cultures were confluent. Confluent primarystromal cells were detached with 0.05% (w/v) trypsin-EDTA (NIH Media Unit, Bethesda, MD) and pooledfrom three rats for subsequent experiments examining[3H]thymidine incorporation or gene expression. Theexperimental protocol used in this study was reviewedand approved by the Animal User and Care Committeeat the Gerontology Research Center, National Instituteon Aging.
2.2. [3H]thymidine incorporation in osteoprogenitorcells
Pooled primary progenitor cells were plated at 50 000cells/well in 24-well dishes in the same media describedabove for primary culture. After 3 h of incubation, themedia were replaced with fresh media without serum.After serum depletion for 20 h, cells were treated withgrowth factors; human recombinant bFGF (BACHEM,INC., Torrance, CA), human recombinant IGF-I(BACHEM, INC., Torrance, CA), human recombinantPDGF (BACHEM, INC., Torrance, CA), for 24 h inserum-free media and pulse labeled with [3H]thymidineduring the last 3 h. Cells were then fixed in 10% coldTCA, lysed in 0.1 N sodium hydroxide, and assayed foracid insoluble [3H]thymidine incorporation.
2.3. Preparation of RNA
Pooled primary cells were plated at a density of4.5×103 cells/cm2 in the same medium as primaryculture. DEX and 1,25(OH)2D3 were added to selecteddishes at a final concentration of 10−8 M. The mediawere changed twice weekly. When cells were confluent

H. Tanaka et al. / Molecular and Cellular Endocrinology 150 (1999) 1–10 3
(approximately 8–10 days), growth factors were addedand incubated for 2 days. Total RNA from progenitorcells was prepared with RNA STAT-60 (Tel-Test,Friendswood, TX) following the instructions providedby the supplier. The purity of RNA was monitored bythe ratio of absorbencies of the samples at 260 and 280nm.
2.4. Northern analysis
Northern blot analysis was carried out as describedpreviously (Liang et al., 1992). Briefly, total RNA (20mg) was denatured, separated on a 1% agarose gel(FMC Co., Rockland, ME) and transferred to GeneScreen paper (New England Nuclear, Boston, MA).The cDNA probes used were rat AP (0.6 kb) (Noda etal., 1987), rat procollagen a1 (I) (1.3 kb) (Genovese etal., 1984), rat OC (0.5 kb) (Yoon et al., 1988) and ratosteopontin (OP) (1.3 kb) (Yoon et al., 1987). 32P-la-beled probes were prepared with a multiprime labelingkit (Amersham, Arlington Height, IL). Blots were hy-bridized at 42°C for 24 h in 50% formamide. Afterhybridization, blots were washed with 2×SSC (0.15 MNaCl and 0.015 M Sodium citrate) and 0.1% SDS twiceat room temperature, 1×SSC and 0.1% SDS at 50°C,and 0.5×SSC and 0.1% SDS at 60°C. For AP and OC,the last wash was carried at 57°C. Autoradiogram wasexposed at −70°C with an intensifying screen. A singleband of 0.6, 1.4 and 2.5 kb was labeled with probes forOC, OP and AP, respectively. Multiple transcripts of5.1 and 5.7 kb were labeled with procollagen a1 (I)probe.
2.5. Dot blot analysis
Total RNA was applied onto Gene Screen membranewith a Minifold apparatus (Schleicher & Schuell,Waburn, MA). The membrane was baked at 80°C for 2h to immobilize RNA and hybridized with variouscDNA probes, as described above. A polythymidylateprobe was used for determination of poly(A)+RNA(Tanaka et al., 1996). Hybridization and washing of thedot blots followed the procedures described above.Radioactivity of each dot on the filter was measuredwith a Betascope 603 Blot analyzer (Betagen Corp.,Waltham, MA). The levels of mRNAs were expressedas the ratio of each mRNA to total poly(A)+RNA.This normalized mRNA level was designated to be 1.0for control cells. In experiments using DEX-treatedcells, the normalized mRNA level for DEX-treated cellswas designated to be 1.0.
2.6. Statistics
Data shown are the mean9S.E. An ANOVA testwas performed to determine the difference between
groups. Statistical difference between two groups wasdetermined by paired and unpaired t-test.
3. Results
3.1. Effect of bFGF on [ 3H]thymidine incorporationinto DNA
The effect of bFGF on DNA synthesis in osteopro-genitor cells was assessed by measuring the incorpora-tion of [3H]thymidine into DNA. bFGF stimulatedDNA synthesis in progenitor cells in a dose-dependentmanner between 0.1–100 ng/ml (Fig. 1). Stimulation of0.9–fold was obtained at 0.1 ng/ml of bFGF. A 5.5-fold increase was achieved with bFGF at 100 ng/ml.
3.2. Effect of bFGF on the expression ofosteoblast-related markers
Confluent progenitor cells were treated with differentdoses of bFGF for 2 days. The levels of OP, type Icollagen (COL I) and AP mRNAs were determined bydot blot analysis and normalized to totalpoly(A)+RNA. Treatment of progenitor cells withbFGF stimulated mRNA expression of OP in a dose-dependent manner (Fig. 2A). Increase of 1.1–4.0-foldwas obtained with 10–250 ng/ml of bFGF. In contrast,bFGF at doses higher than 50 ng/ml decreased COL ImRNA significantly. The AP mRNA was not affected.The stimulatory effect of bFGF on OP mRNA wasconfirmed by Northern blot. A representative Northernblot (Fig. 2B) showed a clear increase in OP mRNA.
Fig. 1. Effect of bFGF on [3H]thymidine incorporation into DNA inrat bone marrow progenitor cells. Primary progenitor cells from threerats were pooled and plated at 5×104 cells/well in 24-well dishes.Serum-starved cells were incubated with growth factor at the concen-tration shown. Details of the experimental protocol are described inSection 2. Values shown are means9S.E. for a representative exper-iment with triplicate determinations. The experiment was repeatedtwice with similar results. P value for ANOVA analysis is shown inthe figure. For comparison between two groups: *P90.05 vs control,paired t-test.
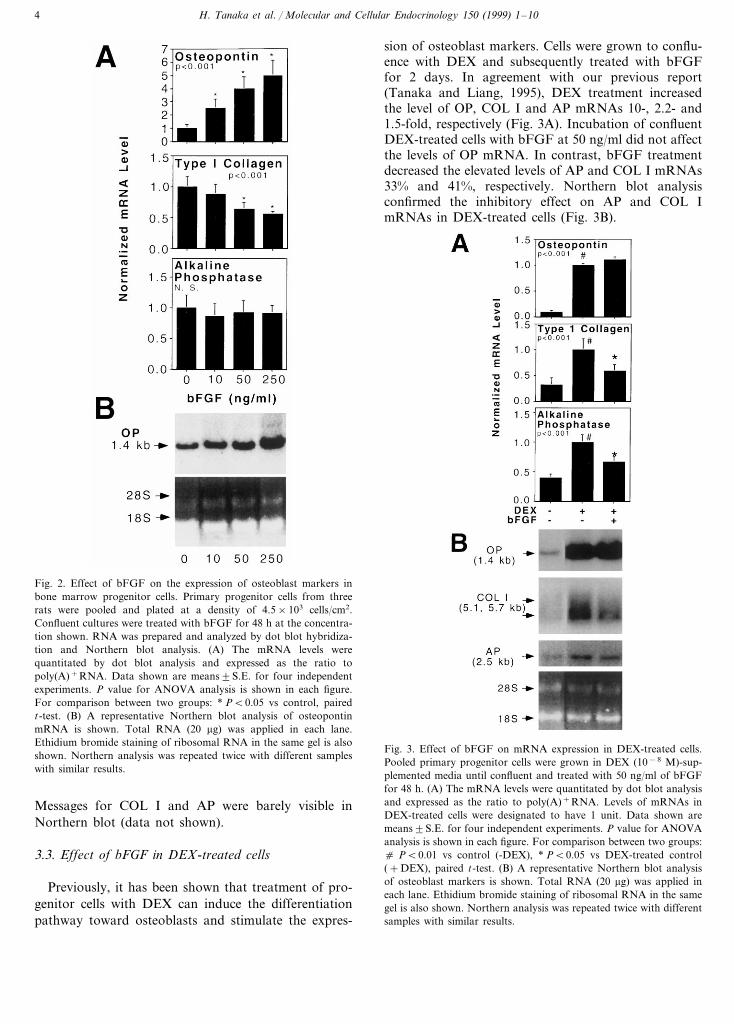
H. Tanaka et al. / Molecular and Cellular Endocrinology 150 (1999) 1–104
Fig. 2. Effect of bFGF on the expression of osteoblast markers inbone marrow progenitor cells. Primary progenitor cells from threerats were pooled and plated at a density of 4.5×103 cells/cm2.Confluent cultures were treated with bFGF for 48 h at the concentra-tion shown. RNA was prepared and analyzed by dot blot hybridiza-tion and Northern blot analysis. (A) The mRNA levels werequantitated by dot blot analysis and expressed as the ratio topoly(A)+RNA. Data shown are means9S.E. for four independentexperiments. P value for ANOVA analysis is shown in each figure.For comparison between two groups: * PB0.05 vs control, pairedt-test. (B) A representative Northern blot analysis of osteopontinmRNA is shown. Total RNA (20 mg) was applied in each lane.Ethidium bromide staining of ribosomal RNA in the same gel is alsoshown. Northern analysis was repeated twice with different sampleswith similar results.
sion of osteoblast markers. Cells were grown to conflu-ence with DEX and subsequently treated with bFGFfor 2 days. In agreement with our previous report(Tanaka and Liang, 1995), DEX treatment increasedthe level of OP, COL I and AP mRNAs 10-, 2.2- and1.5-fold, respectively (Fig. 3A). Incubation of confluentDEX-treated cells with bFGF at 50 ng/ml did not affectthe levels of OP mRNA. In contrast, bFGF treatmentdecreased the elevated levels of AP and COL I mRNAs33% and 41%, respectively. Northern blot analysisconfirmed the inhibitory effect on AP and COL ImRNAs in DEX-treated cells (Fig. 3B).
Fig. 3. Effect of bFGF on mRNA expression in DEX-treated cells.Pooled primary progenitor cells were grown in DEX (10−8 M)-sup-plemented media until confluent and treated with 50 ng/ml of bFGFfor 48 h. (A) The mRNA levels were quantitated by dot blot analysisand expressed as the ratio to poly(A)+RNA. Levels of mRNAs inDEX-treated cells were designated to have 1 unit. Data shown aremeans9S.E. for four independent experiments. P value for ANOVAanalysis is shown in each figure. For comparison between two groups:c PB0.01 vs control (-DEX), * PB0.05 vs DEX-treated control(+DEX), paired t-test. (B) A representative Northern blot analysisof osteoblast markers is shown. Total RNA (20 mg) was applied ineach lane. Ethidium bromide staining of ribosomal RNA in the samegel is also shown. Northern analysis was repeated twice with differentsamples with similar results.
Messages for COL I and AP were barely visible inNorthern blot (data not shown).
3.3. Effect of bFGF in DEX-treated cells
Previously, it has been shown that treatment of pro-genitor cells with DEX can induce the differentiationpathway toward osteoblasts and stimulate the expres-

H. Tanaka et al. / Molecular and Cellular Endocrinology 150 (1999) 1–10 5
Fig. 4. Effect of bFGF on the expression of OC mRNA. Pooledprimary progenitor cells were grown in media supplemented withDEX (10−8 M) and 1,25-(OH)2D3 (10-8 M). Confluent cultures weretreated with 50 ng/ml of bFGF for 48 h. Expression of OC wasanalyzed by Northern blot. Total RNA (20 mg) was applied in eachlane. Ethidium bromide staining of ribosomal RNA in the same gel isalso shown. The experiment was repeated twice with similar results.
adult and old animals. As shown in Fig. 6, the stimula-tion of DNA synthesis was observed at 0.1 ng/ml of
Fig. 5. Effect of bFGF in combination with IGF-I or PDGF onDNA synthesis and OP expression in stromal cells. (A) Serum-starvedbone marrow progenitor cells were incubated with growth factors at100 ng/ml. Details of the experimental protocol are described inSection 2. Values shown are means9S.E. for a representative exper-iment with triplicate determinations. The experiment was repeatedtwice with similar results. P value for ANOVA analysis is shown inthe figure. For comparison between two groups: * PB0.05 vs con-trol, c PB0.05 vs +bFGF or +IGF-I, paired t-test. (B) Conflu-ent progenitor cells were treated with 50 ng/ml of growth factors for48 h. The mRNA levels for OP were quantitated by dot blot analysisand expressed as the ratio to poly(A)+RNA. Data shown aremeans9S.E. for four independent experiments from different ani-mals. P value for ANOVA analysis is shown in the figure. Forcomparison between two groups: * PB0.05 vs control, c PB0.05vs +bFGF or +IGF-I, paired t-test. A representative Northern blotof OP mRNA is shown. Total RNA (20 mg) was applied in each lane.Ethidium bromide staining of ribosomal RNA in the same gel is alsoshown.
The effect of bFGF on the mRNA level OC was alsoexamined. The expression of OC was minimal in con-trol cells and can be induced several fold in cellsmaintained with DEX and 1,25(OH)2D3. As shown inFig. 4, treatment of confluent cultures, maintained inDEX and vitamin D, with bFGF at 50 ng/ml reducedOC mRNA level more than 50%.
3.4. Interactions of bFGF with IGF-I or PDGF onDNA synthesis and the expression of OP
To determine whether bFGF interacts with the re-sponses of osteoprogenitor cells to other local growthfactors, DNA synthesis and the expression of OP wereexamined in cells treated with bFGF and PDGF orbFGF and IGF-I. As shown in Fig. 5A, bFGF, PDGFand IGF-I increased [3H]thymidine incorporation 4.6-,7.2- and 1.2-fold, respectively, over the control. How-ever, the combination of PDGF and bFGF yielded nofurther stimulation above that observed with PDGFalone. In contrast, DNA synthesis was stimulated 8-fold in cells treated with bFGF and IGF-I which wasgreater than the sum of the individual responses (4.6-and 1.2-fold, respectively, for bFGF and IGF-I).
Parallel experiments were carried out to determinewhether bFGF can interact with IGF-I or PDGF onthe expression of OP mRNA. bFGF, PDGF and IGF-Iat 50 ng/ml stimulated OP mRNA expression 2.6-, 2.0-and 0.2-fold over the control, respectively (Fig. 5B).The combined treatment of PDGF and bFGF resultedin no stimulation above that observed with PDGF orbFGF alone. Treatment of cells with IGF-I and bFGFstimulated OP expression 4.5-fold which was higherthan the sum of individual responses (2.6- and 0.1-fold,respectively, for bFGF and IGF-I).
3.5. Effect of age on DNA synthesis in osteoprogenitorcells
DNA synthesis was compared in cells derived from

H. Tanaka et al. / Molecular and Cellular Endocrinology 150 (1999) 1–106
Fig. 6. Effect of age on bFGF-stimulated DNA synthesis in osteopro-genitor cells. DNA [3H]thymidine incorporation was determined asdescribed in the legend to Fig. 1. Data shown are means9S.E. forquadruple measurements. The experiment was repeated three timeswith similar results. *PB0.05 vs cells from adult animals, unpairedt-test.
and osteoblasts in old animals and lead to impairedbone formation activity. The present study was under-taken to gain better insight into the role of bFGF onreplication and the progressive phenotype expression ofrat bone marrow stromal cells which are believed tocontain osteoprogenitor cells. We then examined thepossible defects in these responses in aging.
Initially, we examined the effect of bFGF on DNAsynthesis and the expression of osteoblast markers inprogenitor cells derived from adult rats (6-month old).In cultures of marrow progenitor cells, treatment withbFGF as low as 0.1 ng/ml for 24 h stimulated incorpo-ration of [3H]thymidine into DNA. Similar results havebeen reported in previous studies in cultured fetal ratcalvariae cells (Gospodarowicz et al., 1987; Canalis etal., 1988; McCarthy et al., 1989) and bone marrow
Fig. 7. Effect of age on bFGF-induced changes in gene expression inosteoprogenitor cells. The expression of osteoblast-related genes wasdetermined as described in the legend to Fig. 2. Equal amount ofRNA samples obtained from two age groups were used in the samedot blot analysis. The basal level of mRNA expressed as ratio topoly(A)+RNA was not significantly different between adult and oldanimals. Data shown are means9S. E. for four independent experi-ments. *PB0.01 vs cells treated with 50 ng/ml of bFGF, pairedt-test.
bFGF in cells from both age groups. However, themagnitude of stimulation by bFGF in cells derivedfrom old animals was approximately half of that ob-tained for cells from adult animals.
3.6. Effect of age on the expression ofosteoblast-related genes
The expression of osteoblast-related genes was exam-ined in cells derived from adult and old animals. ThemRNA levels of OP, COL I and AP, measured as ratioto poly(A)+RNA, were not different for cells fromadult and old bones (legend to Fig. 7). bFGF stimu-lated the expression of OP, inhibited that of COL I anddid not significantly activate the expression of AP incells from both age groups (Fig. 7). Age did not affectthese responses significantly. One exception was thatthe stimulation of OP expression was higher in cellsfrom old animals.
4. Discussion
Recently, it has been summarized in a review thatgrowth factors and cytokines play an important role inbone cell metabolism (Canalis et al., 1991). Consider-able interest has also been focused on the effect of theseregulators on proliferation and phenotype expression ofosteoprogenitor cells along the osteogenic pathway. Inaddition, question arises whether the deficiency in theresponse of osteoprogenitor cells to growth factors maycontribute to the insufficient number of progenitor cells

H. Tanaka et al. / Molecular and Cellular Endocrinology 150 (1999) 1–10 7
stromal cells from very young rats (Noff et al., 1989;Pitaru et al., 1993). Based on its effect on the growth ofostoblasts, bFGF is considered as a positive regulatorof bone formation in adult animals.
The effect of bFGF on the expression of osteoblast-related genes was also examined in adult progenitorcells. bFGF at doses higher than 50 ng/ml significantlyreduced the level of COL I mRNA in adult progenitorcells. In contrast, AP mRNA was not affected bybFGF. Osteoprogenitor cells can be induced to commitand differentiate along the osteoblast lineage by treat-ment with DEX (Leboy et al., 1991). Both COL I andAP are considered as markers for preosteoblasts andtheir expression are detected in the early phase of activeproliferation of osteoblasts (Owen et al., 1990; Liang etal., 1992). In the present study, we showed that incuba-tion of confluent DEX-treated progenitor cells with 50ng/ml of bFGF for 2 days reduced the elevated levels ofCOL I and AP mRNAs. The inhibitory effect of bFGFon COL I expression may not be exclusively associatedwith phenotype development of osteoprogenitor cells. Ithas been shown that bFGF inhibits collagen synthesisby a transcriptional mechanism and the COL I genepromoter contains DNA sequences which mediate theinhibitory effect by bFGF (Hurley et al., 1991).
The effect of bFGF on the expression of COL I andAP has been examined previously using cells fromdifferent sources. In rat osteosarcoma cells, bFGF in-hibits the expression of AP and COL I (Rodan et al.,1989). However, bFGF decreases AP activity, but in-creases the level of COL I mRNA in cultures enrichedin parietal derived preosteoblasts and osteoblasts (Mc-Carthy et al., 1989). In bone marrow stromal cellcultures treated with DEX, ascorbic acid and b-glyc-erophosphate, bFGF increases both AP activity and[3H]proline incorporation into collagen (Pitaru et al.,1993). It is not clear whether the discrepancy is theresult of differences in culture condition, cell type, thestage of cells along the osteogenic pathway, the age ofthe animals that these cells were derived from or otherunknown parameters.
Our results clearly showed that bFGF increased OPmRNA in a dose-responsive manner. The stimulatoryeffect can be obtained as low as 10 ng/ml of bFGF. InDEX-treated cells, the expression of OP mRNA wasstimulated 10-fold. The elevated level of OP mRNA inDEX-treated cells was not enhanced further by bFGFtreatment. The expression of OC was not detectable incultured progenitor cells. Combined treatment withDEX and vitamin D enhanced the expression of OCmRNA more than 10-fold. Subsequent incubation with50 ng/ml of bFGF significantly reduced the level of OCmRNA. The significance of the finding that bFGFregulates the expression of OP and OC is not clear.Although OP and OC are postulated to be involved inbone formation, the precise role of these proteins in
bone remodeling have not been defined. In culturedfetal calvarial cells, the expression of OP and OC isstimulated at the onset of mineralization but not duringactive cell proliferation and extracellular matrix matu-ration (Owen et al., 1990). In rat femur, the expressionof OP and OC is elevated during both bone formationand resorption following marrow injury (Liang et al.,1992). Although OP was first isolated from mineralizedbone matrix (Franzen and Heinegard, 1984), it is alsoproduced in many other tissues including skin, kidney,uterus, neural cells and others (Nomura et al., 1988;Young et al., 1990; Tanaka et al., 1996), suggesting thatit may play a wider role than previously realized. OPenhances cell attachment (Oldberg et al., 1986), and hasbeen postulated to act as the anchorage of osteoclaststo bone matrix (Reinholt et al., 1990). A recent studyhas shown that potent bone-resorbing agents such asIL-1, TNF-a, LPS and 1,25(OH)2D3 increase the ex-pression of OP mRNA in both the clonal osteoblasticcells and the primary osteoblast-like cells, suggestingthat OP is a possible candidate involved in the interac-tion between osteoblasts and osteoclasts (Jin et al.,1990). bFGF has been shown to stimulate the resorp-tion of cultured fetal rat long bone (Simmons andRaisz, 1991). It is conceivable that bFGF may stimulatebone resorption via its effect on OP expression.
In bone, osteoprogenitor cells and osteoblasts areexposed to multiple local growth factors constantly andthe effects of one of these multifunctional factors arelikely modulated by the presence of others. In additionto bFGF, IGF-I and PDGF are known to regulate theproliferation and the expression of osteoblast-relatedgenes in stromal cells (Tanaka et al., 1996; Tanaka andLiang, 1996). Here we demonstrate that there is anaugmentative stimulatory effect on DNA synthesis andOP gene expression by IGF-I and bFGF. Interactionbetween IGF-I and bFGF on mitogenic activity hasbeen reported in osteoblasts (Slootweg et al., 1990),chondrocytes (Hiraki et al., 1987), heart mesenchymalcells (Balk et al., 1984) and adipocytes (Butterwith etal., 1992). This type of interaction between growthfactors is not limited to bFGF and IGF-I. PDGF andIGF-I are known to augment each other on cell prolif-eration and OP expression in marrow progenitor cells(Tanaka and Liang, 1995). Synergistic interaction canbe caused by activation of different signal transductionmechanisms or altering the number or the affinity ofthe receptor toward one growth factor by the othergrowth factor. A previous study has shown that bFGFincreases the number of IGF-I receptors in muscle cells(Rosenthal et al., 1991). Failure of bFGF and PDGF toelicit an additional increase in activity over that ob-tained with PDGF alone suggests that these two growthfactors may share the same pathway. However, syner-gistic effect of the same combination has been demon-strated in adipocytes (Butterwith et al., 1992).

H. Tanaka et al. / Molecular and Cellular Endocrinology 150 (1999) 1–108
The effect of bFGF on bone formation has beenexamined in several studies. bFGF stimulate the in-growth rate of new bone in bone induction chambersimplanted in rat tibiae (Wang and Aspenberg, 1996).However, bFGF at high dose is not effective in stimu-lating the ingrowth rate. Systemic administration ofbFGF on bone formation has been examined in grow-ing rats (Nagai et al., 1995). At low dose, bFGFstimulates endosteal and endochondral bone formationand depresses periosteal bone formation. At high doseof bFGF, calcification at the growth plate metaphysealjunction and the endocortical surface are retarded. Thelongitudinal growth rate, endocortical labeled surfaceand bone formation rate are also decreased with highdose of bFGF. The causes of the dose dependentvariation are not discussed in these studies. In trans-genic mice with overexpression of bFGF, skeletal mal-formations including shortening and flattening of longbones were observed (Coffin et al., 1995). In the presentstudy using cultured progenitor cells, we showed thatbFGF can stimulate the proliferation of these osteopro-genitor cells in the range of 0.1–10 ng/ml of bFGF.However, the expression of osteogenic markers such asCOL I and AP was inhibited at higher dose of bFGF(\50 ng/ml). Perhaps the dose dependent effects ofbFGF on bone metabolism observed in in vivo are theresult of the differential effects of bFGF on cell prolif-eration and osteogenic function as shown in our studiesusing cultured cells.
In the current study we also compared the bFGFinduced changes in the mitogenic activity and osteo-genic lineage expression in osteoprogenitor cells derivedfrom marrow of adult and old bones. The effective doseof bFGF to stimulate cell proliferation was not differ-ent between cells from two age groups. However, themaximal mitogenic activity was reduced about half forcells from old animals. In contrast, the expression ofosteoblast-related genes in response to bFGF was, byand large, not altered with age. Similar findings ofage-associated differences in the response of bone pro-genitor cells to growth factors have also been observedin studies with PDGF (Tanaka and Liang, 1995). Eventhough the mitogenic activity of progenitor cells fromold animals in response to bFGF was reduced, positiveincrease in the proliferation of progenitor cells bybFGF was still possible in these animals. Therefore, itcan not exclude the possible application of bFGF intreatment of age-associated osteoporosis. One usefulstrategy would be using the combination of IGF-I anda loss of bFGF which should maximize the mitogenicactivity of progenitor cells. Previously, we showed thatIGF-I infusion can increase bone formation activity inold rats and substantially negate the decrease in trabec-ular bone (Wakisaka et al., 1998). It would be interest-ing to test the combined effect of bFGF and IGF-I inthe same model.
Previously, we and other laboratories have shownthat the number of osteoprogenitor cells reduces withage. This age-related abnormality may be due in part tothe deficient mitogenic response of progenitor cellsfrom old animals to growth factors as described above.It should be noted that despite the consistent resultsfrom numerous animal studies which show that thenumber of osteoprogenitor cells reduces with age, simi-lar findings have not been reported in limited studiesusing human cells. It is unclear whether this is due todifferences in species, experimental protocol or otherunknown factors.
In conclusion, we have demonstrated that bFGFmodulates proliferation and the expression of os-teoblast-related genes in adult bone marrow osteopro-genitor cells. Our findings suggest that bFGF may playa dual role in the complex regulation of bone remodel-ing by local growth factors. We have also shown thatthe interaction between growth factors may augmentthe effect of individual growth factor on bone cellmetabolism. Perhaps, a better strategy can be devel-oped for possible use of growth factors in the treatmentof impaired bone activity associated with osteoporosis.
References
Ashton, B.A., Allen, T.D., Howlett, C.R., Eaglesom, C.C., Hattori,A., Owen, M.E., 1980. Formation of bone and cartilage bymarrow stromal cells in diffusion chambers in vivo. Clin. Orthop.151, 294–307.
Bab, I., Ashton, B.A., Gazit, D., Marx, G., Willamson, M.C., Owen,M.E., 1986. Kinetics and differentiation of stromal cells in diffu-sion chambers in vivo. J. Cell Sci. 84, 139–151.
Balk, S.D., Morisi, A., Gunther, H.S., Svoboda, M.F., Van Wyk,J.J., Nissley, S.P., Scanes, C.G., 1984. Somatomedins (insulin-likegrowth factors), but not growth hormones, are mitogenic forchicken heart mesenchymal cells and act synergistically with epi-dermal growth factor and brain fibroblast growth factor. LifeSciences 35, 335–346.
Butterwith, S.C., Peddie, C.D., Goddard, C., 1992. Regulation ofadipocyte precursor DNA synthesis by acidic and basic fibroblastgrowth factors: interaction with heparin and other growth factors.Endocrinology 137, 369–374.
Canalis, E., Centrella, M., McCarthy, T., 1988. Effect of basicfibroblast growth factor on bone formation in vitro. J. Clin.Invest. 81, 1572–1577.
Canalis, E., McCarthy, T., Centrella, M., 1991. Growth factors andcytokines in bone cell metabolism. Annu. Rev. Med. 42, 17–24.
Coffin, J.D., Florkiewicz, R.Z., Neumann, J., Mort-Hopkins, T.,Dorn, G.W. II, Lightfoot, P., German, R., Howles, P.N., Kier,A., O’Toole, B.A., Sasse, J., Gonzalez, A.M., Baird, A.,Doetschman, T., 1995. Abnormal bone growth and selectivetranslational regulation in basic fibroblast growth factor (FGF-2)transgenic mice. Mol. Biol. Cell 6, 1861–1873.
Franzen, A., Heinegard, D., 1984. Characterization of a proteoglycanfrom the calcified matrix of bovine bone. Biochem. J. 224, 59–66.
Genovese, C., Rowe, D., Kream, B., 1984. Construction of DNAsequences complementary to rat a1 and a2 collagen mRNA andtheir use in studying the regulation of type I collagen synthesis by1,25-dihydroxyvitamin D. Biochemistry 23, 6210–6216.

H. Tanaka et al. / Molecular and Cellular Endocrinology 150 (1999) 1–10 9
Gimenez-Gallego, G., Rodkey, J., Bennett, C., Rios-Candelore, M.,DiSalvo, J., Thomas, K., 1985. Brain derived acidic fibroblastgrowth factor: complete amino acid sequence and homologies.Science 230, 1385–1388.
Globus, R., Plouet, J., Gospodarowicz, D., 1989. Cultured bovinebone cells synthesize fibroblast growth factor and store it in theirextracellular matrix. Endocrinology 124, 1539–1547.
Goshima, J., Goldberg, V.M., Caplan, A.I., 1991. The osteogenicpotential of culture-expanded rat marrow stromal cells assayed invivo in calcium phosphate ceramic blocks. Clin. Orthop. Rel. Res.262, 298–311.
Gospodarowicz, D., 1990. Fibroblast growth factor. Chemical struc-ture and biologic function. Clin. Orthop. 257, 231–248.
Gospodarowicz, D., Ferrera, N., Schweigerer, L., Newfeld, G., 1987.Structural characterization and biological functions of fibroblastgrowth factor. Endocr. Rev. 8, 95–114.
Goustin, A.S., Leof, E.B., Shipley, G.D., Moses, H.L., 1986. Growthfactors and cancer. Cancer Res. 46, 1015–1029.
Hauschka, P.V., Maurakos, A.E., Jafrati, M.D., Doleman, S.E.,Klagsburn, M., 1986. Growth factors in bone matrix: isolation ofmultiple types by affinity chromatography on heparin sepharose.J. Biol. Chem. 261, 12665–12674.
Hiraki, Y., Inoue, H., Kato, Y., Furuya, M., Suzuki, F., 1987.Combined effects of somatomedin-like growth factors with fibrob-last growth factor or epidermal growth factor on DNA synthesisin rabbit chondrocytes. Mol. Cell. Biochem. 76, 185–193.
Hurley, M.M., Abreau, C., Harrison, J., Raisz, L.G., Kream, B.E.,1991. Basic fibroblast growth factors inhibits type 1 collagen geneexpression in osteoblastic MC3T3-E1 cells. J. Bone Miner. Res. 6,S204.
Jin, C.H., Miyaura, C., Ishimi, Y., Hong, M.H., Sato, T., Abe, E.,Suda, T., 1990. Interleukin 1 regulates the expression of osteopon-tin mRNA by osteoblasts. Mol. Cell. Endocr. 74, 221–228.
Leboy, P.S., Beresford, J.N., Devlin, C., Owen, M.E., 1991. Dexam-ethasone induction of osteoblast mRNAs in rat marrow stromalcultures. J. Cell. Physiol. 146, 370–378.
Liang, C.T., Barnes, J., Seeder, J.G., Quartuccio, H.A., Bolander,M., Jeffrey, J.J., Rodan, G.A., 1992. Impaired bone activity inaged rats: alterations at cellular and molecular levels. Bone 13,435–441.
Maniatopoulos, C., Sodek, J., Melcher, A.H., 1988. Bone formationby stromal cells obtained from bone of young adult rats. Cell Tiss.Res. 254, 317–330.
McCarthy, T.L., Centrella, M., Canalis, E., 1989. Effects of fibroblastgrowth factors on deoxyribonucleic acid and collagen synthesis inrat parietal bone cells. Endocrinology 125, 2118–2126.
Nagai, H., Tsukuda, R., Mayahara, H., 1995. Effects of basic fibrob-last growth factor (bFGF) on bone formation in growing rats.Bone 16, 367–373.
Noda, M., Yoon, K., Thiede, M., Buenaga, R., Weiss, M., Henthorn,P., Harris, H., Rodan, G.A., 1987. cDNA cloning of alkalinephosphatase from rat osteosarcoma (ROS 17/2.8) cells. J. BoneMiner. Res. 2, 161–164.
Noff, D., Pitaru, S., Savion, N., 1989. Basic fibroblast growth factorenhances the capacity of bone marrow cells to form bone likenodules in vitro. FEBS Lett. 250, 619–621.
Nomura, S., Willls, A.J., Edwards, D.R., Heath, J.K., Hogan,B.L.M., 1988. Developmental expression of 2ar (osteopontin) andSPARC (osteonectin) RNA as revealed by in situ hybridization. J.Cell Biol. 106, 441–450.
Oldberg, A., Franzen, A., Heinegard, D., 1986. Cloning and sequenceanalysis of rat bone sialoprotein (osteopontin) cDNA reveals anArg-Gly-Asp cell-binding sequence. Proc. Natl. Acad. Sci. USA83, 8819–8823.
Owen, T.A., Aronow, M., Shalhoub, V., Barone, L.M., Wilming, L.,Tassinari. M,.S., Kennedy, M.B., Pockwinse, S., Lian, J.B., Stein,G.S., 1990. Progressive development of the rat osteoblast pheno-
type in vitro: reciprocal relationships in expression of genesassociated with osteoblast proliferation and differentiation duringformation of bone extracellular matrix. J Cell. Physiol. 143,420–430.
Owen, M., Friedenstein, A.J., 1988. Marrow stromal cells: marrowderived osteogenic precursors. In: Evered, D., Harnett, S (Ed.),Foundation Symposium 136: Cell and Molecular Biology ofVertebrate Hard Tissue. Wiley, Chichester, pp. 42–60.
Pitaru, S., Kotev-Emeth, S., Noff, D., Kaffuler, S., Savion, N., 1993.Effect of basic fibroblast growth factor on the growth and differ-entiation of adult stromal bone marrow cells: enhanced develop-ment of mineralized bone-like tissue in culture. J. Bone Miner.Res. 8, 919–929.
Quarto, R., Thomas, D., Liang, C.T., 1995. Bone progenitor celldeficits and the age-associated decline in bone repair capacity.Calcif. Tiss. Int. 56, 123–129.
Reinholt, F.P., Hultenby, K., Oldberg, A., Heinengard, D., 1990.Osteopontin—a possible anchor of osteoclasts to bone. Proc.Natl. Acad. Sci. USA 83, 4473–4475.
Rodan, S.B., Wesolowski, G., Yoon, K., Rodan, G.A., 1989. Oppos-ing effects of fibroblast growth factor and pertussis toxin onalkaline phosphatase, osteopontin, osteocalcin, and type I colla-gen. J. Biol. Chem. 264, 19934–19941.
Rosenthal, S.M., Brown, E.J., Brunetti, A., Goldfine, I.D., 1991.Fibroblast growth factor inhibits insulin-like growth factor-II(IGF-II) gene expression and increases IGF-I receptor abundancein BC3H-1 muscle cells. Mol. Endocrinol. 5, 678–684.
Shing, Y., Folkman, J., Sullivan, R., Butterfield, C., Murray, J.,Klagsburn, M., 1984. Heparin affinity: purification of a tumorderived capillary endothelial cell growth factor. Science 223,1926–1928.
Simmons, H.A., Raisz, L.G., 1991. Effects of acid and basic fibrob-last growth factor and heparin on resorption of cultured fetal ratlong bones. J. Bone Miner. Res. 6, 1301–1305.
Slootweg, M.C., Hoogerbrugge, C.M., de Pooter, T.L., Duursma,S.A., van Buul Offers, S.C., 1990. The presence of classicalinsulin-like growth factor (IGF) type-I and -II receptors on mouseosteoblast:autocrine/paracrine growth effect of IGFs. Endocrinol-ogy 125, 271–277.
Sporn, M.B., Todara, G., 1980. Autocrine secretion and malignanttransformation of cells. N. Engl. J. Med. 303, 878–880.
Tanaka, H., Barnes, J., Liang, C.T., 1996. Effect of age on theexpression of insulin-like growth factor-I, interleukin-6 and trans-forming growth factor-b mRNAs in rat femurs following marrowablation. Bone 18, 473–478.
Tanaka, H., Liang, C.T., 1995. Effect of platelet-derived growthfactor on DNA synthesis and gene expression in bone marrowstromal cells derived from adult and old rats. J. Cell. Physiol. 164,367–375.
Tanaka, H., Liang, C.T., 1996. Mitogenic activity but not phenotypeexpression of rat osteoprogenitor cells in response to IGF-I isimpaired in aged rats. Mech. Ageing Dev. 92, 1–10.
Tanaka, H., Quarto, R., Williams, S., Barnes, J., Liang, C.T., 1994.In vivo and in vitro effects of insulin-like growth factor-I (IGF-I)on femoral mRNA expression in old rats. Bone 15, 647–653.
Wakisaka, A., Tanaka, H., Barnes, J., Liang, C.T., 1998. Effect oflocally infused IGF-I on femoral gene expression and boneturnover activity in old rats. J. Bone Miner. Res. 13, 13–19.
Wang, J.S., Aspenberg, P., 1996. Basic fibroblast growth factorpromotes ingrowth in porous hydroxyapatite. Clin. Orthop. 333,252–260.
Yoon, K., Buenaga, R., Rodan, G.A., 1987. Tissue specificity anddevelopmental expression of rat osteopontin. Biochem. Biophys.Res. Comm. 148, 1129–1136.

H. Tanaka et al. / Molecular and Cellular Endocrinology 150 (1999) 1–1010
Yoon, K., Rutledge, S.J.C., Buenaga, R., Rodan, G.A., 1988. Char-acterization of the rat osteocalcin gene: stimulation of promoteractivity by 1,25-dihydroxyvitamin D3. Biochemistry 27, 8521–8526.
Young, M.F., Kerr, J.M., Termine, J.D., Wewer, U.M., Ge Wang,M., Mcbride, O.W., Fisher, L.W., 1990. cDNa cloning, mRNAdistribution and heterogeneity, chromosomal location, and RFLPanalysis of human osteopontin (OPN). Genomics 7, 491–502.
.
Copyright © 2022 FDOKUMEN




















